

Abstract
For a blood-borne cancer therapeutic agent to be effective, it must cross the blood vessel wall to reach cancer cells in adequate quantities, and it must overcome the resistance conferred by the local microenvironment around cancer cells. The brain microenvironment can thwart the effectiveness of drugs against primary brain tumours as well as brain metastases. In this Review, we highlight the cellular and molecular components of the blood–brain barrier (BBB), a specialized neurovascular unit evolved to maintain brain homeostasis. Tumours are known to compromise the integrity of the BBB, resulting in a vasculature known as the blood–tumour barrier (BTB), which is highly heterogeneous and characterized by numerous distinct features, including non-uniform permeability and active efflux of molecules. We discuss the challenges posed by the BBB and BTB for drug delivery, how multiple cell types dictate BBB function and the role of the BTB in disease progression and treatment. Finally, we highlight emerging molecular, cellular and physical strategies to improve drug delivery across the BBB and BTB and discuss their impact on improving conventional as well as emerging treatments, such as immune checkpoint inhibitors and engineered T cells. A deeper understanding of the BBB and BTB through the application of single-cell sequencing and imaging techniques, and the development of biomarkers of BBB integrity along with systems biology approaches, should enable new personalized treatment strategies for primary brain malignancies and brain metastases.
The blood–brain barrier (BBB) regulates homeostasis of the central nervous system (CNS) by forming a tightly regulated neurovascular unit (NVU) that includes endothelial cells (ECs), pericytes and astrocytic endfeet, which together maintain normal brain function1,2. However, these same features also hinder the delivery of systemic therapies into brain tumours. The BBB is disrupted during tumour progression and is then referred to as the blood–tumour barrier (BTB). Although the BTB is more permeable than the BBB, its heterogeneous permeability to small and large molecules as well as heterogeneous perfusion contributes to suboptimal drug accumulation in brain tumours3–6. As such, the BBB is one of the rate-limiting factors in clinically effective therapy.
In this Review, we discuss how the genotype of brain tumours impacts BBB and BTB (hereafter referred to as BBB/BTB) structure and function, and elaborate on the role of the BBB/BTB in disease progression and treatment. We highlight emerging minimally invasive strategies to improve drug delivery across the BBB/BTB and discuss their potential to improve treatment. Finally, we present emerging insights from systems biology approaches and discuss the impact of the BBB/BTB on immunotherapy, along with recent developments in biomarkers to assess extravasation across the BBB/BTB that should facilitate personalized therapy.
The neurovascular unit
As early as the late 1800s, several studies demonstrated that the CNS vasculature actively maintains CNS homeostasis by tightly regulating molecular and cellular transport across its specialized endothelium7. Preclinical and clinical observations in the following decades helped characterize the structure of capillary beds in the neuroparenchyma, which is composed of ECs connected by tight junctions (TJs), surrounded by a specialized basal lamina that is shared with pericytes and astrocytic endfeet, and sparsely interconnected by neuronal endings and microglia8. Together, these cells dictate the CNS endothelial structure and define the physical properties of the BBB (FIG. 1. Within these cellular and extracellular networks, specifically regulated transport systems allow efficient efflux of toxic cellular byproducts to move back into the circulation while permitting regulated influx of circulating molecules essential for CNS function. Although the CNS is considered immunologically unique, some populations of immune cells can ‘loosen’ and cross the BBB during neuroinflammation, whereas others can repair damaged nervous tissue9,10. Together, the BBB endothelium and neuroparenchymal cells constitute the NVU, which acts as a ‘gatekeeper’ within the CNS that tightly controls transcellular and paracellular crossing of molecules and cells10,11 (FIG. 1).
Fig. 1 |. Overview of the neurovascular unit in healthy and tumour-bearing brains.
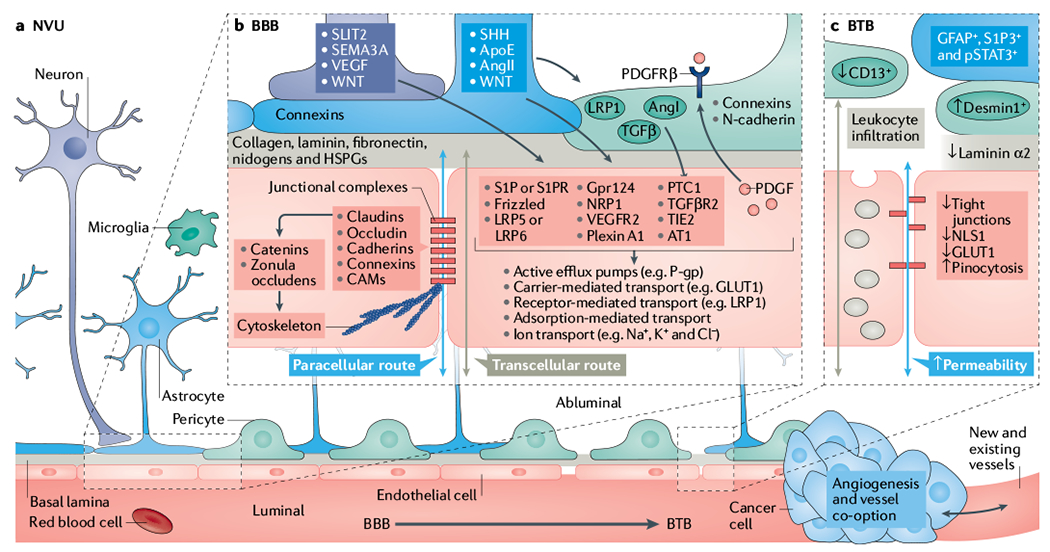
a | Schematic representation of capillaries in the neurovascular unit (NVU) with the intact blood–brain barrier (BBB, bottom left) and the disrupted blood–tumour barrier (BTB, bottom right) in the neuroparenchyma. BBB development and permeability is dictated by signalling and structural mechanisms that are regulated by multiple cells within the NVU. These mechanisms control both paracellular and transcellular routes and, ultimately, vessel permeability in the central nervous system (CNS). b | Schematic representation of notable cellular and molecular components that regulate the development, maturation and function of endothelial cells (ECs) (red) and the NVU (refer to main text for relevant references). Neuronal (purple) and non-neuronal cells regulate the expression of transport and tight junction proteins in ECs, which in turn may ‘loosen’ or ‘tighten’ the BBB. Here, we depict examples of key signalling pathways connecting astrocytes (blue), pericytes (green) and neurons to ECs. Together or individually, these pathways will alter transcellular transport by changing the expression of transporters and the paracellular route by disrupting junctional protein complexes. Of note, ECs reciprocally regulate components of the NVU. For example, EC-secreted transforming growth factor-β (TGFβ) can activate cognate receptor on pericytes. During development and maturation, glial cells, pericytes and neurons regulate EC behaviour via multiple ligands and receptors, which in turn activate downstream signalling cascades (for example, Frizzled, G protein-coupled receptor 124 (GPR124), β-catenin, GLI, PI3K, SRC and the p38 MAPK) that dictate expression of junctional and transcytosis proteins and control CNS homeostasis. For example, astrocytes directly modulate NVU demands such as water content in the neuroparenchymal space via the major water channel protein aquaporin 4 (AQP4), regulate immune cell and cancer cell infiltration via specific chemokine and cytokine production, regulate BBB permeability and integrity in part by angiotensin (AngI and AngII), apolipoprotein E (ApoE) and retinoic acid, and regulate pericyte distribution. c | In the BTB, NVU integrity and endothelial permeability is compromised due to disruption of the NVU, including displacement of astrocytes (blue) and pericytes (green), neurovascular decoupling, altered pericyte populations and changes in EC tight junctions and transcytosis mechanisms. Additional vascular-related phenotypes such as hypoxia, oedema, angiogenesis and tumour-vessel co-option can influence the NVU in brain tumours. Although BBB features remain present during tumour development, in particular at the cancer–neuroparenchyma edge, the BTB displays increased and heterogeneous permeability. Tumour progression leads to BTB structural changes including neuronal death, astrocyte endfeet displacement (from primary and metastatic cancer cells) and heterogenous pericyte and astrocyte subpopulations, all of which can reduce the barrier functions of the CNS endothelium. Intracellular vesicular transport is represented by grey-coloured vesicles in the schematic. AT1, angiotensin II receptor type 1; CAM, cell adhesion molecule; GLUT1, glucose transporter 1; HSPG, heparan sulfate proteoglycan; LRP, low-density lipoprotein receptor-related protein; NLS1, sodium-dependent lysophosphatidylcholine symporter 1; NRP1, neuropilin receptor 1; PDGF, platelet-derived growth factor; PDGFRβ, platelet-derived growth factor receptor β; P-gp, P-glycoprotein; PTC1, protein patched homologue 1; S1P, sphingosine-1-phosphate; S1PR, sphingosine-1-phosphate receptor; SEMA3A, semaphorin 3A; SHH, sonic hedgehog; SLIT2, Slit homologue 2 protein; TGFβR2, transforming growth factor-β receptor 2; TIE2, tyrosine kinase with Ig and EGF homology domains 2; VEGF, vascular endothelial growth factor; VEGFR2, vascular endothelial growth factor receptor 2.
Defects in BBB integrity during development are linked to several neurodegenerative disorders12. Investigation of the developmental expression and function of specific transcription factors, TJ proteins, angiogenic factors and growth factors during mouse development revealed that angiogenesis begins at embryonic day 9 (E9) and barriergenesis, formation of the BBB, ends at E15 (REFS13,14). The final steps of BBB maturation include a decrease in EC fenestrations and the appearance of TJs followed by a reduction in transcytosis. Although functional at birth, the BBB continues to mature postnatally under the influence of pericytes, astrocytes and neurons12. The early onset of the BBB is evident clinically as drug delivery of systemic therapies into the tumour microenvironment of paediatric brain tumours faces similar challenges to those observed in adults15,16.
Molecular profile
Genetic and transcriptomic studies have confirmed activation of signalling pathways, such as WNT–β-catenin and sonic hedgehog (SHH)-dependent signalling in brain ECs within the BBB8. Interestingly, some pathways — including the G protein-coupled receptor (GPCR) GPR124 and WNT–β-catenin axis — also regulate additional features of vascular architecture and function17–19. CNS EC signalling pathways can be directly regulated by pericytes and astrocytes (FIG. 1).
Recent single-cell analyses of the NVU have revealed previously unknown properties of ECs in the BBB. Single-cell RNA sequencing (scRNA-seq) of adult mouse NVU cells, including ECs, pericytes and glia, isolated by specific marker expression (for example, claudin 5 (CLDN5) for ECs and platelet-derived growth factor receptor β (PDGFRβ) for pericytes) has provided distinct cell-specific transcriptional profiles20. As expected, capillary ECs have high expression of nutrient transporters and TJ proteins (for example, CLDN5) when compared with arterial or venous ECs, whereas other factors such as von Willebrand factor (VWF) and vascular cell adhesion protein 1 (VCAM1) were inversely enriched. Interestingly, subpopulations of PDGFRα-positive perivascular fibroblast-like cells are present between the vessel wall and aquaporin 4-positive astrocytic endfeet throughout the CNS vasculature except for capillaries20. A separate single-cell transcriptional profiling study of different areas of the CNS has also confirmed unique molecular signatures of mural cells, fibroblast-like cells and ECs21. Another recent transcriptomic and epigenomic analysis compared ECs isolated from the brain, liver, lung and kidney of postnatal mice22. Brain ECs particularly expressed transporters including sodium-dependent lysophosphatidylcholine symporter 1 (NLS1; also known as MSFD2A), glucose transporter 1 (GLUT1) and solute carrier organic anion transporter family member 1C1 (SLCO1C1), which transport long-chain fatty acids, glucose and organic anions, respectively, across the BBB. By comparing CNS and non-CNS ECs, these authors observed CNS-specific transcription factor activity including T cell factor/lymphoid enhancer-binding factor (TCF/LEF) and the zinc finger protein ZIC3, which implies a role for canonical WNT signalling. Interestingly, there was heterogeneity among CNS capillary ECs reflecting either a more venous-like or more arterial-like transcriptional signature.
ECs are not the only NVU cells that are organotypic. Although pericytes share ubiquitous characteristics throughout the body, mouse brain pericytes express distinct sets of markers when compared with those isolated from the lung20. Current comprehensive single-cell analysis platforms and annotated databases of CNS and non-CNS capillary endothelium are providing organotypic signatures of NVU cells that will directly inform studies evaluating BBB biology and drug delivery across the NVU23,24. Performing single-cell analysis of the BTB during primary brain tumour progression and brain metastasis, and comparing it with normal BBB or extracranial primary endothelium in the case of brain metastases, will surely reveal more unique properties of the NVU and yield novel therapeutic strategies.
Reminiscent of the complex synaptic networks within the CNS, the BBB requires a co-dependent cellular structure to maintain normal function. The BBB can be mimicked ex vivo by co-culturing brain ECs, astrocytes and pericytes, allowing for experimental studies in vitro25. However, to what extent the full repertoire of BBB features, including the expression of BBB-specific transporters, is reproduced in in vitro models will require further analysis. For example, comparisons of the in vivo and in vitro BBB using scRNA-seq have not yet been published. CNS ECs are connected by TJs enriched in transmembrane cell-surface proteins including claudins, occludins and adhesion molecules26 (FIG. 1). Brain EC–cell junctions also contain vascular endothelial cadherin (VE-cadherin)-rich adherens junctions and connexin 43 (Cx43) gap junctions26. Together, these proteins regulate intracellular signalling, via zonula occludin proteins and β-catenin, and modulate actin cytoskeletal rearrangement to ‘tighten’ the extracellular space between ECs, thus preventing paracellular transport27 (FIG. 1).
