

Abstract
Obesity is a serious global health problem because of its increasing prevalence and comorbidities, but its treatments are limited. The serotonin 2C receptor (5-HT2CR), a G-protein-coupled receptor, activates proopiomelanocortin (POMC) neurons in the arcuate nucleus of hypothalamus (ARH) to reduce appetite and weight gain. However, several 5-HT analogs targeting this receptor, e.g., lorcaserin (Lor), suffer from diminished efficacy to reduce weight after prolonged administration. Here, we show that barbadin (Bar), a novel β-arrestin/β2-adaptin inhibitor, can prevent 5-HT2CR internalization in cells and potentiate long-term effects of Lor to reduce appetite and body weight in male mice. Mechanistically, we demonstrate that Bar co-treatment can effectively maintain the sensitivity of the 5-HT2CR in POMCARH neurons, despite prolonged Lor exposure, thereby allowing these neurons to be activated through opening the transient receptor potential cation (TRPC) channels. Thus, our results prove the concept that inhibition of 5-HT2CR desensitization can be a valid strategy to improve the long-term weight loss effects of Lor or other 5-HT2CR agonists, and also provide an intellectual framework to develop effective long-term management of weight by targeting 5-HT2CR desensitization.
SIGNIFICANCE STATEMENT By demonstrating that the combination of barbadin (Bar) with a G-protein-coupled receptor (GPCR) agonist can provide prolonged weight-lowering benefits in a preclinical setting, our work should call for additional efforts to validate Bar as a safe and effective medicine or to use Bar as a lead compound to develop more suitable compounds for obesity treatment. These results prove the concept that inhibition of serotonin 2C receptor (5-HT2CR) desensitization can be a valid strategy to improve the long-term weight loss effects of lorcaserin (Lor) or other 5-HT2CR agonists. Since GPCRs represent a major category as therapeutic targets for various human diseases and desensitization of GPCRs is a common issue, our work may provide a conceptual framework to enhance effects of a broad range of GPCR medicines.
Keywords: 5-HT 2C receptor, food intake, GPCR, hypothalamus, obesity
Introduction
The global prevalence of obesity has increased substantially in the past decades. By 2016, >39% of adults aged 18 years or older were overweight, and 13% were obese. Over 340 million children and adolescents aged 5–19 were overweight or obese worldwide (World Health Organization, 2016). Obesity dramatically increases the risk of cardiometabolic diseases, dementia, depression and even some cancers, thus leading to a decline in life quality and life expectancy (Fontaine et al., 2003; Blüher, 2019). Although obesity is classified as a disease (Kyle et al., 2016), the first option for treating obesity is mainly by intervening lifestyle, which, however, contributes to limited and unabiding outcomes (Diabetes Prevention Program Research Group et al., 2009; Stefan et al., 2018). Therefore, pharmacotherapy is needed to be an adjunctive or even a dominant treatment option.
The serotonin 2C receptor (5-HT2CR) is a Gq-protein-coupled receptor (GPCR) that plays a critical role in regulating energy balance (Heisler et al., 2003). Mice with global 5-HT2CR deficiency develop hyperphagia and obesity (Tecott et al., 1995; Nonogaki et al., 1998). 5-HT2CR's effects on appetite and body weight are largely mediated by proopiomelanocortin (POMC) neurons within the arcuate nucleus of hypothalamus (ARH; Heisler et al., 2002, 2003; Qiu et al., 2007). Notably, re-expressing 5-HT2CRs only in POMC neurons completely normalizes the hyperphagia and obesity in 5-HT2CRs null mice (Xu et al., 2008). Meta-chlorophenylpiperazine, an agonist of 5-HT2CRs, activates POMCARH neurons and induces hypophagia (Vickers et al., 2003; Sohn et al., 2011). Conversely, compounds that suppress brain 5-HT signals produce hyperphagia and weight gain (Ghosh and Parvathy, 1973; Blundell and Leshem, 1974; Geyer et al., 1976; Saller and Stricker, 1976). These previous findings indicate that selective activation of 5-HT2CRs has therapeutic potential for obesity treatment.
In past decades, several weight management drugs targeting the 5-HT2CR-mediated pathway have been most clinically effective for obesity treatment. These include d-fenfluramine, which stimulates the release of 5-HT and inhibits its reuptake into nerve terminals (Vickers et al., 1999), and sibutramine, which inhibits 5-HT and noradrenaline reuptake (Luque and Rey, 1999). However, these two drugs were withdrawn from the clinical use because of their non-specific adverse effects on the cardiovascular system (Connolly et al., 1997; Scheen, 2010). Lorcaserin (Lor), a highly selective agonist of the 5-HT2CR with a minimal risk of causing cardiovascular side effects (Weissman et al., 2013), was approved by the United States Food and Drug Administration (FDA) as an anti-obesity medicine in 2012. While Lor's short-term effects are quite robust, its long-term weight loss efficacy is less than satisfactory. Patients with prolonged Lor treatment show no continuous weight loss but a body weight regain (∼50% of maximum weight loss; Smith et al., 2010). More recently, Lor is reported to be potentially associated with a higher risk of cancers, although this association was not statistically significant (Food and Drug Administration, 2020). However, the FDA withdrew Lor from the United States market because the potential cancer risk outweighs its long-term weight management benefit (Food and Drug Administration, 2020). Putting aside the fact that these drugs were withdrawn because of their unspecific adverse effects, the common issue for these medicines is their modest long-term efficacy to reduce body weight (Douglas et al., 1983; Apfelbaum et al., 1999; James et al., 2010; Smith et al., 2010). Therefore, better understanding the mechanisms by which long-term Lor treatment loses its effects may provide necessary framework to develop strategies to treat obesity with prolonged efficacy.
In the current study, we first demonstrated that barbadin (Bar), a novel β-arrestin/β2-adaptin inhibitor, can prevent 5-HT2CR internalization in cultured cells. We then examined, in mice, the weight-reducing efficacy of a chronic regimen combining Lor and Bar. Then we combined electrophysiology, neuroanatomy, fiber photometry to delineate the actions of Bar on POMCARH neurons' responsiveness to Lor treatment.
Materials and Methods
Mice
All the mice used in the current study were on a C57BL/6J background. For electrophysiological studies, we crossed POMC-CreER (Berglund et al., 2013) with Rosa26-LSL-tdTOMATO allele (Madisen et al., 2010; obtained from The Jackson Laboratory, #007905) to generated POMC-CreER/Rosa26-LSL-tdTOMATO mice. These mice express tdTOMATO selectively in mature POMC neurons after tamoxifen induction (0.2 mg/g, i.p.; at least eight weeks of age). For fiber photometry studies, we crossed C57BL/6J mice with POMC-Cre mice (Balthasar et al., 2004). The wild-type littermates from all above mating were used for metabolic phenotype studies. The mice were fed standard chow diet (6.5% fat, #2920, Harlan-Teklad) or with a high-fat diet (HFD; 60% fat, #D12492i, Research Diets). Water was provided ad libitum.
Plasmid
5-HT2CR was cloned from mouse hypothalamus using the forward primer 5′-ATGGTGAACCTGGGCACTG-3′ and reverse primer 5′-CACACTACTAATCCTCTCGC-3′. The PCR products was amplified with PrimeSTAR Max DNA Polymerase (R045B, Takara) and cloned into EcoRI-XhoI site of pcDNA3.1/myc-his A. Sequencing confirmed the cloned 5-HT2CR is VNV isoform. Then, the VGV isoform was generated by Q5 Site-Directed Mutagenesis kit (E0554S, New England Biolabs) with forward primer 5′-GCTCAACAGGACCACGTACTGCTACATACC-3′ and reverse primer 5′-ATAGCCGGTTCAATTCGCGGAC-3′ (Watanabe et al., 2014). To generate 5-HT2CR-mCherry expression vector, the 5-HT2cR/VGV isoform was sub-cloned into the mCherry2-N1 vector (#54517, Addgene plasmid) which was gift from Michael Davidson.
Cell transfection and confocal fluorescence microscopy imaging
HEK293 cells were grown in DMEM supplemented with 10% (v/v) fetal bovine serum. Cells were plated on gelatin-coated coverslip and then transiently transfected with 5-HT2CR-VGV-mCherry vector using Lipofectamine 3000 reagent (L3000015, Invitrogen). After 48-h transfection, cells were firstly treated with vehicle (Veh; DMSO) or 10 μm Bar (356568-70-2, Axon Medchem) for 30 min and then Veh (DMSO) or 100 μm Lor (A12598, Adoop bioscience) for another 30 min. Then the transfected cells were fixed with 4% paraformaldehyde in PBS for 15 min. After washing in PBS, cells were incubated with 5 mg/ml wheat germ agglutinin Alexa Fluor 633 conjugate (WGA; W32466, Invitrogen) and 1 mg/ml Hoechst 33342 dye (H3570, Invitrogen) at room temperature for 10 min. Images were captured on Olympus confocal microscope. Fluorescence intensity was quantified using ImageJ, and five positive cells were selected for statistical analysis in each group.
Food intake, body weight, and body composition
In chow feeding study, male C57BL/6J mice were fed with chow ad libitum. At 24 weeks of age, these mice were singly housed one week before measuring the chow intake in home cages. For the daily drug treatment, all mice received first intraperitoneal injection (Veh: saline with 1% DMSO, or Bar; 0.3 mg/kg) at 5 P.M., then received second intraperitoneal injection (Veh: saline, or Lor; 9 mg/kg) at 6 P.M. Chow intake and body weight were monitored every 2 d until the end of study. On day 26, body composition (fat mass and lean mass) was determined by quantitative magnetic resonance.
