

Abstract
Background:
The human paracentral lobule (PCL) is the medial continuation of the precentral and postcentral gyri. It has important functional area related to the lower limb and perineum. Its visible surface that corresponds to magnetic resonance imaging scout images varies in morphology, so it requires exact data. Studies related to such data are rare. With such a facile, we studied the morphology and morphometry of PCL.
Materials and Methods:
Fifty formalin-fixed adult human brains dissected in the midsagittal plane were used in this study. First, the morphological types of PCL and its boundary were determined, followed by morphometry of its extrasulcal surface using digital vernier calipers. Measurements were done along the anteroposterior axis (length) and vertical axis (height). In addition to that, the extent of motor and sensory area into PCL was also measured.
Results:
Three distinct morphological types of PCL were found: continuous (2%), partially segmented (91%), and completely segmented type (7%). In completely segmented type, a short transitional lobulolimbic gyrus was also found in three cases. The mean extrasulcal surface of the left PCL was significantly larger, both in males (left 10.67 cm2 vs. right 8.80 cm2) and in females (left 8.80 cm2 vs. right 6.99 cm2). Irrespective of gender and sidedness, motor area was significantly larger than the sensory area.
Conclusion:
Reported data will be useful in diagnosis and treatment of diseases affecting the human PCL. Variations in the distribution of sensorimotor cortex over PCL may help further assessment of hemispheric lateralization and the location of central sulcus as a reliable indicator of cytoarchitectonic borders.
Keywords: Cerebral hemisphere, morphology, morphometry, paracentral lobule
Introduction
Paracentral lobule (PCL) is the continuation of the precentral and postcentral gyri of the superolateral surface into the medial surface of cerebral hemisphere. The PCL controls motor and sensory innervations of the contralateral lower extremity. It is also responsible for cortical control of micturition and defecation. It surrounds the indentation of the central sulcus on its upper border, which divides this lobe in the anterior and posterior regions, respectively. The anterior two-thirds of the PCL (medial surface of the precentral gyrus) belong to Brodmann area 4 (BA4).[1] The thick and agranular cortex of BA4 reflects the primary motor cortex in the PCL for the muscles of contralateral leg, foot,[2] and perineum, making this area functionally important in control of bladder and anal sphincters.[3] BA4 is thus called the cortical center of defecation and micturition. According to Grafton et al.,[4] the leg area of the primary motor cortex may extend more caudal from the central sulcus toward the marginal ramus of cingulate sulcus. The posterior third of the PCL (medial surface of the postcentral gyrus) contains BA3, BA2, and BA1, and inferior to these is the part of BA5.[1] In the PCL, the granular cortex of BA3, BA2, and BA1 reflects the primary somatosensory representation of the leg and foot.[2]
The PCL surrounds the indentation of the central sulcus on its upper border. Central sulcus is a topographic landmark on the superolateral surface of brain, but it also acts as a visible cytoarchitectonic border to divide this lobule into anterior (motor) and posterior (sensory) regions. There is a viewpoint that morphological boundaries of cytoarchitectonic areas do not correspond to brain sulci and gyri.[1] The portion of the sensorimotor cortex with the most interindividual variability is the extension of areas BA4 and BA3 onto the mesial face of the hemisphere in the PCL.[5,6] We have investigated the morphology of this region of the brain in detail to allow an assessment of lateral symmetry in the human primary sensorimotor system.[7] Despite several neuroanatomical studies of the PCL over the last century, we found it necessary to begin by characterizing the gross morphology of this part of the human brain. The classical studies of Cunningham[8] and Campbell[9] have provided detailed descriptions of the PCL; however, the paucity of illustration or quantification in these reports limits their usefulness.[7] In particular, we wished to identify any homologous feature in the two hemispheres that might serve as a basis for quantitative comparison of corresponding segments of the left and right PCL. The need for more complete knowledge of the structure of the pre- and post-central gyri, and a better appreciation of the range of their variation in the human brain, is also necessary to interpret a spate of functional imaging studies of the somatotopic organization of the sensorimotor representations along the central sulcus.[10,11] Thus, our primary objective was to define the morphological variations of PCL, followed by standardization of its boundaries to measure the visible (extrasulcal) surface, and to estimate potential difference in sex and laterality.
