

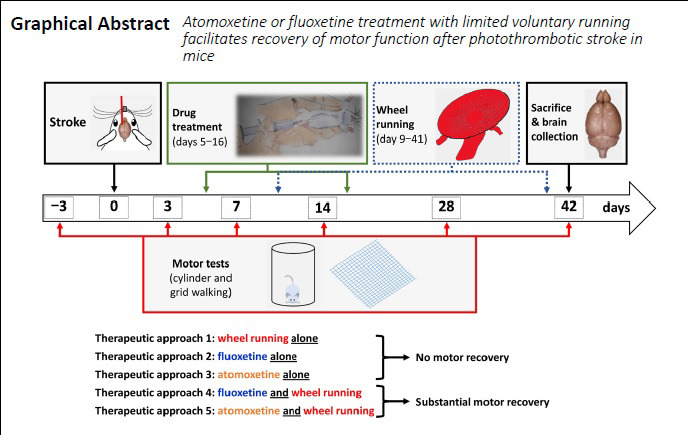
Keywords: drug repurposing, neural repair, physical exercise, physiotherapy, post-stroke recovery, pre-clinical trial, FLAME trial, selective norepinephrine reuptake inhibitor, selective serotonin reuptake inhibitor, stroke pharmacotherapy
Abstract
Currently, there is an unmet need for treatments promoting post-stroke functional recovery. The aim of this study was to evaluate and compare the dose-dependent effect of delayed atomoxetine or fluoxetine therapy (starting on post-stroke day 5), coupled with limited physical exercise (2 hours daily voluntary wheel running; post-stroke days 9 to 42), on motor recovery of adult male mice after photothrombotic stroke. These drugs are selective norepinephrine or serotonin reuptake inhibitors indicated for disorders unrelated to stroke. The predetermined primary end-point for this study was motor function measured in two tasks of spontaneous motor behaviors in grid-walking and cylinder tests. Additionally, we quantified the running distance and speed throughout the study, the number of parvalbumin-positive neurons in the medial agranular cortex and infarct volumes. Both sensorimotor tests revealed that neither limited physical exercise nor a drug treatment alone significantly facilitated motor recovery in mice after stroke. However, combination of physical exercise with either of the drugs promoted restoration of motor function by day 42 post-stroke, with atomoxetine being a more potent drug. This was accompanied by a significant decrease in parvalbumin-positive inhibitory interneurons in the ipsilateral medial agranular cortex of mice with recovering motor function, while infarct volumes were comparable among experimental groups. If further validated in larger studies, our observations suggest that add-on atomoxetine or fluoxetine therapy coupled with limited, structured physical rehabilitation could offer therapeutic modality for stroke survivors who have difficulty to engage in early, high-intensity physiotherapy. Furthermore, in light of the recently completed Assessment oF FluoxetINe In sTroke recoverY (AFFINITY) and Efficacy oF Fluoxetine-a randomisEd Controlled Trial in Stroke (EFFECTS) trials, our observations call for newly designed studies where fluoxetine or atomoxetine pharmacotherapy is evaluated in combination with structured physical rehabilitation rather than alone. This study was approved by the Texas Tech University Health Sciences Center Institutional Animal Care and Use Committee (protocol # 16019).
Chinese Library Classification No. R453; R364; R741
Introduction
The unmet need for treatments promoting functional recovery after stroke has led to an increased research focus on understanding neural repair and post-stroke recovery mechanisms to develop restorative therapies (Bernhardt et al., 2017b, 2019; Corbett et al., 2017; Tsintou et al., 2020). The main therapeutic strategies targeting post-stroke recovery include physical rehabilitation, brain and peripheral stimulation and pharmacological treatment, which are distinctly different from reperfusion and neuroprotective approaches used in acute stroke (Bernhardt et al., 2016, 2017b; Yang et al., 2020). These strategies modulate and/or enhance the cellular and molecular mechanisms of neural plasticity (Murphy and Corbett, 2009; Carmichael, 2016; Hatakeyama et al., 2020), with physiotherapy being the most reproduced intervention with positive impact on post-stroke recovery in preclinical and clinical studies (Krakauer et al., 2012; Bernhardt et al., 2016). Notably, physical rehabilitation is mostly effective at high intensity and early after stroke – a period when there is enhanced neuroplasticity in the brain (Krakauer et al., 2012). Unfortunately, most stroke survivors are unable to meet the requirement of high-intensity and early rehabilitation, because of which their recovery is usually very limited (Krakauer et al., 2012; Nicholson et al., 2013). One potential approach to overcome this problem maybe through add-on pharmacological therapy (Cramer, 2015; Carmichael, 2016).
To this end, the aim of this study was to evaluate the effect of delayed pharmacological therapy by atomoxetine or fluoxetine, coupled with limited physical rehabilitation, on recovery of motor function in mice after ischemic stroke. Atomoxetine is a selective norepinephrine reuptake inhibitor, which elevates synaptic levels of norepinephrine and is approved for management of attention deficit hyperactivity disorder (Christman et al., 2004), whereas fluoxetine is a selective serotonin reuptake inhibitor, which elevates synaptic levels of serotonin and is indicated for management of major depression (Wong et al., 1995).
Our study is the first to directly evaluate and compare the dose-dependent effect of delayed atomoxetine or fluoxetine therapy (starting on post-stroke day 5), coupled with limited physical rehabilitation (2 hours daily voluntary wheel running; post-stroke days 9 to 42), on recovery of motor function in the mouse photothrombotic model of ischemic stroke. We applied rigorous and clinically relevant experimental design and analysis standards to test the efficacy of these therapeutic combinations in adult male mice. The predetermined primary end-point for this study was motor function measured in two tasks of spontaneous motor behaviors of the forelimb in grid-walking and cylinder tests. Mechanistic elucidation of the observed effects was not an objective of this study, because the pharmacological targets and primary molecular signaling pathways of these drugs have been well-defined (Wong et al., 1995; Christman et al., 2004). Our observations indicate that neither limited physiotherapy nor any of the drug treatments alone significantly facilitate motor recovery in mice after stroke. However, the results of both sensorimotor tests reveal that combination of physical rehabilitation with either of the drugs promotes restoration of motor function by day 42 post-stroke, with atomoxetine being the more portent of the two drugs.
Materials and Methods
Animals
Twelve to fourteen week-old, male CD-1 mice (~36 g at the start of the experiments, n = 105) were used in this study which was approved by the Texas Tech University Health Sciences Center Institutional Animal Care and Use Committee (protocol # 16019, last approved on December 13, 2019). The animals were obtained from Charles River Laboratories (Wilmington, MA, USA) and maintained in standard AAALAC-approved animal care facility with controlled temperature and humidity, 12-hour light/dark cycle and ad libitum access to water and chow. To minimize handling stress, mice were individually handled by investigators ~2 minutes once or twice daily for 1 week before evaluation of their baseline motor function in grid-walking and cylinder tests. Mice were housed individually after stroke, and no animal was eliminated from the study.
Study design and treatments
All experimental group assignments (sham vs. stroke, and treatments within stroke animals) and brain processing by completion of the study (fixing vs. fresh tissue collection) were done randomly (https://www.random.org/lists/) (Jayaraman et al., 2020). Experimental groups (n = 12/group; Figure 1) included “Sham” with no stroke or treatments; “Stroke” with stroke but no treatments; “Vehicle” which had stroke, were treated with saline and physical exercise; “Atomo 0.3” – had stroke, were treated with 0.3 mg/kg atomoxetine and physical exercise; “Atomo 1.0” – had stroke, were treated with 1 mg/kg atomoxetine and physical exercise; “Fluox 3.0” – had stroke, were treated with 3 mg/kg fluoxetine and physical exercise; and “Fluox 10.0” – had stroke, were treated with 10 mg/kg fluoxetine and physical exercise. Atomoxetine and fluoxetine (product# A791400 and F597100, Toronto Research Chemicals, Toronto, Canada) were injected intraperitoneally (Al Shoyaib et al., 2019), once daily (in saline, 5 mL/kg volume, ~3 hours before the dark cycle started) between post-stroke days 5 to 16. The doses of atomoxetine (0.3 and 1 mg/kg) used in this study are known to increase extracellular concentrations of norepinephrine in the mouse brain (Koda et al., 2010), and are equivalent to the therapeutically effective doses in humans (Ledbetter, 2006). Likewise, the doses of fluoxetine (3 and 10 mg/kg) used in our study elevate extracellular concentrations of serotonin in the mouse brain (Hodes et al., 2010), and are equivalent to its clinically used doses (Rossi et al., 2004). Daily drug treatments were started on post-stroke day 5 to be outside of neuroprotective time-window (inflection point at ~3 days post-stroke (Clarkson et al., 2010)), and completed on day 16. This drug regimen was selected to primarily modulate spontaneous endogenous plasticity and neurorestorative mechanisms at their activation after stroke through the beginning of their decline (Krakauer et al., 2012; Carmichael, 2016; Karamyan, 2021), and to limit the potential side effects of the drugs upon continuous use (Rossi et al., 2004; Ledbetter, 2006).
