

Abstract
Preeclampsia (PE) is an idiopathic disease that occurs during pregnancy. It comprises multiple organ and system damage, and can seriously threaten the safety of the mother and infant throughout the perinatal period. As the pathogenesis of PE is unclear, there are few specific remedies. Currently, the only way to eliminate the clinical symptoms is to terminate the pregnancy. Although noncoding RNA (ncRNA) was once thought to be the “junk” of gene transcription, it is now known to be widely involved in pathological and physiological processes, including pregnancy-related disorders. Moreover, there is growing evidence that the unbalanced expression of specific ncRNA is involved in the pathogenesis of PE. In the present review, we summarize the expression patterns of ncRNAs, i.e., microRNAs (miRNAs), long noncoding RNAs (lncRNAs), and circular RNAs (circRNAs), and the functional mechanisms by which they affect the development of PE, and examine the clinical significance of ncRNAs as biomarkers for the diagnosis of PE. We also discuss the contributions made by genetic polymorphisms and epigenetic ncRNA regulation to PE. In the present review, we wish to explore and reinforce the clinical value of ncRNAs as noninvasive biomarkers of PE.
Keywords: microRNA, lncRNA, circRNA, Biomarker, Preeclampsia
Introduction
Preeclampsia (PE) is a pregnancy-related disorder that is associated with the unprecedented onset of hypertension (systolic blood pressure ≥ 140 mmHg, diastolic blood pressure ≥ 90 mmHg). It occurs after the 20th week of gestation, and frequently near term. It is estimated that PE occurs in 3–8% of pregnant women globally [1]. Although PE is usually identified by new episodes of hypertension and proteinuria after 20 weeks of gestation, pregnant women without proteinuria may be diagnosed with the disorder if they present with one of the following: thrombocytopenia (a platelet count of less than 100,000 per μL); impaired liver function, such as an abnormal rise in the blood concentration of transaminase (to twice the normal concentration), or renal insufficiency (a serum creatinine concentration greater than 1.1 mg/dL, or in the absence of other kidney disease, doubling of the serum creatinine concentration); pulmonary edema; and the unprecedented onset of headaches that are unresponsive to medication and cannot be accounted for by an alternative diagnoses or visual symptoms [1]. Currently, a strategy for the timely detection and diagnosis of PE is urgently needed. This would avoid emergencies or existing complications with target organs. There are several related theories about the causes of PE, including chronic uterine or placental ischemia, immune disorders, genetic imprinting [2], trophoblast apoptosis and necrosis [3], and excessive trophoblast tolerance to inflammatory reactions [4]. Moreover, previous observations have indicated that an imbalance of angiogenesis factors may also play an important role in the pathogenesis of PE [5]. All these pathogenesis processes may be affected by genetic, epigenetic, environmental, and physiological factors, and there is growing evidence that epigenetics play a role in PE [6]. With regard to epigenetics, modifications to both DNA and histones are intermediately involved in the regulation of gene activity. Furthermore, the regulation of functional noncoding RNAs (ncRNAs) can alter gene activity, which modulates gene expression and transcription, chromatin structure, epigenetic memory, selective RNA splicing, and protein translation [7].
ncRNA is a type of functional RNA molecule that is not usually translated into protein. It accounts for 98% of the human genome, and includes housekeeping ncRNA (transfer RNA (tRNA), ribosomal RNA (rRNA), and small nuclear RNA (snRNA)) and regulatory ncRNA (small interfering RNA (siRNA), microRNA (miRNA), piwi-interacting RNA (piRNA), long noncoding RNA (lncRNA), and circular RNA (circRNA)) [8]. The regulatory ncRNAs can be divided into three types—lncRNA (> 200 nucleotides (nt)), miRNA (< 200 nt), and circRNA (circular structure)—which can regulate cell processes through direct interaction with each other [9]. For example, miRNAs can regulate gene expression by targeting mRNA [10] or by adjusting its stability by targeting circRNAs and lncRNAs [11–13]. Alternatively, circRNAs and lncRNAs can serve as “sponges” to adjust the availability of miRNAs [14, 15]. ncRNAs can also adjust the physiological function of cells by interacting with DNA or proteins [9, 16] The aberrant expression of ncRNAs or their abnormal interactions can lead to the development of various diseases, including PE and cardiovascular disease. There is increasing evidence that miRNAs, lncRNAs, and circRNAs are widely involved in the pathogenesis of PE. In the present review, we summarize the role of these transcripts in the pathogenesis of PE, and highlight the possible use of ncRNA as a noninvasive tool for diagnosing the condition.
miRNAs and preeclampsia
miRNAs are the best studied of the ncRNAs. They are small single-stranded structures of approximately 22–25 nucleotides that can act as regulators at the post-transcriptional level. When miRNA molecule binds to the 3′-untranslated region of mRNA molecule, it induces the degradation of the mRNA or prevents its translation. It is estimated that miRNAs are able to regulate the translation of more than 60% of the protein-coding genes involved in many physiological processes, such as proliferation, differentiation, apoptosis, and development [17]. Numerous studies have revealed abnormal miRNA expression in the placentas or peripheral blood of patients with PE. Aberrant miRNAs can target downstream genes and reduce the migration and invasion of trophoblasts, or increase cell apoptosis, ultimately resulting in PE.
Expression pattern of miRNAs in patients with PE
To date, approximately 70 miRNAs have been reported to be differentially expressed in PE tissues. Table 1 summarizes the differentially expressed miRNAs involved in the pathogenesis of PE. For the first time, Pineles et al. reported seven differentially expressed miRNAs in the placentas of patients with PE who gave birth to either normal babies or babies that were small for their gestational age. Among these miRNAs, only the levels of miR-182 and miR-210 increased significantly relative to the corresponding levels experienced during normal pregnancy [118]. This finding presented new targets for the pathophysiology of PE. In view of the role of miR-210 in PE, related studies with larger sample sizes further confirmed that the placental expression of miR-210 in patients with PE did indeed increase significantly compared to the corresponding levels experienced during normal pregnancy [18–20]. miR-210 can inhibit the proliferation, invasion, and angiogenesis of trophoblasts by acting on the downstream target genes that encode KCMF1 [20], NOTCH1 [21] and MAPK [18]. Upregulated levels of miR-210 have also been detected in peripheral blood serum [18]. It has also been reported that the disturbed expression of miR-182-5p can inhibit trophoblast proliferation, invasion, and migration by acting on the 3′-untranslated region of RND3 [24]. Zhang et al. first reported that miR-155, an inflammation-related miRNA, is overexpressed in the placentas of PE patients, and is involved in the pathogenesis of PE because it downregulates CYR61 [25]. miR-155 can bind the target genes that encode cyclin D1 [16] and eNOS [26] to affect the migration and proliferation of trophoblasts. Differentially expressed miRNAs have been found in exosomes [50, 64, 103, 112] and Mesenchymal stem cells (MSCs) [28, 51, 71, 76] as well as in placental tissues and peripheral serum or plasma. For example, miR-16 upregulation was first confirmed in the placentas of patients with PE [29]. Studies have shown that miR-16 is differentially expressed in the decidual MSCs of patients with severe PE and normal patients, and can inhibit the proliferation and migration of decidual-derived MSCs by targeting cyclin E1 and inducing cell cycle arrest [28]. More interestingly, miR-16 overexpression in decidual-derived MSCs can also reduce the ability of human umbilical vein endothelial cells to form blood vessels [28].
Table 1.
Dysregulation of miRNAs in PE
| MiRNA | Sample type | Status | Targets | Function | Ref. |
|---|---|---|---|---|---|
| miR-210 | Serum/placenta | upregulated | NOTCH1/PTPN2/THSD7A/KCMF1/FOXA1 | Upregulation of miR-210 decreased NOTCH1/PTPN2/THSD7A/KCMF1 expression, impaired HTR-8/SVneo proliferation, migration, invasion, and tube-like formation capabilities, and promoted apoptosis. | [18–23] |
| miR-182-5p | placenta | upregulated | RND3 | the increased miRNA-182-5p expression could inhibit the migratory and invasive ability of trophoblast cells through targeted degrading RND3 protein | [24] |
| miR-155 | placenta/placenta-associated serum exosomes | upregulated | CYR61/Cyclin D1 | Overexpression of miR-155 in HTR-8/SVneo cells inhibited cell invasion, proliferation and increased cell number at the G1 stage in trophoblast cells | [16, 25] |
| miR-155-5p | placenta | upregulated | eNOS | miR-155 inhibited cell invasion in trophoblast cells, and the effect was rescued by over expression of eNOS. | [26] |
| miR-195 | placenta | downregulated | ActRIIB/ActRIIA | miR-195 could promote cell invasion via directly targeting ActRIIB/ActRIIA in human trophoblast cells | [27] |
| miR-16 | placenta/mesenchymal stem cell (MSC) | upregulated | CCNE1 /VEGF-A/Notch2 | over-expressed miR-16 inhibited the proliferation and migration of decidua-derived mesenchymal stem cells /BeWo and JEG-3 cells, and induced cell-cycle arrest by targeting cyclin E1 | [28, 29] |
| miRNA-376c | placenta/plasma/exosome | downregulated | ALK5/ALK7/25-OH-VD | miR-376c inhibits both ALK5 and ALK7 expression to impair transforming growth factor-β/Nodal signaling, leading to increases in cell proliferation and invasion | [30, 31] |
| miR-29b | decidua-derived mesenchymal stem cell (dMSC)/placenta | upregulated | MMP2/MCL1/ VEGFA/ ITGB1/HDAC4 | miR-29b induced apoptosis and inhibited invasion and angiogenesis of trophoblast cells. | [32] |
| miR-101 | placenta | downregulated | ERp44/BRD4/CXCL6 | miR-101 could promote apoptosis and inhibite the proliferation and migration of trophoblasts | [10, 33, 34] |
| miR-18a | placenta/plasma | downregulated | Smad2/ER1/ESRα | miR-18a could promote trophoblast cell invasion and suppress apoptosis of human trophoblast cells | [35–37] |
| miR-20a | placenta | upregulated | Foxa1/VEGF | the upregulated miR-20a in human preeclampsia tissue can inhibit the proliferative and invasive activities of trophoblast cells | [23, 38] |
| miR-125b-1-3p | placenta | upregulated | S1PR1 | miR-125b-1-3p inhibited the invasiveness of human trophoblast cells, | [39] |
| miR-125b | placenta/plasma | upregulated | SGPL1/ STAT3/KCNA1 /GPC1/Trop-2 | upregulated miR-125b can impair endothelial cell function, reduce cell proliferation and invasion and migration | [40–43] |
| miR-126 | placenta | downregulated |
VEGF/ PIK3R2 |
miR-126 enhanced endothelial progenitor cell (EPC) proliferation, differentiation and migration | [44, 45] |
| miR-196b | plasma | downregulated | / | / | [46] |
| miR-206 | plasma | upregulated | VEGF/IGF-1/ET-1 | miR-206 modulated trophoblast cells migration and invasion | [47–49] |
| miR-494 | Mesenchymal stem cells(MSC)/serum exosomes | upregulated | VEGF/CDK6 | supernatant from miR-494-overexpressing dMSCs could reduce HTR-8/SVneo migration, impair HUVEC capillary formation and arrest G1/S transition | [50, 51] |
| miR-519d-3p | placenta | upregulated | MMP-2 | upregulation of miR-519d-3p may contribute to the development of PE by inhibiting trophoblast cell migration and invasion via targeting MMP-2 | [52] |
| miR-335 | placenta | upregulated | eNOS/Sp1 | miR-335-5p could inhibit transforming from epithelial to mesenchymal and cell migration | [53, 54] |
| miR-584 | placenta | upregulated | eNOS | miR-335 inhibited the migratory ability of the trophoblast cells, and the effect was ‘rescued’ by overexpressed eNOS | [53] |
| miR-34a | placenta | upregulated | MYC/BCL-2, Notch-1 | Overexpression of miR-34a inhibited cell proliferation, migration and invasion, and induced trophoblast cell apoptosis by inhibiting expression of BCL-2 protein | [55–57] |
| miR-204 | placenta | upregulated | MMP-9 | miR-204 may contribute to the development of preeclampsia by inhibiting trophoblastic invasion | [58] |
| miR-193b-3p | placenta | upregulated | TGF-β2 | miR-193b-3p could regulate trophoblasts migration and invasion through binding onto the 3’UTR target site of TGF-β2 | [59] |
| miR-193b-5p | placenta | upregulated | APLN/FGF13 | Overexpression of miR-193b-5p inhibited trophoblast cell proliferation and migration | [60] |
| miR-203 | placenta | upregulated | VEGFA/SOCS-3 | miR-203 overexpression inhibited the proliferation, migration and invasion ability of HTR-8/SVneo cells, meanwhile which increased the endothelial inflammatory response | [61–63] |
| miRNA-203a-3p | placental mononuclear cells and exosomes | downregulated | IL24 | microRNA-203a-3p was able to suppress the proliferation capacity of LPS-stimulated mononuclear macrophages, and it exerted anti-inflammatory effects via down-regulating IL24 in THP-1 cells. | [64] |
| miR-885-5p | placenta | upregulated | MMP-9 | / | [65] |
| miR-141 | placenta/plasma | upregulated |
EG-VEGF/ CXCL12β/ CXCR2 / 4 |
miR-141 could inhibit the invasion and vascularization abilities, and promote the apoptosis of HTR-8/SVneo cells | [66, 67] |
| miR-141-5p | placenta | downregulated | ATF2 | miR-141-5p up-regulated transcription factor ATF2 to promote phosphatase DUSP1 expression, which reduces p-MAPK1 and ERK1/2 expression to promote preeclampsia. | [68] |
| miR-128a | placenta | upregulated | Bax | miR-128a induced the apoptosis of HTR-8/SVneo cells by down-regulating Bax through the mitochondrial apoptosis pathway. | [69] |