The secondary objective was to measure the extent of motor (area 4) and sensory (area 3) in the PCL and to estimate the presence of potential difference in sex and laterality.
Materials and Methods
Hundred midsagittal sections obtained from 50 (38 males and 12 females) formalin-fixed cadaveric brains were used in this study. Subjects were aged between 45 and 65 years, with a mean age of 55.84 ± 7.74 years. Cadavers with a history of intracranial lesions, head injury, recorded brain pathology, and visible brain abnormalities were excluded from the study. The method used as follows: In removing the brain, the calvarium was exposed and the skull cap was cut away with a circumferential incision. Since the thickness of the skull varies from one area to another, caution must be taken. The depth of the cut must be regulated by hand to prevent damage to the brain. After removal of skull cap, the dura mater was carefully lifted from the cerebral hemispheres, and using a pointed scissors, it was cut along the line of incision. Finally, the whole brain and remaining spinal cord were gently lifted out of the cranium and placed in brain box.
Immediately after the removal from the cranial vault, they were fixed with basilar suspension in 10% formalin solution for 4 weeks.[12] To carry out morphometric study, brain should be fixed in a satisfactory manner so that the specimen exhibits a natural form after fixation and all the anatomic components fixed completely and uniformly. There should be no distortion of natural shape and border of fixed section. In dissection study, formalin is universally used for fixing tissues. However, formalin embalming process is regarded to cause brain volume shrinkage.[13] Hence, cautions are required for clinical application of quantitatively measured data in embalmed brain. To compensate the defect of this study, we have used 10% formolin solution for 4 weeks as it may be considered as a standardized method of fixation. Thereafter, brains were dissected by using the midsagittal line as an anatomical landmark. The brains were carefully sectioned with brain knife in the midsagittal plane passing through the body of CC, the interhemispheric fissure, septum pellucidum, cavity of the third ventricle, and cerebral aqueduct. Meninges were carefully removed from the medial surface of midsagittal section to be studied.
Morphology of the paracentral lobule
gross examination of the extrasulcal surface of PCL was done to determine its various types. We divided PCL into three morphological types:
Continuous: Precentral and postcentral gyri were in continuity throughout the whole length
Partially segmented: Constituent gyri were in continuity in most of the part but in the lower part separated by the upturned branch of cingulate sulcus
Totally segmented: Constituent gyri were completely separated by the upturned branch of cingulate sulcus.
Determination of paracentral lobule boundaries
Conventional systems of line are often used to delineate different areas of brain. In the present study, we have used the intercommissural line to determine the boundaries of PCL and measure the extrasulcal surface in vertical (Y-axis) and sagittal plane (X-axis). It was defined as a line connecting the most prominent superior point of the anterior commissure (AC) and the most prominent inferior point of the posterior commissure (PC)[14] in the sagittal plane (X-axis), while the Y-axis was defined as the vertical line drawn perpendicular to the AC–PC line extending from the medial end of the central sulcus to the cingulate sulcus. Marginal ramus of the cingulate sulcus forms the posterior boundary of PCL and constant in location. Cingulate sulcus forms the inferior boundary in majority of cases, except in completely segmented PCL. In such cases, inferior boundary was defined by the horizontal Xa line (running above and parallel to the AC–PC line), which connects the two segments of the cingulate sulcus across the region of its missing part between the PCL above and the cingulate gyrus below. Due to the absence of clearly visible anatomic landmark for anterior boundary, anterior boundary was standardized by using the intercommissural or AC–PC line[14] [Figure 1]. Using this means of standardization, the anterior boundary of the PCL was defined as the vertical line (Ya) lying anterior and parallel to Y-axis, extending from the medial end of the precentral sulcus to the cingulate sulcus. The surface located posterior to the Ya line and anterior to the marginal ramus of cingulate sulcus was referred to as the PCL. In a few cases, when the precentral sulcus did not reach the medial surface, we extended its direction by an imaginary straight line that reached the superior border of the hemisphere and then used the corresponding vertical Ya line to the cingulate sulcus. The superior boundary of the PCL was the superior border of the hemisphere.
