

Abstract
Lesions in the Guillain–Mollaret (G–M) triangle frequently cause various types of tremors or tremor-like movements. Nevertheless, we know relatively little about their generation mechanisms. The deep cerebellar nuclei (DCN), which is a primary node of the triangle, has two main output paths: the primary excitatory path to the thalamus, the red nucleus (RN), and other brain stem nuclei, and the secondary inhibitory path to the inferior olive (IO). The inhibitory path contributes to the dentato-olivo-cerebellar loop (the short loop), while the excitatory path contributes to the cerebrocerebellar loop (the long loop). We propose a novel hypothesis: each loop contributes to physiologically distinct type of tremors or tremor-like movements. One type of irregular tremor-like movement is caused by a lesion in the cerebrocerebellar loop, which includes the primary path. A lesion in this loop affects the cerebellar forward model and deteriorates its accuracy of prediction and compensation of the feedback delay, resulting in irregular instability of voluntary motor control, i.e., cerebellar ataxia (CA). Therefore, this type of tremor, such as kinetic tremor, is usually associated with other symptoms of CA such as dysmetria. We call this type of tremor forward model-related tremor. The second type of regular tremor appears to be correlated with synchronized oscillation of IO neurons due, at least in animal models, to reduced degrees of freedom in IO activities. The regular burst activity of IO neurons is precisely transmitted along the cerebellocerebral path to the motor cortex before inducing rhythmical reciprocal activities of agonists and antagonists, i.e., tremor. We call this type of tremor IO-oscillation-related tremor. Although this type of regular tremor does not necessarily accompany ataxia, the aberrant IO activities (i.e., aberrant CS activities) may induce secondary maladaptation of cerebellar forward models through aberrant patterns of long-term depression (LTD) and/or long-term potentiation (LTP) of the cerebellar circuitry. Although our hypothesis does not cover all tremors or tremor-like movement disorders, our approach integrates the latest theories of cerebellar physiology and provides explanations how various lesions in or around the G–M triangle results in tremors or tremor-like movements. We propose that tremor results from errors in predictions carried out by the cerebellar circuitry.
Keywords: tremor, cerebellum, Guillain–Mollaret triangle, predictions, forward model
Introduction
The deep cerebellar nuclei (DCN) represent a primary node of the so-called Guillain–Mollaret (G–M) triangle, an anatomical circuit known to play a major role in tremor genesis both in animal models and in human disorders affecting the posterior fossa (1).
Deep cerebellar nuclei have two main output paths: the primary excitatory path to the thalamus, the red nucleus (RN), and other brain stem nuclei, and the secondary inhibitory path to the inferior olive (IO). The inhibitory path contributes to the dentato-olivo-cerebellar loop (we call it the short loop), while the excitatory path contributes to the cerebrocerebellar loop (we call it the long loop).
We propose a hypothesis according to which each loop contributes to physiologically distinct type of tremors or tremor-like movements. One type of irregular tremor-like movement is caused by a lesion in the cerebrocerebellar loop, which includes the primary path. The second type of regular tremor is correlated with synchronized oscillation of IO neurons due to reduced degrees of freedom in IO activities.
Section I. Phenomenology of Cerebellar Tremors
Cerebellar tremors include diverse phenotypes (2). However, Louis (3) pointed out that, nowadays, “cerebellar tremor is equated exclusively with intention tremor” in an “oversimplified manner” (4). Besides, pathomechanisms underlying essential tremor (ET) have been a focus of debate (2), and thereby roles of IO have likely been overstressed in tremor pathogenesis (5). Due to such a simplification, seminal works by Gordon Holmes appear to be underestimated. This section aims to provide a brief overview of the historical backgrounds and phenomenology of cerebellar tremors.
Kinetic Tremor and Static Tremor in Holmes' Classic Study
Studies of human cerebellar tremors originate from Holmes' works who carefully examined tremor phenomenology in patients with spatially confined lesions in the cerebellum and described two types of tremors, namely, kinetic tremor and static tremor, in the Croonian lectures given in June 1922 (Table 1). Their clinical phenotypes appear different from those we imagine now from the terminology of kinetic or static. Thus, we cite his original descriptions to elucidate their phenomenology (6, 7). One can read his classic papers in an article of Cerebellar Classic (8). It should be acknowledged that these two types of tremors occur concomitantly with deterioration of coordination.
Table 1.
Clinical features of various forms of tremors described by Holmes: summary of Holmes' Croonian lectures given in June 1922.
| Holmes' description | Static/kinetic | Regularity | Target oriented | Reciprocal muscle activities | Contribution of visual feedback | Special features | |
|---|---|---|---|---|---|---|---|
| (1) | Kinetic tremor during motion | Kinetic | Irregular (especially during slow movement) | Yes | Not typical | Yes | Prominent when superior peduncles are damaged. ramya Proximal > distal |
| (2) | Intention tremor | Kinetic | Irregular | Yes | ? | Yes | Tremor associated with disseminated sclerosis. ramya Less sharp than kinetic tremor. |
| (3) | Static tremor/Gravitational irregular tremor | Static/postural | Irregular | ? | No | Yes | Prominent in the extension of both upper limbs. ramya Contribution of fatigue. ramya Proximal joints. |
| (4) | Static (postural)/Regular oscillatory tremor with reciprocal activities of agonists and antagonists | Static/postural | Regular | Yes | Yes | Yes | Prominent in precise maintenance of the limbs accurately in certain positions. ramya “Terminal tremor”-like tremor. ramya PD rest tremor-like tremor. ramya Easily induced in co-contraction of agonist/antagonist. |
Kinetic Tremor in Holmes' Classic Study
Holmes described the tremor during active movement (attempts to bring finger from nose to three points in succession or attempts to touch a series of points alternatively) as follows: “At the commencement of the movement the finger or toe may sway from side to side, or the movement may be broken and jerky, especially when performed slowly (Table 1) (1). There is little irregularity as a rule during its course, but in slow and deliberate movements the rate is irregular or discontinuous, or the finger may swing in any plane from the correct line [page 151 in a reference of Cerebellar Classic (8)].” In addition to the irregular and discontinuous sways, he emphasized the association of two additional features. First, terminal tremor (irregular terminal jerks) occurs, associated with hypermetric and hypometric, for example, “in the former case the finger that has shot past its mark is brought back too far and sways or oscillates about its aim until it touches it; in the latter the limb which is arrested before it has reached it is advanced by a series of irregular jerks” (page 151). Second, continuous sways occur at the target. He described that “Even when the finger comes in contact with the patient's nose or other object it may continue to sway from side to side or in the direction of previous movement owing to inability to maintain the attitude steadily” (page 151).
Notably, this kinetic tremor “was less prominent” in most of his cases with local lesions of the cerebellum than in patients with “the primary atrophies (Table 1) (2).” In other words, this type of tremor is prominent in degenerative cerebellar ataxia (CA), suggesting that its developments might be dependent on cerebellar residual functions. Indeed, Holmes hypothesized kinetic tremor, or tremor during active movement, as follows: tremor “naturally results from the irregularities in the rate of muscle contractions, but errors in the range and direction of movement, necessitating correction, are also factors.”
Static Tremor in Holmes' Classic Study
Holmes described two types of static tremor.
The first subtype has irregular nature (Table 1) (3). Holmes observed this tremor when his patients extended both upper limbs. He described that “Its oscillations are mostly in the line of gravity, and can be seen on careful inspection to be due to a failure in the tonic contractions of the muscles that maintain the attitude, with the result that the limb falls with gravity and is replaced by voluntary efforts” (page 146). It should be noted that the maintenance of the attitude is a highly voluntary process.
The other subtype is characterized by regular oscillations (Table 1) (4). Holmes described conditions in which this tremor preferably occurs: “Another type of tremor, characterized by more regular oscillations of a limb or some of its segments, occurs when the patient attempts to maintain the limb accurately in certain positions, or in postures necessary for the performance of some act” (page 146). Moreover, “It is usually only in attitudes determined by the tonic contractions of opposing groups of muscles that this regular form of tremor develops” (page 146). The lesions of the regular static tremor were ascribed to “the superior peduncles” and “mid-brain lesions that involve these peduncles.”
In summary, the latter type of regular static tremor appears to occur during co-contraction of agonist and antagonist muscles, while the former type of irregular static tremor appears to occur during reciprocal muscle activities for feedback control.
After Holmes
For instance, in the “Handbook of Clinical Neurology” published in 1969, Garcin attributed features of kinetic tremor and irregular static tremor in Holmes' classic study to “disturbed continuity of movement” (9). In this regard, he described more clearly features of irregular static tremor as follows: “the static effort of maintaining posture in fact produces tremors to the same extent as does movement,” and “The tremor is more marked when more motor segments are involved, and this explains the difficulty of maintaining immobility in standing or in keeping the arms widely extended” (page 327). He stressed that irregular static tremor is mostly observed in the initial static phase. For example, he described that “at the moment when a hand grasps the glass: when the first clumsy movement is over there may be a few oscillations of pronation and supination occurs, but the patient can grasp the glass without jerking.” This classification of cerebellar tremor by Holmes appears to be used until the 1970s. However, distinction of these two types of tremors are getting rarer in recent textbooks and review articles (2).
