

Abstract
Phages are viruses of bacteria and are the smallest and most common biological entities in the environment. They can reproduce immediately after infection or integrate as a prophage into their host genome. SPβ is a prophage of the Gram-positive model organism Bacillus subtilis 168, and it has been known for more than 50 years. It is sensitive to dsDNA damage and is induced through exposure to mitomycin C or UV radiation. When induced from the prophage, SPβ requires 90 min to produce and release about 30 virions. Genomes of sequenced related strains range between 128 and 140 kb, and particle-packed dsDNA exhibits terminal redundancy. Formed particles are of the Siphoviridae morphotype. Related isolates are known to infect other B. subtilis clade members. When infecting a new host, SPβ presumably follows a two-step strategy, adsorbing primarily to teichoic acid and secondarily to a yet unknown factor. Once in the host, SPβ-related phages pass through complex lysis–lysogeny decisions and either enter a lytic cycle or integrate as a dormant prophage. As prophages, SPβ-related phages integrate at the host chromosome's replication terminus, and frequently into the spsM or kamA gene. As a prophage, it imparts additional properties to its host via phage-encoded proteins. The most notable of these functional proteins is sublancin 168, which is used as a molecular weapon by the host and ensures prophage maintenance. In this review, we summarise the existing knowledge about the biology of the phage regarding its life cycle and discuss its potential as a research object.
Introduction
Phages are viruses of bacteria and are the smallest and most common biological entities in the environment. As viruses, they depend on the metabolism of their bacterial hosts for reproduction. During the reproductive process, most phage types completely consume the resources of their hosts and kill them when releasing their progeny [1, 2]. Phages that reproduce immediately after infection are called lytic phages (lytic life cycle). However, some phage types are able to reproduce via a temperate life cycle (Fig. 1) in which they insert their genetic information into the genome of the host bacterium, thus becoming prophages, which subsequently multiply passively through the growth of their host. The process of prophage incorporation into the host chromosome is called lysogenisation, and the resulting bacterium with the prophage is called a lysogen. The genetic material of the prophage is transferred to the daughter cells with each cell division. The bacterium can proliferate without any disadvantages (apart from increased energy expenditure for prophage DNA replication), together with its inactive prophage. Sometimes the lysogen acquires a competitive advantage through the presence of its prophage in the form of phage resistance [3, 4] or prototrophy [5, 6]. In rare cases, a prophage can even turn the bacterium into a pathogen (lysogenic conversion). For example, the pathogenicity of enterohemorrhagic Escherichia coli (EHEC) and Vibrio cholerae is due to the presence of a prophage [7, 8]. When the bacterial cell is exposed to stress, the prophage can be activated and enter the lytic life cycle again.
Fig. 1.
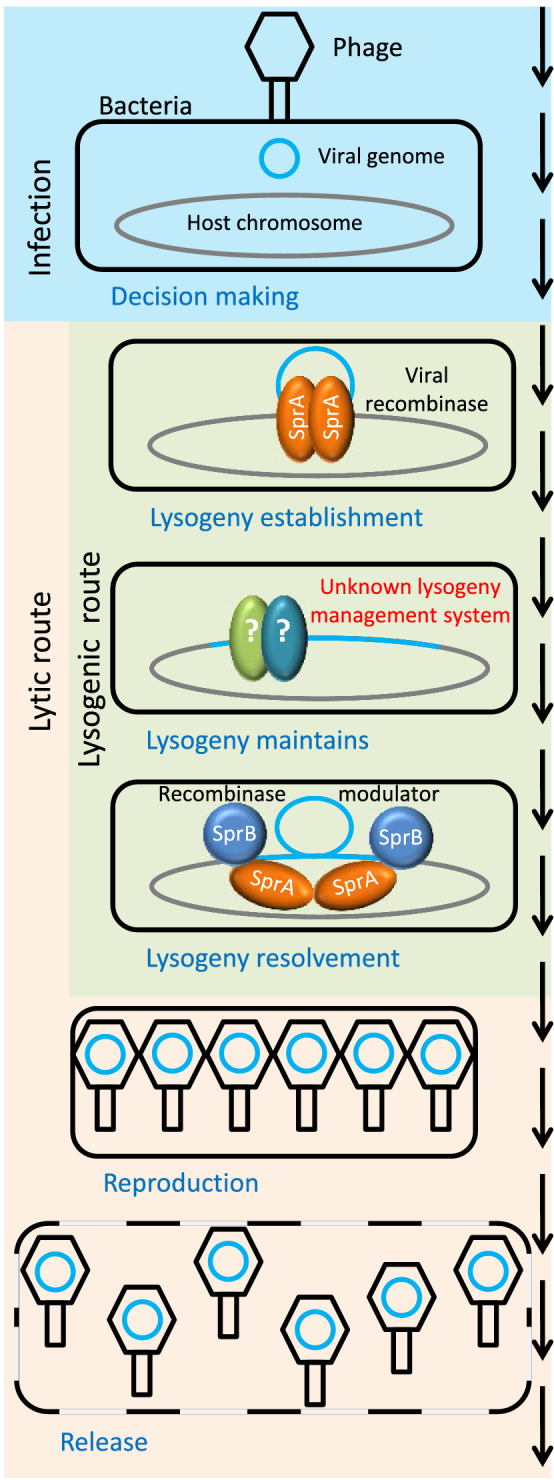
The life cycle of SPβ and related phages
SPβ infects the Gram-positive model bacterium Bacillus subtilis 168 and, as a prophage, significantly impacts the properties of its host. Like its host, it has great potential to serve as a model system for phage biology. In this review, we summarise the existing knowledge about the life cycle of phage SPβ and its impact on its host bacterium.
Host system
Bacillus subtilis was first described by W. Cohn in 1875 as a small sporulating bacterium. The original strain was lost, and in 1930, H. J. Conn proposed the Marburg strain as the new type strain because it best matched the original descriptions of B. subtilis [9], which was soon accepted by the scientific community [10]. In 1947, Burkholder and Giles exposed this strain to a sublethal dose of X-rays and generated a tryptophan-auxotrophic strain named 168 [11]. This strain was distributed worldwide due to its high transformability, as shown by John Spizizen [12]. Thus, B. subtilis 168 became a model organism for many aspects of bacterial molecular biology [13] and one of the most frequently used hosts for B. subtilis phages [10].
The genome sequence of B. subtilis 168 was first published in 1997 by Kunst et al. [14], was resequenced by Barbe et al. in 2009 [15], and has since undergone frequent annotation updates [16, 17], making it one of the best-characterised bacterial genomes. The current version of the genome sequence indicates that it consists of one chromosome 4,215,606 bp in size with 34.98% GC content. It contains 4,325 protein coding regions and encodes 86 tRNAs, 30 rRNAs, two ncRNAs, and 93 small RNAs (misc_RNA). Furthermore, it harbours one integrative and conjugative element (ICEBs1) [18], four prophage-like-regions, and two prophages, known as PBSX [19] and SPβ [20]. All of these alien genomic elements are non-essential for the lysogen and can be removed from the genome of B. subtilis 168 [21].
Bacillus pumilus, Bacillus licheniformis, and Bacillus amyloliquefaciens are other well-known representatives of the B. subtilis clade [22, 23]. These species are morphologically very similar and are mainly mesophiles and neutrophiles [23]. Members of this clade are frequently suitable hosts for phages that were initially isolated on B. subtilis, such as φ29 [24, 25], SP-15 [26], or SPO1 [27], and for diverse SPβ-related phages, as recent bioinformatic analysis has demonstrated [28].
SPβ and related isolates
The characterisation of SPβ was first described in the PhD thesis of F. A. Eiserling at the University of California at Los Angeles in 1964. Two "defective" phages of B. subtilis 168 were investigated and named SPα and SPβ [10]. Edna Seaman and co-workers independently characterised both phages a second time and named them PBSX (SPα) and PBSY (SPβ) [19]. However, the designation SPβ has become generally accepted. SPβ has an icosahedral head (82 to 88 nm in diameter) and a 12-nm-wide and 320-nm-long flexible non-contractile tail, with a 36-nm-wide baseplate exhibiting six equidistant, radial projections [10, 29]. Thus, it resembles the Siphoviridae morphotype (Fig. 2), like the small temperate B. subtilis phage φ105 [30] and the small lytic phage SPP1 [31].
Fig. 2.