Figure 1.
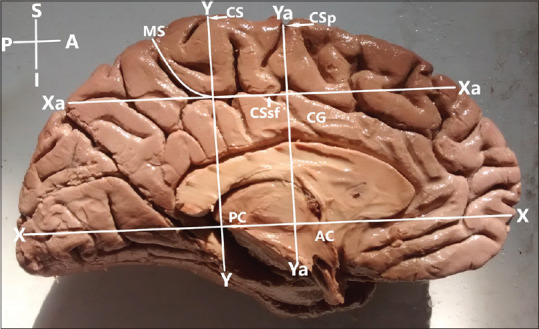
Boundaries of the paracentral lobule in respect to the intercommissural or anterior commissure–posterior commissure line; AC – Anterior commissure; PC – Posterior commissure; X axis – Anterior commissure–posterior commissure line; Xa – Line defines inferior boundary of the paracentral lobule in case of segmented paracentral lobule; Y axis-vertical line intersecting the anterior commissure–posterior commissure line; Ya – Line that defines the anterior boundary of the paracentral lobule; CS – Medial extension of central sulcus; CSp – Medial extension of precentral sulcus; CSsf – Cingulate sulcus (subfrontal part); MS – Cingulate sulcus (marginal part); CG – Cingulate gyrus; Right hemisphere. Orientation: A – Anterior; P – Posterior; S – Superior; I – Inferior
Morphometry of its extrasulcal surface
Measurements of the PCL were done along the anteroposterior axis (length) and vertical axis (height). Using these measurements, the surface area of PCL was thus calculated (length × height). In addition to that, surface area of the motor PCL (area between central and precentral sulcus) and sensory PCL (area between central and marginal branch of the cingulate sulcus) was also measured.
All the measurements were taken using digital vernier calipers (Mitutoyo, Japan) with least count of 0.01 mm. The measurements were repeated two times, and the mean was taken as final for further analysis. Results were expressed as mean ± standard deviation.
Statistical analysis
Statistical analysis was performed using descriptive statistics. The measured parameters of the PCL were compared between hemispheres (interhemispheric differences) and genders (sex differences) using a t-test and P < 0.05 was considered statistically significant.
Results
We found three morphological types of PCL: continuous, partially segmented, and completely segmented. The continuous type of PCL was of rarest variety and found only in two cases (2%) of the investigated brains [Figure 2]. The partially segmented type of PCL was most predominant [Figure 3] and found in 91 cases (91%) of hemispheres. Completely segmented type was found in 7% of cases. Three hemispheres with completely segmented PCL showed the presence of an additional lobulolimbic transitional gyrus (LLTG) interposed between the posterior PCL and cingulate gyrus [Figure 4].
Figure 2.
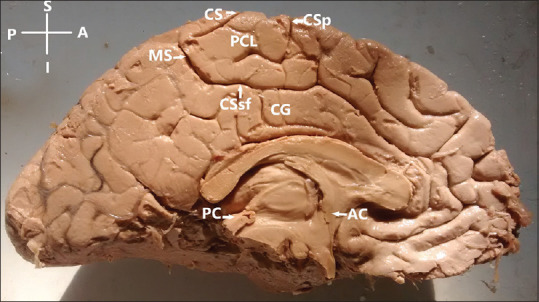
Paracentral lobule-continuous type (precentral and postcentral gyri are in continuity throughout the whole length, found in 2% of cases); AC – Anterior commissure; PC – Posterior commissure; CS – Medial extension of central sulcus; CSp – Medial extension of precentral sulcus; CSsf – Cingulate sulcus (subfrontal part); MS – Cingulate sulcus (marginal part); CG – Cingulate gyrus; Left hemisphere. Orientation: A – Anterior; P – Posterior; S – Superior; I – Inferior
Figure 3.
