

Abstract
Coral disease is a growing problem for coral reefs globally and diseases have been linked to thermal stress, excess nutrients, overfishing and other human impacts. The Red Sea is a unique environment for corals with a strong environmental gradient characterized by temperature extremes and high salinities, but minimal terrestrial runoff or riverine input and their associated pollution. Yet, relatively little is known about coral diseases in this region. Disease surveys were conducted at 22 reefs within three regions (Yanbu, Thuwal, Al Lith) in the central Red Sea along the Saudi Arabian coast. Surveys occurred in October 2015, which coincided with a hyperthermal-induced bleaching event. Our objectives were to 1) document types, prevalence, and distribution of coral diseases in a region with minimal terrestrial input, 2) compare regional differences in diseases and bleaching along a latitudinal gradient of environmental conditions, and 3) use histopathology to characterize disease lesions at the cellular level. Coral reefs of the central Red Sea had a widespread but a surprisingly low prevalence of disease (<0.5%), based on the examination of >75,750 colonies. Twenty diseases were recorded affecting 16 coral taxa and included black band disease, white syndromes, endolithic hypermycosis, skeletal eroding band, growth anomalies and focal bleached patches. The three most common diseases were Acropora white syndrome (59.1% of the survey sites), Porites growth anomalies (40.9%), and Porites white syndrome (31.8%). Sixteen out of 30 coral genera within transects had lesions and Acropora, Millepora and Lobophyllia were the most commonly affected. Cell-associated microbial aggregates were found in four coral genera including a first report in Stylophora. Differences in disease prevalence, coral cover, amount of heat stress as measured by degree heating weeks (DHW) and extent of bleaching was evident among sites. Disease prevalence was not explained by coral cover or DHW, and a negative relationship between coral bleaching and disease prevalence was found. The northern-most sites off the coast of Yanbu had the highest average disease prevalence and highest average DHW values but no bleaching. Our study provides a foundation and baseline data for coral disease prevalence in the central Red Sea, which is projected to increase as a consequence of increased frequency and severity of ocean warming.
Introduction
Coral disease is a significant factor impacting coral reefs with localized disease outbreaks occurring worldwide [1,2]. The single most damaging disease outbreak, stony coral tissue loss disease, has been devastating coral reefs throughout the Florida Reef Tract since 2014 [3–5] and has spread to neighboring Caribbean regions [6]. Outbreaks of coral disease have increased through time [7] and have been linked to anthropogenic impacts such as overfishing [8], plastic pollution [9], dredging activities [10], terrestrial runoff [11,12], and increased ocean temperatures [13,14].
Coral reefs face different threats depending on their geographic region. The Red Sea is a unique environment for corals being a partially enclosed body of water with limited exchange with the Indian Ocean, low influxes of freshwater (~30mm/year) and high evaporation rates [15,16]. The majority of coral reefs around the globe live with temperatures usually not exceeding 29°C and salinities around 36 PSU [17]. In the Red Sea, temperature extremes are the norm with temperatures surpassing 32°C in the summer, around 18°C in the winter and with salinities 40 PSU or higher [18,19]. Yet, the Red Sea has extensive and healthy coral reefs with approximately 346 species of reef corals [20,21]. The Red Sea is characterized by natural north to south gradients of temperature, salinity and nutrient availability [22,23]. As example, in the far north, sea surface temperatures (SSTs) average 26°C (± 1°C) compared to 31.3°C (± 1.1°C) in the south [22]. Numerous bleaching events have occurred on coral reefs in the Red Sea which also show a latitudinal gradient in coral response. During the recent bleaching in 2015, Osman et al. [24] reported that degree heating weeks (DHW) surpassing the bleaching threshold of four (https://coralreefwatch.noaa.gov/product/5km/index_5km_dhw.php) occurred throughout the Red Sea, yet bleaching only occurred in the central and southern parts of the Red Sea and was more severe in the southern end.
Although coral reefs in the Red Sea face temperature and salinity extremes typically not experienced by corals in other ocean basins, they also receive minimal terrestrial runoff or riverine input and their associated sedimentation, turbidity, and nutrient enrichment. Terrestrial runoff degrades local coral reefs [25] and contributes to increased severity and prevalence of coral diseases [10,26–28]. This creates a unique opportunity to examine coral health in an ecosystem with naturally high temperatures and salinities but minimal terrestrial pollution. Only a handful of comprehensive (all diseases recorded) coral disease surveys have been conducted in the Red Sea [29–31] with no known examination of Red Sea coral diseases at the cellular level. To address this paucity of information on coral health, we conducted coral disease surveys in three regions along the Saudi Arabian coast of the Red Sea in October 2015 which coincided with a bleaching event. Our objectives were to 1) document types, prevalence, and distribution of coral diseases in a region with minimal terrestrial input, 2) compare regional differences in diseases along a latitudinal gradient of environmental conditions and a gradient of bleaching response, 3) use histopathology to characterize disease lesions at the cellular level.
Materials and methods
Regional descriptions
We examined three areas that represent the northern (Yanbu), central (Thuwal), and southern (Al Lith) regions within the central Red Sea (Fig 1). The regions differ in environmental factors such as maximum winter and summer temperatures (higher towards the south), and photosynthetic active radiation (PAR, lower toward the south) [32,33] as well as differences in biological components such as water and coral-associated microbial communities [34].
Fig 1. Sites surveyed for coral disease in three regions (Yanbu, Thuwal, Al Lith) along the Saudi Arabian coast within the central Red Sea in Oct-Nov, 2015.
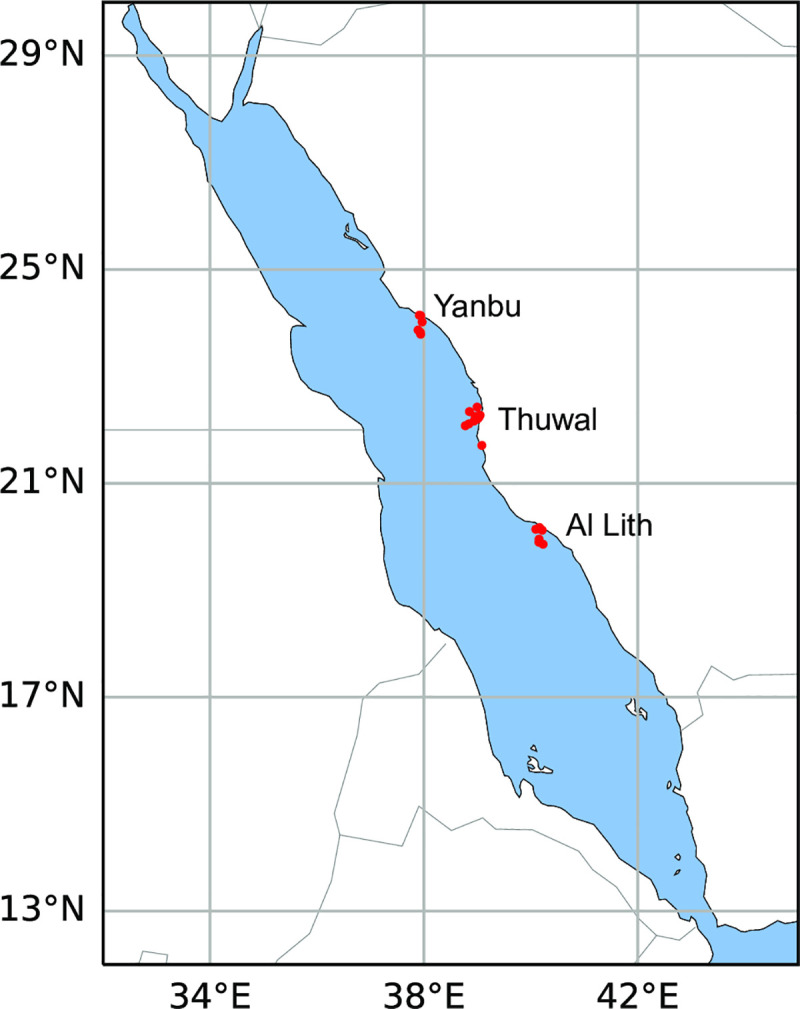
Red dots indicate survey sites within each of the regions. Coral reefs (not shown on map) can be found along its entire coastline.
Disease and bleaching surveys
Coral community structure, disease prevalence and bleaching extent was recorded at 22 sites within three regions (Al Lith, Thuwal, Yanbu) along the Red Sea coast of Saudi Arabia between October 20 and November 9, 2015 (Fig 1). Six sites were surveyed in Al Lith (depths: 4–7.6m), 10 sites in Thuwal (depths: 1.8–6.7m) and six sites in Yanbu (depths: 3.1–5.2m) (S1 Table). At each site, divers surveyed one to two 25m belt transects depending on time availability (S1 Table). Lines were haphazardly deployed and replicate transects were laid end to end separated by approximately five meters. Corals were identified to the genus level along 25 x 1m belts with the exception of some taxa that are difficult to distinguish in the field. As such, Favites and Dipsastraea were combined, Goniopora and Alveopora were combined and Lobophyllia and Symphyllia were combined. Substrate characteristics (hard coral, soft coral, crustose coralline algae, macroalgae, rubble, sand) and bleaching (color loss) were documented by point-intercept method with the substratum underlying the tape measure recorded at 25 cm intervals and coral cover scored as bleached (pale to total color loss) or healthy. Coral lesions were assessed along wider 25 x 6 m belts. Most coral diseases cannot be diagnosed in the field and so gross lesions were classified into three lesion types including tissue loss, discoloration and growth anomaly, and nomenclature for lesions was based on the host genus affected and the lesion type (e.g. Acropora growth anomaly; [35]). Tissue-loss lesions were further classified based on the lesion size, shape, presence of predators, knowledge of what common predation marks look like and evidence of lesion progression based on degree of algal colonization onto the bare coral skeleton. When diseased corals were encountered during surveys, small samples were collected of the lesion and an apparently healthy section of the same colony as a control for histology as described below. Transect lengths, widths and numbers were modified as needed when constrained by dive limits. Under the auspices of KAUST (King Abdullah University of Science and Technology), the Saudi Coastguard Authority issued sailing permits to the sites that include coral collection.
Histopathology of coral lesions
Paired normal and lesion tissues of coral lesions encountered during surveys were sampled for histopathologic analysis to characterize host response and presence of organisms visible on light microscopy [36]. Fragments were fixed in 20% zinc formaldehyde-seawater immediately after the dive and processed for routine histopathology with hematoxylin and eosin staining of sections. On microscopic exam, host response was categorized as to reversible or non-reversible changes. Reversible cellular changes included atrophy, depletion of zooxanthellae from gastrodermis, wound repair, hyperplasia of basal body wall, and inflammation whereas irreversible changes comprised necrosis and fragmentation. Visible organisms associated with lesions were classified as fungi, bacteria, cyanobacteria, sponges, or algae [36]. Tissue-loss lesions not found associated with obvious micro-organisms were termed ‘white syndrome’ indicating a tissue loss disease of unknown etiology. In addition, all samples collected during surveys for histology were also screened for cell-associated microbial aggregates (CAMA). Certain coral genera in the Indo-Pacific contain CAMAs that are proposed to be facultative secondary symbionts important in coral health [37] and so this was an opportunity to examine whether Red Sea corals also contained CAMAs.