Intention Tremor
Intention tremor was first described by Jean-Martin Charcot. In a well-documented lecture on multiple sclerosis (MS) delivered in 1868, he described the presence of CA in patients with MS, now known as the Charcot's triad (intention tremor, scanning speech, and nystagmus) (10).
A consensus statement of the Movement Disorder Society characterizes features of intention tremor as “amplitude increases during visually guided movements toward a target at the termination of the movement” (11) or “a crescendo increase in tremor occurs as the affected body part approaches its visual target” (12). Furthermore, intention tremor is exaggerated in a visually guided target pursuit task but diminished in a memory-guided task (11, 13). In order to emphasize these pathophysiological features, a term of tremor during target-directed movements has been utilized. Thus, intention tremor can be observed in the finger-to-nose maneuver, which requires precise feedback control. Its frequency is mainly <5 Hz, and “the possibility of a position-specific tremor or a postural tremor produced at the beginning or end of a movement is excluded” (11). Rest tremor is commonly not identified (14). There is a consensus that that intention tremor is caused by a lesion in the cerebellothalamic pathway (12, 14, 15). The lesions are usually in the brainstem in the vicinity of the RN (16) or the posterior thalamus (17, 18). Therefore, another term of cerebellar outflow tremor has also been introduced to stress the neural structure for the genesis of intention tremor (19). In contrast, focal lesions in the cerebellar cortex alone usually do not cause this tremor (20).
In Holmes' classic papers, he described both of his kinetic tremor and regular static tremor occurred in patients with lesions in the superior cerebellar peduncles, suggesting that intention tremor has common features with these Holmes' tremors. Notably, there is a description that “In the tremor that is a prominent feature when the superior peduncles are damaged, the deviations are more abrupt and are terminated more suddenly” (page 151).
Late-Onset Cerebellar Tremors: “Holmes' Tremor” and Palatal Tremor
The onset of cerebellar tremor after a stroke is diverse, ranging from the day of a stroke to a few years later (14). The above kinetic and static tremors in Holmes' classic study and intention tremor seem to be present in the acute phase. However, the two types of cerebellar tremors also occur characteristically with some delay after the onset of pathologies (21): “Holmes' tremor” and palatal tremor.
“Holmes' Tremor”
“Holmes' tremor,” as a modern term, is a rare tremor characterized by the following three features: (1) a concomitant expression of rest tremor and intention tremor, involving the proximal and distal part of the upper limbs with large amplitudes, usually associated with postural tremor; (2) slow frequency, usually <4.5 Hz; and (4) in a case when the preceding lesion (e.g., strokes) is identified, a variable delay (usually 4 weeks−2 years) (11, 14). This unique tremor was previously labeled under rubral tremor or midbrain tremor. However, this tremor is also induced by lesions outside these classic locations. For example, one study of three patients with “Holmes' tremor” following stroke showed that the lesions were located in the superior cerebellar peduncle, midbrain tegmentum, and posterior thalamus (22). To avoid topographic names, therefore, “Holmes' tremor” is now used in honor of his first description (11). Holmes' tremor is frequently accompanied by hypertrophy of the inferior olive nucleus (ION) (23).
Palatal Tremor
Palatal tremor is characterized by slow, rhythmic movements of the soft palate (usually, at a frequency of 1–3 Hz) and sometimes of other muscles in the pharynx, larynx, lower face, and trunk (24, 25). Palatal tremor comprises idiopathic and symptomatic types. The causes of symptomatic palatal tremor include stroke, trauma, MS, Behçet's disease, and encephalitis (24), and the most common causes are strokes in the brainstem and the cerebellum (24). The symptomatic palatal tremor usually develops some time (1–49 weeks) after the lesion onset (26), which is associated with cerebellar symptoms (25) and hypertrophy of ION (24, 25).
Taken together, the late-onset nature and the associated ION hypertrophy suggest underlying secondary and compensatory pathological mechanisms in “Holmes' tremor” and palatal tremor (21). The hypertrophy of ION is usually observed as a high signal on T2- or proton density-weighted MR image with the enlargement (24, 25).
Essential Tremor
According to a consensus statement of the Movement Disorder Society, ET is defined by bilateral, largely symmetric postural, or kinetic tremor, at the frequency of 4–12 Hz, involving hands and forearm, with or without head tremor and tremor in other locations (11, 12). The primary clinical phenotype is the postural tremor of the hands (11). The tremor generally persists, although the amplitude fluctuates (11). The tremor may or may not produce disability (11); however, ET is progressive in nature (27). Concomitant manifestation of intention tremor and rest tremor is observed in 50 and 20% of the patients, respectively (27). Due to the heterogeneity, it is proposed that ET comprises a family of diseases rather than a single entity (27). In other words, ET is overlapping clinical phenotypes.
In the 2018 statement, the notion of ET plus was introduced to include patients with neurological signs of uncertain relationship to tremor (i.e., “soft neurological signs”). Notably, soft neurological signs include cerebellar symptoms such as a mild degree of ataxic gait, oculomotor deficits, and impaired motor timing (27). Due to the clinical heterogeneity, Louis et al. (28) proposed that ET may represent a family of diseases rather than a single clinical–pathological entity (28). Our current understanding of the mechanisms behind ET has evolved quickly, thanks to the works of Louis' group with the elucidation that cerebellar cortex shows abnormal features in postmortem material (29). These authors have shown abnormalities in Purkinje cells (PCs: axonal swellings, swellings in and regression of the PC dendritic arbor, and PC death), basket cells, and climbing fibers in individuals with ET.
In conclusion, cerebellar tremors gather various phenotypes (Table 2). Two clinical features will be summarized:
Table 2.
Phenotypes in cerebellar tremors.
| Type of cerebellar tremor | Phenomenology | Responsible region |
|---|---|---|
| Kinetic tremor in Holmes' classic study | • Irregular and discontinuous sways • Sometimes marked at the beginning of the movement |
The cerebellum (probably destruction of the cerebellar cortex and/or the white matter) |
| Static tremor in Holmes' classic study | • Subtype 1: Irregular oscillation in the extension of upper limbs during the maintenance of the limb against gravity • Subtype II: Regular oscillations of a limb or some of its segments during maintenance of the limb accurately in certain positions |
The cerebellum (probably destruction of the cerebellar cortex and/or the white matter) |
| Intention tremor | • Amplitude increase during visually guided movements toward a target at the movement termination | The dentato-rubro-thalamic tract |
| “Holmes' tremor” | • Concomitant expression of rest tremor* and intention tremor with/without postural tremor* • Slow frequency, usually <4.5 Hz • Late onset of pathologies |
Superior peduncle, midbrain tegmentum, and posterior thalamus |
| Palatal tremor | • Rhythmic movements of the soft palate • Late onset of pathologies |
The brainstem and the cerebellum |
| Essential tremor | • Bilateral, largely symmetric postural tremor or kinetic tremor* • Involving hands and forearm, with or without head tremor and tremor in other locations |
Cerebellar cortex |
Kinetic tremor, tremor occurring during any voluntary movement; postural tremor, tremor present while voluntarily maintaining a position against gravity; rest tremor, tremor that occurs in a body part that is not voluntarily activated and is completely supported against gravity.
Definition by Consensus Statement of the Movement Disorder Society on Tremor (11).
Tremor is generally defined as the “involuntary, rhythmic, oscillatory movement of a body part” (11, 12). However, the irregularity in cycle and amplitude is evident in kinetic tremor and irregular static tremor in Holmes' classic study, and sometimes in intention tremor, compared with other types of cerebellar tremors.
In the condition of “Holmes' tremor” and ET, plural pathophysiological mechanisms appear to contribute to their phenotypes of tremor either concomitantly or with the lapse of time.
From physiological and control engineering points of view, difference in regularity and voluntariness strongly suggests contribution of distinct control mechanisms. In addition, difference in onset also suggests distinct pathomechanisms to be factored in. Overall, the three factors, i.e., regularity, voluntariness, and onset, may be key clues for understanding pathophysiology of diverse cerebellar tremors. We will address this issue in section Physiological Backgrounds of Two Types of Cerebellar Tremors.
Section II. Physiological Backgrounds of Two types of Cerebellar Tremors
In the previous section, we traced the historical backgrounds and phenomenology of cerebellar tremors as far back as the original descriptions by Holmes (6, 7). We realized that various phenotypes of “cerebellar tremors” may contain two distinct conditions: involuntary regular tremors and voluntary irregular tremors (or more precisely, tremor-like movements), and each condition may be related to distinct pathology of distinct neuron circuitries. In this section, we will address the two tremor generation mechanisms based on recent physiological, morphological, and clinical findings.
Two Loop Circuitries in the Dentato-Rubro-Olivary (Guillain–Mollaret) Triangle and Their Functions
It has long been established that patients with lesions in or in the vicinity of the G–M triangle (Figure 1) frequently show various types of tremors or tremor-like movements (14). Previous studies established that the G–M triangle contains two distinct loop circuitries: (1) the dentato-olivo-cerebellar loop (we call it the short loop, Figure 1) and (2) the cerebrocerebellar loop (we call it the long loop, Figure 1).