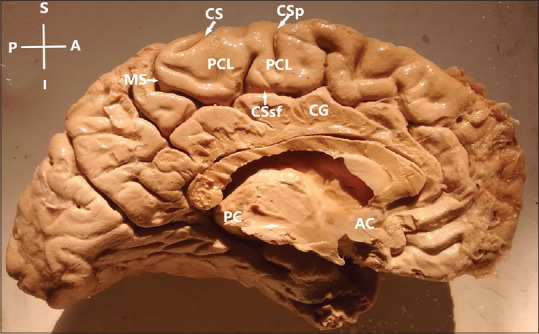
Paracentral lobule-partially segmented type (constituent gyri were in continuity in most of the part but in the lower part separated by the upturned branch of cingulate sulcus, found in 91% of cases); AC – Anterior commissure; PC – Posterior commissure; CS – Medial extension of central sulcus; CSp – Medial extension of precentral sulcus; CSsf – Cingulate sulcus (subfrontal part); MS – Cingulate sulcus (marginal part). CG – Cingulate gyrus; Left hemisphere. Orientation: A – Anterior; P – Posterior; S – Superior; I – Inferior
Figure 4.
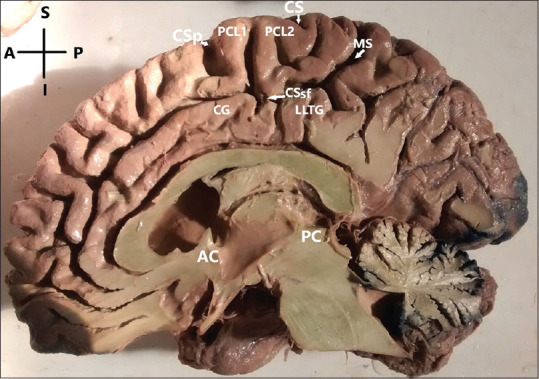
Paracentral lobule-completely segmented type (constituent gyri were completely separated by upturned branch of cingulate sulcus);segmented type (segmented into anterior-PCL1, posterior-PCL2) and lobulo-limbic transitional gyrus: interposed between the PCL2 and cingulate gyrus; AC – Anterior commissure; PC – Posterior commissure; CS – Medial extension of central sulcus; CSp – Medial extension of precentral sulcus; CSsf – Cingulate sulcus (subfrontal part); MS – Cingulate sulcus (marginal part). CG – Cingulate gyrus; Right hemisphere. Orientation: A – Anterior; P – Posterior; S – Superior; I – Inferior
Table 1 depicts the morphometric data of the extrasulcal surface of the PCL, with noted differences related to the side of the hemisphere and sex. In the total sample, the mean surface area of the extrasulcal cortex of the left PCL (10.22 cm2) was significantly larger (P < 0.05) than the extrasulcal surface of the right PCL (8.36 cm2). In males, the average surface of the left PCL extrasulcal cortex (10.67 cm2) was significantly larger (P < 0.05) in relation to the right PCL (8.80 cm2). In females, the average surface of the left PCL extrasulcal cortex (8.80 cm2) was larger than the right PCL (6.99 cm2), and this difference was also statistically significant (P < 0.05). Comparison of the extrasulcal cortical surface areas of the PCL between males and females (separately the left with the left, and the right with the right) showed statistically significant sex differences (P < 0.05). Table 2 depicts the morphometric data of sensorimotor area of PCL with noted differences in relation to side of the hemisphere and sex. The mean value of surface area of motor PCL was significantly larger than the sensory in all the specimens irrespective of sex and sidedness.
