

The overproduction of reactive oxygen species is defined as oxidative stress. While deprived oxygen supply in tissues is known as hypoxia. These mechanisms contribute to the pathogenesis of several retinal diseases, like glaucoma, age-related macular degeneration (AMD), and retinal ischemia. Glaucoma is a neurodegenerative disease defined by a progressive loss of retinal ganglion cells (RGCs) and their axons causing visual field defects, which ultimately leads to blindness. While AMD pathogenesis is further characterized by soft drusen, it involves the retinal pigment epithelium and the Bruch’s membrane - choroid complex. AMD is a disease finally leading to death of the photoreceptors, especially in the macula region, resulting in central vision loss. The pathomechanisms of both diseases are not yet fully understood, but oxidative as well as hypoxic stresses seem to play a crucial. Moreover, hypoxia is also an inducer of oxidative stress by causing cellular stress, followed by mitochondrial stress, in neurodegenerative retinal diseases. In experimental models of retinal degeneration numerous triggers of cell death have been identified, including genetic mutations, intracellularly elevated calcium levels, endoplasmic reticulum stress, as well as hypoxia, and oxidative stress (Grossniklaus et al., 2010; Pang and Clark, 2020).
During hypoxia, the inducible nitric oxide synthase (iNOS) is activated via the hypoxia-inducible factors (HIF) signaling pathway, which subsequently releases large amounts of cytotoxic nitric oxide (NO) (LaManna et al., 2004). With oxidative stress, H2 O2 activates microglia, followed by the secretion of iNOS (Hurst et al., 2017) (Figure 1A). Furthermore, iNOS and HIF-1α are linked by a positive feedback loop and NO plays a role in transcription and stabilization of HIF-1α (Braverman and Stanley, 2017). In result, treatment with an iNOS inhibitor also has an indirect effect on HIF-1α activity and on inflammatory events. In result, iNOS could be a potential option for possible protection of RGCs.
Figure 1.
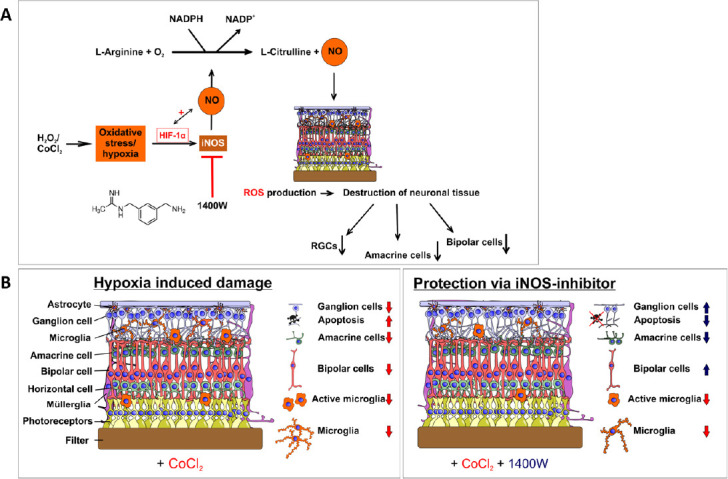
Neuroprotection against hypoxic stress via 1400W.
(A) Schematic overview of cobalt chloride induced hypoxia and the effects on the eye. Cobalt chloride induces hypoxia via hypoxia-inducible factor alpha (Hif-1α), resulting in increased inducible nitric oxide synthase (iNOS) expression, and nitric oxide (NO) production. NO causes an increase in the mRNA expression of HSP70 and iNOS and regulates HIF-1α. This results in a loss of retinal neurons. (B) Overview of the retinal layers and the effects of cobalt chloride and treatment with the iNOS-inhibitor 1400W. Cobalt chloride increased apoptosis and loss of microglia, retinal ganglion, amacrine, and bipolar cells. Basically, all neurons of the inner retina and the microglia were affected. The iNOS-inhibitor prevented apoptosis and loss of retinal ganglion and bipolar cells, but not damage to microglia and amacrine cells. RGC: Retinal ganglion cells; ROS: reactive oxygen species.
In order to simulate hypoxia ex vivo, we adopted a model based on cobalt chloride (CoCl2) induced damage. CoCl2 leads to hypoxia-like processes in retinal cells, like degeneration of RGCs (Kuehn et al., 2017; Maliha et al., 2019). In murine models, low CoCl2 doses cause a pathogenesis similar to inherited retinal degeneration. High concentrations of CoCl2 induce cytotoxic mechanisms by binding the oxygen-dependent region of hypoxia-induced factor-1a (HIF-1α) and therefore preventing the degradation process of HIF-1α, resulting in the induction of different hypoxic genes (Hellwig-Burgel et al., 2005).