| miR-145 | placenta | downregulated | PI3K/MUC1 | miR-145 may serve key roles in the regulation of trophoblast cell proliferation and invasion | [70] |
| miR-136 | Mesenchymal stem cells (MSCs)/serum exosomes | upregulated | BCL2/VEGF | MiR-136 significantly increase the apoptosis and suppress the proliferation of MSCs, and it could also inhibit the capillary formation and trophoblast cell invasion. | [71] |
| miR-520 g | serum | upregulated | MMP-2 | Elevated maternal serum level of miR-520 g level in first trimester could suppress the migration and invasion of trophoblast, and might play a role in the defective spiral artery remodeling, | [72] |
| miR-20b | placentas and peripheral blood | upregulated | MMP-2 | miR-20b inhibited trophoblastic invasion by targeting MMP2 | [73] |
| miR-23a | placenta | upregulated | XIAP/HDAC2 | miR-23a reduced HTR-8/SVneo cell migration and invasion and increased HTR-8/SVneo cell apoptosis | [74, 75] |
| miR-495 | peripheral blood exosomes/umbilical cord mesenchymal stem cells (UCMSCs) | upregulated | Bmi-1 | The supernatants from miR-495-overexpressed inhibited the migration of MSCs and HTR-8/SVneo, invasion of HTR-8/SVneo and tube formation of HUVEC | [76] |
| miR-137 | placenta | upregulated | ERRa | MiRNA-137 significantly reduced the proliferation and migration of placenta trophoblast cells | [77] |
| miR-93 | placenta/plasma | upregulated | MMP-2 | miR-93 reduced migration and invasion of immortalized trophoblast cells. | [78] |
| miR-144 | placenta | downregulated | PTEN | miR-144 induced significant increase in cell proliferation, migration, invasion, and decrease in cell apoptosis, and also affected the cell cycles | [79] |
| miR-144-3p | placenta | downregulated | Cox-2 | Downregulation of miR-144-3p led to systemic inflammation and endothelial cell injury | [80] |
| miR-942 | plasma | downregulated | ENG | Decreased miR-942 expression decreased the invasive ability of TEV-1 cells, and inhibited the HUVEC angiogenesis assay | [81] |
| miR-134 | placenta | upregulated | ITGB1 | MiR-134 suppressed the infiltration of trophoblast cells by targeting ITGB1 | [82] |
| miR-362-3p | placenta | upregulated | Pax3 | miR-362-3p/Pax3 axis regulates cell viability, migration and invasion of HTR8/SVneo cells under hypoxia. | [83] |
| miR-454 | placenta | downregulated | EPHB4/ALK7 | MiR-454 promotes the proliferation and invasion of trophoblast cells, and inhibit the apoptosis | [84, 85] |
| miR-30a-3p | placenta | upregulated | IGF-1 | the over-expression of miR-30a-3p alter the invasive capacity of JEG-3 cells and induce the apoptosis of HTR-8/SVneo cells | [86] |
| miR-31-5p | serum | upregulated | eNOS | NF-κB-responsive miR-31-5p elicits endothelial dysfunction, hypertension, and vascular remodeling via post-transcriptional down-regulation of eNOS | [87] |
| miR-423–5p | plasma | upregulated | IGF2BP1 | MiR-423-5p inhibited migration, invasion and proliferation as well as induced apoptosis in HTR-8/SVneo cells. | [88] |
| miR-299 | placenta | upregulated | HDAC2 | miR-299 suppressed the invasion and migration of trophoblast cells partly via targeting HDAC2 | [89] |
| miR-4421 | placenta | upregulated | CYP11B2 | overexpression of miR-4421 inhibited trophoblast proliferation and significantly blocked cell cycle | [90] |
| miR-320a | placenta | upregulated | IL-4/ERRγ | Excessive miR-320a expression remarkably suppressed trophoblast invasion and proliferation | [12, 91] |
| miR-517-5p | placenta | upregulated | MMP-2 | MiR-517-5p could promote proliferative and invasive abilities of JAR cells by inhibiting ERK/MMP-2 pathway | [92] |
| miR-517c-3p | plasma | upregulated | TNFSF15 | miR-517c-3p could suppress cell growth and proliferation, and promote placental hypoxia, immune response and apoptosis. | [93] |
| miR-let-7d | placenta | upregulated | / | low expression levels of miR-let-7d plays a central role in suppressing apoptosis in addition to promoting the proliferation and invasion of PE TCs. | [94] |
| miR-218-5p | placenta | downregulated | TGF-β2 | miR-218-5p accelerated spiral artery remodeling in a decidua-placenta co-culture and promoted trophoblast invasion and enEVT differentiation | [95] |
| miR-181a-5p | placenta | upregulated | IGF2BP2 | miR-181a-5p may trigger antiproliferation and inhibition of cell cycle progression, induce apoptosis, and suppress invasion in HTR-8/SVneo and JAR cells | [96] |
| miR-142-3p | placenta | upregulated | TGF-β1 | miR-142-3p suppressed cell invasion and migration by reactivating the TGF-β1/Smad3 signaling pathway. | [97] |
| miR-145-5p | placenta | downregulated | FLT1/Cyr61 | miR-145-5p promoted trophoblast cell growth and invasion | [98, 99] |
| miR-30b | placenta | upregulated | MXRA5 | miR-30b might contribute to PE through inhibiting cell viability, invasion while inducing apoptosis of placental trophoblast cells via MAPK pathway by targeting MXRA5. | [100] |
| miR-454 | placenta | downregulated | ALK7/EPHB4 | miR-454 promotes the proliferation and invasion of trophoblast cells by inhibiting EPHB4 /ALK7 | [84, 85] |
| miR-200a | plasma | upregulated | EG-VEGF/TTR | miR-200a suppressed primary cilia formation and inhibited trophoblast invasion. | [101, 102] |
| miR-548c-5p | placenta/ serum exosome | downregulated | PTPRO | miR-548c-5p could inhibit the proliferation and activation of LPS-stimulated macrophages and decrease levels of inflammatory cytokines | [103] |
| miR-342-3p | placenta | upregulated | PDGFRA/ID4 | miR-342-3p was proposed to inhibit the proliferation, migration, invasion and G1/S phase transition of HTR8/SVneo cells | [104, 105] |
| miR-431 | placenta | upregulated | ZEB1 | miR-431 inhibited the migration and invasion of trophoblastic cells | [106] |
| miR-221-3p | placenta | downregulated | THBS2 | miR-221-3p overexpression inhibited apoptosis, increased cell population at S phase, and decreased cell population at G0/G1 phase of HTR-8/SVneo cells | [107] |
| miR-152 | placenta | upregulated | VEGF | the increased miR-152 expression can promote the apoptosis of trophoblast cells. | [108] |
| miR-133 | serum | upregulated | Rho/ROCK | MiR-133 may affect the apoptosis of trophoblasts in the placenta tissues | [109] |
| miR-384 | placenta/plasma | upregulated | PTBP3 | Cell proliferation and migration were inhibited by miR-384 overexpression | [110] |
| miR-21 | Serum/placenta | upregulated | FOXM1 | MiR-21 may regulate placental cell proliferation | [111] |
| miR-125a-5p | placenta/exosome | upregulated | VEGFA | miR-125a-5p might affect HTR8/SVneo cell proliferation and migration and inhibit angiogenesis | [112] |
| miR-214-3p | serum | upregulated | PIGF | Downregulation of miR-214-3p promoted trophoblast invasion into the decidua, as well as spiral artery remodeling | [113] |
| miR-518b | plasma | upregulated | EGR1 | Increased miR-518b inhibited trophoblast migration and angiogenesis | [114] |
| miRNA-646 | serum | upregulated | VEGF-A | miR-646 suppressed endothelial progenitor cells (EPCs) multiplication, differentiation and migration. | [115] |
| miR-215-5p | placenta | upregulated | CDC6 | miR-215-5p inhibited both the migration and invasive potential of trophoblasts, besides decreasing the G1-S transition in HTR-8/SVneo cells | [116] |
| miR-483 | venous blood/ umbilical cord blood / placental tissue | downregulate | IGF1 | miR-483 regulates the expression of PI3K, Akt, and mTOR in endothelial progenitor cells | [117] |
Diagnostic value of miRNAs in PE
Numerous studies have confirmed that miRNAs are involved in the pathogenesis of PE and are differentially expressed in patients with this disease. Some researchers have assessed the diagnostic value of miRNA with regard to PE by drawing receiver operating characteristic (ROC) curves. For example, Zhang et al. showed that the levels of miR-942 decreased significantly in the plasma of patients with PE compared to the corresponding levels in normal patients, and had 65.4% sensitivity and 69.2% specificity with regard to PE diagnosis [81]. Table 2 summarizes the results of research into the value of miRNAs with regard to the diagnosis of PE.
Table 2.
Diagnostic value of miRNAs in PE
| MiRNA | Sample type | Area under curve | Sensitivity | Specificity | Ref. |
|---|---|---|---|---|---|
| miR-31-5p | serum | 0.96 | 95.65% | 92.39% | [113] |
| miR-155-5p | serum | 0.931 | 89.13% | 88.04% | [113] |
| miR-214-3p | serum | 0.924 | 90.22% | 79.35% | [113] |
| miR-1290-3p | serum | 0.957 | 94.57% | 84.78% | [113] |
| miR-210 | serum | 0.75 | / | / | [119] |
| miR-155 | serum | 0.718 | / | / | [119] |
| miR-206 | plasma | 0.94 | 97% | 77.50% | [48] |
| miR-518b | plasma | 0.715 | / | / | [120] |
| miR-31 | plasma | 0.875 | 95.00% | 70.00% | [121] |
| miR-21 | plasma | 0.793 | 65.10% | 90.30% | [121] |
| miRNA-136 | exosome | 1 | 95% | 100% | [50] |
| miRNA-494 | exosome | 0.868 | 86% | 95% | [50] |
| miRNA-495 | exosome | 0.94 | 90% | 83% | [50] |
| miR-195 | placenta | 0.82 | 80% | 80% | [122] |
| miR-423-5p | plasma | 0.844 | 87.50% | 80% | [123] |
| miR-942 | plasma | 0.718 | 65.40% | 69.20% | [81] |
| miR-517-5p | plasma | 0.7 | 42.90% | 86.20% | [124] |
| miR-516-5p | plasma | 0.608 | 38.10% | 84.50% | [124] |
| miR-518b | plasma | 0.55 | 34.40% | 78.70% | [124] |
Association between miRNA variants and the risk of PE
Given that genetic factors play important roles in the occurrence of PE, several studies have focused on the relationship between single nucleotide polymorphisms (SNPs) in miRNAs and the risk of PE. It has also been reported that the miR-146a rs2910164 polymorphism may not be associated with PE susceptibility, cytokines, or related characteristics in black women from South Africa, whereas the GC/CC genotype may reduce susceptibility to severe PE [125]. Interestingly, Salimi et al. discovered that a maternal/placental miR-146a polymorphism (rs2910164) was associated with PE or risk of severe PE, after they genotyped it in the blood samples and placentas from the Asian mainland, using polymerase chain reaction (PCR)–fragment length polymorphism [126]. Table 3 summarizes the reported results.
Table 3.
Association between polymorphisms with SNPs and risk of PE
| MiRNA | Sample type | Risk Variant | Association with PE | minor allele frequencyn n(%) | Ref. |
|---|---|---|---|---|---|
| miRNA-155 | serum | rs767649 | A allele | T allele(39.2) | [127] |
| miRNA-146a | maternal blood | rs2910164 | Negative | C allele(38.7) | [126, 128] |
| miRNA-27a | maternal blood/Placental | rs895819 | GC + CC vs GG | T allele(46) | [129] |
| miRNA-196a2 | maternal blood | rs11614913 | Negative | T allele(38.6) | [130] |
| miR-499 | Placental | rs3746444 | CT + TT vs CC | C allele(37.3) | [130] |
| miRNA-152 | maternal blood | rs12940701 | CC vs TC + TT | T allele(28.9) | [131] |
| miRNA-26a | maternal blood/Placental | rs7372209 | Negative | T allele(16) | [132] |
Demethylation of the miRNA in PE
In addition to the previously reported abnormal expression of miRNA in PE patients, some studies also found that the methylation level of some abnormal expression of miRNA is associated with the risk of PE. Rezaei et al. found that hypomethylation of the miR-34a promoter was associated with the occurrence and severity of PE, when they applied methylation-specific PCR to investigate samples from 104 pregnant women with PE and 119 normotensive pregnant women [133]. Moreover, studies have shown that the abnormal regulation of the miR-let-7 family is related to PE. The methylation status of miR-let-7a in PE was evaluated by methylation-specific PCR and bisulfite sequencing PCR analyses. The results suggested that hypomethylated miR-let-7a promotes PE by downregulating Bcl-xl and YAP1 [134]. The miR-510 promoter region in bisulfite-treated PE DNA samples was also found to be unmethylated, compared to the corresponding region in the control samples [135].
lncRNAs and PE
lncRNAs are composed of more than 200 nucleotides. They promote the development of human diseases by participating in various biological processes, including genomic imprinting, chromatin modification, regulation of transcription and post-transcriptional gene expression, nuclear transport, and other regulatory processes [136]. There is abundant evidence to suggest that lncRNA expression in the placenta and peripheral blood differs between healthy pregnant women and patients with PE. This indicates that abnormal lncRNA expression is involved in the pathogenesis of PE.
Expression pattern of lncRNAs in patients with PE
Table 4 summarizes the differentially expressed lncRNAs that participate in the pathogenesis of PE. For example, Liu et al. discovered that the levels of GASAL1 lncRNA were downregulated in the placental tissues of 30 patients with PE, compared to the corresponding levels in 30 normal controls. They further demonstrated that GASAL1 lncRNA can directly bind to functional RNA-binding protein SRSF1 to promote trophoblastic proliferation and progression from the G1 to the S phase through the mTOR and EBP1 signaling pathways. It can also inhibit trophoblastic apoptosis by downregulating cleaved caspase-3 and upregulating Bcl-2 [174]. Another well-known lncRNA, that of MEG3, is expressed in a variety of normal tissues, but is absent in many tumors and tumor cell lines [189]. Yu et al. found that the expression of lncRNA MEG3 in PE placental tissue decreased significantly by 72% compared with that in the normal controls, and MEG3 interruption induced the expression of E-cadherin but reduced that of N-cadherin. This confirmed that MEG3 inhibits trophoblastic migration and invasion. They also found that MEG3 downregulation affects the TGF-β/Smad pathway by inhibiting Smad7 expression, thereby suppressing epithelial–mesenchymal transition [147]. Wang and Zou also showed that by regulating the expression of NOTCH1, MEG3 can promote the apoptosis of trophoblasts, and inhibit their migration and invasion, thereby inducing PE [148].
Table 4.