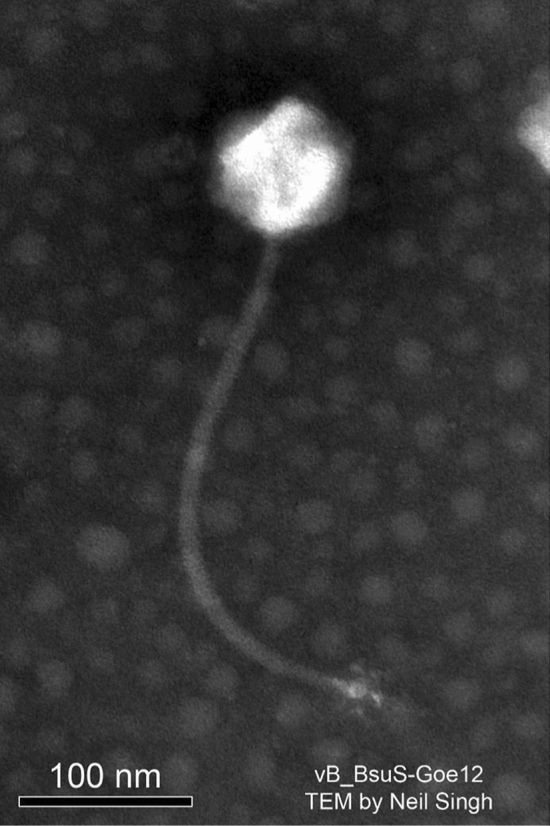
Virion of the SPβ-like phage Goe12 (vB_BsuS-Goe12)
This phage was not further investigated until the discovery of B. subtilis CU1050, a strain that was spontaneously cured of SPβ and is therefore suitable for its lytic replication [29, 32]. Many SPβ-related phage isolates have been reported in the literature, including IG1, IG3, IG4 [33], φ3T [34], Z [33], ρ11 [35], and SPR [36]. Almost all are directly associated with B. subtilis. IG4 can also be propagated on B. pumilus [33], and although H2 originated from a B. amyloliquefaciens lysogen [37], it can also lysogenise B. subtilis [38].
SPβ-related phages fall into three subgroups [39, 40]. Phages IG1, IG3, IG4, ρ11, and φ3T are particularly closely related to SPβ, as SPβ antiserum cross-reacts with all of these phages [33, 39]. SPR belongs to the second subgroup. Antiserum against this phage does not inactivate the above-mentioned strains. In addition, the SPR viral DNA exhibits a unique methylation pattern [36]. Phage H2 represents a third subgroup [37, 38]. It has less genome sequence similarity to other SPβ-like phages and a specific antiserum activity [39]. Three recent isolates, Goe11 [MT601272.1], Goe12 [MT601273.1], and Goe13 [MT601274.1], have been sequenced. Average nucleotide sequence identity analysis shows them to belong to the same species as SPβ and to cluster together with SPβ and φ3T (Fig. 3).
Fig. 3.
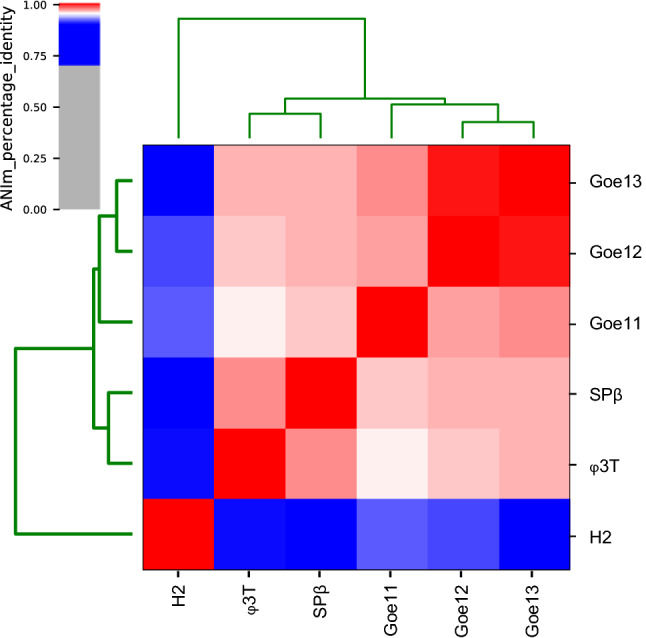
Average nucleotide sequence identity of SPβ-related phages. Blue–white (70–95% identity) indicates affiliation to the same genus; and white–red (95–100% identity), to the same species.
SPβ induction and release
The availability of a suitable host strain has enabled the investigation of SPβ in more detail and and verified it as functional and capable of lytic reproduction. It forms small turbid plaques on the SPβ-free strain CU1050 [29, 32]. As prophage, it is activated by mitomycin C and thus reacts to DNA damage [29]. If induced in B. subtilis 168, it has a latent period of about 90 min and a burst size of 28-36 viable particles. The clear-plaque mutant SPβ c1 is incapable of lysogenising its host. It has a shorter reproduction cycle of only 46 min and a reduced burst size of 16 progeny particles [29]. However, as this phage mutant was never sequenced, the genetic basis of its accelerated reproduction remains unclear.
When mitomycin C enters into bacterial cells, it intercalates into the dsDNA, forming cross-links that in turn cause stalling of the replication fork and RecA activation [41–43]. Activated RecA stimulates proteolytic auto-cleavage of (i) the LexA repressor, thereby activating the SOS response, and (ii) phage repressors, thereby leading to prophage induction, as has been proposed as an induction mechanism for the PBSX prophage [37]. SPβ induction by mitomycin C most likely relies on a more complex mechanism. The YonR protein of SPβ shows pronounced similarity to the lysogenic repressor Xre for lysogeny maintenance of PBSX and YqaE of the skin element, a degenerated prophage of B. subtilis 168. YonR has been proposed to be the lysogenic repressor of SPβ [44]. The d protein (YomJ) was identified as a further lysogeny management component. If expressed ectopically from a plasmid, it conveys resistance against closely related phage strains [45]. However, neither yonR nor yomJ seems to be essential for maintaining the lysogenic state of SPβ, as both could be individually deleted from the prophage genome [46]. These facts suggest a lysogeny control mechanism involving two independent repressors and/or host components, or an activator-based switch to the lytic pathway. Each of the proposed cases would present a new system that is worth scientific attention.
There are indications of the involvement of a host-specific component in SPβ induction. A genomic analysis of SPβ by Lazarevic et al. revealed the presence of four SOS boxes, two in front of the divergently transcribed genes yolC <-> yolD, one at yopS <-> yopT, and one in front of yorB [44]. Further analysis by Au et al. led to the identification of three additional SOS boxes associated with SPβ-related genes (Table 1) [47]. Those SOS boxes provide a binding site for LexA, also known as DinR (damage-inducible regulator) [48]. Au and co-workers were able to confirm in vitro binding of LexA to the promoter regions of the yorB, yolC, and yolD genes. However, it remains unclear how the SOS-box-associated genes of SPβ respond to SOS induction in the host. For technical reasons, B. subtilis YB886, a genetic descendant of the SPβ-free strain CU1050 [29, 32, 49], was used for those experiments. LexA is not responsible for silencing SPβ, as it can be deleted from the host genome without activating the prophage [46]. The possibility that a combination of host- and phage-derived components such as LexA and YonR may handle this task jointly remains to be explored. Another possible host factor involved in SPβ maintenance is the extracytoplasmic function (ECF) sigma factor SigY [50]. Deletion mutants of SigY spontaneously lose the SPβ prophage.
Table 1.
SOS boxes identified in the SPβ genome
| Genea | SOS boxb | Positionc | KdD (nM) | No. of mismatches |
|---|---|---|---|---|
| yokF | aGAACAaAcaTTCt | − 17 | n. d. | 5 (1) |
| yolC (1) | aGAACAaAcGTTCt*1 | − 127 (− 74) | 3.9 | 4 (0) |
| yolC (2) | aGAACAaAaGTTCG*2 | − 97 (− 44) | 3 (0) | |
| yolD (1) | CGAACtTtTGTTCt*2 | − 64 (− 17) | 3.9 | 3 (0) |
| yolD (2) | aGAACgTtTGTTCt*1 | − 34 (+ 14) | 4 (0) | |
| yonT | CGAACATAaGTTtt | − 320 | n. d. | 3 (1) |
| yopS | gGAACgTgcGTTCt*3 | − 119 | n. d. | 5 (0) |
| yopT | aGAACgcAcGTTCc*3 | − 51 | n. d. | 5 (0) |
| yorB | aGAACActTGTTCc | − 62 | 12 | 4 (0) |
| yorL | aGAACtTgTGTTtt | − 15 | n. d. | 5 (1) |
| Consensus | CGAACATATGTTCG |
The data originate from reference [47]
*The operator instance to regulate different genes on the leading and lagging strands
aGenes are listed by the position in the SPβ c2 genome. The numbers in parentheses indicate different operators within the same promoter region
bLowercase nucleotides and uppercase nucleotides indicate nonconsensus and consensus residues, respectively. The underlined sequences were identified previously by Lazarevic et al. [44]
cLocation of the 3' end of the SOS box relative to the ATG codon of the respective gene (and relative to the 3' end of the -10 region of the canonical SigA promoter sequence)
dApparent binding constant of LexA determined by Au et al. [47]
Once SPβ is induced, SprA and SprB proteins manage its excision. The recombination directionality factor SprB promotes synapsis of SprA subunits bound to the attL and attR sites and releases the phage genome with the simultaneous reconstitution of the spsM gene [51, 52]. While the sprA gene is constitutively expressed [53, 54], the expression of the sprB gene depends on the activity of the SigK or SigE sigma factor [52] or a stress-induced SigA promoter [53]. These different promoters enable the excision of the SPβ genome, not only upon induction with mitomycin C (SigA) but also during sporulation (SigK and SigE), where SPβ is removed from the genome to re-establish the SpsM function and thus ensure proper spore surface glycosylation (see below) [53]. A similar prophage excision and gene re-establishment during sporulation was observed with the kamA gene when disrupted by the φ3T prophage [55].