Another cohort of male C57BL/6J mice were used in the HFD feeding study. Briefly, these mice were singly housed since 12 weeks of age and fed with HFD ad libitum to induce an obese phenotype before the drug treatment. After reaching ∼50 g body weight, all mice received same treatment as in the chow-fed mice. HFD intake and body weight were monitored every 2 d until end of study.
Metabolic cages
Physical activity, energy expenditure, and meal patterns were monitored by using a TSE PhenoMaster system. Experiments were performed in temperature-controlled (23°C) cabinets containing 16 TSE PhenoMaster metabolic cages. After finishing the 26-d food intake and body weight study as described above, the same group of chow-fed mice were transported to the metabolic cages to acclimate to the new environment. After acclimation for 2 d, cages were connected to the TSE phenoMaster system for a total of 4 d. For the daily drug treatment, all mice received first intraperitoneal injection (Veh: saline with 1% DMSO, or Bar; 0.3 mg/kg) at 5 P.M., then received second intraperitoneal injection (Veh: saline, or Lor; 9 mg/kg) at 6 P.M. Data of meal pattern, O2 consumption, CO2 production, heat production and activity were automatically recorded by the system, and data collected from the last 2 d were used for analysis. O2 consumption, CO2 production, and heat production were normalized by the lean mass of each mouse. Data for meal analysis was collected as binary data every 10 s. The start of a meal was defined by food removal equal to or larger than 0.1 g and the meal was ended when no further food removal occurred before the end of the inter-meal interval of 30 min. Meal frequency was defined by the total number of food intake events in 1-d cycle, while meal size was defined by the averaged food intake of meals for each mouse.
Electrophysiology
Chow-fed POMC-CreER/Rosa26-LSL-tdTOMATO mice were used for electrophysiology recordings. Male mice at 10–12 weeks of age received daily intraperitoneal injections of various combination of Veh, Lor, and/or Bar for 20 d (as described above) before recordings. Electrophysiology recordings were performed as previously described (He et al., 2020). Briefly, mice were anesthetized with isoflurane and brains were dissected rapidly and immersed in ice-cold and oxygenated cutting solutions (10 mm NaCl, 195 mm sucrose, 2.5 mm KCl, 1.25 mm NaH2PO4, 7 mm MgCl2, 25 mm NaHCO3, 5 mm glucose, 0.5 mm CaCl2, and 2 mm sodium pyruvate, balanced with 95% O2/5% CO2). Coronal brain slices (220 μm) containing the ARH were cut with a Microm HM 650 V vibratome (Thermo Scientific) in oxygenated cutting solution. Slices were then incubated in oxygenated artificial CSF (ACSF; 126 mm NaCl, 2.5 mm KCl, 2.4 mm CaCl2, 1.2 mm NaH2PO4, 1.2 mm MgCl2, 11.1 mm glucose, and 21.4 mm NaHCO3, balanced with 95% O2/5% CO2, pH 7.4) to recover ∼25 min at 32°C and subsequently for ≧1 h at room temperature before recording.
Slices were then transferred to the recording chamber perfused continuously with 32°C artificial cerebrospinal fluid bubbled with 95% O2/5% CO2 to ensure adequate oxygenation of slices. tdTOMATO (+) neurons were identified by using epifluorescence and IR-DIC imaging on an upright microscope (Eclipse FN-1, Nikon) equipped with a moveable stage (MP-285, Sutter Instrument). Patch pipettes with resistances of 3–5 MΩ were filled with intracellular solution (adjusted to pH 7.3) containing 128 mmol/l K gluconate, 10 mmol/l KCl, 10 mmol/l HEPES, 0.1 mmol/l EGTA, 2 mmol/l MgCl2, 0.3 mmol/l Na-GTP, and 3 mmol/l Mg-ATP. Recordings were made using a MultiClamp 700B amplifier (Axon Instrument), sampled using Digidata 1440A and analyzed offline with pClamp 10.3 software (Axon Instrument). Series resistance was monitored during the recording, and the values were generally <10 MΩ and were not compensated. The liquid junction potential was +12.5 mV, and was corrected after the experiment. Data were excluded if the series resistance increased >20% during the experiment or without overshoot for action potential. Currents were amplified, filtered at 1 kHz, and digitized at 20 kHz. Current clamp was engaged to test neural firing frequency at the baseline and after puff delivery of Lor (5 s at a concentration of 100 μm). Each neuron was recorded for at least 1-min baseline, and only the neurons with stable baseline were used to test Lor puff effect. To ensure each recorded neuron receive same amount of Lor, the neurons located on the surface of the slice were selected to record and the puff pipette was always put at a 100-μm horizontal and 50-μm vertical distance from the recorded neurons. The puff strength was maintained at a same level by using a repeatable pressure pulse system (Picospritzer III, Parker). The values for resting membrane potential (RMP) and firing frequency were averaged within 1 min bin at the baseline or after Lor puff. A neuron was considered depolarized if an increase in RMP was ≥2 mV.
To record transient receptor potential cation (TRPC) current, the membrane potential was hold at −60 mV in the voltage-clamp mode in the same neuron recorded in current clamp mode. A continuous current trace was recorded at the baseline and after puff delivery of Lor (5 s at a concentration of 100 μm). The selective TRPC channel blocker 1-[2-(4-methoxyphenyl)−2-[3-(4-methoxyphenyl)propoxy] ethyl-1H-imidazole hydrochloride (SKF96365, #1147, Tocris) was used to identify TRPC currents by perfusion at 100 μm in the ACSF.
Immunohistochemistry
Chow-fed male mice at 10–12 weeks of age received daily intraperitoneal injections of various combination of Veh, Lor, and/or Bar for 1 or 20 d (as described above). One hour after the last injection, mice were anesthetized with inhaled isoflurane, and quickly perfused with saline, followed by 10% formalin. The brain sections were cut at 25 µm and collected into five consecutive series. One series of sections from each mouse were blocked (3% Normal donkey serum) for 1 h, incubated with rabbit anti-β-endorphin antibody (1:10,000; #H-02,233, Phoenix Peptide) and mouse anti-c-Fos antibody (1:1000, Ab208942, Abcam) on shaker at room temperature for overnight, followed by the donkey anti-rabbit Alexa Fluor 488 (1:200, A21206, Invitrogen) and donkey anti-mouse Alexa Fluor 594 (1:200, A21203, Invitrogen) for 2 h. Slides were cover-slipped and analyzed using a fluorescence microscope. The numbers of β-endorphin-positive cells and c-Fos/β-endorphin double positive cells in the ARH were counted in 8–10 brain sections, which contained around 800–1100 β-endorphin-positive cells. The ratio of c-Fos/β-endorphin double positive cells to β-endorphin-positive cells was used to reflect the data value for that mouse. Three or four mice were included in each group for statistical analyses.
Fiber photometry
For the fiber photometry studies, 10-week-old chow-fed male POMC-Cre mice were anesthetized by isoflurane and received stereotaxic injections of pAAV9.Syn.Flex.GCaMP6m WPRE.SV40 virus (200 nl, 3 × 109 VP/ml; Addgene, 100838-AAV9) into the ARH (1.60 mm posterior, 0.30 mm lateral, and 5.90 mm ventral to the bregma, based on Franklin and Paxinos Mouse Brain Atlas). During the same surgery, an optical fiber (fiber: core = 400 μm; 0.48 NA; M3 thread titanium receptacle; Doric Lenses) was implanted over the ARH (1.60 mm posterior, 0.30 mm lateral, and 5.70 mm ventral to the bregma). Fibers were fixed to the skull using dental acrylic and mice were allowed to recover for four to six weeks before fiber photometry recording experiments. The ARH virus injection was confirmed by postmortem GFP fluorescence study, and only mice with GFP expression within the ARH were included in this study.
Mice rested for four to six weeks after surgeries to allow for adequate recovery and GCaMP6m expression to stabilize. Then mice received daily intraperitoneal injections of various combination of Veh, Lor, and/or Bar for 20 d (as described above). On days 1, 10, and 20, the recordings were performed from 5 min before the second injection of that day and for 55 min afterward. As previously described (He et al., 2020), mice could freely move during the recoding. Continuous <20 μW blue LED at 465 nm and UV LED at 405 nm served as excitation light sources, driven by a multichannel hub (Doric Lenses), modulated at 211 Hz and 330 Hz, respectively. Approximately 30-s recoding time was removed from the total recoding period to avoid the noise generated by the intraperitoneal injection. The digital signals were then amplified, demodulated, and collected through a lock-in amplifier (RZ5P, Tucker-Davis Technologies). The fiber photometry data were collected using Synapse 2.0 (Tucker-Davis Technologies) and down sampled to 8 Hz. The values of fluorescence were calculated by (F465 − F405)/F405; for each recording, these values were normalized to the average of the first 5-min baseline (before injections).
Experimental design and statistical analyses
The minimal sample size was predetermined by the nature of experiments. The data are presented as mean ± SEM and/or individual data points. Statistical analyses were performed using GraphPad Prism to evaluate normal distribution and variations within and among groups. Methods of statistical analyses were chosen based on the design of each experiment and are indicated in figure legends; p < 0.05 was considered to be statistically significant.
Study approval
Care of all animals and procedures were approved by the Baylor College of Medicine Institutional Animal Care and Use Committee.