Dysregulation of lncRNAs in PE
| LncRNA | Sample type | Status | Targets | Function | Ref. |
|---|---|---|---|---|---|
| BC030099 | the whole blood (WB) | upregulated | / | / | [137] |
| LOC391533 | placenta | upregulated | sFlt-1 | This protein plays an important role in angiogenesis and vasculogenesis. | [138] |
| LOC284100 | placenta | upregulated | / | / | |
| CEACAMP8 | placenta | upregulated | / | / | |
| HOTAIR | placenta | upregulated | miR-106a | High level of HOTAIR represses the proliferation, migration and invasion of trophoblast cells through downregulating miR-106 in an EZH2-dependent manner. | [139] |
| downregulated | PUM1/HOTAIR | HOTAIR promotes trophoblast invasion by activating PI3K-AKT signaling pathway. | [140] | ||
| AGAP2-AS1 | placenta | downregulated | JDP2 | AGAP2-AS1 knockdown could inhibit trophoblasts proliferation, invasion and spiral artery remodeling and promote cell apoptosis. | [141] |
| HOXA11-AS | placenta | downregulated | miR-15b-5p/HOXA-7/Lsd1 and Ezh2/RND3 | Down-regulated HOXA11-AS inhibits trophoblast cell proliferation, migration and invasion, and promoted cell accumulation in G0–G1 phase and apoptosis. | [142] |
| TUG1 | placenta | downregulated | miR-29b/MCL1/VEGFA /MMP2 | TUG1 could act as a molecular sponge for miR-29b, thus down-regulating MCL1, VEGFA, and MMP2 to promote cell proliferation, invasion, and angiogenesis, while negatively regulated cell apoptosis. | [14] |
| Ezh2/RND3 | Down-regulated TUG1 inhibits trophoblast cell proliferation, migration, invasion and the formation of capillary-like networks and promotes trophoblast cell apoptosis. | [143] | |||
| miR-204-5p | Down-regulated TUG1 negatively regulates trophoblast migration and invasion partly through sponging miR-204-5p and negatively regulating the expression and function of miR-204-5p. | [144] | |||
| SPRY4-IT1 | placenta | upregulated | Bax/Bcl-2 | SPRY4-IT1 overexpression significantly decreases the cell migration, proliferation and spiral artery remodeling, while increases cell apoptosis. | [145] |
| Wnt/β-catenin pathway | Overexpression of SPRY4-IT1 suppresses trophoblast cell migration, invasion and spiral artery remodeling by inducing E-cadherin and β-catenin expression and decreasing vimentin expression. | [146] | |||
| MEG3 | placenta | downregulated | TGF-β/ /E-cadherin/N-cadherin | Down-regulated MEG3 induces E-cadherin upregulation and N-cadherin and slug downregulation in HTR-8/SVneo cells, which inhibits trophoblast cell proliferation, migration, invasion and EMT. | [147] |
| Notch1 | Down-regulation of MEG3 could downregulate Notch1 expression to suppress trophoblast cells migration, invasion and promote its apoptosis. | [148] | |||
| NF-κB/Caspase-3/ Bax | Down-regulation of MEG3 increases apoptosis and decreases migration of trophoblast cells by influencing expression of NF-κB, Caspase-3, and Bax protein expressions. | [149] | |||
| H19 | placenta | downregulated | NOMO1/miR-675 | Lowered expression of H19 participate in the excessive proliferation of trophoblast cells by downregulating miR-675 which targets NOMO1 and interferes with Nodal signaling. | [150] |
| miR-148a-5p/P28/miR-216-3p/EBI3 | Down-regulated could up-regulate the expression of miR-148-5p/miR-216-3p and the expressions of subunits of IL-27, P28 and EBI3 were thus suppressed. | [151] | |||
| miRNAlet-7/the type III TGF-β receptor (TβR3) | H19 repression decreases TGF-β signaling via the Par6/Smurf1/RhoA pathway activated by TβR3, leading to impaired migration and invasion of EVT cells. | [152] | |||
| upregulated | PI3K/AKT/mTOR pathways | LncRNA-H19 overexpression reduces cell viability but increases invasion and autophagy in trophoblast cells by enhancing phosphorylated levels of key kinases in the PI3K/AKT/mTOR pathways. | [153] | ||
| MIR503HG | placenta | upregulated | the matrix metalloproteinase-2/− 9, the snail /E-cadherin/ NF-κB signaling pathway | MIR503HG overexpression suppresses cell proliferation, invasion, and migration, and induces apoptosis and causes cell cycle arrest at the G0/G1 phase of HTR-8/SVneo | [154] |
| DLX6-AS1 | placenta tissues | upregulated | miR-149–5p/ERP44 pathway | DLX6-AS1 inhibits proliferation and invasion of trophoblast cells, and suppresses angiogenesis of HUVEC cells by binding miR-149–5p/ERP44 pathway. | [11] |
| miR-376c/ GADD45A | DLX6-AS1 may contribute to preeclampsia by impairing proliferative, migratory and invasive abilities of trophoblasts via the miR-376c/GADD45A axis. | [155] | |||
| ZEB2-AS1 | placenta | downregulated | miR-149 / PGF | ZEB2-AS1 contributes to PE progression by inhibiting cell proliferative, migratory, invasive capacities and EMT via the miR-149/PGF axis in HTR-8/SVneo cells. | [156] |
| PRNCR1 | placenta | upregulated | MAPK | Overexpression of LncRNA PRNCR1 in PE patients reduces the viability of cells, and is positively correlated with systolic blood pressure, diastolic blood pressure and urine protein | [157] |
| GHET1 | placenta | downregulated | the E-cadherin / vimentin /fibronectin | GHET1 promotes PE by inhibiting cell proliferative, migratory, invasive capacities and EMT by stimulating E-cadherin and suppressing vimentin and fibronectin. | [158] |
| TDRG1 | placenta | downregulated | miR-214-5p/Notch signaling pathway | TDRG1 inhibits cell viability, proliferation, migration, and invasion by stimulating the expression of miR-214-5p and regulating the Notch signaling pathway in trophoblast cells. | [159] |
| MALAT1 | placenta | downregulated | pro-apoptotic proteins | Silencing of MALAT-1 in JEG-3 cells suppresses proliferation, migration and invasion, and induces cell cycle arrest at G0/G1 phase and enhancing apoptosis. | [160] |
| N-cadherin/vimentin /E-cadherin/Hu-antigen R (HuR) /FOS | Down-regulated MALAT1 inhibits trophoblast invasion, migration, epithelial mesenchymal transition (EMT) and spiral artery remodeling by upregulating E-cadherin and downregulating FOS, N-cadherin, and vimentin. | [161] | |||
| miR-206/IGF-1 axis | Down-regulated MALAT1 regulates miR-206/IGF-1 axis and thereby inhibits trophoblast cells migration and invasion through PI3K/Akt signal pathway. | [49] | |||
| TCL6 | placenta | upregulated | PTEN/CDK2 | Overexpression of lncRNA TCL6 is positively correlated with systolic blood pressure, diastolic blood pressure and urine protein, whereas negatively correlated with fetal birth weight of PE patients | [162] |
| uc003fir | placenta /preeclamptic placenta vessels/ vessel-free tissue | upregulated | CCL5/miR-155 | Over-expression of lncRNA uc003fir increases proliferation, migration, and invasion of HTR-8/SVneo cells. | [163] |
| FOXD2-AS1 | placenta | downregulated | miR-3127/MMP2/MMP9 | FOXD2-AS1 silencing decreases the promotion effects on trophoblast cell induced by miR-3127 inhibition, partly mediated by influencing MMP2 and MMP9 levels. | [164] |
| KCNQ1OT1 | placenta | downregulated | miR-146a-3p | KCNQ1OT1 could target the regulation of miR-146a-3p through CXCL12/CXCR4 pathway in the proliferation, invasion and migration of HTR8/SVneo cells. | [165] |
| SNHG5 | placenta | downregulated | miR-26a-5p/N-cadherin | Knockdown of SNHG5 inhibits trophoblast (HTR-8/SVneo) cell proliferation, invasion, and migration, and promotes apoptosis and caused increase of cell population at the G 0 /G 1 phase and decrease of cell population at the S phase. | [166] |
| PVT1 | placenta | downregulated | PI3K/AKT | PVT1 knockdown notably inhibits the proliferation, migration and invasiveness abilities of trophoblast cells, and significantly promotes the apoptosis through PI3K/AKT pathway. | [167] |
| Ezh2/ANGPTL4 | PVT1 knockdown notably inhibits cell proliferation and stimulates cell cycle accumulation and apoptosis by repressing ANGPTL4 transcription through binding with EZH2 in trophoblast cell. | [168] | |||
| WDR86-AS1 | placenta | upregulated | miR-10b-3p / LITAF | WDR86-AS1 downregulates miR-10b-3p but promotes LITAF expression, which controls the inflammatory responses and migration and proliferation of HTR-8/SVneo cells under hypoxia. | [169] |
| ATB | placenta | downregulated | / | Down-regulated lncRNA-ATB decreased migration, proliferation, tube-formation of HTR-8/SVneo cells. | [170] |
| RPAIN | placenta | upregulated | C1q | The increased RPAIN levels may contribute to the development of preeclampsia through regulating trophoblast proliferation, invasion and apoptosis via C1q. | [171] |
| NR_002794 | placenta | upregulated | / | NR_002794 has suppressive effects on proliferation and migration, and results in an increased rate of apoptosis. | [172] |
| MVIH | placenta | downregulated | Jun-B | The silencing of MVIH expression inhibits cell growth, migration, invasion, and angiogenesis in various trophoblast cell lines by modulating Jun-B protein expression. | [173] |
| GASAL1 | placenta | downregulated | SRSF1/mTOR/EBP1/Bcl-2/caspase-3 | lncRNA GASAL1 interacts with SRSF1 to regulate the proliferative, invasive, and apoptotic abilities of trophoblast cells via the mTOR signaling pathway. | [174] |
| MIR193BHG | placenta | upregulated | / | / | [175] |
| PROX1-AS1 | placenta | upregulated | / | / | |
| GATA3-AS1 | placenta | upregulated | / | / | |
| 00511 | placenta | downregulated | miR-31-5p / HOXA7 | Down-regulated lnc00511 suppresses proliferation, invasion and autophagy, and enhances apoptosis in trophoblast cells to mediate pre-eclampsia progression through modulating the miR-31-5p/homeobox protein A7 axis. | [176] |
| placenta | downregulated | miR-29b-3p/ Cyr61 | AP2γ mediates downregulation of lncRNA LINC00511 as a ceRNA suppresses trophoblast invasion, proliferation and migration by regulating miR-29b-3p/Cyr61 axis. | [177] | |
| AK002210 | serum | downregulated | / | AK002210 knockdown suppresses trophoblast invasion, proliferation and migration | [178] |
| ZBTB39 | placenta | upregulated | miR-210/THSD7A | Upregulated LncZBTB39 suppresses trophoblast invasion and migration via antagonizing the inhibition of miR-210 on THSD7A expression. | [179] |
| GAS5 | placenta | upregulated | miR-21/MMP-9/TP53 | GAS5 suppresses trophoblast invasion, proliferation and migration through the regulation of miR-21 and the activation of PI3K/AKT signaling pathway and its downstream proteins covering MMP-9 and TP53. | [180] |
| VIM-AS1 | placenta | downregulated | E-cadherin/Snail/Vimentin | Down-regulated VIM-AS1 suppresses epithelial-to-mesenchymal transition (EMT) by upregulating E-cadherin and downregulating Snail and Vimentin. | [181] |
| FAM99A | placenta | downregulated | Wnt/β-catenin | Down-regulated FAM99A suppresses the invasive and migratory abilities of HTR-8/SVneo, and increases the apoptotic rate. | [182] |
| HIF1A-AS2 | placenta | downregulated | LSD1/PHLDA1 | HIF1A-AS2 suppresses trophoblast cell invasion and proliferation through upregulating PHLDA1 expression. | [183] |
| 00261 | placenta | upregulated | miR-558/TIMP4 | Lnc00261 exerts the suppressive effects on the trophoblast invasion and migration via targeting miR-558/TIMP4 axis, which may involve in the pathogenesis of PE. | [184] |
| Uc.294 | placenta | upregulated | / | Uc.294 inhibits proliferation, invasion and promotes apoptosis of trophoblast cells HTR-8/SVneo. | [185] |
| PSG10P | placenta | upregulated | miR-19a-3p/IL1RAP | IL1RAP promotes the expression of caspase-3 but inhibits MMP9 to suppresses proliferation, migration, and invasion of trophoblast cells. | [186] |
| 00473 | placenta | downregulated | LSD1/TFPI2 | lnc00473 inhibits the expression of tissue factor pathway inhibitor 2 (TFPI2) through binding to lysine-specific demethylase 1 (LSD1) to inhibits cell proliferation and promotes apoptosis. | [187] |
| Uc.187 | placenta | upregulated | PCNA and Ki67/MMP-2/−9 / TIMP-1/ Bcl-2 | Uc.187 suppresses cell proliferation and invasion and promotes the cellular apoptotic response | [188] |
Although there is no report that the methylation level of lncRNA itself is related to PE, abnormally expressed lncRNAs can regulate the proliferation, invasion, and apoptosis of trophoblasts by regulating the methylation of downstream molecules. Zhao et al. found that when lncRNA HOTAIR is expressed at high levels, it targets miR-106 by binding to EZH2 [139], which in turn inhibits the transcription of the target gene by inducing H3K27 methylation in the promoter region, ultimately suppressing the proliferation, migration, and invasion of trophoblasts [190]. Xu et al. further confirmed that EZH2 can directly interact with the promoter region of RND3 by methylating H3K27, the 27th amino acid of histone H3, thereby reducing the expression of RND3 in PE. However, the downregulation of lncRNA TUG1 in placental tissues inhibits the proliferation, invasion, and migration of trophoblasts and promotes their apoptosis and it also obstructs spiral artery remodeling by reducing the transcriptional regulation of RND3, which is mediated by recruited EZH2 proteins [143]. Li et al. reported, for the first time, that TUG1 acts as a molecular sponge for miR-29b, thereby regulating the expression of MCL1, VEGFA, and MMP2; it is therefore involved in the development of PE [14]. Many studies have shown that H19 mutations are closely related to PE. Moreover, lncRNA H19 is upregulated in PE. This activates the PI3K/AKT/mTOR pathway, which reduces trophoblast activity and increases invasion and autophagy [153]. Zuckerwise et al. also proposed that the downregulation of H19 inhibits TGF-β signaling transduction by reducing the Par6/Smurf1/RhoA pathway activated by TβR3 [152], Gao et al. found that this impairs the migration and invasion of extravillous trophoblasts in vitro by upregulating the expression of miR-let-7 and downregulating the expression of miRNA-675 [150]. TβR3 is considered to be a downstream mediator of H19–let-7 interaction. Zhou et al. demonstrated that H19 alters genome-wide methylation levels by regulating the activity of S-adenosyl-l-homocysteine hydrolase, and H19 knockout in the intron region causes the undermethylation of TβR3 [191]. Dey et al. showed that hypermethylation of the H19 promoter is associated with reduced H19 expression [192]. Furthermore, the absence of H19 imprinting in PE placental tissues reduces the invasive ability of trophoblasts, and may be associated with severe hypertension, which exacerbates PE [193].
Diagnostic value of LncRNAs in PE
Table 5 reveals that lncRNAs may serve as potential diagnostic biomarkers for PE through ROC curve analysis. In placenta, Wu et al. have found that ROC of lnc TCL6 could reach to 0.8625 [138]. The ROC values of lnc BC030099 in the whole blood cells were 0.713 [104] and that of serum lnc AF085938, and that of lnc G36948 and lnc AK002210 were 0.7673, 0.7956 and 0.7575 respectively [178].
Table 5.
Diagnostic value of lncRNAs in PE
| LncRNA | Sample type | Area under curve | Sensitivity | Specificity | Ref. |
|---|---|---|---|---|---|
| BC030099 | the wholel blood | 0.713 | / | / | [104] |
| NR_026824.1 | the wholel blood | 0.594 | / | / | [104] |
| AK055151.1 | the wholel blood | 0.512 | / | / | [104] |
| NR_027457 | the wholel blood | 0.532 | / | / | [104] |
| NR_024178 | the wholel blood | 0.542 | / | / | [104] |
| TCL6 | placenta | 0.8625 | / | / | [138] |
| MIR193BHG | placenta | 0.819 | / | / | [159] |
| GATA3-AS1 | placenta | 0.640 | / | / | [159] |
| PROX1-AS1 | placenta | 0.690 | / | / | [159] |
| NR_027457 | serum | 0.5633 | / | / | [178] |
| AF085938 | serum | 0.7673 | / | / | [178] |
| G36948 | serum | 0.7956 | / | / | [178] |
| AK002210 | serum | 0.7575 | / | / | [178] |
Association between lncRNA variants and the risk of PE
Few studies have been conducted on the relationship between lncRNA gene polymorphisms and PE susceptibility. There is a significant correlation between the HOTAIR rs4759314AG genotype and higher PE risk, and the HOTAIR rs10783618 polymorphism is associated with increased PE risk in recessive and allele models. However, HOTAIR gene polymorphisms rs12826786, rs920778, and rs1899663 are not associated with PE susceptibility. The CTGAC haplotype is associated with decreased risk of PE, whereas the CTGAT haplotype is associated with increased risk [194].