So far, members of the SPβ group are the largest known temperate phages of B. subtilis. Their genomes range in size from 128 and 140 kb (SPβ c2, 134 kb [44]; φ3T, 128 kb [56]; H2, 140 kb [positions 1979051..2118711 in CP041693]). Fig. 4 shows a circular map of the SPβ phage genome that is representative of the entire phage group. In the B. subtilis 168 chromosome, SPβ is integrated into the spsM gene, creating two truncated genes: yodU (BSU_19810) and ypqP (BSU_21670). When excised from the bacterial chromosome during sporulation or phage induction, the spsM gene is reconstituted. It codes for a sugar epimerase, which is needed for spore polysaccharide coating by the mother cell during sporulation [53]. The sprA (BSU_21660) and sprB (BSU_19820) genes are essential for the excision process. Both are located at the outer margins of the SPβ prophage and thus mark its boundaries. In the circular SPβ genome, these genes flank the attP site of the phage [53].
Fig. 4.
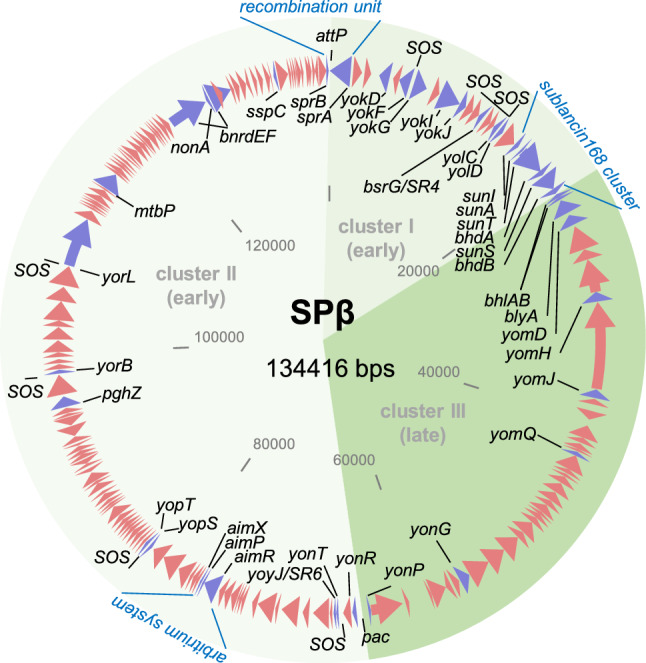
The genome of Bacillus phage SPβ. The genome orientation and the locations of its genomic clusters I–III are defined with respect to phage replication (I, early; II, early; III-late) and adjusted according to Lazarevic et al. [44]. Arrows indicate protein-coding genes. Red arrows represent genes encoding hypothetical proteins not discussed in this review. Purple arrows represent genes discussed in this review to which gene names are connected with a black line. The attP site, the pac site, the SOS boxes, the recombination unit, the sublancin 168 cluster, and the arbitrium system are indicated. The genome map's initial structure was created with Clone Manager 8 (Sci Ed Software, Westminster, Colorado, USA) using the SPβ c2 genome sequence [44] and elaborated further with MS PowerPoint 2019.
Upon induction, SprB modulates SprA to excise the SPβ genome from the host chromosome and presumably circularise it for replication [52, 53]. With the bnrdEF genes, SPβ provides an additional ribonucleotide reductase, likely to meet the increased demand for deoxyribonucleotides during its replication. Apart from the significant similarity of the phage-encoded system to the host system (nrdEF), each of the phage ribonucleotide reductase genes contains an intron, and bnrdE harbours an additional intein-coding sequence. Whether these mobile elements fulfil a function during phage replication or are dispensable remains unclear [57, 58].
The replication processes of SPβ have not yet been investigated. It can just be speculated to be similar to that of SPP1, which is capable of theta and sigma replication (reviewed in reference [59]). Sigma replication, also called rolling-circle replication, results in a phage genome concatemer, which serves as a substrate for genome packaging. Before genome packaging, MtbP of SPβ methylates the first cytosine of the GGCC palindrome at position 5 of the pyrimidine ring [36, 44, 60, 61]. In this way, the SPβ genome is protected from the host's defence systems during the next infection cycle [62].
The genome packaging mechanism of SPβ also remains to be elucidated, but it is also likely to be similar to the one used by SPP1. Different SPβ mutants (SPβ c1 del1, SPβ c2 del2, SPβ c2 del3, and SPβ c2 del4) have been constructed with an about 10% reduced genome size relative to that of the wild type [63, 64]. When a cos site is used for phage genome maturation, the genome is cut into defined units during packaging [65], but in the case of these deletion mutants, the phage head is not filled correctly with DNA due to reduced genome size, resulting in unstable viral particles with reduced viability [66]. However, to our knowledge, such observations have not been reported for SPβ mutants bearing deletions [64]. Alternatively, encapsidation can be initiated at a pac site, employing a head-full mechanism [67]. In this case, the pac site is cleaved, and the phage genome's concatemeric DNA is unidirectionally translocated into the interior of a procapsid until the phage head is full. A sequence-independent DNA cut terminates the process. The encapsidated genome exceeds 100% and thus contains terminal redundancy, which is later necessary for re-circularisation. With phage SPP1, when the first head is filled following the first concatemer cut, the remaining dsDNA is used directly for further encapsidation cycles. As a result, a heterogeneous population of terminally redundant and partially circularly permuted DNA molecules is generated (reviewed in reference 68). Thus, a shortened SPβ genome would lead to elongated terminal redundancies while still forming a correctly filled head and stable particles. Sequence read distribution analysis of new SPβ-like isolates (Goe11, Goe12, and Goe13) strengthens the assumption of a pac site and headful-based genome packaging by SPβ-like phages (our own unpublished data).
Restriction mapping studies of SPβ from particle-packed genomic DNA have indicated the location of the potential pac sites between or around yonR (BSU_21020) and yonP (BSU_21030) [69]. However, a comparison with the SPP1 pac site [70] revealed no sequence similarities (data not shown).
Virion assembly and release
The SPβ gene cluster between yonP and bhlB has been proposed to contain structural genes required for particle assembly [44]. This cluster is transcribed as a single polycistronic mRNA by the SPβ-specific single-subunit RNA polymerase YonO, which is active at the late stage of replication [71].
Almost nothing is known about the virion assembly of the SPβ phage. The yomH gene, annotated as "tail family protein" [NC_001884], and the yomQ gene, annotated as "putative tail phage assembly protein" [NC_000964], are the only two genes where annotation indicates a potential involvement in particle formation. Even an automatic annotation of the recently sequenced genome of a B. subtilis 168 derivative [72] includes no new information about structural genes of SPβ.
The gene cluster from yomD to bhlB (nt 2,262,437-2,265,169 in NC_000964) can be associated with cell lysis required for the release of viral progeny. The yomD gene (BSU_21400) codes for a hypothetical protein, which shows no homology to any known domain or structure (InterProScan 20200716, data not shown). It overlaps with a gene (BSU_21409) that is separated from blyA (yomC) by an 18-bp-long intergenic region. The blyA gene codes for N-acetylmuramoyl-L-alanine amidase, which resembles the phage lysin [73]. The proximity of these three genes implies a joint function, which awaits experimental confirmation. The other two genes of this cluster, bhlA (yomB) and bhlB (yomA), code for potential holin-like proteins, which presumably initiate membrane permeabilisation required for the phage lysin to reach its target and lyse the host cell [73].
Infection and Immunity
Finally released by bursting the host cell, SPβ may start a new infection cycle. This might be challenging, as a B. subtilis population often forms an extracellular matrix consisting of poly-γ-glutamic acid (γ-PGA), which can prevent the phage from gaining access to the host cell [74]. However, the virulent B. subtilis phage phiNIT1 can overcome this barrier by employing a phage-derived γ-PGA hydrolase [74]. The SPβ-derived gene pghZ (BSU_20460) codes for such a functional γ-PGA hydrolase [75], and this might help SPβ to overcome the γ-PGA barrier.
