

Abstract
Combining synthetic biology with adoptive T cell transfer has led to promising advances in the treatment of relapsed refractory B-cell acute lymphoblastic leukemia (R/R B-ALL), diffuse large B-cell lymphoma (DLBCL), and mantle cell lymphoma (MCL). Chimeric antigen receptors (CARs) are synthetic receptors that redirect T cell specificity against cancer. CARs include “built-in” signaling domains that reprogram T cell metabolism, enhance effector function, and support long-term persistence. Despite their success in blood-based malignancies, relapse can occur in CD19-redirected CAR T cell therapies for several reasons including poor engraftment, impaired in vivo proliferation, and T cell senescence. Herein, we explain how subtle alterations in CAR design may overcome barriers to effective adoptive immunotherapy. We also discuss how the physiochemical properties of the single-chain variable fragment (scFv) impact differentiation and persistence. Moreover, we describe innovative advances in CAR engineering and provide insight into the development of humanized scFvs whose proposed benefits include increased persistence and improved clinical outcomes. Tumor cells can evade detection and CAR-mediated elimination due to the emergence or presence of CD19-negative leukemic cell subpopulations. We discuss the opportunities and challenges in targeting other B-ALL-associated antigens. Identifying alternate targets is fundamentally necessary to restore the success of CAR T-cell therapies in CD19-negative B-ALL patients.
Keywords: Chimeric antigen receptor, Acute lymphoblastic leukemia, CD19, Cancer immunotherapy, Adoptive cell therapy, Humanization
1. Introduction
B-cell acute lymphoblastic leukemia (B-ALL) is a hematologic malignancy characterized by the uncontrolled proliferation of progenitor B cells in the bone marrow. B-ALL is particularly insidious in children as well as young adults. Standard treatment regimens for B-ALL include chemotherapy, radiation, and hematopoietic stem cell transplantation. While the five-year overall survival rate is 80–90%, disease recurrence (relapse) can occur and is characterized by a poor response to conventional treatments (refractory). This lack of durable efficacy underscores the need for alternate approaches with enhanced effectiveness against cancer (1,2). Improved outcomes in patients with relapsed or refractory (R/R) B-ALL have been obtained through the use of adoptively transferred T cells. Using advanced principles of synthetic biology, T cells can be genetically reprogramed to recognize and lyse malignant cells in a specific manner. Chimeric antigen receptor (CAR) transgenes have been effectively used to generate tumor-reactive T cells. The unprecedented successes of CAR-expressing T cells have given rise to a succession of US Food and Drug Administration (FDA) approvals including Kymriah™ (Tisagenlecleucel) for pediatric patients and young adults with R/R B-ALL, Yescarta™ (Axicabtagene ciloleucel) and Breyanzi™ (Lisocabtagene maraleucel) for patients with diffuse large B-cell lymphoma (DLBCL), and Tecartus™ (Brexucabtagene autoleucel) for adults with mantle cell lymphoma (MCL) (2,3).
Adoptive T cell therapies (ACT) involve an ordered process whereby patient T cells are isolated and expanded ex vivo in nutrient-rich conditions over 9–14 days before re-infusion (4,5). During this process, T cells are often genetically modified with a transgene to selectively direct their cytolytic activity towards antigens expressed on the surface of tumor cells, known as tumor-associated antigens (TAA) or tumor-specific antigens (TSA). Using viral-mediated gene delivery, transgenes such as CARs are increasingly used not just to redirect T cell specificity but also to enhance effector function, reprogram metabolism, and improve overall persistence in B-ALL. In this review, we provide mechanistic insights into the multi-dimensional benefit of CARs, illuminating their important role against cancer. We will also discuss strategies, explicitly relevant to 1) CAR design, and 2) the ex vivo expansion phase, that support the generation of CAR T cells with enhanced durable efficacy.