CircRNAs and preeclampsia
CircRNAs are covalently closed circular ncRNA molecules. They are resistant to degradation by nucleic acid exonucleases because they lack a 5′ terminal with a cap and a 3′ terminal with a poly(A) tail. These characteristics enable circRNAs to fulfill many biological functions, such as acting as molecular sponges for miRNA, regulating gene transcription and translation, and binding to RNA-binding proteins [195].
Expression pattern of circRNAs in patients with PE
In recent decades, researchers have confirmed that circRNAs are involved in a variety of diseases [196]. However, there have been few studies on the role of circRNAs in the pathogenesis of PE. Ou et al. discovered 49 differentially expressed circRNAs in the placental tissues of patients with severe PE, using RNA sequencing, and further verified the upregulated expression of hsa_circ_0001438, hsa_circ_0001326, and hsa_circ_32340 by quantitative PCR analysis. To determine the interaction between circRNAs and miRNAs, they conducted an analysis of the Kyoto Encyclopedia of Genes and Genomics database. They found that the MAPK signaling pathway was the most enriched pathway in terms of circRNAs, and that the circRNA–miRNA–mRNA interaction network generated by hsa_circ_0001438, hsa_circ_0001326, and hsa_circ_32340 might be involved in the pathogenesis of PE. They also found that miR-145-5p was closely associated with circRNAs and mRNAs [197]. Table 6 generalizes the aberrant expression of circRNA.
Table 6.
Dysregulation of circRNAs in PE
| CircRNA | Sample type | Status | Function | Ref. |
|---|---|---|---|---|
| hsa_circ_0001438 | placenta | upregulated | hsa_circ_0001438, hsa_circ_0001326, and hsa_circ_32340 were upregulated in the sPE patients and the circRNA-miRNA-mRNA interaction network generated with these three circRNAs revealed a broad regulatory network that might be involved in the pathogenesis of sPE | [197] |
| hsa_circ_0001326 | placenta | upregulated | ||
| hsa_circ_32340 | placenta | upregulated | ||
| hsa_circ_101,222 | placenta | upregulated | Plasma protein endoglin in combination with circ-101,222 strengthened the predictive power for pre-eclampsia | [198] |
| hsa_circ_SFXN1 | placenta | upregulated | CircSFXN1 overexpression significantly inhibited the invasion of TEV-1 trophoblasts and blocked the angiogenesis of human umbilical vein endothelial cells | [199] |
| hsa_circ_0011460 | placenta | upregulated | Circ-0011460 were involved in vasodilation, regulation of blood vessel size, protein transport and localization | [200] |
| hsa_circ_0088227 | placenta/plasma | downregulated | Knockdown of circPAPPA led to decreased proliferation and invasion in HTR8-S/Vneo trophoblast cells | [13] |
| hsa_circ_TNRC18 | placenta | upregulated | Circ-TNRC18 enhanced trophoblast cell migration and epithelial-mesenchymal transition | [15] |
| hsa_circ_0014736 | placenta | upregulated | the three altered circ-RNAs had a relationship with transcription regulation, proliferation, protein binding, and response to hypoxia | [201] |
| hsa_circ_0015382 | placenta | upregulated | ||
| hsa_circ_0007121 | placenta | downregulated | ||
| hsa_circ_100782 | placenta | upregulated | / | [202] |
| hsa_circ_102682 | placenta | upregulated | / | |
| hsa_circ_104820 | placenta | upregulated | / | |
| hsa_circ_0001855 | placenta | upregulated | Circ-0004904 and circ-0001855 combined with PAPP-A might be promising biomarkers for the detection of PE | [203] |
| hsa_circ_0004904 | placenta | upregulated | ||
| hsa_circ_3286 | placenta | downregulated | Circ-3286 significantly promoted HTR8/Svneo cell invasion. | [204] |
| hsa_circ_593 | placenta | downregulated | / | |
| hsa_circ_3800 | placenta | downregulated | / | |
| hsa_circ_3286 | plasma | downregulated | / | |
| hsa_circ_0036877 | plasma | upregulated | Circ-0036877 significantly increased apoptosis of syncytial trophoblasts in the PE placenta | [205] |
| placenta | downregulated | |||
| hsa_circ_0036878 | placenta | downregulated | / | |
| hsa_circ_0055724 | placenta | downregulated | / | |
| hsa_circ_0049730 | placenta | downregulated | / | |
| hsa_circ_0036474 | placenta | upregulated | / |
Diagnostic value of circRNAs in PE
In recent years, numerous studies have confirmed the value of plasma circRNAs as potential early biomarkers of PE. Using quantitative reverse transcription PCR, Hu et al. demonstrated that the levels of circ-0036877 in blood samples taken from patients with PE were significantly higher than those in the control group. Furthermore, ROC curve analysis suggested that plasma circ-0036877 is a potential early biomarker of PE: the area under the curve value was 0.846, the sensitivity was 85.3%, and the specificity was 72.7% [205]. Zhang et al. first published reports on the analysis of circRNA expression in blood cells. Analysis of red blood cell samples taken from 32 patients with PE and 32 healthy pregnant women revealed significantly higher circ-101,222 levels in patients with PE than in the healthy women: the area under the ROC curve, the sensitivity, and the specificity were 0.706, 65.61, and 68.54%, respectively [198]. To further verify the value of protein-bound circRNAs in the early diagnosis of PE, Bai et al. combined the plasma protein ENG with circRNAs. They found that the resulting area under the curve value increased to 0.876 (95% confidence interval (CI): 0.816–0.922), sensitivity increased to 70.73%, and specificity increased to 80.49%, compared to the corresponding values for the unbound circRNAs [201]. Table 7 summarizes the reported results.
Table 7.
Diagnostic values of circRNAs in PE
| CircRNA | Sample type | Area under curve | Sensitivity | Specificity | Ref. |
|---|---|---|---|---|---|
| hsa_circ_101,222 | placenta | 0.706 | 65.61% | 68.54% | [198] |
| hsa_circ_0007121 | placenta | 0.72 | 77% | 70% | [201] |
| hsa_circ_100782 | placenta | 0.653 | / | / | [202] |
| hsa_circ_102682 | placenta | 0.774 | / | / | |
| hsa_circ_104820 | placenta | 0.995 | / | / | |
| hsa_circ_0001855 | placenta | 0.621 | 53.33% | 70.00% | [203] |
| hsa_circ_0004904 | placenta | 0.611 | / | / | |
| hsa_circ_3286 | placenta | 0.764 | 80% | 68.60% | [201] |
| hsa_circ_0036877 | plasma | 0.846 | 85.30% | 72.70% | [205] |
Relationship between lncRNAs and miRNAs
As discussed above, both lncRNAs and miRNAs have regulatory effects on the pathogenesis of PE. Moreover, they are interrelated and interactive. lncRNAs can act as competing endogenous RNAs to influence the bioavailability of miRNAs [9]. Gao et al. first reported that H19 promoted the expression of miR-let-7 and downregulated miRNA-675, which resulted in the migration and invasion of extravillous trophoblasts in vitro [150]. Zuckerwise et al. also showed that the downregulation of H19 suppressed the Par6/Smurf1/RhoA pathway activated by TβR3 to reduce TGF-β signaling. TβR3 is considered a downstream mediator of the interaction between H19 and let-7 [152]. There have been numerous investigations into the role of the lncRNA–miRNA–mRNA axis in the pathogenesis of PE. For example, Tan et al. reported that DLX6-AS1 lncRNA may contribute to PE by suppressing the proliferation, migration, and invasion of trophoblasts via the miR-376c–GADD45A axis [155]. Li et al. were the first to report that lncRNA TUG1 causes the development of PE by acting as a molecular sponge for miRNA-29b, thereby regulating the expression of MCL1, VEGFA, and MMP2 [14]. Yu et al. discovered that lncRNA TUG1 can also act as a molecular sponge for miR-204-5p, and downregulated lncRNA TUG1 suppresses trophoblast migration and invasion, partly by sponging miR-204-5p [144]. Figure 1 depicts the molecular mechanism by which ncRNA affects the pathogenesis of PE.
Fig. 1.

lncRNAs and circRNAs can act as a competing endogenous RNA(ceRNA), binding with miRNAs and regulating the effects of miRNAs on target genes, to influence trophoblast cell proliferation, invasion, migration and apoptosis and might play a role in angiogenesis and cell population at the G 0 /G 1 phase
Relationship between circRNAs and miRNAs
CircRNAs that contain miRNA response elements can serve as competing endogenous RNAs by binding with miRNAs. They act as miRNA sponges in cells, thereby regulating the effects of miRNAs on target genes and altering their expression levels [9]. In 2013, Hansen et al. found that ciRS-7, which is a circRNA sponge for miR-7 derived from the CDR1 gene, can bind and adsorb miR-7, thereby reducing its activity and indirectly upregulating the expression of miR-7-related target genes [206]. circRNAs have a stronger potential to adsorb miRNAs in the body than linear mRNAs or lncRNAs, because they are more stable. A few researchers have reported the occurrence of PE involving the interaction between circRNAs and miRNAs. circ-PAPPA, which is downregulated in both the placentas and plasma of patients with PE, can directly target miR-384 and act as a sponge for it. Finally, miR-384 overexpression inhibits the proliferation and invasion of trophoblasts by targeting STAT3 [13]. Shen et al. reported that upregulated circ-TNRC18 in the placental tissues of patients with PE also combined with miR-762 to target GRHL2 protein to regulate trophoblast epithelial–mesenchymal transition and invasion [15]. Although the role of circRNAs in the pathogenesis of PE is not fully understood, there is new evidence that they act as molecular sponges for miRNAs. Figure 2 illustrates the relationships between the various ncRNAs.
Fig. 2.
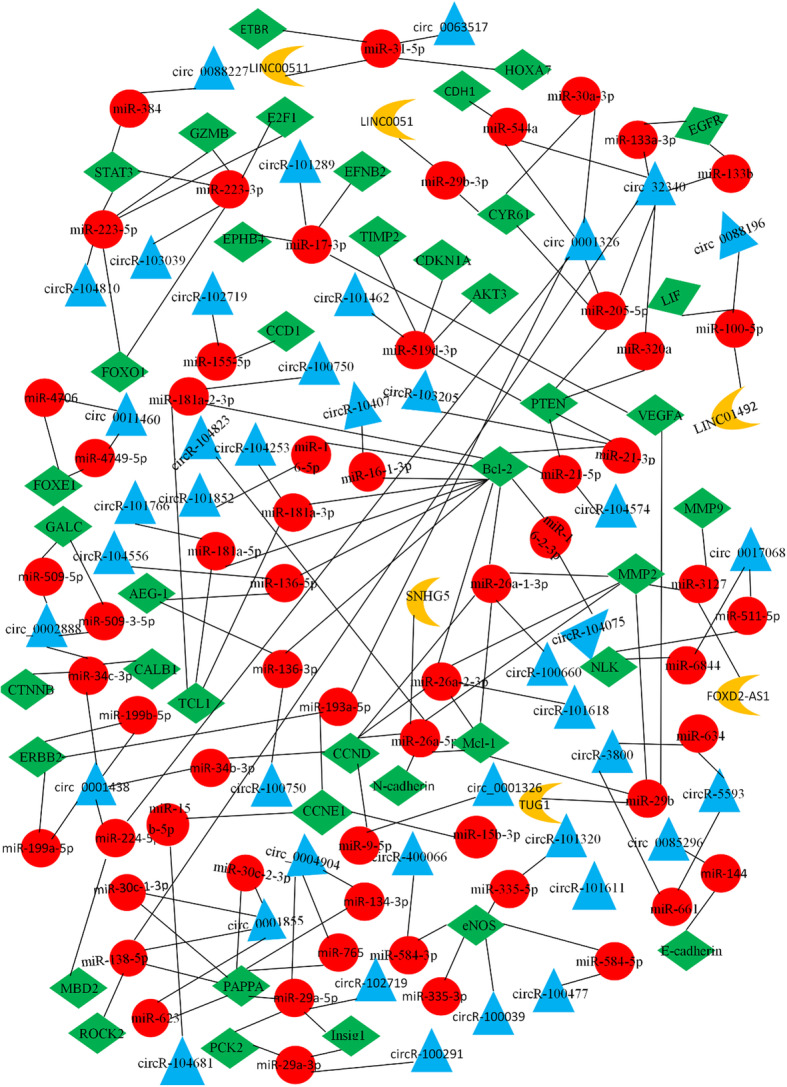
Mechanisms of interaction of ncRNA in PE. Green: protein coding, Yellow: lncRNA, Red: miRNA, Blue: CircRNA
Discussion
The heterogeneity and complexity of PE make its diagnosis, prediction, and treatment difficult. As it is currently not possible to detect the molecular signature of the main affected organ, i.e., the placenta, until the termination of pregnancy, it is difficult to monitor the progression of PE in a timely manner. Therefore, markers that circulate in the peripheral blood have great potential for noninvasive monitoring. It is currently possible to detect numerous biochemical markers in the serum, such as placental growth factor, soluble FMS-like tyrosine kinase receptor 1, placental protein 13, and placental protein A; however, their sensitivity and specificity are low [207]. Molecular biomarkers could provide a more reliable platform for the screening and diagnosis of PE than biochemical markers. In particular, the ncRNAs in the maternal peripheral blood are expected to be useful noninvasive biomarkers.
The differential expression of ncRNAs has been investigated to confirm their involvement in the pathogenesis of PE. Following numerous studies on the abnormal expression of ncRNAs in placental tissues, studies have been carried out to investigate ncRNAs in the peripheral blood of patients with PE. For example, Li et al. separated exosomes from maternal plasma by continuous density gradient hypercentrifugation, and found that seven miRNAs were differentially expressed in the exosomes from women with PE and those from a control group; however, the source of these exosomes was not determined [208]. It has subsequently been reported that exosomes derived from human umbilical cord mesenchymal stem cells that overexpress miR-139-5P, can promote trophoblast migration, invasion, and proliferation, and prevent apoptosis [209]. Studies on placenta-specific miRNA clusters in plasma samples revealed that the overexpression of miR-517-5p, miR-518b, and miR-520 h was associated with the late development of PE, and the screening of plasma miR-517-5p in early pregnancy also identified a proportion of women with subsequent PE [124]. Furthermore, Sun et al. performed univariate and multivariate analyses on the upregulation of lncRNA BC030099 in the whole blood of patients with PE, and determined that lncRNA BC030099 was an independent predictor of PE [137].
