

Abstract
Background: Human amniotic epithelial cells (hAECs) are seed cells used to treat acute myocardial infarction (AMI), but their mechanism remains unclear.
Methods: We cultured hAECs and extracted exosomes from culture supernatants. Next, we established a stable AMI model in rats and treated them with hAECs, exosomes, or PBS. We assess cardiac function after treatment by echocardiography. Additionally, heart tissues were collected and analyzed by Masson’s trichrome staining. We conducted the tube formation and apoptosis assays to explore the potential mechanisms.
Results: Cardiac function was improved, and tissue fibrosis was decreased following implantation of hAECs and their exosomes. Echocardiography showed that the EF and FS were lower in the control group than in the hAEC and exosome groups, and that the LVEDD and LVESD were higher in the control group (P<0.05). Masson’s trichrome staining showed that the fibrotic area was larger in the control group. Tube formation was more efficient in the hAEC and exosome groups (P<0.0001). Additionally, the apoptosis rates of myocardial cells in the hAEC and exosome groups were significantly decreased (P<0.0001).
Conclusions: hAECs and their exosomes improved the cardiac function of rats after AMI by promoting angiogenesis and reducing the apoptosis of cardiac myocytes.
Keywords: human amniotic epithelial cells, exosome, acute myocardial infarction, rat
INTRODUCTION
Mortality from cardiovascular diseases has been increasing recently, accounting for approximately one-third of all deaths in the population, and has posed a hefty burden to the social economy, according to the American Heart Association [1]. Therapies for myocardial infarction have gradually developed in recent years. Thrombolysis, coronary artery bypass grafting (CABG) and percutaneous coronary intervention (PCI) could rescue myocardial cells that are close to death; however, heart failure occurs inevitably [2]. Thus, strategies to repair dying myocardial cells and improve prognosis are needed. Scientists have focused increasingly on cell therapy, and many cell types have been demonstrated to improve cardiac function after acute myocardial infarction (AMI), such as bone marrow-derived stromal cells, embryonic stem cells, mesenchymal stem cells and induced pluripotent stem cells [3]. However, the application of these cells remains limited because the cell survival rate is low in vivo and the cells can become tumorigenic or cause inflammatory problems [4, 5]. Accumulating evidence has shown that stem cells function through autocrine, paracrine and other ways and play a role in infarcted myocardium repair. Scientists have gradually abandoned the concept of stem cells by proliferation differentiation into cardiomyocytes as a protection [6].
Exosomes are important factors secreted by stem cells. Because of their advantages of low immune exclusion and easy access, stem cell-derived exosomes have attracted increasing attention as a new noncellular therapy [7]. Exosomes are disk-like vesicles of varying sizes, with a diameter of approximately 30-150 nm. They play an important role in the intercellular communication process through the lncRNAs, siRNAs, miRNAs and mRNAs contained within them, as well as proteins, DNA, lipids and other substances [8]. The functions of exosomes are gradually being revealed. Studies have reported that exosomes secreted by stem cells have similar biological functions as maternal stem cells [9, 10].
Human amniotic epithelial cells (hAECs) are human amniotic stem cells that can be isolated from human placental tissue and show low immunogenicity, low tumorigenicity and anti-inflammatory properties [11]. Studies by Yi-Sun Song and Yu-Ying Wang have shown that hAECs improve the cardiac function after myocardial infarction in rats by differentiating into cardiac myocytes, but the mechanism remains unclear [12, 13]. Research on exosomes has been increasing since 2010, when Lai et al. isolated exosomes and used them to treat mice with AMI and found that exosomes could significantly reduce the myocardial infarction area and might play a protective role in promoting angiogenesis and inhibiting apoptosis [14]. Lee WH et al. demonstrated that exosomes secreted by embryonic stem cells and human induced pluripotent stem cells contain small RNAs (miRs) and long noncoding RNAs (lncRNAs) that can promote angiogenesis and significantly improve cardiac function after myocardial infarction in nude mice [15].
Exosome and hAECs have good prospects in treating acute myocardial infarction, and the few related studies have shown controversial results. Therefore, we further explored the effect of hAECs and exosomes secreted by hAECs on acute myocardial infarction.
MATERIALS AND METHODS
Cell culture
Human primary amniotic epithelial cells were provided by Icell (Shanghai, China; http://www.icellbioscience.com/index), human umbilical vein endothelial cells (HUVECs) and myocardial cells (H9C2 cells) were purchased from the Stem Cell Bank (Chinese Academy of Science; http://www.cellbank.org.cn/). All the cells were cultured in complete medium comprising 89% Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen) supplemented with high glucose, 10% fetal bovine serum (FBS; GIBCO) and 1% penicillin/streptomycin (GIBCO). We selected P4-P6 hAECs for our next experiment, and the conditioned medium was collected to extract exosomes.