Like some other B. subtilis phages, SPβ primarily adsorbs to the teichoic acid of the host cell wall [40, 76]. The possibility that it uses a protein as a second receptor protein, like the morphologically related lytic phage SPP1 [77], remains open. Phage SPP1 adsorption occurs in two steps, the first of which is reversible but more efficient. The second step is irreversible and associated with the cell wall protein YueB, but it is unlikely to happen without prior reversible association with cell-wall teichoic acid [78]. Without the correct teichoic acid, SPP1 can still bind to YueB, but with reduced efficiency and preferably on solid media [78]. SPβ may follow a similar adsorption strategy. B. subtilis strain IGCgll4, an SPβ-resistant strain, has been reported to harbour a mutation affecting the biosynthesis of teichoic acid. This mutation abolishes the adsorption of SPβ and confers resistance to this phage [40]. However, the observed resistant phenotype may be a misinterpretation. Reduced adsorption strongly impacts plaque formation and lysis of the host culture, mainly because the surrounding host cells grow faster than the phages can infect them [79]. This assumption is supported by transduction experiments showing the ability of SPβ to inject DNA into strain IGCgll4 [39]. These results imply a secondary attachment site for SPβ to which the phage adsorbs poorly.
Once in the host, which lifestyle to choose?
Besides a direct lytic replication a direct lytic replication cycle that kills the host, SPβ, as a temperate phage, can also coexist with the host by becoming a prophage via integration of its DNA into the host chromosome, thereby generating a lysogenic bacterial strain. In its prophage form, SPβ propagates passively through the replication of its host [80].
The choice between a lysogenic and a lytic lifestyle was investigated for φ3T, a close relative of SPβ [56]. The "arbitrium" system of this group of phages is the first step in the decision process. It relies on small-molecule communication to execute lysis–lysogeny decisions [56]. The arbitrium system consists of AimR, a transcription activator; AimP, a quorum-sensing signal peptide; and AimX, a non-coding RNA (ncRNA), which is a lysogeny activator or lysis repressor. AimP is a small peptide with an N-terminal signal sequence that allows it to be secreted into the medium via the B. subtilis Sec pathway. Outside the cell, it is further processed by extracellular proteases to generate the final hexapeptide, which is reimported by the host oligopeptide transporter (OppABCDF) into the cytosol, where it interacts with the AimR regulator. The AimR regulator, when free of AimP, binds as a homodimer to the aimX operator, represented by two hexameric inverted repeats separated by 25 bp [81], and promotes aimX transcription (lytic cycle). The φ3T phage AimR dimer dissociates to monomers after interaction with the AimP peptide, the concentration of which increases during infection of a host population. The monomeric AimR cannot bind its operator and promote aimX transcription [56], thus favouring lysogeny. In SPβ, the association of the AimR dimer with the AimP peptides leads to a closed conformation of the AimR dimer that makes it unable to bind its operator [81] and promote the expression of the aimX ncRNA gene. Crosstalk of the arbitrium systems of different phage strains may be feasible, but it has already been found that φ3T and SPβ have incompatible systems, even though they belong to the same subgroup [56, 81].
Recent findings indicate that the integrative and conjugative elements ICEBs1 may take the choice of lysis–lysogeny from SPβ. The SpbK protein, encoded by the spbK gene on ICEBs1, interferes with the lytic reproduction of SPβ and thus leaves this phage only the option of lysogenisation [82]. This action implies an attempt by the host and its ICEBs1 element to "domesticate" SPβ.
Establishing lysogeny
When establishing lysogeny, a long-term relationship between the host bacterium and the phage in its prophage form, SPβ-related phages preferably integrate at the replication terminus of the bacterial chromosome [5, 28, 83, 84]. The phage attachment site sequence (attP) for SPβ has been identified [52]. On the circularised phage genome, it is located between the sprA (BSU_21660) and sprB (BSU_19820) genes, has an AT-rich core region, and is flanked by inverted repeats. The corresponding bacterial attachment site sequence (attB) is located in spsM, which is interrupted upon integration, resulting in two pseudogenes: ypqP and yodU [53].
Regarding historical SPβ-related strains, the genome sequence is available only for φ3T and H2. Phage φ3T has a different orientation of its sprA gene in the viral genome and integrates its prophage in the kamA gene, coding for lysine 2,3-aminomutase [28]. Its attL and attR sites have a 5-bp conserved core (CCTAC) that is likely to represent the DNA breakpoint for integration. Adjacent to this core sequence, there are imperfect inverted repeat sequences (23-24 bp long) that presumably provide binding sites for a site-specific recombinase [55]. Thus, the φ3T integrase system differs from that of SPβ. This may explain the possibility of φ3T-SPβ double lysogeny, despite the significant similarity between the two phages [29]. Phage H2 lysogenises B. amyloliquefaciens H [CP041693.1] by integrating between two hypothetical genes. However, it also lysogenises B. subtilis and integrates itself between the tyrA and metB loci [38]. A comparison of the H2 core attP site with the relevant region of the B. subtilis genome [NC_000964.3] revealed only a putative imperfect attB sequence (5'-CCCttTAaAAATAACTA-3') at positions 2,333,200-2,333,216. The recent SPβ-like isolates Goe12 [MT601273.1] and Goe13 [MT601274.1] have the same core attP site as SPβ [52] and probably have the same integration locus as SPβ. In contrast, the sprA orientation in Goe11 [MT601272.1] resembles that found in φ3T. Analysis of a Goe11 lysogen revealed a putative integration site identical to that of φ3T (unpublished personal data). Integration site data of SPβ-related isolates are presented in Table 2.
Table 2.
Known core attP sites of SPβ-related phages
| Phage strain | Host | attP sequence (5′– > 3′) | attB sequence (5′– > 3′) | Integration locus |
|---|---|---|---|---|
| SPβ | B. subtilis 168 | ACAGATAA/AGCTGTAT | ACAGATAA/AGCTGTAT | spsM |
| H2 | B. amyloliquefaciens H | CCCTATAAATAACTA | CCCTATAAATAACTA | Intergenic |
| H2 | B. subtilis 168 | CCCTATAAATAACTA* | CCCttTAaAAATAACTA* | Intergenic* |
| φ3T | B. subtilis 168 | aaaatgacataCCTACtgtgttttta | gctatgcggttCCTACctttgtcgtt | kamA |
| Goe11 | B. subtilis ∆6 | aaaatgacataCCTACtgtgtttttt* | gctatgcggttCCTACctttgtcgtt* | kamA* |
| Goe12 | B. subtilis ∆6 | ACAGATAA/AGCTGTAT* | ACAGATAA/AGCTGTAT* | spsM* |
| Goe13 | B. subtilis ∆6 | ACAGATAA/AGCTGTAT* | ACAGATAA/AGCTGTAT* | spsM* |
The SPβ att sites were oriented based on Abe et al., 2017 [52]. Capital letters represent the inverted repeat recognised by SprA, a forward slash indicates the cleavage site, and underlined italic letters represent the 3' overhangs. The att sites of φ3T are presented as described by Suzuki et al., 2020 [55]. Capital letters indicate the conserved core, and lowercase letters indicate the associated imperfect repeat sequences. The att site sequences and integration loci marked with a star (*) are based on personal bioinformatic investigations and have not been confirmed experimentally.
The regulation of the integration process has not yet been investigated. The large serine recombinase SprA is expressed continuously and is responsible for integrating the phage genome into the bacterial chromosome [53], but the prophage itself is established just after the decision by the arbitrium system [56], implying a further layer of regulation.
Not much is known about how SPβ-like phages maintain their lysogenic state. The ~22-kDa d protein of SPβ [GenBank no. M13821.1], when ectopically expressed in an SPβ-sensitive host, prevents lytic replication or lysogenisation of the bacterium by SPβ. This protein seems to be constitutively synthesised, but its production is most likely regulated by other phage- encoded factors. It is either unstable or has an elevated turnover rate in vivo. It has been successfully expressed in a B. subtilis SPβ c2-free background, but it could not be detected via immunoblotting methods in an SPβ c2 lysogen [45]. However, the d protein is functional and biologically active in SPβ c1 and c2. It is neither responsible for the clear-plaque phenotype of SPβ c1/c10 nor the temperature sensitivity of c2, as those strains have been shown to have an intact d gene [45]. A frameshift mutation was engineered to inactivate the d protein. Introduction of the mutated d gene into SPβ-sensitive B. subtilis does not confer resistance to SPβ. When the damaged d gene is introduced into SPβ c1, it no longer has a clear-plaque phenotype but instead produces turbid plaques characteristic of lysogenic bacteria. Those lysogens are unstable and lose the prophage in further passages. In the temperature-sensitive SPβ c2 strain, the introduction of the mutated d gene leads to a clear-plaque phenotype, indicating the inability to form lysogens [45]. Thus, inactivation of the d protein in the clear-plaque mutant c1 and the temperature-sensitive-mutant c2 results in opposite phenotypes. This is intriguing, since complementation experiments suggest that both phenotypes are attributable to the same gene [85]. These observations point to a complex process of lysogeny establishment and maintenance in SPβ-related phages with potentially yet unknown components. Some, for example, may be located between sprA (yokA) and sunI (yolF), as a deletion of this region in the SPβ c2 del3 mutant has been shown to be associated with unstable lysogeny [63].