LncRNAs can regulate miRNA activity, and post-transcriptional regulation will affect the expression and function of their target mRNAs. LncRNAs have been shown to have miRNA binding sites --miRNA responsible elements, and they may potentially sponge the miRNAs, Thus, miRNA-mediated post-transcriptional regulation of target mRNAs was impaired. Dong et al. have demonstrated that LINC00511 regulates the proliferation, apoptosis, invasion and autophagy of trophoblast cells to mediate PE progression through modulating the miR-31-5p/homeobox protein A7 axis through dual luciferase reporter gene analysis [176]. When circRNAs interacted with miRNAs, they formed miRNA molecular sponges that further inhibited the transcript and lead to gene silencing. Due to the complementarity between bases, miRNAs bound to target mRNAs and performed transcriptional silencing to regulate gene expression. However, Li et al. confirmed circ_0063517 acts as ceRNA,targeting the miR-31-5p-ETBR axis to regulate angiogenesis of vascular endothelial cells in PE by dual luciferase reporting system and RNA immunoprecipitation (RIP) analysis [210]. Because lnc00511 and circ0063517 played an important role in the occurrence and development of PE through the bridge relationship of miR-31-5p, we therefore darw the conclusion that circRNA was associated with lncRNA through miRNA. In addition to that, lnc00511 functioned as a molecular spong for miR-29b-3p,antagonizing its ability to repress Cyr61 protein translation, and meanwhile overexpression of lnc00511 promoted trophoblast cell proliferation, migration and invasion [177].It is through this network that miRNA, lncRNA and circRNA are inseparable and jointly promote the occurrence and development of PE.
Although numerous studies have confirmed the differential expression of ncRNAs in placental tissues, and their pathogenic mechanism in PE, studies on ncRNAs in the peripheral blood, especially circRNAs and lncRNAs, remain scarce. More research is required to elucidate the key role of ncRNAs in PE, because they are potential stable biomarkers for the diagnosis of the disorder.
Conclusions
The present review summarizes the expression patterns of ncRNAs, i.e., microRNAs (miRNAs), long noncoding RNAs (lncRNAs), and circular RNAs (circRNAs), and the mechanisms by which they affect PE. We examine the clinical significance of ncRNAs as biomarkers for the diagnosis of PE, and discuss the contributions made to PE by genetic polymorphisms and epigenetic ncRNA regulation. We believe that our study makes a significant contribution to the literature because it highlights the clinical value of ncRNAs as noninvasive biomarkers of PE.
Acknowledgements
The authors thank Editage (www.editage.cn) for English language editing.
Abbreviations
- CircRNA
Circular RNA
- LncRNA
Long noncoding RNA
- MSCs
Mesenchymal stem cells
- MiRNA
MicroRNA
- NcRNA
Noncoding RNA
- PiRNA
Piwi-interacting RNA
- PE
Preeclampsia
- ROC
Receiver operating characteristic
- SNP
Single nucleotide polymorphism
- SiRNA
Small interfering RNA
- SnRNA
Small nuclear RNA
Authors’ contributions
Ningxia Sun and Shiting Qin contributed to data collection and article writing. Lu Zhang and Shiguo Liu assisted in designing the study and revising the article. All authors have read and approved the content of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China [No. 82071667]; Natural Science Fund Project of Shandong Province [ZR2019MH127]; and Key research and development plan of Shandong Province [2019GSF108106].
Availability of data and materials
Datasets are available through the corresponding author upon reasonable request.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Ningxia Sun and Shiting Qin contributed equally to this work.
Contributor Information
Lu Zhang, Email: 0130aici@163.com.
Shiguo Liu, Email: sarah9262020@163.com.
References
- 1.Wolters Kluwer health. ACOG Practice Bulletin No. 202: gestational hypertension and preeclampsia. Obstet Gynecol. 2019;133(1):e1–e25. [DOI] [PubMed]
- 2.Dekker GA, Sibai BM. Etiology and pathogenesis of preeclampsia: current concepts. Am J Obstet Gynecol. 1998;179(5):1359–1375. doi: 10.1016/S0002-9378(98)70160-7. [DOI] [PubMed] [Google Scholar]
- 3.Leung DN, et al. Increased placental apoptosis in pregnancies complicated by preeclampsia. Am J Obstet Gynecol. 2001;184(6):1249–1250. doi: 10.1067/mob.2001.112906. [DOI] [PubMed] [Google Scholar]
- 4.Sargent IL, et al. Trophoblast deportation and the maternal inflammatory response in pre-eclampsia. J Reprod Immunol. 2003;59(2):153–160. doi: 10.1016/S0165-0378(03)00044-5. [DOI] [PubMed] [Google Scholar]
- 5.Levine RJ, Lam C, Qian C, Yu KF, Maynard SE, Sachs BP, Sibai BM, Epstein FH, Romero R, Thadhani R, Karumanchi SA. Soluble endoglin and other circulating antiangiogenic factors in preeclampsia. N Engl J Med. 2006;355(10):992–1005. doi: 10.1056/NEJMoa055352. [DOI] [PubMed] [Google Scholar]
- 6.Cai M, Kolluru GK, Ahmed A. Small Molecule, Big Prospects: MicroRNA in Pregnancy and Its Complications. J Pregnancy. 2017;2017:6972732. doi: 10.1155/2017/6972732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bonasio R, Tu S, Reinberg D. Molecular signals of epigenetic states. Science. 2010;330(6004):612–616. doi: 10.1126/science.1191078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hemberg M, et al. Integrated genome analysis suggests that most conserved non-coding sequences are regulatory factor binding sites. Nucleic Acids Res. 2012;40(16):7858–7869. doi: 10.1093/nar/gks477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Anastasiadou E, Jacob LS, Slack FJ. Non-coding RNA networks in cancer. Nat Rev Cancer. 2018;18(1):5–18. doi: 10.1038/nrc.2017.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang S, et al. MiR-101 inhibits migration and invasion of trophoblast HTR-8/SVneo cells by targeting CXCL6 in preeclampsia. Minerva Med. 2021;112(2):302-3. [DOI] [PubMed]
- 11.Liu R, Wang X, Yan Q. The regulatory network of lncRNA DLX6-AS1/miR-149-5p/ERP44 is possibly related to the progression of preeclampsia. Placenta. 2020;93:34–42. doi: 10.1016/j.placenta.2020.02.001. [DOI] [PubMed] [Google Scholar]
- 12.Xie N, Jia Z, Li L. miR-320a upregulation contributes to the development of preeclampsia by inhibiting the growth and invasion of trophoblast cells by targeting interleukin 4. Mol Med Rep. 2019;20(4):3256–3264. doi: 10.3892/mmr.2019.10574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhou W, et al. Down-regulated circPAPPA suppresses the proliferation and invasion of trophoblast cells via the miR-384/STAT3 pathway. Biosci Rep. 2019;39(9). [DOI] [PMC free article] [PubMed]
- 14.Li Q, et al. lncRNA TUG1 modulates proliferation, apoptosis, invasion, and angiogenesis via targeting miR-29b in trophoblast cells. Hum Genomics. 2019;13(1):50. doi: 10.1186/s40246-019-0237-z. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 15.Shen XY, et al. CircTRNC18 inhibits trophoblast cell migration and epithelial-mesenchymal transition by regulating miR-762/Grhl2 pathway in pre-eclampsia. RNA Biol. 2019;16(11):1565–1573. doi: 10.1080/15476286.2019.1644591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dai Y, et al. MicroRNA-155 inhibits proliferation and migration of human extravillous trophoblast derived HTR-8/SVneo cells via down-regulating cyclin D1. Placenta. 2012;33(10):824–829. doi: 10.1016/j.placenta.2012.07.012. [DOI] [PubMed] [Google Scholar]
- 17.Esteller M. Non-coding RNAs in human disease. Nat Rev Genet. 2011;12(12):861–874. doi: 10.1038/nrg3074. [DOI] [PubMed] [Google Scholar]
- 18.Anton L, et al. miR-210 inhibits trophoblast invasion and is a serum biomarker for preeclampsia. Am J Pathol. 2013;183(5):1437–1445. doi: 10.1016/j.ajpath.2013.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li J, et al. Roles of miR-210 in the pathogenesis of pre-eclampsia. Arch Med Sci. 2019;15(1):183–190. doi: 10.5114/aoms.2018.73129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Luo R, et al. MicroRNA-210 contributes to preeclampsia by downregulating potassium channel modulatory factor 1. Hypertension. 2014;64(4):839–845. doi: 10.1161/HYPERTENSIONAHA.114.03530. [DOI] [PubMed] [Google Scholar]
- 21.Wang R, et al. MicroRNA-210 regulates human trophoblast cell line HTR-8/SVneo function by attenuating Notch1 expression: implications for the role of microRNA-210 in pre-eclampsia. Mol Reprod Dev. 2019;86(7):896–907. doi: 10.1002/mrd.23154. [DOI] [PubMed] [Google Scholar]
- 22.Luo R, et al. Hypoxia-inducible miR-210 contributes to preeclampsia via targeting thrombospondin type I domain containing 7A. Sci Rep. 2016;6:19588. doi: 10.1038/srep19588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang Y, et al. Aberrantly up-regulated miR-20a in pre-eclampsic placenta compromised the proliferative and invasive behaviors of trophoblast cells by targeting forkhead box protein A1. Int J Biol Sci. 2014;10(9):973–982. doi: 10.7150/ijbs.9088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fang YN, et al. Highly expressed miR-182-5p can promote preeclampsia progression by degrading RND3 and inhibiting HTR-8/SVneo cell invasion. Eur Rev Med Pharmacol Sci. 2018;22(20):6583–6590. doi: 10.26355/eurrev_201810_16132. [DOI] [PubMed] [Google Scholar]
- 25.Zhang Y, et al. MicroRNA-155 contributes to preeclampsia by down-regulating CYR61. Am J Obstet Gynecol. 2010;202(5):466.e1–466.e7. doi: 10.1016/j.ajog.2010.01.057. [DOI] [PubMed] [Google Scholar]
- 26.Li X, et al. MicroRNA-155 inhibits migration of trophoblast cells and contributes to the pathogenesis of severe preeclampsia by regulating endothelial nitric oxide synthase. Mol Med Rep. 2014;10(1):550–554. doi: 10.3892/mmr.2014.2214. [DOI] [PubMed] [Google Scholar]
- 27.Wu H, et al. MiR-195 participates in the placental disorder of preeclampsia via targeting activin receptor type-2B in trophoblastic cells. J Hypertens. 2016;34(7):1371–1379. doi: 10.1097/HJH.0000000000000948. [DOI] [PubMed] [Google Scholar]
- 28.Wang Y, et al. miR-16 inhibits the proliferation and angiogenesis-regulating potential of mesenchymal stem cells in severe pre-eclampsia. FEBS J. 2012;279(24):4510–24. doi: 10.1111/febs.12037. [DOI] [PubMed] [Google Scholar]
- 29.Yuan Y, et al. MicroRNA-16 is involved in the pathogenesis of pre-eclampsia via regulation of Notch2. J Cell Physiol. 2020;235(5):4530–4544. doi: 10.1002/jcp.29330. [DOI] [PubMed] [Google Scholar]
- 30.Fu G, et al. MicroRNA-376c impairs transforming growth factor-β and nodal signaling to promote trophoblast cell proliferation and invasion. Hypertension. 2013;61(4):864–872. doi: 10.1161/HYPERTENSIONAHA.111.203489. [DOI] [PubMed] [Google Scholar]
- 31.Li J, et al. Expression of miR-376 in blood of pregnant women with preeclampsia and its effect on 25-hydroxyvitamin D. Exp Ther Med. 2018;16(3):1701–1706. doi: 10.3892/etm.2018.6394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li P, et al. microRNA-29b contributes to pre-eclampsia through its effects on apoptosis, invasion and angiogenesis of trophoblast cells. Clin Sci (Lond) 2013;124(1):27–40. doi: 10.1042/CS20120121. [DOI] [PubMed] [Google Scholar]
- 33.Cui J, et al. MiR-101-containing extracellular vesicles bind to BRD4 and enhance proliferation and migration of trophoblasts in preeclampsia. Stem Cell Res Ther. 2020;11(1):231. doi: 10.1186/s13287-020-01720-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zou Y, et al. MiR-101 regulates apoptosis of trophoblast HTR-8/SVneo cells by targeting endoplasmic reticulum (ER) protein 44 during preeclampsia. J Hum Hypertens. 2014;28(10):610–616. doi: 10.1038/jhh.2014.35. [DOI] [PubMed] [Google Scholar]
- 35.Yang Y, et al. Inhibition of miR-18a increases expression of estrogen receptor 1 and promotes apoptosis in human HTR8 trophoblasts. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2017;33(8):1102–1107. [PubMed] [Google Scholar]
- 36.Zhu X, et al. Suppression of microRNA-18a expression inhibits invasion and promotes apoptosis of human trophoblast cells by targeting the estrogen receptor α gene. Mol Med Rep. 2015;12(2):2701–2706. doi: 10.3892/mmr.2015.3724. [DOI] [PubMed] [Google Scholar]
- 37.Xu P, et al. Variations of microRNAs in human placentas and plasma from preeclamptic pregnancy. Hypertension. 2014;63(6):1276–1284. doi: 10.1161/HYPERTENSIONAHA.113.02647. [DOI] [PubMed] [Google Scholar]
- 38.Hu TX, et al. MiR 20a,-20b and -200c are involved in hydrogen sulfide stimulation of VEGF production in human placental trophoblasts. Placenta. 2016;39:101–110. doi: 10.1016/j.placenta.2016.01.019. [DOI] [PubMed] [Google Scholar]
- 39.Li Q, et al. miR-125b-1-3p inhibits trophoblast cell invasion by targeting sphingosine-1-phosphate receptor 1 in preeclampsia. Biochem Biophys Res Commun. 2014;453(1):57–63. doi: 10.1016/j.bbrc.2014.09.059. [DOI] [PubMed] [Google Scholar]
- 40.Li Q, et al. Elevated microRNA-125b inhibits cytotrophoblast invasion and impairs endothelial cell function in preeclampsia. Cell Death Discov. 2020;6:35. doi: 10.1038/s41420-020-0269-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tang J, et al. MiR-125b participates in the occurrence of preeclampsia by regulating the migration and invasion of extravillous trophoblastic cells through STAT3 signaling pathway. J Recept Signal Transduct Res. 2021;41(2):202-8. [DOI] [PubMed]