EdU labeling and hAEC staining
The P4-P6 hAECs were labeled with EdU, an analogue of thymidine nucleoside (T). In the cell proliferation phase, EdU is substituted for T infiltration during the replication of the DNA molecule. Based on the specificity of the fluorescent dyes in the EdU and using the Apollo® system, cell proliferation could be detected using the EdU tag. We added EdU according to the manufacturer’s instructions of the EdU kit (Ribobio, https://www.ribobio.com/).
Isolation and identification of exosomes
The conditioned medium of P4-P6 hAECs was collected in a 50-ml centrifuge tube and centrifuged at 2000 g for 30 minutes to remove dead cells and other cell fragments. Next, the supernatant was transferred to another aseptic centrifuge tube, followed by the addition of 0.5 volumes of exosome isolation reagent (Thermo Fisher Scientific) at 4° C. After 24 hours, the mixture was passed through a 0.22-μm cell filter, and then we placed the mixture in a centrifuge for 1 hour at 10000 g, discarded the supernatant, leaving the exosomes on the wall of the centrifuge tube. Phosphate-buffered saline (PBS) was used to resuspend the exosomes and store them at -80° C for the next step. The protein concentration of the extracted component was detected using the bicinchoninic acid (BCA) protein assay (Solarbio, China). CD63(BD) and CD81(BD) were identified by flow cytometry (Accuri C6 flow cytometer; BD). We observed the lipid bilayer vesicles using electron microscopy (H-7650; Hitachi) and conducted nanoparticle tracking analysis using the ZETASIZER Nano series-Nano-ZS.
PKH-26 labeling and exosome staining
The exosomes were labeled with PKH-26 (Sigma), a fluorescent dye widely used to label exosomes, according to the manufacturer’s instructions. Next, the labeled exosomes were stored at -80° C for implantation into rats with acute myocardial infarction.
Induction of myocardial infarction, cells and exosome transplantation
We purchased SD rats (male, 270–320 g) from Soochow University Experimental Animal Center and conducted animal experiments according to the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH publication No. 85–23, revised 1996). Twenty-four rats were randomly divided into three groups: a control group treated with PBS, a group treated with hAECs and a third group treated with exosomes. We generated the acute myocardial infarction model as previously reported [16] with a slight modification. Briefly, the rats were anesthetized with 4% chloral hydrate. First, we performed tracheotomy and connected a breathing machine (instrument plant of Chengdu, China), and then electrocardiography (ECG) followed by thoracotomy was conducted. After exposing the heart well, we ligated it to the left anterior descending (LAD) coronary artery. We estimated the modeling by the changes in the ventricular wall activity and color and ST segment changes of ECG. We transplanted 150 μl of PBS, 1.5*106 hAECs suspended with 150 μl of PBS or 300 μg of exosomes suspended with 150 μl of PBS into 3 sites around the infarcted heart tissue 30 minutes after myocardial infarction. Finally, the thoracic incision was closed carefully. After awakening, the rats were placed back into the cage for intensive monitoring.
Measurement of cardiac function and myocardial infarction size
Echocardiography was performed at baseline and 7, 14 and 28 days after MI surgery by a blinded researcher well trained in the procedure. We used a 250-Hz transducer (SONOS 7500; Philips Medical Systems) and measured the left ventricular end-diastolic diameters (LVEDDs), left ventricular end-systolic diameters (LVESDs) and ejection fraction (EF) and fraction shortening (FS) in 3 consecutive cardiac cycles.
The rats were euthanized 28 days after myocardial infarction, and their heart tissue for Masson’s trichrome staining (Sigma) was removed to measure the size of the infarcted tissue. Briefly, the harvested heart tissues were trimmed carefully and immersed in 4% formaldehyde for 2 days at 4° C and then were transferred to 10%, 20% and 30% sucrose solution for gradual dehydration at 4° C, over a 24-hour period. Next, we used OCT (Sakura) component to embed the tissues and then froze them in dry ice. We then prepared frozen sections of heart tissue perpendicular to the left anterior descending coronary artery with a slice thickness of approximately 6 mm. We stained the sections according to standard procedures, and then the area of infarction was measured using ImageJ pro software (http://www.mediacy.com).