Figure 1.
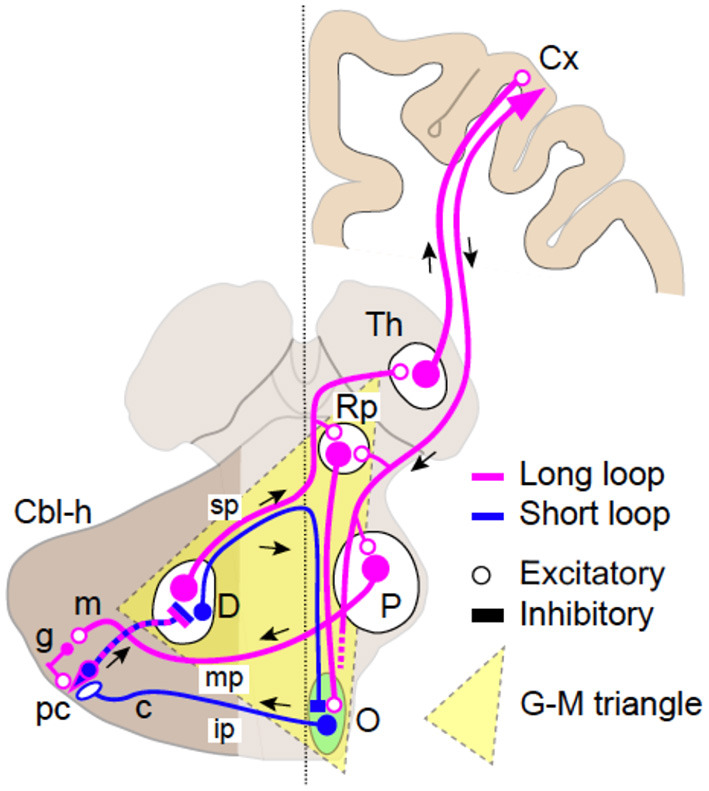
Schematics of the two loop circuits in the Guillain–Mollaret triangle. The dentato-olivo-cerebellar loop (short loop, blue) and the cerebrocerebellar loop (long loop, magenta). Smaller GABAergic (inhibitory) cells in the dentate nucleus (D) pass through the superior cerebellar peduncle (SCP) (sp), cross the midline, and project directly to the contralateral inferior olivary nucleus (O). Efferent fibers from IO pass through the inferior cerebellar peduncle (ip) and project to Purkinje cells (PC, pc) in the contralateral cerebellar hemisphere (Cbl-h). PCs then project to DN cells to close the loop. The long loop is almost identical to the cerebrocerebellar loop. Larger excitatory DN cells pass through (SCP, sp), cross the midline, and project to the contralateral parvocellular red nucleus (RNp), and the thalamus (Th) with collaterals. Thalamocortical neurons relay the cerebellar inputs to various cortical areas (Cx). The return path to the cerebellum is the cortico-ponto-cerebellar tract, which originates from a various parts of the cerebral cortex. The corticofugal axons project directly to the PN (P) and finally arrive at the contralateral cerebellar hemisphere (Cbl-h) as mossy fibers (MFs) via the middle cerebellar peduncle (mp) to close the loop.
Anatomy of the Long Loop
The long loop is almost identical to the cerebrocerebellar loop (30–32). Larger excitatory dentate nucleus (DN) cells, after passing through SCP (Figure 1, sp) and crossing the midline, project to the contralateral RNp and the thalamus (Figure 1, Th) with collaterals. Thalamocortical neurons relays the cerebellar inputs to various cortical areas (Figure 1, Cx). The return path to the cerebellum is the cortico-ponto-cerebellar tract, which originates from various parts of the cerebral cortex (30–32). The corticofugal axons project directly to the pontine nuclei (PN, Figure 1, P) and finally arrive at the contralateral cerebellar hemisphere (Figure 1, Cbl-h) as mossy fibers (MFs) via the middle cerebellar peduncle (Figure 1, mp) to close the loop (30, 31).
Physiological Operation of the Short Loop
The DN contains two distinct types of neurons. Larger excitatory neurons project to the parvocellular part of the red nucleus (RNp) and the thalamus (Th), while smaller inhibitory (GABAergic) neurons project directly to the IO to inhibit IO neurons (33). The GABAergic terminals in IO are concentrated around gap junctions between the IO neurons (34) and reduces their conductance, thereby reducing synchronous activities of the IO neurons (35). On the other hand, IO neurons also receive excitatory inputs from PNp (35–37). The excitatory terminals are concentrated around the gap junctions and are presumed to facilitate synchronous activities of the IO neurons (34, 35). In summary, the IO neurons receive two distinct types of inputs; one facilitates, and the other suppresses synchronous activities of IO neurons.
A Putative Servo-Like Mechanism to Limit the Synchrony of IO Neurons
In physiological conditions, the inhibitory input from DN and the excitatory input from RNp to the gap junctions between the IO neurons appear to be balanced. For instance, when DN cells get more active, the direct inhibition from DN to IO increases, while the disynaptic excitatory input from RNp to IO also increases concomitantly. In contrast, when DN cells get inhibited, the direct inhibition from DN to IO decreases (i.e., disinhibition), while the disynaptic excitation from RNp to IO decreases concomitantly. In summary, regardless of the alteration of output from DN, modulations of inhibitory and excitatory inputs to IO appear to cancel each other. Overall, the synchrony between IO neurons appears to be limited within a certain range in physiological conditions with this servo-like mechanism.
Physiological Operation of the Long Loop: the Cerebrocerebellum as a Site of Forward Models
One critical problem in biological motor control is that afferent sensory signals have inevitable temporal delays in reaching the central nervous system. In other words, the brain always observes “the past” of its own body and environments. A visual signal, for instance, arrive at the primary visual cortex about 30 ms later and at the parietal cortex about 80 ms later than an onset of the signal (38). Among the factors contributing to the feedback delay, such as a synaptic delay or an electro-mechanical delay, the dominant factor is the nerve conduction delay, ranging about 10 ms for a shrew to about 100 ms for an elephant. Sensory delays are comparable to typical time scales of rapid movements and hence not negligible.
The delay in sensory feedback is problematic not only in sensing the body and the environments but also in controlling the body. It is well known in control engineering that feedback control based on a previous state causes oscillatory and unstable movements if the delay in feedback control is of the order of or larger than a time constant of a controlled plant (39). The delays in visual feedback are comparable to the movement time of rapid reaching movement of the upper limb (about a few hundred milliseconds) and of saccadic eye movements (typically <50 ms). Therefore, in biological motor control, feedback control based on delayed sensory signals would result in unstable movements. Nonetheless, animals can perform a fast movement without losing its stability. Biological motor control must be equipped with a mechanism to compensate the sensory delay for a fast and stable movement.
One mechanism proposed to cope with the delay in sensory feedback is to compute a future state of the body based on a current estimate of the body and an efferent signal of motor control. This predictive computation internally emulates or models an actual movement of the body by essentially solving an equation of motion of the body forward in time, thereby known as an internal forward model (40, 41). An internal forward model predicts the state of the body time by time that is then used by a feedback controller, thereby allowing fast and stable movements. The feedback control based on the prediction of internal forward model is called internal feedback. There are lines of evidence supporting the hypothesis of predictive forward model and internal feedback from neuroimaging studies (42, 43), non-invasive stimulation studies (44, 45), and psychophysics studies (46–48) in human.
Previous studies repeatedly suggested the cerebrocerebellum as a potential site of the forward model based on neuroanatomical data and clinical observations [e.g., (39, 49–52)]. A forward model requires two distinct inputs: (a) a set of sensory feedback signals, which are necessary to update the forward model and (b) the copy of descending motor commands. The two inputs are integrated in the forward model to generate the state estimate. In fact, the cerebellum receives both of these inputs. It receives inputs from cortical motor areas via the PN (53, 54), and these inputs represent the efference copy of descending motor commands (55–57). The cerebellum also receives somatosensory inputs directly from the ascending spinocerebellar tracts and indirectly via brain stem nuclei, such as the cuneate nucleus or the lateral reticular nucleus. These sensory inputs may provide an update on the state to be estimated. The above argument may appear to support the cerebellar forward model hypothesis. However, in reality, it is on insufficient grounds because the two lines of inputs are primarily separate in the cerebellar cortex. The MF inputs from the cortical motor areas (via PN) distribute mainly in the hemispheric (i.e., lateral) part (58), while the sensory MF inputs from the spinal cord or the brain stem nuclei distribute in more rostral and medial part (the anterior lobe and the intermediate zone) [e.g., (59)] of the cerebellar cortex. Therefore, one may expect a convergence of the two MF inputs only in a minor part of the intermediate zone. Unfortunately, the simple summation of the two MF inputs is not consistent with their asymmetric roles in the forward model. The efference copy plays an essential role in a state prediction, while the sensory input plays a critical role in an update of the prediction, as will be discussed later.