Statistical analyses
Underwater time constraints prevented enumeration of all coral colonies within the wider belt transects surveyed for disease. Hence, prevalence of lesions was determined by extrapolating colony counts within the 25 x 1 m transect to the wider 25 x 6 m disease survey area and by using this as the denominator of prevalence calculations (e.g. (number of colonies with lesions/total number of estimated colonies) * 100). Overall prevalence was the percentage of colonies surveyed that had a particular lesion type with all surveys combined. The frequency of disease occurrence (FOC) reflects the spatial distribution of diseases on reefs and was defined as the number of sites having corals with lesions divided by total number of sites surveyed. The denominator for FOC calculations were limited to sites that had the specific coral taxa exhibiting lesions. All calculations for disease prevalence or FOC were by coral genera (e.g. prevalence of Porites growth anomalies was calculated as the total number of affected Porites colonies divided by the total number of Porites colonies surveyed, multiplied by 100. The potential for damage to coral reefs from specific diseases would depend on the spatial distribution (frequency of occurrence) as well as the proportion of colonies surveyed that were affected (prevalence). For example, a disease that is widespread within a region but affects few colonies (low prevalence) may not impact coral reef health whereas diseases that are widespread and have a high prevalence would be of concern. As such, we report both spatial distribution and prevalence of each disease. Percent bleaching was calculated from point-intercept data with the number of points with bleached cover divided by the total number of points (bleached + healthy). Climatology data for each survey site were obtained from NOAA’s Coral Reef Watch Product Suite Version 3.1 [38,39] and include the average minimum and maximum sea surface temperatures (SSTs) over the last 25 years (historical SSTs), and the degree heating weeks (DHW) for the 12 week period prior to Oct. 1, 2015.
Data were not normally distributed, even with transformation, so non-parametric analyses were used. A Kruskal-Wallis test and Dunn’s post hoc tests were used to examine regional (Al Lith, Thuwal, Yanbu) differences in coral cover, number of coral genera, colony densities, disease prevalence, percent bleaching and degree heating weeks (DHW). A pairwise analysis of similarities (ANOSIM) was performed (999 permutations) on weighted (presence and abundance of coral taxa) Bray-Curtis similarity matrices to test for significant differences in coral communities among (Al Lith, Thuwal, Yanbu) using PRIMER-E v7 (Primer-E Ltd.). To correct for multiple comparisons, a Bonferroni correction was applied to all p-values generated. Weighted nMDS plots based on Bray-Curtis similarity matrices were produced to visualize regional differences. A Simpson’s index of coral diversity, which includes species richness and evenness, was calculated for each site and ranges from 0 (low diversity) to 1 (high diversity). Disease susceptibility among coral taxa was examined using a chi-square test for equality of distributions comparing the distribution of the number of diseased versus healthy colonies among the coral genera affected by disease. A non-parametric Spearman’s rank order correlation was used to examine the relationship between disease prevalence and three potential co-factors: coral cover, percent bleaching and DHW. Non-parametric statistics were performed using JMP Pro 13 statistical software (SAS Institute Inc., Buckinghamshire, UK). The map indicating survey locations was created using reefMapMaker [40].
Results
Coral reef characteristics and coral community structure
For the 22 sites surveyed, overall average coral cover was 44.7% (range 3–83%), average soft coral cover was 13.7% (range 0–45%), average crustose coralline algae (CCA) cover was 7.2% (range 0–30%), and average macroalgae cover was <1%. Average colony density was 16.2/m2 (range 7.3–28.2). Across all sites, 30 coral genera were identified with the three dominant coral taxa being Porites (20.6% of the community), Pocillopora (14.9%) and Favites/Dipsastraea (9.5%) (S2 Table). Coral species diversity across sites was high with 20 out of 22 sites having indices greater than 0.8 (S2 Table).
Regional differences in coral cover, colony densities and genera richness
There were significant differences among the three regions (Al Lith, Thuwal, Yanbu) in coral cover (Kruskal-Wallis, X2 = 11.0, df = 2, p = 0.004), coral colony densities (Kruskal-Wallis, X2 = 7.2, df = 2, p = 0.03) and number of coral genera (Kruskal-Wallis, X2 = 13.1, df = 2, p = 0.001) (Fig 2). Coral communities were also significantly different among regions (ANOSIM, Global R = 0.13, p = 0.047) (Fig 3). Coral communities in Al Lith differed significantly from Yanbu (ANOSIM, Global R = 0.415, p = 0.013) but Thuwal was not significantly different from Yanbu (ANOSIM, Global R = 0.046, p = 0.301) or Al Lith (ANOSIM, Global R = 0.097, p = 0.169).
Fig 2.

Regional differences in coral cover (a), colony densities (b) and coral genus richness (c). Letters above bars indicate results of Dunn’s multiple group comparison tests. Symbols overlaying the bar graphs indicate the data points for all sites in each region. Six sites each were surveyed in Al Lith (triangles) and Yanbu (circles) and ten sites in Thuwal (squares).
Fig 3.

A) Average relative abundance of coral taxa in three regions surveyed for coral disease in 2015. Data show the average percent of the coral community represented by each coral genus. B) A non-metric multi-dimensional scaling (nMDS plot) illustrating the differences in coral communities among regions. Six sites each were surveyed in Al Lith and Yanbu and ten sites in Thuwal.
Regional differences in percent bleached coral and amount of heat stress
There were significant differences among the three regions (Al Lith, Thuwal, Yanbu) in percent bleached coral (Kruskal-Wallis, X2 = 14.0, df = 2, p = 0.0009) (Table 1). Sites in the Al Lith region had the highest proportion of bleached coral cover (avg. 33.5%) followed by sites in Thuwal (avg. 13.1%) with no bleaching found at sites in Yanbu. There were also significant differences among the three regions in DHW (Kruskal-Wallis, X2 = 13.4, df = 2, p = 0.001) (Table 1). Sites in Yanbu had the highest DHW (avg. 4.9), followed by Al Lith (avg. 4.4) and Thuwal (avg. 3.6). No relationship was found between percent bleached coral and DHW (Spearman’s rank, Pho = -0.24, p = 0.3).
Table 1. Summary of temperature variability and bleaching response in different study sites along the Saudi coast of the Red Sea.
| Historical SSTs | |||||
|---|---|---|---|---|---|
| Region | Site | Min (˚C) | Max (˚C) | DHW (Oct 1, 2015) | Bleached coral (%) |
| Al Lith | Al Lith 3 | 26.4 | 31.2 | 4.0 | 33.0 |
| Al Lith | Al Lith 2 | 26.3 | 31.2 | 3.8 | 34.1 |
| Al Lith | Al Lith 1 | 26.3 | 31.2 | 4.3 | 14.1 |
| Al Lith | Al Lith 6 | 26.3 | 31.1 | 4.8 | 9.6 |
| Al Lith | Al Lith 4 | 26.2 | 31.1 | 4.6 | 53.7 |
| Al Lith | Al Lith 5 | 26.2 | 31.1 | 4.7 | 36.3 |
| Al Lith | Average | 26.3 | 31.1 | 4.4 | 33.5 |
| Thuwal | La Plage | NA | NA | NA | 3.4 |
| Thuwal | Abu Madafi | 24.7 | 30.2 | 2.5 | 0.0 |
| Thuwal | Al-Mashpah | 24.7 | 30.2 | 3.1 | 9.6 |
| Thuwal | Um Alkthal | 24.6 | 30.2 | 3.7 | 0.8 |
| Thuwal | Shaab | 24.6 | 30.2 | 3.8 | 1.6 |
| Thuwal | Inner Fsar | 24.5 | 30.2 | 4.1 | 44.9 |
| Thuwal | Al Fahal | 24.5 | 30.2 | 3.9 | 19.5 |
| Thuwal | Tahlah | 24.5 | 30.2 | 4.1 | 32.3 |
| Thuwal | Shi’b Nazar | 24.5 | 30.2 | 2.9 | 14.1 |
| Thuwal | Qita al Kirsh | 24.5 | 30.2 | 3.9 | 5.0 |
| Thuwal | Average | 24.6 | 30.2 | 3.6 | 13.1 |
| Yanbu | Yanbu 3 | 24.0 | 29.9 | 5.3 | 0.0 |
| Yanbu | Yanbu 2 | 24.0 | 29.8 | 5.2 | 0.0 |
| Yanbu | Yanbu 1 | 24.0 | 29.8 | 5.1 | 0.0 |
| Yanbu | Yanbu 5 | 23.9 | 29.7 | 5.1 | 0.0 |
| Yanbu | Yanbu 4 | 23.9 | 29.7 | 4.5 | 0.0 |
| Yanbu | Yanbu 6 | 23.9 | 29.7 | 4.5 | 0.0 |
| Yanbu | Average | 24.0 | 29.7 | 4.9 | 0.0 |
Avg. minimum and maximum SSTs over the last 25 years, the Degree Heating Weeks (DHW) for the 12-week period prior to Oct. 1, 2015, and degree of bleaching is reported for each survey site.
Disease prevalence, frequency of occurrence and types of diseases
An estimated 75,750 coral colonies were examined for disease and the overall disease prevalence (all sites combined) was 0.17%. A total of 20 diseases in 16 coral taxa were recorded (Table 2). Lesion types included tissue loss diseases of unknown etiology (white syndromes), growth anomalies, distinct focal bleached patches, skeletal eroding band (folliculinid ciliate disease), black band disease (tissue loss due to microbial consortium dominated by filamentous cyanobacteria) and endolithic hypermycosis (purple discoloration due to endolithic fungal infection) (Fig 4). The three most common diseases were Acropora white syndrome found at 59.1% of the survey sites, Porites growth anomalies found at 40.9% of the sites, and Porites white syndrome found at 31.8% of the sites.
