

Abstract
Background
Emerging evidence suggests that inflammation induced by the inflammasome plays a crucial role in the course of Parkinson’s disease (PD). Ulinastatin (UTI) has shown significant anti-inflammatory effects. However, few studies have examined whether UTI protects neurons through its anti-inflammatory effects in PD. The purpose of this study is to determine whether UTI exerts neuroprotection in a PD cell model and to explore the mechanisms.
Methods
SH-SY5Y cells and nerve growth factor (NGF)-treated PC12 cells were used to establish MPP+ induced PD cell models. Cells were pre-treated with UTI, then cell viabilities were detected using the MTT assay. Lactate dehydrogenase (LDH) release was detected using the LDH release assay kit. Inflammatory factors such as IL-1β, IL-6, and TNF-α were detected using ELISA. The expression levels of TH, NLRP3, caspase-1, ASC, IL-1β, and IL-18 were measured using western blotting, and DA release was detected using HPLC. A NLRP3 activator Nigericin was used to verify the effect of NLRP3 in the neuroprotective mechanism of UTI.
Results
We observed decreased cell viability, increased apoptosis, and increased inflammatory factors such as IL-1β, IL-6, and TNF-α in the MPP+ induced PD model. We also found decreased DA secretion and TH expression, as well as increased NLRP3, caspase-1, ASC, IL-1α, and IL-18 expression in the MPP+ induced PD model. These changes were significantly attenuated by UTI pre-treatment in a dose dependent manner. NLRP3 activator Nigericin markedly increased LDH release, accelerated apoptosis, increased inflammation levels and decreased DA secretion and TH expression, suggesting that Nigericin eliminated the neuroprotective effect of UTI on MPP+ treated cells.
Conclusions
Our data demonstrated that UTI pre-treatment performed a neuroprotective role in the MPP+ induced PD cell models by inhibiting the NLRP3 pathway.
Keywords: Ulinastatin (UTI), Parkinson’s disease (PD), NOD-leucine rich repeat and pyrin containing protein 3 (NLRP3), inflammation, neuroprotection
Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disease, after Alzheimer disease (1). In addition to classic motor symptoms, such as tremors, rigidity, and slow movement, some non-motor symptoms, such as emotional, cognitive, and olfactory disorders also appear in the early stages of PD. This is accompanied by the apoptosis of dopaminergic neurons, which eventually leads to paralysis and even death, seriously affecting people’s quality of life (2-7). However, current treatments for PD, including deep brain stimulation therapy and dopaminergic replacement therapy, have failed to halt or slow down the progression of PD (8). Long-term use of levodopa is associated with severe side effects, including the emergence of drug-induced involuntary movements and motor response fluctuation (9). Finding new potential drugs will provide more options for PD therapy.
As a urinary trypsin inhibitor, ulinastatin (UTI) is a glycoprotein isolated from healthy human urine or synthetically produced. Highly purified UTI has been used clinically for the treatment of acute pancreatitis, chronic pancreatitis, Stevens-Johnson syndrome, and toxic epidermal necrolysis (10,11). Clinical study found that Ulinastatin can reduce postoperative bleeding and cognitive dysfunction in patients undergoing cardiac surgery (12,13). Ulinastatin treatment could alleviate sepsis shock of CLP rats by suppressing the inflammatory responses of the body and strengthening the immune system (14). Additionally, ulinastatin ameliorated the severity of intestinal damage in necrotizing enterocolitis (NEC) by inhibiting apoptosis and suppressing the levels of inflammation cytokines (15). Studies have shown that Ulinastatin can also reduce the level of inflammatory cytokines and inhibit oxidative stress (16,17).
PD pathology is characterized by the loss of dopaminergic neurons in the brain and the accumulation of α-synuclein (18). Substantial evidence in autopsies, brain imaging, and biomarkers in bodily fluid have shown that neuroinflammation is a significant feature of PD (19). The activation of glial cells and the release of inflammatory factors have also been significantly observed in PD animal models induced by neurotoxins such as MPTP or 6-OHDA (20). Inflammasomes are polyprotein complexes in the cytosome, including NLRPI, NALP2, NLRC4, AIM2, etc., of which the most studied is the NOD-leucine rich repeat and pyrin containing protein 3 (NLRP3) (21). NLPR3 is a multi-protein complex in cells, mainly composed of receptor molecules (PYD), adaptive proteins (ASCs), and effect proteins (caspase-1) (22). Previous studies have shown that NLRP3 palys a vital role in neurodegenerative diseases (23,24). α-synuclein promoted the progression of PD by activating NLRP3 inflammasome in astrocytes (25). The NLRP3 inflammasome is activated and the NLRP3 mRNA expression was up-regulated in peripheral blood mononuclear cells (PBMCs) from PD patients (26). Downregulation of NLRP3 could inhibit inflammation and alleviate MPTP-induced degeneration of DA neurons in MPTP-induced PD mice (27). Hence, studying whether UTI can alleviate PD by regulating NLRP3 might provide theoretical reference for the treatment of PD.