Figure 1.
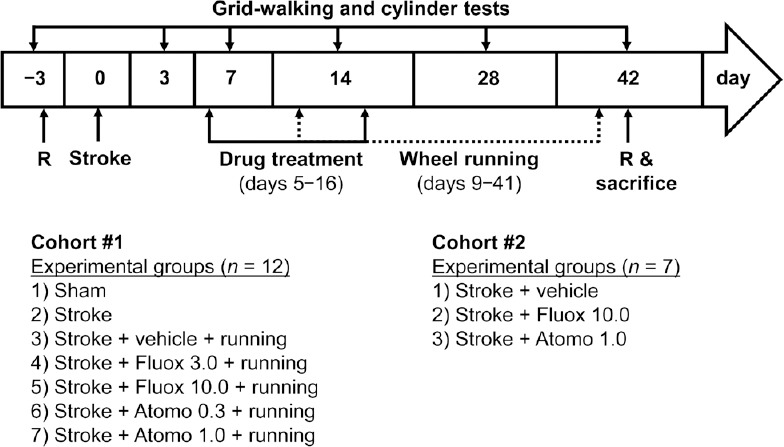
Schematic representation of the study design.
Atomo 0.3: 0.3 mg/kg atomoxetine; Atomo 1.0: 1.0 mg/kg atomoxetine; Fluox 3.0: 3 mg/kg fluoxetine; Fluox 10.0: 10 mg/kg fluoxetine; R: randomization.
Physical exercise as a rehabilitative treatment in our study consisted of voluntary wheel running in animal’s home cage for 2 hours at the start of the dark cycle (6 days per week with 1 day off on the day before behavioral tests), starting from post-stroke day 9 until the end of the study. Each running wheel (Innowheel, InnoVive, San Diego, CA) was equipped with a custom-designed counter to document the running distance. In addition, running of each animal was video recorded in weeks 4, 5 and 6 to subsequently measure the running speed. In one separate set of experiments, mice were randomly assigned to three groups (n = 7/group) and treated daily with vehicle, atomoxetine (1 mg/kg) or fluoxetine (10 mg/kg) on post-stroke days 5 to 16. Here, the animals did not run on wheels, i.e. no physical rehabilitation, to allow evaluation of the pharmacological intervention alone.
Motor function of the mice was evaluated during the light cycle (8 to 11 a.m.), on post-stroke days 3, 7, 14, 28 and 42. Association of animals with a specific experimental group was blinded from experimenters.
Photothrombotic stroke model
Photothrombosis was induced 3 days after baseline evaluation of the motor function as detailed in our earlier publication (Alamri et al., 2018). In brief, the right hemisphere (1.5 mm lateral from Bregma 0) was illuminated through intact skull with cold light (2-mm diameter, 15-minute irradiation, fiber optic illuminator light source with a halogen lamp) starting 5 minutes after intraperitoneal administration of Rose Bengal solution (8 mg/mL, 10 mL/kg) in anesthetized mice maintained at 36.9 ± 0.5°C. After surgery, mice were kept in a recovery chamber (~37°C) for ~1.5 hours followed by individual housing. Sham animals underwent the same procedure except for the light illumination. Throughout the manuscript, the left (contralateral) forelimb of all experimental mice is referred to as ‘affected forelimb’, whereas the right (ipsilateral) forelimb as ‘unaffected forelimb’.
Grid-walking test
This task was carried out as previously described in our publications (Alamri et al., 2018; Syeara et al., 2019). Footfaults for each forelimb and the total normal steps on an elevated wire grid were counted from a 5-minute video recording. Affected or unaffected footfault percentage was calculated by: number of affected or unaffected forepaw faults/number of normal steps ×100.
Cylinder test
This task was also conducted as previously described (Alamri et al., 2018; Syeara et al., 2019). To determine forelimb symmetry in exploratory rearing the use of affected, unaffected or both forelimbs was counted for each rearing in a 5-minute session from a video recording. Forelimb use symmetry index was calculated by: (number of affected forelimb use − number of unaffected forelimb use)/(number of affected forelimb use + number of unaffected forelimb use + number of use of both forelimbs).
Brain collection and infarct size evaluation
On post-stroke day 42, mice were anesthetized with isoflurane (Henry Schein Animal Health, Dublin, OH, USA) to either cardially perfuse with phosphate-buffered saline and 4% paraformaldehyde (PFA) for fixing and cryosectioning (immunostaining and infarct evaluation, n = 6/group), or to decapitate and dissect the brain to isolate 1–2 mm of peri-infarct cortex together with infarct core for western blotting (n = 6/group). The brains of PFA-perfused mice were subsequently incubated in 4% PFA overnight, sucrose cryopreserved, and cryosectioned (coronal plane, 50 μm thickness). Cresyl violet staining was carried out following a standard protocol and stained sections were digitized for volumetric analysis (Syeara et al., 2019). Quantification of infarct volume was done by multiplying the lesion area by the thickness of each section plus the distance between sections.
Immunoblotting and immunofluorescence staining
For western blotting, each collected cortical sample was processed according to our published protocol to obtain total cell lysate (Jayaraman et al., 2020), followed by standard SDS-PAGE and immunoblotting as described earlier (Wangler et al., 2012; Rashid et al., 2014). Primary antibodies used were anti-GAP-43 (growth associated protein 43), anti-synaptophysin and anti-PSD-95 (postsynaptic density protein 95) (product# D9C8, D35E4 and D27E11; Cell Signaling Technology, Danvers, MA, USA), whereas the secondary antibody was HRP-conjugated goat anti-rabbit immunoglobulin (product# 170-6515, Bio-Rad Laboratories, Hercules, CA, USA).
For immunostaining, we used free floating cryopreserved brain sections to label parvalbumin containing cells (primary antibody: mouse anti-parvalbumin antibody, 1:150 dilution, overnight incubation at room temperature, product# P3088, Sigma-Aldrich, St. Louis, MO, USA; secondary antibody, donkey AlexaFluorTM 488 IgG, 1:2000 dilution, overnight incubation at room temperature, product# A21202, Thermo Fisher Scientific, Waltham, MA, USA) following standard procedures (Jayaraman et al., 2020). Fluorescence microscopic images were acquired using a Nikon A1R MP confocal microscope to include previously defined boundaries of medial agranular cortex (AGm; Ng et al., 2015) in both hemispheres throughout the entire thickness of each coronal brain section (z-stack imaging). The acquired z-stack images were saved at maximal projections as a 2D image for each coronal section and a 400 × 400 µm2 subarea of AGm, extending to medial and dorsal pial boundaries (Figure 2), was selected in each hemisphere. Within the selected subarea, the number of immunofluorescently labeled parvalbumin-positive cells were counted in a blinded manner. For each brain, we used two coronal sections (300 µm apart in rostral-caudal axis) and a cell was counted as positive if it had any immunofluorescent label for parvalbumin.
Figure 2.
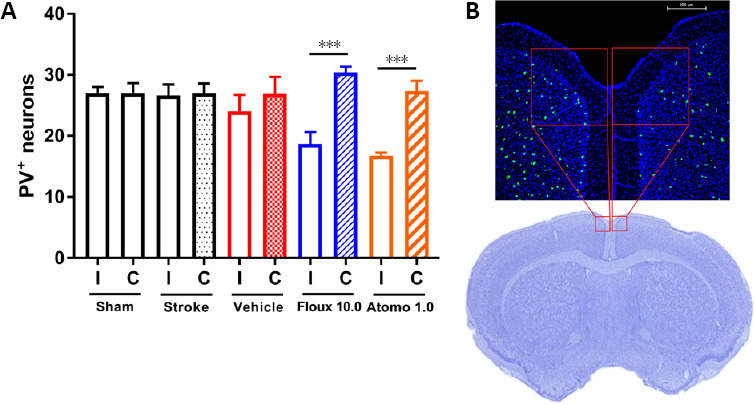
Motor recovery augmented by atomoxetine or fluoxetine is associated with decreased number of inhibitory interneurons.