Table 2. Frequency of occurrence (FOC) and average prevalence of coral diseases recorded during surveys.
| Coral Genus | Lesion type | FOC (%) | Average Prevalence (%) |
|---|---|---|---|
| Acropora | WS | 59.1 | 2.01 (1.19) |
| GA | 9.1 | 0.95 (0.35) | |
| Astreopora | BBD | 13.6 | 0.90 (0.42) |
| WS | 9.1 | 1.43 (0.24) | |
| Echinopora | WS | 9.1 | 4.87 (1.54) |
| Favites/Dipsastraea | BBD | 13.6 | 2.57 (1.92) |
| GA | 4.5 | 0.198 | |
| Gardinoseris | BBD | 13.6 | 2.31 (0.54) |
| Goniastrea | WS | 4.5 | 1.74 |
| Goniopora | BBD | 4.5 | 0 |
| Lobophyllia | WS | 4.5 | 1.85 |
| Millepora | WS | 9.1 | 2.6 (2.0) |
| GA | 4.5 | 0.758 | |
| EH | 4.5 | 0.463 | |
| Montipora | GA | 13.6 | 0.72 (0.14) |
| WS | 4.5 | 6.41 | |
| BBD | 4.5 | 2.33 | |
| Pavona | BBD | 4.5 | 10.2 |
| EH | 4.5 | 0.249 | |
| Platygyra | BBD | 4.5 | 1.86 |
| Pocillopora | WS | 18.2 | 2.21 (1.56) |
| SEB | 13.6 | 0.29 (0.08) | |
| Porites | GA | 40.9 | 1.29 (0.62) |
| WS | 31.8 | 0.40 (0.13) | |
| Bl patch | 13.6 | 2.11 (1.70) | |
| Psammocora | WS | 4.5 | 0.08 |
| BBD | 4.5 | 4.8 | |
| EH | 4.5 | 0 | |
| Stylophora | WS | 27.3 | 7.57 (6.62) |
FOC represents the proportion of sites containing corals with each respective disease within and outside of the belt transects. Average prevalence (standard error in parentheses) calculated exclusively from the sites containing each respective disease. Absence of a standard error indicates the disease was found at a single survey site. Prevalence data includes diseased colonies only within transects and so will differ from frequency of disease occurrence data. WS = white syndrome, GA = growth anomaly, SEB = skeletal eroding band (ciliates), BBD = black band disease, Bl patch = focal bleached area, EH = endolithic hypermycosis. The most common diseases are in bold.
Fig 4. Examples of different coral diseases encountered during disease surveys along the Saudi coast of the Red Sea.

WS = white syndrome, GA = growth anomaly, EH = endolithic hypermycosis, Bl = bleached. A) black band disease, B) Acropora WS, C) Acropora GA, D) Astreopora WS, E) Echinopora WS, F) Favites GA, G) Goniastrea WS, H) Lobophyllia WS, I) Millepora WS, J) Millepora GA, K) Millepora EH, L) Pavona EH, M) Montipora WS, N) Montipora GA, O) Pocillopora WS, P) Pocillopora SEB, Q) Porites WS, R) Porites GA, S) Porites GA, T) Porites Bl patch, U) Psammocora WS, V) Psammocora EH, W) Stylophora WS.
Histopathology of coral lesions
Paired normal and abnormal tissues collected from 43 colonies representing 15 coral genera were examined (S3 Table). This included samples from 13 Porites colonies (30% of the samples), 6 Stylophora (14%), 5 Pocillopora (12%), 3 colonies each from Acropora, Astreopora, and Psammocora (7% each), 2 Gardinoseris (5%), and 1 colony each from Dipsastraea, Echinopora, Favites, Goniastrea, Goniopora, Leptoseris, Montipora, and Sarcophyton (2% each). The most common lesion sampled for histology was tissue loss (67%) (Fig 5A–5C), followed by growth anomalies (16.5%) (Fig 5D and 5E) and discoloration (16.5%) (Fig 5F). Of the 29 colonies with tissue loss, the tissue loss lesions were further subdivided based on evidence of rate of tissue loss, e.g. amount of bare, white skeleton. Rate of tissue loss was arbitrarily scored as subacute (1-5cm white skeleton)(n = 14) (Fig 5A), acute (>5cm white skeleton)(n = 10) (Fig 5B), chronic (<1cm white skeleton)(n = 3) (Fig 5C), or a combination of the three (n = 2). Histology samples originated from Yanbu (n = 13), Al Lith (n = 11), and Thuwal (n = 19).
Fig 5. Representative types of lesions sampled for histopathology.

A) Acute tissue loss (>5cm bare, white skeleton), Dipsastraea sp., B) Acute tissue loss (>5cm bare, white skeleton), Goniastrea sp., C) Chronic tissue loss (<1cm bare, white skeleton) with discoloration, Porites sp., D) Growth anomaly, Montipora sp., E) Growth anomaly, Porites sp., F) Multifocal pink discoloration with tissue swelling, Porites sp.
A total of 87 tissue samples were collected from colonies manifesting tissue loss (N = 29), growth anomalies (n = 9) and discoloration (n = 7) with the remainder being from grossly apparently normal tissues (controls). Of 29 histology sections of lesions from colonies with tissue loss, 12 had necrosis either alone or associated with fungi, cyanobacteria, algae, or sponges, 6 sections had atrophy of tissues with depletion of zooxanthellae, 5 had no evident microscopic changes, and 3 had varying degrees of inflammation associated with algae, fungi, or cyanobacteria. For 9 samples from growth anomalies, 5 had no evident changes, 3 had hyperplasia of basal body wall, and 1 had necrosis with algae. Of 7 sections with discoloration, all but 2 had necrosis with inflammation, fungi, or algae with the 2 remaining with no evident lesions. Of 42 apparently normal fragments, 16 had no evident changes, 12 had necrosis associated with fungi, cyanobacteria, algae or sponges, 8 had atrophy and depletion of zooxanthellae, 4 had inflammation sometimes associated with algae, and 1 had endolithic fungi. Of organisms associated with host response (inflammation, necrosis), fungi was the most common organism found (n = 12) followed by cyanobacteria (n = 6), algae (n = 3), and sponges (n = 1).
Cell associated microbial aggregates (CAMA)
A total of 87 coral fragments from 15 genera were examined histologically for CAMAs. CAMAs were found in four coral genera, including 5 out of 6 of the Stylophora fragments (83% of the samples examined) (Fig 6A), three out of 13 Porites fragments (21%) (Fig 6B), two out of three Acropora fragments (67%) (Fig 6C) and two out of five Pocillopora fragments (33%) (Fig 6D).
Fig 6.
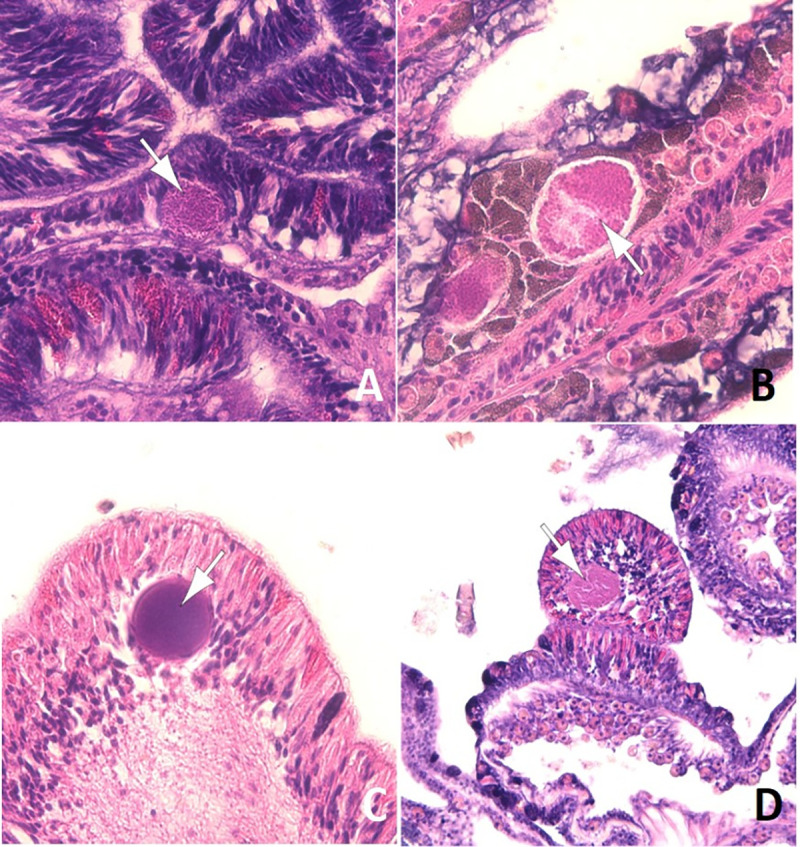
Cell associated microbial aggregates (arrows) in epidermis (A, C, D) and gastrodermis (B) of Stylophora (A), Porites (B), Acropora (C) and Pocillopora (D).
Differences in disease prevalence among coral genera
Out of 30 coral genera found within transects, 16 had lesions indicative of disease. Disease prevalence diffed among coral genera (X2 = 90.3, df = 16, p<0.005) with Acropora having the highest overall disease prevalence (0.54%), followed by Millepora (0.44%) and Lobophyllia (0.38%) (Fig 7).
Fig 7. Differences in overall disease prevalence among coral taxa.

Data show overall prevalence with all surveys combined.
Differences in disease among regions
Average disease prevalence differed significantly among regions (Kruskal-Wallis, X2 = 6.6, df = 2, p = 0.04) (Fig 8) with differences among specific diseases in the frequency of occurrence and average prevalence (Table 3). Of the three most common lesions (BBD, WS, growth anomalies), there were significant regional differences in black band disease (Kruskal-Wallis, X2 = 6.3, df = 2, p = 0.04), and white syndrome (Kruskal-Wallis, X2 = 10.2, df = 2, p = 0.006) but not growth anomalies (Kruskal-Wallis, X2 = 4.6, df = 2, p = 0.1). Average BBD prevalence was highest in Al Lith (0.33% SE±0.3), although mainly due to one outbreak site, followed by Thuwal (0.009% SE±0.005), and 0% in Yanbu. Average white syndrome prevalence was highest in Yanbu (0.47% SE±0.15) followed by Al Lith (0.06% SE±0.02) and Thuwal (0.07% SE±0.03). Average prevalence of growth anomalies was 0.11% (SE±0.04) in Yanbu, 0.04% (SE±0.02) in Thuwal and 0.003% (SE±0.003) in Al Lith.
Fig 8. Regional differences in disease prevalence at sites surveyed along the Saudi Arabian coast of the Red Sea in October-November 2015.

Letters indicate results of Dunn’s multiple group comparison tests. Symbols overlaying the bar graphs indicate the data points for all sites in each region. Six sites each were surveyed in Al Lith (triangles) and Yanbu (circles) and ten sites in Thuwal (squares).