2. CAR design and structure
CARs are modular polypeptides encoding 3 canonical components. The CAR ectodomain contains an extracellular antigen-binding domain which is essential for tumor reactivity (6). This antigen-recognition domain is usually derived from a monoclonal antibody single-chain variable fragment (scFv) (6). The scFv contains heavy and light chain moieties which are connected by a linker region and anchored via a hinge region to the plasma membrane. The scFv is fused to an intracellular costimulatory signaling domain expressed in tandem with the CD3ζ chain from the T cell receptor (TCR) (Fig. 1) (7). There is renewed interest in understanding how the physiochemical properties of these individual components, without any modification to the antigen-binding region, can influence the efficacy of the modified T cells. For this reason, we will discuss how subtle modifications in CAR design and structure influence its overall function.
Figure 1.
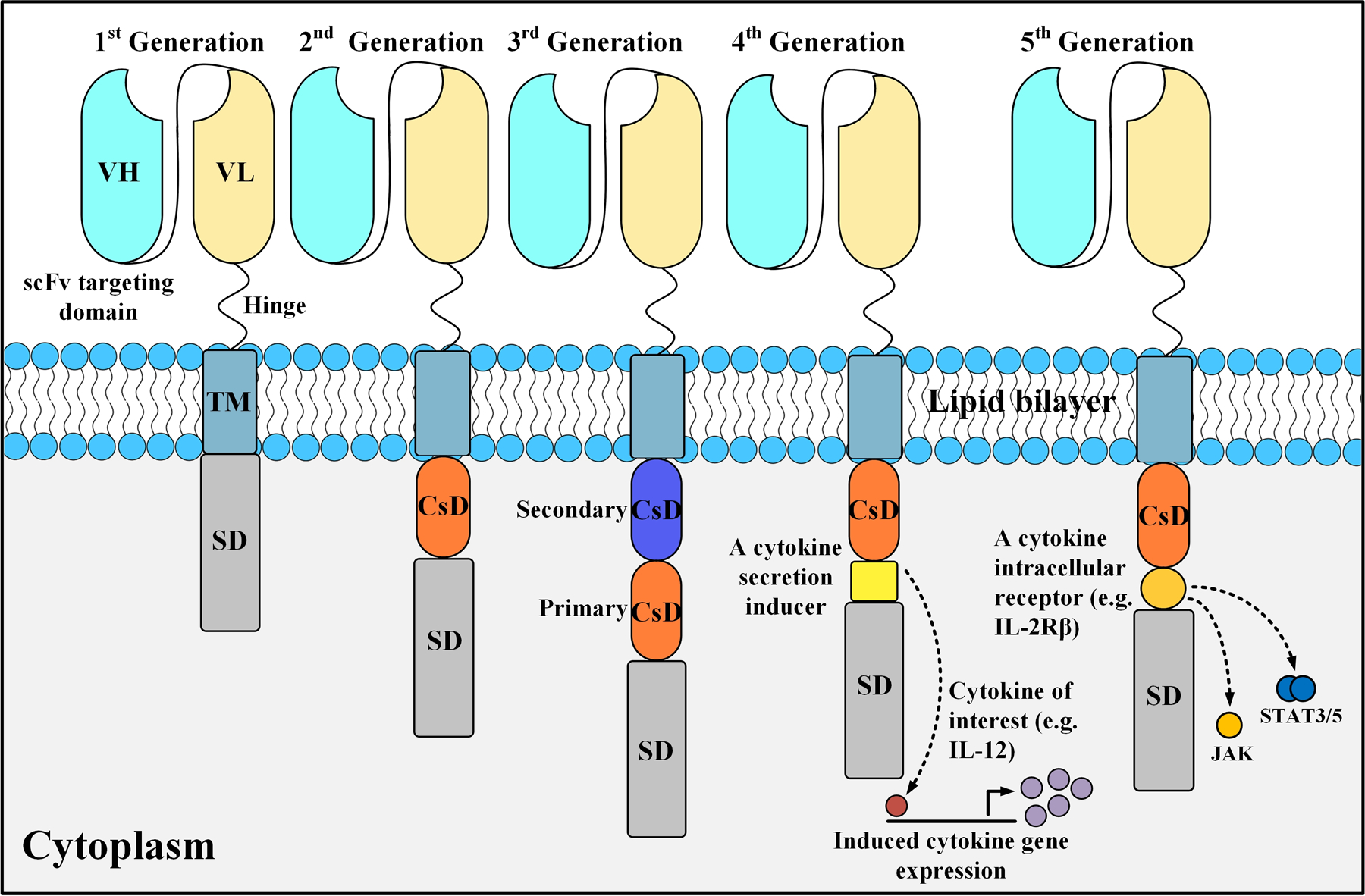
A detailed anatomy of different CAR generations. First-generation CARs contain only a primary stimulation domain (e.g. CD3ζ). Second-generation CARs harbor one costimulatory domain (CsD) whereas third-generation CARs have two CsDs (a primary CsD and a secondary CsD). Fourth-generation CARs, also known as T cell redirected for universal cytokine-mediated killing (TRUCKs) or armored CARs, are based on second-generation CARs and are paired with a cytokine secretion inducer (e.g. IL-12). Fifth-generation CARs are also tailored versions of second-generation CARs as they contain an intracellular domain of a cytokine receptor (e.g. IL-2Rβ). SD, stimulation domain.
2.1. The extracellular domain