NLRP3 activation is thought to require a two-step process: start-up and activation. The initial process increases the expression of various parts of the inflammasome. There are three models for the progression of NLRP3 inflammasome: reactive oxygen species (ROS) generation by pathogen-associated molecular patterns (PAMPs) and damage associated molecular pattern (DAMPs) (28-32); outflow of potassium ions (K+) by extracellular Adenosine Triphosphate (ATP) through purinergic 2X7 receptor (P2X7R) (33), and disruption of phagocytic vesicles including crystals by lysosomes (34). Thioredoxin-interacting protein (TXNIP) induces the activation of NLRP3 inflammasome in a ROS-sensitive manner as an endogenous antagonist of thioredoxin (35). NLRP3 agonists such as ATP and nigericin cause mitochondrial damage and decrease of NAD+, thereby promoting the activation of NLRP3 inflammasome (36). The second step is the NLRP3 inflammasome oligomerization, which mediates the proteolysis of pro-caspase-1 into active caspase-1, thereby activating pro-IL-1β and pro-IL-18 into IL-1β and IL-18 (37,38). Furthermore, LPS in the cytotype binds directly to mouse caspase-11 or human caspase-4/5, which is a non-classical pathway that activates the NLRP3 (39,40).
Given the important role of neuroinflammation in the PD process and the significant anti-inflammatory effects of UTI, we speculate that UTI plays a neuroprotective role by inhibiting NLPR3. To test this hypothesis, we firstly identified the effects of different concentrations of UTI on cell viability and lactate dehydrogenase (LDH) release using PC12 and SH-SY5Y cells. The protective effect of UTI on MPP+ treated cells was then assessed, and inflammatory factors were detected using ELISA. Finally, HPLC and western blot methods were used to detect dopamine (DA) secretion and tyrosine hydroxylase (TH) protein expression. Through the above experiments, we expected to identify the neuroprotective effects of UTI in vitro and explore the underlying mechanisms.
We present the following article in accordance with the MDAR checklist (available at http://dx.doi.org/10.21037/atm-21-1882).
Methods
The following materials were used: RPMI 1640 medium (Solarbio Life Sciences, Beijing, China), PC12 cell (Mlbio, ml052695, Shanghai, China), SH-SY5Y cells (Mlbio, ml052084, Shanghai, China), MPP+ (Sigma-Aldrich, MO, USA), microplate reader (Thermo Fisher Scientific, MA, USA), LDH assay kit (Beyotime, Shanghai, China), ELISA kits (R&D Systems, Minneapolis, USA), RIPA total protein lysate (Abcam, MA, USA), PVDF membranes (Thermo Fisher Scientific, USA), anti-TH (sc-25269, Santa Cruz Biotechnology, CA, USA), anti-NLRP3 (SAB1410191, Sigma-Aldrich, USA), anti-caspase-1 (PRS3459, Sigma-Aldrich, USA), anti-ASC (sc-514414, Santa Cruz Biotechnology, USA), anti-IL-1β (sc-12742, Santa Cruz Biotechnology, USA), anti-IL-18 (PA5-79481, Thermo Fisher Scientific, USA), and anti-GAPDH (G9545, Sigma-Aldrich, USA), HRP-conjugated secondary antibody (sc-2357, sc-2789, Santa Cruz Biotechnology, USA), chemiluminescent imaging system (Syngene, Cambridge, UK), HPLC (Agilent Technologies, CA, USA).
Cell cultures and treatments
The PC12 cells were cultured in RPMI 1640 medium with 5% fetal bovine serum (FBS). After incubation for 24 hours, nerve growth factor (NGF) at a concentration of 50 ng/mL was added to the medium to induce differentiation. After NGF addition for 4 days, subsequent experiments were performed. SH-SY5Y cells were cultured in DMEM including 1% streptomycin/penicillin as well as 10% FBS.
To detect whether UTI affects cell viability and LDH release, PC12 cells and SH-SY5Y cells were treated with different concentrations of UTI (0, 100, 200, 500, 1,000 µM) for 6 hours, then the cell viability and LDH release were detected. To produce the experimental PD cell model, PC12 and SH-SY5Y cells were exposed to 100 µM MPP+ for 24 hours. Different concentrations of UTI were used to process cells before the PD cell model was established.
Cell viability by the MTT cell assay
Cell viability was detected by the MTT cell assay. Cells were seeded into 96-well plates and incubated under standard culture conditions for 72 hours. After UTI or MPP+ treatment, the cells were incubated for 5 hours with MTT (5 mg/mL). The absorbance of individual wells was read at 570 nm using a microplate reader. Cell viability was determined by absorbance values.
LDH release assay
The LDH release assay was performed to assess the neuroprotective effects of UTI on PC12 or SH-SY5Y cells (41). Cells were seeded in 24-well plates at a concentration of 2×105 cells/mL. The amount of LDH was detected using the LDH assay kit according to the instructions of the manufacturer.
Enzyme-linked immunosorbent assay (ELISA)
The levels of interleukin-1 beta (IL-1β), IL-6, and TNF-α were measured in PC12 and SH-SY5Y cell culture medium using ELISA kits according to the manufacturer’s instructions (42). Optical density values were measured at 450 mm with a microplate reader in each well.
Western blot assay
Cellular proteins were extracted using RIPA total protein lysate, then separated by electrophoresis by SDS-PAGE and transferred onto PVDF. After incubation with blocking solution, the membranes were separately incubated with anti-TH, anti-NLRP3, anti-caspase-1, anti-ASC, anti-IL-1β, anti-IL-18, and anti-GAPDH at room temperature for 3 hours. Then, the HRP-conjugated secondary antibody was incubated with the blot for 1.5 hours. Image acquisition was performed by a chemiluminescent imaging system.
High performance liquid chromatography (HPLC)
The level of DA was measured by HPLC. After UTI or MPP+ treatment, extracellular buffer was collected and stored at −20 °C until analysis. On the day of analysis, the samples were centrifuged at 10,000 g for 10 min, and 3 µL of supernatant was injected into the HPLC to measure DA. To quantify the sample peaks, each chemical species (DA) was compared to the external standards.