Table 3. Regional differences in frequency of occurrence (FOC) and average prevalence (±SEM) of different coral diseases found during surveys in the central Red Sea.
| FOC (%) | Average Disease Prevalence (%) | ||||||
|---|---|---|---|---|---|---|---|
| Coral Genus | Lesion Type | Yanbu | Thuwal | Al Lith | Yanbu | Thuwal | Al Lith |
| Montipora | BBD | 0 | 0 | 16.7 | 0 | 0 | 2.33 |
| Favites/Dipsastraea | BBD | 0 | 10 | 33.3 | 0 | 0.5 | 3.6 (2.8) |
| Psammocora | BBD | 0 | 0 | 16.7 | 0 | 0 | 4.8 |
| Gardinoseris | BBD | 0 | 10 | 33 | 0 | 3.3 | 1.81 (0.29) |
| Astreopora | BBD | 0 | 10 | 33.3 | 0 | 0.32 | 1.19 (0.52) |
| Pavona | BBD | 0 | 0 | 16.7 | 0 | 0 | 10.2 |
| Platygyra | BBD | 0 | 0 | 16.7 | 0 | 0 | 1.9 |
| Goniopora | BBD | 0 | 0 | 16.7 | 0 | 0 | 0 |
| Porites | Bl patch | 16.7 | 20 | 0 | 5.1 | 0.42 (0.19) | 0 |
| Millepora | EH | 0 | 10 | 0 | 0 | 0.8 | 0 |
| Psammocora | EH | 0 | 10 | 0 | 0 | 0 | 0 |
| Pavona | EH | 0 | 10 | 0 | 0 | 0.25 | 0 |
| Acropora | GA | 0 | 20 | 0 | 0 | 0.95 (0.35) | 0 |
| Montipora | GA | 0 | 10 | 0 | 0 | 0.72 (0.14) | 0 |
| Porites | GA | 66.7 | 40 | 0 | 2.15 (1.13) | 0.56 (0.23) | 0 |
| Millepora | GA | 0 | 10 | 0 | 0 | 0.8 | 0 |
| Favites/Dipsastraea | GA | 0 | 10 | 0 | 0 | 0.2 | 0 |
| Pocillopora | SEB | 33.3 | 10 | 0 | 0.33 (0.12) | 0.2 | 0 |
| Acropora | WS | 33.3 | 80 | 50 | 8.4 (7.6) | 0.98 (0.42) | 0.5 (0.23) |
| Pocillopora | WS | 50 | 0 | 16.7 | 0.87 (0.55) | 0 | 8.3 |
| Montipora | WS | 33.3 | 0 | 0 | 6.41 | 0 | 0 |
| Porites | WS | 50 | 20 | 16.7 | 0.67 (0.04) | 0.04 (0.04) | 0.15 (0.03) |
| Echinopora | WS | 33.3 | 0 | 0 | 4.86 (1.56) | 0 | 0 |
| Millepora | WS | 33.3 | 0 | 0 | 2.6 (2.0) | 0 | 0 |
| Goniastrea | WS | 16.7 | 0 | 0 | 1.74 | 0 | 0 |
| Psammocora | WS | 0 | 0 | 16.7 | 0 | 0 | 0.08 |
| Lobophyllia | WS | 0 | 0 | 16.7 | 0 | 0 | 1.85 |
| Astreopora | WS | 0 | 10 | 16.7 | 0 | 0 | 1.67 |
| Stylophora | WS | 16.7 | 10 | 0 | 1.74 (0.74) | 0.19 (0.16) | 0.08 |
Prevalence data includes diseased colonies only within transects and so will differ from frequency of disease occurrence data. WS = white syndrome, GA = growth anomaly, SEB = skeletal eroding band (ciliates), BBD = black band disease, Bl patch = focal bleached area, EH = endolithic hypermycosis. Six sites each were surveyed in Al Lith and Yanbu and ten sites in Thuwal.
Relationship between disease prevalence and environmental factors
We examined the relationship between disease prevalence and coral cover, percent bleaching and degree heating weeks (DHW). No significant relationship was found between disease prevalence and coral cover (Spearman’s rank, rs = -0.06, p = 0.79) or DHW (Spearman’s rank, rs = 0.35, p = 0.12). A negative relationship was found between disease prevalence and coral bleaching (Spearman’s rank, rs = -0.51, p = 0.02) (Fig 9). The analyses were also done excluding the BBD outbreak and similar results found. No significant relationship was found between disease prevalence and coral cover (Spearman’s rank, rs = 0.04, p = 0.84) or DHW (Spearman’s rank, rs = 0.28, p = 0.23). A negative relationship was found between disease prevalence and coral bleaching (Spearman’s rank, rs = -0.58, p = 0.005).
Fig 9. Relationship between coral disease and level of bleaching for 22 sites surveyed in 2015 along the Red Sea coast of Saudi Arabia.

Discussion
Coral reefs are in decline globally and disease has played a significant factor in that decline [1,41,42]. Comparatively little research has been conducted on coral disease in the Red Sea and our study presents important information on types of diseases present on coral reefs along the Saudi Arabian Red Sea coast, prevalence of diseases, susceptible coral taxa within this region and a description of the histology of different coral lesions. The current study is particularly relevant considering the planned mega-building projects such as NEOM (https://www.neom.com) and the Red Sea project (https://visiontoreality.theredsea.sa), which are expected to exert heavy impacts on surrounding coral reef ecosystems. Only two prior disease studies have been carried out along the Red Sea Saudi Arabian coast with one being in the 1980s [29] and the second between 2006 and 2009 [31]. Many regions in the world, including the Red Sea, still lack basic information on coral diseases on local reefs and that is problematic. Knowing what the local threats are in a region would be important for resource managers as they develop strategies to maintain coral reefs in the face of global climate change. For this study, we surveyed twenty-two reefs in the Red Sea along the Saudi Arabian coast, and found robust hard coral (45%) and soft coral (14%) cover, and very low levels of macroalgae cover (<1%). Coral reefs in the Red Sea have experienced significant coral loss from bleaching events and crown of thorn seastar outbreaks [31,43] and our findings of notable hard coral cover suggests a degree of resilience for these reefs in the central Red Sea. Thirty hard coral genera were found within transects with the genus, Porites, numerically dominant which is consistent with other studies in the Red Sea [31,43]. Porites are known to be stress tolerant and are also dominant in other extreme environments such as the Persian/Arabian Gulf which has extreme seasonal temperature fluctuations and chronic hypersalinity [44–46].
Coral reefs had widespread but overall low prevalence of disease (<0.5%) with 20 diseases recorded affecting 16 coral taxa and disease lesions found on corals at all sites surveyed. Coral reefs in the Red Sea had diseases typical of many regions including black band disease, white syndromes, endolithic hypermycosis, skeletal eroding band, growth anomalies and distinct focal bleached patches. Identifying specific diseases in situ is difficult and should be limited to diseases with agents large enough to be seen with the naked eye, (black band disease, brown band disease, skeletal eroding band) or with lesions with a consistent microscopic presentation (endolithic hypermycosis). As example, ‘white syndromes’ are the classic example of a lesion type that is known to have different underlying etiologies, ecologies and differences in host specificity, even though the gross lesions (i.e. lesion determined underwater) may look similar [47]. In contrast, black band disease presents as progressive tissue loss associated with a microbial consortium visually dominated by filamentous cyanobacteria, which is consistent in the Indo-Pacific [48], Red Sea [49,50], and the Caribbean [51,52]. The ecology of BBD is also consistent among regions with BBD affecting a wide range of coral genera [53], is more prevalent during warm-water months [54], and has a consistent etiology [50]. Until more is known about the etiology and ecology of other coral diseases, researchers are advised to report diseases by host genera affected and lesion type [35]. As such, below we describe lesion types and report on which coral genera are affected.
Overview of diseases affecting corals in the Red Sea
Black band disease (BBD)
BBD has been reported from coral reefs across the world [55] including the Red Sea [34,50]. BBD typically remains at low background levels [56,57] with seasonal outbreaks occurring [58–60]. At our sites in the Red Sea, there was a similar pattern with a low prevalence of infected corals (0.02–0.12%) found at seven out of 22 sites which is similar to BBD levels found by Antonius [29] along the Saudi Arabian coast in the 1980s. We also documented outbreak levels (disease prevalence higher than the regional average) at one site in the Al Lith region, which has been described elsewhere [50]. In fact, 26 of the 29 BBD cases we recorded during surveys occurred at sites in Al Lith (4 out of 6 sites had colonies with BBD), with only 3 BBD cases found at sites in Thuwal (3 out of 10 sites with BBD) and no cases were found in the northern-most region (Yanbu). BBD is sensitive to water temperatures becoming more common in summer months when water temperature and light levels are higher and usually disappears during colder winter months [54,61,62]. BBD infections also appear following bleaching events[63,64]. Al Lith has higher average SST ranges (26.3˚C to 31.1˚C) as compared to Yanbu with average SSTs ranging from 24.0˚C to 29.7˚C. Al Lith also had the highest level of bleaching among the regions surveyed whereas no bleaching was found in Yanbu. Thus, higher average SSTs combined with a higher bleaching potential may leave reefs at Al Lith particularly vulnerable to BBD infections.
White syndromes (WS)
Tissue loss diseases of unknown etiology (white syndromes) are commonly found on a multitude of species on reefs throughout the world [47] and white syndromes were found in all three regions affecting 10 coral genera. Acropora white syndrome was the most widespread disease occurring in all regions at 13 of the 17 reefs surveyed. In addition, Acropora white syndrome occurred at outbreak levels at three sites with the highest level at Yanbu 2 (16% prevalence), followed by Abu Madafi (3.6% prevalence) and Al-Mashpah (1.9% prevalence) (both Thuwal region). We defined a site as having a localized disease outbreak if prevalence was higher than the overall prevalence for this region which was <1%. Acropora is an exceptionally vulnerable coral genus to tissue loss diseases throughout the world [47] and the Red Sea is no exception.
Endolithic hypermycosis (EH)
Endolithic hypermycosis was an uncommon disease with only three cases noted within three different coral genera (Millepora, Psammocora, Pavona). In other regions, this lesion is associated with overgrowth of coral tissue by endolithic fungi [65–67] and the samples we examined also had a consistent histological diagnosis of endolithic fungal invasion. This disease has been reported from American Samoa [68], Hawaii [65], Micronesia [67] and New Caledonia [66] and so the present study extends this disease to the reefs of the Red Sea (biogeographic range extension). There are no prior reports of EH in Millepora and so the present report also potentially extends the affected taxa to include Millepora, a skeleton-forming hydrozoan. However, although the gross lesion on Millepora was consistent with endolithic hypermycosis, histology was not done; therefore, confirming presence of the fungus requires future microscopic examination.