As for the output from a forward model, we expect it to correlate with the future state of the motor apparatus (39). In principle, we should examine the output from the cerebrocerebellum in the DN because it is the sole output node from the cerebrocerebellum. Nevertheless, previous studies tried to address this issue by analyzing the PC activities. Note that the PCs' activity represents an intermediate representation of the cerebellar circuitry and is not suitable for characterizing the output of a forward model. In this regard, few studies are eligible to discuss the output of the cerebellar forward models (60–62).
System Identification of the Transformation in the Cerebrocerebellum—Its Similarity to Kalman Filter
If the cerebrocerebellum functions as a forward model, it is expected that the current output from DN should contain predictive information about the future MF input. Therefore, in our previous study (63, 64), we examined the relationship between activities of MFs (cerebellar inputs), PCs (intermediate representation), and DNCs (cerebellar outputs) (Figure 3). Briefly, we demonstrated that the activities of individual PCs were reconstructed precisely as a weighted sum of those of MFs. Similarly, the activities of individual DNCs were reconstructed strictly as a weighted sum of those of PCs and MFs. We further proved that the activities of DNCs contained predictive information about future MF inputs (63, 64). Namely, the output from the cerebrocerebellum is capable of predicting 200 ms into the future to compensate for the delay of sensory feedback. We finally note that the linear relationship between MF, PC, and DNC activities resemble an optimal linear estimator known as the Kalman filter [(63–65)].
The functional similarity of the cerebellum to the Kalman filter has already been suggested in some previous studies. Most notably, Paulin (66, 67) indicated that the cerebellum could be a neural analog of a Kalman filter. Droulez and Cornílleau-Pérèz (68) drew attention to the relevance of multisensory integration in the moving organism to the Kalman filter. Nevertheless, the suggested analogy was only at the functional level and totally lacked correspondence to the cerebellar network. In our study, we demonstrated the three computational steps in the cerebellar circuit that are compatible with the Kalman filter (63, 64) (Figure 2): (1) the PCs compute a predictive state from a current estimate conveyed by the MFs (prediction step); (2) the DNCs combine the predicted state from the PCs and sensory feedback from the MFs (Filtering step); and (4) the DNCs represent future activities of MFs (cerebellar prediction).
Figure 2.

Equivalence of the cerebrocerebellar circuitry to a Kalman filter [reproduced with permission from Tanaka et al. (63)]. Schematic of the Kalman filter model of the cerebrocerebellum overlaid on the cerebellar circuit. MF, mossy fiber (red); PC, Purkinje cell (green); DC, dentate cell (light blue). Granule cells (orange) and inhibitory interneurons (blue) that are not analyzed in this work are included to show the basic structure of the cerebellar neuron circuitry. Three stages of linear computation obtained in our analysis are accompanied with the three types of computation of Kalman filter explained in the text. Reproduced from Tanaka et al. (63) under CC-BY license.
Overall, the cerebellum appears to perform not only an internal-forward-model prediction but also an optimal integration of a predicted state and sensory feedback signals, in a way that is equivalent to Kalman filter as demonstrated in Tanaka et al. (63, 64) (Figure 2).
Interaction Between the Two Loops
It should be noted that the two loops are not independent to each other as clearly depicted in Figure 1. First, they share the same PCs in the hemispheric part of the cerebellar cortex. Second, the long loop has a side path to modulate activities of IO cells through RNp. Therefore, the two loops are interactive and dependent to each other. An unstable loop may therefore impact on the physiological behavior of the second loop. Abnormal discharges may emerge from altered PCs (see the example of ET), and this will impact on both loops.
Generation of Two Types of Tremors
We underline that both loops are designed to avoid tremor or instability as described above. Indeed, the short loop has a neural mechanism to avoid synchronous discharges of IO neurons, while the long loop has evolved to function as a forward model to avoid instability of control. Nevertheless, in pathological conditions, each safety mechanism fails, resulting in the generation of a characteristic type of tremor.
Failure of the Short Loop Results in Regular Oscillatory Tremors
As reviewed in section Phenomenology of cerebellar tremors, the modern definition of the term “tremor” is “the involuntary, rhythmic, oscillatory movement of a body part” (11, 12). Naturally, a number of previous studies, both basic and clinical, addressed the location of the oscillator. There is a consensus that IO plays an essential role in the generation of the regular tremors (35, 70, 71). For instance, harmaline-induced tremor in rodents has been extensively used as an animal model for ET. Cheng et al. (72) made a subcutaneous injection of harmaline hydrochloride (20 mg/kg) in mice and then videotaped the responses. Regular action and postural tremors in the mouse began no more than 5 min after harmaline injection and peaked at approximately 30 min. The forelimb tremor was postural or action tremor, similar to that observed in ET. In these model animals, a large population of IO neurons appear to discharge in synchrony and rhythmically (73–75), thereby inducing synchronized complex spikes (CSs) of Purkinje cells. Then, the synchronized CSs ignite synchronized rebound excitation of DN cells (71, 76), and the cerebellar output finally induces, through the thalamocortical pathway, rhythmical and reciprocal discharges of agonists and antagonists muscles, i.e., tremor. As described in Physiological operation of the short loop, there is a mechanism to avoid synchronous discharges of IO neurons in physiological conditions. Nevertheless, in pathological conditions and for specific posture and/or movement, IO neurons are somehow switched into a synchronization mode to induce rhythmical discharges, resulting in regular tremors. We infer that involuntary and regular tremors, such as static tremor described by Holmes (6, 7), rest tremor and postural tremor of “Holmes' tremor,” and ET, are likely to depend on the same mechanism described above. We also infer that “Holmes' tremor” and palatal tremor depend on the same mechanism, although the efferent pathway of the palatal tremor appears to spare the Vim nucleus of the thalamus because Vim thalamotomy is ineffective to palatal tremor, while it is effective to “Holmes' tremor” (77).
Generation of Irregular Tremor-Like Movement and Its Relevance to the Forward Model Hypothesis of the Cerebellum
Not all tremors or tremor-like movements are regular or oscillatory (see section Phenomenology of Cerebellar Tremors) as noted by Holmes himself (6, 7). The irregularity in cycle and amplitude is crucial because it strongly suggests different generation mechanisms from that of the regular tremors described above. Moreover, it should be noted that the irregularity appears during voluntary movement, as exemplified in their names “kinetic” or “intention.” Here, we explain the irregularity (i.e., kinetic tremor in Holmes' classic study and intention tremor) as malfunction of the cerebellar forward model.
In our previous study (69), we demonstrated a clinical evidence that supported the cerebellar forward model hypothesis [e.g., (44, 51)]. A series of studies from our group confirmed the impaired predictive control in movements of patients with degenerative CA. We first decomposed the muscle activities for the wrist movement into a low-frequency ( ≤ 0.5 Hz) component (F1) and a high-frequency (>0.5 Hz) component (F2), each of which represented the predictive control and the feedback correction, respectively (69). Then, for each component, we identified a recipe of muscle activities by analyzing a relationship between the muscle tension and movement kinematics [the wrist angle θ(t) and the wrist angular velocity ] weighted by the coefficients of Kr (the elastic term) and Br (the viscous term) (69, 78–80). Importantly, the ratio of Br/Kr characterized the recipe of muscle activities for the predictive and corrective components. In control subjects, the Br/Kr ratio for the predictive (F1) component demonstrated a higher value (Figure 3A), suggesting the velocity control dominance. On the other hand, the Br/Kr ratio for the corrective (F2) component demonstrated a much smaller value (Figure 3A), suggesting the role of F2 component in correction of positional errors (69). In contrast, CAs showed a selective decrease in the Br/Kr ratio for the predictive (F1) component (Figure 3A), suggesting poor recruitment of the predictive velocity control and compensatory dependence on the position-dependent pursuit (69). The loss of component-specific differences in the Br/Kr ratio suggests impairment of predictive control in CA. Indeed, the decrease in Br/Kr ratio in CA correlated with the increase in error in the predictive (F1) movement (Figure 3B) (69). Another critical difference between the control and CA was the increased delay of the predictive (F1) component in CA (Figure 3C). In the control subjects, the predictive (F1) movement lagged the target motion only by 66 ms, which was too small to be a visual feedback delay (i.e., a proof of prediction) (69). In contrast, in patients with CA, the delay increased by more than 100 ms, as much as 172 ms. The increased delay is comparable to a visual feedback delay, demonstrating lack of compensation of feedback delay in CA patients. In summary, ataxic movements are consistent with an impairment of a forward model in terms of accuracy and delay of state prediction. As mentioned already, the delay in prediction alone provides instability in control of goal-directed movement. Moreover, the increase in prediction error makes the oscillatory movement irregular because it makes uncertainty of each corrective (i.e., feedback) movement unreliable due to increased uncertainty of both current and future states. The residual errors trigger a chain of irregular corrective movements around the target trajectory (Figure 3D, CA wrist movement). Note that the chain of corrective movements (i.e., the tremor-like movement in Figure 3D) is voluntary in nature, although it must be far from what CA patients intended to do.
Figure 3.