CARs combine two important aspects of an adaptive, cell-mediated immune response. In a humoral immune response, differentiated B cells recognize unique antigens through the production, and secretion, of antibodies. T cells also recognize unique antigens but do so through the assembly of a multi-dimensional T cell receptor complex. T cell activation occurs in a multi-step process: antigenic peptides are internalized by antigen-presenting cells (APCs), proteolytically processed, and presented to T cells in the context of major histocompatibility complexes (MHC). CARs combine the high selectivity of antibodies with T cell cytolytic function. By using scFvs as the antigen-binding moiety, CARs effectively redirect T cell cytolytic activity to a specific epitope found on tumor cells, in an MHC-independent manner.
The scFv used in Kymriah and Yescarta is derived from the FMC63 murine antibody that recognizes human CD19. FMC63 targets a specific epitope found on exon 4 of the CD19 gene (8). There are inherent limitations in the use of murine scFv-based CARs. Non-native scFvs may contain immunogenic epitopes that trigger an immune response. The potential for immunogenicity is increased when murine sequences are encoded in synthetic receptors, such as CARs, and expressed in human T cells. HLA-restricted T cell-mediated immune responses against specific epitopes in the FMC63 scFv have been reported (9,10). Importantly, the use of FMC63 has been increasingly recognized as an important parameter influencing CAR T cell engraftment in the treatment of B-ALL. Similarly, the use of murine-based scFvs such as the SS1 CAR has also been associated with decreased persistence and impaired clinical efficacy in T cell therapies against mesothelioma (11).
Antibody humanization is a useful strategy to overcome the inherent limitations of murine scFvs. To initiate this process, framework regions of the murine scFv are replaced with human coding sequences. We comprehensively describe the humanization process of a murine scFv and the proposed benefits of humanized CAR T cell therapies against cancer (Fig. 2).
Figure 2.