Skeletal eroding band (SEB)
Skeletal eroding band (SEB) was found exclusively on pocilloporids. SEB is caused by folliculinid ciliates with tissue loss occurring when motile larval stages migrate into the tissue edges, and secrete pseudochitinous loricae which embed in the coral skeleton [69]. The disease is characterized by a dark band of varied width, adjacent to the healthy tissue, with the denuded skeleton behind the band littered with discarded black loricae [70]. Folliculinid ciliates readily colonize recently exposed coral skeletons [71] so presence of ciliates on coral skeletons does not necessarily indicate ciliate disease. Hence, we only scored a lesion as SEB if we found ciliates within millimeters of live tissue and did not include tissue loss lesions with loricae in dead skeleton further from the lesion edge. However, as we did not follow tissue loss lesions through time, we cannot rule out that lesions that we scored as SEB, were opportunistic colonization of ciliates following tissue lost to other processes. Unfortunately, verified SEB infections have not been characterized histologically, so the role of folliculinids in contributing to gross lesions remains speculative.
SEB can be quite common on reefs affecting numerous coral taxa. For example, Page and Willis [71] found SEB at 90–100% of their survey reefs affecting at least 82 scleractinian species across the GBR. Winkler et al. [72] surveyed corals in the Gulf of Aqaba, Red Sea and found 28 coral taxa affected by SEB with an overall prevalence of 29% of the total colonies surveyed. In contrast, we only found SEB affecting pocilloporids, which are one of the most commonly affected coral genera [70] but no other coral taxa had SEB lesions. We also only found SEB infections at 3 of 22 survey sites with a prevalence of <0.5% at affected sites. The low SEB prevalence we found could be due to a more conservative approach to field diagnosis of the disease or the environmental conditions on the reefs we surveyed were not conducive to SEB infections. Transmission experiments conducted by Page and Willis [71] showed that ciliates could not colonize intact coral tissue but infections were initiated in coral with injuries. Page et al. [70] suggested that co-infection involving other pathogens and/or stress under specific environmental factors may be required for ciliates to become pathogenic.
Growth anomalies (GA)
Growth anomalies were found in five coral genera (Porites, Acropora, Montipora, Millepora, and Favites/Dipsastraea). Prior to our study, the only coral genus reported to be affected by GAs in the Red Sea was Platygyra sp. [73]. Our study now expands the biogeographic range of GAs in Porites, Acropora, Montipora, and Favites/Dipsastraea to the Red Sea. GAs in Millepora have not been reported elsewhere and so our study also expands the host range of GAs. In many regions of the world, Acropora and Porites have been found disproportionately affected by growth anomalies in the field [74] and consistent with this, Porites was the most common coral taxon affected by GAs at our sites. In contrast, acroporids were the 5th most abundant coral taxa at our sites, yet Acropora GAs were only found at two of 22 sites. Aeby et al. [75] found that the environmental predictors of GAs differed between Porites and Acropora and so the environmental conditions in the Red Sea may be conducive to GAs in Porites but not Acropora. Alternatively, different species within a single genus can differ in their disease susceptibilities [76] so perhaps the specific Acropora species found in the Red Sea are less prone to growth anomalies compared to those in other regions.
Focal bleached patches
Discrete, focal bleached patches were found in Porites spp. at three sites. Porites bleached patches have been reported from the Persian Gulf and Oman Sea [46], New Caledonia [66], and the GBR [77]. This study now extends this disease to the Red Sea (biogeographic range extension). Bleached patches are thought to be due to a viral infection of symbiotic zooxanthellae [78,79] but little else is known about this disease.
Histology shows healthy tissues compromised and presence of CAMAs
Samples for histology were collected following periods of increased sea temperatures and accumulated heat stress in all three regions surveyed and bleaching occurred in two of the regions. Reflecting this stress, we found that 61% of normal fragments (no gross lesion) had some sort of microscopic lesion, mainly necrosis, half of which were associated with a microorganism or atrophy of tissues with depletion of zooxanthellae (bleaching). Indeed, the breakdown of histologic lesions for apparently normal coral fragments was not much different than those associated with tissue loss lesions. Microscopic lesions in normal fragments is not uncommon and has been documented elsewhere. For instance, in a coral disease survey from Micronesia [67] or New Caledonia [80], ca. 26% and 28%, respectively, of normal fragments had microscopic lesions comprising changes similar to those seen here. Fungi were the dominant organisms associated with tissue loss in Saudi Arabia in most species examined. In contrast, when organisms were associated with tissue loss lesions, ciliates dominated for Acropora in the Pacific [81] whereas chimeric parasitic corals dominated for Montipora in Hawaii [82]. Cell associated microbial aggregates (CAMAs) were associated with Pocilloporidae, Poritidae, and Acroporidae thus extending a pattern similar to that in the Indo-Pacific where CAMA infect these same coral families [37]. Presence of CAMA in Stylophora has not been previously described and adds another genus of Pocilloporidae to the list of members of this family infected with bacterial aggregates. It is suggested that CAMA are facultative secondary endosymbionts of select coral genera that may be playing an important ecological role [37]. Little is known about the ecology or distribution of CAMAs but this study now reports the presence of CAMAs in corals to the Red Sea (biogeographic range expansion).
Coral taxa differ in disease susceptibility
There were differences in disease prevalence among coral genera with Acropora, Favites/Dipsastraea and Millepora having a higher disease prevalence than expected based on their abundance in the field. This is consistent with other regions of the world where disease susceptibility differs across families or genera [46,56,68,83,84]. Our study differed from other regions, in that, over half of the coral genera within transects had signs of disease (16 out of 30 coral genera). In contrast, Aeby et al. [46] found seven out of 25 coral genera with disease signs in the Persian Gulf and Williams et al. [84] found five affected coral genera in the Line Islands which has approximately 31 coral genera on its reefs [85].
Regional differences in disease and potential environmental co-factors
The survey sites were spread out along a latitudinal gradient spanning from 19˚ to 24˚N. Among the survey sites, coral cover (a measure of host abundance) ranged from 3% to 83%, degree of heat stress, as measured by DHW, ranged from 2.5 to 5.3 and amount of bleaching ranged from 0% to 54%. All three of these co-factors would be expected to affect subsequent disease prevalence and as expected, disease prevalence ranged from healthy reefs with no disease to reefs with almost 2% of the corals affected. However, no significant relationship was found between disease prevalence and coral cover or DHW. And even more surprising, there was a weak negative relationship, not a positive one as expected, between percent coral bleaching and disease prevalence. This is in contrast to what has been found on coral reefs in other regions. A positive relationship between host density and disease prevalence is considered a hallmark of the infectious process of density-dependent diseases, whereby higher host abundance results in greater rates of transmission and localized increases in prevalence [86]. In the Indo-Pacific, an association between coral cover and coral disease prevalence has been found in numerous regions [13,87–89]. Warmer temperatures, heat stress and bleaching have also been linked with higher disease prevalence [2,13,14,56]. For example, in the Persian Gulf, white syndrome outbreaks coincide with annual thermal heating events [90]. In the Caribbean, bleaching extent was linked to increased disease incidence [91] and tissue-loss disease outbreaks frequently follow bleaching events [92–94]. Our study suggests that a different pattern may be emerging for the Red Sea. The northern-most sites along the coast of Yanbu had the highest disease levels despite no bleaching occurring within transects and although heat stress was higher in this region, DHW alone was not a significant factor explaining disease prevalence.
Reef corals in the northern Red Sea have extraordinarily high thermal tolerances in relation to the ambient temperatures they usually experience [24] and our study supports this. Significant bleaching is expected when the DHW value reaches 4˚C-weeks (https://www.coralreefwatch.noaa.gov/product/5km/index_5km_dhw.php) yet our sites in Yanbu had DHW values over 4 but no bleaching was observed. Thermal tolerance in corals has been linked to host factors [95–99], Symbiodiniaceae partners [100–102] or resident microbial communities [103]. In some coral species, thermal tolerance comes at the expense of increased disease susceptibility [104,105] and this has also been suggested as a possible explanation for high disease levels found in corals in the Persian Gulf [46]. Whether there are trade-offs between disease susceptibility and thermal tolerance in corals in the central Red Sea is a hypothesis worth exploring. No work as yet been done on latitudinal gradients of microbial communities or host adaptations on corals in the Red Sea. However, within the Red Sea, the main Symbiodiniaceae genus in Porites changed from Durusdinium (D1) at warmer nearshore location to Cladocopium (C15) at cooler offshore locations [106] suggesting that differences in Symbiodiniaceae could be influencing spatial patterns of disease occurrence in this region.
Disease prevalence is low despite environmental challenges
Compared to other ocean basins, the Red Sea experiences extreme temperature variation and regularly exceeds 32˚C in the summers [18,19,107,108] which are conditions not well tolerated by most other corals. Chronic temperature stress can exert significant energetic costs on corals resulting in reduced growth and reproduction [109–111] and an increased prevalence of coral diseases [2,13]. Yet, we found corals existing in this challenging environment to have surprisingly low disease levels (<1%). Moreover, our surveys were conducted in the midst of a bleaching event. This suggests acclimation/adaptation to prevailing environmental conditions [19,112] and notably, the Red Sea is an arid region with minimal terrestrial run-off and almost no riverine input [18]. At our sites, we saw little evidence of sedimentation, water clarity was optimal with little macroalgae on the reef (<1%). Coastal coral reefs in other regions are increasingly exposed to excess nutrients, sediments, and pollutants discharged from land which are known to degrade local reefs [25] including increasing coral disease prevalence. Haapkyla et al. [11] found a 10-fold greater mean abundance of disease on reefs during the rainy summer months and concluded that rainfall and associated runoff were facilitating disease outbreaks. Laboratory studies showed that the rate of tissue loss from BBD increased with nutrient enrichment [26] and increased BBD prevalence in the field is associated with sewage effluent [113]. An experimental in situ nutrient enrichment of reefs was conducted in the Caribbean and corals exposed to chronic nutrient stress suffered a 3.5-fold increase in bleaching frequency and a two-fold increase in prevalence and severity of disease, compared to corals in control plots [27]. Terrestrial run-off also promotes the growth of macroalgae on coral reefs [25] which are major competitors with corals [114,115]. Additionally, macroalgae exude dissolved organic carbon which can disrupt the function of the coral holobiont and promote potential bacterial pathogens [116,117]. The coral reefs in the Red Sea have to contend with high temperatures and salinity, which appear to be countered by the lack of terrestrial run-off and its associated problems. As a comparison, the Persian Gulf is also an arid region but has riverine input, and massive coastal habitat modification by dredging and converting shallow, productive marine areas into land for homes, recreation, and industrial activities [118]. Resuspension of sediments is an ongoing stress for coral reefs in this region as well [119]. Under these conditions, higher coral disease levels were found and attributed to environmental stress [120]. Compared to our study, reefs along the northeastern Arabian Peninsula show a 6-fold higher disease prevalence with a high number of localized disease outbreaks [46]. Reefs surrounding Kish Island, off the coast of Iran, showed a 20-fold increase in disease [45]. Disease is a serious problem in other world regions [3,12,56] and our study suggests that a reduction in human impacts and improvement in water quality may be effective management strategies giving corals increased capacity to withstand the warming oceans predicted with global climate change.