Deficits of forward models in patients with cerebellar ataxia (CA). (A) Comparison of the Br/Kr ratios that represents recipe of the motor commands for the F1 and F2 components between the controls and the cerebellar patients. Controls: Br/Kr ratios of the control subjects for the F1 component (top) and the F2 component (bottom) (n = 13). Note the highly significant difference between the two components. Patients: Br/Kr ratios of the patients for the F1 (top) and the F2 (bottom) components (n = 19). Note the selective decrease in Br/Kr ratios for the F1 component in the patients. (B) Correlation between the Br/Kr ratios for F1 component and cursor–target error for F1 (F1 error, in short). The F1 error is defined as an average error between the target motion and the F1 component of the movement. Note the negative correlation. (C) Delay of the predictive (F1) component of the movement relative to the target motion calculated with a cross-correlation analysis for controls (n = 13) and patients (n = 19). (D) A highly ataxic wrist movement of a CA patient. Note the irregular tremor-like movement trajectory. Adapted from Kakei et al. (69) under CC-BY license.
As demonstrated in Figure 1, the long loop could be disrupted at any point along the loop. In addition, the disruption may vary from a partial one to a complete one. In case of a complete disruption, malfunction of the forward model may be irreversible, and the resultant irregular tremor must be severe and persisting because the cerebellar reserve (81) is unavailable. In contrast, in case of a partial disruption, the initial irregular tremor may recover partially or completely depending on the level of compensation with the cerebellar reserve. For instance, Sasaki and his colleagues made cerebellar hemispherectomy in monkeys trained for skilled hand movements and observed CA for many months (82, 83). When the lesion involved both DN and interpositus nuclei (IN), the monkeys revealed typical cerebellar symptoms, hypotonia, asthenia, awkwardness, dysmetria, and kinetic and/or static tremor. These symptoms lasted for several months until the animals were sacrificed. However, in the cases in which the lesion involved DN but spared IN, the symptoms disappeared in a few weeks.
These studies suggest that cerebellar reserve is damaged more severely in a lesion in the SCP than in a lesion in the cerebellar hemisphere. Thus, tremor in the former lesion (e.g., intention tremor) develops more irregular and abrupt natures compared with tremor in the latter lesion (e.g., kinetic tremor in Holmes' classic study). In this regard, this type of irregular tremor may disappear in a short period when the cerebellar reserve is available, as typically seen in patients with a localized cerebellar stroke.
Impairments in “G–M Triangle”
Disruptions of the Two Loops in the “G–M Triangle”
The G–M triangle includes vital parts of the long loop and the short loop. In particular, both loops are packed into the same bundle in SCP (Figure 1, sp). On the other hand, after crossing the midline, SCP is divided into the ascending branch and the descending branch (84). The ascending branch mainly contains thicker excitatory fibers from DN, while the descending branch mainly contains finer inhibitory fibers from DN (34). Therefore, a focal lesion of SCP or a large lesion in the G–M triangle may disrupt both loops. On the other hand, a localized lesion of the ascending branch or the descending branch may disrupt the long loop or the short loop separately.
For instance, a selective disruption of the long loop disorganizes the online operation of cerebellar forward model and leads to manifestation of irregular tremors, including kinetic tremor in Holmes' classic study and intention tremor, when the dysfunction exceeds a threshold. We also hypothesize that the disruption of the short loop (i.e., removal of inhibition on the gap junctions between IO neurons) shifts IO activities toward the synchronous mode like a local injection of bicuculine into IO (85) to cause regular tremors such as regular postural tremor in Holmes' classic study.
It has been a focus of debate why “Holmes' tremor” exhibits diverse types of tremors (i.e., rest, postural, and intention tremors) after a period of time. “Holmes' tremor” (midbrain tremor) was previously called cerebellar outflow tremor, whose causal lesions include SCP, midbrain tegmentum, or posterior thalamus. These foci are aligned on the dentato-thalamic (DN-Th) tract and are in or close to the G–M triangle (Figure 1). A lesion in the G–M triangle may well disrupt the two loops in a complicated manner, causing the diverse types of tremors (Figure 4).
Figure 4.

Summary diagram. A lesion in the G–M triangle may well-disrupt the short loop (left panel) and the long loop (right panel) to cause the diverse types of tremors. In addition, the aberrant activities in the short loop (i.e., aberrant complex spike activities) may induce secondary maladaptation of cerebellar forward models through aberrant patterns of LTD and/or LTP of the cerebellar circuitry (dashed arrow).
Reorganization and Maladaptation in the G–M Triangle
Reorganization in the Short Loop
Emergence of regular rest or postural tremors in “Holmes' tremor” needs several weeks or longer (usually 4 weeks−2 years) after disruption of the short loop. The longer latent period may correspond to the time required for synaptic reorganization around the gap junctions of IO neurons, i.e., reduction or disappearance of inhibitory terminals and concomitant sprouting of excitatory terminals (86, 87). However, this hypothesis does not exclude possibility of regular tremors during acute phases (14). For instance, the above-mentioned harmaline-induced tremor model clearly suggests the existence of a switch to ignite regular tremors without chronic reorganizations of neuron circuitries.
Induction of Maladaptation Caused by Regular Tremors
The regular tremor is accompanied by abnormal synchronized IO activities. The aberrant IO activities (i.e., aberrant CS activities) may induce secondary maladaptation of cerebellar forward models through aberrant patterns of LTD and/or LTP of the cerebellar circuitry (Figure 4, dashed arrow). The problem may be twofold. First, during a regular tremor, average CS activities (>4 Hz) are much higher than normal levels of CS activities (~1 Hz). Therefore, CS activities are corrupted by increased noise (i.e., low S/N ratio) during regular tremors. Second, Hoang et al. (85) recently found that high coupling strengths of IO neurons induce their synchronous firing and decrease the amount of information encoded by firing dynamics of IO neurons. The two mechanisms may gradually deteriorate the forward model and increase its prediction error, resulting in irregular tremor. In this regard, it may be possible to explain the intention tremor of “Holmes' tremor” with this mechanism.
In conclusion, it is important to note that in “Holmes' tremor,” or more generally tremors induced by lesions in the G–M triangle, disruptions of the two loops coexist and induce the regular and irregular types of tremors in various combinations depending on the location and size of the lesion. In addition, the complex pathological condition is further prone to secondary changes such as reorganization and maladaptation.
Consideration of Neuroimaging Studies
Our proposal of a dual pathogenesis will now require an in-depth multimodal assessment to establish how it can be translated into a direct clinical practice. This ambitious goal will likely remain a highly challenging task. For the time being, let us conclude this manuscript with a brief consideration of neuroimaging studies because it allows to assess the morphological and functional aspects in cerebellar tremor patients. Structural imaging by MRI provides insights for focal or diffuse anatomical lesions, complemented in particular by diffusion imaging (DTI), fMRI, and assessment of metabolic brain networks (88, 89). Diffusion tractography shows the neuronal connections in the brain and allows to draw conclusions in terms of deafferentation following a focal lesion such as a stroke and infer on remote effects of this connection.
One typical example was provided by Seidel et al. who reported the case of a 20-year-old patient with right-sided Holmes' tremor 9 months after a midbrain/pontine hemorrhage (90). Tractography demonstrated a reduced fiber connectivity of the superior and middle cerebellar peduncles on the lesioned side. The hemorrhage affected the RN directly and impacted on nigro-striatal projections and the cortico-rubro-cerebellar loop, underlining that tremor was probably due to a deafferentation mechanism (88). These findings are coincident with the present proposal of reorganizations in the short loop (see section Reorganization and Maladaptation in the G–M Triangle). Tractography has been used successfully to target the dentato-rubro-thalamic tract to plan the implantation of electrodes for deep brain stimulation in combination with traditional landmark-based targeting techniques (91).
In ET, a functional disconnection of dentate nuclei with cortical, subcortical, and cerebellar areas has been demonstrated recently (92). Changes in the cerebellum positively correlated with tremor amplitude, in contrast with changes in the bilateral thalamus that negatively correlated with tremor amplitude. The functional connectivity with the supplementary motor area, precentral and postcentral gyri, and prefrontal cortex negatively correlated with tremor scores. These observations confirm the importance of the cerebello-thalamo-cortical pathway in tremor genesis. These, from imaging studies, favor the present hypothesis that a pathological synchronization of IO neurons sparks a chain reaction in the cerebello-cerebral circuits (e.g., synchronous CS, rebound potentiation of DN neurons, and finally rhythmical activation of M1 through the cerebello-thalamo-cortical pathway) (see section Failure of the Short Loop Results in Regular Oscillatory Tremors). In the systematic literature search by Ceresa–Quattrone, who combined the terms ET with the following keywords MRI, VBM, MRS, DTI, fMRI, PET, and SPECT, a total of 51 neuroimaging studies met search criteria, divided into 19 structural and 32 functional studies (93). The studies showed similar findings but without defining a clear topography of the neurodegenerative process. The majority of studies identified functional and structural abnormalities in several portions of the anterior and posterior cerebellar lobules, but the authors stressed the absence of correlation between these neural changes and the clinical symptoms of ET. The authors also highlighted the high variability in results.