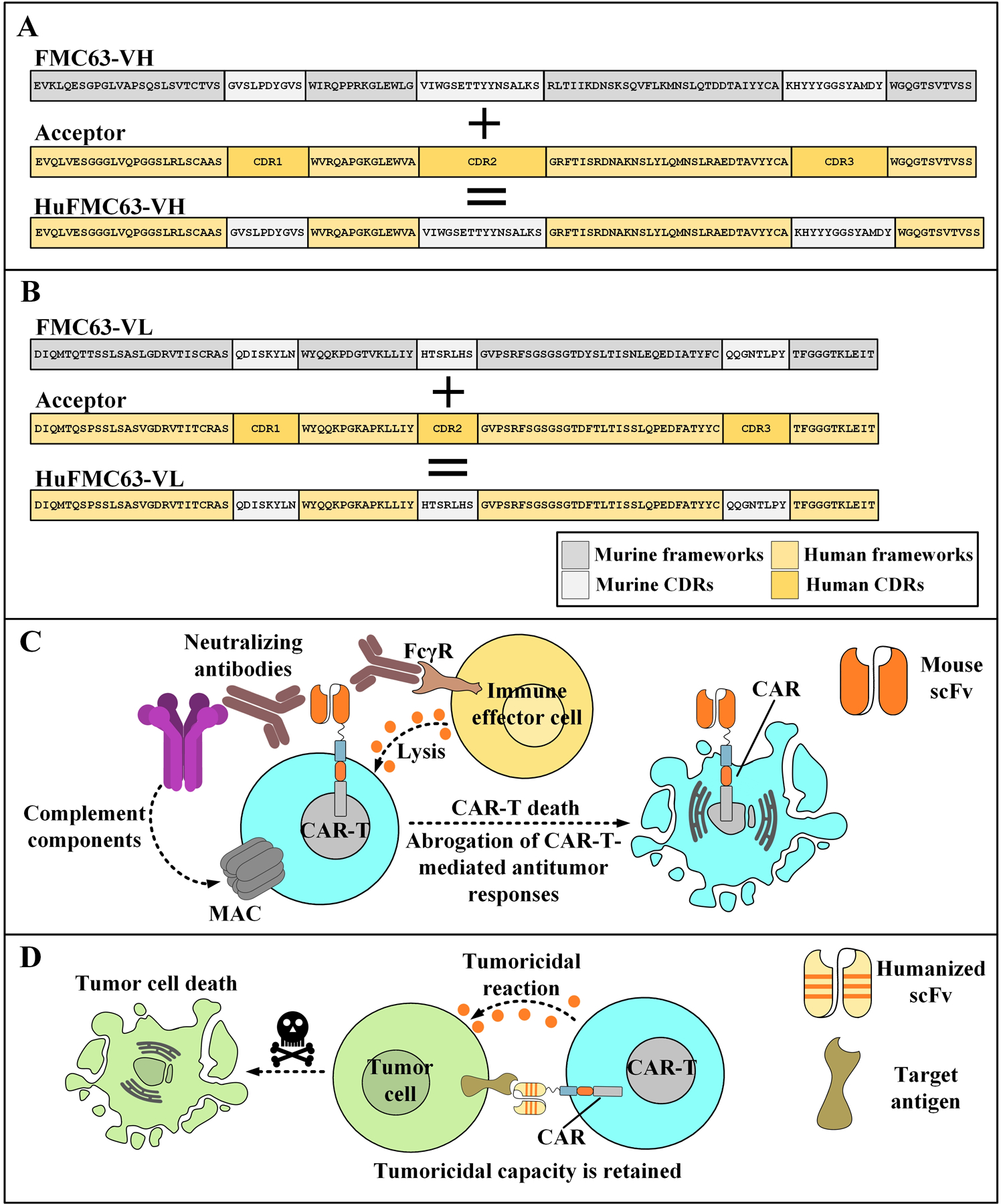
The humanization process of the murine CD19-specific scFv, FMC63, (as proposed by Qian et al. (117)) and the mechanism of action underlying the elimination of CAR T cells that harbor an animal-derived targeting domain. A: The CDRs of the FMC63-VH are grafted onto the framework regions of an acceptor human antibody with a high-rate of amino acid sequence similarity. In some cases, a few framework residues, such as those positioned in framework 2, might get back-mutated to the native animal residues since their substitutions are considered detrimental for the antigen affinity of the antibody. B: The CDR grafting of the FMC63-VL onto the acceptor frameworks of a human antibody. C: Neutralizing antibodies are produced by the immune system of the recipient against the murine-derived targeting domain of the CAR T cells leading to their elimination and subsequent dismantling of all CAR T cell-related tumoricidal responses. D: As opposed to the previous situation, CAR T cells that harbor humanized targeting domains are much less likely to encounter neutralizing antibodies formed against them interfering with their antitumor capacity. Therefore, prolonged antitumor responses are a possible scenario. VH, variable regions of the heavy chains; VL, variable regions of the light chains; CDR, complementarity-determining regions; HuFMC63-VH, humanized FMC63-VH; HuFMC63-VL, humanized FMC63-VL; FcγR, Fcγ receptor; scFv, single-chain variable fragment; MAC, membrane attack complex; CAR, chimeric antigen receptor.