Supporting information
(XLSX)
(XLSX)
(XLSX)
Acknowledgments
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government. We would like to thank the invaluable assistance of CMOR staff who helped with diving operations and boating.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
The author(s) received no specific funding for this work.
References
- 1.Harvell CD, Jordan-Dahlgren E, Merkel S, Rosenberg E, Raymundo L, Smith G, et al. Coral disease, environmental drivers and the balance between coral and microbial associates. Oceanography. 2007;20:172–95. [Google Scholar]
- 2.Heron SF, Willis BL, Skirving WJ, Eakin CM, Page CA, Miller IR. Summer hot snaps and winter conditions: modelling white syndrome outbreaks on Great Barrier Reef corals. PLoS ONE. 2010;5:e12210. doi: 10.1371/journal.pone.0012210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Precht WF, Gintert BE, Robbart ML, Fura R, van Woesik R. Unprecedented disease-related coral mortality in southeastern Florida. Scientific Reports. 2016;6:31374. doi: 10.1038/srep31374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Walton CJ, Hayes NK, Gilliam DS. Impacts of a regional, multi-year, multi-species coral disease outbreak in southeast Florida. Frontiers in Marine Science. 2018;5:323. [Google Scholar]
- 5.Muller EM, Sartor C, Alcaraz N, van Woesik R. Spatial epidemiology of the stony coral tissue loss disease in Florida. Frontiers in Marine Science. 2020;7:163. doi: 10.1371/journal.pone.0241871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Alvarez-Filip L, Estrada-Saldívar N, Pérez-Cervantes E, Molina-Hernández A, González-Barrios FJ. A rapid spread of the stony coral tissue loss disease outbreak in the Mexican Caribbean. PeerJ. 2019;7:e8069. doi: 10.7717/peerj.8069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ward JR, Lafferty KD. The elusive baseline of marine disease: are diseases in ocean ecosystems increasing? PLoS Biology. 2004;2:e120. doi: 10.1371/journal.pbio.0020120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Raymundo LJ, Halford AR, Maypa AP, Kerr AM. Functionally diverse reef-fish communities ameliorate coral disease. PNAS. 2009;106:17067–70. doi: 10.1073/pnas.0900365106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lamb JB, Willis BL, Fiorenza EA, Couch CS, Howard R, Rader DN, et al. Plastic waste associated with disease on coral reefs. Science. 2018;359:460–2. doi: 10.1126/science.aar3320 [DOI] [PubMed] [Google Scholar]
- 10.Pollock FJ, Lamb JB, Field SN, Heron SF, Schaffelke B, Shedrawi G, et al. Sediment and turbidity associated with offshore dredging increase coral disease prevalence on nearby reefs. PLoS ONE. 2014;9:e102498. doi: 10.1371/journal.pone.0102498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Haapkylä J, Unsworth RKF, Flavell M, Bourne DG, Schaffelke B, Willis BL. Seasonal rainfall and runoff promote coral disease on an inshore reef. PLoS ONE. 2011;6:e16893. doi: 10.1371/journal.pone.0016893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Aeby GS, Callahan S, Cox E, Runyon C, Smith A, Stanton F, et al. Emerging coral diseases in Kāne‘ohe Bay, O‘ahu, Hawai‘i (USA): two major disease outbreaks of acute Montipora white syndrome. Dis Aquat Org. 2016;119:189–98. doi: 10.3354/dao02996 [DOI] [PubMed] [Google Scholar]
- 13.Bruno JF, Selig ER, Casey KS, Page CA, Willis BL, Harvell CD, et al. Thermal Stress and coral cover as drivers of coral disease outbreaks. PLoS Biology. 2007;5:e124. doi: 10.1371/journal.pbio.0050124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Maynard J, van Hooidonk R, Eakin CM, Puotinen M, Garren M, Williams G, et al. Projections of climate conditions that increase coral disease susceptibility and pathogen abundance and virulence. Nature Clim Change. 2015;5:688–94. [Google Scholar]
- 15.Sheppard C, Price A, Roberts C. Marine ecology of the Arabian region: patterns and processes in extreme tropical environments. London: Academic Press; 1992. [Google Scholar]
- 16.Berman T, Paldor N, Brenner S. Annual SST cycle in the Eastern Mediterranean, Red Sea and Gulf of Elat: annual cycle of SST. Geophys Res Lett. 2003;30:n/a-n/a. [Google Scholar]
- 17.Kleypas JA, Mcmanus JW, Meñez LAB. Environmental limits to coral reef development: where do we draw the line? Am Zool. 1999;39:146–59. [Google Scholar]
- 18.Roik A, Röthig T, Roder C, Ziegler M, Kremb SG, Voolstra CR. Year-long monitoring of physico-chemical and biological variables provide a comparative baseline of coral reef functioning in the central Red Sea. PLoS ONE. 2016;11:e0163939. doi: 10.1371/journal.pone.0163939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Voolstra CR, Buitrago‐López C, Perna G, Cárdenas A, Hume BCC, Rädecker N, et al. Standardized short‐term acute heat stress assays resolve historical differences in coral thermotolerance across microhabitat reef sites. Glob Change Biol. 2020;26:4328–43. doi: 10.1111/gcb.15148 [DOI] [PubMed] [Google Scholar]
- 20.DiBattista JD, Roberts MB, Bouwmeester J, Bowen BW, Coker DJ, Lozano-Cortés DF, et al. A review of contemporary patterns of endemism for shallow water reef fauna in the Red Sea. J Biogeogr. 2016;43:423–39. [Google Scholar]
- 21.Berumen ML, Voolstra CR, Daffonchio D, Agusti S, Aranda M, Irigoien X, et al. The Red Sea: environmental gradients shape a natural laboratory in a nascent ocean. Coral Reefs of the Red Sea Coral Reefs of the World. Springer, Cham; 2019. Available from: 10.1007/978-3-030-05802-9_1. [DOI] [Google Scholar]
- 22.Sofianos SS. An oceanic general circulation model (OGCM) investigation of the Red Sea circulation: 2. Three-dimensional circulation in the Red Sea. J Geophys Res. 2003;108:3066. [Google Scholar]
- 23.Sawall Y, Al-Sofyani A, Hohn S, Banguera-Hinestroza E, Voolstra CR, Wahl M. Extensive phenotypic plasticity of a Red Sea coral over a strong latitudinal temperature gradient suggests limited acclimatization potential to warming. Scientific Reports. 2015;9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Osman EO, Smith DJ, Ziegler M, Kürten B, Conrad C, El‐Haddad KM, et al. Thermal refugia against coral bleaching throughout the northern Red Sea. Glob Change Biol. 2018;24:e474–84. doi: 10.1111/gcb.13895 [DOI] [PubMed] [Google Scholar]
- 25.Fabricius KE. Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Marine Pollution Bulletin. 2005;50:125–46. doi: 10.1016/j.marpolbul.2004.11.028 [DOI] [PubMed] [Google Scholar]
- 26.Voss JD, Richardson LL. Nutrient enrichment enhances black band disease progression in corals. Coral Reefs. 2006;25:569–76. [Google Scholar]
- 27.Vega Thurber RL, Burkepile DE, Fuchs C, Shantz AA, McMinds R, Zaneveld JR. Chronic nutrient enrichment increases prevalence and severity of coral disease and bleaching. Glob Change Biol. 2014;20:544–54. doi: 10.1111/gcb.12450 [DOI] [PubMed] [Google Scholar]
- 28.Shore-Maggio A, Aeby GS, Callahan SM. Influence of salinity and sedimentation on Vibrio infection of the Hawaiian coral Montipora capitata. Dis Aquat Org. 2018;128:63–71. doi: 10.3354/dao03213 [DOI] [PubMed] [Google Scholar]
- 29.Antonius A. Distribution and dynamics of coral diseases in the Eastern Red Sea. Proceedings of the 6th International Coral Reef Symposium. 1988;2:293–8. [Google Scholar]
- 30.Mohamed AR, Ali A-Ham, Abdel-Salam HA. Status of coral reef health in the northern Red Sea, Egypt. Proceedings of the 12th International Coral Reef Symposium. 2012;5. [Google Scholar]
- 31.Bruckner AW, Dempsey AC. The status, threats, and resilience of reef-building corals of the Saudi Arabian Red Sea. In: Rasul NMA, Stewart ICF, editors. The Red Sea. Berlin, Heidelberg: Springer Berlin Heidelberg; 2015. p. 471–86. Available from: http://link.springer.com/10.1007/978-3-662-45201-1_27. [Google Scholar]
- 32.Sawall Y, Al-Sofyani A, Banguera-Hinestroza E, Voolstra CR. Spatio-temporal analyses of Symbiodinium physiology of the coral Pocillopora verrucosa along large-scale nutrient and temperature gradients in the Red Sea. PLoS ONE. 2014;9:e103179. doi: 10.1371/journal.pone.0103179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sawall Y, Al-Sofyani A, Hohn S, Banguera-Hinestroza E, Voolstra CR, Wahl M. Extensive phenotypic plasticity of a Red Sea coral over a strong latitudinal temperature gradient suggests limited acclimatization potential to warming. Sci Rep. 2015;5:1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Neave MJ, Apprill A, Aeby G, Miyake S, Voolstra CR. Microbial communities of Red Sea coral reefs. In: Voolstra CR, Berumen ML, editors. Coral Reefs of the Red Sea. Cham: Springer International Publishing. p. 53–68. Available from: http://link.springer.com/10.1007/978-3-030-05802-9_4. [Google Scholar]
- 35.Work T, Aeby G. Systematically describing gross lesions in corals. Dis Aquat Org. 2006;70:155–60. doi: 10.3354/dao070155 [DOI] [PubMed] [Google Scholar]
- 36.Work TM, Russell R, Aeby GS. Tissue loss (white syndrome) in the coral Montipora capitata is a dynamic disease with multiple host responses and potential causes. Proceedings of the Royal Society B: Biological Sciences. 2012;279:4334–41. doi: 10.1098/rspb.2012.1827 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Work TM, Aeby GS. Microbial aggregates within tissues infect a diversity of corals throughout the Indo-Pacific. MEPS. 2014;500:1–9. [Google Scholar]
- 38.Skirving W, Marsh B, De La Cour J, Liu G, Harris A, Maturi E, et al. CoralTemp and the Coral Reef Watch Coral Bleaching Heat Stress Product Suite Version 3.1. Remote Sensing. 2020;12:3856. [Google Scholar]