Application of CoCl2 on pig retinal organ cultures resulted in a strong retina degeneration, especially a RGC loss (Kuehn et al., 2017). This degeneration was already visible after 4 days and progressed up to the later investigated time point (8 days). A loss of amacrine and bipolar cells was also noted in porcine organ cultures treated with CoCl2. Additionally, mRNA expression levels of HIF-1α and heat shock protein 70 (HSP70) were increased.
The iNOS-inhibitor 1400W: 1400W (N-(3-aminomethyl)-benzylacetamidine) is a specific irreversible inhibitor of iNOS (Zhu et al., 2005). It has a molecular weight of 250.17 g/mol with a structural formula of C10 H15 N3 × 2 HCl. The iNOS-catalyzed reaction of L-arginine to L-citrulline and NO is inhibited (Garvey et al., 1997). Hence, iNOS cannot release large amounts of cytotoxic NO in response to hypoxia or oxidative stress (Betteridge, 2000). Consequently, the iNOS-mediated cell death should be reduced by this inhibitor. Furthermore, the iNOS-inhibitor itself prevents the formation of pathophysiological amounts of NO (Figure 1A).
Inhibiting iNOS in retinal degeneration: The neuroprotective effect of the iNOS inhibitor 1400W has already been proven in various studies. For example, in a cerebral ischemia rat model, the use of 1400W resulted in a reduced infarct size, an improved neurological score, and a reduced weight loss of the animals (Parmentier et al., 1999). The role of iNOS was also investigated in iNOS knock-out mice or by chemical inhibition via 1400W after ischemic retinopathy. In both models, the role of iNOS in this disease via a reduced apoptosis rate and less angiogenesis was shown (Sennlaub et al., 2002).
We previously evaluated the neuroprotective effect of the iNOS-inhibitor 1400W on porcine retinae organ cultures after CoCl2 induced neurodegeneration (Hurst et al., 2020). The inhibitor was designed to minimize the potential damage to RGCs caused by the enzyme iNOS. Based on our previous studies, a cultivation period of four and eight days was determined. Damage with CoCl2 occurs already at day one in this organ culture cultivation. Therefore, treatment with the iNOS-inhibitor was initiated on the first day and lasted for 72 hours. In a preceding study, we compared different CoCl2 concentrations, namely 100, 300 and 500 μM, in regard to their damaging effects on porcine retina organ cultures (Kuehn et al., 2017). Based on the results from this study, it was concluded, that 300 μM CoCl2 should be used for studies analyzing possible neuroprotective agents. In another pilot experiment, different concentrations of 1400W were applied to CoCl2 damaged retinae.
CoCl2 leads to an increase in oxidative stress and activates transcription of HIF-1α (up to 1.9-fold). We analyzed the expression of HIF-1α after treatment with the iNOS-inhibitor and a significant reduction of HIF-1α was found. On the other hand, due to CoCl2 application, iNOS (5.2-fold) and HSP70 (up to 38.6-fold) expression increased. This effect was counteracted by the iNOS-inhibitor (down to 1.7-fold for iNOS and 2.5-fold for HSP70) (Hurst et al., 2020).
Regarding RGCs, after four days of cultivation, the CoCl2 group showed a significant RGC loss of 25.6 ± 2.5 RBPMS +cells/mm (P = 0.0002) compared to the control group with 40.2 ± 1.0 RBPMS +cells/mm. Strong neuroprotection was evident in the group treated with CoCl2 + iNOS inhibitor with 33.9 ± 2.2 RBPMS +cells/mm compared to CoCl2 damaged retinae (P = 0.02). The control group and the CoCl2 + iNOS inhibitor were comparable in cell numbers (P = 0.10) over time. The control group had the most RGCs (31.7 ± 2.0 RBPMS +cells/mm) followed by the iNOS inhibitor treated group (28.5 ± 1.8 RBPMS +cells/mm). The group damaged through CoCl2 displayed significantly fewer cells with 19.4 ± 0.9 RBPMS +cells/mm (P = 0.0001), suggesting neuroprotective effects of 1400W on RGCs (Hurst et al., 2020).
The use of the iNOS inhibitor in the CoCl2 degeneration model also prevented the loss of bipolar cells. Although no differences were found after four days, the degenerative effect of CoCl2 after eight days were diminished. In controls, we detected 59.2 ± 7.8 PKCa +cells/mm, whereas CoCl2 samples had 17.1 ± 14.9 PKCa +cells/mm (P = 0.0001) and 45.5 ± 7.1 PKCa +cells/mm were seen after 1400W treatment (P = 0.0017). However, no protective effect on the microglia and amacrine cells could be observed (Hurst et al., 2020).