We did not expand here on the numerous MRI reports describing the location of lesions in the G–M triangle and the involvement of the central tegmental tract, the dentatorubrothalamic tract, the transaxonal degeneration, and Wallerian degeneration [see the recent work of Raeder et al. (94) focusing on imaging characteristics of transaxonal degenerations involving cerebellar connections].
Conclusion
We tried to explain complex phenotypes of tremors or tremor-like movements with two physiological principles related to the G–M triangle, pointing out the abnormal motor behavior on the basis of errors in feedforward and feedback loops. The G–M triangle appears in our view as an interface between sensory and motor processes. Tremor is viewed as the result of errors in predictions executed by the posterior fossa structures including the cerebellum, causing an unstable state. Although our hypothesis may not cover all tremors or tremor-like movement disorders, our approach integrates the latest theories of cerebellar physiology and provides explanations how various lesions in or around the G–M triangle results in tremors or tremor-like movements. These two elemental mechanisms can be extrapolated to the loops between dentate nuclei and reticular nuclei in the brainstem acting as reverberation (95). We did not speculate on the neurobiological mechanisms underlying the aberrant synaptogenesis in the G–M triangle (96).
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Prof. Koji Ito for his invaluable comments and discussions.
Footnotes
Funding. This work was supported by Grants-in-aid from the Ministry of Education, Culture, Sports, Science and Technology in Japan (MEXT) (http://www.mext.go.jp/) (Nos. 26120003, 14580784, 15016008, 16015212, 20033029, and 21500319 to SK; Nos. 25430007, 26120005, and 16K12476 to HT), the Japan Science and Technology Agency (A-STEP) to SK (http://www.jst.go.jp/), the Japan Science and Technology Agency (PRESTO: Intelligent Cooperation and Control) (SK), NBRP Japanese Monkeys through the National BioResource Project of MEXT, the JSPS Programs (Program for Advancing Strategic International Networks to Accelerate the Circulation of Talented Researchers, and Embodied-Brain Systems Science) (HT), and the Hitachi-Kurata and the Tateishi Science Foundations (HT). This research was also supported by AMED under Grant Number 16ek0109048h0003 to SK.
References
- 1.Elble RJ. Animal models of action tremor. Mov Disord. (1998) 13(Suppl 3):35–9. 10.1002/mds.870131306 [DOI] [PubMed] [Google Scholar]
- 2.Lenka A, Louis ED. Revisiting the clinical phenomenology of “cerebellar tremor”. Beyond intention tremor. Cerebellum. (2019) 18:565–74. 10.1007/s12311-018-0994-6 [DOI] [PubMed] [Google Scholar]
- 3.Louis ED Essential tremor: a common disorder of Purkinje neurons? Neuroscientist . (2016) 22:108–18. 10.1177/1073858415590351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bodranghien F, Bastian A, Casali C, Hallett M, Louis ED, Manto M, et al. Consensus paper: revisiting the symptoms and signs of cerebellar syndrome. Cerebellum. (2016) 15:369–91. 10.1007/s12311-015-0687-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Louis ED, Lenka A. The olivary hypothesis of essential tremor: time to lay this model to rest? Tremor Other Hyperkinet Mov (NY). (2017) 7:473. 10.5334/tohm.380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Holmes G. Clinical symptoms of cerebellar disease and their interpretation, Lecture I. Lancet. (1922) 202:1178–82.17566248 [Google Scholar]
- 7.Holmes G. Clinical symptoms of cerebellar disease and their interpretation, Lecture II. Lancet. (1922) 202:1232–7. [Google Scholar]
- 8.Haines DE, Manto MU. Cerebellar classic II. Cerebellum. (2007) 6:141–56. 10.1080/14734220701415208 [DOI] [Google Scholar]
- 9.Garcin R. The ataxias. In: Vinken PJ, Bruyn WG, editors. Handbook of Clinical Neurology, Vol. 1, Disturbances of Nervous Function. Amsterdam: North Holland Publishing Company; (1969). p. 309–35. [Google Scholar]
- 10.Charcot JM. Séance du 14mars. C R Soc Biol (Paris). (1868) 20:13. [Google Scholar]
- 11.Deuschl G, Bain P, Brin M. Consensus statement of the movement disorder society on tremor. Ad Hoc Scientific Committee. Mov Disord. (1998) 13(Suppl 3):2–23. 10.1002/mds.870131303 [DOI] [PubMed] [Google Scholar]
- 12.Bhatia KP, Bain P, Bajaj N, Elble RJ, Hallett M, Louis ED, et al. Consensus statement on the classification of tremors. From task force on tremor of the International Parkinson and Movement Disorder Society. Mov Disord. (2018) 33:75–87. 10.1002/mds.27121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mitoma H. Intention tremor exaggerated by visually guided movement. Eur Neurol. (1996) 36:177–8. 10.1159/000117241 [DOI] [PubMed] [Google Scholar]
- 14.Choi SM. Movement disorders following cerebrovascular lesions in cerebellar circuits. J Mov Disord. (2016) 9:80–8. 10.14802/jmd.16004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Boonstra F, Florescu G, Evans A, Steward C, Mitchell P, Desmond P, et al. Tremor in multiple sclerosis is associated with cerebello-thalamic pathology. J Neural Transm (Vienna). (2017) 124:1509–14. 10.1007/s00702-017-1798-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lehéricy S, Grand S, Pollak P, Poupon F, Le Bas JF, Limousin P, et al. Clinical characteristics and topography of lesions in movement disorders due to thalamic lesions. Neurology. (2001) 57:1055–66. 10.1212/WNL.57.6.1055 [DOI] [PubMed] [Google Scholar]
- 17.Bastian AJ, Thach WT. Cerebelalr outflow lesions: a comparison of movement deficits resulting from lesions at the levels of the cerebellum and thalamus. Ann Neurol. (1995) 38:881–92. 10.1002/ana.410380608 [DOI] [PubMed] [Google Scholar]
- 18.Krauss JK, Wakhloo AK, Nobbe F, Tränkle R, Mundinger F, Seeger W. Lesion of dentatothalamic pathways in severe post-traumatic tremor. Neurol Res. (1995) 17:409–16. 10.1080/01616412.1995.11740353 [DOI] [PubMed] [Google Scholar]
- 19.Solomon DH, Barohn RJ, Bazan C, Grissom J. The thalamic ataxia syndrome. Neurology. (1994) 44:810–4. 10.1212/WNL.44.5.810 [DOI] [PubMed] [Google Scholar]
- 20.Fahn S, Jankovic J, Hallett M. Motor control: physiology of voluntary and involuntary movements. In: Fahn S, Jankovic J, Hallett M, editors. Principles and Practice of Movement Disorders. 2nd ed. Philadelphia, PA: Elsevier Saunders; (2011). p. 36–54. 10.1016/B978-1-4377-2369-4.00002-0 [DOI] [Google Scholar]
- 21.Lawrenson C, Bares M, Kamondi A, Kovács A, Lumb B, Apps R, et al. The mystery of the cerebellum: clues from experimental and clinical observations. Cerebellum Ataxias. (2018) 5:8. 10.1186/s40673-018-0087-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Berkovic SF, Bladin PF. Rubral tremor: clinical features and treatment of three cases. Clin Exp Neurol. (1984) 20:119–28. [PubMed] [Google Scholar]
- 23.Kipfer S, Frigerio SB. Post-ischemic stroke Holmes' tremor of the upper limb. Mov Disord. (2013) 28:1347. 10.1002/mds.25621 [DOI] [PubMed] [Google Scholar]
- 24.Deuschl G, Toro C, Valls-Solé J, Zeffiro T, Zee DS, Hallett M. Symptomatic and essential palatal tremor. 1. Clinical, physiological and MRI analysis. Brain. (1994) 117:775–88. 10.1093/brain/117.4.775 [DOI] [PubMed] [Google Scholar]
- 25.Goyal M, Versnick E, Tuite P, Cyr JS, Kucharczyk W, Montanera W, et al. Hypertrophic olivary degeneration: meta-analysis of the temporal evolution of MR findings. AJNR Am J Neuroradiol. (2000) 21:1073–7. PMCID: PMC7973904 [PMC free article] [PubMed] [Google Scholar]
- 26.Lehéricy S, Gerardin E, Poline JB, Meunier S, Van de Moortele PF, Le Bihan D, et al. Motor execution and imagination networks in post-stroke dystonia. Neuroreport. (2004) 15:1887–90. 10.1097/00001756-200408260-00010 [DOI] [PubMed] [Google Scholar]