Fluorescence immunohistochemistry
To detect the distribution and activity of transplanted cells and exosomes, the rats were euthanized 7 days after myocardial infarction and the heart tissue was removed for fluorescence immunohistochemistry. The specific treatment of the heart tissue was the same as above. The frozen sections from the hAEC group were stained with EdU, followed by observation under a fluorescence microscope (Olympus). Because PKH-26 is fluorescent, the frozen sections from the exosome group could be observed directly under a fluorescence microscope.
Tube formation assay
Before the tube formation assay, we co-cultured hAECs (cell group) and exosomes (exosome group) using the same generation of HUVECs and then set up a control group, HUVECs without treatment. After coculturing for 48 hours, the prepared HUVECs were used in the tube formation assay. First, we added 50 μl of growth factor-reduced Matrigel (BD Biosciences) to 96-well plates and then stored it in an incubator for 30 minutes at 37° C. At the same time, we prepared HUVECs at the appropriate concentration and then the prepared HUVECs were seeded at a density of 50,000 cells per well of a 96-well plate. After the Matrigel solidified, we incubated the plate at 37° C. After 8 hours, we observed the images of tube formation using an inverted microscope (Olympus) and counted all the interconnecting tubes in each group using ImageJ pro software.
Apoptosis detection of myocardial cells
We co-cultured hAECs (cell group) and exosomes (exosome group) with the same generation of H9C2 cells for 48 hours and set up a control group, H9C2 cells without treatment. Next, we replaced the medium with sugar-free medium and placed the H9C2 cells of each group into the oxygen-free environment for 5 hours at 37° C. We detected the apoptosis of H9C2 cells by flow cytometry. Annexin V-FITC/PI dyes distinguish dead cells, apoptosis cells and live cells according to different staining patterns. We stained H9C2 cells according to the manufacturer’s instructions of the Annexin V-FITC/PI kit (BD) and used flow cytometry (Beckman FC 500) to analyze the rate of apoptosis cells.
Statistical analysis
All the data in the experiment were presented as means ± standard deviation, and t test was used for comparison analysis between the experimental and control groups. P<0.05 indicated statistical significance between the groups. One-way ANOVA was used for comparison among the groups. SPSS 15.0 and GraphPad Prism 8.0 statistical software packages were used to analyze the data.
RESULTS
hAECs were cultured, and the exosomes were extracted
hAECs were polygonal and showed adherent growth ability (Figure 1A). The cell proliferation rate slowed down significantly after P8, and individual cells grew larger and flattened. Therefore, P4-P6 cells were selected for all experiments. We labeled the hAECs to be implanted in vivo by EdU to identify them by in vitro immunofluorescence (Figure 1B). We extracted exosomes from 300 μg of the supernatants from each T75 cell culture bottle according to the manufacturer’s instructions of the exosome extraction kit (Figure 2A) and confirmed their identity in subsequent assays. Electron microscopy showed that the exosomes were oval and round, with variable sizes of lipid-containing cystic vesicles (Figure 2B). The results of surface marker antigen staining of CD63 and CD81 showed that the positive labeling rates were 86.2±1.4% and 94.9±1.1%, respectively (Figure 2C), and the average value (128.5±8.2 nm) and main distribution (136.2±1.7 nm) of the extracted material were within the range of exosomes (Figure 2D). These results prove that exosomes were extracted, provided a reliable guarantee for subsequent analysis.
Figure 1.

Characterization and labeling of human amniotic epithelial cells (hAECs). (A) The morphology of hAECs was observed under a microscope. (B) Immunofluorescence of hAECs labeled with EdU in vitro.
Figure 2.

Identification of exosomes deprived from hAECs. (A) Protein concentration curve from the BCA test. (B) The morphology of exosomes was observed under an electron microscope. (C) The phenotype of exosomes for CD63 and CD81 was identified by flow cytometry. (D) The diameter distribution of exosomes was measured by nanoparticle tracking analysis.
The cardiac function is improved by implanting hAECs and their exosomes in a rat acute myocardial infarction model
We established the stable model of acute myocardial infarction in rats. After ligating the LAD coronary artery, ECG showed typical dynamic changes such as changes in ST. Compared with the ST of ECG before coronary artery ligation (Figure 3A), the ST of ECG was elevated approximately 5 minutes after ligating the LAD (Figure 3B). Next, we implanted hAECs and exosomes from the hAEC culture supernatant into the myocardium of acute myocardial infarction rats and set up a control group to avoid the influence of other factors on the results. We then supervised the alteration of cardiac function via echocardiography (Figure 4). The typical graph of each group in the M-mode is shown in Figure 4A, and the ultrasound data were measured at baseline and 7, 14 and 28 days after MI surgery. The postoperative EF and FS were lower in the control group than those in the therapy group (P<0.0001). The LVEDD and LVESD measured at 14 and 28 days postoperatively were significantly higher in the control group than those in the therapy group (P<0.05; Figure 4B). Fluorescence immunohistochemistry confirmed that the hAECs and exosomes were scattered in the myocardial tissue at 7 days after surgery (Figure 5A). Masson’s trichrome staining revealed that the area of tissue fibrosis in the control group was larger at 28 days after surgery, and the difference between groups was significant (P<0.0001; Figure 5B). The infarction area is displayed in Figure 5C. Taken together, the results showed that the hAECs and exosomes secreted by hAECs could improve cardiac function after AMI and decrease the infarction area.