During antibody humanization, sequence modifications are limited to the frameworks rather than CDRs. For this reason, the binding affinities of CARs engineered with a humanized scFv should be minimally affected. In support of this premise, Qian et al. showed that CARs designed with a humanized scFv supported T cell proliferation and cytokine production following co-culture with CD19+ target cell lines Nalm-6 and Daudi (12). In a human xenograft model of lymphoma, they showed that CAR T cells expressing a humanized scFv (hCARTs) displayed similar anti-tumor efficacy and superior survival as those of CAR T cells expressing murine scFv. These encouraging results advanced the use of humanized scFvs in preclinical models and clinical studies further.
Cao et al. evaluated the therapeutic efficacy of hCARTs clinically, following infusion into R/R ALL patients. Interestingly, a subset of patients in their trial had undergone prior treatment with the FMC63 murine-based CAR T cells (13). Based on tumor clearance from the bone marrow, central nervous system, and testis they showed that trafficking and overall cytolytic activity of hCARTs were superior to those of their murine-based derivatives (14). Conclusions from this trial show that hCARTs have a decreased potential for immunogenicity, improved therapeutic effectiveness, and enhanced persistence. Moreover, hCART19s can effectively treat pediatric or adult R/R CD19+ B-ALL, even following relapse from CARs engineered with the murine-based scFv.
In a separate clinical trial (NCT02349698), Heng et al. evaluated the efficacy and safety of CD19-redirected hCAR-Ts in R/R B-ALL patients. They reported enhanced long-term persistence of hCAR-Ts; exemplified by a complete remission (CR) in 60% of the treated patients (15). As persistence, correlates with enhanced durable efficacy, their study provides mechanistic insights into the beneficial attributes of hCAR-Ts; ultimately culminating in lower relapse rates as well as improved clinical impact in the treatment of R/R ALL patients. Collectively, these studies show that scFv humanization is an important optimization strategy to increase CAR T cell bioactivity in preclinical and clinical subjects. As survival is substantially influenced by engraftment, persistence, and durable efficacy, these findings encourage the humanization of other scFvs used in T cell-based therapies.
Generally, B-cell aplasia, as characterized by CAR T cell-mediated elimination of healthy CD19-expressing B cells, is an established hallmark of successful CAR T therapy (2). However, B-cell aplasia can result in hypogammaglobulinemia and increased risk of viral as well as bacterial infections (2). Such patients often require precise monitoring and (healthy donor-derived) plasma to complete their CAR T cell treatment. Following antigen engagement, activated CAR T cells increase IFNγ, TNF, and GM-CSF cytokine production; macrophages respond to this inflammatory cascade by secreting IL-1 and IL-6-collectively this process manifests as cytokine release syndrome (CRS) (2,16). Various therapeutic modalities have been developed and/or repurposed to counter the deleterious effects of CAR T cell-induced CRS and neurotoxicity, including the administration of tocilizumab, anakinra, lenzilumab, metyrosine, or dasatinib, all of which have shown promise in a case-dependent manner (2,16). Interestingly, CAR T cells can also be engineered to secrete neutralizing antibodies or even IL-6R- or GM-CSF-specific nanobodies to reduce the life-threatening effects of CRS and neurotoxicity (2,16). Unsurprisingly, there is a direct relationship between the severity of toxicity and the persistence of the infused cells. Since humanized hCARTs exhibit higher persistence relative to their murine counterparts, their toxicity burden must be followed closely. To our knowledge, no clinical study has yet compared the differences between the toxicities of these two CAR T cell platforms. In one study with eighteen R/R ALL patients, resolvable neurotoxicity was reported in 1 recipient while grade 1–2 and grade 3–5 CRS was observed in 13 and 4 recipients, respectively, after a single round of CD19-redirected hCART infusion (1 × 106/kg) (17). In a separate study (ChiCTR1800017401) comprising two previously-untreated B-ALL patients, both recipients showed signs of grade 1–2 CRS (for which tocilizumab or corticosteroids was not advised) after a single infusion of CD19-redirected hCARTs (1 × 106/kg) (18). In the Phase I trials ChiCTR1800014761 and ChiCTR1800017439, five B-ALL patients treated with CD19-redirected hCARTs, experienced mild adverse events (including fever and grade 1 CRS). However, the humanized version conferred a six-fold increase in affinity to CD19 (19) which likely impacted the magnitude, and rate, of the response independent of persistence. More research is necessary to fully understand how antibody humanization in the CAR format impacts the overall response and toxicity profiles.