(A) Fluoxetine or atomoxetine-treated mice have statistically significantly decreased number of parvalbumin positive (PV+) neurons in ipsilateral (I) medial premotor area (AGm, medial agranular cortex) compared to that of the contralateral (C) hemisphere (n = 6 per group; unpaired t-test, ***P < 0.001). Values are expressed as mean ± standard error. (B) A representative immunofluorescence confocal microscopy image of a mouse brain section on day 42 after stroke. PV signal is depicted in green and cell nuclei (stained with DAPI) in blue. Note, the representative Cresyl violet-stained brain section is shown to illustrate the imaged medial premotor area (red boxes). Atomo 1.0: 1.0 mg/kg atomoxetine; Fluox 10.0: 10 mg/kg fluoxetine.
Statistical analysis
Data from motor tests were analyzed using two-way repeated measures analysis of variance (ANOVA) and followed by Dunnett’s post hoc test for multiple comparisons (GraphPad Prism 7.05 software, GraphPad Software Inc., La Jolla, CA, USA). For all other data, means from two experimental groups were compared using two-tailed Student’s t-test (paired t-test for body weight comparisons), and means from three or more groups using one-way ANOVA followed by Dunnett’s multiple comparisons test. A P-value < 0.05 was considered statistically significant. Expressed values are mean ± standard error.
Results
Running distance and speed
To mimic mild physical rehabilitation, all vehicle and drug-treated animals received access to a running wheel in their home cage for 2 hours daily, from post-stroke day 9 through completion of the study. Because of voluntary running, on average it took about 1 week (between post-stroke days 9 to 16) for vehicle-treated mice to gradually increase their covered distance and consistently run ~2 km per session/day (Figure 3A). On the contrary, it took about 2 weeks (post-stroke days 9 to 22) for drug-treated mice to reach consistent running (Figure 3A), however they covered shorter distance compared to vehicle-treated mice (Figure 3B). No significant difference was observed in the running speed of mice among experimental groups (Figure 3C). Notably, wheel running prevented body weight gain in experimental animals, except 10 mg/kg fluoxetine group (Figure 3D). Compared to baseline body weight values, high dose fluoxetine-treated mice recorded gain of ~3 g on post-stroke day 42, similar to “sham” and “stroke” groups.
Figure 3.

Voluntary running distance and speed.
Daily running distance (panel A) and total average running (B) of mice in experimental groups. Mice treated with atomoxetine and fluoxetine covered shorter distance of running compared to vehicle-treated animals (one - (B) or two- (A) way repeated measures ANOVA followed by Dunnett’s multiple comparison, *P < 0.05, ***P < 0.001, vs. vehicle-treated group). (C) Speed of running in experimental groups did not differ significantly during the last three weeks of the study (one-way ANOVA followed by Dunnett’s multiple comparison). (D) Compared to their baseline body weights sham and stroke (sP < 0.001) and 10 mg/kg fluoxetine-treated animals (fP = 0.014) gained substantial weight by completion of the study (paired t-test). Values are expressed as mean ± standard error. n = 12/group in all panels. ANOVA: Analysis of variance, Atomo 0.3: 0.3 mg/kg atomoxetine; Atomo 1.0: 1.0 mg/kg atomoxetine; Fluox 3.0: 3 mg/kg fluoxetine; Fluox 10.0: 10 mg/kg fluoxetine.
Motor recovery in grid-walking test
To evaluate the effect of atomoxetine or fluoxetine, in combination with wheel running, on recovery of motor function after stroke, spontaneous motor behavior of the forelimb in gait was monitored in grid-walking test. As expected, focal cerebral stroke caused a significant and sustained deficit in the contralateral, i.e. affected, forelimb function of mice in the grid-walking test (Figure 4A; n = 12/group; group × day interaction F(30, 385)= 5.805, P < 0.0001). Within each stroke-affected experimental group, post hoc analyses with Dunnett’s correction revealed statistically significant differences in the affected forelimb function between baseline and corresponding post-stroke evaluation days (P = 0.0001). Within the sham group, no statistically significant difference was observed in the function of the affected forelimb between baseline and post-stroke evaluation days (P > 0.05). Gradual recovery of the impaired function was observed in all stroke-affected animals between post-stroke days 3 and 14, which continued improvement for higher dose atomoxetine and fluoxetine-treated mice, but plateaued for vehicle-treated group similar to observations by other researchers (Birjandi et al., 2020; Trout et al., 2020). Within day comparisons of experimental groups (vehicle-treated vs others; post hoc analyses with Dunnett’s correction) showed statistically significant difference of the forelimb function only between vehicle and higher dose drug-treated mice on day 42 post-stroke (Figure 4B; P = 0.007 for 1 mg/kg atomoxetine, and P = 0.011 for 10 mg/kg fluoxetine). As expected, comparison of sham-operated and vehicle-treated mice also showed statistically significant difference of the forelimb function on all evaluation days (P < 0.0001) except the baseline (P > 0.05). Lastly, similar comparisons done for the ipsilateral, i.e. unaffected, forelimb did not reveal statistically significant difference between any of the groups on any of the evaluation days (Figure 4C; P > 0.05).
Figure 4.

Enhanced motor recovery with atomoxetine or fluoxetine treatment in grid-walking test.
(A, B) Following photothrombotic stroke mice treated with atomoxetine or fluoxetine exhibited dose-dependent improvement in affected forelimb motor function (i.e., decreased number of footfaults; n = 12 per group). Compared to the vehicle-treated group, mice treated with 1 mg/kg atomoxetine (aP = 0.007) or 10 mg/kg fluoxetine (fP = 0.011) showed statistically significantly improved motor function on day 42 after stroke (two-way repeated measures ANOVA followed by Dunnett’s multiple comparison). (C) No functional deficit was observed in the unaffected (i.e., ipsilateral) forepaw of mice in experimental groups (two-way repeated measures ANOVA followed by Dunnett’s multiple comparison, P > 0.05 for all within group and within day comparisons). Values are expressed as mean ± standard error. ANOVA: Analysis of variance. Atomo 0.3: 0.3 mg/kg atomoxetine; Atomo 1.0: 1.0 mg/kg atomoxetine; Fluox 3.0: 3 mg/kg fluoxetine; Fluox 10.0: 10 mg/kg fluoxetine.
Motor recovery in cylinder test
For the same purpose, we used cylinder test to monitor spontaneous motor behavior of the forelimb in rearing. Similar to the grid-walking test, cylinder test revealed significant deficit in the function of the contralateral, i.e. stroke-affected, forelimb after stroke (Figure 5A; n = 12/group; group × day interaction F(30, 385)= 9.572, P < 0.0001). Post hoc analyses with Dunnett’s correction confirmed statistically significant differences between affected forelimb function at baseline and corresponding post-stroke evaluation days for each of stroke-affected experimental groups (P < 0.001). No statistically significant difference was observed in the function of the affected forelimb between baseline and any post-stroke evaluation day within the sham group (P > 0.05). Here too, gradual recovery of the impaired function was documented in all stroke-affected animals during the first 2 weeks post-stroke, which continued in higher dose drug-treated mice but plateaued at various degrees for others. Within day comparisons (vehicle-treated vs. other experimental groups; post hoc analyses with Dunnett’s correction) showed statistically significant difference of the forelimb function only between vehicle and higher dose drug-treated mice on day 42 post-stroke (Figure 5B; P < 0.001 for 1 mg/kg atomoxetine, and P = 0.04 for 10 mg/kg fluoxetine). Comparison of sham-operated and vehicle-treated mice also showed statistically significant difference of the forelimb function on all evaluation days (P < 0.0001) except the baseline (P > 0.05).
Figure 5.