Figure 3.
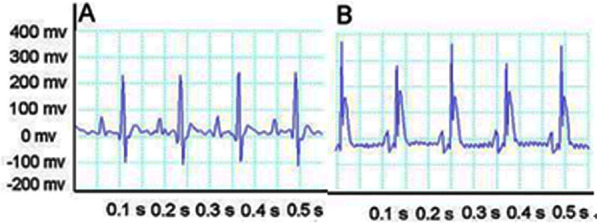
ECG shows typical dynamic changes. (A) ECG before ligating the coronary artery. (B) After ligation, the ST of ECG is elevated.
Figure 4.

Echocardiography shows that cardiac function is improved by implanting hAECs and their exosomes in a rat acute myocardial infarction model. (A) Representative M-mode echocardiography of SD rats before and after MI in the hAEC treatment group, exosome group and control group. (B) The line charts shows the analysis of the left ventricular ejection fraction (EF), left ventricular fractional shortening (FS), left ventricular end-diastolic diameter (LVEDD) and left ventricular end-systolic diameter (LVESD) in SD rats 7, 14 and 28 days after MI, respectively. The data are expressed as the means ± SD, n=8 per group, *P<0.05.
Figure 5.

hAECs and exosomes implanted in the heart decrease the fibrosis area. (A) Immunofluorescence of hAECs in vivo 7 days after MI are shown. (B) Immunofluorescence of exosomes in vivo 7 days after MI are shown. (C) Representative images of four consecutive myocardial slices stained with Masson’s trichrome in the hAEC, exosome and control groups 28 days after MI are shown. (D) The bar graph shows quantitative analysis of the LV fibrosis area. The data are shown as the means ± SD, n=4, *P<0.05.
The protective mechanism is explored in vitro
Finally, the therapeutic mechanisms were explored. We conducted the tube formation assay to assess the potential mechanism of HUVECs and found that both hAECs and exosomes promoted lumen formation ability. In the control group, the HUVECs rarely formed tubes (Figure 6). The lumen lengths measured using image J pro Plus 6.0 were as follows: cell group (Figure 6A): 5251±166.09 μm; exosome group (Figure 6B): 4755.8±157.52 μm; control group (Figure 6C): 3669±236.03 μm. The difference between the experimental and control groups was statistically significant (P<0.05; Figure 6D).
Figure 6.

Lumen formation and quantitative analysis of the lumen length. Both hAECs and exosomes promote lumen formation. (A) Treatment with hAECs. (B) Treatment with exosomes. (C) Control group. (D) The bar graph shows quantitative analysis of the tube length. The data are shown as the means ± SD, n=4, *P<0.05.
Apoptosis plays a major role in AMI. In this experiment, we mainly explored the protection of hAECs and exosomes. The results of cardiomyocyte apoptosis were similar to those of tube formation. Following co-culture with hAECs and exosomes, the apoptosis rate of H9C2 decreased significantly compared with that of the control group. In an oxygen-free environment for 5 hours, the apoptosis rate of each group was as follows: cell group (Figure 7A): 16.76±1.77%; exosome group (Figure 7B): 22.04±1.71%; control group (Figure 7C): 37.46±1.4%. The difference between each group was statistically significant (P<0.05; Figure 7D).
Figure 7.

The apoptosis rate of cardiomyocytes was measured by flow cytometry. The results showed that both hAECs and exosomes decrease apoptosis. (A) Treatment with hAECs. (B) Treatment with exosomes. (C) Control group. (D) The bar graph shows quantitative analysis of early apoptosis. The data are shown as the means ± SD, n=4, *P<0.05.