- 42.Yang W, et al. miR-125b Enhances IL-8 Production in Early-Onset Severe Preeclampsia by Targeting Sphingosine-1-Phosphate Lyase 1. PLoS One. 2016;11(12):e0166940. doi: 10.1371/journal.pone.0166940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Licini C, et al. Pre-eclampsia predictive ability of maternal miR-125b: a clinical and experimental study. Transl Res. 2021;228:13-27. [DOI] [PubMed]
- 44.Hong F, Li Y, Xu Y. Decreased placental miR-126 expression and vascular endothelial growth factor levels in patients with pre-eclampsia. J Int Med Res. 2014;42(6):1243–1251. doi: 10.1177/0300060514540627. [DOI] [PubMed] [Google Scholar]
- 45.Yan T, et al. MicroRNA-126 regulates EPCs function: implications for a role of miR-126 in preeclampsia. J Cell Biochem. 2013;114(9):2148–2159. doi: 10.1002/jcb.24563. [DOI] [PubMed] [Google Scholar]
- 46.Campos CB, et al. Reduced circulating miR-196b levels is associated with preeclampsia. Pregnancy Hypertens. 2014;4(1):11–13. doi: 10.1016/j.preghy.2013.10.002. [DOI] [PubMed] [Google Scholar]
- 47.Akehurst C, et al. Differential expression of microRNA-206 and its target genes in preeclampsia. J Hypertens. 2015;33(10):2068–2074. doi: 10.1097/HJH.0000000000000656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sheng C, Zhao Y, Zhu L. Down-regulation of EDN1 gene expression by circulating miR-206 is associated with risk of preeclampsia. Medicine (Baltimore) 2020;99(22):e20319. doi: 10.1097/MD.0000000000020319. [DOI] [PubMed] [Google Scholar]
- 49.Wu HY, et al. LncRNA MALAT1 regulates trophoblast cells migration and invasion via miR-206/IGF-1 axis. Cell Cycle. 2020;19(1):39–52. doi: 10.1080/15384101.2019.1691787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Motawi TMK, et al. Role of mesenchymal stem cells exosomes derived microRNAs; miR-136, miR-494 and miR-495 in pre-eclampsia diagnosis and evaluation. Arch Biochem Biophys. 2018;659:13–21. doi: 10.1016/j.abb.2018.09.023. [DOI] [PubMed] [Google Scholar]
- 51.Chen S, et al. MicroRNA-494 inhibits the growth and angiogenesis-regulating potential of mesenchymal stem cells. FEBS Lett. 2015;589(6):710–717. doi: 10.1016/j.febslet.2015.01.038. [DOI] [PubMed] [Google Scholar]
- 52.Ding J, et al. MiR-519d-3p suppresses invasion and migration of trophoblast cells via targeting MMP-2. PLoS One. 2015;10(3):e0120321. doi: 10.1371/journal.pone.0120321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Jiang F, et al. Upregulation of microRNA-335 and microRNA-584 contributes to the pathogenesis of severe preeclampsia through downregulation of endothelial nitric oxide synthase. Mol Med Rep. 2015;12(4):5383–5390. doi: 10.3892/mmr.2015.4018. [DOI] [PubMed] [Google Scholar]
- 54.Lu W, et al. ROS/p53/miR-335-5p/Sp1 axis modulates the migration and epithelial to mesenchymal transition of JEG-3 cells. Mol Med Rep. 2020;21(3):1208–1216. doi: 10.3892/mmr.2019.10901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sun M, et al. MicroRNA-34a inhibits human trophoblast cell invasion by targeting MYC. BMC Cell Biol. 2015;16:21. doi: 10.1186/s12860-015-0068-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Guo M, et al. Elevated microRNA-34a contributes to trophoblast cell apoptosis in preeclampsia by targeting BCL-2. J Hum Hypertens. 2017;31(12):815–820. doi: 10.1038/jhh.2017.65. [DOI] [PubMed] [Google Scholar]
- 57.Liu JJ, et al. Influence of miR-34a on preeclampsia through the notch signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(3):923–931. doi: 10.26355/eurrev_201902_16978. [DOI] [PubMed] [Google Scholar]
- 58.Yu Y, et al. MicroRNA-204 suppresses trophoblast-like cell invasion by targeting matrix metalloproteinase-9. Biochem Biophys Res Commun. 2015;463(3):285–291. doi: 10.1016/j.bbrc.2015.05.052. [DOI] [PubMed] [Google Scholar]
- 59.Zhou X, et al. The aberrantly expressed miR-193b-3p contributes to preeclampsia through regulating transforming growth factor-β signaling. Sci Rep. 2016;6:19910. doi: 10.1038/srep19910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Awamleh Z, Han VKM. Potential pathophysiological role of microRNA 193b-5p in human placentae from pregnancies complicated by preeclampsia and intrauterine growth restriction. Mol Biol Rep. 2020;47(9):6531-44. [DOI] [PubMed]
- 61.Dong Q, et al. OS073. Over-expression of MIRNA-203 results in increased inflammatory response in endothelial cells: a mechanism of increased endothelial inflammatory response in preeclampsia. Pregnancy Hypertens. 2012;2(3):217. doi: 10.1016/j.preghy.2012.04.074. [DOI] [PubMed] [Google Scholar]
- 62.Liu F, et al. miR-203 contributes to pre-eclampsia via inhibition of VEGFA expression. Mol Med Rep. 2018;17(4):5627–5634. doi: 10.3892/mmr.2018.8558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang Y, et al. Up-regulation of miR-203 expression induces endothelial inflammatory response: potential role in preeclampsia. Am J Reprod Immunol. 2016;76(6):482–490. doi: 10.1111/aji.12589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ma HY, et al. MiRNA-203a-3p inhibits inflammatory response in preeclampsia through regulating IL24. Eur Rev Med Pharmacol Sci. 2020;24(10):5223–5230. doi: 10.26355/eurrev_202005_21304. [DOI] [PubMed] [Google Scholar]
- 65.Sandrim VC, et al. Circulating microRNA expression profiles in pre-eclampsia: evidence of increased miR-885-5p levels. Bjog. 2016;123(13):2120–2128. doi: 10.1111/1471-0528.13903. [DOI] [PubMed] [Google Scholar]
- 66.Ospina-Prieto S, et al. MicroRNA-141 is upregulated in preeclamptic placentae and regulates trophoblast invasion and intercellular communication. Transl Res. 2016;172:61–72. doi: 10.1016/j.trsl.2016.02.012. [DOI] [PubMed] [Google Scholar]
- 67.Wu D, et al. Hypoxia-induced microRNA-141 regulates trophoblast apoptosis, invasion, and vascularization by blocking CXCL12β/CXCR2/4 signal transduction. Biomed Pharmacother. 2019;116:108836. doi: 10.1016/j.biopha.2019.108836. [DOI] [PubMed] [Google Scholar]
- 68.Wang Y, et al. miR-141-5p regulate ATF2 via effecting MAPK1/ERK2 signaling to promote preeclampsia. Biomed Pharmacother. 2019;115:108953. doi: 10.1016/j.biopha.2019.108953. [DOI] [PubMed] [Google Scholar]
- 69.Ding GC, et al. MicroRNA-128a-induced apoptosis in HTR-8/SVneo trophoblast cells contributes to pre-eclampsia. Biomed Pharmacother. 2016;81:63–70. doi: 10.1016/j.biopha.2016.03.040. [DOI] [PubMed] [Google Scholar]
- 70.Chi Z, Zhang M. Exploration of the regulation and control mechanisms of miR-145 in trophoblast cell proliferation and invasion. Exp Ther Med. 2018;16(6):5298–5304. doi: 10.3892/etm.2018.6890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ji L, et al. MiR-136 contributes to pre-eclampsia through its effects on apoptosis and angiogenesis of mesenchymal stem cells. Placenta. 2017;50:102–109. doi: 10.1016/j.placenta.2017.01.102. [DOI] [PubMed] [Google Scholar]
- 72.Jiang L, et al. Elevated microRNA-520g in pre-eclampsia inhibits migration and invasion of trophoblasts. Placenta. 2017;51:70–75. doi: 10.1016/j.placenta.2017.02.001. [DOI] [PubMed] [Google Scholar]
- 73.Jin M, et al. MicroRNA-20b inhibits trophoblast cell migration and invasion by targeting MMP-2. Int J Clin Exp Pathol. 2017;10(11):10901–10909. [PMC free article] [PubMed] [Google Scholar]
- 74.Fan Y, et al. Elevated miR-23a impairs trophoblast migration and invasiveness through HDAC2 inhibition and NF-κB activation. Life Sci. 2020;261:118358. doi: 10.1016/j.lfs.2020.118358. [DOI] [PubMed] [Google Scholar]
- 75.Li L, et al. Lentivirus-mediated miR-23a overexpression induces trophoblast cell apoptosis through inhibiting X-linked inhibitor of apoptosis. Biomed Pharmacother. 2017;94:412–417. doi: 10.1016/j.biopha.2017.07.082. [DOI] [PubMed] [Google Scholar]
- 76.Li X, et al. MiR-495 promotes senescence of mesenchymal stem cells by targeting Bmi-1. Cell Physiol Biochem. 2017;42(2):780–796. doi: 10.1159/000478069. [DOI] [PubMed] [Google Scholar]
- 77.Lu TM, Lu W, Zhao LJ. MicroRNA-137 affects proliferation and migration of placenta trophoblast cells in preeclampsia by targeting ERRα. Reprod Sci. 2017;24(1):85–96. doi: 10.1177/1933719116650754. [DOI] [PubMed] [Google Scholar]
- 78.Pan Q, et al. Invasion of trophoblast cell lines is inhibited by miR-93 via MMP-2. Placenta. 2017;53:48–53. doi: 10.1016/j.placenta.2017.03.008. [DOI] [PubMed] [Google Scholar]
- 79.Xiao J, et al. miR-144 may regulate the proliferation, migration and invasion of trophoblastic cells through targeting PTEN in preeclampsia. Biomed Pharmacother. 2017;94:341–353. doi: 10.1016/j.biopha.2017.07.130. [DOI] [PubMed] [Google Scholar]
- 80.Hu S, et al. MicroRNA-144-3p may participate in the pathogenesis of preeclampsia by targeting Cox-2. Mol Med Rep. 2019;19(6):4655–4662. doi: 10.3892/mmr.2019.10150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Zhang Y, Huang G, Zhang Y, Yang H, Long Y, Liang Q, Zheng Z. MiR-942 decreased before 20 weeks gestation in women with preeclampsia and was associated with the pathophysiology of preeclampsia in vitro. Clin Exp Hypertens. 2017;39(2):108–113. doi: 10.1080/10641963.2016.1210619. [DOI] [PubMed] [Google Scholar]
- 82.Zou AX, et al. MiR-134 inhibits infiltration of trophoblast cells in placenta of patients with preeclampsia by decreasing ITGB1 expression. Eur Rev Med Pharmacol Sci. 2018;22(8):2199–2206. doi: 10.26355/eurrev_201804_14804. [DOI] [PubMed] [Google Scholar]
- 83.Wang N, et al. miR-362-3p regulates cell proliferation, migration and invasion of trophoblastic cells under hypoxia through targeting Pax3. Biomed Pharmacother. 2018;99:462–468. doi: 10.1016/j.biopha.2018.01.089. [DOI] [PubMed] [Google Scholar]
- 84.Shi Z, et al. MicroRNA-454 contributes to sustaining the proliferation and invasion of trophoblast cells through inhibiting nodal/ALK7 signaling in pre-eclampsia. Chem Biol Interact. 2019;298:8–14. doi: 10.1016/j.cbi.2018.10.012. [DOI] [PubMed] [Google Scholar]
- 85.Wang F, Yan J. MicroRNA-454 is involved in regulating trophoblast cell proliferation, apoptosis, and invasion in preeclampsia by modulating the expression of ephrin receptor B4. Biomed Pharmacother. 2018;107:746–753. doi: 10.1016/j.biopha.2018.08.055. [DOI] [PubMed] [Google Scholar]
- 86.Niu ZR, et al. MicroRNA-30a-3p is overexpressed in the placentas of patients with preeclampsia and affects trophoblast invasion and apoptosis by its effects on IGF-1. Am J Obstet Gynecol. 2018;218(2):249.e1–249.e12. doi: 10.1016/j.ajog.2017.11.568. [DOI] [PubMed] [Google Scholar]
- 87.Kim S, et al. NF-κB-responsive miRNA-31-5p elicits endothelial dysfunction associated with preeclampsia via down-regulation of endothelial nitric-oxide synthase. J Biol Chem. 2018;293(49):18989–19000. doi: 10.1074/jbc.RA118.005197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Guo L, et al. MicroRNA-423-5p inhibits the progression of trophoblast cells via targeting IGF2BP1. Placenta. 2018;74:1–8. doi: 10.1016/j.placenta.2018.12.003. [DOI] [PubMed] [Google Scholar]
- 89.Gao Y, et al. Up-regulation of miR-299 suppressed the invasion and migration of HTR-8/SVneo trophoblast cells partly via targeting HDAC2 in pre-eclampsia. Biomed Pharmacother. 2018;97:1222–1228. doi: 10.1016/j.biopha.2017.11.053. [DOI] [PubMed] [Google Scholar]
- 90.Gao X, Li H, Wei JX. MiR-4421 regulates the progression of preeclampsia by regulating CYP11B2. Eur Rev Med Pharmacol Sci. 2018;22(6):1533–1540. doi: 10.26355/eurrev_201803_14557. [DOI] [PubMed] [Google Scholar]
- 91.Gao T, Deng M, Wang Q. MiRNA-320a inhibits trophoblast cell invasion by targeting estrogen-related receptor-gamma. J Obstet Gynaecol Res. 2018;44(4):756–763. doi: 10.1111/jog.13560. [DOI] [PubMed] [Google Scholar]
- 92.Fu JY, et al. Up-regulation of miR-517-5p inhibits ERK/MMP-2 pathway: potential role in preeclampsia. Eur Rev Med Pharmacol Sci. 2018;22(20):6599–6608. doi: 10.26355/eurrev_201810_16134. [DOI] [PubMed] [Google Scholar]
- 93.Nejad RMA, et al. Quantification of circulating miR-517c-3p and miR-210-3p levels in preeclampsia. Pregnancy Hypertens. 2019;16:75–78. doi: 10.1016/j.preghy.2019.03.004. [DOI] [PubMed] [Google Scholar]
- 94.Dai X, Cai Y. Down-regulation of microRNA let-7d inhibits the proliferation and invasion of trophoblast cells in preeclampsia. J Cell Biochem. 2018;119(1):1141–1151. doi: 10.1002/jcb.26282. [DOI] [PubMed] [Google Scholar]
- 95.Brkić J, et al. MicroRNA-218-5p promotes endovascular trophoblast differentiation and spiral artery remodeling. Mol Ther. 2018;26(9):2189–2205. doi: 10.1016/j.ymthe.2018.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Huang X, et al. Elevated MicroRNA-181a-5p contributes to trophoblast dysfunction and preeclampsia. Reprod Sci. 2019;26(8):1121–1129. doi: 10.1177/1933719118808916. [DOI] [PubMed] [Google Scholar]
- 97.Liu E, et al. MicroRNA-142-3p inhibits trophoblast cell migration and invasion by disrupting the TGF-β1/Smad3 signaling pathway. Mol Med Rep. 2019;19(5):3775–3782. doi: 10.3892/mmr.2019.9997. [DOI] [PubMed] [Google Scholar]
- 98.Lv Y, et al. miR-145-5p promotes trophoblast cell growth and invasion by targeting FLT1. Life Sci. 2019;239:117008. doi: 10.1016/j.lfs.2019.117008. [DOI] [PubMed] [Google Scholar]
- 99.Wen Z, et al. Tumor necrosis factor-alpha suppresses the invasion of HTR-8/SVneo trophoblast cells through microRNA-145-5p-mediated downregulation of Cyr61. Life Sci. 2018;209:132–139. doi: 10.1016/j.lfs.2018.08.005. [DOI] [PubMed] [Google Scholar]