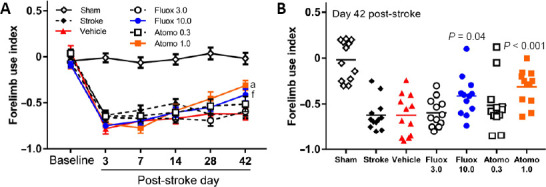
Enhanced motor recovery with atomoxetine or fluoxetine treatment in cylinder test.
(A, B) Following photothrombotic stroke mice treated with atomoxetine or fluoxetine exhibited dose-dependent improvement in affected forelimb motor function (n = 12 per group). Compared to vehicle-treated group, mice treated with 1 mg/kg atomoxetine (aP < 0.001) or 10 mg/kg fluoxetine (fP = 0.04) showed statistically significantly improved motor function on day 42 after stroke (two-way repeated measures analysis of variance followed by Dunnett’s multiple comparison). Values are expressed as mean ± standard error. Atomo 0.3: 0.3 mg/kg atomoxetine; Atomo 1.0: 1.0 mg/kg atomoxetine; Fluox 3.0: 3 mg/kg fluoxetine; Fluox 10.0: 10 mg/kg fluoxetine.
Correlation of molecular and cellular markers with motor function
In addition to functional outcomes, we evaluated three molecular and one molecular-cellular outcome measure of post-stroke motor recovery in experimental animals on day 42 post-stroke. For molecular outcome measures, the levels of regeneration-associated protein GAP-43, and synaptogenesis-associated proteins synaptophysin and PSD-95 were evaluated in protein lysates of peri-infarct cortical samples using western blotting and no significant differences were observed among experimental groups (data not shown). For the molecular-cellular outcome measure, the number of parvalbumin-positive neurons in medial premotor area (AGm) of ipsilateral and contralateral hemispheres was evaluated using immunofluorescence labeling. Our observations revealed visually apparent and statistically significant decrease in the number of parvalbumin-positive inhibitory interneurons in the ipsilateral AGm of higher dose drug-treated mice but not other experimental groups (Figure 2; P < 0.001).
Infarct location and volume
To evaluate the location of cerebral infarction and lesion volume on day 42 post-stroke, PFA-fixed brain sections of all experimental groups were stained with Cresyl violet, followed by histologic examination (Figure 6). As expected, cerebral infarction involved the primary motor cortex in all stroke-affected experimental groups. Volumetric measurements revealed no significant differences in the cerebral infarct volume among stroke-affected experimental groups (Figure 6; P > 0.05, 0.33–0.59 mm3 average stroke volume).
Figure 6.
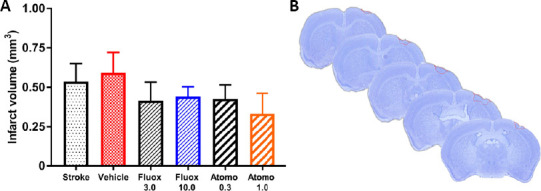
Infarct location and volume.
(A) Volumetric measurements of brain infarction did not reveal statistically significant differences among experimental groups on day 42 after stroke (n = 6 per group; one-way analysis of variance followed by Dunnett’s multiple comparison, P > 0.05). Values are expressed as mean ± standard error. (B) A representative Cresyl violet-stained mouse brain on day 42 after stroke, indicating location of infarction (outlined in red dotted line) in the primary motor cortex. Atomo 0.3: 0.3 mg/kg atomoxetine; Atomo 1.0: 1.0 mg/kg atomoxetine; Fluox 3.0: 3 mg/kg fluoxetine; Fluox 10.0: 10 mg/kg fluoxetine. Similar results were observed in the second cohort of experimental animals which received high dose drug treatments but no physical rehabilitation.
Motor function in post-stroke mice treated with atomoxetine or fluoxetine alone
In a separate set of experiments where the effect of high dose atomoxetine or fluoxetine treatment alone (i.e. no wheel running) was evaluated on motor recovery, mice showed a sustained deficit in the stroke-affected forelimb function in the grid-walking test, however no statistically significant group by day interaction was documented (Figure 7A; n = 7/group; group × day interaction F(10, 90)= 0.9223, P = 0.52). Within each experimental group, post hoc analyses with Dunnett’s correction revealed statistically significant differences in the affected forelimb function between baseline and corresponding post-stroke evaluation days (P = 0.0001). However, within day comparisons showed no significant difference in the stroke-affected forelimb function of vehicle vs. drug-treated animals (Figure 7A; P > 0.05). As expected, similar comparisons done for the unaffected forelimb did not reveal statistically significant difference between any of the groups on any of the evaluation days (Figure 7B; P > 0.05). Similarly, these animals showed significant deficit in the function of the stroke-affected forelimb in cylinder test (Figure 7C; n = 7/group; group × day interaction F(10, 90)= 2.278, P = 0.0198). Within experimental group comparisons revealed statistically significant difference in the affected forelimb function between baseline and corresponding post-stroke evaluation days (P = 0.0001). Whereas, within day comparisons revealed statistically significant difference only for high-dose atomoxetine vs vehicle-treated groups on day 14 (P = 0.026), indicating slightly worsened affected forelimb function in atomoxetine-treated animals (Figure 7C).
Figure 7.

Lack of motor recovery with atomoxetine or fluoxetine in the absence of physical rehabilitation.
(A, B) Following photothrombotic stroke mice treated with fluoxetine (10 mg/kg) or atomoxetine (1 mg/kg) did not show improvement in the affected forelimb motor function in comparison to vehicle-treated mice in grid-walking test (n = 7 per group, two-way repeated measures analysis of variance followed by Dunnett’s multiple comparison, P > 0.05). (C) No improvement was observed in the affected forelimb motor function of the same animals in cylinder test (n = 7 per group; two-way repeated measures analysis of variance followed by Dunnett’s multiple comparison, *P < 0.05 for vehicle vs. Atomo 1.0 on post-stroke day 14). Values are expressed as mean ± standard error. Atomo 1.0: 1.0 mg/kg atomoxetine; Fluox 10.0: 10 mg/kg fluoxetine.
Discussion
It is generally accepted that higher intensity and earlier physical rehabilitation leads to better functional outcomes after stroke (Egan et al., 2014; Bernhardt et al., 2017a) by enhancing spontaneous endogenous plasticity and facilitating neural repair (Krakauer et al., 2012; Bernhardt et al., 2017a). Most of these endogenous neurorestorative mechanisms are activated a few days after stroke and peak within the first 2 weeks post-stroke, making physiotherapy and other potential therapeutic interventions especially effective during this time-window (Murphy and Corbett, 2009; Lenz et al., 2015; Carmichael, 2016). Because most stroke survivors are unable to meet the requirement of high-intensity and early rehabilitation, one potential approach to overcome this problem is through add-on pharmacological therapy aimed at prolonging the enhanced neuroplasticity of the post-stroke brain and/or enhancing the effects of lower-intensity physical rehabilitation (Ng et al., 2015; Carmichael, 2016).
To expand on this idea, we evaluated the potential of atomoxetine and fluoxetine, in combination with wheel running as a form of physiotherapy, to promote motor recovery in adult male mice after ischemic stroke. Atomoxetine and fluoxetine enhance noradrenergic and serotonergic neurotransmission, and are indicated for treatment of ADHD and depression, respectively (Wong et al., 1995; Christman et al., 2004). Our rationale for focusing on these two neurotransmitter systems was because of a large body of literature indicating involvement of noradrenergic and serotonergic systems in modulation of brain plasticity, motor learning and memory (Tully and Bolshakov, 2010; Rossi, 2016; Borodovitsyna et al., 2017; Kraus et al., 2017). Although, there are other drug classes which enhance noradrenergic or serotonergic neurotransmission, we focused on reuptake inhibitors because of their high target selectivity and long history of clinical use (Lopez-Munoz and Alamo, 2009).
It is noteworthy, that there are several published experimental studies which evaluated the effects of fluoxetine on post-stroke motor recovery in rodents and reported mixed results (Windle and Corbett, 2005; Ng et al., 2015; Sun et al., 2016; Hu et al., 2020). Two other experimental studies documented beneficial effect of fluoxetine on cognitive recovery after stroke (Li et al., 2009; Vahid-Ansari and Albert, 2018). One the contrary, we are unaware of a preclinical/basic study focusing on atomoxetine for post-stroke recovery.