Table 1.
The morphometric data of the extrasulcal surface of the paracentral lobule, with noted differences related to the side of the hemisphere in each gender
| Statistical parameters | Males (n=38) | Female (n=12) | ||
|---|---|---|---|---|
| Right | Left | Right | Left | |
| Mean (cm2) | 8.80 | 10.68 | 6.99 | 8.80 |
| Range | 8.00-9.50 | 10.23-11.50 | 6.75-7.30 | 8.50-9.32 |
| SD (cm2) | 0.40 | 0.34 | 0.18 | 0.26 |
| CV (%) | 4.50 | 3.18 | 2.57 | 2.95 |
| P | 0.000 | 0.000 | ||
SD - Standard deviation; CV - Coefficient of variation
Table 2.
The morphometric data of the sensorimotor area of the paracentral lobule, with noted differences related to the side of the hemisphere in each gender
| Statistical parameters | Males (n=38) | Female (n=12) | ||||||
|---|---|---|---|---|---|---|---|---|
| Right | Left | Right | Left | |||||
| Sensory area | Motor area | Sensory area | Motor area | Sensory area | Motor area | Sensory area | Motor area | |
| Mean (cm2) | 4.08 | 4.55 | 4.72 | 6.12 | 3.19 | 3.97 | 3.79 | 4.83 |
| Range | 3.25-4.50 | 4.00-5.26 | 4.20-5.50 | 5.26-6.52 | 3.00-3.50 | 3.50-4.15 | 3.50-4.16 | 4.40-5.20 |
| SD (cm2) | 0.22 | 0.40 | 0.36 | 0.27 | 0.15 | 0.21 | 0.24 | 0.24 |
| CV (%) | 5.39 | 8.79 | 7.62 | 4.41 | 4.70 | 5.29 | 6.33 | 4.97 |
| P | 0.000 | 0.000 | 0.000 | 0.000 | ||||
SD - Standard deviation; CV - Coefficient of variation
Discussion
In the present study, we have investigated the PCL in 100 hemispheres obtained from 50 human brains. Our purpose in the present work was twofold. We wished to examine its gross morphological structure and variations. In addition, we wished to characterize the visible boundaries of PCL to measure its extrasulcal cortical surface area. Subsequently, we have taken measurements of the extent of area 4 and 3 into the medial surface, to evaluate the degree of lateral symmetry in the human sensorimotor system in PCL.[7]
In the present study, three distinct morphological types of the human PCL were found. Partially segmented PCL was found predominantly (91%), followed by completely segmented (7%) and continuous type (2%). Three hemispheres with segmented cingulated sulcus showed the presence of the lobulolimbic gyri between the PCL proper and supratruncal region of the cingulate gyrus.
Spasojević et al.[15] studied the morphology of PCL on 84 hemispheres and found two distinct morphological types of the human PCL: continuous type, which was predominant (95.2%), and rare segmented type (4.8%). In hemispheres with segmented cingulate sulcus, they also found the short transitional lobulolimbic gyrus (13.1%).
The difference in the morphological findings is actually due to the fact that Spasojevic et al. have considered both partially segmented and continuous type in a single category of continuous type, whereas in the present study, we have taken them as separate entity as we found them morphologically distinct.
According to Fischl et al.,[16] the predominance of continuous morphological type confirms that primary and secondary areas exhibit less variability than higher orders cortical areas.
Stanczyk[17] studied the variation of the gyrus and sulcus cinguli. They reported short LLTG interposed between PCL and segmented cingulate sulcus in 37% of cases, whereas Spasojevic et al.[15] found LLTG in 13.1%. In our specimens, it was found in 3% of cases only. These variations in PCL morphology may be due to difference in race and ethnicity of the specimens studied.