- 27.Louis ED. Essential tremor: from bedside to bench and back to bedside. Curr Opin Neurol. (2014) 27:461–7. 10.1097/WCO.0000000000000115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Louis ED, Kuo SH, Vonsattel JP, Faust PL. Torpedo formation and Purkinje cell loss: modeling their relationship in cerebellar disease. Cerebellum. (2014) 13:433–9. 10.1007/s12311-014-0556-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Louis ED, Faust PL. Essential tremor within the broader context of other forms of cerebellar degeneration. Cerebellum. (2020) 19:879–96. 10.1007/s12311-020-01160-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kelly RM, Strick PL. Cerebellar loops with motor cortex and prefrontal cortex of a nonhuman primate. J Neurosci. (2003) 23:8432–44. 10.1523/JNEUROSCI.23-23-08432.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Strick PL, Dum RP, Fiez JA. Cerebellum and nonmotor function. Annu Rev Neurosci. (2009) 32:413–34. 10.1146/annurev.neuro.31.060407.125606 [DOI] [PubMed] [Google Scholar]
- 32.Bostan AC, Dum RP, Strick PL. Cerebellar networks with the cerebral cortex and basal ganglia. Trends Cogn Sci. (2013) 17:241–54. 10.1016/j.tics.2013.03.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Garifoli A, Scardilli G, Perciavalle V. Effects of cerebellar dentate nucleus GABAergic cells on rat inferior olivary neurons. Neuroreport. (2001) 12:3709–13. 10.1097/00001756-200112040-00021 [DOI] [PubMed] [Google Scholar]
- 34.De Zeeuw CI, Simpson JI, Hoogenraad CC, Galjart N, Koekkoek SKE, Ruigrok TJH. Microcircuitry and function of the inferior olive. Trends Neurosci. (1998) 21:391–400. 10.1016/S0166-2236(98)01310-1 [DOI] [PubMed] [Google Scholar]
- 35.Bazzigaluppi P Ruigrok T Saisan P De Zeeuw CI de Jeu M. Properties of the Nucleo-Olivary pathway: an in vivo whole-cell patch clamp study. PLoS ONE. (2012). 7:e46360. 10.1371/journal.pone.0046360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.De Zeeuw CI, Holstege JC, Ruigrok TJ, Voogd J. Mesodiencephalic and cerebellar terminals terminate upon the same dendritic spines in the glomeruli of the cat and rat inferior olive: an ultrastructural study using a combination of [3H]leucine and wheat germ agglutinin coupled horseradish peroxidase anterograde tracing. Neuroscience. (1990) 34:645–55. 10.1016/0306-4522(90)90171-Y [DOI] [PubMed] [Google Scholar]
- 37.Ruigrok TJH, Voogd J. Cerebellar influence on olivary excitability in the cat. Eur J Neurosci. (1995) 7:679–93. 10.1111/j.1460-9568.1995.tb00672.x [DOI] [PubMed] [Google Scholar]
- 38.Schmolesky MT, Wang Y, Hanes DP, Thompson KG, Leutgeb S, Schall JD, et al. Signal timing across the macaque visual system. J Neurophysiol. (1998) 79:3272–8. 10.1152/jn.1998.79.6.3272 [DOI] [PubMed] [Google Scholar]
- 39.Wolpert DM, Miall RC. Forward models for physiological motor control. Neural Netw. (1996) 9:1265–79. 10.1016/S0893-6080(96)00035-4 [DOI] [PubMed] [Google Scholar]
- 40.Wolpert DM, Miall RC, Kawato M. Internal models in the cerebellum. Trends Cogn Sci. (1998) 2:338–47. 10.1016/S1364-6613(98)01221-2 [DOI] [PubMed] [Google Scholar]
- 41.McNamee D, Wolpert DM. Internal models in biological control. Annu Rev Control Robot Auton Syst. (2019) 2:339–64. 10.1146/annurev-control-060117-105206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Heinks-Maldonado TH, Nagarajan SS, Houde JF. Magnetoencephalographic evidence for a precise forward model in speech production. Neuroreport. (2006) 17:1375–9. 10.1097/01.wnr.0000233102.43526.e9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bass P, Jacobsen T, Schroger E. Suppression of the auditory N1 event-related potential component with unpredictable self-initiated tones: evidence for internal forward models with dynamic stimulation. Int J Psychophysiol. (2008) 70:137–43. 10.1016/j.ijpsycho.2008.06.005 [DOI] [PubMed] [Google Scholar]
- 44.Miall RC, Christensen LO, Cain O, Stanley J. Disruption of state estimation in the human lateral cerebellum. PLoS Biol. (2007) 5:e316. 10.1371/journal.pbio.0050316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lesage E, Morgan BE, Olson AC, Meyer AS, Miall RC. Cerebellar rTMS disrupts predictive language processing. Curr. Biol. (2012) 22:R794–5. 10.1016/j.cub.2012.07.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lang CE, Bastian AJ, Cerebellar subjects show impaired adaptation of anticipatory EMG during catching . J Neurophysiol. (1999) 82:2108–19. 10.1152/jn.1999.82.5.2108 [DOI] [PubMed] [Google Scholar]
- 47.Nowak DA, Hermsdörfer J, Rost K, Timmann D, Topka H, Predictive and reactive finger force control during catching in cerebellar degeneration . Cerebellum. (2004) 3:227–35. 10.1080/14734220410019057 [DOI] [PubMed] [Google Scholar]
- 48.Nowak DA, Timmann D, Hermsdörfer J, Dexterity in cerebellar agenesis . Neuropsychologia. (2007) 45:696–703. 10.1016/j.neuropsychologia.2006.08.011 [DOI] [PubMed] [Google Scholar]
- 49.Miall RC, Weir DJ, Wolpert DM, Stein J. Is the cerebellum a Smith predictor? J Mot Behav. (1993) 25:203–16. 10.1080/00222895.1993.9942050 [DOI] [PubMed] [Google Scholar]
- 50.Haggard P, Wing A. Coordinate responses following mechanical perturbations of the arm during prehension. Exp Brain Res. (1995) 102:483–94. 10.1007/BF00230652 [DOI] [PubMed] [Google Scholar]
- 51.Bastian AJ. Learning to predict the future: the cerebellum adapts feedforward movement control. Curr Opin Neurobiol. (2006) 16:645–9. 10.1016/j.conb.2006.08.016 [DOI] [PubMed] [Google Scholar]
- 52.Ebner TJ, Pasalar S. Cerebellum predicts the future motor state. Cerebellum. (2008) 7:583–8. 10.1007/s12311-008-0059-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Brodal P, Bjaalie JG. Organization of the pontine nuclei. Neurosci Res. (2003) 13:83–118. 10.1016/0168-0102(92)90092-Q [DOI] [PubMed] [Google Scholar]
- 54.Schmahmann JD. Disorders of the cerebellum: ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J Neuropsychiatry Clin Neurosci. (2004) 16:367–78. 10.1176/jnp.16.3.367 [DOI] [PubMed] [Google Scholar]
- 55.Ishikawa T, Tomatsu S, Tsunoda Y, Lee J, Hoffman DS, Kakei S. Releasing dentate nucleus cells from Purkinje cell inhibition generates output from the cerebrocerebellum. PLoS ONE. (2014) 9:e108774. 10.1371/journal.pone.0108774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ishikawa T, Tomatsu S, Izawa J, Kakei S. The cerebro-cerebellum: could it be loci of forward models? Neurosci Res. (2016) 104:72–9. 10.1016/j.neures.2015.12.003 [DOI] [PubMed] [Google Scholar]
- 57.Tomatsu S, Ishikawa T, Tsunoda Y, Lee J, Hoffman DS, Kakei S. Information processing in the hemisphere of the cerebellar cortex for control of wrist movement. J Neurophysiol. (2016) 115:255–70. 10.1152/jn.00530.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Na J, Sugihara I, Shinoda Y, The entire trajectories of single pontocerebellar axons and their lobular and longitudinal terminal distribution patterns in multiple aldolase C-positive compartments of the rat cerebellar cortex . J. Comp. Neurol. (2019) 527:2488–511. 10.1002/cne.24685 [DOI] [PubMed] [Google Scholar]
- 59.Wu HS, Sugihara I, Shinoda Y. Projection patterns of single mossy fibers originating from the lateral reticular nucleus in the rat cerebellar cortex and nuclei. J Comp Neurol. (1999) 411:97–118. [DOI] [PubMed] [Google Scholar]
- 60.Thach WT. Timing of activity in cerebellar dentate nucleus and cerebral motor cortex during prompt volitional movement. Brain Res. (1975) 88:233–41. 10.1016/0006-8993(75)90387-X [DOI] [PubMed] [Google Scholar]
- 61.Thach WT. Correlation of neural discharge with pattern and force of muscular activity, joint position, and direction of intended next movement in motor cortex and cerebellum. J Neurophysiol. (1978) 41:654–76. 10.1152/jn.1978.41.3.654 [DOI] [PubMed] [Google Scholar]
- 62.Thier P, Markanday A. Role of the vermal cerebellum in visually guided eye movements and visual motion perception. Annu Rev Vis Sci. (2019) 5:247–68. 10.1146/annurev-vision-091718-015000 [DOI] [PubMed] [Google Scholar]
- 63.Tanaka H, Ishikawa T, Kakei S. Neural evidence of the cerebellum as a state predictor. Cerebellum. (2019) 18:349–71. 10.1007/s12311-018-0996-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tanaka H, Ishikawa T, Lee J, Kakei S. The cerebre-cerebellum as a locus of forward model; a review. Front Syst Neurosci. (2020) 14:19. 10.3389/fnsys.2020.00019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kalman RE, Bucy RS, New results in linear filtering and prediction theory . J. Basic Eng. (1961) 83:95–108. 10.1115/1.3658902 [DOI] [Google Scholar]