Result
Bar inhibits Lor-induced 5-HT2CRs internalization in HEK293 cells
It is well known that long-term (many minutes to hours or days) agonist exposure induces GPCR internalization from the cell membrane, which leads to the desensitization of the receptor (Rajagopal and Shenoy, 2018). To examine whether Lor exposure induces 5-HT2CR internalization, we used Lor to treat HEK293 cells transiently transfected with mCherry-tagged 5-HT2CR-VGV (Fig. 1A). We found that 30-min Lor (100 μm) treatment induced a robust internalization of 5-HT2CRs compared with the Veh-treated group (Fig. 1A,B). Bar is a novel selective β-arrestin/β2-adaptin inhibitor that has been reported to block agonist-induced internalization of multiple GPCRs (Beautrait et al., 2017). We next tested whether Bar inhibits Lor-induced 5-HT2CR internalization. We treated the cells with Bar (10 μm) for 30 min followed by 30-min treatment of Lor. We observed that pretreatment with Bar significantly prevented Lor-induced 5-HT2CR internalization (Fig. 1A,B). This data indicates that Bar inhibits Lor-induced 5-HT2CRs internalization and may be combined with Lor to enhance its long-term efficacy.
Figure 1.

Bar inhibits Lor-induced 5-HT2CR internalization in HEK293 cells. A, Representative confocal fluorescence images in Veh + Veh, Veh + Lor, Bar + Veh, and Bar + Lor groups. mCherry (red): 5-HT2CR; WGA (green): cell membrane; Hoechst (blue): cell nucleus. B, Quantification of fluorescence intensity of 5-HT2CR-VGV-mCherry on cell membrane. Two-way ANOVA, Lor × Bar: F(1,16) = 11.92, p = 0.0033, post hoc Sidak's multiple comparisons test, **p < 0.01, ***p < 0.001. N = 5 for each group.
Bar enhances Lor's effects on food intake and body weight
To determine the long-term effects of Lor on body weight, we performed daily intraperitoneal injection of Lor (9 mg/kg) in chow-fed mice for 26 d and measured the body weight and food intake every 2 d (Fig. 2A). During the first 14 d of treatment, we found that Lor significantly decreased body weight and food intake (Fig. 2B–D). However, continuous treatment with Lor during day 15 to day 26 failed to decrease food intake and body weight (Fig. 2B,C,E). Food intake in Lor-treated group eventually showed no difference compared with the control group on day 26 (Fig. 2C). The result is consistent with a previous study showing that continuous Lor treatment for the first two weeks induces hypophagia and body weight loss in rats, whereas this effect is blunted afterward (Thomsen et al., 2008). In line with these observations in animals, clinical studies showed a clear halt of body weight loss in obese patients who took Lor for more than six months (Fidler et al., 2011). These results indicate that long-term Lor administration fails to maintain its efficacy to reduce food intake and body weight.
Figure 2.

Bar enhances Lor's effects on chow intake and body weight. A, Experimental scheme. B, C, Body weight change (B) and chow intake (C) during 26-d treatment. Two-way repeated measures ANOVA, body weight: time × group: F(39,312) = 23.17, p < 0.0001; food intake: time × group: F(42,336) = 23.17, p < 0.0001, post hoc Sidak's multiple comparisons test; *p < 0.05, **p < 0.01, ***p < 0.001 (blue asterisk: Veh + Veh vs Veh + Lor, Bar + Veh vs Veh + Lor; pink asterisk: Veh + Veh vs Bar + Lor, Bar + Veh vs Bar + Lor); #p < 0.05, ##p <0.01, ###p < 0.001 (Veh + Lor vs Bar + Lor). N = 9 mice per group. Data are presented as mean ± SEM. D, E, Cumulative food intake during day 1 to day 14 (D) and during day 15 to day 26 (E). Two-way ANOVA, Lor × Bar (day 1 to day 14): F(1,32) = 0.768, p = 0.387; Lor × Bar (day 15 to day 26): F(1,32) = 6.12, p = 0.019, post hoc Sidak's multiple comparisons test, **p < 0.01, ***p < 0.001. N = 9 mice per group. Data are presented as mean ± SEM with individual data points. F, G, Meal frequency (F) and meal size (G) measured by the TSE PhenoMaster system during day 27 to day 32. Two-way ANOVA, meal frequency: Lor × Bar (total day: F(1,24) = 0.18, p = 0.672; dark phase: F(1,24) = 0.01, p = 0.912; light phase: F(1,24) = 0.03, p = 0.87), meal size: Lor × Bar (total day: F(1,24) = 4.54, p = 0.0435; dark phase: F(1,24) = 2.71, p = 0.1126; light phase: F(1,24) = 1.49, p = 0.234), with post hoc Sidak's multiple comparisons test; *p < 0.05. N = 7 mice per group. Data are presented as mean ± SEM with individual data points. H, I, Body fat mass (H) and lean mass (I) measured on day 26. Two-way ANOVA, fat mess: Lor × bar F(1,32) = 0.265, p = 0.61; lean mess: F(1,32) = 0.036, p = 0.85, with post hoc Sidak's multiple comparisons test; *p < 0.05. N = 9 mice per group. Data are presented as mean ± SEM with individual data points.
To test whether Bar enhances long-term Lor's effect on food intake and body weight, we applied Bar (0.3 mg/kg, i.p.) 1 h before Lor injection every day. We observed that the application of Lor in Bar-pretreated group exerted a similar effect on body weight loss and food intake reduction during the first two weeks (Fig. 2B–D). Interestingly, during day 15 to day 26, Lor maintained its effect on body weight and food intake in the Bar-pretreated group (Fig. 2B,C,E). We further found that the lower food intake in the Bar-pretreated group was because of decreased meal size but not meal frequency (Fig. 2F,G). In addition, fat mass, but not lean mass, was significantly decreased by the Bar/Lor combination (Fig. 2H,I). Importantly, the application of Bar alone did not alter body weight, food intake, fat mass, and lean mass in mice (Fig. 2B–G). We found that Lor did not change energy expenditure (Fig. 3A–F), although there were significant decreases in activities at the xy- and z-axis at a few time points during the dark cycle (Fig. 3G–J). Similarly, in HFD-induced obese mice, pretreatment with Bar significantly potentiated anorexia and weight loss induced by Lor, while Bar alone did not alter food intake and body weight (Fig. 4A,B). Taken together, these data indicate that Bar enhances long-term Lor efficacy to reduce food intake and body weight in both chow and HFD-fed mice.
Figure 3.

Bar does not change energy expenditure and physical activity. A, C, E, G, I, Temporal changes in oxygen consumption (A), carbon dioxide production (C), heat production (E), xy-axis activity (G), and z-axis activity (I) in Veh + Veh, Veh + Lor, Bar + Veh, and Bar + Lor groups. N = 7 mice per group. Data are presented as mean ± SEM. Two-way repeated measures ANOVA, [dot]VO2: time × group (dark phase: F(39,234) = 1.43, p = 0.058; light phase: F(36,216) = 1.23, p = 0.19); [dot]VCO2: time × group (dark phase: F(39,234) = 1.42, p = 0.065; light phase: F(36,216) = 1.24, p = 0.18); heat: time × group (dark phase: F(39,234) = 1.44, p = 0.056; light phase: F(36,216) = 1.24, p = 0.18); heat: time × group (dark phase: F(39,234) = 1.44, p = 0.056; light phase: F(36,216) = 1.24, p = 0.18); xy-axis activity: time × group (dark phase: F(39,234) = 2.26, p < 0.0001; light phase: F(36,216) = 0.967, p = 0.53); z-axis activity: time × group (dark phase: F(39,234) = 2.01, p = 0.0008; light phase: F(36,216) = 0.94, p = 0.58), with post hoc Sidak's multiple comparisons test. *p < 0.05, **p < 0.01, ***p < 0.001. N = 7 mice per group. Data are presented as mean ± SEM. B, D, F, H, J, Average dark and light oxygen consumption (B), carbon dioxide production (D), heat production (F), xy-axis activity (H), and z-axis activity (J) in Veh + Veh, Veh + Lor, Bar + Veh, and Bar + Lor groups. Two-way ANOVA, Lor × Bar: ([dot]VO2: dark phase: F(1,24) = 0.32, p = 0.57; light phase: F(1,24) = 0.58, p = 0.45; [dot]VCO2: dark phase: F(1,24) = 0.71, p = 0.41; light phase: F(1,24) = 0.95, p = 0.34; heat: dark phase: F(1,24) = 0.33, p = 0.57; light phase: F(1,24) = 0.10, p = 0.74; xy-axis activity: dark phase: F(1,24) = 0.93, p = 0.34; light phase: F(1,24) = 0.08, p = 0.78; Y-axis activity: dark phase: F(1,24) = 0.09, p = 0.76; light phase: F(1,24) = 0.10, p = 0.75), with post hoc Sidak's multiple comparisons test. N = 7 mice per group. Data are presented as mean ± SEM with individual data points.
Figure 4.

Bar enhances Lor's effects on HFD intake and body weight. A, Body weight change during 24-d treatment. Two-way repeated measures ANOVA, time × group: F(36,180) = 12.89, p < 0.0001, with post hoc Sidak's multiple comparisons test. ***p < 0.001 (blue asterisk: Veh + Veh vs Veh + Lor, Bar + Veh vs Veh + Lor; pink asterisk: Veh + Veh vs Bar + Lor, Bar + Veh vs Bar + Lor); ###p < 0.001 (Veh + Lor vs Bar + Lor). B, HFD intake during 24-d treatment. Two-way repeated measures ANOVA, time × group: F(42,210) = 1.33, p = 0.098, with post hoc Sidak's multiple comparisons test. *p < 0.05, ***p < 0.001 (blue asterisk: Veh + Veh vs Veh + Lor, Bar + Veh vs Veh + Lor; pink asterisk: Veh + Veh or Bar + Veh vs Bar + Lor); #p < 0.05, ##p < 0.01 (Veh + Lor vs Bar + Lor).