Host conversion
Lysogenic conversion is the alteration of the properties of the host by its prophage, which introduces new genetic information into the bacterial chromosome. As mentioned above, this phenomenon is sometimes associated with the transformation of harmless bacteria into pathogens [86]. It is unlikely that SPβ-related phages turn their hosts into pathogens, as B. subtilis is generally recognised as safe and is even used for food production [87]. However, SPβ carries a gene (yokG) encoding a protein similar to the insecticidal delta endotoxin of B. thuringiensis, but its biological role remains unclear [44].
Regarding nucleotide metabolism, φ3T can convert its host from deoxyribosylthymine auxotrophy to prototrophy through the phage-encoded thymidylate synthase gene thyP3 [5, 6]. SPβ does not have a thymidylate synthase gene, while its original host B. subtilis 168 contains two genes for this function (thyA -BSU_17680 and thyB - BSU_21820) [14, 15]. Still, SPβ has the ability to acquire the thyP3 gene from φ3T through phage-phage homologous recombination during a mixed infection, or through the transformation of B. subtilis 168 with the corresponding φ3T DNA fragment [88, 89].
The sspC gene, provided by SPβ [44], encodes an α/β-type small acid-soluble spore protein (SASP) [90, 91], which stabilises spore DNA and increases spore resistance to UV light [92]. Its transcription is regulated by the forespore-specific SigG factor and is thus part of the host sporulation regulatory network [93].
SPβ contains the genes sunI (yolF), sunA (yolG), sunT (yolH), bhdA (yolI), sunS (yolU), and bdbB (yolK), with which it can weaponise the host with sublancin 168 [94, 95]. The sunA gene codes for the pre-peptide [96] which is post-translationally glycosylated by SunS at its cysteine residue 22 [97]. The remaining four cysteines are oxidised by BdbA and, in particular, by BdbB, resulting in two disulfide bounds [98, 99]. SunT exports this glycopeptide by proteolytic cleavage of the leader peptide [98]. The exported sublancin 168 inhibits the growth of many Gram-positive bacteria [100]. The active form of sublancin 168 is imported into sensitive cells by a glucose phosphate transferase system and inhibits synthesis of DNA, RNA, and protein [101]. SunI provides immunity against sublancin 168 and thus protects the SPβ lysogen [102]. While the sunA gene is regulated, sunI is constitutively expressed [54, 102]. The differential expression of the sunA gene in specific cell types during the late exponential and stationary growth phase enables B. subtilis to compete for its resources [103].
The SPβ yokD gene codes for a putative aminoglycoside N3'-acetyltransferase. This typically bacterial enzyme can confer resistance to aminoglycoside antibiotics, such as gentamicin or kanamycin [104, 105]. However, it remains unclear whether YokD plays a particular role in SPβ biology.
The SPβ nonA gene endows its host with the capability of protecting itself against unrelated lytic phages such as SP10 by aborting their infections. The small transmembrane protein NonA inhibits the synthesis of the capsid protein of SP10 and is regulated by sigma factors encoded by SP10 itself (Orf199-Orf200) [3, 4].
A selfish phage
The production of sublancin 168 by SPβ may benefit the host but may also turn against it. During periods of adequate nutrient supply and logarithmic growth, B. subtilis frequently loses the SPβ prophage [50]. When entering the stationary phase, the remaining lysogens activate the sublancin 168 production and thereby eliminate all SPβ-free cells [103]. This reveals that sublancin 168 is also part of the prophage maintenance system.
An even more selfish aspect of SPβ can be found in its three toxin-antitoxin (TA) systems. Such two-component systems consist of a stable toxin, which kills the cell or causes growth stasis if produced in a certain amount, and an unstable antitoxin that controls the toxin. TA systems were first discovered to be encoded by plasmids and were shown to contribute to their maintenance by killing the host if the plasmid got lost [106]. In such a case, the antitoxin levels decrease rapidly, allowing the stable toxin to kill the plasmid-free cell (reviewed in reference 107). The TA systems of SPβ may have a similar function.
The bsrG/SR4 system is a type I TA system consisting of an antisense ncRNA (SR4), which controls a toxin (BsrG) mRNA [108]. SR4 acts through RNA-RNA interaction by inhibiting the translation of bsrG and promoting its degradation [109]. The yonT/yoyJ/SR6 system is the second type I toxin-antitoxin (TA) system. The toxin genes yonT and yoyJ act independently but are transcribed on one polycistronic mRNA. The antisense ncRNA SR6 resides on the DNA strand opposite to that of the toxins genes and partially covers the coding region of both toxins. It interacts with the 3' untranslated region of yonT mRNA, thereby promoting its degradation by RNase III, and controls the translation of yoyJ by directly binding to its ribosome-binding site [110]. The only type II TA system identified in SPβ consists of YokI and YokJ proteins [111, 112]. Further investigation is needed to elucidate how those TAs are implemented in the regulatory circuits of SPβ and how they fulfil their biological function.
Concluding remarks
Despite half a century of research on SPβ and the fact that its host, B. subtilis 168, is one of the best-studied prokaryotic model systems, there are still blind spots in our understanding of its biology. We barely know how SPβ maintains its prophage status and how it decides to enter the lytic cycle. Not much more is known about the replication of its genome and the assembly of the infectious virion. For the majority of genes, no function has been assigned. It is also not known which genes are essential for lytic or lysogenic replication. A plethora of open questions remain to be answered to understand the biology of phage SPβ.
Acknowledgements
We thank Birthe Nordmann, Neil Singh, and Michael Hoppert for their help with the Goe12 micrograph, and Fabian M. Commichau and Vladimir Lazarevic for stimulating discussions, suggestions, and constructive revision of the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL. This work was supported by the Georg-August University of Göttingen, the Brandenburg Technische Universität Cottbus-Senftenberg, and the Volkswagen Foundation (Re. 94045).