Data processing and analysis
Statistical analysis was performed using GraphPad Prism software (version 7.0). Differences between groups were analyzed using one-way analysis of variance (one-way ANOVA) followed by a Bonferroni post-hoc test. Data were considered significantly different when P<0.05. All experiments in this study were performed in triplicate.
Results
Low concentration of UTI has no significant effect on cell viability
To verify whether different concentrations of UTI affected cell state, we first treated PC12 and SH-SY5Y cells with UTI at the concentrations of 100, 200, 500, and 1,000 µM. After 6 hours of UTI treatment, cell viability was detected via the MTT cell assay, and LDH release was also detected. As shown in Figure 1, UTI treatment at concentrations of 100, 200, and 500 µM did not significantly alter cell viability and LDH release compared to the control group (one-way ANOVA). Additionally, with 1,000 µM UTI treatment, the cell viability of PC12 cells and SH-SY5Y cells decreased significantly (P<0.01) and LDH release increased significantly (P<0.01). These results indicated that low concentrations (100, 200, 500 µM) of UTI had no significant effect on cell viability and did not affect LDH release levels. Since the high concentration of UTI affected the viability and LDH level of PC12 cells and SH-SY5Y cells, 100, 200, and 500 µM UTI treatments were used in subsequent experiments.
Figure 1.
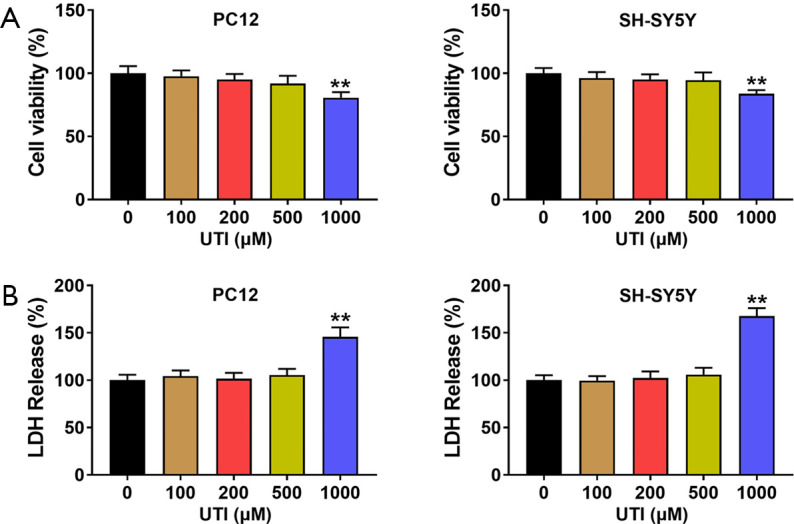
The effect of UTI on PC12 and SH-SY5Y cells. (A) The effect of different concentrations of UTI on cell viability. Concentrations of 0–500 µM UTI had no significant effect on cell viability, while 1,000 µM UTI significantly reduced the cell viability. (B) The effect of different concentrations of UTI on LDH release. Concentrations of 0–500 µM UTI had no significant effect on LDH release, while 1,000 µM UTI significantly increased the LDH of PC12 and SH-SY5Y cells. Since a high concentration of UTI affected the viability and LDH level of PC12 cells and SH-SY5Y cells, 100, 200, and 500 µM UTI treatments were used in subsequent experiments. **, P<0.01. UTI, ulinastatin; LDH, lactate dehydrogenase.
UTI increases the viability of MPP+ treated cells and inhibits apoptosis
In this study, PC12 and SH-SY5Y cells treated with MPP+ at a concentration of 100 µM were used as PD cell models. In order to assess the protective effect on cells, different concentrations of UTI were used to treat cells before modeling. As shown in Figure 2A, the MTT assay demonstrated that MPP+ significantly reduced cell viability, while different concentrations of UTI pre-treatment significantly enhanced the viability of MPP+ treated PC12 and SH-SY5Y cells (one-way ANOVA, P<0.01). Using the higher UTI concentration, the viability of the MPP+ treated cells was closer to that of the control group. Correspondingly, as shown in Figure 2B, UTI pre-treatment also significantly reversed the increase in LDH release caused by MPP+ (one-way ANOVA, P<0.01). Flow cytometric analysis showed that MPP+ induced cell apoptosis, while UTI pre-treatment significantly reduced apoptosis, as shown in Figure 2C,D. It is worth noting that the protective effect of UTI was dose-dependent (one-way ANOVA, P<0.01). The dose dependence of the protective effect of UTI was also reflected in subsequent experiments.
Figure 2.

UTI enhances cell viability after MPP+ treatment and inhibits apoptosis. (A) MPP+ reduced PC12 and SH-SY5Y cell viabilities, while UTI pre-treatment significantly enhanced PC12 and SH-SY5Y cell viabilities; (B) MPP+ increased LDH release in PC12 and SH-SY5Y cells, and UTI pre-treatment inhibited the release of LDH; (C,D) UTI pre-treatment significantly inhibited apoptosis of PC12 and SH-SY5Y cells induced by MPP+. The protective effect of UTI increased with the increase in concentration. **, P<0.01 vs. Normal group; ##, P<0.01 vs. MMP+ group. UTI, ulinastatin; LDH, lactate dehydrogenase.