Functional profile
The BBB is surrounded by the basal lamina, an extracellular matrix (ECM) formed predominantly of glycoproteins that can be proteolytically cleaved to influence BBB function in health and disease28. ECs, pericytes and astrocytes contribute to the basal lamina, which provides ligands that activate multiple signalling pathways in the NVU and regulate BBB function. CNS ECs are considered non-fenestrated, as they display a reduced number of pores in their cellular membrane, and have limited intracellular vesicular trafficking and pinocytosis29,30. Transcellular movement of cargo across the BBB takes the form of either vesicle-mediated transcytosis, involving receptor-mediated or adsorptive routes, or carrier-mediated transcytosis, which regulates transport of hormones, transferrin and plasma proteins. Passive diffusion across the BBB is largely dependent upon the lipophilicity of small molecules. Small lipophilic molecules such as oxygen and caffeine diffuse through cell membranes. The CNS endothelium displays polarized cellular transporters to dynamically regulate influx and efflux between the neuroparenchyma and blood (FIGS 1,2). Small hydrophilic molecules such as glucose and amino acids cross the BBB via solute carrier (SLC) proteins. Active transport occurs across the BBB endothelium by polarized expression of ATP-binding cassette transporters (ABC transporters) present on the luminal and abluminal sides of vessel walls31 (FIG. 2). These transporters mediate efflux of xenobiotics and toxins from the endothelium away from the neuroparenchymal space, effectively clearing these agents into the luminal compartment32. Most anti-neoplastic low molecular weight drugs are substrates for ABC proteins. For example, several targeted therapies for gliomas and brain metastases have affinity for the multidrug-resistant (MDR) ABC transporters, P-glycoprotein (P-gp; also known as ABCB1), breast cancer resistance protein (BCRP; also known as ABCG2) and multidrug resistance proteins (MRPs), which are all expressed in the BBB33. Furthermore, some transporters are also expressed in other NVU cells including astrocytes, microglia and neurons34. The above transport mechanisms form a delicate balance between influx and efflux routes that is essential for neuroparenchymal homeostasis. However, this homeostatic mechanism will also directly affect any therapeutic delivery.
Fig. 2 |. Physical and chemical properties of the BBB.
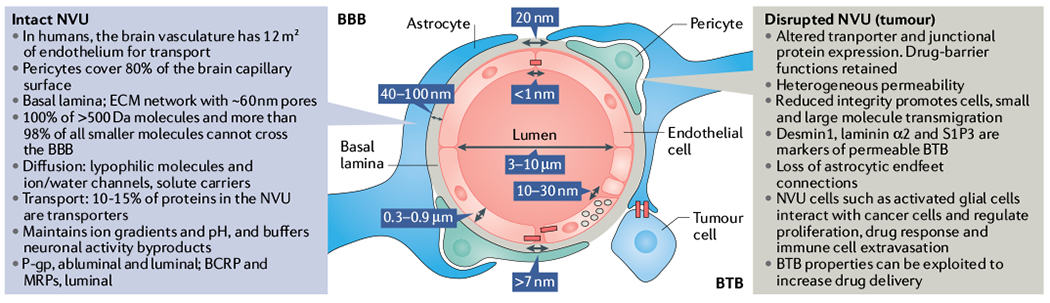
The blood–brain barrier (BBB) structure displays unique physical properties that tightly regulate molecular and cellular flow in the neuroparenchyma. The brain barrier’s importance is evident from its functional conservation across organisms, from fruit flies to humans1. As the BBB develops and matures, endothelial cell fenestrations decrease, and the subsequent appearance of tight junctions is followed by a reduction in transcytosis. Centre panel: cross-section of a central nervous system (CNS) capillary depicting estimated distances and spaces within the BBB, which circulating drugs are required to overcome to permeate the brain parenchyma. Left panel: in the non-diseased brain, the neurovascular unit (NVU) includes an intact BBB that displays multiple characteristics that limit drug permeability into the CNS. Right panel: during tumour progression, stroma–cancer cell interactions in the brain tumour microenvironment dictate vessel permeability and cancer cell proliferation. The blood–tumour barrier (BTB) characteristics listed here contribute to the heterogeneous permeability observed in the disrupted NVU. BCRP, breast cancer resistance protein; ECM, extracellular matrix; MRP, multidrug resistance protein; P-gp, P-glycoprotein; S1P3, sphingosine 1-phosphate 3.
Pericytes at the abluminal side of the endothelium are key regulators of vascular function throughout the body and control vessel function, vessel remodelling and neuroinflammation during ageing35–38. In addition to communicating with astrocytes within the NVU (FIG. 1), pericytes support BBB maintenance in the postnatal brain. Pericyte-deficient mutant mice showed increased BBB permeability to low and high molecular weight tracers36,38. Pericytes can directly regulate the expression of transporters. NLS1, which regulates transport of docosahexaenoic acid (DHA, an omega-3 fatty acid) across the BBB, is necessary for neuronal function and neuroprotection39. Loss of NLS1 expression in ECs leads to increased transcytosis and a leaky BBB. Pericytes wrap the CNS endothelium and together they produce a basal lamina that attracts astrocytic endfeet during development38,40. Astrocytes are the most abundant cell type in the brain, are metabolic sensors and play an integral role in BBB development and function41. Astrocytes appose their endfeet and cover the majority of the abluminal surface area of the BBB, and regulate signalling pathways that maintain junctional complexes and produce additional basal lamina. By connecting via gap junctions and TJs, astrocytes can form an additional barrier in the CNS called the glia limitans42,43. Interestingly, astrocytic endfeet removed by laser ablation repopulated blood vessels in 50% of cases44. Despite studies suggesting a key role for astrocytes in BBB function, this study showed limited BBB disruption after removing astrocytic input in the adult mouse brain. Together, astrocytes and pericytes play a plethora of direct and indirect roles in regulating BBB integrity during development, adulthood and disease progression.
BBB function is dynamic and can be influenced by local and circulating factors. Microglia are the most abundant innate immune cells in the neuroparenchyma and contribute to early CNS vasculogenesis45,46. Microglia activation can cause BBB disruption or repair and, during inflammation, microglia and peripheral immune cells such as leukocytes can increase BBB permeability through interleukin-1β (IL-1β) secretion or through the adhesion proteins intercellular adhesion molecule 1 (ICAM1), VCAM1 or E-selectin47. The BBB can also be innervated by synaptic endings. GABA-ergic, cholinergic, noradrenergic and serotonergic neurons can directly contact the CNS endothelium to regulate blood flow, neurovascular coupling and BBB permeability48,49. In sum, BBB development is modulated by intracranial cues, which control the underlying molecular pathways. In addition, the maintenance of BBB integrity is likely regulated on multiple fronts ranging from direct cellular contact to long-range factors including signalling proteins and molecules, cells and even exosomes, which may help extravasation into the neuroparenchyma50.
BBB heterogeneity in brain tumours
Owing to the confined space within the brain, tumour growth can impair blood flow even in peritumoral regions by compressing vessels51. Furthermore, as brain tumour lesions expand, the NVU gains different properties in the tumour core when compared with the periphery of the tumour and the neuroparenchyma, with the latter harbouring an intact BBB52. During primary brain tumour progression and the development of brain metastasis, the tumour vasculature becomes increasingly heterogeneous. An expanding neoplastic lesion causes local and distal changes that can directly compromise neuronal viability and vascular function51 (FIGS 1,2). As the vasculature dramatically changes during tumour expansion, nutritional demand of proliferating cancer cells requires co-opting existing vessels and/or creating new ones via angiogenesis. In addition, tumours can increase the blood vessel supply via other mechanisms: postnatal vasculogenesis, intussusception, vascular mimicry and transdifferentiation53,54.
Tumour vessels are tortuous and the architecture can differ between tumours of the brain and other tumour types4,55. Vascular dysfunction during tumour progression, in part mediated by deregulated expression of angiogenic factors such as vascular endothelial growth factor (VEGF), leads to hypoxia and an acidic microenvironment that fuels tumour progression in part through hypoxia-inducible factor 1α (HIF1α)-induced transcriptional programmes52,56,57. Blocking VEGF signalling transiently prunes the immature and leaky vessels of brain tumours in mice and actively remodels the remaining vasculature so that it more closely resembles the normal vasculature. We have shown the survival benefit of vascular normalization in patients with newly diagnosed as well as recurrent glioblastoma receiving antiangiogenic agents58,59. However, an adverse consequence of anti-angiogenic therapy, at least at high doses, is that the resulting hypoxia can increase invasiveness of cancer cells60. Furthermore, VEGF itself can regulate BBB permeability; therefore, anti-angiogenics at high doses might decrease BTB permeability, which could potentially affect delivery of other therapeutics61,62. Realizing an optimal balance between these vascular phenotypes remains a challenge for anti-angiogenic therapies of primary brain tumours and brain metastases.
The BTB
BBB disruption is evident from the higher drug accumulation within brain tumours as compared with the unaffected brain and, indirectly, by the detection of brain tumour markers in the circulation, such as circulating tumour cells from gliomas. The BTB is generally considered ‘leakier’ than the BBB (BOX 1). Imaging techniques including magnetic resonance imaging (MRI) and positron emission tomography (PET) have confirmed disruption of the BTB, in particular with high-grade brain tumours such as glioblastoma6. Despite these observations, it is clear that there can be an intact BBB in progressive glioblastomas displaying a range of efflux transporters that prevent the entry of anticancer agents6,63. The BTB is characterized by aberrant pericyte distribution and loss of astrocytic endfeet and neuronal connections64. Furthermore, invading glioma cells can physically displace astrocytic endfeet and disrupt BBB integrity65. T cell subpopulations and peripheral monocytes are detected in brain tumours, indicating permeability of the NVU to circulating immune cells66. Moreover, junctional proteins decrease in BTB ECs and the intratumoural vasculature never fully re-establishes a normal BBB in brain metastases67. Despite being characterized as a disrupted NVU, the BTB retains critical aspects of the BBB including expression of active efflux transporters in ECs and tumour cells63.
Box 1 |. Examples of BBB/BTB heterogeneity in preclinical and clinical studies.
Preclinical mouse models
Glioma
Delivery can vary greatly across a primary brain tumour lesion such that the centre of the tumour will demonstrate higher leakiness when compared with the peritumoral region and the surrounding brain microenvironment. In the intracranial GBM8401 glioma model, the distribution of liposomes containing doxorubicin is higher within the tumour when compared with the surrounding brain tissue; however, focused ultrasound further increases drug accumulation indicating that the blood–tumour barrier (BTB) is a compromised blood–brain barrier (BBB)197. Delivery of the targeted therapy dasatanib is heterogeneous within the tumour lesion and higher than the corresponding normal brain tissue in an oncogene-induced spontaneous model of malignant glioma198.
The orthotopic GL261 mouse glioblastoma model was studied using contrast-enhanced magnetic resonance imaging (MRI), Evans blue staining and histopathology. GL261 tumours displayed increased BBB disruption as they progressed, whereas the Hs683 human oligodendroglioma model had less disruption199,200. The U251 xenograft model has an intense rim on contrast-enhanced MRI representing vasogenic oedema (disrupted BBB and vasculature)201.
Medulloblastoma
All subtypes, with the exception of the WNT subtype, have an intact functional BBB in preclinical models and patients15. This could explain why the WNT subgroup has the best prognosis owing to improved drug delivery.
Brain metastasis
In the intracardiac injection (ICD) brain metastasis model using breast cancer cell lines 231-BR-Her2 and 4T1-BR5, BTB permeability is highly heterogeneous72. The 231-BR brain metastasis model displays areas of higher permeability due to sphingosine 1-phosphate receptor 3 (S1PR3)-positive astrocyte-dependent disruption of BTB integrity75.
In the ICD brain metastasis model using breast cancer cell lines SUM190-BR3 and JIMT-1-BR3, the BBB is more permeable than that with the 231-BR-Her2 cell line202.
In an intracranial injection brain metastasis model using the human breast cancer cell line BT474 (oestrogen receptor-positive, human epidermal growth factor receptor 2 (HER2)-amplified), the BBB is permeable to chemotherapy and antibodies but BBB/BTB disruption significantly increases delivery128. This preclinical model recapitulates clinically observed subpar central nervous system (CNS) delivery of targeted therapies such as lapatinib when compared with the site where the primary tumour arose203. Brain metastases formed from BT474 cells injected intracranially, intravenously or intracardially display some differences in the delivery of therapeutic nanoparticles. The intracranial model system is ‘leakier’ as more drug uptake and inhibition is observed204.
In the ICD brain metastasis model using the melanoma cell line A2058, there are regions with intact BBB112,205.
Intracranial brain metastasis models using melanoma cells (MDA-MB-435 and A2058) display a vessel co-option phenotype in the brain, whereas those using lung carcinoma cells (PC14-PE6 and HTB177) harbour more angiogenesis82.
In the intracranial brain metastasis model using the ALK-mutated lung adenocarcinoma cell line H3122 EML4-ALKL1196M, brain lesions respond to brain-penetrant ALK inhibitor (PF-06463922)206. These observations highlight the benefit of designing systemic therapies that bypass barriers of the neurovascular unit (NVU). Designing brain-permeable drugs is essential when targeting non-permeable micrometastases and macrometastases, for example, those observed in the ICD model of melanoma brain metastasis112.
Patient-derived xenograft models of brain metastases (from the breast, lung, melanoma, neuroendocrine, prostate) display reduced sodium-dependent lysophosphatidylcholine symporter 1 (NLS1) expression when compared with normal brain. Breast cancer brain metastases display heterogeneously disrupted BBB74.
Clinical data
Glioma
Glioblastomas (high-grade gliomas) display heterogeneous permeability to circulating drugs. Contrast-enhanced MRI of high-grade gliomas suggests there are areas of vasogenic oedema5,6. Together with positron emission tomography (PET) imaging and MRI, and analysis of surgically removed tissues, it has been shown that some regions of gliomas also display an intact BBB.
Contrast-enhanced MRI and immunohistochemistry (IHC) analyses (GFAP, AQ4, CD31) suggest that there are regions of intact BBB in low-grade and high-grade gliomas207.
Medulloblastoma
Contrast-enhanced MRI has revealed that the BBB/BTB is tumour subtype specific with high variability within the subgroups and heterogeneity across the tumour208,209. These observations highlight that genetic and molecular phenotypes may influence barrier function and dictate drug delivery.