Declarations
Conflict of interest
The author declares no conflict of interest.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Salmond GPC, Fineran PC. A century of the phage: past, present and future. Nat Rev Microbiol. 2015;13:777–786. doi: 10.1038/nrmicro3564. [DOI] [PubMed] [Google Scholar]
- 2.Dion MB, Oechslin F, Moineau S. Phage diversity, genomics and phylogeny. Nat Rev Microbiol. 2020;18:125–138. doi: 10.1038/s41579-019-0311-5. [DOI] [PubMed] [Google Scholar]
- 3.Yamamoto T, Obana N, Yee LM, et al. SP10 infectivity is aborted after bacteriophage SP10 infection induces nonA transcription on the prophage SPβ region of the Bacillus subtilis genome. J Bacteriol. 2014;196:693–706. doi: 10.1128/JB.01240-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rettenmier CW, Gingell B, Hemphill HE. The role of temperate bacteriophage SPβ in prophage-mediated interference in Bacillus subtilis. Can J Microbiol. 1979;25:1345–1351. doi: 10.1139/m79-212. [DOI] [PubMed] [Google Scholar]
- 5.Williams MT, Young FE. Temperate Bacillus subtilis bacteriophage φ3T: chromosomal attachment site and comparison with temperate bacteriophages φ105 and SPO2. J Virol. 1977;21:522–529. doi: 10.1128/JVI.21.2.522-529.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gardner AL, Aronson AI. Expression of the Bacillus subtilis glutamine synthetase gene in Escherichia coli. J Bacteriol. 1984;158:967–971. doi: 10.1128/JB.158.3.967-971.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Asadulghani M, Ogura Y, Ooka T, et al. The defective prophage pool of Escherichia coli O157: prophage–prophage interactions potentiate horizontal transfer of virulence determinants. PLoS Pathog. 2009;5:e1000408. doi: 10.1371/journal.ppat.1000408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kim EJ, Yu HJ, Lee JH, et al. Replication of Vibrio cholerae classical CTX phage. Proc Natl Acad Sci USA. 2017;114:2343–2348. doi: 10.1073/pnas.1701335114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Conn HJ. The identity of Bacillus subtilis. Médecine Mal Infect. 1971;1:45–50. doi: 10.1016/S0399-077X(71)80199-3. [DOI] [Google Scholar]
- 10.Hemphill HE, Whiteley HR. Bacteriophages of Bacillus subtilis. Bacteriol Rev. 1975;39:257–315. doi: 10.1128/br.39.3.257-315.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Burkholder PR, Giles NH. Induced biochemical mutations in Bacillus subtilis. Am J Bot. 1947;34:345–348. doi: 10.2307/2437147. [DOI] [PubMed] [Google Scholar]
- 12.Spizizen J. Transformation of biochemically deficient strains of Bacillus subtilis by deoxyribonucleate. Proc Natl Acad Sci USA. 1958;44:1072–1078. doi: 10.1073/pnas.44.10.1072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sonenshein AL, Hoch JA, Losick R. Bacillus subtilis and its closest relatives. Washington: ASM Press; 2001. [Google Scholar]
- 14.Kunst F, Ogasawara N, Moszer I, et al. The complete genome sequence of the Gram-positive bacterium Bacillus subtilis. Nature. 1997;390:249–256. doi: 10.1038/36786. [DOI] [PubMed] [Google Scholar]
- 15.Barbe V, Cruveiller S, Kunst F, et al. From a consortium sequence to a unified sequence: the Bacillus subtilis 168 reference genome a decade later. Microbiology. 2009;155:1758–1775. doi: 10.1099/mic.0.027839-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Belda E, Sekowska A, Le Fèvre F, et al. An updated metabolic view of the Bacillus subtilis 168 genome. Microbiology. 2013;159:757–770. doi: 10.1099/mic.0.064691-0. [DOI] [PubMed] [Google Scholar]
- 17.Borriss R, Danchin A, Harwood CR, et al. Bacillus subtilis, the model Gram-positive bacterium: 20 years of annotation refinement. Microb Biotechnol. 2018;11:3–17. doi: 10.1111/1751-7915.13043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Auchtung JM, Aleksanyan N, Bulku A, Berkmen MB. Biology of ICE Bs1, an integrative and conjugative element in Bacillus subtilis. Plasmid. 2016;86:14–25. doi: 10.1016/j.plasmid.2016.07.001. [DOI] [PubMed] [Google Scholar]
- 19.Seaman E, Tarmy E, Marmur J. Inducible phages of Bacillus subtilis. Biochemistry. 1964;3:607–613. doi: 10.1021/bi00893a001. [DOI] [PubMed] [Google Scholar]
- 20.Brodetsky AM, Romig WR. Characterization of Bacillus subtilis Bacteriophages. J Bacteriol. 1965;90:1655–1663. doi: 10.1128/JB.90.6.1655-1663.1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Westers H, Dorenbos R, van Dijl JM, et al. Genome engineering reveals large dispensable regions in Bacillus subtilis. Mol Biol Evol. 2003;20:2076–2090. doi: 10.1093/molbev/msg219. [DOI] [PubMed] [Google Scholar]
- 22.Rooney AP, Price NPJ, Ehrhardt C, et al. Phylogeny and molecular taxonomy of the Bacillus subtilis species complex and description of Bacillus subtilis subsp. inaquosorum subsp. nov. Int J Syst Evol Microbiol. 2009;59:2429–2436. doi: 10.1099/ijs.0.009126-0. [DOI] [PubMed] [Google Scholar]
- 23.Fritze D. Taxonomy of the genus Bacillus and related genera: the aerobic endospore-forming bacteria. Phytopathology. 2004;94:1245–1248. doi: 10.1094/PHYTO.2004.94.11.1245. [DOI] [PubMed] [Google Scholar]
- 24.Meijer WJJ, Horcajadas JA, Salas M. φ29 family of Phages. Microbiol Mol Biol Rev. 2001;65:261–287. doi: 10.1128/MMBR.65.2.261-287.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Reilly BE, Spizizen J. Bacteriophage deoxyribonucleate infection of competent Bacillus subtilis. J Bacteriol. 1965;89:782–790. doi: 10.1128/JB.89.3.782-790.1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Taylor MJ, Thorne CB. Transduction of Bacillus licheniformis and Bacillus subtilis by each of two Phages1. J Bacteriol. 1963;86:452–461. doi: 10.1128/JB.86.3.452-461.1963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Klumpp J, Lavigne R, Loessner MJ, Ackermann HW. The SPO1-related bacteriophages. Arch Virol. 2010;155:1547–1561. doi: 10.1007/s00705-010-0783-0. [DOI] [PubMed] [Google Scholar]
- 28.Dragoš A, Priyadarshini B, Hasan Z, et al. Pervasive prophage recombination occurs during evolution of spore-forming Bacilli. ISME J. 2020 doi: 10.1038/s41396-020-00854-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Warner FD, Kitos GA, Romano MP, Hemphill HE. Characterization of SPβ: a temperate bacteriophage from Bacillus subtilis 168M. Can J Microbiol. 1977;23:45–51. doi: 10.1139/m77-006. [DOI] [Google Scholar]
- 30.Zeigler DR. Complete genome sequence of Bacillus subtilis phage φ105. Genome Announc. 2013;1:3401. doi: 10.1128/genomeA.e00641-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Godinho LM, El Sadek FM, Monniot C, et al. The revisited genome of Bacillus subtilis Bacteriophage SPP1. Viruses. 2018;10:705. doi: 10.3390/v10120705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Johnson CM, Grossman AD. Complete genome sequence of Bacillus subtilis strain CU1050, which is sensitive to phage SPβ. Genome Announc. 2016;4:e00262–e316. doi: 10.1128/genomeA.00262-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fernandes RM, de Lencastre H, Archer LJ. Three new temperate phages of Bacillus subtilis. Microbiology. 1986;132:661–668. doi: 10.1099/00221287-132-3-661. [DOI] [PubMed] [Google Scholar]
- 34.Tucker RG. Acquisition of thymidylate synthetase activity by a thymine-requiring mutant of Bacillus subtilis following infection by the temperate phage φ3. J Gen Virol. 1969;4:489–504. doi: 10.1099/0022-1317-4-4-489. [DOI] [PubMed] [Google Scholar]
- 35.Dean DH, Orrego JC, Hutchison KW, Halvorson HO. New temperate bacteriophage for Bacillus subtilis, ρ11. J Virol. 1976;20:509–519. doi: 10.1128/JVI.20.2.509-519.1976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Noyer-Weidner M, Jentsch S, Pawlek B, et al. Restriction and modification in Bacillus subtilis: DNA methylation potential of the related bacteriophages Z, SPR, SPβ, φ3T, and ρ11. J Virol. 1983;46:446–453. doi: 10.1128/JVI.46.2.446-453.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zahler SA, Korman RZ, Thomas C, Odebralski JM. Temperate Bacteriophages of Bacillus amylolquefaciens. Microbiology. 1987;133:2933–2935. doi: 10.1099/00221287-133-10-2933. [DOI] [PubMed] [Google Scholar]
- 38.Zahler SA, Korman RZ, Thomas C, et al. H2, a temperate bacteriophage isolated from Bacillus amyloliquefaciens strain H. Microbiology. 1987;133:2937–2944. doi: 10.1099/00221287-133-10-2937. [DOI] [PubMed] [Google Scholar]
- 39.Weiner MP, Zahler SA. Genome homology and host range of some SPβ-related Bacteriophages of Bacillus subtilis and Bacillus amyloliquefaciens. J Gen Virol. 1988;69:1307–1315. doi: 10.1099/0022-1317-69-6-1307. [DOI] [PubMed] [Google Scholar]