UTI inhibits the inflammatory response induced by MPP+
MPP+ induces apoptosis by activating inflammatory responses that stimulate cells to produce inflammatory factors. We used ELISA to detect inflammatory factors such as IL-1β, IL-6, and TNF-α in PC12 cells and SH-SY5Y cells. For PC12 cells, compared to the control group, the levels of inflammatory factors increased significantly in the MPP+ group, and UTI pre-treatment significantly reduced the levels of inflammatory factors (Figure 3, one-way ANOVA, P<0.01). For SH-SY5Y cells, UTI pre-treatment also showed similar anti-inflammatory effects (Figure 3, one-way ANOVA, P<0.01).
Figure 3.
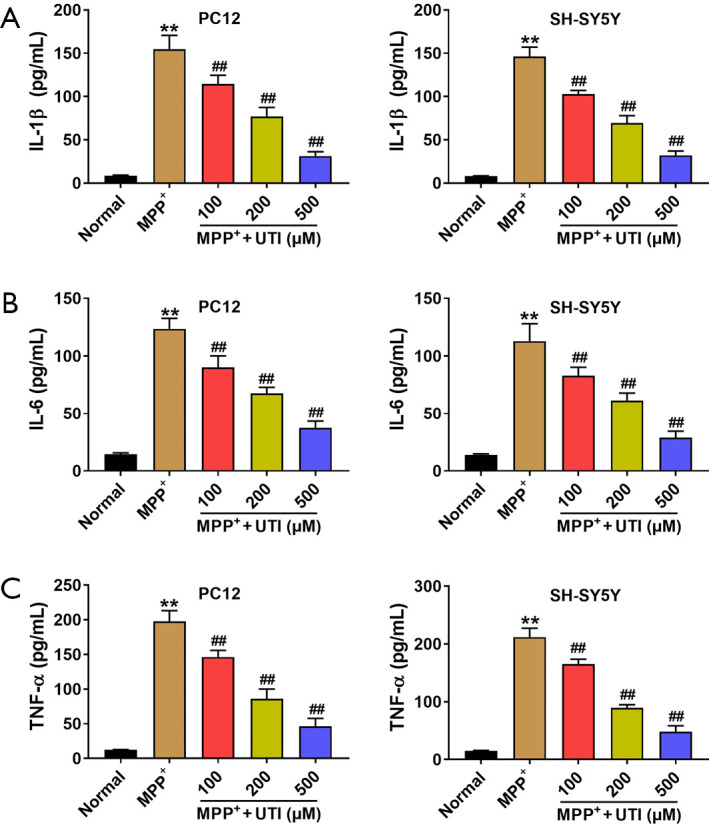
UTI inhibits the inflammatory response induced by MPP+. (A) MPP+ significantly increased IL-1β levels of PC12 and SH-SY5Y cells, while UTI pre-treatment significantly reduced IL-1β levels in a concentration dependent manner; (B) MPP+ increased the level of IL-6 in PC12 and SH-SY5Y cells, and UTI pre-treatment significantly reduced IL-6 levels in a concentration dependent manner; (C) UTI pre-treatment reduced TNF-α levels that were increased by MPP+ in PC12 and SH-SY5Y cells in a concentration dependent manner. **, P<0.01 vs. Normal group; ##, P<0.01 vs. MPP+ group. UTI, ulinastatin.
UTI enhances DA release and TH expression in MPP+ treated cells
In the above experiments, we demonstrated that UTI inhibited the inflammatory response induced by MPP+, thereby enhancing cell viability, inhibiting cell death, and playing a neuroprotective role. Another question was whether UTI protected the function of dopaminergic neurons. To explore this question, DA secretion levels were detected by HPLC and TH expression levels were detected via western blotting in PC12 and SH-SY5Y cells. As demonstrated in Figure 4A, in PC12 cells, MPP+ significantly reduced DA secretion, and UTI pre-treatment attenuated the inhibition of MPP+ (one-way ANOVA, P<0.01). The western blotting image of TH is on the left side of Figure 4B. In PC12 cells, MPP+ significantly reduced TH expression and UTI pre-treatment also reversed the inhibition of MPP+ (one-way ANOVA, P<0.05). The same trend was found in SH-SY5Y cells, as shown in Figure 4C,D (DA secretion test: one-way ANOVA, P<0.01, TH expression test: one-way ANOVA, P<0.05).
Figure 4.

UTI increases the DA and TH levels of the PC12 and SH-SY5Y cells treated with MPP+. (A,C) MPP+ treatment significantly reduced the DA levels of PC12 cells and SH-SY5Y cells, while UTI pre-treatment attenuated DA levels in a concentration dependent manner; (B,D) MPP+ treatment significantly inhibited TH expression in PC12 cells and SH-SY5Y cells, and UTI pre-treatment increased TH expression in a concentration dependent manner. **, P<0.01 vs. Normal group; #, P<0.05 vs. MPP+ group; ##, P<0.01 vs. MPP+ group. UTI, ulinastatin.
UTI inhibits the NLRP3 pathway
To further explore the mechanism by which UTI inhibited MPP+ induced inflammatory responses, we detected the protein levels of NLRP3 and related proteins by western blotting. As shown in Figure 5A, in PC12 cells, MPP+ treatment significantly increased NLRP3, caspase-1, ASC, IL-1α, and IL-18 expression, and UTI pre-treatment reduced the over-expression of these proteins (one-way ANOVA, P<0.01). Similarly, in SH-SY5Y cells, MPP+ treatment significantly increased the expression levels of NLRP3, caspase-1, ASC, IL-1α, and IL-18, and UTI pre-treatment reduced the over-expression of these proteins (Figure 5B; one-way ANOVA, P<0.01). These results indicate that the anti-inflammatory effects exerted by UTI were achieved through inhibiting the NLRP3 pathway.