Brain metastasis
BBB function is tumour subtype specific: triple-negative or basal-type tumours often disrupt the BBB, whereas HER2-positive tumours tend to preserve the BBB68. HER2-positive breast cancer brain metastases express high levels of glucose transporter 1 (GLUT1) and breast cancer resistance protein (BCRP) compared with brain metastases derived from other breast cancer subtypes.
Desmin-positive pericytes are present in specimens of human brain metastases. This phenotype correlates with areas of high permeability in preclinical models of breast cancer brain metastasis71.
Lapatinib concentration in breast cancer brain metastases ranges from 1.0 to 6.5 μM (REF.210). PET studies show higher levels of lapatinib in tumour lesion than normal brain211 and higher trastuzumab accumulation in breast cancer brain metastases212. Although tumour lesions maintain BBB features, the BTB ‘leakiness’ may allow more drug accumulation compared with unaffected brain parenchyma. Brain tumour permeability to targeting antibodies such as trastuzumab is evident in human and mouse preclinical orthotopic models of brain metastasis89,203,213,214.
Dynamic contrast-enhanced MRI has also been used to assess barrier integrity disruption in patients with brain metastases and revealed heterogeneity215.
BBB/BTB structural integrity is heterogeneous between metastatic lesions and between tumour types (BOX 1). For example, characteristics of BBB ECs such as fenestration differ between four molecular subtypes of medulloblastoma, thus directly impacting transcytosis of drugs across the BTB and therapeutic efficacy; the WNT subtype of medulloblastoma has a fenestrated vasculature, allowing higher accumulation of chemotherapeutic agents in the neuroparenchyma, whereas the SHH medulloblastoma subtype has an intact BBB15. The extent of BBB function and characteristics can vary among brain metastases from different subtypes of breast cancer68. Human epidermal growth factor receptor 2 (HER2, also known as ERBB2)-positive breast cancer brain metastases generally express more GLUT1 and BCRP than other subtypes. Although vessels are considered abnormal, the BTB may retain expression of efflux pumps such as BCRP.
In addition to genetic phenotypes of brain tumours, brain stroma signalling cascades that are important during barriergenesis can also influence progression of primary brain tumours. The Norrin–Frizzled 4–β-catenin signalling cascade (WNT pathway) can control tumour burden when activated in ECs69,70. Norrin–Frizzled 4 suppresses neoplastic progression of medulloblastoma by maintaining an antitumour stroma61. WNT signalling through the endothelial GPCR GPR124 regulates TJs and microvascular haemorrhage during glioblastoma progression in mice to prevent BBB dysfunction and oedema70. Although it is still unclear how molecular subtypes of gliomas might differentially dictate BTB function, emerging data are providing new insights. For example, oligodendrocyte transcription factor 2 (OLIG2)-positive precursor-like glioma cells co-opt blood vessels by activating WNT–β-catenin signalling17, which could influence BTB integrity.
BTB heterogeneity.
Elegant preclinical studies have shown that despite being leakier on average, the BTB also displays heterogeneous permeability (BOX 1). For example, low molecular weight compounds are unevenly distributed within the tumour lesion when injected systemically in mouse models of brain metastasis71,72. In preclinical models, the lipophilic low molecular weight inhibitor lapatinib displays increased heterogeneous distribution in HER2-positive breast cancer brain metastasis lesions when compared with the surrounding unaffected brain73. Molecular and cellular components of the NVU are disrupted in brain metastases. Pericyte subpopulations are altered in mouse and human brain metastasis, where desmin-positive pericytes largely populate the drug-permeable BTB when compared with unaffected vessel areas71. Reactive astrocytes lose connections to the ECs, reducing the expression of the lipid transporter NLS1 in ECs, which directly disrupts transport of DHA into the brain tumour microenvironment74. Furthermore, the sphingosine 1-phosphate receptor 3 (S1PR3) that is expressed in reactive astrocytes and brain metastatic cancer cells loosens the BTB via astrocytic IL-6 and CC-chemokine ligand 2 (CCL2) secretion75. EC S1PR1 signalling can also regulate BBB integrity within the mature CNS76. In glioblastoma, the BTB also displays abnormal barrier features such as reduced TJs, non-uniform pericyte vessel coverage and stem cell-derived pericytes, which disrupt vascular integrity during tumour progression64,77. Of particular note, glioma stem cells, which are considered refractory to therapies, reside in perivascular, hypoxic and invasive niches that maintain ‘sternness’ and also display abnormal barrier integrity78. Leakier and dysfunctional vessels cause water and metabolic waste retention in the neuroparenchymal space, increasing the interstitial and intracranial fluid pressure, often requiring glucocorticoid administration to reduce oedema in malignant brain tumours79. Interestingly, targeting glioma stem cell-derived pericytes in a preclinical model of glioma disrupts the BTB and enhances drug delivery. Furthermore, pericyte coverage is inversely correlated with the prognosis of patients with glioblastoma treated with chemotherapy80. Thus, the structural and functional heterogeneity of the BBB/BTB in the brain tumour microenvironment must be considered for developing effective systemic therapies.
Although primary brain tumours arise from CNS cells, metastatic cells must first cross the intact BBB. Elegant studies have demonstrated that specific circulating neoplastic cells express cell-surface markers that allow their extravasation into the brain parenchyma by facilitating transcellular transport81. Real-time imaging of a mouse brain metastasis model showed early extravasation and proximity to microvessels as a necessary element for colonization82. Metastatic cells that breach the BBB can disrupt CLDN5 TJs, and this can be reduced by blocking expression of angiopoietin 2 in ECs83. Once arrested within the CNS capillary bed, brain-tropic metastatic cells that successfully colonize the neuroparenchyma express cell-surface proteins, including ligands and proteases, which mediate extravasation across BBB and survival in the brain tumour microenvironment (FIG. 3). For example, expression of the 2,6-sialyltransferase ST6GALNAC5 on circulating metastatic breast cancer cells facilitates paracellular crossing of the BBB84. Brain metastatic cells can also express proteases such as cathepsin S to degrade the junctional adhesion molecule JAM2 (REF.85). Multiple steps during brain tumour progression, including the brain metastatic cascade and tumour expansion in the neuroparenchyma, alter BBB integrity, permeability and structural composition67. Although the BBB is regarded as the main barrier to successful therapy of brain tumours, the local microenvironment must be considered to improve patient survival86,87. Indeed, preclinical and clinical findings demonstrate that cancer cells that colonize the brain microenvironment display enhanced oncogenic and pro-survival signalling that can counteract the efficacy of targeted therapies and chemotherapies88–90.
Fig. 3 |. Improving drug delivery through the BBB/BTB.
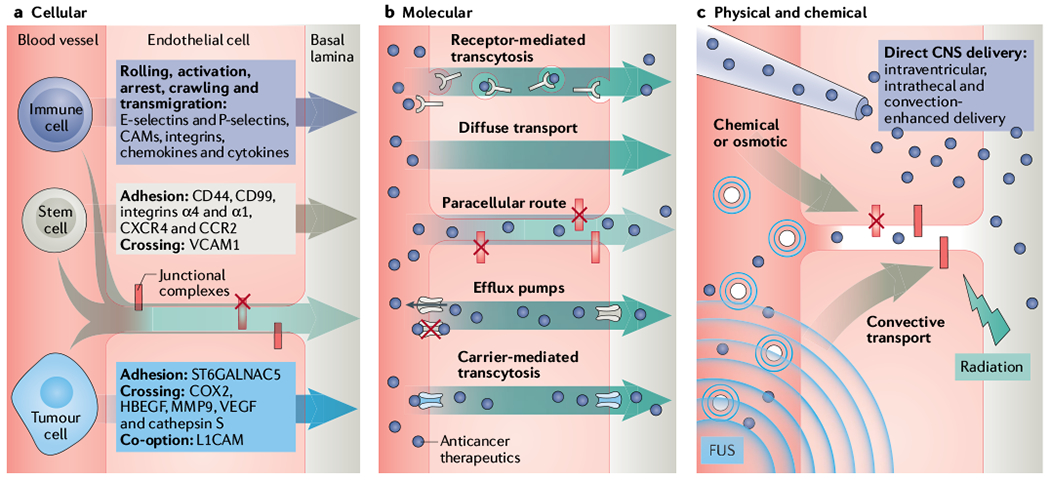
Schematic presentation of key molecular, cellular and physical mechanisms and systems to overcome the blood–brain barrier and blood–tumour barrier (BBB/BTB). a | The BBB prevents cellular extravasation into the neuroparenchyma unless compromised by circulating cells equipped with necessary ‘brain-tropic’ molecular components197, including soluble and membrane-bound proteins, to disturb the BBB integrity. Immune cell extravasation into the central nervous system (CNS) occurs by the following steps: rolling, activation, arrest, crawling, transmigration. The transcellular route is preferred when the BBB is intact, whereas the paracellular route is preferred when there is reduced tight junction integrity (red X = disrupted junction) and formation of intercellular gaps. Transmigration across the BBB is mediated by actin-containing protrusive structures and occurs on the timescale of minutes. Stem cells that have been engineered to contain anticancer cargo localize to sites of neuroinflammation and display coordinated rolling and adhesion behaviour, and transcellular and paracellular transmigration. In contrast to immune cells, mesenchymal stem cell (MSC) transmigration does not involve substantial lateral crawling. Stem cells migrate by the paracellular or transcellular route through discrete gaps or pores in the CNS endothelium. Stem cell transmigration is mediated by membrane blebbing and occurs on the timescale of hours. In contrast, circulating metastatic cancer cells that accumulate in the CNS capillary bed must express specific proteins in order to adhere and breach the BBB. Metastatic cells express proteases that disrupt junctional complexes. Although preclinical studies show that this process may occur within days, the time course of symptomatic brain metastasis in patients varies greatly (from months to years after metastatic dissemination of primary tumour). b | Several molecular strategies are employed to hijack or bypass barriers posed by the neurovascular unit (NVU) (described in main text and TABLE 1). Anticancer therapeutics (purple circles) can be designed with low affinity to efflux pumps or may hijack carriers or receptor-mediated transcytosis mechanisms. Alternatively, inhibitors of efflux pump or junctional complexes can be used to limit the clearance of drugs from the tumours. c | Direct delivery into the neuroparenchyma and physical disruption of the BBB/BTB. Specifically, focused ultrasound (FUS, indicated by the waves) with microbubbles (concentric small blue circles), radiation, osmotic, direct and convective-mediated delivery of therapeutics into the CNS is depicted. CAM, cell adhesion molecule; CCR2, CC-chemokine receptor 2; COX2, cyclooxygenase 2; CXCR4, CXC-chemokine receptor 4; HBECF, proheparin-binding ECF-like growth factor; L1CAM, L1 cell adhesion molecule; ST6GALNAC5, a 2,6-sialyltransferase; MMP9, matrix metalloproteinase 9; VCAM1, vascular cell adhesion protein 1; VECF, vascular endothelial growth factor.
Drug delivery across the BBB/BTB
Although essential for normal CNS function, the BBB represents a formidable barrier during therapeutic intervention. However, emerging insights into BBB/BTB structure and function have yielded novel strategies to overcome this barrier and deliver drugs to brain tumours, including those infiltrating the peritumoral regions. All the facets described above are directly or indirectly responsible for poor drug delivery within brain tumours. Circulating drugs are subject to multiple barriers posed by the NVU: reduced paracellular transport of hydrophilic molecules, reduced transcytosis and regulated polarized efflux transporters that prevent neuroparenchyma access to lipophilic synthetic molecules91. With the BBB intact, hydrophobic molecules with molecular weight <500 Da (smaller than 1 nm) diffuse transcellularly into the neuroparenchyma, whereas small hydrophilic compounds can enter the brain via the paracellular route92. However, many circulating pharmacological compounds have affinity for the multidrug resistance ABC transporters, as discussed above. As a result, ABC transporters are often responsible for decreasing the uptake rate of potential drugs crossing the BBB, as well as enhancing the barrier properties of the BTB53. The most common ABC transporters are P-gp, responsible for the poor penetration of large (>500 Da) hydrophobic drugs in the brain, the BCRP, conferring resistance to non-chemotherapeutic drugs and xenobiotics, and several members of the MDR ABC transporter family93–97.
Multiple strategies are being developed or optimized to bypass or hijack the cellular and molecular barriers of the BBB (FIG. 3). The various approaches to improve drug delivery across the BBB/BTB can be categorized as invasive or non-invasive, with minimally invasive included in the latter category. Although invasive approaches, which involve direct access to the disease site, have not lived up to their full potential, they are currently undergoing extensive refinements and optimization with promising preclinical findings (BOX 2). In addition to refining invasive methods, many minimally invasive (and non-invasive) approaches are currently in preclinical or early-stage clinical evaluation with encouraging findings (TABLE 1). Below, we will discuss these approaches.
Box 2 |. Invasive approaches to improve drug delivery to brain tumours.
Convection-enhanced delivery by direct injection
Direct intraparenchymal infusion of a drug via a cannula is used to bypass the blood–brain barrier (BBB) and promote bulk convective flow of the drug in the interstitial space. Although several phase I–III clinical trials have been carried out in patients with glioblastoma, the improvement in survival remains modest216. Optimization of the catheters and their placement, the administration protocol (for example, infusion rate and so on) and the physicochemical properties of the drug, along with methods to track it, may improve drug penetration and uptake217,218.
Intrathecal and intraventricular injection
The drug is administered to the lumbar subarachnoid space or ventricular system, which contains cerebrospinal fluid (CSF), and diffuses to the brain through the brain–CSF barrier. Intrathecal chemotherapy is currently the main treatment for patients with neoplastic meningitis219. Intrathecal or intraventricular administration delivers much higher doses to the leptomeningeal space than systemically administered chemotherapies219, but drug penetration to the brain parenchyma is limited220.