- 100.Qian S, Liu R. miR-30b facilitates preeclampsia through targeting MXRA5 to inhibit the viability, invasion and apoptosis of placental trophoblast cells. Int J Clin Exp Pathol. 2019;12(11):4057–4065. [PMC free article] [PubMed] [Google Scholar]
- 101.Cao G, et al. MicroRNA regulation of transthyretin in trophoblast biofunction and preeclampsia. Arch Biochem Biophys. 2019;676:108129. doi: 10.1016/j.abb.2019.108129. [DOI] [PubMed] [Google Scholar]
- 102.Wang CY, et al. Elevated miR-200a and miR-141 inhibit endocrine gland-derived vascular endothelial growth factor expression and ciliogenesis in preeclampsia. J Physiol. 2019;597(12):3069–3083. doi: 10.1113/JP277704. [DOI] [PubMed] [Google Scholar]
- 103.Wang Z, et al. MiRNA-548c-5p downregulates inflammatory response in preeclampsia via targeting PTPRO. J Cell Physiol. 2019;234(7):11149–11155. doi: 10.1002/jcp.27758. [DOI] [PubMed] [Google Scholar]
- 104.Han X, et al. MiR-342-3p inhibition promotes cell proliferation and invasion by directly targeting ID4 in pre-eclampsia. J Obstet Gynaecol Res. 2020;46(1):49–57. doi: 10.1111/jog.14150. [DOI] [PubMed] [Google Scholar]
- 105.Yang X, Guo F. miR-342-3p suppresses cell migration and invasion in preeclampsia by targeting platelet-derived growth factor receptor α. Mol Med Rep. 2019;20(2):1772–1780. doi: 10.3892/mmr.2019.10372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Yang X, Meng T. MicroRNA-431 affects trophoblast migration and invasion by targeting ZEB1 in preeclampsia. Gene. 2019;683:225–232. doi: 10.1016/j.gene.2018.10.015. [DOI] [PubMed] [Google Scholar]
- 107.Yang Y, et al. MiR-221-3p is down-regulated in preeclampsia and affects trophoblast growth, invasion and migration partly via targeting thrombospondin 2. Biomed Pharmacother. 2019;109:127–134. doi: 10.1016/j.biopha.2018.10.009. [DOI] [PubMed] [Google Scholar]
- 108.Zhang L, et al. Correlation of MiR-152 expression with VEGF expression in placental tissue of preeclampsia rat and its influence on apoptosis of trophoblast cells. Eur Rev Med Pharmacol Sci. 2019;23(9):3553–3560. doi: 10.26355/eurrev_201905_17776. [DOI] [PubMed] [Google Scholar]
- 109.Zhang WM, et al. Effect of miR-133 on apoptosis of trophoblasts in human placenta tissues via rho/ROCK signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(24):10600–10608. doi: 10.26355/eurrev_201912_19755. [DOI] [PubMed] [Google Scholar]
- 110.Zhou W, et al. MiR-384 inhibits proliferation and migration of trophoblast cells via targeting PTBP3. Pregnancy Hypertens. 2020;21:132–138. doi: 10.1016/j.preghy.2020.05.017. [DOI] [PubMed] [Google Scholar]
- 111.Zhou F, et al. microRNA-21 regulates the proliferation of placental cells via FOXM1 in preeclampsia. Exp Ther Med. 2020;20(3):1871–1878. doi: 10.3892/etm.2020.8930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Xueya Z, et al. Exosomal encapsulation of miR-125a-5p inhibited trophoblast cell migration and proliferation by regulating the expression of VEGFA in preeclampsia. Biochem Biophys Res Commun. 2020;525(3):646–653. doi: 10.1016/j.bbrc.2020.02.137. [DOI] [PubMed] [Google Scholar]
- 113.Kim S, et al. Circulating miRNAs Associated with Dysregulated Vascular and Trophoblast Function as Target-Based Diagnostic Biomarkers for Preeclampsia. Cells. 2020;9(9). [DOI] [PMC free article] [PubMed]
- 114.Yang W, et al. Increased miRNA-518b inhibits trophoblast migration and angiogenesis by targeting EGR1 in early embryonic arrest†. Biol Reprod. 2019;101(4):664–674. doi: 10.1093/biolre/ioz109. [DOI] [PubMed] [Google Scholar]
- 115.Dong D, et al. microRNA-646 inhibits angiogenesis of endothelial progenitor cells in pre-eclamptic pregnancy by targeting the VEGF-A/HIF-1α axis. Exp Ther Med. 2020;20(3):1879–1888. doi: 10.3892/etm.2020.8929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Yang X, Meng T. miR-215-5p decreases migration and invasion of trophoblast cells through regulating CDC6 in preeclampsia. Cell Biochem Funct. 2020;38(4):472–479. doi: 10.1002/cbf.3492. [DOI] [PubMed] [Google Scholar]
- 117.Han L, et al. miR-483 is downregulated in pre-eclampsia via targeting insulin-like growth factor 1 (IGF1) and regulates the PI3K/Akt/mTOR pathway of endothelial progenitor cells. J Obstet Gynaecol Res. 2021;47(1):63-72. [DOI] [PubMed]
- 118.Pineles BL, et al. Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia. Am J Obstet Gynecol. 2007;196(3):261.e1–261.e6. doi: 10.1016/j.ajog.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 119.Gan L, et al. MiR-210 and miR-155 as potential diagnostic markers for pre-eclampsia pregnancies. Medicine (Baltimore) 2017;96(28):e7515. doi: 10.1097/MD.0000000000007515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Jelena M, et al. Placenta-specific plasma miR518b is a potential biomarker for preeclampsia. Clin Biochem. 2020;79:28–33. doi: 10.1016/j.clinbiochem.2020.02.012. [DOI] [PubMed] [Google Scholar]
- 121.Dong K, et al. Downregulations of circulating miR-31 and miR-21 are associated with preeclampsia. Pregnancy Hypertens. 2019;17:59–63. doi: 10.1016/j.preghy.2019.05.013. [DOI] [PubMed] [Google Scholar]
- 122.Gunel T, et al. Regulatory effect of miR-195 in the placental dysfunction of preeclampsia. J Matern Fetal Neonatal Med. 2020;33(6):901–908. doi: 10.1080/14767058.2018.1508439. [DOI] [PubMed] [Google Scholar]
- 123.Timofeeva AV, et al. Identification of potential early biomarkers of preeclampsia. Placenta. 2018;61:61–71. doi: 10.1016/j.placenta.2017.11.011. [DOI] [PubMed] [Google Scholar]
- 124.Hromadnikova I, et al. First trimester screening of circulating C19MC microRNAs and the evaluation of their potential to predict the onset of preeclampsia and IUGR. PLoS One. 2017;12(2):e0171756. doi: 10.1371/journal.pone.0171756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Maharaj NR, et al. MicroRNA-146a rs2910164 is associated with severe preeclampsia in black south African women on HAART. BMC Genet. 2017;18(1):5. doi: 10.1186/s12863-016-0469-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Salimi S, et al. The effect of miR-146a rs2910164 and miR-149 rs2292832 polymorphisms on preeclampsia susceptibility. Mol Biol Rep. 2019;46(4):4529–4536. doi: 10.1007/s11033-019-04908-2. [DOI] [PubMed] [Google Scholar]
- 127.Ayoub SE, et al. Association of MicroRNA-155rs767649 polymorphism with susceptibility to preeclampsia. Int J Mol Cell Med. 2019;8(4):247–257. doi: 10.22088/IJMCM.BUMS.8.4.247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Abo-Elmatty DM, Mehanna ET. MIR146A rs2910164 (G/C) polymorphism is associated with incidence of preeclampsia in gestational diabetes patients. Biochem Genet. 2019;57(2):222–233. doi: 10.1007/s10528-018-9886-1. [DOI] [PubMed] [Google Scholar]
- 129.Maharaj NR, et al. microRNA-27a rs895819 is associated with obesity in HIV infected preeclamptic Black South African women on HAART. BMC Med Genet. 2016;17(1):92. doi: 10.1186/s12881-016-0353-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Asadi-Tarani M, et al. The relationships between maternal and placental polymorphisms of miR-196a2 and miRNA-499 genes and preeclampsia. Br J Biomed Sci. 2020;77(4):191-5. [DOI] [PubMed]
- 131.Rokni M, et al. Association between miRNA-152 polymorphism and risk of preeclampsia susceptibility. Arch Gynecol Obstet. 2019;299(2):475–480. doi: 10.1007/s00404-018-4979-y. [DOI] [PubMed] [Google Scholar]
- 132.Eskandari F, et al. The association of pri-miRNA- 26a1 rs7372209 polymorphism and preeclampsia susceptibility. Clin Exp Hypertens. 2019;41(3):268–273. doi: 10.1080/10641963.2018.1469643. [DOI] [PubMed] [Google Scholar]
- 133.Rezaei M, et al. Hypomethylation of the miRNA-34a gene promoter is associated with Severe Preeclampsia. Clin Exp Hypertens. 2019;41(2):118–22. [DOI] [PubMed]
- 134.Zha W, et al. Let-7a inhibits Bcl-xl and YAP1 expression to induce apoptosis of trophoblast cells in early-onset severe preeclampsia. Sci Total Environ. 2020;745:139919. doi: 10.1016/j.scitotenv.2020.139919. [DOI] [PubMed] [Google Scholar]
- 135.Sekar D, et al. Methylation-dependent circulating microRNA 510 in preeclampsia patients. Hypertens Res. 2019;42(10):1647–1648. doi: 10.1038/s41440-019-0269-8. [DOI] [PubMed] [Google Scholar]
- 136.Sun Q, Hao Q, Prasanth KV. Nuclear long noncoding RNAs: key regulators of gene expression. Trends Genet. 2018;34(2):142–157. doi: 10.1016/j.tig.2017.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Sun Y, et al. Circulating lncRNA BC030099 increases in preeclampsia patients. Mol Ther Nucleic Acids. 2019;14:562–566. doi: 10.1016/j.omtn.2019.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.He X, et al. LncRNAs expression in preeclampsia placenta reveals the potential role of LncRNAs contributing to preeclampsia pathogenesis. PLoS One. 2013;8(11):e81437. doi: 10.1371/journal.pone.0081437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Zhao YH, et al. Long non-coding RNA HOTAIR modulates the progression of preeclampsia through inhibiting miR-106 in an EZH2-dependent manner. Life Sci. 2020;253:117668. doi: 10.1016/j.lfs.2020.117668. [DOI] [PubMed] [Google Scholar]
- 140.Zhang Y, He XY, Qin S, Mo HQ, Li X, Wu F, Zhang J, Li X, Mao L, Peng YQ, Guo YN, Lin Y, Tian FJ. Upregulation of PUM1 expression in preeclampsia impairs trophoblast invasion by negatively regulating the expression of the lncRNA HOTAIR. Mol Ther. 2020;28(2):631–641. doi: 10.1016/j.ymthe.2019.11.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Xu Y, et al. Down-regulated lncRNA AGAP2-AS1 contributes to pre-eclampsia AS a competing endogenous RNA for JDP2 by impairing trophoblastic phenotype. J Cell Mol Med. 2020;24(8):4557–4568. doi: 10.1111/jcmm.15113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Xu Y, et al. Downregulated lncRNA HOXA11-AS affects trophoblast cell proliferation and migration by regulating RND3 and HOXA7 expression in PE. Mol Ther Nucleic Acids. 2018;12:195–206. doi: 10.1016/j.omtn.2018.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Xu Y, Ge Z, Zhang E, Zuo Q, Huang S, Yang N, Wu D, Zhang Y, Chen Y, Xu H, Huang H, Jiang Z, Sun L. The lncRNA TUG1 modulates proliferation in trophoblast cells via epigenetic suppression of RND3. Cell Death Dis. 2017;8(10):e3104. doi: 10.1038/cddis.2017.503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Yu Y, et al. Long non-coding RNA TUG1 regulates the migration and invasion of trophoblast-like cells through sponging miR-204-5p. Clin Exp Pharmacol Physiol. 2019;46(4):380-88. [DOI] [PubMed]
- 145.Zou Y, et al. Upregulation of long noncoding RNA SPRY4-IT1 modulates proliferation, migration, apoptosis, and network formation in trophoblast cells HTR-8SV/neo. PLoS One. 2013;8(11):e79598. doi: 10.1371/journal.pone.0079598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Zuo Q, et al. The Lnc RNA SPRY4-IT1 modulates trophoblast cell invasion and migration by affecting the epithelial-mesenchymal transition. Sci Rep. 2016;6:37183. doi: 10.1038/srep37183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Yu L, et al. The role and molecular mechanism of long Nocoding RNA-MEG3 in the pathogenesis of preeclampsia. Reprod Sci. 2018;25(12):1619–1628. doi: 10.1177/1933719117749753. [DOI] [PubMed] [Google Scholar]
- 148.Wang R, Zou L. Downregulation of LncRNA-MEG3 promotes HTR8/SVneo cells apoptosis and attenuates its migration by repressing Notch1 signal in preeclampsia. Reproduction. 2020;160(1):21–29. doi: 10.1530/REP-19-0614. [DOI] [PubMed] [Google Scholar]
- 149.Zhang Y, et al. Down-regulated long non-coding RNA MEG3 and its effect on promoting apoptosis and suppressing migration of trophoblast cells. J Cell Biochem. 2015;116(4):542–550. doi: 10.1002/jcb.25004. [DOI] [PubMed] [Google Scholar]
- 150.Gao WL, et al. The imprinted H19 gene regulates human placental trophoblast cell proliferation via encoding miR-675 that targets nodal modulator 1 (NOMO1) RNA Biol. 2012;9(7):1002–1010. doi: 10.4161/rna.20807. [DOI] [PubMed] [Google Scholar]
- 151.Shu C, et al. Metformin exhibits its therapeutic effect in the treatment of pre-eclampsia via modulating the met/H19/miR-148a-5p/P28 and met/H19/miR-216-3p/EBI3 signaling pathways. Int Immunopharmacol. 2019;74:105693. doi: 10.1016/j.intimp.2019.105693. [DOI] [PubMed] [Google Scholar]
- 152.Zuckerwise L, et al. H19 long noncoding RNA alters trophoblast cell migration and invasion by regulating TβR3 in placentae with fetal growth restriction. Oncotarget. 2016;7(25):38398–38407. doi: 10.18632/oncotarget.9534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Xu J, et al. Overexpression of long non-coding RNA H19 promotes invasion and autophagy via the PI3K/AKT/mTOR pathways in trophoblast cells. Biomed Pharmacother. 2018;101:691–697. doi: 10.1016/j.biopha.2018.02.134. [DOI] [PubMed] [Google Scholar]
- 154.Cheng D, et al. The increased lncRNA MIR503HG in preeclampsia modulated trophoblast cell proliferation, invasion, and migration via regulating matrix metalloproteinases and NF-κB signaling. Dis Markers. 2019;2019:4976845. doi: 10.1155/2019/4976845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Tan Y, et al. Long non-coding RNA DLX6-AS1 is upregulated in preeclampsia and modulates migration and invasion of trophoblasts through the miR-376c/GADD45A axis. Exp Cell Res. 2018;370(2):718–724. doi: 10.1016/j.yexcr.2018.07.039. [DOI] [PubMed] [Google Scholar]