Figure 5.

UTI inhibits the NLRP3 pathway. MPP+ activated the NLRP3 pathway in PC12 cells (A) and SH-SY5Y cells (B), promoting NLRP3, caspase-1, ASC, IL-1β, and IL-18 expression, while UTI pre-treatment significantly attenuated changes in the NRPL3 pathway. **, P<0.01 vs. Normal group; ##, P<0.01 vs. MPP+ group. UTI, ulinastatin; NLRP3, NOD-leucine rich repeat and pyrin containing protein 3.
Activation of NLRP3 eliminates the protective effect of UTI on MPP+ treated cells
To confirm that UTI protected MPP+ treated cells by inhibiting the activation of NLRP3, we detected the changes of cells after treated with NLRP3 activator Nigericin (5 µM). As shown in Figure 6A, UTI pre-treatment (500 µM) markedly decreased NLRP3, caspase-1, ASC, IL-1α, and IL-18 expression induced by MPP+ treatment in PC12 and SH-SY5Y cells (one-way ANOVA, P<0.01). Compared with MPP+ + UTI group, Nigericin treatment significantly up-regulated NLRP3, caspase-1, ASC, IL-1α, and IL-18 expression (one-way ANOVA, P<0.01). The results in Figure 6B showed that Nigericin treatment significantly increased LDH release and eliminated the suppressed effect of UTI pre-treatment on LDH release (one-way ANOVA, P<0.01). Flow cytometric analysis showed that Nigericin treatment accelerated apoptosis and eliminated the inhibited effect of UTI on apoptosis of MPP+ treated PC12 and SH-SY5Y cells (Figure 6C and D, one-way ANOVA, P<0.01). Additionally, we detected inflammatory factors in PC12 cells and SH-SY5Y cells after Nigericin treatment. Compared to the MPP++UTI group, the levels of IL-1β, IL-6, and TNF-αsignificantly increased In the MPP+ + UTI + Nigericin group (Figure 6E, one-way ANOVA, P<0.01). As shown in Figure 6F and G, Nigericin treatment significantly decreased the levels of DA secretion and TH expression in MPP++UTI group, indicating that Nigericin eliminated the promoting effect of UTI on DA secretion and TH expression of MPP+ treated PC12 and SH-SY5Y cells (one-way ANOVA, P<0.01).
Figure 6.

Activation of NLRP3 eliminates the protective effect of UTI on MPP+ treated cells. (A) The NLRP3 activator Migericin significantly increased the protein expression of NLRP3, caspase-1, ASC, IL-1β, and IL-18, and eliminated the inhibited effect of UTI on NLRP3 pathway in MPP+ treated PC12 cells and SH-SY5Y cells. (B) Nigericin increased LDH release and eliminated the inhibited effect of UTI on LDH release in PC12 and SH-SY5Y cells. (C,D) Nigericin accelerated cell apoptosis eliminated the inhibited effect of UTI on apoptosis of PC12 and SH-SY5Y cells. (E) Nigericin significantly increased the levels of IL-1β, IL-6, and TNF-α, and eliminated the inhibited effect of UTI on inflammation of PC12 and SH-SY5Y cells. (F) Nigericin significantly increased DA levels and eliminated the promoting effect of UTI on DA of PC12 and SH-SY5Y cells. (G) Nigericin significantly increased TH protein expression and eliminated the promoting effect of UTI on TH protein of PC12 and SH-SY5Y cells. **, P<0.01 vs. Normal group; ##, P<0.01 vs. MPP+ group, &&, P<0.01 vs. MPP+ + UTI group. UTI, ulinastatin; LDH, lactate dehydrogenase; NLRP3, NOD-leucine rich repeat and pyrin containing protein 3.
Discussion
In this study, we first treated cells with different concentrations of UTI to verify that low concentrations of UTI had no significant effect on cell viability. Subsequently, in the MPP+ induced PD cell model, different concentrations of UTI pre-treatment were shown to enhance cell viability and inhibit apoptosis. Furthermore, we used ELISA to measure the levels of IL-1β, IL-6, and TNF-α, and the results showed that UTI pre-treatment significantly reduced the levels of inflammatory factors. On the other hand, to detect whether UTI could protect the function of neurons in the PD model, we used HPLC to detect DA secretion and western blot to detect cell TH expression, and found that UTI pre-treatment improved DA secretion and TH expression in PD model cells. To further explore the mechanism by which UTI inhibited MPP+ induced inflammatory responses, we tested NLRP3 and related protein expression with western blotting. UTI was found to significantly reduce the expression of NLRP3, caspase-1, ASC, IL-1α, and IL-18 in MPP+ treated cells. To confirm the role of NLRP3 in the protective mechanism of UTI on MPP+ treated cells, the NLRP3 activator Nigericin was used to treat cells. The results showed that Nigericin treatment significantly increased the expression of NLRP3 related proteins, accelerated apoptosis, increased levels of inflammatory factors, and decreased DA secretion and TH expression. These results showed that UTI played a neuroprotective role in PD model cells by inhibiting the NLRP3 pathway.