DISCUSSION
We showed that both hAECs and exosomes improved the cardiac function of rats after acute myocardial infarction and that cardiac function and the myocardial infarction area were similar between treatment with cells or exosomes. Thus, hAECs play a protective role in the treatment of acute myocardial infarction by secreting exosomes. Although many studies have reported that exosomes improve cardiac function, the specific underlying mechanisms remain unclear [17–19]. We conducted the apoptosis detection of myocardial cells and tube formation assay to explore the potential mechanisms in vitro and found that promoting angiogenesis and inhibiting cardiac apoptosis were the protective mechanisms.
hAECs, with strong plasticity, multidirectional differentiation potential, low immunogenicity, low tumorigenic and anti-inflammatory characteristics, have been considered a potential candidate to treat myocardial infarction [11]. Recent studies have demonstrated that hAECs improve cardiac function after myocardial infarction in rats by differentiating into cardiomyocytes [12, 13]; however, the mechanism requires further exploration. Exosome-related studies have gradually increased in recent years, and scientists have reached a consensus that the protection of stem cells is not only related to paracrine and autocrine factors but also influenced by extracellular vesicles, such as exosomes, which are secreted by stem cells [19–21]. Exosomes, whose components include microRNAs, lncRNAs, and siRNAs, are considered a novel therapy in regenerative medicine [22, 23]. Echocardiography showed that the EF and FS in the therapy group were significantly higher than those in the control group 7, 14 and 28 days after surgery, indicating that the treatment was effective. However, the meaningful decrease in the EF and FS at 28 days compared with 7 days was likely due to the single treatment, ventricular remodeling and expansion of the cardiac cavity, which reduced cardiac function. The LVEDD and LVESD measured 14 and 28 days postoperatively were obviously reduced in the therapy group compared with that in the control group, suggesting that the implanted hAECs and exosomes secreted by hAECs after myocardial infarction can inhibit ventricular remodeling and improve heart function in rats. By contrast, no conspicuous difference was observed in 7 days; ventricular remodeling was not evident, and the measurements indicated no prominent changes. After 28 days postoperatively, although the LVEDD and LVESD in the therapy group were significantly improved, ventricular remodeling still occurred; thus, the cardiac function decreased over time, a finding that was similar to the previous research results of Wang et al. [13]. These results offer new evidence that cardiac function is improved by implanting hAECs and their exosomes in the rat model of acute myocardial infarction.
To clarify the phenomenon demonstrated in vivo and explore the potential mechanisms, we performed relevant in vitro studies. We simulated myocardial infarction in vitro by building an H9C2 ischemic hypoxemia model [24], in which the tube formation assay for angiogenesis [25] was also conducted. We observed that hAECs and exosomes decreased apoptosis and increased angiogenesis in vitro. Additionally, treatment with hAECs was more effective than with exosomes. However, the in vivo results differed, showing that treatment with hAECs and exosomes had similar efficacies. In vivo, we implanted hAECs and exosomes at different doses, and the in vitro intervention procedure was the same. The concentration of implanted exosomes was 300 μg and that of implanted hAECs was 1.5*106 in vivo. However, 1.5*106 hAECs could extract only 100 μg of exosomes, likely explaining the divergent outcomes. We considered that exosomes play a positive role in acute myocardial infarction, in addition to other bioactive factors, such as vascular endothelial growth factor, fibroblast growth factor and circulating microRNAs, which could improve cardiac function [26–30]. We deemed that the role of hAECs was not only to produce exosomes in a protective function but also to cooperate with various biological activity factors. The mystery of exosomes needs more basic experiments to be gradually uncovered. Additionally, the stability, safety and bioavailability of exosomes require further study [31, 32].
We demonstrated that hAECs improved cardiac function after acute myocardial infarction, and the mechanisms of protection include inhibiting apoptosis and promoting angiogenesis. Exosomes, as an important component of stem cell secretion, with many protective substances, high availability, and low immunogenicity, are a potential candidate for therapy [15, 20, 22, 31–33]. Many studies have investigated exosomes. Zhao et al. found that rat bone marrow mesenchymal-derived exosomes can reduce inflammatory infiltration and fibrosis in the myocardial infarction area and promote the survival of rat embryonic myocardial cells (H9C2) under the oxidative stress of H2O2. The specific mechanism is that these exosomes can upregulate mir-24 and mir-29, downregulate mir-130 and mir-34, and further affect Ras, PI3K/Akt, mammalian target protein of rapamycin and other pathways to play a protective role [33]. Arslan et al. demonstrated that exosomes derived from mesenchymal stem cells could enhance myocardial vitality, prevent adverse remodeling after myocardial ischemia/reperfusion injury, and activate the PI3K/Akt pathway by increasing the ATP level to reduce oxidative stress [34]. Therefore, exosomes have great potential in regenerative medicine [35].