Implantation of wafers, gels and microchips
Macromolecules incorporated in biodegradable polymers are released by a combination of diffusion and polymer degradation. Microchips can provide even more control over the release. Biodegradable discs or wafers that contain the chemotherapy drug carmustine led to modest improvement in survival in patients with high-grade glioma221 but were also associated with high complication rates222. Current, preclinical investigations with injectable gel-like lipid capsules and microchips have shown promising results223–225. Evaluation of these methods in large animal models (for example, dogs with spontaneous gliomas) to demonstrate their improved ability to deliver drugs at larger distances, similar to those found in humans, is needed.
Table 1 |.
Emerging minimally invasive approaches to improve transport across the BBB/BTB
| Method | Route | Compound | Clinical trial | Main findings and comments | Refs |
|---|---|---|---|---|---|
| Molecular | |||||
| Receptor mediated | Transcellular (transcytosis) | LRP1-targeting peptide bound to paclitaxel | Phase II | Similar toxicity to paclitaxel alone, increased delivery and doubled the median survival in patients with breast cancer brain metastasis | 101,102 |
| Lipid soluble | Transcellular (diffusion) | Enzastaurin — a synthetic bisindolylmaleimide that targets/inhibits both the PKC and the PI3K–AKT pathways | Phase III | Well tolerated but did not have superior efficacy compared with chemotherapeutic agent lomustine in patients with recurrent glioblastoma | 226,227 |
| Evade active efflux | Transcellular | None | NA | None | |
| Tight junction pathways | Paracellular (diffusion) | None | NA | None | |
| Biological | |||||
| Cellular | Paracellular and transcellular (diapedesis) | NSCs expressing cytosine deaminase plus 5-fluorocytosine prodrug | Phase I | Therapy was well tolerated in trials against glioblastoma; intracerebral microdialysis revealed that NSCs produced 5-fluoruracil in the brain in a dose-dependent manner | 120 |
| Viral | Paracellular (diffusion) and transcellular (receptor-mediated endocytosis: infect and spread) | Adenovirus-mediated gene therapy | Phase III | Marginal improvement in overall survival in patients with newly diagnosed glioblastoma with more treatment-related adverse events, including hemiparesis and aphasia, as compared with the control group | 228 |
| Physical | |||||
| Focused ultrasound with microbubbles | Paracellular and transcellular (diffusion and convection) | Carboplatin | Phase II | Demonstrated safety and provided preliminary evidence of its efficacy in patients with glioblastoma | 138 |
| Radiation | Paracellular and transcellular | None | NA | Inconsistent results; more rigorous assessment on the direct versus indirect effects (for example, inflammation) of radiation on blood–brain barrier is needed | 154–156 |
| Nanoparticles | Paracellular (diffusion) and transcellular (transcytosis) | Liposomal doxorubicin plus radiotherapy and temozolomide | Phase II | Well tolerated but did not add any significant clinical benefit in patients with glioblastoma | 161–163 |
BBB, blood–brain barrier; BTB, blood–tumour barrier; LRP1, lipoprotein receptor-related protein 1; NA, not available; NSC, neural stem cell; PKC, protein kianse C.
Molecular approaches
Hijacking the endogenous influx transport.
Knowledge of the proteins expressed on ECs that line the BBB/BTB has been exploited for receptor-mediated BBB/BTB transcytosis (that is, a transcellular route)98. In this case, a targeting ligand (for example, a monoclonal antibody) binds to its receptor to trigger endocytosis, using the vesicular trafficking machinery to transport to the abluminal surface. A drug can be molecularly linked to this ligand, thus enabling transport across the BBB/BTB. Some well-characterized receptors that have been used for this approach include transferrin, insulin and insulin-like growth factor 1 receptors99,100. Although receptor-mediated transport is a very promising strategy for the delivery of macromolecular pharmaceuticals (up to 80 nm in diameter; see below for a discussion of nanoparticles)99 in the treatment of brain tumours, the widespread expression of these receptors in other tissues, the small dissociation rate and potential toxicity need to be carefully considered. Other targeted approaches take advantage of the endogenously expressed BBB/BTB receptor low-density lipoprotein receptor-related protein 1 (LRP1). LRP1-targeted peptide–chemotherapy conjugates, consisting of three paclitaxel molecules covalently linked to angiopep-2, a peptide designed to utilize the LRP1 transport system, increased delivery (>50-fold) into brain metastases and improved median survival in preclinical and clinical studies101,102.
Another approach is to employ the SLC proteins on the endothelial surface to cross the BBB/BTB. Examples include the glucose transporter GLUT1 that is responsible for the passive transport of glucose through the cell membrane and LAT1 that transports neutral amino acids bidirectionally between the blood and the brain93. Interestingly, overexpression of GLUT1 is correlated with poor survival in most solid tumours, including glioblastoma103, supporting further exploration of GLUT1 targeting to improve penetration across the BBB/BTB and enhance cancer cell uptake. Likewise, LAT1 could be targeted as its overexpression has been correlated with the malignant phenotype and proliferation of gliomas, although its function might be sensitive to hypoxia104.
Overcoming the efflux pumps.
Efflux pumps contribute to poor brain-to-blood drug ratios by actively pumping out pharmacological compounds with sufficient binding affinity. Efflux transporters including P-gp and BCRP have affinity for several FDA (US Food and Drug Administration)-approved drugs, including chemotherapeutics and various targeted therapeutics. They are often overexpressed in tumour vessels when compared with unaffected brain and keep drug levels well below the required effective dose in the brain tumour microenvironment105. ABC transporters (for example, P-gp and BCRP) are expressed in the BBB/BTB as well as in tumour cells of mouse and human brain tumours, which may exclude drugs from the cellular cytoplasm106–108.
To overcome efflux transporter-mediated drug reduction in the brain and reach sufficient bioactive concentrations in the brain tumour microenvironment, preclinical studies have focused on the co-administration of drugs with transporter inhibitors (for example, inhibitors of ABC transporters). This approach has shown significant enhancement in the brain concentrations of concurrently administered chemotherapeutic agents and targeted therapies (1.5-fold for temozolomide (TMZ)105, 5-fold for the poly(ADP ribose) polymerase (PARP) inhibitor ABT-888 (REF.106) and 40-fold for the mutant BRAF inhibitor vemurafenib109). Although the effectiveness of these strategies in the clinics has been questioned in the past110 (this was most likely due to methods used at that time), the increasingly recognized role of the efflux transporters in BBB and brain tumours combined with current knowledge31,111 suggests that more potent inhibitors with high specificity that concurrently target multiple pathways can lead to increased penetration of substrate drugs across the BBB/BTB31. In addition to blocking ABC transporters, structural refinement of molecularly targeted agents such as kinase inhibitors (for example, PI3K and mTOR inhibitors), with reduced affinity to P-gp and BCRP, is a promising alternative approach33,112. This approach may not only improve penetration across the BBB but also lead to improved cancer cell uptake106,113.
Cellular approaches
Harnessing the homing ability of certain stem cells is another exciting option for enabling therapeutic delivery across the BBB/BTB. These cells can disseminate in solid tumours, thus enabling site-specific delivery, and potentially circumvent the short half-lives that many chemotherapeutic agents exhibit by stably expressing or releasing various anticancer agents114.
A wide range of stem cell-based systems has been tested against brain malignancies; however, the high tumour tropism and ability to cross the BBB/BTB of neural stem cells (NSCs) and mesenchymal stem cells (MSCs) makes them the preferred carriers of therapeutics against primary brain tumours and brain metastases115,116. How stem cells migrate across the BBB is a controversial topic, but there are many reported similarities to immune cell infiltration (FIG. 3), including rolling on and adhesion to the endothelium and transmigration across the BBB. However, the specific molecular mechanisms might be different117,118. Importantly, stem cells can migrate along metastatic or invasive tumour borders, even when implanted intracranially at distant sites from the tumour, providing a promising platform to target brain micrometastases119. Although the reasons for the high tumour tropism of NSCs and MSCs are not yet fully elucidated, it is assumed that the chemoattractants and cytokines released by the tumour microenvironment play a central role114.
Current approaches for cell-mediated drug delivery involve the genetic modification of the carrier cell to secrete anticancer proteins, antiangiogenic factors or immunosupportive factors, such as IL-12 (REF.114). The NSC-mediated expression of an enzyme that converts a separately administered non-toxic pro-drug into a cytotoxic drug via the bystander effect is another approach that was also found to be well tolerated in a recent phase I trial in glioblastoma120. MSCs can shield oncolytic viruses from the host immune system to maintain long-term reservoirs of the therapeutic virus in the tumour microenvironment121. Nanoparticles can also be loaded into stem cells via several mechanisms including passive and/or caveolin-assisted or clathrin-assisted uptake122. When these cells reach the tumour, unloading takes place via exocytosis (passively or in response to external stimuli) or by membrane disruption due to stem cell death123.
Looking forward, analysis of stem cell migration kinetics and their short-term and long-term fate124, along with further understanding of the signalling pathways mediating stem cell migration and their interaction with cancer cells, may not only offer the means of increasing the migrating pool of stem cells but also further potentiate this promising therapeutic strategy.
Physical approaches: focused ultrasound
The combination of low-intensity focused ultrasound (FUS) pulses and circulating microbubbles — lipid, albumin or polymer-shelled gas pockets (1–10 μm) that scatter sound and vibrate in response to ultrasound waves — provides a physical method to transiently disrupt the BBB/BTB (for 6–24 h) and increase the permeability of the BBB/BTB125–127. Although the exact mechanism of action of FUS-mediated BBB/BTB disruption remains unknown, it appears that the forces exerted by the microbubbles in the brain vessels while they vibrate in response to ultrasound pressure waves (the microbubbles expand during peak negative pressures and contract during peak positive pressures) can change vessel permeability. Recent studies indicate that FUS and microbubbles can increase the BBB/BTB permeability to small molecules by 2–4-fold in tumours in rats and mice128,129, and experiments in the brains of healthy animals have shown that molecules with a diameter around 10 nm can readily extravasate130,131. Cellular and molecular evidence suggests that microbubble oscillation in the vessels can promote paracellular transport via transient reorganization of the TJs and facilitate transcellular passage through caveolae-dependent vesicular transport125. Although it is not well understood how the microbubble oscillation promotes the above changes, it appears that the microbubble dose and FUS exposure settings can be optimized to modulate transvascular transport, while limiting micro-haemorrhages and other adverse effects132,133. In addition to increasing BBB/BTB permeability125, FUS and microbubble-mediated BBB/BTB disruption induces a shift from diffusive transport to convective transport in the tumour interstitial space128. Due to their very low diffusivity, the transport of large molecules is primarily mediated by convection134, supporting the use of this technology with larger therapeutic agents, including antibodies (3–9 nm) and nanoparticles (10–60 nm)125.
Of interest, this minimally invasive method to disrupt the BBB/BTB can be repeated numerous times over a long period without any apparent functional deficit, delivering agents to white matter structures in Rhesus macaques135. Whereas FUS is a focal therapy, intraoperative MRI of the tumours combined with fast electronic beam steering can be used to target and treat a broad region of the brain135,136. Extensive preclinical research has shown that the method of FUS with microbubbles can lead to a more than 4-fold increase in the delivery and penetration of a range of intravenously administered anticancer agents in brain tumours125. This improvement in the delivery of anticancer agents has also led to a significant increase in the median survival time (3-fold) in multiple orthotopic murine tumour models, including glioma and breast cancer brain metastasis128. Numerous phase I clinical trials have confirmed the preclinical findings of increased BBB permeability by FUS in combination with microbubbles, demonstrated its safety and provided preliminary evidence for its efficacy137,138.
Identifying optimum drug combinations to use with FUS and microbubbles is a critical next step for evaluating the therapeutic efficacy of this method. Effective integration of quantitative imaging, molecular analysis, minimally invasive assays and physiologically accurate mathematical modelling should help realize this goal128. Moreover, methods for imaging the cerebrovascular microbubble dynamics — a highly non-linear and, to some extent, stochastic physical process — through the skull along with methods to control them (that is, promote stable oscillation) are important for safe and effective BBB/BTB disruption139–142.
Other strategies
Manipulate EC signalling pathways to increase BBB porosity.
Intravenously injected angubindin 1, a protein which binds to the tricellular TJ protein lipolysis-stimulated lipoprotein receptor (LSR)143, can increase the paracellular transport between brain microvascular ECs144. This increased transport across the BBB enabled the transient delivery of antisense oligonucleotides into the mouse brain that silenced a target RNA without any overt adverse effects144. Another approach includes the use of RNA interference (RNAi) to reduce the expression levels of TJ proteins and transiently modulate BBB permeability145. Although these methods have so far only been explored in the context of neurological disorders, they may provide a novel approach for targeting brain tumours and metastases. Moreover, exploiting the role of VEGF receptors to destabilize the TJs is an alternative method to increase BBB/BTB vessel permeability61. In addition to manipulating TJ pathways, modulating WNT–β-catenin signalling to induce fenestrations in ECs, as observed in the WNT subtype of medulloblastoma, could also provide a viable strategy to increase BBB/BTB leakiness146.
Radiation.
In addition to killing cancer cells, radiation is known to induce functional and structural changes in the tumour micreonvironment147. Recent investigations in glioma-bearing rats have demonstrated increased BBB/BTB permeability148–150, increased levels of an MRI contrast agent151 and increased nanoparticle delivery after radiation152 (dose >5 Gy; typical radiotherapy doses range from 30–50 Gy at 5–25 dose fractionation)153, implicating radiation in changes of BBB/BTB permeability151,152. These findings are corroborated by recent clinical trials showing increased levels of gefitinib in cerebrospinal fluid in a dose-dependent manner in patients with brain metastasis154. Although these findings have opened up the possibility that radiation therapy could also improve therapeutic delivery across the BBB/BTB, other studies have failed to reproduce these findings155. Further research is needed to resolve whether enhancement in therapeutic delivery indeed can occur with radiotherapy, and if so, whether it is driven by direct effects of radiation on the vasculature or by other effects of radiation on the microenvironment, such as radiation-induced (neuro) inflammation156 or other mechanisms.
Nanoparticles.