In summary, treatment with the iNOS inhibitor 1400W showed a positive effect on RGCs and bipolar cells after both 4 and 8 days and ensured their survival under hypoxic stress (Figure 1B; Hurst et al., 2020).
1400W and hypoxia stress: Hydrogen peroxide (H2 O2) can be used to produce radical oxygen species and thereby induce oxidative stress ex vivo (Hurst et al., 2017). The application of H2 O2 to porcine retinal organ cultures resulted in severe RGC degeneration with persistent apoptosis after 4 and 8 days. In addition, a strong microglial response was observable, which was indicated by increased expression of microglial markers and stress factors (Hurst et al., 2017). The iNOS inhibitor 1400W significantly protected the RGCs from H2 O2 induced oxidative stress after 4 and 8 days. In addition, the destruction of bipolar cells could be reversed by treatment with the inhibitor. In contrast, the number of amacrine cells remained unchanged both by the H2 O2 damage and by treatment with the inhibitor (Joachim et al., 2020).
Conclusion: Both noxious agents were able to induce degeneration in porcine ex vivo cultured retinae, which corresponds to the pathophysiology of various diseases, like glaucoma. To this end, the focus of these studies was on the irreversible loss of RGCs. The evaluation of this therapeutic approach leads to interesting and promising results: On one hand, we noted that the loss of RGCs can be reduced and on the other hand, these ex vivo degeneration models of the porcine retina are suitable for an initial therapy testing. In the future, this organ culture system could be a reliable treatment-screening model.
Hypothesis and future directions: The advantage of this iNOS-inhibitor is that it tackles both, hypoxia and oxidative stress. Both factors play a crucial role in several neuronal diseases especially in disease progression and extent. We previously observed that the inhibitor in two stress models protected RGC death. It would be interesting to combine the two stressors, CoCl2 and H2 O2 , in one approach. In addition, in vivo experiments are necessary to confirm the protective effects of iNOS inhibitors.
Although AMD and glaucoma are very different diseases, both stressors play a crucial role in these diseases. Oxidative stress and hypoxia also play a crucial role in AMD. AMD is the main cause for blindness in the industrialized countries with an estimated 67 million people currently affected in the EU. This number is steadily increasing with the ageing population. AMD is a multifactorial disease; besides hypoxia and oxidative stress, further pathomechanisms include inflammation, complement dysregulation, retinal lipid alterations, and pro-angiogenic signaling. Genetic disposition as well as poor nutrition, smoking, and the lack of physical exercise are also factors affecting AMD. At a cellular level, AMD pathogenesis takes place as interaction between choroid, Bruch’s membrane, retinal pigment epithelium, and photoreceptors. The degeneration of the retinal pigment epithelium is partially caused by oxidative damage and accumulation of undigested material in and under the retinal pigment epithelium, leading to deterioration of nutrient supply, maintenance of photoreceptors, and the choriocapillaris, which worsens the degeneration exponentially. Subsequently, hypoxia in the tissue leads to compensatory release of pro-angiogenic cytokines. Furthermore, the degenerative processes activate inflammatory pathways, which attract cells of the monocytic lineage and lead to additional pro-inflammatory cytokine secretion. All together, these effects finally lead to irreversible cell death of the photoreceptors. In spite of the introduction of the anti-vascular endothelial growth factors therapy, which slows down the progression of one form of AMD, the therapeutic options are still far from satisfactory. No therapeutic options exist to slow or halt the progression from early to late AMD, and treatment possibilities for the early forms of AMD are lacking. Consequently, there is an urgent need for the development of novel therapeutic approaches that would allow fighting blindness in an early stage. Inhibiting iNOS via 1400W or another inhibitor would address both pathways and offer chances of treating retinal diseases like AMD. Studies evaluating these effects in AMD models are currently being conducted should help to develop novel treatment options.
We thank all the collaborators and coauthors who thereby contributed to this perspective article, especially José Hurst and Ana Mueller-Buehl. We also thank Sami Ullah and Teresa Tsai for their support concerning the figures as well as for critical proof reading.
This work was supported in part by the set Stiftung.
Additional file: Open peer review report 1 (84.4KB, pdf) .
Footnotes
P-Reviewer: Liu X; C-Editors: Zhao M, Li JY; T-Editor: Jia Y
Copyright license agreement: The Copyright License Agreement has been signed by both authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewer: Xiaorong Liu, University of Virginia, USA.