A previous study found that inflammatory cytokine IL-1β levels increased in the serum of patients with PD (28). Another group found that elevated levels of IL-1β in serum were mainly associated with the activation of NLRP3 in the brain and small glial cells in PD patients (43). The activation of NLRP3 promotes the maturation and secretion of IL-1β and increases neuroinflammation, thus promoting the development of PD (31,44-46). The main pathological changes in PD are the degeneration and disappearance of DA neurons in the substantia nigra, and neuroinflammation mediated by NLRP3 may lead to dopaminergic neuron degeneration in PD patients, aggravating the process of PD (47). A histological study from 2018 found an increase in NLRP3 expression in dopaminergic neurons in the brains of PD patients (48). These studies suggest that NLRP3 is a reasonable candidate target for PD.
As a protease inhibitor isolated from the urine of healthy adult males, UTI is safe to use and does not easily induce allergies. UTI reduces the inflammatory response through several processes such as anti-proteolysis, inhibition of inflammatory mediator release, and antioxidant reactions (49,50). By activating the heme oxygenase 1 antioxidant pathway, UTI attenuates allergic inflammation (33). By reducing the translocation of lipopolysaccharide and systemic inflammation, UTI improves the cognitive function of elderly patients with spinal surgery (51). Besides, UTI could have a neuroprotective effect in the rat model of neuropathic pain (52,53). Ulinastatin may attenuate the development of neuropathic pain when it was administered before the development of neuropathic pain (54). In our study, UTI pre-treatment inhibited the activation of NLRP3 and suppressed the expression of NLRP3 related proteins and the levels of inflammatory factors. We found that UTI significantly inhibited LDH release and cell apoptosis that induced by MPP+. Additionally, UTI increased DA secretion and TH expression of MPP+ treated PC12 and SH-SY5Y cells. However, after treated with Nigericin, a NLRP3 activator, the inhibited effect of UTI on the levels of inflammatory factors, LDH release, and cell apoptosis of MPP+ treated cells were mostly eliminated. These results indicated that UTI performed a neuroprotective effect on MPP+ induced PD cell models by inhibiting the activation of NLRP3.
One of the limitations of this study is the PD cell models used in this study. It should be noted that at the tissue or overall level, neurons are in a more complex environment, and the neuroprotective effect of UTI needs to be further verified in PD animal models. Secondly, it is well known that the development and processes of PD are associated with the release of a variety of inflammatory factors. In this study, we focused only on the NLRP3 pathway. Therefore, suppressing these cytokines by inhibition of the NLRP3 pathway may be only one part of UTI’s neuroprotective mechanism, and further research is needed to fully elucidate these mechanisms.
In conclusion, UTI significantly decreased LDH release, cell apoptosis and levels of inflammatory factors, improved cell viability and DA secretion. Its neuroprotective effect was achieved by inhibiting the NLRP3 pathway in the MPP+ induced PD cell model.
Supplementary
The article’s supplementary files as
Acknowledgments
Funding: None.
Ethical Statement: The authors are accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Reporting Checklist: The authors have completed the MDAR checklist. Available at http://dx.doi.org/10.21037/atm-21-1882
Data Sharing Statement: Available at http://dx.doi.org/10.21037/atm-21-1882
Conflicts of Interest: All authors have completed the ICMJE uniform disclosure form (available at http://dx.doi.org/10.21037/atm-21-1882). The authors have no conflicts of interest to declare.
(English Language Editor: C. Betlazar-Maseh)
References
- 1.Guo M. Molecular pathways to Parkinson’s disease. Mol Neurodegener 2012;7 Suppl 1:L13. 10.1186/1750-1326-7-S1-L13 [DOI] [Google Scholar]
- 2.Shah RS, Chang SY, Min HK, et al. Deep Brain Stimulation: Technology at the Cutting Edge. J Clin Neurol 2010;6:167-82. 10.3988/jcn.2010.6.4.167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dai HD, Otten B, Mehrkens JH, et al. A Novel Glove Monitoring System Used to Quantify Neurological Symptoms During Deep-Brain Stimulation Surgery. IEEE Sensors Journal 2013;13:3193-202. 10.1109/JSEN.2013.2271775 [DOI] [Google Scholar]
- 4.Rissanen SM, Kankaanpaa M, Tarvainen MP, et al. Analysis of EMG and acceleration signals for quantifying the effects of deep brain stimulation in Parkinson's disease. IEEE Trans Biomed Eng 2011;58:2545-53. 10.1109/TBME.2011.2159380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Stacy M. Iconography: Medical Treatment of Parkinson Disease. [DOI] [PubMed]
- 6.Giuffrida JP, Riley DE, Maddux BN, et al. Clinically deployable Kinesia technology for automated tremor assessment. Mov Disord 2009;24:723-30. 10.1002/mds.22445 [DOI] [PubMed] [Google Scholar]