Because of experimental limitations, our experiment only proves that hAECs and their exosomes can improve cardiac function after myocardial infarction in rats by promoting angiogenesis and reducing myocardial cell apoptosis; the specific underlying pathway and other potential protective mechanisms have not been explored. In the future, the source of exosomes and mechanism of cell protection and differentiation mediated by exosomes in cardiovascular diseases will be explored.
Footnotes
AUTHOR CONTRIBUTIONS: Yi-Qing Zhang, Lu Hong, Yu-Feng Jiang, Kang-Yun Sun and Ya-Feng Zhou contributed to the conception of the study; Yi-Qing Zhang, Lu Hong, Sheng-Da Hu and Hong-Xia Li performed the experiment; Yi-Qing Zhang, Lu Hong and Ya-Feng Zhou contributed significantly to analysis and manuscript preparation; Yi-Qing Zhang, Lu Hong and Yu-Feng Jiang performed the data analyses and wrote the manuscript; Hong-Xia Li, Ya-Feng Zhou, Gui-Dong Xu, Nan-Nan Zhang and Lang-Biao Xu helped perform the analysis with constructive discussions.
CONFLICTS OF INTEREST: We declare that we do not have any commercial or associative interest that represents a conflicts of interest in connection with the work submitted.
FUNDING: This work was supported by grants from National Natural Science Foundation of China (81170174), Natural Scientific Fund of Jiangsu province (BK20161226), Jiangsu Province’s Key Provincial Talents Program (ZDRCA2016043), Jiangsu Province’s 333 High-Level Talents Project (BRA2017539), Gusu School, Nanjing Medical University Scientific Research Program (GSKY20210202), Suzhou Science and Technology Development Plan Project (SYSD2020143). The funders had no roles in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
This corresponding author has a verified history of publications using a personal email address for correspondence
REFERENCES
- 1.Warner JJ, Benjamin IJ, Churchwell K, Firestone G, Gardner TJ, Johnson JC, Ng-Osorio J, Rodriguez CJ, Todman L, Yaffe K, Yancy CW, Harrington RA, and American Heart Association Advocacy Coordinating Committee. Advancing Healthcare Reform: The American Heart Association’s 2020 Statement of Principles for Adequate, Accessible, and Affordable Health Care: A Presidential Advisory From the American Heart Association. Circulation. 2020; 141:e601–14. 10.1161/CIR.0000000000000759 [DOI] [PubMed] [Google Scholar]
- 2.Nabel EG, Braunwald E. A tale of coronary artery disease and myocardial infarction. N Engl J Med. 2012; 366:54–63. 10.1056/NEJMra1112570 [DOI] [PubMed] [Google Scholar]
- 3.Bolli R, Chugh AR, D’Amario D, Loughran JH, Stoddard MF, Ikram S, Beache GM, Wagner SG, Leri A, Hosoda T, Sanada F, Elmore JB, Goichberg P, et al. Cardiac stem cells in patients with ischaemic cardiomyopathy (SCIPIO): initial results of a randomised phase 1 trial. Lancet. 2011; 378:1847–57. 10.1016/S0140-6736(11)61590-0 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 4.Toma C, Pittenger MF, Cahill KS, Byrne BJ, Kessler PD. Human mesenchymal stem cells differentiate to a cardiomyocyte phenotype in the adult murine heart. Circulation. 2002; 105:93–98. 10.1161/hc0102.101442 [DOI] [PubMed] [Google Scholar]
- 5.Mirotsou M, Jayawardena TM, Schmeckpeper J, Gnecchi M, Dzau VJ. Paracrine mechanisms of stem cell reparative and regenerative actions in the heart. J Mol Cell Cardiol. 2011; 50:280–89. 10.1016/j.yjmcc.2010.08.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pearl JI, Lee AS, Leveson-Gower DB, Sun N, Ghosh Z, Lan F, Ransohoff J, Negrin RS, Davis MM, Wu JC. Short-term immunosuppression promotes engraftment of embryonic and induced pluripotent stem cells. Cell Stem Cell. 2011; 8:309–17. 10.1016/j.stem.2011.01.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Huang L, Ma W, Ma Y, Feng D, Chen H, Cai B. Exosomes in mesenchymal stem cells, a new therapeutic strategy for cardiovascular diseases? Int J Biol Sci. 2015; 11:238–45. 10.7150/ijbs.10725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Simpson RJ, Jensen SS, Lim JW. Proteomic profiling of exosomes: current perspectives. Proteomics. 2008; 8:4083–99. 10.1002/pmic.200800109 [DOI] [PubMed] [Google Scholar]