- 39.Liu G, Skirving WJ, Geiger EF, Heron SF, Tirak KV, Strong AE, et al. NOAA Coral Reef Watch’s 5km Satellite Coral Bleaching Heat Stress Monitoring Product Suite Version 3 and Four-Month Outlook Version 4. 2017;32:7. [Google Scholar]
- 40.Hume BCC, Voolstra CR. reefMapMaker—convenient creation of user-defined regional maps with coral reef locations (Version v0.1.4). Zenodo. 2021; Available from: 10.5281/zenodo.4415326. [DOI] [Google Scholar]
- 41.Harvell CD, Kim K, Burkholder JM, Colwell RR, Epstein PR, Grimes DJ, et al. Emerging marine diseases: climate links and anthropogenic factors. Science. 1999;285:1505–10. doi: 10.1126/science.285.5433.1505 [DOI] [PubMed] [Google Scholar]
- 42.Maynard JA, Anthony KRN, Harvell CD, Burgman MA, Beeden R, Sweatman H, et al. Predicting outbreaks of a climate-driven coral disease in the Great Barrier Reef. Coral Reefs. 2010;30:485–95. [Google Scholar]
- 43.Riegl BM, Bruckner AW, Rowlands GP, Purkis SJ, Renaud P. Red Sea coral reef trajectories over 2 decades suggest increasing community homogenization and decline in coral size. PLoS ONE. 2012;7:e38396. doi: 10.1371/journal.pone.0038396 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Burt J. Persian Gulf reefs: an important asset for climate science in urgent need of pro- tection. Ocean Challenge. 2014;20:49–56. [Google Scholar]
- 45.Alidoost Salimi M, Mostafavi P, Fatemi S, Aeby G. Health status of corals surrounding Kish Island, Persian Gulf. Dis Aquat Org. 2017;124:77–84. doi: 10.3354/dao03105 [DOI] [PubMed] [Google Scholar]
- 46.Aeby GS, Howells E, Work T, Abrego D, Williams GJ, Wedding LM, et al. Localized outbreaks of coral disease on Arabian reefs are linked to extreme temperatures and environmental stressors. Coral Reefs. 2020;39:829–46. [Google Scholar]
- 47.Bourne DG, Ainsworth TD, Willis BL. White syndromes of Indo-Pacific corals. Diseases of Coral. Wiley-Blackwell; 2016. p. 300–15. [Google Scholar]
- 48.Sato Y, Willis BL, Bourne DG. Successional changes in bacterial communities during the development of black band disease on the reef coral, Montipora hispida. ISME J. 2010;4:203–14. doi: 10.1038/ismej.2009.103 [DOI] [PubMed] [Google Scholar]
- 49.Barneah O, Ben-Dov E, Kramarsky-Winter E, Kushmaro A. Characterization of black band disease in Red Sea stony corals. Environ Microbiol. 2007;9:1995–2006. doi: 10.1111/j.1462-2920.2007.01315.x [DOI] [PubMed] [Google Scholar]
- 50.Hadaidi G, Ziegler M, Shore-Maggio A, Jensen T, Aeby G, Voolstra CR. Ecological and molecular characterization of a coral black band disease outbreak in the Red Sea during a bleaching event. PeerJ. 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cooney RP, Pantos O, Tissier MDAL, Barer MR, O’Donnell AG, Bythell JC. Characterization of the bacterial consortium associated with black band disease in coral using molecular microbiological techniques. Environ Microbiol. 2002;4:401–13. doi: 10.1046/j.1462-2920.2002.00308.x [DOI] [PubMed] [Google Scholar]
- 52.Richardson L, Miller A, Broderick E, Kaczmarsky L, Gantar M, Stanic D, et al. Sulfide, microcystin, and the etiology of black band disease. Dis Aquat Org. 2009;87:79–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sutherland K, Porter J, Torres C. Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Marine Ecology Progress Series. 2004;266:273–302. [Google Scholar]
- 54.Sato Y, Bourne DG, Willis BL. Dynamics of seasonal outbreaks of black band disease in an assemblage of Montipora species at Pelorus Island (Great Barrier Reef, Australia). Proceedings of the Royal Society B: Biological Sciences. 2009;276:2795–803. doi: 10.1098/rspb.2009.0481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Raymundo L, Weil E. Indo-Pacific coloured-band diseases of corals. Diseases of corals. Hoboken, NJ: Wiley-Blackwell; 2015. [Google Scholar]
- 56.Willis BL, Page CA, Dinsdale EA. Coral Disease on the Great Barrier Reef. In: Rosenberg E, Loya Y, editors. Coral Health and Disease [Internet]. Berlin, Heidelberg: Springer Berlin Heidelberg. Available from: http://link.springer.com/10.1007/978-3-662-06414-6_3. [Google Scholar]
- 57.Page C, Willis B. Distribution, host range and large-scale spatial variability in black band disease prevalence on the Great Barrier Reef, Australia. Dis Aquat Org. 2006;69:41–51. [DOI] [PubMed] [Google Scholar]
- 58.Green EP, Bruckner AW. The significance of coral disease epizootiology for coral reef conservation. Biological Conservation. 2000;96:347–61. [Google Scholar]
- 59.Al-Moghrabi S. Unusual black band disease (BBD) outbreak in the northern tip of the Gulf of Aqaba (Jordan). Coral Reefs. 2001;19:330–1. [Google Scholar]
- 60.Aeby GS, Work TM, Runyon CM, Shore-Maggio A, Ushijima B, Videau P, et al. First Record of Black Band Disease in the Hawaiian Archipelago: Response, Outbreak Status, Virulence, and a Method of Treatment. PLOS ONE. 2015;10:e0120853. doi: 10.1371/journal.pone.0120853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kuta KG, Richardson LL. Abundance and distribution of black band disease on coral reefs in the northern Florida Keys. Coral Reefs. 1996;15:219–23. [Google Scholar]
- 62.Boyett HV, Bourne DG, Willis BL. Elevated temperature and light enhance progression and spread of black band disease on staghorn corals of the Great Barrier Reef. Mar Biol. 2007;151:1711–20. [Google Scholar]
- 63.Lewis CL. Temporal dynamics of black band disease affecting pillar coral (Dendrogyra cylindrus) following two consecutive hyperthermal events on the Florida Reef Tract. Coral Reefs. 2017;6. [Google Scholar]
- 64.Kubomura T, Yamashiro H, Reimer J. Appearance of an anomalous black band disease at upper mesophotic depths after coral bleaching. Dis Aquat Org. 2018;131:245–50. doi: 10.3354/dao03292 [DOI] [PubMed] [Google Scholar]
- 65.Work TM, Aeby GS, Stanton FG, Fenner D. Overgrowth of fungi (endolithic hypermycosis) associated with multifocal to diffuse distinct amorphous dark discoloration of corals in the Indo-Pacific. Coral Reefs. 2008;27:663–663. [Google Scholar]
- 66.Aeby G, Tribollet A, Lasne G, Work T. Assessing threats from coral and crustose coralline algae disease on the reefs of New Caledonia. Marine and Freshwater Research. 2015;67:455–65. [Google Scholar]
- 67.Work TM, Aeby GS, Hughen KA. Gross and microscopic lesions in corals from Micronesia. Vet Pathol. 2016;53:153–62. doi: 10.1177/0300985815571669 [DOI] [PubMed] [Google Scholar]
- 68.Aeby G, Work T, Fenner D, Didonato E. Coral and crustose coralline algae disease on the reefs of American Samoa. Proceedings of the 11th International Coral Reef Symposium. 2008;7:197–201. [Google Scholar]
- 69.Antonius AA, Lipscomb D. First protozoan coral-killer identified in the Indo-Pacific. Atoll Research Bulletin. 2001;481:1–21. [Google Scholar]
- 70.Page CA, Cróquer A, Bastidas C, Rodríguez S, Neale SJ, Weil E, et al. Halofolliculina ciliate infections on corals (Skeletal Eroding Disease). In: Woodley CM, Downs CA, Bruckner AW, Porter JW, Galloway SB, editors. Diseases of Coral. Hoboken, NJ: John Wiley & Sons, Inc; 2015. p. 361–75. Available from: http://doi.wiley.com/10.1002/9781118828502.ch26. [Google Scholar]
- 71.Page CA, Willis BL. Epidemiology of skeletal eroding band on the Great Barrier Reef and the role of injury in the initiation of this widespread coral disease. Coral Reefs. 2008;27:257–72. [Google Scholar]
- 72.Winkler R, Antonius A, Abigail Renegar D. The skeleton eroding band disease on coral reefs of Aqaba, Red Sea. Marine Ecology. 2004;25:129–44. [Google Scholar]
- 73.Loya Y. The coral reefs of Eilat—past, present and future: three decades of coral community structure studies. Coral Health and disease. Springer. 2004; p. 1–34. [Google Scholar]
- 74.Work TM, Kaczmarsky L, Peters EC. Skeletal growth anomalies in corals. In: Woodley CM, Downs CA, Bruckner AW, Porter JW, Galloway SB, editors. Diseases of Coral. Hoboken, NJ: John Wiley & Sons, Inc; 2016. p. 291–9. [Google Scholar]
- 75.Aeby GS, Williams GJ, Franklin EC, Haapkyla J, Harvell CD, Neale S, et al. Growth anomalies on the coral genera Acropora and Porites are strongly associated with host density and human population size across the Indo-Pacific. PLoS ONE. 2011;6:e16887. doi: 10.1371/journal.pone.0016887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Aeby GS, Bourne DG, Wilson B, Work TM. Coral diversity and the severity of disease outbreaks: A cross-regional comparison of Acropora white syndrome in a species-rich region (American Samoa) with a species-poor region (Northwestern Hawaiian Islands). Journal of Marine Biology. 2011;2011:1–8. [Google Scholar]
- 77.Roff G, Ulstrup KE, Fine M, Ralph PJ, Hoegh-Guldberg O. Spatial heterogeneity of photosynthetic activity within diseased corals from the Great Barrier Reef. Journal of Phycology. 2008;44:526–38. doi: 10.1111/j.1529-8817.2008.00480.x [DOI] [PubMed] [Google Scholar]
- 78.Lawrence SA, Davy JE, Wilson WH, Hoegh-Guldberg O, Davy SK. Porites white patch syndrome: associated viruses and disease physiology. Coral Reefs. 2015;34:249–57. [Google Scholar]
- 79.Kenkel CD, Mocellin VJL, Bay LK. Global gene expression patterns in Porites white patch syndrome: Disentangling symbiont loss from the thermal stress response in reef‐building coral. Mol Ecol. 2020;29:3907–20. doi: 10.1111/mec.15608 [DOI] [PubMed] [Google Scholar]
- 80.Work TM, Aeby GS, Lasne G, Tribollet A. Gross and microscopic pathology of hard and soft corals in New Caledonia. Journal of Invertebrate Pathology. 2014;120:50–8. doi: 10.1016/j.jip.2014.05.007 [DOI] [PubMed] [Google Scholar]