Bar maintains POMCARH neuron responses to Lor puff in vitro
POMCARH neurons co-expressing 5-HT2CRs are shown to be a principal mediator of 5-HT2CR's effects on metabolic functions (Xu et al., 2008, 2010; Fidler et al., 2011). Acute exposure to Lor activates POMCARH neurons and induces hypophagia in mice (D'Agostino et al., 2018). The blunted effect of Lor on food intake and body weight may be because of desensitized actions of 5-HT2CRs in POMCARH neurons. To test this possibility, POMC-CreER/Rosa26-LSL-tdTOMATO mice (after tamoxifen induction) received intraperitoneal injections of Veh or Bar followed by Veh or Lor for 20 consecutive days, and we then performed the whole-cell electrophysiology recordings in tdTOMATO-labeled POMCARH neurons (Fig. 5A,B). We first compared the baseline excitability of POMCARH neurons in all groups, and found that 20-d Lor application significantly reduced RMP compared with the Veh-treated group, and firing frequency was slightly reduced with no statistical significance; interestingly, pretreatment with Bar rescued these reductions (Fig. 5C,D). We then examined responses of POMCARH neurons to a 5-s puff of 100 μm Lor. In mice receiving 20-d intraperitoneal injections of Veh and Veh, 52.63% POMCARH neurons were activated by the Lor puff, as demonstrated by >2 mV depolarization in RMP (Fig. 5E,K). However, the same Lor puff only activated 24.49% POMCARH neurons in mice receiving 20-d intraperitoneal injections of Veh and Lor (p = 0.0069 in χ2 test compared with Veh + Veh group; Fig. 5F,K). Moreover, we found that the depolarization and increases in firing frequency after Lor puff were significantly lower in mice receiving 20-d intraperitoneal injections of Lor than other groups (Fig. 5F,I,J). We further analyzed the changes induced by Lor puff only in depolarized neurons, and found that both the magnitude of depolarization and increases in firing frequency induced by Lor puff were significantly reduced in mice receiving 20-d intraperitoneal injections of Veh and Lor (Fig. 5L,M). These results indicate that the long-term Lor intraperitoneal injections significantly reduced the 5-HT2CR actions in POMCARH neurons. Interestingly, in mice receiving 20-d intraperitoneal injections of Bar and Lor, effects of Lor puff to activate POMCARH neurons were completely restored (Fig. 5G,H). Importantly, in mice receiving 20-d intraperitoneal injections of Bar and Veh, effects of Lor puff were comparable to those in mice injected with Veh and Veh (Fig. 5I–M). These data indicate that long-term systemic Lor treatment (via intraperitoneal injections) can desensitize 5-HT2CR actions in POMCARH neurons, which can be fully restored by preinjections of Bar.
Figure 5.

Bar maintains POMCARH neuron responses to Lor puff in vitro. A, Experimental scheme. B, tdTomato fluorescent labeled POMC neurons in the ARH (upper panel) and positioning of puff and recording electrodes as illustrated by white lines. C, Quantification of RMP in all recorded neurons in various groups before Lor treatment. Two-way ANOVA, Lor × Bar: F(1,163) = 4.65, p = 0.033, with post hoc Sidak's multiple comparisons test. *p < 0.05. N = 38–52 neurons per group from at least 3 mice. Data are presented as mean ± SEM with individual data points. D, Quantification of firing rate in all recorded neurons in various groups before Lor treatment. Two-way ANOVA, Lor × Bar: F(1,163) = 0.98, p = 0.32, with post hoc Sidak's multiple comparisons test. **p < 0.05. N = 38–52 neurons per group from at least 3 mice. Data are presented as mean ± SEM with individual data points. E–H, Representative traces of action firing potential in POMCARH neurons before and after Lor treatment (100 μm, 5-s puff) in Veh + Veh (E), Veh + Lor (F), Bar + Veh (G), and Bar + Lor (H) groups. I, Change in membrane potential in all recorded neurons treated by Lor in various groups. Two-way ANOVA, Lor × Bar: F(1,162) = 7.24, p = 0.0079, with post hoc Sidak's multiple comparisons test. **p < 0.01, ***p < 0.001. N = 38–49 neurons per group from at least 3 mice. Data are presented as mean ± SEM with individual data points. J, Changes in firing rate in all recorded neurons treated by Lor in various groups. Two-way ANOVA, Lor × Bar: F(1,162) = 8.58, p = 0.0039, with post hoc Sidak's multiple comparisons test. **p < 0.01. N = 38–49 neurons per group from at least 3 mice. K, Percentage of neurons with depolarization or no response induced by Lor in various groups. χ2 test; **p < 0.01. L, Magnitude of depolarization in activated neurons induced by Lor in various groups. Two-way ANOVA, Lor × Bar: F(1,68) = 5.18, p = 0.026, with post hoc Sidak's multiple comparisons test. **p < 0.01. N = 12–20 neurons per group from at least 3 mice. Data are presented as mean ± SEM with individual data points. M, Changes in firing rate in activated neurons induced by Lor in various groups. Two-way ANOVA, Lor × Bar: F(1,68) = 5.68, p = 0.02; with post hoc Sidak's multiple comparisons test. *p < 0.05. N = 12–20 neurons per group from at least 3 mice. Data are presented as mean ± SEM with individual data points.
It has been reported that 5-HT2CRs activate POMCARH neurons at least partly via putative TRPC channels (Sohn et al., 2011). In mice receiving 20-d intraperitoneal injections of Veh and Veh, we observed that Lor puff induced clear inward currents in activated POMCARH neurons which were fully blocked by a TRPC channel blocker, SKF96365 (Fig. 6A,E); in addition, we observed that repeated Lor puffs (with 5-min interval) reproducibly triggered the inward currents in POMCARH neurons (data not shown). Importantly, such currents were minimal in unresponsive POMCARH neurons (Fig. 6F). Interestingly, in mice receiving 20-d intraperitoneal injections of Veh and Lor, effects of Lor puff to induce the SKF96365-sensitive inward currents were significantly decreased, which however were largely restored in mice receiving 20-d intraperitoneal injections of Bar and Lor (Fig. 6B,D,F). Notably, effects of Lor puff to induce the inward currents in mice receiving 20-d intraperitoneal injections of Bar and Veh were comparable to those in mice injected with Veh and Veh (Fig. 6C,F). Further, we noted that the depolarization induced by Lor puff was positively correlated with the currents (Fig. 6G), suggesting that these SKF96365-sensitive currents mediate the Lor-induced depolarization. Supporting this notion, SKF96365 can fully block Lor's activation effect on POMCARH neurons (Fig. 6H–J). Taken together, these results indicate that long-term systemic Lor treatment can desensitize 5-HT2CR actions to open the TRPC channel, which can be restored by preinjections of Bar.
Figure 6.
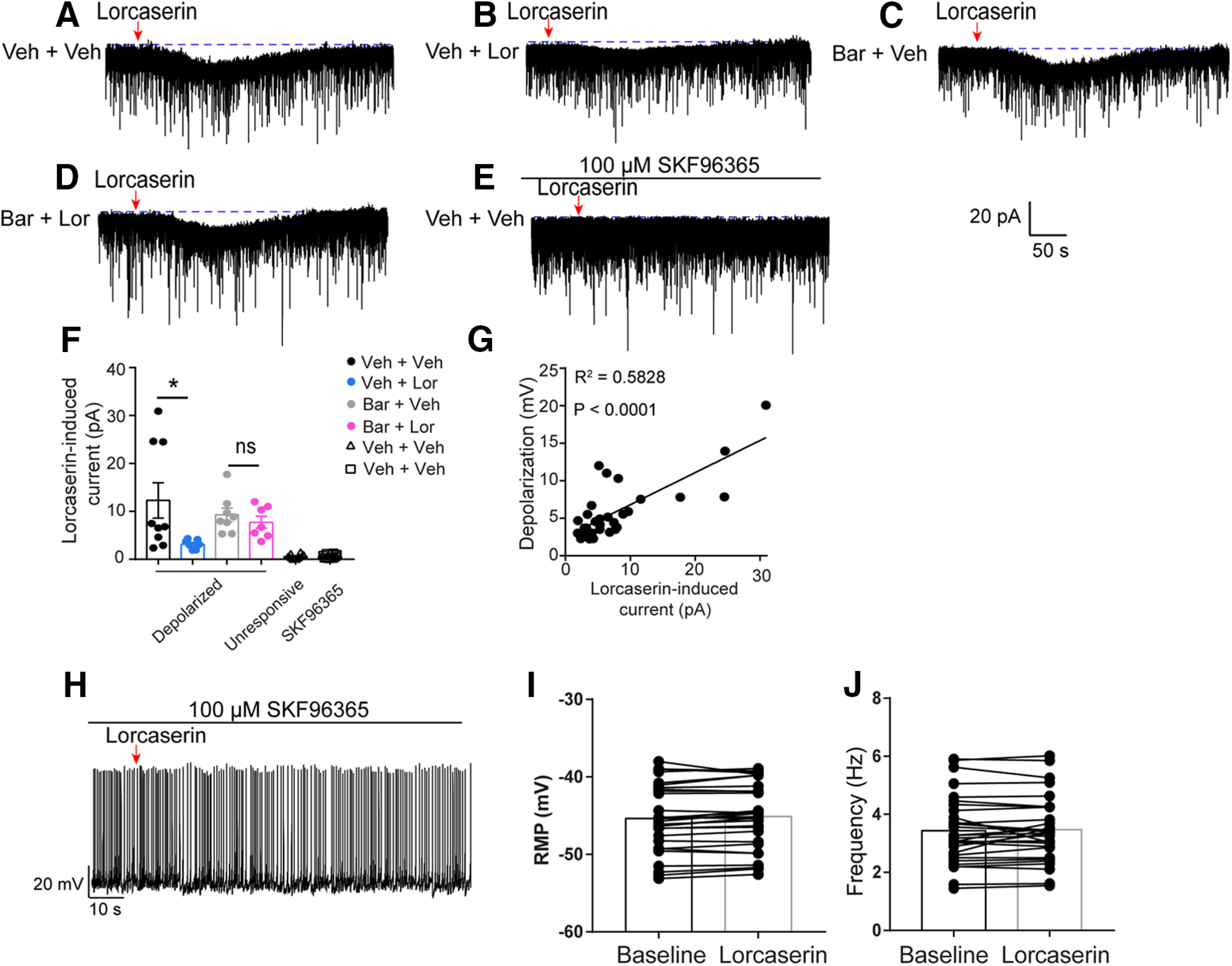
Lor depolarizes POMCARH neurons via TRPC channels. A–E, Representative traces of Lor-induced inward currents in POMCARH neurons after Lor treatment (100 μm, 5-s puff) in the absence of SKF96365 in Veh + Veh (A), Veh + Lor (B), Bar + Veh (C), Bar + Lor (D), or in the presence of SKF96365 in Veh + Veh (E) groups. F, Quantification of TRPC currents induced by Lor in various groups. Two-way ANOVA, Lor × Bar: F(1,27) = 5.68, p = 0.12, with post hoc Sidak's multiple comparisons test. *p < 0.05. N = 7–10 neurons per group from at least 3 mice. Data are presented as mean ± SEM with individual data points. G, Linear-regression analysis of the correlation between the depolarization level and the TRPC currents induced by Lor. Linear regression; R2 = 0.5828, p < 0.0001. H, Representative traces of action firing potential in POMCARH neurons before and after Lor treatment (100 μm, 5-s puff) in the presence of SKF96365. I–J, Quantification of the RMP (I) and firing frequency (J) of neurons treated by Lor in the presence of SKF96365. N = 30 neurons from at least 3 mice. Two tailed unpaired t tests. Data are presented as mean with single dots and line. ns, not significant.