2.2. Advanced CAR Design: Understanding how the physiochemical properties of the ectodomain influence CAR function
Nonvariable regions confined to the CAR ectodomain can overtly contribute to the overall biophysical balance, thermodynamic stability, and tertiary structure of the scFv. The CAR ectodomain provides 1) the necessary framework to anchor the scFv to the plasma membrane and 2) confers the necessary flexibility to adopt select conformational states upon antigen engagement. To optimize CAR efficacy, it is important to understand how subtle modifications in the framework regions of the CAR targeting domains can influence anti-tumor function.
The intrinsic stability of VH and VL chains is decreased in the scFv format. This instability is exemplified by a tendency to unfold, aggregate, and oligomerize with the scFvs contained within adjacent CARs. Several approaches have been developed to counteract the inherent thermodynamic instability of scFvs which include the inclusion of disulfide bridges to reinforce the VH and VL domains. In the format of a CAR, the aggregation of scFvs leads to CAR clustering, spontaneous T cell activation and cytokine production in the absence of antigen engagement (which is recognized as tonic signaling). Sustained tonic signaling promotes progressive T cell dysfunction, characterized by impaired engraftment, diminished proliferation, poor cytokine production, and decreased anti-tumor function.
In an exemplary study, Mackall and coworkers showed how the physiochemical properties of the scFv framework impact overall CAR T cell function (20). They compared the ability of T cells that were selectively engineered with either a GD2-specific CAR (containing the 14g2a scFv) or CD19-specific CAR (using the FMC63 scFv) to eliminate tumors and persist as memory cells (20). GD2 is a membrane-bound disialoganglioside overexpressed in osteosarcoma (21), melanoma (22) and neuroblastoma (23). CD19 expression is restricted to the B cell lineage. To control for inter-tumor heterogeneity, they ectopically expressed CD19 into GD2-expressing osteosarcoma cells. Immunodeficient mice were engrafted with these target cells. The ability of CAR T cells with directed specificity against GD2 or CD19 to control tumor size was examined (20). As GD2- and CD19-redirected CAR T cells encountered identical tumor burdens, the observed variations in tumoricidal efficacy were attributed to intrinsic factors contained within the CAR design (20). In contrast to CD19-redirected CAR T cells, GD2-redirected CAR T cells lacked efficacy and displayed phenotypic features of exhaustion (20). Using a series of approaches including confocal microscopy, in vitro killing assays, cytokine production, and xenograft models, the authors attributed this antigen-independent anergy to the aggregation of CAR molecules that lead to spontaneous cytokine production, and ultimately effector cell exhaustion (20). Furthermore, the authors concluded that CAR aggregation is strictly driven by the interactions between the framework residues of the scFvs of different GD2-specific CARs on the surface of an engineered T cell (20). These data support earlier findings showing an inherent propensity for antibody (as well as antibody-derived fragments such as scFv) oligomerization (24,25). In general, the organization of antibodies into higher order complexes reduces their free energy cost. Initial findings suggest that 4–1BB mitigates the phenotypic and functional features of exhaustion that result from scFv-induced CAR aggregation (20). These findings are consistent with the proposed ability of 4–1BB to counter anergy in the context of chronic viral infections (20,26,27), and also complement previous work demonstrating antigen-independent signaling contributes to CD28-CD3ζ CAR T cell exhaustion in vivo (28). Taken together, these findings highlight the importance of meticulous CAR design and the need to understand how all components of a synthetic molecule impact therapeutic efficacy.