- 66.Paulin M. A Kalman filter theory of the cerebellum. In: Arbib MA, Amari S, editors. Dynamic Interactions in Neural Networks: Models and Data. New York, NY: Springer; (1989). p. 239–59. 10.1007/978-1-4612-4536-0_15 [DOI] [Google Scholar]
- 67.Paulin M. Neural representations of moving systems. In: Schmahmann JD, editor. The Cerebellum and Cognition. San Diego, CA: Academic Press; (1997). p. 515–33. 10.1016/S0074-7742(08)60368-2 [DOI] [PubMed] [Google Scholar]
- 68.Droulez J, Cornílleau-Pérèz V. Application of the coherence scheme to the multisensory fusion problem. In: Berthoz A, editor. Multisensory Control of Movement. Oxford: Oxford University Press; (1993). p. 485–501. 10.1093/acprof:oso/9780198547853.003.0234 [DOI] [Google Scholar]
- 69.Kakei S, Lee J, Mitoma H, Tanaka H, Manto M, Hampe CS. Contribution of the cerebellum to predictive motor control and its evaluation in ataxic patients. Front Hum Neurosci. (2019) 13:216. 10.3389/fnhum.2019.00216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Baumel Y, Jacobson GA, Cohen D. Implications of functional anatomy on information processing in the deep cerebellar nuclei. Front Cell Neurosci. (2009) 3:14. 10.3389/neuro.03.014.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hoebeek FE, Witter L, Ruigrok TJH, De Zeeuw CI. Differential olivo-cerebellar cortical control of rebound activity in the cerebellar nuclei. Proc Natl Acad Sci USA. (2010) 107:8410–5. 10.1073/pnas.0907118107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Cheng MM, Tang G, Kuo SH. Harmaline-induced tremor in mice: videotape documentation and open questions about the model. Tremor Other Hyperkinet Mov (NY). (2013) 3:tre-03-205-4668-1. 10.5334/tohm.139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Handforth A. Linking essential tremor to the cerebellum-animal model evidence. Cerebellum. (2016) 15:285–98. 10.1007/s12311-015-0750-0 [DOI] [PubMed] [Google Scholar]
- 74.Zhang X, Santaniello S. Role of cerebellar GABAergic dysfunctions in the origins of essential tremor. Proc. Natl. Acad. Sci. USA. (2019) 116:13592–601. 10.1073/pnas.1817689116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Handforth A, Lang EJ. Increased Purkinje cell complex spike and deep cerebellar nucleus synchrony as a potential basis for syndromic essential tremor. A review and synthesis of the literature. Cerebellum. (2021) 20:266–81. 10.1007/s12311-020-01197-5 [DOI] [PubMed] [Google Scholar]
- 76.Witter L, Canto CB, Hoogland TM, de Gruijl JR, De Zeeuw CI. Strength and timing of motor responses mediated by rebound firing in the cerebellar nuclei after Purkinje cell activation. Front Neural Cricuits. (2013) 7:133. 10.3389/fncir.2013.00133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Maki F, Sato S, Watanabe K, Yanagisawa T, Hagiwara Y, Shimizu T, et al. Vim thalamotomy in a patient with Holmes' tremor and palatal tremor – pathophysiological considerations. BMC Neurol. (2015) 15:26. 10.1186/s12883-015-0277-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lee J, Kagamihara Y, Tomatsu S Kakei S. The functional role of the cerebellum in visually guided tracking movement. Cerebellum. (2012) 11:426–33. 10.1007/s12311-012-0370-x [DOI] [PubMed] [Google Scholar]
- 79.Lee J, Kagamihara Y, Kakei S. A new method for functional evaluation of motor commands in patients with cerebellar ataxia. PLoS ONE. (2015) 10:e0132983. 10.1371/journal.pone.0132983 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Mitoma H, Adhikari K, Aeschlimann D, Chattopadhyay P, Hadjivassiliou M, Hampe CS, et al. Consensus paper: Neuroimmune mechanisms of cerebellar ataxias. Cerebellum. (2016) 15:213–32. 10.1007/s12311-015-0664-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Mitoma H, Buffo A, Gelfo F, Guell X, Fucà E, Kakei S, et al. Consensus paper. Cerebellar reserve: from cerebellar physiology to cerebellar disorders. Cerebellum. (2019) 19:131–53. 10.1007/s12311-019-01091-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sasaki K, Gemba H. Cortical field potentials preceding self-paced and visually initiated hand movements in one and the same monkey and influences of cerebellar hemispherectomy upon the potentials. Neurosci Lett. (1981) 25:287–92. 10.1016/0304-3940(81)90406-7 [DOI] [PubMed] [Google Scholar]
- 83.Sasaki K, Gemba H, Mizuno N. Cortical field potentials preceding visually initiated hand movements and cerebellar actions in the monkey. Exp Brain Res. (1982) 46:29–36. 10.1007/BF00238095 [DOI] [PubMed] [Google Scholar]
- 84.Ruigrok TJH, Voogd J. Cerebellar nucleo-olivary projections in the rat: an anterograde tracing study with Phaseolus vulgaris-leucoagglutinin (PHA-L). J Comp Neurol. (1990) 298:315–33. 10.1002/cne.902980305 [DOI] [PubMed] [Google Scholar]
- 85.Hoang H, Lang EJ, Hirata Y, Tokuda IT, Aihara K, Toyama K. Electrical coupling controls dimensionality and chaotic firing of inferior olive neurons. PLoS Comp Biol. (2020) 16:e1008075. 10.1371/journal.pcbi.1008075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Tsukahara N, Hultborn H, Murakami F, Fujito Y. Electrophysiological study of formation of new synapses and collateral sprouting in red nucleus neurons after partial denervation. J Neurophysiol. (1975) 38:1359–72. 10.1152/jn.1975.38.6.1359 [DOI] [PubMed] [Google Scholar]
- 87.Katsumaru H, Murakami F, Wu JY, Tsukahara N. Sprouting of GABAergic synapses in the red nucleus after lesions of the nucleus interpositus in the cat. J Neurosci. (1986) 6:2864–74. 10.1523/JNEUROSCI.06-10-02864.1986 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Klein JC. Diffusion imaging in tremor. In: Grimaldi G, Manto M, editor. Mechanisms and Emerging Therapies in Tremor Disorders, New York, NY: Springer. (2013). p. 391–401. 10.1007/978-1-4614-4027-7_21 [DOI] [Google Scholar]
- 89.Pourfar M, Niethammer M, Eidelberg D. Metabolic networks in Parkinson's disease. In: Grimaldi G, Manto M, editor. Mechanisms and Emerging Therapies in Tremor Disorders. New York, NY: Springer; (2013). p. 403–15. 10.1007/978-1-4614-4027-7_22 [DOI] [Google Scholar]
- 90.Seidel S, Kasprian G, Leutmezer F, Prayer D, Auff E. Disruption of nigrostriatal and cerebellothalamic pathways in dopamine responsive Holmes' tremor. J Neurol Neurosurg Psychiatry. (2009) 80:921–3. 10.1136/jnnp.2008.146324 [DOI] [PubMed] [Google Scholar]
- 91.Coenen VA, Allert N, Mädler B. A role of diffusion tensor imaging fiber tracking in deep brain stimulation surgery: DBS of the dentato-rubro-thalamic tract (drt) for the treatment of therapy-refractory tremor. Acta Neurochir (Wien). (2011) 153:1579–85. 10.1007/s00701-011-1036-z [DOI] [PubMed] [Google Scholar]
- 92.Tikoo S, Pietracupa S, Tommasin S, Bologna M, Petsas N, Bharti K, et al. Functional disconnection of the dentate nucleus in essential tremor. J Neurol. (2020) 267:1358–67. 10.1007/s00415-020-09711-9 [DOI] [PubMed] [Google Scholar]
- 93.Ceresa A, Quattrone A. Linking essential tremor to the cerebellum : neuroimaging evidence. Cerebellum. (2016) 15:263–75. 10.1007/s12311-015-0739-8 [DOI] [PubMed] [Google Scholar]
- 94.Raeder MTL, Reis EP, Campos BM, Zamilute IAG, França Júnior MC, Reis F. Transaxonal degenerations of cerebellar connections: the value of anatomical knowledge. Arq Neuropsiquiatr. (2020) 78:301–6. 10.1590/0004-282x20200021 [DOI] [PubMed] [Google Scholar]
- 95.Dietrichs E, Heier MS, Faye-Lund H, Nakstad PH. Bulbar myoclonus without palatal myoclonus. A hypothesis on pathophysiology. Eur J Neurol. (1999) 6:367–70. 10.1046/j.1468-1331.1999.630367.x [DOI] [PubMed] [Google Scholar]
- 96.Sarnat HB, Flores-Sarnat L, Auer RN. Sequence of synaptogenesis in the fetal and neonatal cerebellar system - part 1: Guillain-Mollaret triangle (dentato-rubro-olivo-cerebellar circuit). Dev Neurosci. (2013) 35:69–81. 10.1159/000350503 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.