Bar maintains Lor-induced c-Fos immunoreactivity in POMCARH neurons
To further determine the Lor effects on POMCARH neuron activity in vivo, we examined the c-Fos immunoreactivity in POMCARH neurons in mice that received intraperitoneal injections of Veh or Bar followed by Veh or Lor for either 1 d or for 20 consecutive days (Fig. 7A). Consistent with a previous report (Burke et al., 2017), we found that Lor treatment on day 1 increased the number of POMCARH neurons that co-express c-Fos (p < 0.01 in two tailed unpaired t tests but nonsignificant in two-way ANOVA analysis, Fig. 7B,C). However, after a 20-d continuous Lor daily injections, the last Lor injection failed to induce c-Fos in POMCARH neurons (Fig. 7D,E), indicating that 5-HT2CRs in POMCARH neurons no longer responded to Lor after a long-term Lor administration. Interestingly, in mice receiving pretreatment of Bar every day, Lor significantly induced c-Fos in POMCARH neurons (Fig. 7D,E). Notably, the pretreatment of Bar can further potentiate the effect of Lor to induce c-Fos on day 1, and Bar alone did not affect the c-Fos in POMCARH neurons on either day 1 or 20 (day 1: 16.8 ± 3.4% vs day 20: 18.6 ± 0.5%, p > 0.05; Fig. 7B,E). These results indicate that Bar can maintain Lor's effect to activate POMCARH neurons likely through preventing the 5-HT2CR desensitization.
Figure 7.

Bar maintains Lor-induced c-Fos immunoreactivity in POMCARH neurons. A, Experimental scheme. B, D, c-fos immunoreactivity in POMCARH neurons in Veh + Veh, Veh + Lor, Bar + Veh, and Bar + Lor on day 1 (B) and day 20 (D) after injection of Lor (9 mg/kg) for 1 h. Arrowheads point to double-labeled neurons. C, E, Quantification of c-fos immunoreactivity in POMCARH neurons in various groups on day 1 (C) and day 20 (E) after injection of Lor. Two-way ANOVA, day 1: Lor × Bar: F(1 8) = 20.11, p = 0.002; day 20: Lor × Bar: F(1 9) = 7.04, p = 0.03, with post hoc Sidak's multiple comparisons test; **p < 0.01, ***p < 0.001. $$p < 0.01 in two tailed unpaired t tests, which was not detected by the two-way ANOVA analysis. N = 3 or 4 mice per group. Data are presented as mean ± SEM with individual data points. The right panels in B, D are higher magnification images. ns, not significant.
Bar maintains POMCARH neuron responses to Lor treatment in vivo
We further used fiber photometry to record Lor-induced POMCARH neuron activity in vivo. To this end, POMC-Cre mice received stereotaxic injections of a Cre-dependent viral vector expressing a calcium indicator GCaMP6m into the ARH, and an optic fiber detector was implanted above the ARH to allow recording of GCaMP6m fluorescence as a measure of POMCARH neuron activity (Fig. 8A,B). After recovery, these mice were subjected to the 20-d intraperitoneal injection regimen: Veh or Bar followed by Veh or Lor (Fig. 8A). The fiber photometry recordings were performed on days 1, 10, and 20 (Fig. 8A,B). The combination of Veh and Veh injection, or that of Bar and Veh, did not change the POMCARH neuron activity on day 1, 10, or 20 (Fig. 8C,E,G). In mice receiving the combination of Veh and Lor, the Lor intraperitoneal injection induced a significant and robust increase in POMCARH neuron activity only on day 1, and this response was largely attenuated on days 10 and 20 (day 1: 1.25 ± 0.058 vs day 20: 1.04 ± 0.021, p < 0.05; Fig. 8D,G). On the contrary, in mice receiving the combination of Bar and Lor, the Lor intraperitoneal injection induced a significant and robust increase in POMCARH neuron activity on day 1, and this response persisted on days 10 and 20 (Fig. 8F,G). Taken together, these results further confirm in vivo that long-term systemic Lor treatment can desensitize 5-HT2CR actions in POMCARH neurons, which can be fully restored by preinjections of Bar.
Figure 8.
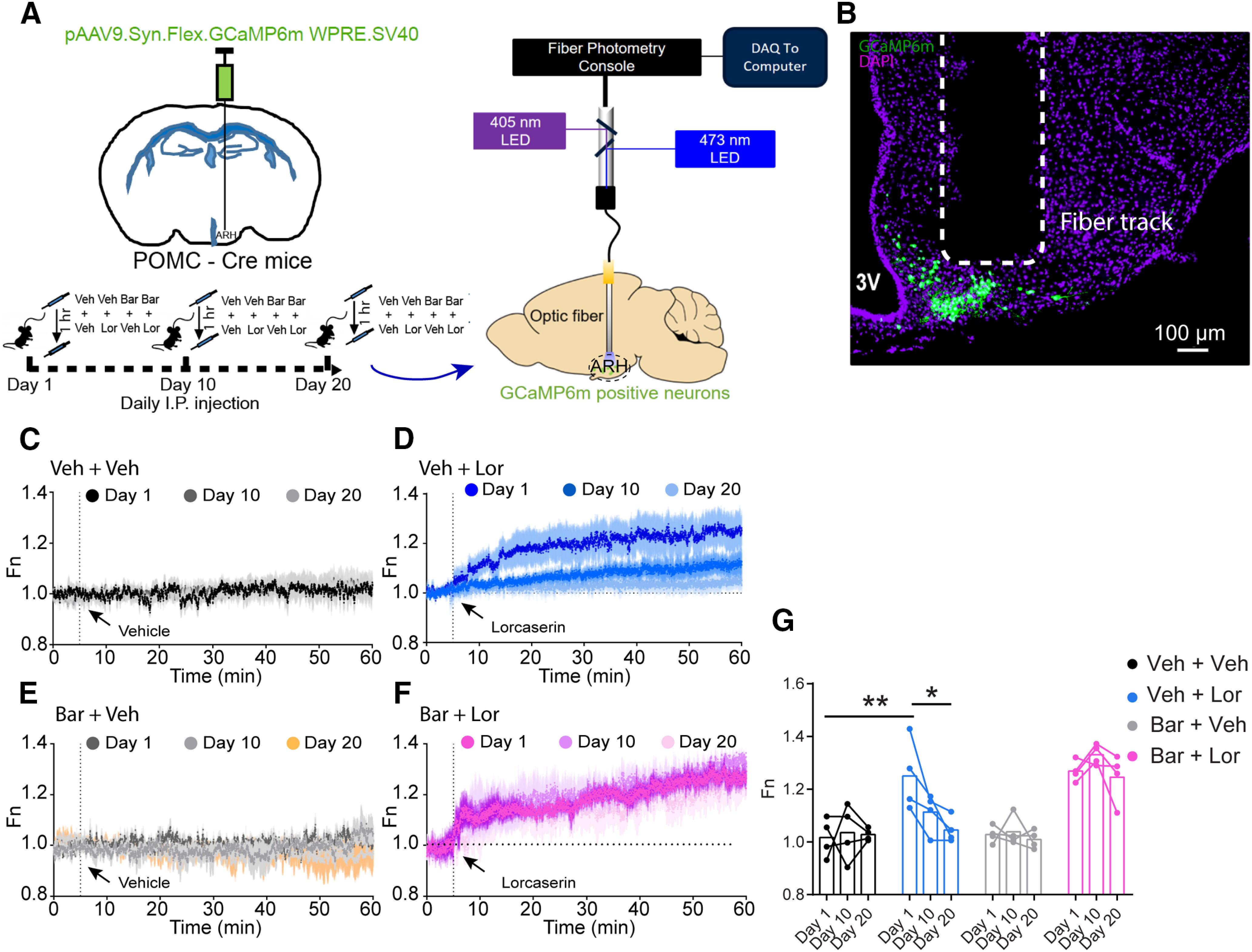
Bar maintains POMCARH neuron responses to Lor treatment in vivo. A, Experimental scheme. B, Representative image showing dual immunofluorescence for GCaMP6m (green) and DAPI (purple) in the ARH of a POMC-Cre mouse. C–E, Quantification of neural activity of POMCARH neurons in response to intraperitoneal Veh or intraperitoneal Lor (9 mg/kg) in mice treated with Veh + Veh (C), Veh + Lor (D), Bar + Veh (E), and Bar + Lor (F) on days 1, 10, and 20. Two-way repeated measures ANOVA, time × group: Veh + Veh: F(7198,21594) = 0.76, p > 0.99, Veh + Lor: F(7198,21594) = 2.66, p < 0.0001, Bar + Veh: F(7198,21594) = 0.99, p = 0.77, Bar + Lor: F(7198,21594) = 0.81, p > 0.99, with post hoc Sidak's multiple comparisons test. Data are presented as mean ± SEM. G, Quantification of the average Fn value in last 5 min. C–E, Two-way ANOVA, time × group: F(6,36) = 2.42, p = 0.046, with post hoc Sidak's multiple comparisons test. *p < 0.05, **p < 0.01. N = 4 mice per group. Data are presented as mean ± SEM with individual data points.