- 7.Jahanshahi M. Effects of deep brain stimulation of the subthalamic nucleus on inhibitory and executive control over prepotent responses in Parkinson's disease. Front Syst Neurosci 2013;7:118. 10.3389/fnsys.2013.00118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhou H, Shao M, Yang X, et al. Tetramethylpyrazine Analogue T-006 Exerts Neuroprotective Effects against 6-Hydroxydopamine-Induced Parkinson's Disease In Vitro and In Vivo. Oxid Med Cell Longev 2019;2019:8169125. 10.1155/2019/8169125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hwang TY, Song MA, Ahn S, et al. Effects of Combined Treatment with Acupuncture and Chunggan Formula in a Mouse Model of Parkinson's Disease. Evid Based Complement Alternat Med 2019;2019:3612587. 10.1155/2019/3612587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xu Q, Yan Q, Chen S. Ulinastatin is effective in reducing mortality for critically ill patients with sepsis: a causal mediation analysis. Sci Rep 2018;8:14360. 10.1038/s41598-018-32533-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Del Pozzo-Magana BR, Lazo-Langner A, Carleton B, et al. A systematic review of treatment of drug-induced Stevens-Johnson syndrome and toxic epidermal necrolysis in children. J Popul Ther Clin Pharmacol 2011;18:e121. [PubMed] [Google Scholar]
- 12.Yao YT, Fang NX, Liu DH, et al. Ulinastatin reduces postoperative bleeding and red blood cell transfusion in patients undergoing cardiac surgery: A PRISMA-compliant systematic review and meta-analysis. Medicine 2020;99:e19184. 10.1097/MD.0000000000019184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhou M, Lyu Y, Zhu Y, et al. Effect of Ulinastatin Combined With Dexmedetomidine on Postoperative Cognitive Dysfunction in Patients Who Underwent Cardiac Surgery. Front Neurol 2019;10:1293. 10.3389/fneur.2019.01293 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang J, Zhou J, Bai S. Combination of Glutamine and Ulinastatin Treatments Greatly Improves Sepsis Outcomes. J Inflamm Res 2020;13:109-15. 10.2147/JIR.S234122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liang S, Lai P, Li X, et al. Ulinastatin Reduces the Severity of Intestinal Damage in the Neonatal Rat Model of Necrotizing Enterocolitis. Med Sci Monit 2019;25:9123-30. 10.12659/MSM.919413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Li C, Ma D, Chen M, et al. Ulinastatin attenuates LPS-induced human endothelial cells oxidative damage through suppressing JNK/c-Jun signaling pathway. Biochem Biophys Res Commun 2016;474:572-8. 10.1016/j.bbrc.2016.04.104 [DOI] [PubMed] [Google Scholar]
- 17.Wei F, Liu S, Luo L, et al. Anti-inflammatory mechanism of ulinastatin: Inhibiting the hyperpermeability of vascular endothelial cells induced by TNF-α via the RhoA/ROCK signal pathway. Int Immunopharmacol 2017;46:220-7. 10.1016/j.intimp.2017.03.007 [DOI] [PubMed] [Google Scholar]
- 18.Lotharius J, Brundin P. Pathogenesis of Parkinson's disease: dopamine, vesicles and alpha-synuclein. Nat Rev Neurosci 2002;3:932-42. 10.1038/nrn983 [DOI] [PubMed] [Google Scholar]
- 19.Wang Z, Luo XG, Gao C. Utility of susceptibility-weighted imaging in Parkinson's disease and atypical Parkinsonian disorders. Transl Neurodegener 2016;5:17. 10.1186/s40035-016-0064-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wang Q, Liu Y, Zhou J. Neuroinflammation in Parkinson's disease and its potential as therapeutic target. Transl Neurodegener 2015;4:19. 10.1186/s40035-015-0042-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.He Y, Hara H, Nunez G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem Sci 2016;41:1012-21. 10.1016/j.tibs.2016.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Davis BK, Wen H, Ting PY. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu Rev Immunol 2011;29:707-35. 10.1146/annurev-immunol-031210-101405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Voet S, Srinivasan S, Lamkanfi M, et al. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO Mol Med 2019;11:e10248. 10.15252/emmm.201810248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Duan Y, Kelley N, He Y. Role of the NLRP3 inflammasome in neurodegenerative diseases and therapeutic implications. Neural Regen Res 2020;15:1249-50. 10.4103/1673-5374.272576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang X, Chi J, Huang D, et al. α-synuclein promotes progression of Parkinson's disease by upregulating autophagy signaling pathway to activate NLRP3 inflammasome. Exp Ther Med 2020;19:931-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fan Z, Pan YT, Zhang ZY, et al. Systemic activation of NLRP3 inflammasome and plasma α-synuclein levels are correlated with motor severity and progression in Parkinson's disease. J Neuroinflammation 2020;17:11. 10.1186/s12974-019-1670-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang LY, Yu X, Li XX, et al. Catalpol Exerts a Neuroprotective Effect in the MPTP Mouse Model of Parkinson’s Disease. Front Aging Neurosci 2019;11:316. 10.3389/fnagi.2019.00316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Qiao C, Zhang Q, Jiang QL, et al. Inhibition of the hepatic Nlrp3 protects dopaminergic neurons via attenuating systemic inflammation in a MPTP/p mouse model of Parkinson's disease. J Neuroinflammation 2018;15:193. 10.1186/s12974-018-1236-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhu J, Hu Z, Han X, et al. Dopamine D2 receptor restricts astrocytic NLRP3 inflammasome activation via enhancing the interaction of β-arrestin2 and NLRP3. Cell Death Differ 2018;25:2037-49. 10.1038/s41418-018-0127-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chhabra A, Ring AM, Weiskopf K, et al. Hematopoietic stem cell transplantation in immunocompetent hosts without radiation or chemotherapy. Sci Transl Med 2016;8:351ra105. 10.1126/scitranslmed.aae0501 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bauernfeind FG, Horvath G, Stutz A, et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol 2009;183:787-91. 10.4049/jimmunol.0901363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Liu W, Yin Y, Zhou Z, et al. OxLDL-induced IL-1 beta secretion promoting foam cells formation was mainly via CD36 mediated ROS production leading to NLRP3 inflammasome activation. Inflamm Res 2014;63:33-43. 10.1007/s00011-013-0667-3 [DOI] [PubMed] [Google Scholar]