Our morphological observations add to these early descriptions[15,16] by illustrating and documenting the features of the PCL in sagittal plane of cerebral hemisphere. In particular, the ability to view this region in the axial plane has facilitated comparison of our morphological data with neurosurgical and radiological studies of the somatotopic organization of the human sensorimotor cortex in medial surface of brain. We believe that the findings about the role of the PCL in micturition control[3] should be considered in the context of its morphological variability, close relationships to the medial frontal and anterior cingulated cortex, and absence of its clear anterior boundaries. The presence of completely segmented PCL, which we found only in 7% of hemispheres, correlates with the report that hemisphere with interruption and duplication of the cingulate sulcus had increased amount of blood flow in the sensorimotor area located more rostrally.[4]
We have measured the area of extrasulcal surface of PCL as this area corresponds to the magnetic resonance imaging (MRI) scout images. Due to the absence of a clearly visible anterior boundary of the PCL, we standardized it using the intercommissural line.[14] Conventional lines are often used by previous researchers in delineation of the brain lobes also, including the boundary between the frontal and parietal lobes passing just through the PCL.
The AC–PC or similar exact systems of lines for delineation of the PCL or other brain areas are often used[6,18,19] to increase precision of data.
In a functional MRI study, Deskin et al.[20] delineated the PCL as the structure located posterior to the medial (superior) frontal gyrus (or the precentral gyrus when visible), anterior to the precuneus (or the postcentral gyri when visible), superior to the cingulate gyrus, and inferior to the superior hemispheric border.
PCL includes portions of the frontal and parietal lobes: while the boundary between the lobes, the central sulcus, is easy to locate on the lateral surface of the cerebral hemispheres, this boundary is often discerned in a cytoarchitectonic manner in cases where the central sulcus is not visible on the medial surface.[20]
The extrasulcal surface of PCL is clearly visible on routine MRI scout images, so our morphometric data can be used for delineation of the PCL and its further studies. Our finding showed significantly larger extrasulcal surface of the left PCL (10.22 cm2) compared to right PCL (8.36 cm2), which was in consonance with the findings of Spasojević et al.[15] This may be due to the fact that majority of the population (90%–95%) is predominantly right handed, and in them, left hemisphere acts as dominant hemisphere for many important functions.[21] The region of the left PCL showed greater activation during the knee task compared with the ankle tasks[22] and also when subjects performed saccadic eye movements.[18,19]
We also found statistically significant differences in the extrasulcal surface area of the PCL between males and females, but Spasojević et al.[15] did not find statistically significant sex differences (P > 0.05). Morphological and morphometric gender differences have various aspects and are more complex than usually expected.[23,24]
Subsequently, we measured the area 4 and 3 to evaluate the degree of lateral symmetry in the human sensorimotor system in PCL.[7,15]
In the current study, the extent of the motor area (BA4) was significantly greater than the sensory area (BA3) in all the hemispheres, irrespective sex and side.
Oliveira Neto et al.[25] measured the length of motor and sensory area of PCL on 42 hemispheres in the sagittal and coronal axis and found that the length of motor area was larger than the sensory area, but the difference was not significant unlike our study. Significant statistical correlations between these measures could be inferred through studies with larger sample correlating variables such as age, body weight, height, race, and education level to the length of the measures analyzed in this study.
White et al.[7] studied the cytoarchitecture of primary sensorimotor cortex on 40 cerebral hemispheres and found that the cytoarchitectonic borders between BA4 and BA3 in the PCL showed considerable interindividual and interhemispheric variations in the way that a significant portion of BA4 extends into the PCL in comparison to a very small portion of BA3. Although the positions of the cytoarchitectonic boundaries in the PCL showed considerable interindividual variation, the locations of the borders of areas 4 and 3 along the course of the sulcus were similar among the 40 hemispheres examined. These observations provide a basis for evaluating lateral symmetry of the human primary sensorimotor cortex in the PCL.
Rademacher et al.[5] also found variation in the distribution of sensorimotor cortex in the PCL and concluded that the visible topographic landmarks of the central sulcus provide a reliable indication of the cytoarchitectonic borders.