- 40.Estrela AI, De Lencastre H, Archer LJ. Resistance of a Bacillus subtilis mutant to a group of temperate bacteriophages. Microbiology. 1986;132:411–415. doi: 10.1099/00221287-132-2-411. [DOI] [PubMed] [Google Scholar]
- 41.Lovett CM, O’Gara TM, Woodruff JN. Analysis of the SOS inducing signal in Bacillus subtilis using Escherichia coli LexA as a probe. J Bacteriol. 1994;176:4914–4923. doi: 10.1128/JB.176.16.4914-4923.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sassanfar M, Roberts JW. Nature of the SOS-inducing signal in Escherichia coli. J Mol Biol. 1990;212:79–96. doi: 10.1016/0022-2836(90)90306-7. [DOI] [PubMed] [Google Scholar]
- 43.Tomasz M. Mitomycin C: small, fast and deadly (but very selective) Chem Biol. 1995;2:575–579. doi: 10.1016/1074-5521(95)90120-5. [DOI] [PubMed] [Google Scholar]
- 44.Lazarevic V, Düsterhöft A, Soldo B, et al. Nucleotide sequence of the Bacillus subtilis temperate bacteriophage SPβc2. Microbiology. 1999;145(Pt 5):1055–1067. doi: 10.1099/13500872-145-5-1055. [DOI] [PubMed] [Google Scholar]
- 45.McLaughlin JR, Wong HC, Ting YE, et al. Control of lysogeny and immunity of Bacillus subtilis temperate bacteriophage SPβ by its d gene. J Bacteriol. 1986;167:952–959. doi: 10.1128/jb.167.3.952-959.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Koo B, Kritikos G, Farelli JD, et al. Construction and analysis of two genome-scale deletion libraries for Bacillus subtilis. Cell Syst. 2017;4:291–305.e7. doi: 10.1016/j.cels.2016.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Au N, Kuester-Schoeck E, Mandava V, et al. Genetic composition of the Bacillus subtilis SOS system. J Bacteriol. 2005;187:7655–7666. doi: 10.1128/JB.187.22.7655-7666.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Miller MC, Resnick JB, Smith BT, Lovett CM. The Bacillus subtilis dinR gene codes for the analogue of Escherichia coli LexA. J Biol Chem. 1996;271:33502–33508. doi: 10.1074/jbc.271.52.33502. [DOI] [PubMed] [Google Scholar]
- 49.Yasbin RE, Fields PI, Andersen BJ. Properties of Bacillus subtilis 168 derivatives freed of their natural prophages. Gene. 1980;12:155–159. doi: 10.1016/0378-1119(80)90026-8. [DOI] [PubMed] [Google Scholar]
- 50.Mendez R, Gutierrez A, Reyes J, Márquez-Magaña L. The extracytoplasmic function sigma factor SigY is important for efficient maintenance of the Spβ prophage that encodes sublancin in Bacillus subtilis. DNA Cell Biol. 2012;31:946–955. doi: 10.1089/dna.2011.1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Abe K, Takahashi T, Sato T. Extreme C-terminal element of SprA serine integrase is a potential component of the “molecular toggle switch” which controls the recombination and its directionality. Mol Microbiol. 2020 doi: 10.1111/mmi.14654. [DOI] [PubMed] [Google Scholar]
- 52.Abe K, Takamatsu T, Sato T. Mechanism of bacterial gene rearrangement: SprA-catalyzed precise DNA recombination and its directionality control by SprB ensure the gene rearrangement and stable expression of spsM during sporulation in Bacillus subtilis. Nucleic Acids Res. 2017;45:6669–6683. doi: 10.1093/nar/gkx466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Abe K, Kawano Y, Iwamoto K, et al. Developmentally-regulated excision of the SPβ Prophage reconstitutes a gene required for spore envelope maturation in Bacillus subtilis. PLoS Genet. 2014;10:e1004636. doi: 10.1371/journal.pgen.1004636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nicolas P, Mäder U, Dervyn E, et al. Condition-dependent transcriptome reveals high-level regulatory architecture in Bacillus subtilis. Science. 2012;335:1103–1106. doi: 10.1126/science.1206848. [DOI] [PubMed] [Google Scholar]
- 55.Suzuki S, Yoshikawa M, Imamura D, et al. Compatibility of site-specific recombination units between mobile genetic elements. iScience. 2020;23:100805. doi: 10.1016/j.isci.2019.100805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Erez Z, Steinberger-Levy I, Shamir M, et al. Communication between viruses guides lysis–lysogeny decisions. Nature. 2017;541:488–493. doi: 10.1038/nature21049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lazarevic V. Ribonucleotide reductase genes of Bacillus prophages: a refuge to introns and intein coding sequences. Nucleic Acids Res. 2001;29:3212–3218. doi: 10.1093/nar/29.15.3212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lazarevic V, Soldo B, Düsterhöft A, et al. Introns and intein coding sequence in the ribonucleotide reductase genes of Bacillus subtilis temperate bacteriophage SPβ. Proc Natl Acad Sci USA. 1998;95:1692–1697. doi: 10.1073/pnas.95.4.1692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lo Piano A, Martínez-Jiménez MI, Zecchi L, Ayora S. Recombination-dependent concatemeric viral DNA replication. Virus Res. 2011;160:1–14. doi: 10.1016/j.virusres.2011.06.009. [DOI] [PubMed] [Google Scholar]
- 60.Jentsch S, Günthert U, Trautner TA. DNA methyltransferases affecting the sequence 5’CCGG. Nucleic Acids Res. 1981;9:2753–2759. doi: 10.1093/nar/9.12.2753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Tran-Betcke A, Behrens B, Noyer-Weidner M, Trautner TA. DNA methyltransferase genes of Bacillus subtilis phages: comparison of their nucleotide sequences. Gene. 1986;42:89–96. doi: 10.1016/0378-1119(86)90153-8. [DOI] [PubMed] [Google Scholar]
- 62.Trautner TA, Pawlek B, Günthert U, et al. Restriction and modification in Bacillus subtilis: identification of a gene in the temperate phage SPβ coding for a BsuR specific modification methyltransferase. Mol Gen Genet MGG. 1980;180:361–367. doi: 10.1007/BF00425849. [DOI] [PubMed] [Google Scholar]
- 63.Spancake GA, Hemphill HE. Deletion mutants of Bacillus subtilis bacteriophage SPβ. J Virol. 1985;55:39–44. doi: 10.1128/JVI.55.1.39-44.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fink PS, Korman RZ, Odebralski JM, Zahler SA. Bacillus subtilis bacteriophage SPβc1 is a deletion mutant of SPβ. Mol Gen Genet MGG. 1981;182:514–515. doi: 10.1007/BF00293946. [DOI] [PubMed] [Google Scholar]
- 65.Wu R, Taylor E. Nucleotide sequence analysis of DNA. II. Complete nucleotide sequence of the cohesive ends of bacteriophage lambda DNA. J Mol Biol. 1971;57:491–511. doi: 10.1016/0022-2836(71)90105-7. [DOI] [PubMed] [Google Scholar]
- 66.Murialdo H. Bacteriophage lambda DNA maturation and packaging. Annu Rev Biochem. 1991;60:125–153. doi: 10.1146/annurev.bi.60.070191.001013. [DOI] [PubMed] [Google Scholar]
- 67.Deichelbohrer I, Messer W, Trautner TA. Genome of Bacillus subtilis bacteriophage SPP1: structure and nucleotide sequence of pac, the origin of DNA packaging. J Virol. 1982;42:83–90. doi: 10.1128/JVI.42.1.83-90.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Oliveira L, Tavares P, Alonso JC. Headful DNA packaging: bacteriophage SPP1 as a model system. Virus Res. 2013;173:247–259. doi: 10.1016/j.virusres.2013.01.021. [DOI] [PubMed] [Google Scholar]
- 69.Fink PS, Zahler SA. Restriction fragment maps of the genome of Bacillus subtilis bacteriophage SPβ. Gene. 1982;19:235–238. doi: 10.1016/0378-1119(82)90012-9. [DOI] [PubMed] [Google Scholar]
- 70.Chai S, Bravo A, Lüder G, et al. Molecular analysis of the Bacillus subtilis bacteriophage SPP1 region encompassing genes 1 to 6. J Mol Biol. 1992;224:87–102. doi: 10.1016/0022-2836(92)90578-8. [DOI] [PubMed] [Google Scholar]
- 71.Forrest D, James K, Yuzenkova Y, Zenkin N. Single-peptide DNA-dependent RNA polymerase homologous to multi-subunit RNA polymerase. Nat Commun. 2017;8:1–8. doi: 10.1038/ncomms15774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Richts B, Hertel R, Potot S, et al. Complete genome sequence of the prototrophic Bacillus subtilis subsp. subtilis strain SP1. Microbiol Resour Announc. 2020 doi: 10.1128/MRA.00825-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Regamey A, Karamata D. The n-acetylmuramoyl-l-alanine amidase encoded by the Bacillus subtilis 168 prophage SPβ. Microbiology. 1998;144:885–893. doi: 10.1099/00221287-144-4-885. [DOI] [PubMed] [Google Scholar]
- 74.Kimura K, Itoh Y. Characterization of poly-γ-glutamate hydrolase encoded by a bacteriophage genome: possible role in phage infection of Bacillus subtilis encapsulated with poly-γ-glutamate. Appl Environ Microbiol. 2003;69:2491–2497. doi: 10.1128/AEM.69.5.2491-2497.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Mamberti S, Prati P, Cremaschi P, et al. γ-PGA hydrolases of phage origin in Bacillus subtilis and other microbial genomes. PLoS ONE. 2015;10:e0130810. doi: 10.1371/journal.pone.0130810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Soldo B, Lazarevic V, Margot P, Karamata D. Sequencing and analysis of the divergon comprising gtaB, the structural gene of UDP-glucose pyrophosphorylase of Bacillus subtilis 168. J Gen Microbiol. 1993;139:3185–3195. doi: 10.1099/00221287-139-12-3185. [DOI] [PubMed] [Google Scholar]