- 33.Hussen J, Düvel A, Koy M, et al. Inflammasome activation in bovine monocytes by extracellular ATP does not require the purinergic receptor P2X7. Dev Comp Immunol 2012;38:312-20. 10.1016/j.dci.2012.06.004 [DOI] [PubMed] [Google Scholar]
- 34.van Bruggen R, Köker MY, Jansen M, et al. Human NLRP3 inflammasome activation is Nox1-4 independent. Blood 2010;115:5398-400. 10.1182/blood-2009-10-250803 [DOI] [PubMed] [Google Scholar]
- 35.Zhou R, Tardivel A, Thorens B, et al. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat Immunol 2010;11:136-40. 10.1038/ni.1831 [DOI] [PubMed] [Google Scholar]
- 36.Misawa T, Takahama M, Kozaki T, et al. Microtubule-driven spatial arrangement of mitochondria promotes activation of the NLRP3 inflammasome. Nat Immunol 2013;14:454-60. 10.1038/ni.2550 [DOI] [PubMed] [Google Scholar]
- 37.Lamkanfi M, Dixit VM. Mechanisms and functions of inflammasomes. Cell 2014;157:1013-22. 10.1016/j.cell.2014.04.007 [DOI] [PubMed] [Google Scholar]
- 38.Vanaja SK, Rathinam VAK, Fitzgerald KA. Mechanisms of inflammasome activation: recent advances and novel insights. Trends Cell Biol 2015;25:308-15. 10.1016/j.tcb.2014.12.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.de Carvalho RVH, Andrade WA, Lima-Junior DS, et al. Leishmania Lipophosphoglycan Triggers Caspase-11 and the Non-canonical Activation of the NLRP3 Inflammasome. Cell Rep 2019;26:429-37.e5. 10.1016/j.celrep.2018.12.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Baker PJ, Boucher D, Bierschenk D, et al. NLRP3 inflammasome activation downstream of cytoplasmic LPS recognition by both caspase-4 and caspase-5. Eur J Immunol 2015;45:2918-26. 10.1002/eji.201545655 [DOI] [PubMed] [Google Scholar]
- 41.Wu L, Jiang C, Kang Y, et al. Curcumin exerts protective effects against hypoxia-reoxygenation injury via the enhancement of apurinic/apyrimidinic endonuclease 1 in SH-SY5Y cells: Involvement of the PI3K/AKT pathway. Int J Mol Med 2020;45:993-1004. 10.3892/ijmm.2020.4483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xu J, Sun Y, Lu J. Knockdown of Long Noncoding RNA (lncRNA) AK094457 Relieved Angiotensin II Induced Vascular Endothelial Cell Injury. Med Sci Monit 2020;26:e919854. 10.12659/MSM.919854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dinarello CA, Simon A, van der Meer JW. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat Rev Drug Discov 2012;11:633-52. 10.1038/nrd3800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Próchnicki T, Mangan MS, Latz E. Recent insights into the molecular mechanisms of the NLRP3 inflammasome activation. F1000Res 2016;5:1469. 10.12688/f1000research.8614.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Burre J, Sharma M, Sudhof TC. Definition of a Molecular Pathway Mediating α-Synuclein Neurotoxicity. J Neurosci 2015;35:5221-32. 10.1523/JNEUROSCI.4650-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Codolo G, Plotegher N, Pozzobon T, et al. Triggering of Inflammasome by Aggregated α–Synuclein, an Inflammatory Response in Synucleinopathies. PLoS One 2013;8:e55375. 10.1371/journal.pone.0055375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liu Y, Fiskum G, Schubert D. Generation of reactive oxygen species by the mitochondrial electron transport chain. J Neurochem 2002;80:780-7. 10.1046/j.0022-3042.2002.00744.x [DOI] [PubMed] [Google Scholar]
- 48.Verma D, Lerm M, Blomgran Julinder R, et al. Gene polymorphisms in the NALP3 inflammasome are associated with interleukin-1 production and severe inflammation: relation to common inflammatory diseases? Arthritis Rheum 2008;58:888-94. 10.1002/art.23286 [DOI] [PubMed] [Google Scholar]
- 49.Lv ZT, Huang JM, Zhang JM, et al. Effect of Ulinastatin in the Treatment of Postperative Cognitive Dysfunction: Review of Current Literature. Biomed Res Int 2016;2016:2571080. 10.1155/2016/2571080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Song D, Song G, Niu Y, et al. Ulinastatin Activates Heme Oxygenase-1 Antioxidant Pathway and Attenuates Allergic Inflammation. Br J Pharmacol 2014;171:4399-412. 10.1111/bph.12780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhang M, Zhang YH, Fu HQ, et al. Ulinastatin May Significantly Improve Postoperative Cognitive Function of Elderly Patients Undergoing Spinal Surgery by Reducing the Translocation of Lipopolysaccharide and Systemic Inflammation. Front Pharmacol 2018;9:1007. 10.3389/fphar.2018.01007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jung KT, Lee HY, Yoon MH, et al. The effect of urinary trypsin inhibitor against neuropathic pain in rat models. Korean J Pain 2013;26:356-60. 10.3344/kjp.2013.26.4.356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Ouyang H, Nie B, Wang P, et al. Ulinastatin attenuates neuropathic pain induced by L5-VRT via the calcineurin/IL-10 pathway. Mol Pain 2016;12:1744806916646785. 10.1177/1744806916646785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Oh SH, So HJ, Lee HY, et al. Urinary trypsin inhibitor attenuates the development of neuropathic pain following spinal nerve ligation. Neurosci Lett 2015;590:150-5. 10.1016/j.neulet.2015.01.070 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The article’s supplementary files as