In the current study, daily treatment with atomoxetine or fluoxetine started on post-stroke day 5, past the acute neuroprotective time-window and beginning of subchronic phase, and lasted until day 16 to include the period during which endogenous post-stroke brain plasticity is most active (Krakauer et al., 2012; Carmichael, 2016). Physiotherapy, which consisted of voluntary wheel running (2 hours/day, 6 days/week) in animal’s home cage, started on post-stroke day 9 and lasted until the end of the study. Post-stroke day 9 is comparable to the time when most stroke survivors start some degree of regular physical rehabilitation (the 2nd week), which however is likely delayed and suboptimal (Lay et al., 2016; Fini et al., 2017). Our rationale for combing drug treatment with physiotherapy was because of the standard of care in most developed countries, where majority of stroke survivors go through some type of physical rehabilitation (Winstein et al., 2016). Although, the practice of physical rehabilitation greatly varies among providers, the current recommendation for drug testing intended for post-stroke recovery is trending towards combination therapy with physical rehabilitation (Kwakkel et al., 2020).
It is noteworthy, that both atomoxetine and fluoxetine affected voluntary running of animals. Mice treated with either drug covered shorter distance in comparison to the vehicle-treated group, despite similarities in their speed of running. This effect was especially evident during daily drug treatments (post-stroke days 9 to 16), after which the difference gradually dissipated. Notably, our results are in line with earlier observations indicating decreased locomotion/running of experimental animals in the result of chronic atomoxetine or fluoxetine treatment (Klenotich et al., 2012; Moon et al., 2014).
The daily average distance ran by animals in our study was 1.5–2 km during the 2-hour rehabilitation period, which is 5 to 10-fold shorter compared to the distance ran by mice with overnight access to the wheels (Karamyan Al-Shoyaib, unpublished observations). This amount of physical activity prevented body weight gain in all experimental groups, except 10 mg/kg fluoxetine-treated animals, which gained ~3.0 g over the course of our study, similar to “sham” and “stroke” groups. Higher-dose atomoxetine-treated animals also gained weight throughout the study (~1.2 g), however it was not at a statistically significant level. Although, food consumption of the animals was not measured in our study, we speculate that the observed weigh gain in high dose drug-treated animals has to do with shorter distance of running during the first 3 weeks of the study.
In our study, functional recovery of motor control was determined in two tasks of spontaneous motor behaviors of the forelimb, one during gait (grid-walking test) and another during exploratory rearing (cylinder test). Both tests revealed dose-dependent potential of atomoxetine and fluoxetine to enhance motor recovery by day 42 post-stroke, while atomoxetine being more potent considering its lower effective dose. Importantly, limited amount of wheel running alone was insufficient to promote motor recovery in mice, but was critical to augment the effect of high dose atomoxetine or fluoxetine. Likewise, high-dose drug treatments alone were insufficient to enhance functional recovery in the experimental animals. This synergetic effect of drug treatment plus physiotherapy is of great interest, and our ongoing pharmacological, molecular-genetic and biochemical studies should provide details about the molecular mechanism(s) responsible for this observation. It is unlikely that one mechanism is responsible for the observed effects, but rather multiple mechanisms involving enhanced axonal sprouting and synaptogenesis, angiogenesis and neurogenesis are likely involved it the observed effects. Notably, these processes are intertwined and inherently linked to neural repair and post-stroke recovery (Carmichael, 2016). Physical exercise (Christie et al., 2008; van Praag, 2008; Liu and Nusslock, 2018) as well as noradrenergic (Marzo et al., 2009; Tully and Bolshakov, 2010) and serotonergic (Lesch and Waider, 2012; Sobrido-Camean et al., 2018) neurotransmission are well-known to modulate these processes. Furthermore, fluoxetine has been shown to potentiate brain plasticity processes upon continuous use in numerous experimental studies (Levy et al., 2019; Song et al., 2019; Steinzeig et al., 2019), and there is some evidence for similar effects with chronic treatment of atomoxetine (Fumagalli et al., 2010; Pina et al., 2020).
In this study, we also evaluated the expression levels of GAP-43, synaptophysin, and PSD-95 in the peri-infarct cortical samples of experimental animals collected on post-stroke day 42. These molecular targets maybe causally associated with neural repair mechanisms (Wang et al., 2019), however, we did not observe significant differences in their expression levels among experimental groups (data not shown). In addition, we evaluated the number of parvalbumin-expressing neurons in the AGm area (medial premotor cortex) of ipsi- and contralateral hemispheres of the experimental animals. Parvalbumin-expressing neurons in AGm are primarily GABAergic inhibitory interneurons, and their reduced excitability and/or number is causally linked to motor learning and functional recovery after stroke (Zeiler et al., 2013; Ng et al., 2015). Notably, our results are in line with these observations indicating statistically significant reduction of parvalbumin-expressing neurons in ipsilateral AGm area of high-dose atomoxetine or fluoxetine-treated animals but not other experimental groups.
It is noteworthy, that fluoxetine has been used in several clinical trials for stroke recovery, including well-publicized FLAME (FLuoxetine on motor rehabilitation After ischeMic strokE) and FOCUS (Fluoxetine Or Control Under Supervision) trials (Chollet et al., 2011; Collaboration, 2019). FLAME study suggested that fluoxetine improves motor recovery in post-stroke patients, whereas FOCUS study found no significant difference in motor recovery of patients receiving fluoxetine and placebo. Among notable differences in these two clinical trials, which, in our opinion, are the likely reasons for the conflicting results are the lack of regular physical rehabilitation and use of a less rigorous/quantitative method (i.e., modified Rankin scale) for functional assessment of patients in FOCUS study. Unfortunately, a very similar study design and the same deficiencies were present in newly completed Assessment oF FluoxetINe In sTroke recoverY (AFFINITY) (Collaboration, 2020a) and Efficacy oF Fluoxetine-a randomisEd Controlled Trial in Stroke (EFFECTS) (Collaboration, 2020b) clinical trials, which again did not find beneficial effects of fluoxetine therapy for post-stroke recovery.
The relevance of our study to these contradicting reports is the finding that combination of fluoxetine with physical rehabilitation is necessary for motor recovery of mice after stroke. In fact, in addition to FLAME trial, two smaller and less publicized clinical studies also combined fluoxetine therapy with physical rehabilitation and documented improved motor function in stroke patients (Guo et al., 2016; Asadollahi et al., 2018). Collectively, our observations and the results of these clinical studies highlight the main deficiency of FOCUS, AFFINITY and EFFECTS trials, and suggest the need for combining fluoxetine therapy with physical rehabilitation in a future large clinical trial, before moving away from this drug (Kwakkel et al., 2020).
Contrary to fluoxetine, little is known about atomoxetine in a stroke setting. Only two pilot studies with handful of stroke patients were conducted so far, reporting improvement of motor function with atomoxetine therapy (Kinoshita et al., 2016; Ward et al., 2017), whereas another study reported improvement of post-stroke aphasia with atomoxetine therapy (Yamada et al., 2016).
In summary, our results indicate that pharmacological modulation of noradrenergic or serotonergic systems with atomoxetine and fluoxetine, in combination with limited voluntary running, promotes motor recovery in mice after ischemic stroke. This is the first study to document the potential of atomoxetine to facilitate post-stroke functional recovery in a pre-clinical setting. Our observations point out that enhanced noradrenergic neurotransmission by atomoxetine is likely more effective in facilitating post-stroke recovery than enhanced serotonergic neurotransmission by fluoxetine. This question is a subject of our ongoing research, which also aims to reveal the molecular signaling pathways and potential link between these two neurotransmitter systems in mechanisms related to functional recovery after stroke. Importantly, several published studies suggest that norepinephrine maybe more central in modulating neural repair mechanisms than serotonin. For example, norepinephrine but not serotonin appears to directly activate neural precursor cells (Jhaveri et al., 2010) and modulate long-term potentiation (Stanton and Sarvey, 1985). Another study suggested that noradrenergic neurotransmission maybe involved in noradrenergic and serotonergic axonal regeneration, whereas serotonergic neurotransmission only for serotonergic axonal regrowth (Liu et al., 2003). Lastly, one more study documented that exercise-induced BDNF upregulation is modulated by noradrenergic but not serotonergic neurotransmission (Garcia et al., 2003).