- 9.Chen L, Wang Y, Pan Y, Zhang L, Shen C, Qin G, Ashraf M, Weintraub N, Ma G, Tang Y. Cardiac progenitor-derived exosomes protect ischemic myocardium from acute ischemia/reperfusion injury. Biochem Biophys Res Commun. 2013; 431:566–71. 10.1016/j.bbrc.2013.01.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xin H, Li Y, Liu Z, Wang X, Shang X, Cui Y, Zhang ZG, Chopp M. MiR-133b promotes neural plasticity and functional recovery after treatment of stroke with multipotent mesenchymal stromal cells in rats via transfer of exosome-enriched extracellular particles. Stem Cells. 2013; 31:2737–46. 10.1002/stem.1409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sakuragawa N, Tohyama J, Yamamoto H. Immunostaining of human amniotic epithelial cells: possible use as a transgene carrier in gene therapy for inborn errors of metabolism. Cell Transplant. 1995; 4:343–46. 10.1016/0963-6897(95)00008-l [DOI] [PubMed] [Google Scholar]
- 12.Song YS, Joo HW, Park IH, Shen GY, Lee Y, Shin JH, Kim H, Shin IS, Kim KS. Transplanted Human Amniotic Epithelial Cells Secrete Paracrine Proangiogenic Cytokines in Rat Model of Myocardial Infarction. Cell Transplant. 2015; 24:2055–64. 10.3727/096368914X685609 [DOI] [PubMed] [Google Scholar]
- 13.Wang Y, Fang N, Chen D, Yu L, Zhang T, Zhao C. Human amnion-derived stem cells differentiate into myocardial cell delay ventricular remodeling and improve Cardiac function in rats with acute myocardial infarction. Chinese Journal of Ger. 2013; 33:1583–86. 10.7666/d.y1736499 [DOI] [Google Scholar]
- 14.Lai RC, Arslan F, Lee MM, Sze NS, Choo A, Chen TS, Salto-Tellez M, Timmers L, Lee CN, El Oakley RM, Pasterkamp G, de Kleijn DP, Lim SK. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010; 4:214–22. 10.1016/j.scr.2009.12.003 [DOI] [PubMed] [Google Scholar]
- 15.Lee WH, Chen WY, Shao NY, Xiao D, Qin X, Baker N, Bae HR, Wei TT, Wang Y, Shukla P, Wu H, Kodo K, Ong SG, Wu JC. Comparison of Non-Coding RNAs in Exosomes and Functional Efficacy of Human Embryonic Stem Cell- versus Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Stem Cells. 2017; 35:2138–49. 10.1002/stem.2669 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Iso Y, Rao KS, Poole CN, Zaman AK, Curril I, Sobel BE, Kajstura J, Anversa P, Spees JL. Priming with ligands secreted by human stromal progenitor cells promotes grafts of cardiac stem/progenitor cells after myocardial infarction. Stem Cells. 2014; 32:674–83. 10.1002/stem.1546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Xin H, Li Y, Cui Y, Yang JJ, Zhang ZG, Chopp M. Systemic administration of exosomes released from mesenchymal stromal cells promote functional recovery and neurovascular plasticity after stroke in rats. J Cereb Blood Flow Metab. 2013; 33:1711–15. 10.1038/jcbfm.2013.152 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vandergriff AC, de Andrade JB, Tang J, Hensley MT, Piedrahita JA, Caranasos TG, Cheng K. Intravenous Cardiac Stem Cell-Derived Exosomes Ameliorate Cardiac Dysfunction in Doxorubicin Induced Dilated Cardiomyopathy. Stem Cells Int. 2015; 2015:960926. 10.1155/2015/960926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cervio E, Barile L, Moccetti T, Vassalli G. Exosomes for Intramyocardial Intercellular Communication. Stem Cells Int. 2015; 2015:482171. 10.1155/2015/482171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lyu L, Wang H, Li B, Qin Q, Qi L, Nagarkatti M, Nagarkatti P, Janicki JS, Wang XL, Cui T. A critical role of cardiac fibroblast-derived exosomes in activating renin angiotensin system in cardiomyocytes. J Mol Cell Cardiol. 2015; 89:268–79. 10.1016/j.yjmcc.2015.10.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Garcia NA, Moncayo-Arlandi J, Sepulveda P, Diez-Juan A. Cardiomyocyte exosomes regulate glycolytic flux in endothelium by direct transfer of GLUT transporters and glycolytic enzymes. Cardiovasc Res. 2016; 109:397–408. 10.1093/cvr/cvv260 [DOI] [PubMed] [Google Scholar]