- 156.Gao Y, et al. The decreased lncRNA ZEB2-AS1 in pre-eclampsia controls the trophoblastic cell line HTR-8/SVneo's invasive and migratory abilities via the miR-149/PGF axis. J Cell Biochem. 2019;120(10):17677–17686. doi: 10.1002/jcb.29034. [DOI] [PubMed] [Google Scholar]
- 157.Jiao S, Wang SY, Huang Y. LncRNA PRNCR1 promoted the progression of eclampsia by regulating the MAPK signal pathway. Eur Rev Med Pharmacol Sci. 2018;22(12):3635–3642. doi: 10.26355/eurrev_201806_15240. [DOI] [PubMed] [Google Scholar]
- 158.Li S, et al. lncRNA GHET1 has effects in development of pre-eclampsia. J Cell Biochem. 2019;120(8):12647–12652. doi: 10.1002/jcb.28531. [DOI] [PubMed] [Google Scholar]
- 159.Gong F, et al. LncRNA TDRG1/miR-214-5p axis affects preeclampsia by modulating trophoblast cells. Cell Biochem Funct. 2020;38(4):352–361. doi: 10.1002/cbf.3480. [DOI] [PubMed] [Google Scholar]
- 160.Chen H, et al. Long non-coding RNA MALAT-1 is downregulated in preeclampsia and regulates proliferation, apoptosis, migration and invasion of JEG-3 trophoblast cells. Int J Clin Exp Pathol. 2015;8(10):12718–12727. [PMC free article] [PubMed] [Google Scholar]
- 161.Li Q, et al. LncRNA MALAT1 affects the migration and invasion of trophoblast cells by regulating FOS expression in early-onset preeclampsia. Pregnancy Hypertens. 2020;21:50–57. doi: 10.1016/j.preghy.2020.05.001. [DOI] [PubMed] [Google Scholar]
- 162.Wu JL, et al. Overexpression of lncRNA TCL6 promotes preeclampsia progression by regulating PTEN. Eur Rev Med Pharmacol Sci. 2019;23(10):4066–4072. doi: 10.26355/eurrev_201905_17907. [DOI] [PubMed] [Google Scholar]
- 163.Pengjie Z, et al. LncRNA uc003fir promotes CCL5 expression and negatively affects proliferation and migration of trophoblast cells in preeclampsia. Pregnancy Hypertens. 2018;14:90–96. doi: 10.1016/j.preghy.2018.08.449. [DOI] [PubMed] [Google Scholar]
- 164.Zhang Y, Chen X. lncRNA FOXD2-AS1 affects trophoblast cell proliferation, invasion and migration through targeting miRNA. Zygote. 2020;28(2):131–8. [DOI] [PubMed]
- 165.Chen FR, et al. Regulatory relationship between lncRNA KCNQ1OT1 and miR-146a-3p in preeclampsia. Zhonghua Fu Chan Ke Za Zhi. 2020;55(8):535–543. doi: 10.3760/cma.j.cn112141-20200322-00246. [DOI] [PubMed] [Google Scholar]
- 166.Yang Y, et al. The lncRNA small nucleolar RNA host gene 5 regulates trophoblast cell proliferation, invasion, and migration via modulating miR-26a-5p/N-cadherin axis. J Cell Biochem. 2019;120(3):3173–3184. doi: 10.1002/jcb.27583. [DOI] [PubMed] [Google Scholar]
- 167.Wang Q, et al. Down-regulated long non-coding RNA PVT1 contributes to gestational diabetes mellitus and preeclampsia via regulation of human trophoblast cells. Biomed Pharmacother. 2019;120:109501. doi: 10.1016/j.biopha.2019.109501. [DOI] [PubMed] [Google Scholar]
- 168.Xu Y, et al. The long non-coding RNA PVT1 represses ANGPTL4 transcription through binding with EZH2 in trophoblast cell. J Cell Mol Med. 2018;22(2):1272–1282. doi: 10.1111/jcmm.13405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Li R, et al. A potential regulatory network among WDR86-AS1, miR-10b-3p, and LITAF is possibly involved in preeclampsia pathogenesis. Cell Signal. 2019;55:40–52. doi: 10.1016/j.cellsig.2018.12.006. [DOI] [PubMed] [Google Scholar]
- 170.Liu X, et al. Down-regulated long non-coding RNA-ATB in preeclampsia and its effect on suppressing migration, proliferation, and tube formation of trophoblast cells. Placenta. 2017;49:80–87. doi: 10.1016/j.placenta.2016.12.004. [DOI] [PubMed] [Google Scholar]
- 171.Song X, et al. Long non-coding RNA RPAIN regulates the invasion and apoptosis of trophoblast cell lines via complement protein C1q. Oncotarget. 2017;8(5):7637–7646. doi: 10.18632/oncotarget.13826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Ma Y, et al. Long non-coding RNA NR_002794 is upregulated in pre-eclampsia and regulates the proliferation, apoptosis and invasion of trophoblast cells. Mol Med Rep. 2019;20(5):4567–4575. doi: 10.3892/mmr.2019.10701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Zou Y, et al. Promotion of trophoblast invasion by lncRNA MVIH through inducing Jun-B. J Cell Mol Med. 2018;22(2):1214–1223. doi: 10.1111/jcmm.13400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Liu J, Zhang Q, Ma N. LncRNA GASAL1 interacts with SRSF1 to regulate trophoblast cell proliferation, invasion, and apoptosis via the mTOR signaling pathway. Cell Transplant. 2020;29:963689720965182. doi: 10.1177/0963689720965182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Zhang Z, et al. Identification of key genes and long noncoding RNA-associated competing endogenous RNA (ceRNA) networks in early-onset preeclampsia. Biomed Res Int. 2020;2020:1673486. doi: 10.1155/2020/1673486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Dong X, et al. Long noncoding RNA LINC00511 regulates the proliferation, apoptosis, invasion and autophagy of trophoblast cells to mediate pre-eclampsia progression through modulating the miR-31-5p/homeobox protein A7 axis. J Obstet Gynaecol Res. 2020;46(8):1298–1309. doi: 10.1111/jog.14344. [DOI] [PubMed] [Google Scholar]
- 177.Quan X, et al. AP2γ mediated downregulation of lncRNA LINC00511 as a ceRNA suppresses trophoblast invasion by regulating miR-29b-3p/Cyr61 axis. Biomed Pharmacother. 2019;120:109269. doi: 10.1016/j.biopha.2019.109269. [DOI] [PubMed] [Google Scholar]
- 178.Luo X, Li X. Long non-coding RNAs serve as diagnostic biomarkers of preeclampsia and modulate migration and invasiveness of trophoblast cells. Med Sci Monit. 2018;24:84–91. doi: 10.12659/MSM.907808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Tian J, et al. Upregulated LncZBTB39 in pre-eclampsia and its effects on trophoblast invasion and migration via antagonizing the inhibition of miR-210 on THSD7A expression. Eur J Obstet Gynecol Reprod Biol. 2020;248:164–171. doi: 10.1016/j.ejogrb.2020.03.035. [DOI] [PubMed] [Google Scholar]
- 180.Zheng D, et al. Long non-coding RNA Gas5 is associated with preeclampsia and regulates biological behaviors of trophoblast via MicroRNA-21. Front Genet. 2020;11:188. doi: 10.3389/fgene.2020.00188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Zhao X, et al. Long noncoding RNA VIM antisense RNA 1 (VIM-AS1) plays an important role in development of preeclampsia by regulation of epithelial mesenchymal transition. Med Sci Monit. 2019;25:8306–8314. doi: 10.12659/MSM.916601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.He T, et al. lncRNA FAM99A is downregulated in preeclampsia and exerts a regulatory effect on trophoblast cell invasion, migration and apoptosis. Mol Med Rep. 2019;20(2):1451–1458. doi: 10.3892/mmr.2019.10350. [DOI] [PubMed] [Google Scholar]
- 183.Wu D, et al. lncRNA HIF1A antisense RNA 2 modulates trophoblast cell invasion and proliferation through upregulating PHLDA1 expression. Mol Ther Nucleic Acids. 2019;16:605–615. doi: 10.1016/j.omtn.2019.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Cheng D, et al. Upregulated long noncoding RNA Linc00261 in pre-eclampsia and its effect on trophoblast invasion and migration via regulating miR-558/TIMP4 signaling pathway. J Cell Biochem. 2019;120(8):13243–13253. doi: 10.1002/jcb.28598. [DOI] [PubMed] [Google Scholar]
- 185.Song X, et al. The long noncoding RNA uc.294 is upregulated in early-onset pre-eclampsia and inhibits proliferation, invasion of trophoblast cells (HTR-8/SVneo) J Cell Physiol. 2019;234(7):11001–11008. doi: 10.1002/jcp.27916. [DOI] [PubMed] [Google Scholar]
- 186.Wang N, Li R, Xue M. Potential regulatory network in the PSG10P/miR-19a-3p/IL1RAP pathway is possibly involved in preeclampsia pathogenesis. J Cell Mol Med. 2019;23(2):852–864. doi: 10.1111/jcmm.13985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Wu D, et al. Long noncoding RNA 00473 is involved in preeclampsia by LSD1 binding-regulated TFPI2 transcription in trophoblast cells. Mol Ther Nucleic Acids. 2018;12:381–392. doi: 10.1016/j.omtn.2018.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Cao C, et al. Long non-coding RNA Uc.187 is upregulated in preeclampsia and modulates proliferation, apoptosis, and invasion of HTR-8/SVneo trophoblast cells. J Cell Biochem. 2017;118(6):1462–1470. doi: 10.1002/jcb.25805. [DOI] [PubMed] [Google Scholar]
- 189.Song X, et al. Dysregulation of LncRNAs in placenta and pathogenesis of preeclampsia. Curr Drug Targets. 2017;18(10):1165–1170. doi: 10.2174/1389450118666170404160000. [DOI] [PubMed] [Google Scholar]
- 190.Ling Z, et al. Involvement of aberrantly activated HOTAIR/EZH2/miR-193a feedback loop in progression of prostate cancer. J Exp Clin Cancer Res. 2017;36(1):159. doi: 10.1186/s13046-017-0629-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Zhou J, Yang L, Zhong T, Mueller M, Men Y, Zhang N, Xie J, Giang K, Chung H, Sun X, Lu L, Carmichael GG, Taylor HS, Huang Y. H19 lncRNA alters DNA methylation genome wide by regulating S-adenosylhomocysteine hydrolase. Nat Commun. 2015;6(1):10221. doi: 10.1038/ncomms10221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Dey BK, Pfeifer K, Dutta A. The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014;28(5):491–501. doi: 10.1101/gad.234419.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Yu L, et al. The H19 gene imprinting in normal pregnancy and pre-eclampsia. Placenta. 2009;30(5):443–447. doi: 10.1016/j.placenta.2009.02.011. [DOI] [PubMed] [Google Scholar]
- 194.Mohammadpour-Gharehbagh A, et al. Impact of HOTAIR variants on preeclampsia susceptibility based on blood and placenta and in silico analysis. IUBMB Life. 2019;71(9):1367–1381. doi: 10.1002/iub.2095. [DOI] [PubMed] [Google Scholar]
- 195.Barrett SP, Salzman J. Circular RNAs: analysis, expression and potential functions. Development. 2016;143(11):1838–1847. doi: 10.1242/dev.128074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Han B, Chao J, Yao H. Circular RNA and its mechanisms in disease: from the bench to the clinic. Pharmacol Ther. 2018;187:31–44. doi: 10.1016/j.pharmthera.2018.01.010. [DOI] [PubMed] [Google Scholar]
- 197.Ou Y, et al. The expression profile of circRNA and its potential regulatory targets in the placentas of severe pre-eclampsia. Taiwan J Obstet Gynecol. 2019;58(6):769–777. doi: 10.1016/j.tjog.2019.09.007. [DOI] [PubMed] [Google Scholar]
- 198.Zhang YG, et al. Circular RNA in blood corpuscles combined with plasma protein factor for early prediction of pre-eclampsia. Bjog. 2016;123(13):2113–2118. doi: 10.1111/1471-0528.13897. [DOI] [PubMed] [Google Scholar]
- 199.Zhang Y, et al. CircSFXN1 regulates the behaviour of trophoblasts and likely mediates preeclampsia. Placenta. 2020;101:115–123. doi: 10.1016/j.placenta.2020.09.012. [DOI] [PubMed] [Google Scholar]
- 200.Deng N, et al. Circular RNA expression profiling identifies hsa_circ_0011460 as a novel molecule in severe preeclampsia. Pregnancy Hypertens. 2019;17:216–225. doi: 10.1016/j.preghy.2019.06.009. [DOI] [PubMed] [Google Scholar]
- 201.Bai Y, et al. Profiles of circular RNAs in human placenta and their potential roles related to preeclampsia. Biol Reprod. 2018;98(5):705–712. doi: 10.1093/biolre/ioy034. [DOI] [PubMed] [Google Scholar]
- 202.Qian Y, et al. Potential significance of circular RNA in human placental tissue for patients with preeclampsia. Cell Physiol Biochem. 2016;39(4):1380–1390. doi: 10.1159/000447842. [DOI] [PubMed] [Google Scholar]
- 203.Jiang M, et al. CircRNA-0004904, CircRNA-0001855, and PAPP-A: potential novel biomarkers for the prediction of preeclampsia. Cell Physiol Biochem. 2018;46(6):2576–2586. doi: 10.1159/000489685. [DOI] [PubMed] [Google Scholar]
- 204.Zhou W, et al. The profile analysis of circular RNAs in human placenta of preeclampsia. Exp Biol Med (Maywood) 2018;243(14):1109–1117. doi: 10.1177/1535370218813525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Hu X, et al. Competing endogenous RNA expression profiling in pre-eclampsia identifies hsa_circ_0036877 as a potential novel blood biomarker for early pre-eclampsia. Clin Epigenetics. 2018;10:48. doi: 10.1186/s13148-018-0482-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Hansen TB, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495(7441):384–388. doi: 10.1038/nature11993. [DOI] [PubMed] [Google Scholar]
- 207.Laganà AS, et al. Early serum markers of pre-eclampsia: are we stepping forward? J Matern Fetal Neonatal Med. 2016;29(18):3019–3023. doi: 10.3109/14767058.2015.1113522. [DOI] [PubMed] [Google Scholar]
- 208.Li H, et al. Unique microRNA signals in plasma exosomes from pregnancies complicated by preeclampsia. Hypertension. 2020;75(3):762–771. doi: 10.1161/HYPERTENSIONAHA.119.14081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Liu H, et al. Exosomal microRNA-139-5p from mesenchymal stem cells accelerates trophoblast cell invasion and migration by motivation of the ERK/MMP-2 pathway via downregulation of protein tyrosine phosphatase. J Obstet Gynaecol Res. 2020;46(12):2561–72. [DOI] [PMC free article] [PubMed]
- 210.Li W, et al. Circ_0063517 acts as ceRNA, targeting the miR-31-5p-ETBR axis to regulate angiogenesis of vascular endothelial cells in preeclampsia. Life Sci. 2020;244:117306. doi: 10.1016/j.lfs.2020.117306. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Datasets are available through the corresponding author upon reasonable request.