Discussion
Our work demonstrates that long-term Lor administration induces aberrant 5-HT2CR desensitization in POMCARH neurons, which blunts Lor's long-term efficacy to reduce body weight. Bar can prevent 5-HT2CR internalization and maintain Lor potency to activate POMCARH neurons via the TRPC-mediated mechanisms and to reduce body weight. Our findings provide a necessary framework to target 5-HT2CR desensitization pathway for effective long-term treatment of obesity.
The major shortcoming of the several 5-HT2CR agonists is the modest long-term efficacy to reduce body weight. Data from multiple clinical trials clearly show that the Lor's efficacy gradually reached a plateau after ∼24 weeks of treatment (Smith et al., 2010; Fidler et al., 2011; O'Neil et al., 2012), and even an apparent body weight rebound existed in obese patients who still received continuous Lor treatment (Smith et al., 2010). Interestingly, this plateau phenomenon also existed in the clinical trials with >24 weeks of treatment with d-fenfluramine or sibutramine (Douglas et al., 1983; Apfelbaum et al., 1999; James et al., 2010). The animal studies also showed that Lor, as well as other 5-HT analogs, significantly reduced food intake over the first two weeks of dosing, after which time there was a progressive loss of this hypophagia (Mccann et al., 1997; Vickers et al., 2000; Thomsen et al., 2008). Likewise, our results also showed that the Lor gradually lose its effect after two weeks of administration. It has been well documented that Lor and other 5-HT analogs reduce body weight through 5-HT2CR-mediated activation of POMCARH neurons (Heisler et al., 2002, 2003; Vickers et al., 2003; Sohn et al., 2011; D'Agostino et al., 2018), indicating that POMCARH neuron activity is essential for weight-lowering effects of these compounds. Notably, using slice electrophysiology, c-Fos immunoreactivity and fiber photometry, we consistently demonstrated that the Lor treatment hardly activated POMCARH neurons after prolonged Lor exposure. Thus, we suggest that the loss of Lor's efficacy to reduce body weight and food intake is largely attributed to the desensitization of 5-HT2CRs in POMCARH neurons.
The GPCRs desensitization is a complicated process which requires the coordination of multiple proteins, including β-arrestins (β-arrestin-1 and β-arrestin-2), clathrin and adaptor protein AP2 (Ferguson et al., 1996; Goodman et al., 1996; Laporte et al., 1999). The role of β-arrestins in regulating GPCR signaling and trafficking has been well studied (Shenoy and Lefkowitz, 2011). β-Arrestin-1 null mice showed less β-adrenergic receptors desensitization and more sensitive to the agonist stimulation (Conner et al., 1997). Mice lacking β-arrestin-2 exhibited enhanced and prolonged morphine analgesia because of reduced µ-opioid receptor desensitization (Bohn et al., 1999, 2000). A recent study reported that β-arrestin-1, but not β-arrestin-2, in hypothalamic agouti-related peptide neurons prevents obesity-associated metabolic impairments (Pydi et al., 2020). Importantly, the interaction between β-arrestins and adaptor protein AP2 is indispensable for the process of GPCR desensitization (Ferguson et al., 1996; Goodman et al., 1996; Laporte et al., 1999). Bar, a novel inhibitor of β-arrestin/AP2 complex, blocks agonist-promoted internalization of various GPCRs at the cellular level (Beautrait et al., 2017). Bar was reported to ameliorate the opiate drug tolerance by blocking β-arrestin-mediated µ-opioid receptor internalization (Aberoumandi et al., 2019). Here, we demonstrated that Bar can prevent internalization of 5-HT2CRs in cells exposed to Lor, the selective 5-HT2CR agonist. In addition, in mice, we found that Bar co-treatment with prolonged Lor exposure can effectively maintain the sensitivity of POMCARH neurons to respond to Lor, indicating that Bar prevents desensitization of the 5-HT2CR. Importantly, these effects were associated with potentiated Lor efficacy to reduce body weight and food intake in mice either fed with chow diet or HFD. It is important to point out that Bar was only recently identified, and its safety profile has not been characterized yet (Beautrait et al., 2017). Further, Bar is shown to prevent desensitization of multiple GPCRs (Beautrait et al., 2017), not just the 5-HT2CR. Thus, we do not intend to imply that Bar is ready to be used to treat patients with obesity. Rather, our studies used Bar as a tool to provide proof of the principle that inhibition of 5-HT2CR desensitization pathway can be a valid strategy to improve the long-term weight loss effects of Lor or other 5-HT2CR agonists.
At the mechanistic level, the TRPC channels in POMCARH neurons have been shown to mediate activation effects of multiple hormones or neural signals, including leptin (Qiu et al., 2010, 2018; Gao et al., 2017), insulin (Qiu et al., 2018), pituitary adenylate cyclase activating polypeptide (Chang et al., 2021), and 5-HT (Sohn et al., 2011; Gao et al., 2017). Consistently, here we found that Lor induced inward currents that were abolished by the selective TRPC inhibitor, indicating that these currents are largely mediated by the TRPC channels. The TRPC inhibitor also abolished Lor-induced activation of POMCARH neurons, confirming the role of the TRPC channels in mediating Lor's effects to activate these neurons. Interestingly, the Lor-induced TRPC currents underwent similar “desensitization” in mice with prolonged Lor exposure, which were restored by Bar co-treatment. The restoration of TRPC currents is not likely because of a direct effect of Bar on TRPC channels. Rather, we suggest that the different responses in TRPC currents were secondary to alterations in 5-HT2CR sensitivity, given the tight coupling of TRPC channels with 5-HT2CR singling in POMCARH neurons (Sohn et al., 2011; Gao et al., 2017). One limitation of our study is that the systemic administration of Lor is expected to act on many other cell types that express 5-HT2CRs, and therefore may have a wide range of effects other than anorexia and weight loss. In addition, since we observed that Bar inhibits Lor-induced 5-HT2CR internalization in HEK293 cells which derived from non-neuronal cells, we suggest that Bar may regulate 5-HT2CR internalization in other cells that co-express the receptor and β-arrestin/β2-adaptin.
In the current study, we show that Bar is sufficient to enhance the long-term Lor efficacy to reduce body weight and food intake in mice. We provide evidence that the body weight benefits of this prolonged combination regimen is associated with the prevention of 5-HT2CR desensitization in POMCARH neurons. Our findings provide a necessary framework to further characterize the mechanisms and regulations of 5-HT2CR desensitization and to develop compounds that may specifically target this process as an effective strategy for the long-term management of weight.
Notably, the FDA requested a voluntary withdrawal of Lor from the United States market because Lor was potentially associated with increased cancer risk (Bohula et al., 2018; Food and Drug Administration, 2020). It was reported that more patients taking Lor were diagnosed with cancers (n = 462; 7.7%) compared with those taking a placebo treatment (n = 423; 7.1%; Food and Drug Administration, 2020). It is noteworthy that the difference in cancer cases was minimal and did not reach statistical significance, and the FDA did not conclude that Lor contributed to the cancer risk (Food and Drug Administration, 2020). While the potential association between the Lor regimen and cancer risk warrants further investigations, our findings suggest that targeting the 5-HT2CR desensitization pathway may lower the effective dose of Lor for obesity and therefore reduce the potential cancer risk.
Footnotes
This work was supported by grants from National Institutes of Health Grants R01DK114279, R01DK109934, and R21NS108091 (to Q.T.), K01DK119471 (to C.W.), R01DK109194 and R56DK109194 (to Q.W.), R01ES027544 and R01DK111436 (to Z.S.), P01DK113954, R01DK115761, R01DK117281, and R01DK125480 (to Y.X.), and R01DK120858 (to Q.T. and Y.X.); the Pew Charitable Trust Award 0026188 (to Q.W.); Baylor Collaborative Faculty Research Investment Program grants (Q.W.); the United States Department of Agriculture/Current Research Information System Grant 51000-064-01S (to Y.X. and Q.W.); American Diabetes Association Grants 7-13-JF-61 (to Q.W.) and 1-15-BS-184 (to Q.T.); and the American Heart Association Grant 20POST35120600 (to Yang He).