Several passive and active strategies (as discussed earlier and in FIG. 3) based on different nanoparticle formulations have been proposed for improving drug delivery across the BBB/BTB157–159. Unfortunately, nanoparticle accumulation in the brain and brain tumours is very low160, potentially explaining their limited therapeutic efficacy in clinical studies of high-grade gliomas161–163. Although several improvements in nanoparticle formulation for enhancing drug delivery in brain tumours have been proposed, most efforts either led to marginal improvements or were limited by the capabilities of the nanoparticle technology employed157–159. Although new and multifunctional nanoparticles are constantly being developed164, a transition from formulation-driven designs to tumour microenvironment-based development that takes into account BBB/BTB heterogeneity and tumour biology may provide an alternative path towards identifying optimum nanoparticle designs165–167. Such approaches would first establish the rules for design of BBB/BTB-penetrable nanoparticles, such as identifying the optimum nanoparticle size168 and shape165, and should then fuel technological developments that are more targeted to the brain tumour microenvironment. Tracking the nanoparticles and their payload is also important for gaining insights into nanoparticle penetration and cargo delivery to the brain tumour microenvironment, and further support the refinement of nanoparticle designs for maximum drug delivery to brain tumours167. In concert with improved nanoparticle designs and understanding of the mechanism for drug delivery, remodelling of the tumour microenvironment to promote nanoparticle accumulation169, potentially combined with triggered-release mechanisms and other strategies employed in extracranial malignancies170, may further improve the cargo delivery and penetration by supporting higher concentration gradients from the blood to the tumour core (that is, diffusive transport).
Future directions
BBB and immunotherapy
Although the BBB plays a key role in limiting antigen presentation and immune cell infiltration, both of which are immunological hallmarks of brain tumours and especially glioblastomas52,171,172, recent investigations have revealed potential strategies to alleviate its immunoprotective function. In this Review we focus only on the BBB/BTB, so for an extended review on brain cancer immunotherapy we direct the reader to a companion review in this issue by Sampson et al.173.
The compromised BBB may facilitate the presentation of tumour-associated antigens, as implied by a recent clinical trial showing improved responses when immune checkpoint inhibitors were administered before surgery in patients with glioblastoma174. Consequently, the physical approaches discussed above, such as FUS, might enhance antigen presentation as well as enable immune checkpoint inhibitor penetration across the BBB/BTB and increase immune cell infiltration to further sensitize brain tumours and micrometastases to immunotherapies175,176. Although these approaches seem promising, the relative contribution of reprogramming immune cells within the brain tumours versus systemically needs further exploration52. For example, the penetration of anti-PD1 immune checkpoint antibodies through the BBB/BTB in medulloblastomas in mice did not seem necessary for an effective antitumour response177. Nevertheless, there is a clear need for strategies that can deliver immunotherapeutic agents into tumours that have high immune cell infiltration (that is, CD8+ T cells) but no antitumour immunity and/or can improve T cell infiltration into the brain tumour microenvironment. Approaches that will limit or avoid the use of corticosteroids to control oedema178, which causes immunosuppression and reduces BBB/BTB permeability179,180, will also be crucial for effective immunotherapy of brain malignancies181.
Brain tumours and metastases also pose a substantial challenge for adoptively transferred T cell therapy and chimeric antigen receptor (CAR) T cell therapy182,183. T cell infiltration from the circulation into the brain involves several stages, including adhesion to the endothelium and transmigration across the BBB/BTB (FIG. 3). However, in brain tumours, such as glioblastoma, the ECs express little or no cell adhesion molecules52. Recent studies indicate that administration of CAR T cells in the circulation elicited inferior antitumour responses when compared with intracranial tumour infusions and local intracranial delivery184, further supporting the rate-limiting role of the BBB/BTB in T cell therapy. Hence, strategies that enhance the ability of T cells to penetrate the BBB/BTB185 and/or change the BBB/BTB phenotype to become more amenable to immune cell infiltration could lead to enhanced tumour control.
Biomarkers of BBB integrity
Assessment and quantification of BBB/BTB heterogeneity pre and post therapy to identify appropriate treatment windows has emerged as a clinical priority186. Several methods and biomarkers to assess extravasation across or disruption of the BBB have been proposed (summarized in TABLE 2). Contrast-enhanced MRI is the gold standard method for assessing BBB dysfunction. Most notably, Ktrans — a bulk transport parameter obtained using dynamic contrast-enhanced (DCE) MRI that is dependent on both capillary permeability and perfusion — provides a semi-quantitative assessment of BBB permeability. This parameter has been used to quantify changes in BBB/BTB permeability after anti-angiogenic therapy (drop in Ktrans)187 and characterize the effectiveness of targeted BBB/BTB disruption methods (increase in Ktrans)129. PET is another modality for delineating brain tumours and assessing the extent of BBB disruption. Whereas the most widely used PET tracers are those that penetrate the BBB (for example, 18F-fluorodeoxyglucose (FDG), O-(2-18F-fluoroethyl)-l-tyrosine (18F-FET)), 18F-3′-deoxy-3′-fluorothymidine (18F-FLT) might be of interest for assessing BBB integrity as FLT uptake is dependent on BBB integrity188.
Table 2|.
Biomarkers of BBB/BTB permeability
| Method | Agent or molecule | Measurement of BBB permeability | Clinical approval | Refs |
|---|---|---|---|---|
| Intravital microscopy | Fluorescent molecules | • Direct quantification of BBB/BTB permeability • Direct measurement of the extravasation of fluorescently labelled drugs and cells |
No | 128,229 |
| MRI (contrast enhanced) | Gadolinium or magnetic nanoparticles | • Semi-quantitative assessment of BBB permeability (Ktrans) • Indirect assessment of the extravasation of drugs that have similar permeability with gadolinium or magnetic nanoparticles • Overall limited correlation with the extravasation of drugs with different physiochemical properties from those of the agents |
Yes | 129,187 |
| PET | 18F | • Relative assessment of BBB permeability (extravasation of imaging tracer) • Direct measurement of radiolabelled drug and cell accumulation in the brain |
Yes | 230–233 |
BBB, blood–brain barrier; BTB, blood–tumour barrier; Ktrans, bulk transport parameter obtained using dynamic contrast-enhanced MRI that is dependent on both capillary permeability and perfusion; MRI, magnetic resonance imaging; PET, positron emission tomography.
Systems biology and the BBB
Although still in an early phase, systems biology approaches may provide a quantitative framework to meaningfully combine the increasing amount of data and information gathered during cancer diagnosis and lead to the development of rational strategies to overcome the BBB/BTB. Most notably, physiologically based pharmacokinetics modelling expanded to include new knowledge in cancer biology, including mechanics, blood vessel function and tissue oxygenation, can be used to identify effective new treatment regimens and drug combinations along with vascular remodelling strategies for effective delivery of nanomedicines169. Mathematical models combined with quantitative imaging can also be used to analyse drug transport across the BBB/BTB after modulating the paracellular and transcellular routes (for example, using FUS and microbubbles) to capture agent-specific transport parameters at the cellular level and optimize therapeutic interventions128. Moreover, by incorporating human-derived in vitro (that is, human cell lines) and ex vivo data on drug interaction with the efflux and uptake transporters and drug metabolism, model analysis can suggest in which patients or for which drugs the BBB/BTB may not pose major transport obstacles189. Direct assessment of drug concentrations via microdialysis methods or biopsy, during and after drug administration, respectively, may further increase the precision of these methods190,191. Likewise, data from The Cancer Genome Atlas may also be used to develop computational approaches to identify rate-limiting factors for effective antitumour immunity, including the type and abundance of immune cell populations that can cross the BBB/BTB, in order to inform effective cancer vaccine and immune checkpoint inhibitor therapies192. Incorporating data from in vivo measurements, including single cell profiles of the evolving NVU during primary brain tumour progression and metastasis, is a critical next step to improve these potentially personalized approaches.
Conclusion and perspectives
The BBB/BTB harbours considerable structural and functional heterogeneity within the microenvironment of the same lesion and across different cancer subtypes. Emerging insight into cellular, molecular and tumour subtype-specific features of the BBB/BTB has led to improved understanding of its role during tumour progression and treatment. These findings highlight the need to optimize and define tumour-specific therapeutic windows to disrupt CNS barriers and increase drug efficacy in the CNS with minimal side effects. Combining the strategies to improve penetration of anticancer agents across the BBB/BTB with biomarkers of BBB integrity may allow better delivery of therapeutics across the BBB/BTB and improved treatment outcomes.
Acknowledgements
The authors would like to thank C. Betsholtz, S. Chatterjee, M. Datta, Z. Amoozgar and S. Krishnan for their helpful input. C.D.A.’s research in this area is supported by NIH (National Institutes of Health) Grant R00EB016971 (National Institute of Biomedical Imaging and Bioengineering), NSF (National Science Foundation) Grants 1933158 and 1830577 (Leading Engineering for America’s Prosperity, Health, and Infrastructure) and CURE Childhood Cancer. G.B.F. received a fellowship from Susan G. Komen for the Cure. R.K.J.’s research in this general area is supported by an Outstanding Investigator Award from the National Cancer Institute (R35CA197743) and grants from the National Foundation for Cancer Research, the Ludwig Center at Harvard, the Advanced Medical Research Foundation, the Ellison Foundation, the Jane’s Trust Foundation and the Koch Institute-Dana Farber/Harvard Cancer Center Bridge Project Award.
Competing Interests
R.K.J. received honorarium from Amgen; consultant fees from Chugai, Enlight, Ophthotech, SPARC and SynDevRx; owns equity in Enlight, Ophthotech and SynDevRx; and serves on the Boards of Trustees of Tekla Healthcare Investors, Tekla Life Sciences Investors, Tekla Healthcare Opportunities Fund and Tekla World Healthcare Fund. The other authors declare no competing interests.
Glossary
- Pericytes
Mural cells that regulate development, permeability and function of the microcirculation.
- Astrocytic endfeet
Extensions of astrocytes into the perivascular space, covering the majority of the central nervous system endothelium and regulating development and function of the microcirculation.
- Microglia
Resident myeloid cells in the central nervous system (CNS) that regulate CNS function and homeostasis.
- Astrocytes
Major glial cells in the central nervous system (CNS) that regulate CNS function and homeostasis.
- Mural cells
Vascular-associated cells that include pericytes and smooth muscle cells.
- Pinocytosis
Small particles or molecules suspended in extracellular fluid that are brought into the cell through cell membrane invagination.
- ATP-binding cassette transporters
(ABC transporters). Active transporters that use energy in the form of ATP to transport molecules across the cell membrane against their concentration gradient.
- Diffusive transport
The net movement of molecules from a region of higher concentration to a region of lower concentration.
- Convective transport
Mass transport mediated by bulk fluid flow that is driven by a pressure gradient.
- Fast electronic beam steering
Changing the direction of the ultrasound focus or pattern by changing the relative phases of the radiofrequency signals driving tile transducer elements. Multiphase array transducers that are composed of hundreds of elements are able to perform electronic beam steering within a few microseconds.
Footnotes
Intriguingly, the recent discovery of bona fide functional synapses between gliomas and neurons that are associated with tumour proliferation and growth193,194 suggests that it is possible for brain tumours to hijack CNS pathways to control, and not necessarily disrupt, the NVU in order to support their metabolic demands and fuel tumour growth. Interestingly, another finding elegantly demonstrates that breast cancer cells that colonize the brain can replace astrocytes in tripartite synaptic complexes by forming pseudo-tripartite synapses with neurons195.
Another study published after this Review was accepted demonstrated that tumour-derived extracellular vesicles can breach the intact BBB in vivo by decreasing the expression of the late endosomal marker RAB7 in brain ECs and thereby increasing transcytosis196.
Peer review information
Nature Reviews Cancer thanks W. Elmquist, T. Phoenix and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
References
- 1.O’Brown NM, Pfau SJ & Gu C Bridging barriers: a comparative look at the blood–brain barrier across organisms. Genes Dev 32, 466–478 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]; This review describes the conservation and variation of BBB function across organisms.
- 2.Abbott NJ Blood–brain barrier structure and function and the challenges for CNS drug delivery. J. Inherit. Metab. Dis 36, 437–449 (2013). [DOI] [PubMed] [Google Scholar]
- 3.Hobbs SK et al. Regulation of transport pathways in tumor vessels: role of tumor type and microenvironment. Proc. Natl Acad. Sci. USA 95, 4607–4612 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study presents the first systematic examination of the role of the brain versus the subcutaneous microenvironment on permeability of tumour vessels.
- 4.Monsky WL et al. Role of host microenvironment in angiogenesis and microvascular functions in human breast cancer xenografts: mammary fat pad versus cranial tumors. Clin. Cancer Res 8, 1008–1013 (2002). [PubMed] [Google Scholar]; This study demonstrates the differential response of blood vessels in tumours growing in the brain versus in the subcutaneous microenvironment.
- 5.Pitz MW, Desai A, Grossman SA & Blakeley JO Tissue concentration of systemically administered antineoplastic agents in human brain tumors. J. Neurooncol 104, 629–638 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sarkaria JN et al. Is the blood–brain barrier really disrupted in all glioblastomas? A critical assessment of existing clinical data. Neuro. Oncol 20, 184–191 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Saunders NR et al. The rights and wrongs of blood–brain barrier permeability studies: a walk through 100 years of history. Front. Neurosci 8, 404 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Daneman R & Prat A The blood–brain barrier. Cold Spring Harb. Perspect. Biol 7, a020412 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dombrowski Y et al. Regulatory T cells promote myelin regeneration in the central nervous system. Nat. Neurosci 20, 674–680 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Prinz M & Priller J The role of peripheral immune cells in the CNS in steady state and disease. Nat. Neurosci 20, 136–144 (2017). [DOI] [PubMed] [Google Scholar]
- 11.Haddad-Tovolli R, Dragano NRV, Ramalho AFS & Velloso LA Development and function of the blood–brain barrier in the context of metabolic control. Front. Neurosci 11, 224 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sweeney MD, Zhao Z, Montagne A, Nelson AR & Zlokovic BV Blood–brain barrier: from physiology to disease and back. Physiol. Rev 99, 21–78 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhao Z, Nelson AR, Betsholtz C & Zlokovic BV Establishment and dysfunction of the blood–brain barrier. Cell 163, 1064–1078 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Møllgård K, Dziegielewska KM, Holst CB, Habgood MD & Saunders NR Brain barriers and functional interfaces with sequential appearance of ABC efflux transporters during human development. Sci. Rep 7, 11603 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Phoenix TN et al. Medulloblastoma genotype dictates blood brain barrier phenotype. Cancer Cell 29, 508–522 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]; This paper demonstrates that the WNT subtype of medulloblastoma produces antagonists of WNT7A that cause fenestrations in the ECs of the BBB.