Thus, we confirm and extend the important observations of previous authors[6,7,24] concerning the variation in the distribution of areas 4 (BA4) and 3 (BA3) in the PCL.
The central portion of the PCL occupies the medial portion of BA4 (primary somatomotor cortex) without limiting the sulcal landmark and without consistent association between the individual sulci and cytoarchitectonic borders.[6]
Limitations
Since formalin fixation causes some amount of brain shrinkage, the measured parameters may deviate to some extent than the actual value. Moreover, MRI revelation of the measurements can be more helpful.
Conclusion
We can conclude that the extrasulcal (visible) cortical surface of the left PCL was significantly larger than right in both the sexes, and in males, the value was significantly larger than female. Reported data will be useful in diagnosis and treatment of diseases affecting the human PCL. Variations in the distribution of sensorimotor cortex over PCL may help further assessment of hemispheric lateralization and the location of central sulcus as a reliable indicator of cytoarchitectonic borders. These morphological observations extend the classical anatomical studies on the human PCL and provide a basis for evaluating lateral symmetry of the human primary sensorimotor cortex in the mesial surface.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest.
References
- 1.Brodmann K. Comparative localization theory of the cerebral cortex. Leipzig: Verlag JA Barth; 1925. pp. 43–69. [Google Scholar]
- 2.Sakakibara R, Fowler CJ, Hattori T. Voiding and MRI analysis of the brain. Int Urogynecol J Pelvic Floor Dysfunct. 1999;10:192–9. doi: 10.1007/s001920050044. [DOI] [PubMed] [Google Scholar]
- 3.Fowler CJ, Griffiths D, de Groat WC. The neural control of micturition. Nat Rev Neurosci. 2008;9:453–66. doi: 10.1038/nrn2401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Grafton ST, Fagg AH, Woods RP, Arbib MA. Functional anatomy of pointing and grasping in humans. Cereb Cortex. 1996;6:226–37. doi: 10.1093/cercor/6.2.226. [DOI] [PubMed] [Google Scholar]
- 5.Rademacher J, Bürgel U, Geyer S, Schormann T, Schleicher A, Freund HJ, et al. Variability and asymmetry in the human precentral motor system. A cytoarchitectonic and myeloarchitectonic brain mapping study. Brain. 2001;124:2232–58. doi: 10.1093/brain/124.11.2232. [DOI] [PubMed] [Google Scholar]
- 6.White LE, Andrews TJ, Hulette C, Richards A, Groelle M, Paydarfar J, et al. Structure of the human sensorimotor system. I: Morphology and cytoarchitecture of the central sulcus. Cereb Cortex. 1997;7:18–30. doi: 10.1093/cercor/7.1.18. [DOI] [PubMed] [Google Scholar]
- 7.White LE, Andrews TJ, Hulette C, Richards A, Groelle M, Paydarfar J, et al. Structure of the human sensorimotor system. II: Lateral symmetry. Cereb Cortex. 1997;7:31–47. doi: 10.1093/cercor/7.1.31. [DOI] [PubMed] [Google Scholar]
- 8.Cunningham DJ. Surface Anatomy of the Cerebral Hemispheres. Dublin: Academy House; 1892. [Google Scholar]
- 9.Campbell AW. Histological Studies on the Localisation of Cerebral Function. New York: Cambridge University Press; 1905. [Google Scholar]
- 10.Rumeau C, Tzourio N, Murayama N, Peretti-Viton P, Levrier O, Joliot M, et al. Location of hand function in the sensorimotor cortex: MR and functional correlation. AJNR Am J Neuroradiol. 1994;15:567–72. [PMC free article] [PubMed] [Google Scholar]