References
- 1.Betteridge DJ. What is oxidative stress. Metabolism. 2000;49:3–8. doi: 10.1016/s0026-0495(00)80077-3. [DOI] [PubMed] [Google Scholar]
- 2.Braverman J, Stanley SA. Nitric oxide modulates macrophage responses to mycobacterium tuberculosis infection through activation of HIF-1α and repression of NF-κB. J Immunol. 2017;199:1805–1816. doi: 10.4049/jimmunol.1700515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Garvey EP, Oplinger JA, Furfine ES, Kiff RJ, Laszlo F, Whittle BJ, Knowles RG. 1400W is a, tight binding, and highly selective inhibitor of inducible nitric-oxide synthase in vitro and in vivo. J Biol Chem. 1997;272:4959–4963. doi: 10.1074/jbc.272.8.4959. [DOI] [PubMed] [Google Scholar]
- 4.Grossniklaus HE, Kang SJ, Berglin L. Animal models of choroidal and retinal neovascularization. Prog Retin Eye Res. 2010;29:500–519. doi: 10.1016/j.preteyeres.2010.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hellwig-Burgel T, Stiehl DP, Wagner AE, Metzen E, Jelkmann W. Review: hypoxia-inducible factor-1 (HIF-1): a novel transcription factor in immune reactions. J Interferon Cytokine Res. 2005;25:297–310. doi: 10.1089/jir.2005.25.297. [DOI] [PubMed] [Google Scholar]
- 6.Hurst J, Kuehn S, Jashari A, Tsai T, Bartz-Schmidt KU, Schnichels S, Joachim SC. A novel porcine ex vivo retina culture model for oxidative stress induced by H(2)O(2) Altern Lab Anim. 2017;45:11–25. doi: 10.1177/026119291704500105. [DOI] [PubMed] [Google Scholar]
- 7.Hurst J, Mueller-Buehl AM, Hofmann L, Kuehn S, Herms F, Schnichels S, Joachim SC. iNOS-inhibitor driven neuroprotection in a porcine retina organ culture model. J Cell Mol Med. 2020;24:4312–4323. doi: 10.1111/jcmm.15091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Joachim SC, Müller-Bühl AM, Hurst J, Theiss C, Hofmann L, Peters L, Herms F, Kuehn S, Dick HB, Schnichels S. Reduced apoptosis and autophagy in an oxidative stress retina organ culture model through an iNOS-inhibitor. Invest Ophthalmol Vis Sci. 2020;61:2473–2473. [Google Scholar]
- 9.Kuehn S, Hurst J, Rensinghoff F, Tsai T, Grauthoff S, Satgunarajah Y, Dick HB, Schnichels S, Joachim SC. Degenerative effects of cobalt-chloride treatment on neurons and microglia in a porcine retina organ culture model. Exp Eye Res. 2017;155:107–120. doi: 10.1016/j.exer.2017.01.003. [DOI] [PubMed] [Google Scholar]
- 10.LaManna JC, Chavez JC, Pichiule P. Structural and functional adaptation to hypoxia in the rat brain. J Exp Biol. 2004;207:3163–3169. doi: 10.1242/jeb.00976. [DOI] [PubMed] [Google Scholar]
- 11.Maliha AM, Kuehn S, Hurst J, Herms F, Fehr M, Bartz-Schmidt KU, Dick HB, Joachim SC, Schnichels S. Diminished apoptosis in hypoxic porcine retina explant cultures through hypothermia. Sci Rep. 2019;9:4898. doi: 10.1038/s41598-019-41113-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Parmentier S, Böhme GA, Lerouet D, Damour D, Stutzmann JM, Margaill I, Plotkine M. Selective inhibition of inducible nitric oxide synthase prevents ischaemic brain injury. Br J Pharmacol. 1999;127:546–552. doi: 10.1038/sj.bjp.0702549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Pang IH, Clark AF. Inducible rodent models of glaucoma. Prog Retin Eye Res. 2020;75:100799. doi: 10.1016/j.preteyeres.2019.100799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sennlaub F, Courtois Y, Goureau O. Inducible nitric oxide synthase mediates retinal apoptosis in ischemic proliferative retinopathy. J Neurosci. 2002;22:3987–3993. doi: 10.1523/JNEUROSCI.22-10-03987.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zhu Y, Nikolic D, Van Breemen RB, Silverman RB. Mechanism of inactivation of inducible nitric oxide synthase by amidines. Irreversible enzyme inactivation without inactivator modification. J Am Chem Soc. 2005;127:858–868. doi: 10.1021/ja0445645. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.