The authors declare no competing financial interests.
References
- Aberoumandi SM, Vousooghi N, Tabrizi BA, Karimi P (2019) Heroin-based crack induces hyperalgesia through β-arrestin 2 redistribution and phosphorylation of Erk1/2 and JNK in the periaqueductal gray area. Neurosci Lett 698:133–139. 10.1016/j.neulet.2019.01.023 [DOI] [PubMed] [Google Scholar]
- Apfelbaum M, Vague P, Ziegler O, Hanotin C, Thomas F, Leutenegger E (1999) Long-term maintenance of weight loss after a very-low-calorie diet: a randomized blinded trial of the efficacy and tolerability of sibutramine. Am J Med 106:179–184. 10.1016/S0002-9343(98)00411-2 [DOI] [PubMed] [Google Scholar]
- Balthasar N, Coppari R, McMinn J, Liu SM, Lee CE, Tang V, Kenny CD, McGovern RA, Chua SC Jr, Elmquist JK, Lowell BB (2004) Leptin receptor signaling in POMC neurons is required for normal body weight homeostasis. Neuron 42:983–991. 10.1016/j.neuron.2004.06.004 [DOI] [PubMed] [Google Scholar]
- Beautrait A, Paradis JS, Zimmerman B, Giubilaro J, Nikolajev L, Armando S, Kobayashi H, Yamani L, Namkung Y, Heydenreich FM, Khoury E, Audet M, Roux PP, Veprintsev DB, Laporte SA, Bouvier M (2017) A new inhibitor of the β-arrestin/AP2 endocytic complex reveals interplay between GPCR internalization and signalling. Nat Commun 8:15054. 10.1038/ncomms15054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berglund ED, Liu C, Sohn JW, Liu T, Kim MH, Lee CE, Vianna CR, Williams KW, Xu Y, Elmquist JK (2013) Serotonin 2C receptors in pro-opiomelanocortin neurons regulate energy and glucose homeostasis. J Clin Invest 123:5061–5070. 10.1172/JCI70338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blüher M (2019) Obesity: global epidemiology and pathogenesis. Nat Rev Endocrinol 15:288–298. 10.1038/s41574-019-0176-8 [DOI] [PubMed] [Google Scholar]
- Blundell JE, Leshem MB (1974) Central action of anorexic agents: effects of amphetamine and fenfluramine in rats with lateral hypothalamic lesions. Eur J Pharmacol 28:81–88. 10.1016/0014-2999(74)90115-0 [DOI] [PubMed] [Google Scholar]
- Bohn LM, Lefkowitz RJ, Gainetdinov RR, Peppel K, Caron MG, Lin FT (1999) Enhanced morphine analgesia in mice lacking β-arrestin 2. Science 286:2495–2498. 10.1126/science.286.5449.2495 [DOI] [PubMed] [Google Scholar]
- Bohn LM, Gainetdinov RR, Lin F-T, Lefkowitz RJ, Caron MG (2000) μ-Opioid receptor desensitization by β-arrestin-2 determines morphine tolerance but not dependence. Nature 408:720–723. 10.1038/35047086 [DOI] [PubMed] [Google Scholar]
- Bohula EA, Scirica BM, Inzucchi SE, McGuire DK, Keech AC, Smith SR, Kanevsky E, Murphy SA, Leiter LA, Dwyer JP, Corbalan R, Hamm C, Kaplan L, Nicolau JC, Ophuis TO, Ray KK, Ruda M, Spinar J, Patel T, Miao W, et al. (2018) Effect of lorcaserin on prevention and remission of type 2 diabetes in overweight and obese patients (CAMELLIA-TIMI 61): a randomised, placebo-controlled trial. Lancet 392:2269–2279. 10.1016/S0140-6736(18)32328-6 [DOI] [PubMed] [Google Scholar]
- Burke LK, Ogunnowo-Bada E, Georgescu T, Cristiano C, de Morentin PBM, Valencia Torres L, D'Agostino G, Riches C, Heeley N, Ruan Y, Rubinstein M, Low MJ, Myers MG, Rochford JJ, Evans ML, Heisler LK (2017) Lorcaserin improves glycemic control via a melanocortin neurocircuit. Mol Metab 6:1092–1102. 10.1016/j.molmet.2017.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang R, Hernandez J, Gastelum C, Guadagno K, Perez L, Wagner EJ (2021) Pituitary adenylate cyclase activating polypeptide excites proopiomelanocortin neurons: implications for the regulation of energy homeostasis. Neuroendocrinology 111:45–69. 10.1159/000506367 [DOI] [PubMed] [Google Scholar]
- Conner DA, Mathier MA, Mortensen RM, Christe M, Vatner SF, Seidman CE, Seidman J (1997) β-Arrestin1 knockout mice appear normal but demonstrate altered cardiac responses to β-adrenergic stimulation. Circ Res 81:1021–1026. 10.1161/01.res.81.6.1021 [DOI] [PubMed] [Google Scholar]
- Connolly HM, Crary JL, McGoon MD, Hensrud DD, Edwards BS, Edwards WD, Schaff HV (1997) Valvular heart disease associated with fenfluramine–phentermine. N Engl J Med 337:581–588. 10.1056/NEJM199708283370901 [DOI] [PubMed] [Google Scholar]
- D'Agostino G, Lyons D, Cristiano C, Lettieri M, Olarte-Sanchez C, Burke LK, Greenwald-Yarnell M, Cansell C, Doslikova B, Georgescu T, Martinez de Morentin PB, Myers MG, Rochford JJ, Heisler LK (2018) Nucleus of the solitary tract serotonin 5-HT2C receptors modulate food intake. Cell Metab 28:619–630.e5. 10.1016/j.cmet.2018.07.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diabetes Prevention Program Research Group, Knowler WC, Fowler SE, Hamman RF, Christophi CA, Hoffman HJ, Brenneman AT, Brown-Friday JO, Goldberg R, Venditti E, Nathan DM (2009) 10-year follow-up of diabetes incidence and weight loss in the Diabetes Prevention Program Outcomes Study. Lancet 374:1677–1686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Douglas J, Preston P, Haslett C, Gough J, Frazer I, Chalmers S, Munro J (1983) Long-term efficacy of fenfluramine in treatment of obesity. Lancet 321:384–386. 10.1016/S0140-6736(83)91501-5 [DOI] [PubMed] [Google Scholar]
- Ferguson SS, Downey WE, Colapietro AM, Barak LS, Ménard L, Caron MG (1996) Role of β-arrestin in mediating agonist-promoted G protein-coupled receptor internalization. Science 271:363–366. 10.1126/science.271.5247.363 [DOI] [PubMed] [Google Scholar]
- Fidler MC, Sanchez M, Raether B, Weissman NJ, Smith SR, Shanahan WR, Anderson CM (2011) A one-year randomized trial of lorcaserin for weight loss in obese and overweight adults: the BLOSSOM trial. J Clin Endocrinol Metab 96:3067–3077. 10.1210/jc.2011-1256 [DOI] [PubMed] [Google Scholar]
- Fontaine KR, Redden DT, Wang C, Westfall AO, Allison DB (2003) Years of life lost due to obesity. JAMA 289:187–193. 10.1001/jama.289.2.187 [DOI] [PubMed] [Google Scholar]
- Food and Drug Administration (2020) Safety clinical trial shows possible increased risk of cancer with weight-loss medicine Belviq, Belviq XR (lorcaserin). Available from https://www.fda.gov/drugs/drug-safety-and-availability/safety-clinical-trial-shows-possible-increased-risk-cancer-weight-loss-medicine-belviq-belviq-xr.