- 11.Sanes JN, Donoghue JP, Thangaraj V, Edelman RR, Warach S. Shared neural substrates controlling hand movements in human motor cortex. Science. 1995;268:1775–7. doi: 10.1126/science.7792606. [DOI] [PubMed] [Google Scholar]
- 12.Anagnostopoulou S, Mourgel S, Katritsi D. Morphometry of corpus callosum: An anatomical study. Neuroanat. 2006;5:20–3. [Google Scholar]
- 13.Schulz G, Crooijmans HJ, Germann M, Scheffler K, Müller-Gerbl M, Müller B. Three-dimensional strain fields in human brain resulting from formalin fixation. J Neurosci Methods. 2011;202:17–27. doi: 10.1016/j.jneumeth.2011.08.031. [DOI] [PubMed] [Google Scholar]
- 14.Talairach J, Tournoux P. Co-Planar Stereotaxic Atlas of the Human Brain. New York: Thieme Medical Publishers; 1988. [Google Scholar]
- 15.Spasojević G, Malobabic S, Pilipović-Spasojević O, Djukić-Macut N, Maliković A. Morphology and digitally aided morphometry of the human paracentral lobule. Folia Morphol (Warsz) 2013;72:10–6. doi: 10.5603/fm.2013.0002. [DOI] [PubMed] [Google Scholar]
- 16.Fischl B, Rajendran N, Busa E, Augustinack J, Hinds OB, Yeo T, et al. Cortical folding patterns and predicting cytoarchitecture. Cereb Cortex. 2008;18:1973–80. doi: 10.1093/cercor/bhm225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Stanczyk LJ. Variation of the gyrus and sulcus cinguli in phylo and ontogenesis. Folia Morphol. 1983;42:243–59. [PubMed] [Google Scholar]
- 18.Grosbras MH, Lobel E, van de Moortele PF, Le Bihan D, Berthoz A. An anatomical landmark for the supplementary eye field in human revealed with functional magnetic resonance imaging. Cereb Cortex. 1999;9:705–711. doi: 10.1093/cercor/9.7.705. [DOI] [PubMed] [Google Scholar]
- 19.Hamani C, Mayberg H, Snyder B, Giacobbe P, Kennedy S, Lozano AM. Deep brain stimulation of the subcallosal cingulate gyrus for depression: Anatomical location of active contacts in clinical responders and a suggested guideline for targeting. J Neurosurg. 2009;111:1209–15. doi: 10.3171/2008.10.JNS08763. [DOI] [PubMed] [Google Scholar]
- 20.Desikan SR, Segonne F, Fischl B, Quinn TB, Dickerson CB, Blacker D, et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuro Image. 2006;31:968–80. doi: 10.1016/j.neuroimage.2006.01.021. [DOI] [PubMed] [Google Scholar]
- 21.Conn PM. Neuroscience in Medicine. Totowa, New Jersey: Humana Press; 2003. p. 348. [Google Scholar]
- 22.Witelson SF, Kigar DL. Asymmetry in brain function follows asymmetry in anatomical form: Gross, microscopic, postmortem and imaging studies. In: Boller F, Grafman J, editors. Handbook of Neuropsychology. Ch. 6. Vol. 1. New York: Elsevier; 1998. pp. 111–42. [Google Scholar]
- 23.Newton MJ, Dong Y, Hidler J, Plummer D, 'Amato P, Marehbian J, Albistegui-DuBois MR, et al. Reliable assessment of lower limb motor representation with fMRI: Use of a novel MR compatible device for real-time monitoring of ankle, knee and hip torques. Neuroimage. 2008;43:36–146. doi: 10.1016/j.neuroimage.2008.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Spasojević G, Malobabić S, Suscević D, Miljković Z. Morphometric variability of precuneus in relation to gender and the hemisphere of human brain. Vojnosanit Pregl. 2004;61:365–70. doi: 10.2298/vsp0404365s. [DOI] [PubMed] [Google Scholar]
- 25.Oliveira Neto M, Nascimento PA, Rodrigues RB, Machado WB, Oliveira LS, Silva Filho AR. Morphometric analysis of the paracentral lobe. J Health Biol Sci. 2014;2:19–21. [Google Scholar]