- 77.São-José C, Baptista C, Santos MA. Bacillus subtilis operon encoding a membrane receptor for bacteriophage SPP1. J Bacteriol. 2004;186:8337–8346. doi: 10.1128/JB.186.24.8337-8346.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Baptista C, Santos MA, São-José C. Phage SPP1 reversible adsorption to Bacillus subtilis cell wall teichoic acids accelerates virus recognition of membrane receptor YueB. J Bacteriol. 2008;190:4989–4996. doi: 10.1128/JB.00349-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Willms IM, Hoppert M, Hertel R. Characterization of Bacillus subtilis viruses vB_BsuM-Goe2 and vB_BsuM-Goe3. Viruses. 2017;9:146. doi: 10.3390/v9060146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Fink PS, Zahler SA. The bacteriophages. 2. Oxford: Springer; 2005. Temperate bacteriophages of Bacillus subtilis; pp. 557–571. [Google Scholar]
- 81.Gallego Del Sol F, Penadés JR, Marina A. Deciphering the molecular mechanism underpinning phage arbitrium communication systems. Mol Cell. 2019;74:59–72.e3. doi: 10.1016/j.molcel.2019.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Johnson CM, Harden MM, Grossman AD (2020) An integrative and conjugative element encodes an abortive infection system to protect host cells from predation by a bacteriophage. biorxiv. 10.1101/2020.12.13.422588
- 83.Zahler SA, Korman RZ, Rosenthal R, Hemphill HE. Bacillus subtilis bacteriophage SPβ: localization of the prophage attachment site, and specialized transduction. J Bacteriol. 1977;129:556–558. doi: 10.1128/JB.129.1.556-558.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Fernandes RM, de Lencastre H, Archer LJ. Specialized transduction in Bacillus subtilis by the phages IG1, IG3, and IG4. Arch Virol. 1989;105:137–140. doi: 10.1007/bf0131112410.1007/bf01311124. [DOI] [PubMed] [Google Scholar]
- 85.Rosenthal R, Toye PA, Korman RZ, Zahler SA. The prophage of SPβc2dcitK1, a defective specialized transducing phage of Bacillus subtilis. Genetics. 1979;92:721–739. doi: 10.1093/genetics/92.3.721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Wagner PL, Waldor MK. Bacteriophage control of bacterial virulence. Infect Immun. 2002;70:3985–3993. doi: 10.1128/IAI.70.8.3985-3993.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Cui W, Han L, Suo F, et al. Exploitation of Bacillus subtilis as a robust workhorse for production of heterologous proteins and beyond. World J Microbiol Biotechnol. 2018;34:145. doi: 10.1007/s11274-018-2531-7. [DOI] [PubMed] [Google Scholar]
- 88.Spancake GA, Daignault SD, Hemphill HE. Genome homology and divergence in the SPβ-related bacteriophages of Bacillus subtilis. Can J Microbiol. 1987;33:249–255. doi: 10.1139/m87-042. [DOI] [PubMed] [Google Scholar]
- 89.Spancake GA, Hemphill HE, Fink PS. Genome organization of SPβ c2 bacteriophage carrying the thyP3 gene. J Bacteriol. 1984;157:428–434. doi: 10.1128/JB.157.2.428-434.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Connors MJ, Setlow P. Cloning of a small, acid-soluble spore protein gene from Bacillus subtilis and determination of its complete nucleotide sequence. J Bacteriol. 1985;161:333–339. doi: 10.1128/JB.161.1.333-339.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Frenkiel-Krispin D, Sack R, Englander J, et al. Structure of the DNA-SspC complex: implications for DNA packaging, protection, and repair in bacterial spores. J Bacteriol. 2004;186:3525–3530. doi: 10.1128/JB.186.11.3525-3530.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Tovar-Rojo F, Setlow P. Effects of mutant small, acid-soluble spore proteins from Bacillus subtilis on DNA in vivo and in vitro. J Bacteriol. 1991;173:4827–4835. doi: 10.1128/JB.173.15.4827-4835.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Nicholson WL, Sun DX, Setlow B, Setlow P. Promoter specificity of σG-containing RNA polymerase from sporulating cells of Bacillus subtilis: identification of a group of forespore-specific promoters. J Bacteriol. 1989;171:2708–2718. doi: 10.1128/JB.171.5.2708-2718.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Hemphill HE, Gage I, Zahler SA, Korman RZ. Prophage-mediated production of a bacteriocinlike substance by SPβ lysogens of Bacillus subtilis. Can J Microbiol. 1980;26:1328–1333. doi: 10.1139/m80-220. [DOI] [PubMed] [Google Scholar]
- 95.Dragoš A, Andersen AJC, Lozano-Andrade CN, et al (2020) Phages weaponize their bacteria with biosynthetic gene clusters. bioRxiv. 10.1101/2020.10.01.322628
- 96.Paik SH, Chakicherla A, Hansen JN. Identification and characterization of the structural and transporter genes for, and the chemical and biological properties of, Sublancin 168, a novel lantibiotic produced by Bacillus subtilis 168. J Biol Chem. 1998;273:23134–23142. doi: 10.1074/jbc.273.36.23134. [DOI] [PubMed] [Google Scholar]
- 97.Oman TJ, Boettcher JM, Wang H, et al. Sublancin is not a lantibiotic but an S-linked glycopeptide HHS public access author manuscript. Nat Chem Biol. 2011;7:78–80. doi: 10.1038/nchembio.509.Sublancin. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Dorenbos R, Stein T, Kabel J, et al. Thiol-disulfide oxidoreductases are essential for the production of the lantibiotic sublancin 168. J Biol Chem. 2002;277:16682–16688. doi: 10.1074/jbc.M201158200. [DOI] [PubMed] [Google Scholar]
- 99.Kouwen TRHM, Van Der Goot A, Dorenbos R, et al. Thiol-disulphide oxidoreductase modules in the low-GC Gram-positive bacteria. Mol Microbiol. 2007;64:984–999. doi: 10.1111/j.1365-2958.2007.05707.x. [DOI] [PubMed] [Google Scholar]
- 100.Wang Q, Zeng X, Wang S, et al. The bacteriocin sublancin attenuates intestinal injury in young mice infected with Staphylococcus aureus. Anat Rec. 2014;297:1454–1461. doi: 10.1002/ar.22941. [DOI] [PubMed] [Google Scholar]
- 101.Wu C, Biswas S, Garcia De Gonzalo CV, Van Der Donk WA. Investigations into the mechanism of action of sublancin. ACS Infect Dis. 2019;5:454–459. doi: 10.1021/acsinfecdis.8b00320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Dubois J-YF, Kouwen TRHM, Schurich AKC, et al. Immunity to the bacteriocin sublancin 168 is determined by the SunI (YolF) protein of Bacillus subtilis. Antimicrob Agents Chemother. 2009;53:651–661. doi: 10.1128/AAC.01189-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Denham EL, Piersma S, Rinket M, et al. Differential expression of a prophage-encoded glycocin and its immunity protein suggests a mutualistic strategy of a phage and its host. Sci Rep. 2019;9:2845. doi: 10.1038/s41598-019-39169-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Klimecka MM, Chruszcz M, Font J, et al. Structural analysis of a putative aminoglycoside N-Acetyltransferase from Bacillus anthracis. J Mol Biol. 2011;410:411–423. doi: 10.1016/j.jmb.2011.04.076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Galimand M, Fishovitz J, Lambert T, et al. AAC(3)-XI, a new aminoglycoside 3-N-acetyltransferase from Corynebacterium striatum. Antimicrob Agents Chemother. 2015;59:5647–5653. doi: 10.1128/AAC.01203-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Cooper TF, Heinemann JA. Postsegregational killing does not increase plasmid stability but acts to mediate the exclusion of competing plasmids. Proc Natl Acad Sci USA. 2000;97:12643–12648. doi: 10.1073/pnas.220077897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Brantl S, Müller P. Toxin-antitoxin systems in Bacillus subtilis. Toxins (Basel) 2019;11:262. doi: 10.3390/toxins11050262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Brantl S. Bacterial type I toxin-antitoxin systems. RNA Biol. 2012;9:1488–1490. doi: 10.4161/rna.23045. [DOI] [PubMed] [Google Scholar]
- 109.Jahn N, Brantl S. One antitoxin—two functions: SR4 controls toxin mRNA decay and translation. Nucleic Acids Res. 2013;41:9870–9880. doi: 10.1093/nar/gkt735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Reif C, Löser C, Brantl S. Bacillus subtilis Type I antitoxin SR6 promotes degradation of toxin yonT mRNA and is required to prevent Toxic yoyJ overexpression. Toxins (Basel) 2018;10:74. doi: 10.3390/toxins10020074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Holberger LE, Garza-Sánchez F, Lamoureux J, et al. A novel family of toxin/antitoxin proteins in Bacillus species. FEBS Lett. 2012;586:132–136. doi: 10.1016/j.febslet.2011.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Van Melderen L. Toxin–antitoxin systems: why so many, what for? Curr Opin Microbiol. 2010;13:781–785. doi: 10.1016/j.mib.2010.10.006. [DOI] [PubMed] [Google Scholar]