- Gao Y, Yao T, Deng Z, Sohn JW, Sun J, Huang Y, Kong X, Yu KJ, Wang RT, Chen H, Guo H, Yan J, Cunningham KA, Chang Y, Liu T, Williams KW (2017) TrpC5 mediates acute leptin and serotonin effects via POMC neurons. Cell Rep 18:583–592. 10.1016/j.celrep.2016.12.072 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geyer MA, Puerto A, Menkes DB, Segal DS, Mandell AJ (1976) Behavioral studies following lesions of the mesolimbic and mesostriatal serotonergic pathways. Brain Res 106:257–269. 10.1016/0006-8993(76)91024-6 [DOI] [PubMed] [Google Scholar]
- Ghosh MN, Parvathy S (1973) The effect of cyproheptadine on water and food intake and on body weight in the fasted adult and weanling rats. Br J Pharmacol 48:328P–329P. [PMC free article] [PubMed] [Google Scholar]
- Goodman OB, Krupnick JG, Santini F, Gurevich VV, Penn RB, Gagnon AW, Keen JH, Benovic JL (1996) β-Arrestin acts as a clathrin adaptor in endocytosis of the β 2-adrenergic receptor. Nature 383:447–450. 10.1038/383447a0 [DOI] [PubMed] [Google Scholar]
- He Y, Xu P, Wang C, Xia Y, Yu M, Yang Y, Yu K, Cai X, Qu N, Saito K, Wang J, Hyseni I, Robertson M, Piyarathna B, Gao M, Khan SA, Liu F, Chen R, Coarfa C, Zhao Z, et al. (2020) Estrogen receptor-α expressing neurons in the ventrolateral VMH regulate glucose balance. Nat Commun 11:2165. 10.1038/s41467-020-15982-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heisler LK, Cowley MA, Tecott LH, Fan W, Low MJ, Smart JL, Rubinstein M, Tatro JB, Marcus JN, Holstege H, Lee CE, Cone RD, Elmquist JK (2002) Activation of central melanocortin pathways by fenfluramine. Science 297:609–611. 10.1126/science.1072327 [DOI] [PubMed] [Google Scholar]
- Heisler LK, Cowley MA, Kishi T, Tecott LH, Fan W, Low MJ, Smart JL, Rubinstein M, Tatro JB, Zigman JM (2003) Central serotonin and melanocortin pathways regulating energy homeostasis. Ann NY Acad Sci 994:169–174. [DOI] [PubMed] [Google Scholar]
- James WPT, Caterson ID, Coutinho W, Finer N, Van Gaal LF, Maggioni AP, Torp-Pedersen C, Sharma AM, Shepherd GM, Rode RA, Renz CL (2010) Effect of sibutramine on cardiovascular outcomes in overweight and obese subjects. N Engl J Med 363:905–917. 10.1056/NEJMoa1003114 [DOI] [PubMed] [Google Scholar]
- Kyle TK, Dhurandhar EJ, Allison DB (2016) Regarding obesity as a disease: evolving policies and their implications. Endocrinol Metab Clin North Am 45:511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laporte SA, Oakley RH, Zhang J, Holt JA, Ferguson SS, Caron MG, Barak LS (1999) The beta2-adrenergic receptor/betaarrestin complex recruits the clathrin adaptor AP-2 during endocytosis. Proc Natl Acad Sci USA 96:3712–3717. 10.1073/pnas.96.7.3712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luque CA, Rey JA (1999) Sibutramine: a serotonin–norepinephrine reuptake-inhibitor for the treatment of obesity. Ann Pharmacother 33:968–978. 10.1345/aph.18319 [DOI] [PubMed] [Google Scholar]
- Madisen L, Zwingman TA, Sunkin SM, Oh SW, Zariwala HA, Gu H, Ng LL, Palmiter RD, Hawrylycz MJ, Jones AR, Lein ES, Zeng H (2010) A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci 13:133–140. 10.1038/nn.2467 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mccann UD, Yuan J, Hatzidimitriou G, Ricaurte GA (1997) Selective serotonin reuptake inhibitors dissociate fenfluramine's anorectic and neurotoxic effects: importance of dose, species and drug. J Pharmacol Exp Ther 281:1487–1498. [PubMed] [Google Scholar]
- Nonogaki K, Strack AM, Dallman MF, Tecott LH (1998) Leptin-independent hyperphagia and type 2 diabetes in mice with a mutated serotonin 5-HT 2C receptor gene. Nat Med 4:1152–1156. 10.1038/2647 [DOI] [PubMed] [Google Scholar]
- O'Neil PM, Smith SR, Weissman NJ, Fidler MC, Sanchez M, Zhang J, Raether B, Anderson CM, Shanahan WR (2012) Randomized placebo-controlled clinical trial of lorcaserin for weight loss in type 2 diabetes mellitus: the BLOOM-DM study. Obesity 20:1426–1436. 10.1038/oby.2012.66 [DOI] [PubMed] [Google Scholar]
- Pydi SP, Cui Z, He Z, Barella LF, Pham J, Cui Y, Oberlin DJ, Egritag HE, Urs N, Gavrilova O, Schwartz GJ, Buettner C, Williams KW, Wess J (2020) Beneficial metabolic role of β-arrestin-1 expressed by AgRP neurons. Sci Adv 6:eaaz1341. 10.1126/sciadv.aaz1341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiu J, Xue C, Bosch MA, Murphy JG, Fan W, Rønnekleiv OK, Kelly MJ (2007) Serotonin 5-hydroxytryptamine2C receptor signaling in hypothalamic proopiomelanocortin neurons: role in energy homeostasis in females. Mol Pharmacol 72:885–896. 10.1124/mol.107.038083 [DOI] [PubMed] [Google Scholar]
- Qiu J, Fang Y, Rønnekleiv OK, Kelly MJ (2010) Leptin excites proopiomelanocortin neurons via activation of TRPC channels. J Neurosci 30:1560–1565. 10.1523/JNEUROSCI.4816-09.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiu J, Wagner EJ, Rønnekleiv OK, Kelly MJ (2018) Insulin and leptin excite anorexigenic pro-opiomelanocortin neurones via activation of TRPC5 channels. J Neuroendocrinol 30:e12501. 10.1111/jne.12501 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajagopal S, Shenoy SK (2018) GPCR desensitization: acute and prolonged phases. Cell Signal 41:9–16. 10.1016/j.cellsig.2017.01.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saller CF, Stricker EM (1976) Hyperphagia and increased growth in rats after intraventricular injection of 5,7-dihydroxytryptamine. Science 192:385–387. 10.1126/science.1257774 [DOI] [PubMed] [Google Scholar]
- Scheen AJ (2010) Cardiovascular risk-benefit profile of sibutramine. Am J Cardiovasc Drugs 10:321–334. [DOI] [PubMed] [Google Scholar]
- Shenoy SK, Lefkowitz RJ (2011) β-Arrestin-mediated receptor trafficking and signal transduction. Trends Pharmacol Sci 32:521–533. 10.1016/j.tips.2011.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SR, Weissman NJ, Anderson CM, Sanchez M, Chuang E, Stubbe S, Bays H, Shanahan WR, Modification B, Overweight L (2010) Multicenter, placebo-controlled trial of lorcaserin for weight management. N Engl J Med 363:245–256. 10.1056/NEJMoa0909809 [DOI] [PubMed] [Google Scholar]
- Sohn JW, Xu Y, Jones JE, Wickman K, Williams KW, Elmquist JK (2011) Serotonin 2C receptor activates a distinct population of arcuate pro-opiomelanocortin neurons via TRPC channels. Neuron 71:488–497. 10.1016/j.neuron.2011.06.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefan N, Häring HU, Schulze MB (2018) Metabolically healthy obesity: the low-hanging fruit in obesity treatment? Lancet Diabetes Endocrinol 6:249–258. 10.1016/S2213-8587(17)30292-9 [DOI] [PubMed] [Google Scholar]
- Tecott LH, Sun LM, Akana SF, Strack AM, Lowenstein DH, Dallman MF, Julius D (1995) Eating disorder and epilepsy in mice lacking 5-HT2c serotonin receptors. Nature 374:542–546. 10.1038/374542a0 [DOI] [PubMed] [Google Scholar]
- Thomsen WJ, Grottick AJ, Menzaghi F, Reyes-Saldana H, Espitia S, Yuskin D, Whelan K, Martin M, Morgan M, Chen W (2008) Lorcaserin, a novel selective human 5-hydroxytryptamine2C agonist: in vitro and in vivo pharmacological characterization. J Pharmacol Exp Ther 325:577–587. 10.1124/jpet.107.133348 [DOI] [PubMed] [Google Scholar]
- Vickers SP, Clifton PG, Dourish CT, Tecott LH (1999) Reduced satiating effect of d-fenfluramine in serotonin 5-HT(2C) receptor mutant mice. Psychopharmacology 143:309–314. 10.1007/s002130050952 [DOI] [PubMed] [Google Scholar]
- Vickers S, Benwell K, Porter R, Bickerdike M, Kennett G, Dourish C (2000) Comparative effects of continuous infusion of mCPP, Ro 60-0175 and d-fenfluramine on food intake, water intake, body weight and locomotor activity in rats. Br J Pharmacol 130:1305–1314. 10.1038/sj.bjp.0703443 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vickers S, Easton N, Webster L, Wyatt A, Bickerdike M, Dourish C, Kennett G (2003) Oral administration of the 5-HT 2C receptor agonist, mCPP, reduces body weight gain in rats over 28 days as a result of maintained hypophagia. Psychopharmacology 167:274–280. 10.1007/s00213-002-1378-6 [DOI] [PubMed] [Google Scholar]
- Watanabe Y, Yoshimoto K, Tatebe H, Kita M, Nishikura K, Kimura M, Tanaka MJ (2014) Enhancement of alcohol drinking in mice depends on alterations in RNA editing of serotonin 2C receptors. Int J Neuropsychopharmacol 17:739–751. 10.1017/S1461145713001545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weissman NJ, Sanchez M, Koch GG, Smith SR, Shanahan WR, Anderson CM (2013) Echocardiographic assessment of cardiac valvular regurgitation with lorcaserin from analysis of 3 phase 3 clinical trials. Circ Cardiovasc Imaging 6:560–567. 10.1161/CIRCIMAGING.112.000128 [DOI] [PubMed] [Google Scholar]
- World Health Organization (2016) Noncommunicable diseases: obesity among adults. Available from https://www.who.int/gho/ncd/risk_factors/overweight_obesity/obesity_adults/en/.
- Xu Y, Jones JE, Kohno D, Williams KW, Lee CE, Choi MJ, Anderson JG, Heisler LK, Zigman JM, Lowell BB (2008) 5-HT2CRs expressed by pro-opiomelanocortin neurons regulate energy homeostasis. Neuron 60:582–589. 10.1016/j.neuron.2008.09.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu Y, Jones JE, Lauzon DA, Anderson JG, Balthasar N, Heisler LK, Zinn AR, Lowell BB, Elmquist JK (2010) A serotonin and melanocortin circuit mediates D-fenfluramine anorexia. J Neurosci 30:14630–14634. 10.1523/JNEUROSCI.5412-09.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]