- 16.Triarico S et al. Improving the brain delivery of chemotherapeutic drugs in childhood brain tumors. Cancers 11, E824 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Griveau A et al. A glial signature and Wnt7 signaling regulate glioma–vascular interactions and tumor microenvironment. Cancer Cell 33, 874–889.e7 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study shows the molecular mechanisms of vessel co-option versus angiogenesis by different subtypes of brain tumours.
- 18.Kuhnert F et al. Essential regulation of CNS angiogenesis by the orphan G protein-coupled receptor GPR124. Science 330, 985–989 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Umans RA et al. CNS angiogenesis and barriergenesis occur simultaneously. Dev. Biol 425, 101–108 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Vanlandewijck M et al. A molecular atlas of cell types and zonation in the brain vasculature. Nature 554, 475–480 (2018). [DOI] [PubMed] [Google Scholar]; Using vascular single-cell transcriptomics, this paper provides molecular definitions for the principal types of blood vascular and vessel-associated cells in the adult mouse brain.
- 21.Saunders A et al. Molecular diversity and specializations among the cells of the adult mouse brain. Cell 174, 1015–1030.e16 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sabbagh MF et al. Transcriptional and epigenomic landscapes of CNS and non-CNS vascular endothelial cells. eLife 7, e36187 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Khan S et al. EndoDB: a database of endothelial cell transcriptomics data. Nucleic Acids Res 47, D736–D744 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.He L et al. Single-cell RNA sequencing of mouse brain and lung vascular and vessel-associated cell types. Sci. Data 5, 180160 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kaisar MA et al. New experimental models of the blood–brain barrier for CNS drug discovery. Expert Opin. Drug Discov. 12, 89–103 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tietz S & Engelhardt B Brain barriers: crosstalk between complex tight junctions and adherens junctions. J. Cell Biol 209, 493–506 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Stamatovic SM, Johnson AM, Keep RF & Andjelkovic AV Junctional proteins of the blood–brain barrier: new insights into function and dysfunction. Tissue Barriers 4, e1154641 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Thomsen MS, Routhe LJ & Moos T The vascular basement membrane in the healthy and pathological brain. J. Cereb. Blood Flow Metab 37, 3300–3317 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ayloo S & Gu C Transcytosis at the blood–brain barrier. Curr. Opin. Neurobiol 57, 32–38 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chow BW & Gu C The molecular constituents of the blood–brain barrier. Trends Neurosci 38, 598–608 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Robey RW et al. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 18, 452 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Schinkel AH et al. Disruption of the mouse mdr1a P-glycoprotein gene leads to a deficiency in the blood-brain barrier and to increased sensitivity to drugs. Cell 77, 491–502 (1994). [DOI] [PubMed] [Google Scholar]
- 33.Oberoi RK et al. Strategies to improve delivery of anticancer drugs across the blood–brain barrier to treat glioblastoma. Neuro Oncol 18, 27–36 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sanchez-Covarrubias L, Slosky LM, Thompson BJ, Davis TP & Ronaldson PT Transporters at CNS barrier sites: obstacles or opportunities for drug delivery? Curr Pharm. Des 20, 1422–1449 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bell RD et al. Pericytes control key neurovascular functions and neuronal phenotype in the adult brain and during brain aging. Neuron 68, 409–427 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Armulik A, Genové G & Betsholtz C Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell 21 , 193–215 (2011). [DOI] [PubMed] [Google Scholar]
- 37.Daneman R, Zhou L, Kebede AA & Barres BA Pericytes are required for blood–brain barrier integrity during embryogenesis. Nature 468, 562–566 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Armulik A et al. Pericytes regulate the blood–brain barrier. Nature 468, 557–561 (2010). [DOI] [PubMed] [Google Scholar]
- 39.Ben-Zvi A et al. MSFD2A is critical for the formation and function of the blood–brain barrier. Nature 509, 507–511 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.ElAli A, Thériault P & Rivest S The role of pericytes in neurovascular unit remodeling in brain disorders. Int. J. Mol. Sci 15, 6453–6474 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Giaume C, Koulakoff A, Roux L, Holcman D & Rouach N Astroglial networks: a step further in neuroglial and gliovascular interactions. Nat. Rev. Neurosci 11, 87–99 (2010). [DOI] [PubMed] [Google Scholar]
- 42.Alvarez JI, Katayama T & Prat A Glial influence on the blood–brain barrier. Glia 61, 1939–1958 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Horng S et al. Astrocytic tight junctions control inflammatory CNS lesion pathogenesis. J. Clin. Invest 127, 3136–3151 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kubotera H et al. Astrocytic endfeet re-cover blood vessels after removal by laser ablation. Sci. Rep 9, 1263 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lannes N, Eppler E, Etemad S, Yotovski P & Filgueira L Microglia at center stage: a comprehensive review about the versatile and unique residential macrophages of the central nervous system. Oncotarget 8, 114393–114413 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shemer A, Erny D, Jung S & Prinz M Microglia plasticity during health and disease: an immunological perspective. Trends Immunol 36, 614–624 (2015). [DOI] [PubMed] [Google Scholar]
- 47.Sarelius IH & Glading AJ Control of vascular permeability by adhesion molecules. Tissue Barriers 3, e985954 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mizee MR & de Vries HE Blood–brain barrier regulation. Tissue Barriers 1, e26882 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Iadecola C The neurovascular unit coming of age: a journey through neurovascular coupling in health and disease. Neuron 96, 17–42 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Tominaga N et al. Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing blood–brain barrier. Nat. Commun. 6, 6716 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Seano G et al. Solid stress in brain tumours causes neuronal loss and neurological dysfunction and can be reversed by lithium. Nat. Biomed. Eng 3, 230 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Quail DF & Joyce JA The microenvironmental landscape of brain tumors. Cancer Cell 31, 326–341 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Carmeliet P & Jain RK Molecular mechanisms and clinical applications of angiogenesis. Nature 473, 298–307 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]; This review discusses how tumours acquire blood vessels by six different mechanisms, including angiogenesis and vessel co-option.
- 54.Voutouri C et al. Experimental and computational analyses reveal dynamics of tumor vessel cooption and optimal treatment strategies. Proc. Natl Acad. Sci. USA 116, 2662–2671 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study presents a systematic analysis of vessel co-option by tumours and how to overcome this barrier to increase drug delivery.
- 55.Vakoc BJ et al. Three-dimensional microscopy of the tumor microenvironment in vivo using optical frequency domain imaging. Nat. Med 15, 1219–1223 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Argaw AT et al. IL-1β regulates blood–brain barrier permeability via reactivation of the hypoxia-angiogenesis program. J. Immunol 177, 5574–5584 (2006). [DOI] [PubMed] [Google Scholar]
- 57.Engelhardt S, Patkar S & Ogunshola OO Cell-specific blood–brain barrier regulation in health and disease: a focus on hypoxia. Br. J. Pharmacol 171, 1210–1230 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Batchelor TT et al. Improved tumor oxygenation and survival in glioblastoma patients who show increased blood perfusion after cediranib and chemoradiation. Proc. Natl Acad. Sci. USA 110, 19059–19064 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Emblem KE et al. Vessel architectural imaging identifies cancer patient responders to anti-angiogenic therapy. Nat. Med 19, 1178–1183 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pàez-Ribes M et al. Antiangiogenic therapy elicits malignant progression of tumors to increased local invasion and distant metastasis. Cancer Cell 15, 220–231 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Argaw AT, Gurfein BT, Zhang Y, Zameer A & John GR VEGF-mediated disruption of endothelial CLN-5 promotes blood–brain barrier breakdown. Proc. Natl Acad. Sci. USA 106, 1977–1982 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Argaw AT et al. Astrocyte-derived VEGF-A drives blood–brain barrier disruption in CNS inflammatory disease. J. Clin. Invest 122, 2454–2468 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.van Tellingen O et al. Overcoming the blood–brain tumor barrier for effective glioblastoma treatment. Drug Resist. Updat 19, 1–12 (2015). [DOI] [PubMed] [Google Scholar]
- 64.Dubois LG et al. Gliomas and the vascular fragility of the blood–brain barrier. Front. Cell. Neurosci 8, 418 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Watkins S et al. Disruption of astrocyte–vascular coupling and the blood–brain barrier by invading glioma cells. Nat. Commun 5, 4196 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study systematically investigates the interaction of invading glioma cells with host cerebral blood vessels during the early stages of disease.
- 66.Ratnam NM, Gilbert MR & Giles AJ Immunotherapy in CNS cancers: the role of immune cell trafficking. Neuro Oncol 21,37–46 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Achrol AS et al. Brain metastases. Nat. Rev. Dis. Primers 5, 5 (2019). [DOI] [PubMed] [Google Scholar]
- 68.Yonemori K et al. Disruption of the blood–brain barrier by brain metastases of triple-negative and basal-type breast cancer but not HER2/neu-positive breast cancer. Cancer 116, 302–308 (2010). [DOI] [PubMed] [Google Scholar]
- 69.Bassett EA et al. Norrin/Frizzled4 signalling in the preneoplastic niche blocks medulloblastoma initiation. eLife 5, e16764 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Chang J et al. Gpr124 is essential for blood–brain barrier integrity in central nervous system disease. Nat. Med 23, 450 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lyle LT et al. Alterations in pericyte subpopulations are associated with elevated blood–tumor barrier permeability in experimental brain metastasis of breast cancer. Clin. Cancer Res 22, 5287–5299 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lockman PR et al. Heterogeneous blood–tumor barrier permeability determines drug efficacy in experimental brain metastases of breast cancer. Clin. Cancer Res 16, 5664–5678 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Taskar KS et al. Lapatinib distribution in HER2 overexpressing experimental brain metastases of breast cancer. Pharm. Res 29, 770–781 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Tiwary S et al. Metastatic brain tumors disrupt the blood–brain barrier and alter lipid metabolism by inhibiting expression of the endothelial cell fatty acid transporter Mfsd2a. Sci. Rep 8, 8267 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Gril B et al. Reactive astrocytic S1P3 signaling modulates the blood–tumor barrier in brain metastases. Nat. Commun 9, 2705 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Yanagida K et al. Size-selective opening of the blood–brain barrier by targeting endothelial sphingosine 1-phosphate receptor 1. Proc. Natl Acad. Sci. USA 114, 4531–4536 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cheng L et al. Glioblastoma stem cells generate vascular pericytes to support vessel function and tumor growth. Cell 153, 139–152 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hambardzumyan D & Bergers G Glioblastoma: defining tumor niches. Trends Cancer 1, 252–265 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Boucher Y, Salehi H, Witwer B, Harsh GR & Jain RK Interstitial fluid pressure in intracranial tumours in patients and in rodents. Br. J. Cancer 75, 829–836 (1997). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zhou W et al. Targeting glioma stem cell-derived pericytes disrupts the blood–tumor barrier and improves chemotherapeutic efficacy. Cell Stem Cell 21 , 591–603.e4 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Valiente M et al. The evolving landscape of brain metastasis. Trends Cancer 4, 176–196 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kienast Y et al. Real-time imaging reveals the single steps of brain metastasis formation. Nat. Med 16, 116–122 (2010). [DOI] [PubMed] [Google Scholar]
- 83.Avraham HK et al. Angiopoietin-2 mediates blood–brain barrier impairment and colonization of triple-negative breast cancer cells in brain. J. Pathol 232, 369–381 (2014). [DOI] [PubMed] [Google Scholar]
- 84.Bos PD et al. Genes that mediate breast cancer metastasis to the brain. Nature 459, 1005–1009 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sevenich L et al. Analysis of tumour- and stroma-supplied proteolytic networks reveals a brain-metastasis-promoting role for cathepsin S. Nat. Cell Biol 16, 876–888 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Kodack DP, Askoxylakis V, Ferraro GB, Fukumura D & Jain RK Emerging strategies for treating brain metastases from breast cancer. Cancer Cell 27, 163–175 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]; This review discusses recent advances in breast cancer brain metastasis therapy and potential approaches for successful treatment.
- 87.Kabraji S et al. Drug resistance in HER2-positive breast cancer brain metastases: blame the barrier or the brain? Clin Cancer Res 24, 1795–1804 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ni J et al. Combination inhibition of PI3K and mTORC1 yields durable remissions in mice bearing orthotopic patient-derived xenografts of HER2-positive breast cancer brain metastases. Nat. Med 22, 723–726 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Kodack DP et al. The brain microenvironment mediates resistance in luminal breast cancer to PI3K inhibition through HER3 activation. Sci. Transl Med 9, eaal4682 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]; This paper systematically investigates how the brain microenvironment confers resistance to low and high molecular weight therapeutics — targeted against oncogenic pathways — even when these therapeutics accrue within brain metastases.