It is noteworthy, that our study has key limitations, including the use of adult male but not female and older animals, a need to determine the optimal time-window of pharmacotherapy and whether physiotherapy should be continuous to maintain improved function, and extension of studies beyond 6 weeks. Another important question to address experimentally is the extent of physiotherapy, i.e. its threshold, below which the added pharmacotherapy does not promote functional recovery. If further validated in larger studies, our observations suggest that add-on atomoxetine or fluoxetine therapy coupled with limited physical rehabilitation could offer therapeutic modality for stroke survivors who have difficulty to engage in early, high-intensity physiotherapy.
Footnotes
C-Editors; Zhao M, Li CH; T-Editor: Jia Y
Conflicts of interest: The authors declare no commercial conflict of interest.
Financial support: This work was partly supported by a National Institutes of Health Research Grant to VTK (1R01NS106879).
Institutional review board statement: This study was approved by the Texas Tech University Health Sciences Center Institutional Animal Care and Use Committee (protocol # 16019).
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Funding: This work was partly supported by a National Institutes of Health Research Grant to VTK (1R01NS106879).
References
- 1.Al Shoyaib A, Archie SR, Karamyan VT. Intraperitoneal route of drug administration: should it be used in experimental animal studies. Pharm Res. 2019;37:12. doi: 10.1007/s11095-019-2745-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Alamri FF, Shoyaib AA, Biggers A, Jayaraman S, Guindon J, Karamyan VT. Applicability of the grip strength and automated von Frey tactile sensitivity tests in the mouse photothrombotic model of stroke. Behav Brain Res. 2018;336:250–255. doi: 10.1016/j.bbr.2017.09.008. [DOI] [PubMed] [Google Scholar]
- 3.Asadollahi M, Ramezani M, Khanmoradi Z, Karimialavijeh E. The efficacy comparison of citalopram, fluoxetine, and placebo on motor recovery after ischemic stroke: a double-blind placebo-controlled randomized controlled trial. Clin Rehabil. 2018;32:1069–1075. doi: 10.1177/0269215518777791. [DOI] [PubMed] [Google Scholar]
- 4.Bernhardt J, Godecke E, Johnson L, Langhorne P. Early rehabilitation after stroke. Curr Opin Neurol. 2017a;30:48–54. doi: 10.1097/WCO.0000000000000404. [DOI] [PubMed] [Google Scholar]
- 5.Bernhardt J, Hayward KS, Kwakkel G, Ward NS, Wolf SL, Borschmann K, Krakauer JW, Boyd LA, Carmichael ST, Corbett D, Cramer SC. Agreed definitions and a shared vision for new standards in stroke recovery research: The Stroke Recovery and Rehabilitation Roundtable taskforce. Int J Stroke. 2017b;12:444–450. doi: 10.1177/1747493017711816. [DOI] [PubMed] [Google Scholar]
- 6.Bernhardt J, Borschmann K, Boyd L, Thomas Carmichael S, Corbett D, Cramer SC, Hoffmann T, Kwakkel G, Savitz SI, Saposnik G, Walker M, Ward N. Moving rehabilitation research forward: Developing consensus statements for rehabilitation and recovery research. Int J Stroke. 2016;11:454–458. doi: 10.1177/1747493016643851. [DOI] [PubMed] [Google Scholar]
- 7.Bernhardt J, Hayward KS, Dancause N, Lannin NA, Ward NS, Nudo RJ, Farrin A, Churilov L, Boyd LA, Jones TA, Carmichael ST, Corbett D, Cramer SC. A stroke recovery trial development framework: Consensus-based core recommendations from the Second Stroke Recovery and Rehabilitation Roundtable. Int J Stroke. 2019;14:792–802. doi: 10.1177/1747493019879657. [DOI] [PubMed] [Google Scholar]
- 8.Birjandi SZ, Abduljawad N, Nair S, Dehghani M, Suzuki K, Kimura H, Carmichael ST. Phosphodiesterase 10A inhibition leads to brain region-specific recovery based on stroke type. Transl Stroke Res. 2020 doi: 10.1007/s12975-020-00819-8. doi: 101007/s12975-020-00819-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Borodovitsyna O, Flamini M, Chandler D. Noradrenergic modulation of cognition in health and disease. Neural Plast. 2017;2017:6031478. doi: 10.1155/2017/6031478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Carmichael ST. Emergent properties of neural repair: elemental biology to therapeutic concepts. Ann Neurol. 2016;79:895–906. doi: 10.1002/ana.24653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chollet F, Tardy J, Albucher JF, Thalamas C, Berard E, Lamy C, Bejot Y, Deltour S, Jaillard A, Niclot P, Guillon B, Moulin T, Marque P, Pariente J, Arnaud C, Loubinoux I. Fluoxetine for motor recovery after acute ischaemic stroke (FLAME): a randomised placebo-controlled trial. Lancet Neurol. 2011;10:123–130. doi: 10.1016/S1474-4422(10)70314-8. [DOI] [PubMed] [Google Scholar]
- 12.Christie BR, Eadie BD, Kannangara TS, Robillard JM, Shin J, Titterness AK. Exercising our brains: how physical activity impacts synaptic plasticity in the dentate gyrus. Neuromolecular Med. 2008;10:47–58. doi: 10.1007/s12017-008-8033-2. [DOI] [PubMed] [Google Scholar]
- 13.Christman AK, Fermo JD, Markowitz JS. Atomoxetine, a novel treatment for attention-deficit-hyperactivity disorder. Pharmacotherapy. 2004;24:1020–1036. doi: 10.1592/phco.24.11.1020.36146. [DOI] [PubMed] [Google Scholar]
- 14.Clarkson AN, Huang BS, Macisaac SE, Mody I, Carmichael ST. Reducing excessive GABA-mediated tonic inhibition promotes functional recovery after stroke. Nature. 2010;468:305–309. doi: 10.1038/nature09511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Collaboration AT. Safety and efficacy of fluoxetine on functional outcome after acute stroke (AFFINITY): a randomised ,double-blind, placebo-controlled trial. Lancet Neurol. 2020a;19:651–660. doi: 10.1016/S1474-4422(20)30207-6. [DOI] [PubMed] [Google Scholar]
- 16.Collaboration ET. Safety and efficacy of fluoxetine on functional recovery after acute stroke (EFFECTS): a randomised, double-blind, placebo-controlled trial. Lancet Neurol. 2020b;19:661–669. doi: 10.1016/S1474-4422(20)30219-2. [DOI] [PubMed] [Google Scholar]
- 17.Collaboration FT. Effects of fluoxetine on functional outcomes after acute stroke (FOCUS): a pragmatic, double-blind, randomised, controlled trial. Lancet. 2019;393:265–274. doi: 10.1016/S0140-6736(18)32823-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Corbett D, Carmichael ST, Murphy TH, Jones TA, Schwab ME, Jolkkonen J, Clarkson AN, Dancause N, Weiloch T, Johansen-Berg H, Nilsson M, McCullough LD, Joy MT. Enhancing the alignment of the preclinical and clinical stroke recovery research pipeline: Consensus-based core recommendations from the Stroke Recovery and Rehabilitation Roundtable translational working group. Int J Stroke. 2017;12:462–471. doi: 10.1177/1747493017711814. [DOI] [PubMed] [Google Scholar]
- 19.Cramer SC. Drugs to enhance motor recovery after stroke. Stroke. 2015;46:2998–3005. doi: 10.1161/STROKEAHA.115.007433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Egan KJ, Janssen H, Sena ES, Longley L, Speare S, Howells DW, Spratt NJ, Macleod MR, Mead GE, Bernhardt J. Exercise reduces infarct volume and facilitates neurobehavioral recovery: results from a systematic review and meta-analysis of exercise in experimental models of focal ischemia. Neurorehabil Neural Repair. 2014;28:800–812. doi: 10.1177/1545968314521694. [DOI] [PubMed] [Google Scholar]
- 21.Fini NA, Holland AE, Keating J, Simek J, Bernhardt J. How physically active are people following stroke. Systematic review and quantitative synthesis. Phys Ther. 2017;97:707–717. doi: 10.1093/ptj/pzx038. [DOI] [PubMed] [Google Scholar]