- 22.Ailawadi S, Wang X, Gu H, Fan GC. Pathologic function and therapeutic potential of exosomes in cardiovascular disease. Biochim Biophys Acta. 2015; 1852:1–11. 10.1016/j.bbadis.2014.10.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Motavaf M, Pakravan K, Babashah S, Malekvandfard F, Masoumi M, Sadeghizadeh M. Therapeutic application of mesenchymal stem cell-derived exosomes: A promising cell-free therapeutic strategy in regenerative medicine. Cell Mol Biol (Noisy-le-grand). 2016; 62:74–79. [PubMed] [Google Scholar]
- 24.Li R, Geng HH, Xiao J, Qin XT, Wang F, Xing JH, Xia YF, Mao Y, Liang JW, Ji XP. miR-7a/b attenuates post-myocardial infarction remodeling and protects H9c2 cardiomyoblast against hypoxia-induced apoptosis involving Sp1 and PARP-1. Sci Rep. 2016; 6:29082. 10.1038/srep29082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhang B, Wu X, Zhang X, Sun Y, Yan Y, Shi H, Zhu Y, Wu L, Pan Z, Zhu W, Qian H, Xu W. Human umbilical cord mesenchymal stem cell exosomes enhance angiogenesis through the Wnt4/β-catenin pathway. Stem Cells Transl Med. 2015; 4:513–22. 10.5966/sctm.2014-0267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Creemers EE, Tijsen AJ, Pinto YM. Circulating microRNAs: novel biomarkers and extracellular communicators in cardiovascular disease? Circ Res. 2012; 110:483–95. 10.1161/CIRCRESAHA.111.247452 [DOI] [PubMed] [Google Scholar]
- 27.Kuehbacher A, Urbich C, Zeiher AM, Dimmeler S. Role of Dicer and Drosha for endothelial microRNA expression and angiogenesis. Circ Res. 2007; 101:59–68. 10.1161/CIRCRESAHA.107.153916 [DOI] [PubMed] [Google Scholar]
- 28.Welten SM, Goossens EA, Quax PH, Nossent AY. The multifactorial nature of microRNAs in vascular remodelling. Cardiovasc Res. 2016; 110:6–22. 10.1093/cvr/cvw039 [DOI] [PubMed] [Google Scholar]
- 29.Bussche L, Van de Walle GR. Peripheral Blood-Derived Mesenchymal Stromal Cells Promote Angiogenesis via Paracrine Stimulation of Vascular Endothelial Growth Factor Secretion in the Equine Model. Stem Cells Transl Med. 2014; 3:1514–25. 10.5966/sctm.2014-0138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Deuse T, Peter C, Fedak PW, Doyle T, Reichenspurner H, Zimmermann WH, Eschenhagen T, Stein W, Wu JC, Robbins RC, Schrepfer S. Hepatocyte growth factor or vascular endothelial growth factor gene transfer maximizes mesenchymal stem cell-based myocardial salvage after acute myocardial infarction. Circulation. 2009. (Suppl 11); 120:S247–54. 10.1161/CIRCULATIONAHA.108.843680 [DOI] [PubMed] [Google Scholar]
- 31.Barile L, Moccetti T, Marbán E, Vassalli G. Roles of exosomes in cardioprotection. Eur Heart J. 2017; 38:1372–79. 10.1093/eurheartj/ehw304 [DOI] [PubMed] [Google Scholar]
- 32.Kishore R, Khan M. More Than Tiny Sacks: Stem Cell Exosomes as Cell-Free Modality for Cardiac Repair. Circ Res. 2016; 118:330–43. 10.1161/CIRCRESAHA.115.307654 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shao L, Zhang Y, Lan B, Wang J, Zhang Z, Zhang L, Xiao P, Meng Q, Geng YJ, Yu XY, Li Y. MiRNA-Sequence Indicates That Mesenchymal Stem Cells and Exosomes Have Similar Mechanism to Enhance Cardiac Repair. Biomed Res Int. 2017; 2017:4150705. 10.1155/2017/4150705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Arslan F, Lai RC, Smeets MB, Akeroyd L, Choo A, Aguor EN, Timmers L, van Rijen HV, Doevendans PA, Pasterkamp G, Lim SK, de Kleijn DP. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013; 10:301–12. 10.1016/j.scr.2013.01.002 [DOI] [PubMed] [Google Scholar]
- 35.Konala VB, Mamidi MK, Bhonde R, Das AK, Pochampally R, Pal R. The current landscape of the mesenchymal stromal cell secretome: A new paradigm for cell-free regeneration. Cytotherapy. 2016; 18:13–24. 10.1016/j.jcyt.2015.10.008 [DOI] [PMC free article] [PubMed] [Google Scholar]