- 90.Lin NU et al. Multicenter phase II study of lapatinib in patients with brain metastases from HER2-positive breast cancer. Clin. Cancer Res 15, 1452–1459 (2009). [DOI] [PubMed] [Google Scholar]
- 91.Banks WA From blood–brain barrier to blood–brain interface: new opportunities for CNS drug delivery. Nat. Rev. Drug Discov 15, 275–292 (2016). [DOI] [PubMed] [Google Scholar]
- 92.Banks WA Characteristics of compounds that cross the blood–brain barrier. BMC Neurol 9, S3 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Nałęcz KA Solute carriers in the blood–brain barrier: safety in abundance. Neurochem. Res 42, 795–809 (2017). [DOI] [PubMed] [Google Scholar]
- 94.Seelig A The role of size and charge for blood–brain barrier permeation of drugs and fatty acids. J. Mol. Neurosci 33, 32–41 (2007). [DOI] [PubMed] [Google Scholar]
- 95.Schinkel AH P-Glycoprotein, a gatekeeper in the blood–brain barrier. Adv. Drug Delivery Rev. 36, 179–194 (1999). [DOI] [PubMed] [Google Scholar]
- 96.Ni Z, Bikadi Z, Rosenberg MF & Mao Q Structure and function of the human breast cancer resistance protein (BCRP/ABCG2). Curr. Drug Metab 11, 603–617 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Smith MW & Gumbleton M Endocytosis at the blood–brain barrier: from basic understanding to drug delivery strategies. J. Drug Target 14, 191–214 (2006). [DOI] [PubMed] [Google Scholar]
- 98.Deeken JF & Löscher W The blood–brain barrier and cancer: transporters, treatment, and trojan horses. Clin. Cancer Res 13, 1663–1674 (2007). [DOI] [PubMed] [Google Scholar]
- 99.Lajoie JM & Shusta EV Targeting receptor-mediated transport for delivery of biologics across the blood–brain barrier. Annu. Rev. Pharmacol. Toxicol 55, 613–631 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Pardridge WM Drug transport across the blood–brain barrier. J. Cereb. Blood Flow Metab 32, 1959–1972 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Drappatz J et al. Phase I study of GRN1005 in recurrent malignant glioma. Clin. Cancer Res 19, 1567–1576 (2013). [DOI] [PubMed] [Google Scholar]
- 102.Tang S-C et al. ANG1005, a novel peptide-paclitaxel conjugate crosses the BBB and shows activity in patients with recurrent CNS metastasis from breast cancer, results from a phase II clinical study. Ann. Oncol 27, 103–113 (2016). [Google Scholar]
- 103.Wang J et al. Glucose transporter GLUT1 expression and clinical outcome in solid tumors: a systematic review and meta-analysis. Oncotarget 8, 16875–16886 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Youland RS et al. The role of LAT1 in 18F-DOPA uptake in malignant gliomas. J. Neurooncol 111, 11–18 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.de Gooijer MC et al. Improved brain penetration and antitumor efficacy of temozolomide by inhibition of ABCB1 and ABCG2. Neoplasia 20, 710–720 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lin F et al. ABCB1, ABCG2, and PTEN determine the response of glioblastoma to temozolomide and ABT-888 therapy. Clin. Cancer Res 20, 2703–2713 (2014). [DOI] [PubMed] [Google Scholar]
- 107.Wijaya J, Fukuda Y & Schuetz JD Obstacles to brain tumor therapy: key ABC transporters. Int. J. Mol. Sci 18, 2544 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Dréan A et al. ATP binding cassette (ABC) transporters: expression and clinical value in glioblastoma. J. Neurooncol 138, 479–486 (2018). [DOI] [PubMed] [Google Scholar]
- 109.Durmus S, Sparidans RW, Wagenaar E, Beijnen JH & Schinkel AH Oral availability and brain penetration of the B-RAFV600E inhibitor vemurafenib can be enhanced by the P-GLYCOprotein (ABCB1) and breast cancer resistance protein (ABCG2) inhibitor elacridar. Mol. Pharm 9, 3236–3245 (2012). [DOI] [PubMed] [Google Scholar]
- 110.Kalvass JC et al. Why clinical modulation of efflux transport at the human blood–brain barrier is unlikely: the ITC evidence-based position. Clin. Pharmacol. Ther 94, 80–94 (2013). [DOI] [PubMed] [Google Scholar]
- 111.Kim M et al. Barriers to effective drug treatment for brain metastases: a multifactorial problem in the delivery of precision medicine. Pharm. Res 35, 177 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study provides direct evidence of heterogeneity in the BBB/BTB and its impact on the therapeutic efficacy of small-molecule inhibitors.
- 112.Osswald M et al. Impact of blood–brain barrier integrity on tumor growth and therapy response in brain metastases. Clin. Cancer Res 22, 6078–6087 (2016). [DOI] [PubMed] [Google Scholar]
- 113.Agarwal S, Sane R, Oberoi R, Ohlfest JR & Elmquist W Delivery of molecularly targeted therapy to malignant glioma, a disease of the whole brain. Expert Rev. Mol. Med 13, e17 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Stuckey DW & Shah K Stem cell-based therapies for cancer treatment: separating hope from hype. Nat. Rev. Cancer 14, 683–691 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Bovenberg MSS, Degeling MH & Tannous BA Advances in stem cell therapy against gliomas. Trends Mol. Med 19, 281–291 (2013). [DOI] [PubMed] [Google Scholar]
- 116.Mooney R, Hammad M, Batalla-Covello J, Majid AA & Aboody KS Concise review: neural stem cell-mediated targeted cancer therapies. Stem Cells Transl Med 7, 740–747 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Liu L et al. From blood to the brain: can systemically transplanted mesenchymal stem cells cross the blood–brain barrier? Stem Cells Int. 2013, 435093 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Rüster B et al. Mesenchymal stem cells display coordinated rolling and adhesion behavior on endothelial cells. Blood 108, 3938–3944 (2006). [DOI] [PubMed] [Google Scholar]
- 119.Aboody KS et al. Neural stem cells display extensive tropism for pathology in adult brain: evidence from intracranial gliomas. Proc. Natl Acad. Sci. USA 97, 12846–12851 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]; This paper is the first to demonstrate that NSCs can migrate into brain tumours.
- 120.Portnow J et al. Neural stem cell-based anticancer gene therapy: a first-in-human study in recurrent high-grade glioma patients. Clin. Cancer Res 23, 2951–2960 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Sonabend AM et al. Mesenchymal stem cells effectively deliver an oncolytic adenovirus to intracranial glioma. Stem Cells 26, 831–841 (2008). [DOI] [PubMed] [Google Scholar]
- 122.Auffinger B et al. Drug-loaded nanoparticle systems and adult stem cells: a potential marriage for the treatment of malignant glioma? Oncotarget 4, 378–396 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Batrakova EV, Gendelman HE & Kabanov AV Cell-mediated drugs delivery. Expert Opin. Drug Deliv 8, 415–433 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Lee AS, Tang C, Rao MS, Weissman IL & Wu JC Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat. Med 19, 998–1004 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Aryal M, Arvanitis CD, Alexander PM & McDannold N Ultrasound-mediated blood–brain barrier disruption for targeted drug delivery in the central nervous system. Adv. Drug Deliv. Rev 72, 94–109 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Hynynen K, McDannold N, Vykhodtseva N & Jolesz FA Noninvasive MR imaging-guided focal opening of the blood–brain barrier in rabbits. Radiology 220, 640–646 (2001). [DOI] [PubMed] [Google Scholar]; This paper is the first to demonstrate that FUS with microbubbles can induce focal BBB disruption.
- 127.Meairs S Facilitation of drug transport across the blood–brain barrier with ultrasound and microbubbles. Pharmaceutics 7, 275–293 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Arvanitis CD et al. Mechanisms of enhanced drug delivery in brain metastases with focused ultrasound-induced blood–tumor barrier disruption. Proc. Natl Acad. Sci. USA 115, E8717–E8726 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]; This study demonstrates that BBB/BTB disruption mediated by FUS with microbubbles induces a shift from diffusive to convective transport in the tumour interstitial space.
- 129.Park J, Aryal M, Vykhodtseva N, Zhang Y-Z & McDannold N Evaluation of permeability, doxorubicin delivery, and drug retention in a rat brain tumor model after ultrasound-induced blood–tumor barrier disruption. J. Control. Release 250, 77–85 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Marty B et al. Dynamic study of blood–brain barrier closure after its disruption using ultrasound: a quantitative analysis. J. Cereb. Blood Flow Metab 32, 1948–1958 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Choi JJ, Wang S, Tung Y-S, Morrison B III & Konofagou EE Molecules of various pharmacologically-relevant sizes can cross the ultrasound-induced blood–brain barrier opening in vivo. Ultrasound Med. Biol 36, 58–67 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.McMahon D & Hynynen K Acute inflammatory response following increased blood–brain barrier permeability induced by focused ultrasound is dependent on microbubble dose. Theranostics 7, 3989–4000 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Kovacs ZI et al. Disrupting the blood–brain barrier by focused ultrasound induces sterile inflammation. Proc. Natl Acad. Sci. USA 114, E75–E84 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Schmidt MM & Wittrup KD A modeling analysis of the effects of molecular size and binding affinity on tumor targeting. Mol. Cancer Ther 8, 2861–2871 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.McDannold N, Arvanitis CD, Vykhodtseva N & Livingstone MS Temporary disruption of the blood–brain barrier by use of ultrasound and microbubbles: safety and efficacy evaluation in rhesus macaques. Cancer Res 72, 3652–3663 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Mainprize T et al. Blood–brain barrier opening in primary brain tumors with non-invasive MR-guided focused ultrasound: a clinical safety and feasibility study. Sci. Rep 9, 321 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Carpentier A et al. Clinical trial of blood–brain barrier disruption by pulsed ultrasound. Sci. Transl Med 8, 343re2 (2016). [DOI] [PubMed] [Google Scholar]
- 138.Idbaih A et al. Safety and feasibility of repeated and transient blood–brain barrier disruption by pulsed ultrasound in patients with recurrent glioblastoma. Clin. Cancer Res 25, 3793–3801 (2019). [DOI] [PubMed] [Google Scholar]
- 139.Arvanitis CD, Crake C, McDannold N & Clement GT Passive acoustic mapping with the angular spectrum method. IEEE Trans. Med. Imaging 36, 983–993 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Jones RM & Hynynen K Advances in acoustic monitoring and control of focused ultrasound-mediated increases in blood–brain barrier permeability. Br. J. Radiol 92, 20180601 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Patel A, Schoen SJ & Arvanitis CD Closed loop spatial and temporal control of cavitation activity with passive acoustic mapping. IEEE Trans. Biomed. Eng 66, 2022–2031 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Sun T et al. Closed-loop control of targeted ultrasound drug delivery across the blood–brain/tumor barriers in a rat glioma model. Proc. Natl Acad. Sci. USA 114, E10281–E10290 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Krug SM et al. Angubindin-1, a novel paracellular absorption enhancer acting at the tricellular tight junction. J. Control. Release 260, 1–11 (2017). [DOI] [PubMed] [Google Scholar]
- 144.Zeniya S et al. Angubindin-1 opens the blood–brain barrier in vivo for delivery of antisense oligonucleotide to the central nervous system. J. Control. Release 283, 126–134 (2018). [DOI] [PubMed] [Google Scholar]
- 145.Greene C & Campbell M Tight junction modulation of the blood brain barrier: CNS delivery of small molecules. Tissue Barriers 4, e1138017 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Guerit S & Liebner S Blood–brain barrier breakdown determines differential therapeutic outcome in genetically diverse forms of medulloblastoma. Cancer Cell 29, 427–429 (2016). [DOI] [PubMed] [Google Scholar]
- 147.Stapleton S, Jaffray D & Milosevic M Radiation effects on the tumor microenvironment: implications for nanomedicine delivery. Adv. Drug Deliv. Rev 109, 119–130 (2017). [DOI] [PubMed] [Google Scholar]
- 148.Bouchet A et al. Permeability of brain tumor vessels induced by uniform or spatially microfractionated synchrotron radiation therapies. Int. J. Radiat. Oncol. Biol. Phys 98, 1174–1182 (2017). [DOI] [PubMed] [Google Scholar]
- 149.Teng F, Tsien CI, Lawrence TS & Cao Y Blood–tumor barrier opening changes in brain metastases from pre to one-month post radiation therapy. Radiother. Oncol 125, 89–93 (2017). [DOI] [PubMed] [Google Scholar]
- 150.Crowe W et al. MRI evaluation of the effects of whole brain radiotherapy on breast cancer brain metastasis. Int. J. Radiat. Biol 95, 338–346 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Lemasson B et al. Monitoring blood–brain barrier status in a rat model of glioma receiving therapy: dual injection of low-molecular-weight and macromolecular MR contrast media. Radiology 257, 342–352 (2010). [DOI] [PubMed] [Google Scholar]
- 152.Erel-Akbaba G et al. Radiation-induced targeted nanoparticle-based gene delivery for brain tumor therapy. ACS Nano 13, 4028–4040 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Mann J, Ramakrishna R, Magge R & Wernicke AG Advances in radiotherapy for glioblastoma. Front. Neurol 8, 748 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Zeng Y et al. Blood–brain barrier permeability of gefitinib in patients with brain metastases from non-small-cell lung cancer before and during whole brain radiation therapy. Oncotarget 6, 8366–8376 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Fang L et al. Whole-brain radiation fails to boost intracerebral gefitinib concentration in patients with brain metastatic non-small cell lung cancer: a self-controlled, pilot study. Cancer Chemother. Pharmacol 76, 873–877 (2015). [DOI] [PubMed] [Google Scholar]
- 156.Miller MA et al. Radiation therapy primes tumors for nanotherapeutic delivery via macrophage-mediated vascular bursts. Sci. Transl Med 9, eaal0225 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Kreuter J Drug delivery to the central nervous system by polymeric nanoparticles: what do we know? Adv. Drug Deliv. Rev 71, 2–14 (2014). [DOI] [PubMed] [Google Scholar]
- 158.Tapeinos C, Battaglini M & Ciofani G Advances in the design of solid lipid nanoparticles and nanostructured lipid carriers for targeting brain diseases. J. Control. Release 264, 306–332 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Bertrand N, Wu J, Xu X, Kamaly N & Farokhzad OC Cancer nanotechnology: the impact of passive and active targeting in the era of modern cancer biology. Adv. Drug Deliv. Rev 66, 2–25 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Wilhelm S et al. Analysis of nanoparticle delivery to tumours. Nat. Rev. Mater 1, 16014 (2016). [Google Scholar]