- 22.Fumagalli F, Cattaneo A, Caffino L, Ibba M, Racagni G, Carboni E, Gennarelli M, Riva MA. Sub-chronic exposure to atomoxetine up-regulates BDNF expression and signalling in the brain of adolescent spontaneously hypertensive rats: comparison with methylphenidate. Pharmacol Res. 2010;62:523–529. doi: 10.1016/j.phrs.2010.07.009. [DOI] [PubMed] [Google Scholar]
- 23.Garcia C, Chen MJ, Garza AA, Cotman CW, Russo-Neustadt A. The influence of specific noradrenergic and serotonergic lesions on the expression of hippocampal brain-derived neurotrophic factor transcripts following voluntary physical activity. Neuroscience. 2003;119:721–732. doi: 10.1016/s0306-4522(03)00192-1. [DOI] [PubMed] [Google Scholar]
- 24.Guo Y, He Y, Tang B, Ma K, Cai Z, Zeng S, Zhang Y, Jiang X. Effect of using fluoxetine at different time windows on neurological functional prognosis after ischemic stroke. Restor Neurol Neurosci. 2016;34:177–187. doi: 10.3233/RNN-150535. [DOI] [PubMed] [Google Scholar]
- 25.Hatakeyama M, Ninomiya I, Kanazawa M. Angiogenesis and neuronal remodeling after ischemic stroke. Neural Regen Res. 2020;15:16–19. doi: 10.4103/1673-5374.264442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hodes GE, Hill-Smith TE, Lucki I. Fluoxetine treatment induces dose dependent alterations in depression associated behavior and neural plasticity in female mice. Neurosci Lett. 2010;484:12–16. doi: 10.1016/j.neulet.2010.07.084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hu Q, Liu L, Zhou L, Lu H, Wang J, Chen X, Wang Q. Effect of fluoxetine on HIF-1alpha- Netrin/VEGF cascade, angiogenesis and neuroprotection in a rat model of transient middle cerebral artery occlusion. Exp Neurol. 2020;329:113312. doi: 10.1016/j.expneurol.2020.113312. [DOI] [PubMed] [Google Scholar]
- 28.Jayaraman S, Al Shoyaib A, Kocot J, Villalba H, Alamri FF, Rashid M, Wangler NJ, Chowdhury EA, German N, Arumugam TV, Abbruscato TJ, Karamyan VT. Peptidase neurolysin functions to preserve the brain after ischemic stroke in male mice. J Neurochem. 2020;153:120–137. doi: 10.1111/jnc.14864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jhaveri DJ, Mackay EW, Hamlin AS, Marathe SV, Nandam LS, Vaidya VA, Bartlett PF. Norepinephrine directly activates adult hippocampal precursors via beta3-adrenergic receptors. J Neurosci. 2010;30:2795–2806. doi: 10.1523/JNEUROSCI.3780-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Karamyan VT. The role of peptidase neurolysin in neuroprotection and neural repair after stroke. Neural Regen Res. 2021;16:21–25. doi: 10.4103/1673-5374.284904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kinoshita S, Kakuda W, Yamada N, Momosaki R, Okuma R, Watanabe S, Abo M. Therapeutic administration of atomoxetine combined with rTMS and occupational therapy for upper limb hemiparesis after stroke: a case series study of three patients. Acta Neurol Belg. 2016;116:31–37. doi: 10.1007/s13760-015-0503-3. [DOI] [PubMed] [Google Scholar]
- 32.Klenotich SJ, Seiglie MP, McMurray MS, Roitman JD, Le Grange D, Dugad P, Dulawa SC. Olanzapine, but not fluoxetine, treatment increases survival in activity-based anorexia in mice. Neuropsychopharmacology. 2012;37:1620–1631. doi: 10.1038/npp.2012.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Koda K, Ago Y, Cong Y, Kita Y, Takuma K, Matsuda T. Effects of acute and chronic administration of atomoxetine and methylphenidate on extracellular levels of noradrenaline, dopamine and serotonin in the prefrontal cortex and striatum of mice. J Neurochem. 2010;114:259–270. doi: 10.1111/j.1471-4159.2010.06750.x. [DOI] [PubMed] [Google Scholar]
- 34.Krakauer JW, Carmichael ST, Corbett D, Wittenberg GF. Getting neurorehabilitation right: what can be learned from animal models. Neurorehabil Neural Repair. 2012;26:923–931. doi: 10.1177/1545968312440745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kraus C, Castren E, Kasper S, Lanzenberger R. Serotonin and neuroplasticity - Links between molecular, functional and structural pathophysiology in depression. Neurosci Biobehav Rev. 2017;77:317–326. doi: 10.1016/j.neubiorev.2017.03.007. [DOI] [PubMed] [Google Scholar]
- 36.Kwakkel G, Meskers C, Ward NS. Time for the next stage of stroke recovery trials. Lancet Neurol. 2020;19:636–637. doi: 10.1016/S1474-4422(20)30218-0. [DOI] [PubMed] [Google Scholar]
- 37.Lay S, Bernhardt J, West T, Churilov L, Dart A, Hayes K, Cumming TB. Is early rehabilitation a myth. Physical inactivity in the first week after myocardial infarction and stroke. Disabil Rehabil. 2016;38:1493–1499. doi: 10.3109/09638288.2015.1106598. [DOI] [PubMed] [Google Scholar]
- 38.Ledbetter M. Atomoxetine: a novel treatment for child and adult ADHD. Neuropsychiatr Dis Treat. 2006;2:455–466. doi: 10.2147/nedt.2006.2.4.455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lenz M, Vlachos A, Maggio N. Ischemic long-term-potentiation (iLTP): perspectives to set the threshold of neural plasticity toward therapy. Neural Regen Res. 2015;10:1537–1539. doi: 10.4103/1673-5374.165215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lesch KP, Waider J. Serotonin in the modulation of neural plasticity and networks: implications for neurodevelopmental disorders. Neuron. 2012;76:175–191. doi: 10.1016/j.neuron.2012.09.013. [DOI] [PubMed] [Google Scholar]
- 41.Levy MJF, Boulle F, Emerit MB, Poilbout C, Steinbusch HWM, Van den Hove DLA, Kenis G, Lanfumey L. 5-HTT independent effects of fluoxetine on neuroplasticity. Sci Rep. 2019;9:6311. doi: 10.1038/s41598-019-42775-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Li WL, Cai HH, Wang B, Chen L, Zhou QG, Luo CX, Liu N, Ding XS, Zhu DY. Chronic fluoxetine treatment improves ischemia-induced spatial cognitive deficits through increasing hippocampal neurogenesis after stroke. J Neurosci Res. 2009;87:112–122. doi: 10.1002/jnr.21829. [DOI] [PubMed] [Google Scholar]
- 43.Liu PZ, Nusslock R. Exercise and hippocampal neurogenesis: a dogma re-examined and lessons learned. Neural Regen Res. 2018;13:1354–1355. doi: 10.4103/1673-5374.235225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liu Y, Ishida Y, Shinoda K, Nakamura S. Interaction between serotonergic and noradrenergic axons during axonal regeneration. Exp Neurol. 2003;184:169–178. doi: 10.1016/s0014-4886(03)00221-8. [DOI] [PubMed] [Google Scholar]
- 45.Lopez-Munoz F, Alamo C. Monoaminergic neurotransmission: the history of the discovery of antidepressants from 1950s until today. Curr Pharm Des. 2009;15:1563–1586. doi: 10.2174/138161209788168001. [DOI] [PubMed] [Google Scholar]
- 46.Marzo A, Bai J, Otani S. Neuroplasticity regulation by noradrenaline in mammalian brain. Curr Neuropharmacol. 2009;7:286–295. doi: 10.2174/157015909790031193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Moon SJ, Kim CJ, Lee YJ, Hong M, Han J, Bahn GH. Effect of atomoxetine on hyperactivity in an animal model of attention-deficit/hyperactivity disorder (ADHD) PLoS One. 2014;9:e108918. doi: 10.1371/journal.pone.0108918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Murphy TH, Corbett D. Plasticity during stroke recovery: from synapse to behaviour. Nat Rev Neurosci. 2009;10:861–872. doi: 10.1038/nrn2735. [DOI] [PubMed] [Google Scholar]