- 81.Work TM. Pathology of tissue loss (white syndrome) in Acropora sp. corals from the Central Pacific. Journal of Invertebrate Pathology. 2011;5. doi: 10.1016/j.jip.2011.03.009 [DOI] [PubMed] [Google Scholar]
- 82.Work TM, Forsman ZH, Szabó Z, Lewis TD, Aeby GS, Toonen RJ. Inter-specific coral chimerism: genetically distinct multicellular structures associated with tissue loss in Montipora capitata. PLoS ONE. 2011;6:e22869. doi: 10.1371/journal.pone.0022869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Aeby GS, Williams GJ, Franklin EC, Kenyon J, Cox EF, Coles S, et al. Patterns of coral disease across the Hawaiian Archipelago: relating disease to environment. PLoS ONE. 2011;6:e20370. doi: 10.1371/journal.pone.0020370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Williams G, Knapp I, Aeby G, Davy S. Spatial and temporal patterns of scleractinian coral, soft coral, and zoanthid disease on a remote, near-pristine coral reef (Palmyra Atoll, central Pacific). Dis Aquat Org. 2011;94:89–100. [DOI] [PubMed] [Google Scholar]
- 85.Williams GJ, Maragos JE, Davy SK. Characterization of the coral communities at Palmyra Atoll in the remote central Pacific Ocean. Atoll Research Bulletin. 2008;557:1–32. [Google Scholar]
- 86.Lafferty KD, Gerber LR. Good medicine for conservation biology: the intersection of epidemiology and conservation theory. Conservation Biology. 2002;16:12. [Google Scholar]
- 87.Haapkylä J, Unsworth R, Seymour A, Melbourne-Thomas J, Flavell M, Willis B, et al. Spatio-temporal coral disease dynamics in the Wakatobi Marine National Park, South-East Sulawesi, Indonesia. Dis Aquat Org. 2009;87:105–15. doi: 10.3354/dao02160 [DOI] [PubMed] [Google Scholar]
- 88.Myers R, Raymundo L. Coral disease in Micronesian reefs: a link between disease prevalence and host abundance. Dis Aquat Org. 2009;87:97–104. doi: 10.3354/dao02139 [DOI] [PubMed] [Google Scholar]
- 89.Aeby GS, Ross M, Williams GJ, Lewis TD, Work TM. Disease dynamics of Montipora white syndrome within Kaneohe Bay, Oahu, Hawaii: distribution, seasonality, virulence, and transmissibility. Dis Aquat Org. 2010;91:1–8. [DOI] [PubMed] [Google Scholar]
- 90.Howells EJ, Vaughan GO, Work TM, Burt JA, Abrego D. Annual outbreaks of coral disease coincide with extreme seasonal warming. Coral Reefs. 2020;39:771–81. [Google Scholar]
- 91.Brandt ME, Mcmanus JW. Disease incidence is related to bleaching extent in reef-building corals. 90:10. doi: 10.1890/08-0445.1 [DOI] [PubMed] [Google Scholar]
- 92.Miller J, Muller E, Rogers C, Waara R, Atkinson A, Whelan KRT, et al. Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs. 2009;28:925–37. [Google Scholar]
- 93.Muller EM, Rogers CS, Spitzack AS, van Woesik R. Bleaching increases likelihood of disease on Acropora palmata (Lamarck) in Hawksnest Bay, St John, US Virgin Islands. Coral Reefs. 2008;27:191–5. [Google Scholar]
- 94.Cróquer A, Weil E. Changes in Caribbean coral disease prevalence after the 2005 bleaching event. Dis Aquat Org. 2009;87:33–43. doi: 10.3354/dao02164 [DOI] [PubMed] [Google Scholar]
- 95.Jokiel PL. Temperature Stress and Coral Bleaching. In: Rosenberg E, Loya Y, editors. Coral Health and Disease. Berlin, Heidelberg: Springer Berlin Heidelberg; 2004. p. 401–25. Available from: http://link.springer.com/10.1007/978-3-662-06414-6_23. [Google Scholar]
- 96.Dove SG, Hoegh-Guldberg O. The cell physiology of coral bleaching. In: Phinney JT, Hoegh‐Guldberg O, Kleypas J, Skirving W, Strong A, editors. Coastal and Estuarine Studies]. Washington, D. C.: American Geophysical Union; 2006. p. 55–71. Available from: http://www.agu.org/books/ce/v061/61CE05/61CE05.shtml. [Google Scholar]
- 97.Baird AH, Bhagooli R, Ralph PJ, Takahashi S. Coral bleaching: the role of the host. Trends in Ecology & Evolution. 2009;24:16–20. doi: 10.1016/j.tree.2008.09.005 [DOI] [PubMed] [Google Scholar]
- 98.Fitt WK, Gates RD, Hoegh-Guldberg O, Bythell JC, Jatkar A, Grottoli AG, et al. Response of two species of Indo-Pacific corals, Porites cylindrica and Stylophora pistillata, to short-term thermal stress: the host does matter in determining the tolerance of corals to bleaching. Journal of Experimental Marine Biology and Ecology. 2009;373:102–10. [Google Scholar]
- 99.Howells EJ, Abrego D, Meyer E, Kirk NL, Burt JA. Host adaptation and unexpected symbiont partners enable reef-building corals to tolerate extreme temperatures. Glob Change Biol. 2016;22:2702–14. doi: 10.1111/gcb.13250 [DOI] [PubMed] [Google Scholar]
- 100.Ulstrup K, Berkelmans R, Ralph P, van Oppen M. Variation in bleaching sensitivity of two coral species across a latitudinal gradient on the Great Barrier Reef: the role of zooxanthellae. Mar Ecol Prog Ser. 2006;314:135–48. [Google Scholar]
- 101.Berkelmans R, van Oppen MJH. The role of zooxanthellae in the thermal tolerance of corals: a ‘nugget of hope’ for coral reefs in an era of climate change. Proc R Soc B. 2006;273:2305–12. doi: 10.1098/rspb.2006.3567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Bay LK, Doyle J, Logan M, Berkelmans R. Recovery from bleaching is mediated by threshold densities of background thermo-tolerant symbiont types in a reef-building coral. R Soc open sci. 2016;3:160322. doi: 10.1098/rsos.160322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ziegler M. Bacterial community dynamics are linked to patterns of coral heat tolerance. Nature Communications. 2017;8:1–8. doi: 10.1038/s41467-016-0009-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Shore-Maggio A, Callahan S, Aeby GS. Trade-offs in disease and bleaching susceptibility among two color morphs of the Hawaiian reef coral, Montipora capitata. Coral Reefs. 2018;37:507–17. [Google Scholar]
- 105.Merselis DG, Lirman D, Rodriguez-Lanetty M. Symbiotic immuno-suppression: is disease susceptibility the price of bleaching resistance? PeerJ. 2018;6:e4494. doi: 10.7717/peerj.4494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Ziegler M, Roder C, Büchel C, Voolstra C. Niche acclimatization in Red Sea corals is dependent on flexibility of host-symbiont association. Mar Ecol Prog Ser. 2015;533:149–61. [Google Scholar]
- 107.Davis KA, Lentz SJ, Pineda J, Farrar JT, Starczak VR, Churchill JH. Observations of the thermal environment on Red Sea platform reefs: a heat budget analysis. Coral Reefs. 2011;30:25–36. [Google Scholar]
- 108.Chaidez V, Dreano D, Agusti S, Duarte CM, Hoteit I. Decadal trends in Red Sea. Scientific Reports. 2017;7:8144. doi: 10.1038/s41598-017-08146-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Porter JW, Lewis SK, Porter KG. The effect of multiple stressors on the Florida Keys coral reef ecosystem: A landscape hypothesis and a physiological test. Limnol Oceanogr. 1999;44:941–9. [Google Scholar]
- 110.De’ath G, Lough JM, Fabricius KE. Declining Coral Calcification on the Great Barrier Reef. Science. 2009;323:116–9. doi: 10.1126/science.1165283 [DOI] [PubMed] [Google Scholar]
- 111.Cantin NE, Cohen A, Karnauskas K, Tarrant A, McCorkle D. Ocean warming slows coral growth in the Central Red Sea. Science. 2010;329:322–5. doi: 10.1126/science.1190182 [DOI] [PubMed] [Google Scholar]
- 112.Voolstra C, Valenzuela J, Turkarslan S, Cardenas A, Hume B, Perna G, et al. Contrasting heat stress response patterns of coral holobionts across the Red Sea suggest distinct mechanisms of thermal tolerance. In Review; 2020. Dec. Available from: https://www.researchsquare.com/article/rs-117181/v1. [DOI] [PubMed] [Google Scholar]
- 113.Kaczmarsky LT, Draud M, Williams EH. Is there a relationship between proximity to sewage effluent and the prevalence of coral disease? Caribbean Journal of Science. 2005;41:14. [Google Scholar]
- 114.Tanner JE. Competition between scleractinian corals and macroalgae: An experimental investigation of coral growth, survival and reproduction. Journal of Experimental Marine Biology and Ecology. 1995;190:151–68. [Google Scholar]
- 115.McCook L, Jompa J, Diaz-Pulido G. Competition between corals and algae on coral reefs: a review of evidence and mechanisms. Coral Reefs. 2001;19:400–17. [Google Scholar]
- 116.Haas AF, Nelson CE, Wegley Kelly L, Carlson CA, Rohwer F, Leichter JJ, et al. Effects of coral reef benthic primary producers on dissolved organic carbon and microbial activity. PLoS ONE. 2011;6:e27973. doi: 10.1371/journal.pone.0027973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Barott KL, Rohwer FL. Unseen players shape benthic competition on coral reefs. Trends in Microbiology. 2012;20:621–8. doi: 10.1016/j.tim.2012.08.004 [DOI] [PubMed] [Google Scholar]
- 118.Sheppard C, Al-Husiani M, Al-Jamali F, Al-Yamani F, Baldwin R, Bishop J, et al. The Gulf: A young sea in decline. Marine Pollution Bulletin. 2010;60:13–38. doi: 10.1016/j.marpolbul.2009.10.017 [DOI] [PubMed] [Google Scholar]
- 119.Riegl B. Corals in a non-reef setting in the southern Arabian Gulf (Dubai, UAE): fauna and community structure in response to recurring mass mortality. Coral Reefs. 1999;18:63–73. [Google Scholar]
- 120.Riegl BM, Bruckner AW, Samimi-Namin K, Purkis SJ. Diseases, harmful algae blooms (HABs) and their effects on Gulf coral populations and communities. In: Riegl BM, Purkis SJ, editors. Coral Reefs of the Gulf. Dordrecht: Springer Netherlands; 2012. p. 107–25. Available from: http://www.springerlink.com/index/10.1007/978-94-007-3008-3_7. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.