- 161.Hau P et al. Pegylated liposomal doxorubicin-efficacy in patients with recurrent high-grade glioma. Cancer 100, 1199–1207 (2004). [DOI] [PubMed] [Google Scholar]
- 162.Beier CP et al. RNOP-09: pegylated liposomal doxorubicine and prolonged temozolomide in addition to radiotherapy in newly diagnosed glioblastoma — a phase II study. BMC Cancer 9, 308 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Ananda S et al. Phase 2 trial of temozolomide and pegylated liposomal doxorubicin in the treatment of patients with glioblastoma multiforme following concurrent radiotherapy and chemotherapy. J. Clin. Neurosci. 18, 1444–1448 (2011). [DOI] [PubMed] [Google Scholar]
- 164.Johnsen KB et al. Modulating the antibody density changes the uptake and transport at the blood–brain barrier of both transferrin receptor-targeted gold nanoparticles and liposomal cargo. J. Control. Release 295, 237–249 (2019). [DOI] [PubMed] [Google Scholar]
- 165.Chauhan VP & Jain RK Strategies for advancing cancer nanomedicine. Nat. Mater 12, 958–962 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Hare JI et al. Challenges and strategies in anticancer nanomedicine development: an industry perspective. Adv. Drug Deliv. Rev 108, 25–38 (2017). [DOI] [PubMed] [Google Scholar]
- 167.Stylianopoulos T & Jain RK Design considerations for nanotherapeutics in oncology. Nanomedicine 11, 1893–1907 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Thorne RG & Nicholson C In vivo diffusion analysis with quantum dots and dextrans predicts the width of brain extracellular space. Proc. Natl Acad. Sci. USA 103, 5567–5572 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Stylianopoulos T, Munn LL & Jain RK Reengineering the physical microenvironment of tumors to improve drug delivery and efficacy: from mathematical modeling to bench to bedside. Trends Cancer 4, 292–319 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Lyon PC et al. Safety and feasibility of ultrasound-triggered targeted drug delivery of doxorubicin from thermosensitive liposomes in liver tumours (TARDOX): a single-centre, open-label, phase 1 trial. Lancet Oncol 19, 1027–1039 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Lim M, Xia Y, Bettegowda C & Weller M Current state of immunotherapy for glioblastoma. Nat. Rev. Clin. Oncol 15, 422–442 (2018). [DOI] [PubMed] [Google Scholar]
- 172.Thorsson V et al. The immune landscape of cancer. Immunity 48, 812–830.e14 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Sampson J Brain immunology and immunotherapy in brain tumours. Nat. Rev. Cancer (in the press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Cloughesy TF et al. Neoadjuvant anti-PD-1 immunotherapy promotes a survival benefit with intratumoral and systemic immune responses in recurrent glioblastoma. Nat. Med 25, 477 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Sevenich L Turning “cold” into “hot” tumors—opportunities and challenges for radio-immunotherapy against primary and metastatic brain cancers. Front. Oncol 9, 163 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Curley CT, Sheybani ND, Bullock TN & Price RJ Focused ultrasound immunotherapy for central nervous system pathologies: challenges and opportunities. Theranostics 7, 3608–3623 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Pham CD et al. Differential immune microenvironments and response to immune checkpoint blockade among molecular subtypes of murine medulloblastoma. Clin. Cancer Res 22, 582–595 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Okada H et al. Immunotherapy response assessment in neuro-oncology: a report of the RANO working group. Lancet Oncol 16, e534–e542 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Kamoun WS et al. Edema control by cediranib, a vascular endothelial growth factor receptor–targeted kinase inhibitor, prolongs survival despite persistent brain tumor growth in mice. J. Clin. Oncol 27, 2542–2552 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Stegmayr C et al. Influence of blood–brain barrier permeability on O-(2-18F-fluoroethyl)-l-tyrosine uptake in rat gliomas. Eur. J. Nucl. Med. Mol. Imaging 44, 408–416 (2017). [DOI] [PubMed] [Google Scholar]
- 181.Postow MA, Sidlow R & Hellmann MD Immune-related adverse events associated with immune checkpoint blockade. N. Engl. J. Med 378, 158–168 (2018). [DOI] [PubMed] [Google Scholar]
- 182.Platten M & Reardon DA Concepts for immunotherapies in gliomas. Semin. Neurol 38, 62–72 (2018). [DOI] [PubMed] [Google Scholar]
- 183.Brown CE et al. Regression of glioblastoma after chimeric antigen receptor T-cell therapy. N. Engl. J. Med 375, 2561–2569 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Brown CE et al. Optimization of IL13Rα2-targeted chimeric antigen receptor T cells for improved antitumor efficacy against glioblastoma. Mol. Ther 26, 31–44 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Kebir H et al. Human TH17 lymphocytes promote blood–brain barrier disruption and central nervous system inflammation. Nat. Med 13, 1173 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Levin VA et al. CNS anticancer drug discovery and development conference white paper. Neuro Oncol 17, vi1–vi26 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Sorensen AG et al. A “vascular normalization index” as potential mechanistic biomarker to predict survival after a single dose of cediranib in recurrent glioblastoma patients. Cancer Res 69, 5296–5300 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Langen K-J, Galldiks N, Hattingen E & Shah NJ Advances in neuro-oncology imaging. Nat. Rev. Neurol 13, 279–289 (2017). [DOI] [PubMed] [Google Scholar]
- 189.Li J et al. Quantitative and mechanistic understanding of AZD1775 penetration across human blood–brain barrier in glioblastoma patients using an IVIVE–PBPK modeling approach. Clin. Cancer Res 23, 7454–7466 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Lee EQ et al. Phase I and biomarker study of plerixafor and bevacizumab in recurrent high-grade glioma. Clin. Cancer Res. 24, 4643–4649 (2018). [DOI] [PubMed] [Google Scholar]
- 191.Kennedy RT Emerging trends in in vivo neurochemical monitoring by microdialysis. Curr. Opin. Chem. Biol 17, 860–867 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Li B et al. Comprehensive analyses of tumor immunity: implications for cancer immunotherapy. Genome Biol 17, 174 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Venkataramani V et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature 573, 532–538 (2019). [DOI] [PubMed] [Google Scholar]
- 194.Venkatesh HS et al. Electrical and synaptic integration of glioma into neural circuits. Nature 573, 539–545 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Zeng Q et al. Synaptic proximity enables NMDAR signalling to promote brain metastasis. Nature 573, 526–531 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Morad G et al. Tumor-derived extracellular vesicles breach the intact blood–brain barrier via transcytosis. ACS Nano 10.1021/acsnano.9b04397 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Yang F-Y et al. Focused ultrasound and interleukin-4 receptor-targeted liposomal doxorubicin for enhanced targeted drug delivery and antitumor effect in glioblastoma multiforme. J. Control. Release 160, 652–658 (2012). [DOI] [PubMed] [Google Scholar]
- 198.Agarwal S et al. Active efflux of dasatinib from the brain limits efficacy against murine glioblastoma: broad implications for the clinical use of molecularly targeted agents. Mol. Cancer Ther 11,2183–2192 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Cha S et al. Dynamic, contrast-enhanced perfusion MRI in mouse gliomas: correlation with histopathology. Magn. Reson. Med 49, 848–855 (2003). [DOI] [PubMed] [Google Scholar]
- 200.Leten C, Struys T, Dresselaers T & Himmelreich U In vivo and ex vivo assessment of the blood–brain barrier integrity in different glioblastoma animal models. J. Neurooncol 119, 297–306 (2014). [DOI] [PubMed] [Google Scholar]
- 201.Jacobs VL, Valdes PA, Hickey WF & De Leo JA Current review of in vivo GBM rodent models: emphasis on the CNS-1 tumour model. ASN Neuro 3, e00063 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Murrell DH et al. Understanding heterogeneity and permeability of brain metastases in murine models of HER2-positive breast cancer through magnetic resonance imaging: implications for detection and therapy. Transl Oncol 8, 176 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Kodack DP et al. Combined targeting of HER2 and VEGFR2 for effective treatment of HER2-amplified breast cancer brain metastases. Proc. Natl Acad. Sci. USA 109, E3119–E3127 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Wyatt EA & Davis ME Method of establishing breast cancer brain metastases affects brain uptake and efficacy of targeted, therapeutic nanoparticles. Bioeng. Transl Med. 4, 30–37 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Henry MN, Chen Y, McFadden CD, Simedrea FC & Foster PJ In-vivo longitudinal MRI study: an assessment of melanoma brain metastases in a clinically relevant mouse model. Melanoma Res 25, 127–137 (2015). [DOI] [PubMed] [Google Scholar]
- 206.Zou HY et al. PF-06463922, an ALK/ROS1 inhibitor, overcomes resistance to first and second generation ALK inhibitors in preclinical models. Cancer Cell 28, 70–81 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Nduom EK, Yang C, Merrill MJ, Zhuang Z & Lonser RR Characterization of the blood–brain barrier of metastatic and primary malignant neoplasms: laboratory investigation. J. Neurosurg 119, 427 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Perreault S et al. MRI surrogates for molecular subgroups of medulloblastoma. Am. J. Neuroradiol 35, 1263–1269 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Dangouloff-Ros V et al. Imaging features of medulloblastoma: conventional imaging, diffusion-weighted imaging, perfusion-weighted imaging, and spectroscopy: from general features to subtypes and characteristics. Neurochirurgie S0028-3770, 30178–30179 (2018). [DOI] [PubMed] [Google Scholar]
- 210.Morikawa A et al. Capecitabine and lapatinib uptake in surgically resected brain metastases from metastatic breast cancer patients: a prospective study. Neuro Oncol 17, 289–295 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Saleem A et al. Lapatinib access into normal brain and brain metastases in patients with Her-2 overexpressing breast cancer. EJNMMI Res 5, 30 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Tamura K et al. 64Cu-DOTA-trastuzumab PET imaging in patients with HER2-positive breast cancer. J. Nucl. Med 54, 1869–1875 (2013). [DOI] [PubMed] [Google Scholar]
- 213.Lewis GP et al. Trastuzumab uptake and its relation to efficacy in an animal model of HER2-positive breast cancer brain metastasis. Breast Cancer Res. Treat 164, 581–591 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Askoxylakis V et al. Preclinical efficacy of ado-trastuzumab emtansine in the brain microenvironment. J. Natl Cancer Inst 108, djv313 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Heye AK, Culling RD, Valdés CHM, Thrippleton MJ & Wardlaw JM Assessment of blood–brain barrier disruption using dynamic contrast-enhanced MRI. A systematic review. Neuroimage Clin 6, 262–274 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Jahangiri A et al. Convection-enhanced delivery in glioblastoma: a review of preclinical and clinical studies. J. Neurosurg 126, 191–200 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Lonser RR, Sarntinoranont M, Morrison PF & Oldfield EH Convection-enhanced delivery to the central nervous system. J. Neurosurg 122, 697–706 (2015). [DOI] [PubMed] [Google Scholar]
- 218.Zhou Z, Singh R & Souweidane MM Convection-enhanced delivery for diffuse intrinsic pontine glioma treatment. Curr. Neuropharmacol 15, 116–128 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Beauchesne P Intrathecal chemotherapy for treatment of leptomeningeal dissemination of metastatic tumours. Lancet Oncol 11, 871–879 (2010). [DOI] [PubMed] [Google Scholar]
- 220.Groothuis DR The blood–brain and blood–tumor barriers: a review of strategies for increasing drug delivery. Neuro Oncol 2, 45–49 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Chowdhary SA, Ryken T & Newton HB Survival outcomes and safety of carmustine wafers in the treatment of high-grade gliomas: a meta-analysis. J. Neurooncol 122, 367–382 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Bregy A et al. The role of Gliadel wafers in the treatment of high-grade gliomas. Expert Rev. Anticancer Ther 13, 1453–1461 (2013). [DOI] [PubMed] [Google Scholar]
- 223.Masi BC et al. Intracranial MEMS based temozolomide delivery in a 9L rat gliosarcoma model. Biomaterials 33, 5768–5775 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Vellimana AK et al. Combination of paclitaxel thermal gel depot with temozolomide and radiotherapy significantly prolongs survival in an experimental rodent glioma model. J. Neurooncol 111, 229–236 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Bastiancich C et al. Lauroyl-gemcitabine-loaded lipid nanocapsule hydrogel for the treatment of glioblastoma. J. Control. Release 225, 283–293 (2016). [DOI] [PubMed] [Google Scholar]
- 226.Wick W et al. Phase III study of enzastaurin compared with lomustine in the treatment of recurrent intracranial glioblastoma. J. Clin. Oncol. 28, 1168–1174 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Lau D, Magill ST & Aghi MK Molecularly targeted therapies for recurrent glioblastoma: current and future targets. Neurosurg. Focus 37, E15 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Westphal M et al. Adenovirus-mediated gene therapy with sitimagene ceradenovec followed by intravenous ganciclovir for patients with operable high-grade glioma (ASPECT): a randomised, open-label, phase 3 trial. Lancet Oncol 14, 823–833 (2013). [DOI] [PubMed] [Google Scholar]
- 229.Jain RK, Munn LL & Fukumura D Dissecting tumour pathophysiology using intravital microscopy. Nat. Rev. Cancer 2, 266–276 (2002). [DOI] [PubMed] [Google Scholar]
- 230.Galldiks N, Law I, Pope WB, Arbizu J & Langen K-J The use of amino acid PET and conventional MRI for monitoring of brain tumor therapy. Neuroimage Clin 13, 386–394 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Dhermain FG, Hau P, Lanfermann H, Jacobs AH & van den Bent MJ Advanced MRI and PET imaging for assessment of treatment response in patients with gliomas. Lancet Neurol 9, 906–920 (2010). [DOI] [PubMed] [Google Scholar]
- 232.Niemeijer AN et al. Whole body PD-1 and PD-L1 positron emission tomography in patients with non-small-cell lung cancer. Nat. Commun 9, 4664 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Keu KV et al. Reporter gene imaging of targeted T cell immunotherapy in recurrent glioma. Sci. Transl Med 9, eaag2196 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]