- 49.Ng KL, Gibson EM, Hubbard R, Yang J, Caffo B, O’Brien RJ, Krakauer JW, Zeiler SR. Fluoxetine maintains a state of heightened responsiveness to motor training early after stroke in a mouse model. Stroke. 2015;46:2951–2960. doi: 10.1161/STROKEAHA.115.010471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Nicholson S, Sniehotta FF, van Wijck F, Greig CA, Johnston M, McMurdo ME, Dennis M, Mead GE. A systematic review of perceived barriers and motivators to physical activity after stroke. Int J Stroke. 2013;8:357–364. doi: 10.1111/j.1747-4949.2012.00880.x. [DOI] [PubMed] [Google Scholar]
- 51.Pina R, Rozas C, Contreras D, Hardy P, Ugarte G, Zeise ML, Rojas P, Morales B. Atomoxetine reestablishes long term potentiation in a mouse model of attention deficit/hyperactivity disorder. Neuroscience. 2020;439:268–274. doi: 10.1016/j.neuroscience.2019.10.040. [DOI] [PubMed] [Google Scholar]
- 52.Rashid M, Wangler NJ, Yang L, Shah K, Arumugam TV, Abbruscato TJ, Karamyan VT. Functional up-regulation of endopeptidase neurolysin during post-acute and early recovery phases of experimental stroke in mouse brain. J Neurochem. 2014;129:179–189. doi: 10.1111/jnc.12513. [DOI] [PubMed] [Google Scholar]
- 53.Rossi A, Barraco A, Donda P. Fluoxetine: a review on evidence based medicine. Ann Gen Hosp Psychiatry. 2004;3:2. doi: 10.1186/1475-2832-3-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Rossi FM. Analysis of fluoxetine-induced plasticity mechanisms as a strategy for understanding plasticity related neural disorders. Neural Regen Res. 2016;11:547–548. doi: 10.4103/1673-5374.180731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sobrido-Camean D, Rodicio MC, Barreiro-Iglesias A. Serotonin controls axon and neuronal regeneration in the nervous system: lessons from regenerating animal models. Neural Regen Res. 2018;13:237–238. doi: 10.4103/1673-5374.226387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Song T, Wu H, Li R, Xu H, Rao X, Gao L, Zou Y, Lei H. Repeated fluoxetine treatment induces long-lasting neurotrophic changes in the medial prefrontal cortex of adult rats. Behav Brain Res. 2019;365:114–124. doi: 10.1016/j.bbr.2019.03.009. [DOI] [PubMed] [Google Scholar]
- 57.Stanton PK, Sarvey JM. Depletion of norepinephrine, but not serotonin, reduces long-term potentiation in the dentate gyrus of rat hippocampal slices. J Neurosci. 1985;5:2169–2176. doi: 10.1523/JNEUROSCI.05-08-02169.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Steinzeig A, Cannarozzo C, Castren E. Fluoxetine-induced plasticity in the visual cortex outlasts the duration of the naturally occurring critical period. Eur J Neurosci. 2019;50:3663–3673. doi: 10.1111/ejn.14512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sun X, Zhou Z, Liu T, Zhao M, Zhao S, Xiao T, Jolkkonen J, Zhao C. Fluoxetine enhances neurogenesis in aged rats with cortical infarcts, but this is not reflected in a behavioral recovery. J Mol Neurosci. 2016;58:233–242. doi: 10.1007/s12031-015-0662-y. [DOI] [PubMed] [Google Scholar]
- 60.Syeara N, Alamri FF, Jayaraman S, Lee P, Karamyan ST, Arumugam TV, Karamyan VT. Motor deficit in the mouse ferric chloride-induced distal middle cerebral artery occlusion model of stroke. Behav Brain Res. 2019;380:112418. doi: 10.1016/j.bbr.2019.112418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Trout AL, Kahle MP, Roberts JM, Marcelo A, de Hoog L, Boychuk JA, Grupke SL, Berretta A, Gowing EK, Boychuk CR, Gorman AA, Edwards DN, Rutkai I, Biose IJ, Ishibashi-Ueda H, Ihara M, Smith BN, Clarkson AN, Bix GJ. Perlecan domain-V Enhances neurogenic brain repair after stroke in mice. Transl Stroke Res. 2020 doi: 10.1007/s12975-020-00800-5. doi: 101007/s12975-020-00800-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tsintou M, Dalamagkas K, Makris N. Taking central nervous system regenerative therapies to the clinic: curing rodents versus nonhuman primates versus humans. Neural Regen Res. 2020;15:425–437. doi: 10.4103/1673-5374.266048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Tully K, Bolshakov VY. Emotional enhancement of memory: how norepinephrine enables synaptic plasticity. Mol Brain. 2010;3:15. doi: 10.1186/1756-6606-3-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Vahid-Ansari F, Albert PR. Chronic fluoxetine induces activity changes in recovery from poststroke anxiety, depression, and cognitive impairment. Neurotherapeutics. 2018;15:200–215. doi: 10.1007/s13311-017-0590-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.van Praag H. Neurogenesis and exercise: past and future directions. Neuromolecular Med. 2008;10:128–140. doi: 10.1007/s12017-008-8028-z. [DOI] [PubMed] [Google Scholar]
- 66.Wang CJ, Wu Y, Zhang Q, Yu KW, Wang YY. An enriched environment promotes synaptic plasticity and cognitive recovery after permanent middle cerebral artery occlusion in mice. Neural Regen Res. 2019;14:462–469. doi: 10.4103/1673-5374.245470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wangler NJ, Santos KL, Schadock I, Hagen FK, Escher E, Bader M, Speth RC, Karamyan VT. Identification of membrane-bound variant of metalloendopeptidase neurolysin (EC 3.42416) as the non-angiotensin type 1 (Non-AT1), Non-AT2 angiotensin binding site. J Biol Chem. 2012;287:114–122. doi: 10.1074/jbc.M111.273052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ward A, Carrico C, Powell E, Westgate PM, Nichols L, Fleischer A, Sawaki L. Safety and improvement of movement function after stroke with atomoxetine: A pilot randomized trial. Restor Neurol Neurosci. 2017;35:1–10. doi: 10.3233/RNN-160673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Windle V, Corbett D. Fluoxetine and recovery of motor function after focal ischemia in rats. Brain Res. 2005;1044:25–32. doi: 10.1016/j.brainres.2005.02.060. [DOI] [PubMed] [Google Scholar]
- 70.Winstein CJ, Stein J, Arena R, Bates B, Cherney LR, Cramer SC, Deruyter F, Eng JJ, Fisher B, Harvey RL, Lang CE, MacKay-Lyons M, Ottenbacher KJ, Pugh S, Reeves MJ, Richards LG, Stiers W, Zorowitz RD American Heart Association Stroke Council, Council on Cardiovascular and Stroke Nursing, Council on Clinical Cardiology and Council on Quality of Care and Outcomes Research. Guidelines for adult stroke rehabilitation and recovery: a guideline for healthcare professionals from the American Heart Association/American Stroke Association. Stroke. 2016;47:e98–169. doi: 10.1161/STR.0000000000000098. [DOI] [PubMed] [Google Scholar]
- 71.Wong DT, Bymaster FP, Engleman EA. Prozac (fluoxetine, Lilly 110140), the first selective serotonin uptake inhibitor and an antidepressant drug: twenty years since its first publication. Life Sci. 1995;57:411–441. doi: 10.1016/0024-3205(95)00209-o. [DOI] [PubMed] [Google Scholar]
- 72.Yamada N, Kakuda W, Yamamoto K, Momosaki R, Abo M. Atomoxetine administration combined with intensive speech therapy for post-stroke aphasia: evaluation by a novel SPECT method. Int J Neurosci. 2016;126:829–838. doi: 10.3109/00207454.2015.1074226. [DOI] [PubMed] [Google Scholar]
- 73.Yang YW, Pan WX, Xie Q. Combined effect of repetitive transcranial magnetic stimulation and physical exercise on cortical plasticity. Neural Regen Res. 2020;15:1986–1994. doi: 10.4103/1673-5374.282239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zeiler SR, Gibson EM, Hoesch RE, Li MY, Worley PF, O’Brien RJ, Krakauer JW. Medial premotor cortex shows a reduction in inhibitory markers and mediates recovery in a mouse model of focal stroke. Stroke. 2013;44:483–489. doi: 10.1161/STROKEAHA.112.676940. [DOI] [PMC free article] [PubMed] [Google Scholar]