To overcome the intermolecular clustering of a toxin-specific scFv, Chen and colleagues introduced a disulfide bond between position 44 and 100 of the heavy and light chains, respectively. Replacing amino acids glycine and glutamine with cysteine increased the overall stability of the scFv, without any adverse effect on antigen-binding activity (29). Although speculative, there might be an additive benefit to such structural reconfigurations that enhance scFv stability, along with optimal costimulatory domain to enhance anti-tumor function following adoptive transfer.
Similarly, a recent study showed how the biophysical balance of the CAR ectodomain is an important determinant of CAR T cell efficacy (30). Using the 763.74 scFv, Dotti and colleagues developed a CAR against chondroitin sulphate proteoglycan 4 (CSPG4), which is a glycosylated transmembrane protein overexpressed in malignant melanoma. Regardless of the costimulatory endodomain (4-1BB or CD28), the inherent instability of the scFv culminated in spontaneous cytokine release, tonic signaling, and progressive dysfunction over time (30). Using computational modeling strategies, the authors identified critical residues within the framework regions of the CAR targeting domain (isoleucine 123 as well as glutamine 127) that regulated the scFv stability (30). Advanced structural analyses using Eris software, revealed a number of amino acid substitutions that would further stabilize the underlying framework of the scFv; attenuate tonic signaling, and enhance the overall anti-tumor function of CSPG4-specific CAR T cells (30). As the authors found no connection between CD28 vs 4-1BB endodomains and the degree of tonic signaling, future studies are necessary to understand how CAR costimulation impacts CAR desensitization in the absence of antigen engagement (30). These findings highlight the benefit of structural modeling to design CARs that can overcome the inherent instability of scFvs in the CAR format, and enhance persistence following adoptive transfer (30).
3. Strategies for overcoming antigen escape
Several parameters intrinsic to CAR T cells including CAR expression, T cell trafficking, in vivo expansion, and persistence influence the overall outcome (including relapse) of adoptive transfer. Tumor-intrinsic mechanisms also contribute to relapse following CAR T treatment. In the context of B-ALL, 60% of CAR T cases exhibit relapse due to CD19 antigen loss (31). While the expression of a chimeric entity provides specificity against a given tumor antigen, loss of the corresponding antigen renders the very same CAR T cells obsolete.
There are several established mechanisms by which tumor cells achieve antigen loss and/or antigen escape. Intuitively, tumor cells can decrease CD19 expression on their cell surface. This impedes CAR T cell activation, resulting in a low level of antitumor function (32,33). Mechanistically, tumor cells can achieve this through alternate splicing at the CD19 locus. Splice variants can give rise to truncated CD19 proteins which are undetectable by flow cytometry, but present by western blot analyses. Such a dichotomy , exemplified by the expression of variants lacking the necessary epitope to interact with the corresponding scFv used in flow cytometry reflects antigen escape rather than antigen loss.
Additional mechanisms contributing to relapse following CD19-redirected CAR T cells have been recently identified. CD123 has a low-level expression on normal hematopoietic stem cells but is upregulated in several hematologic malignancies such as acute myeloid leukemia (AML) (34), hairy cell leukemia (35), blastic plasmacytoid dendritic cell neoplasm (36,37), and systemic mastocytosis (38). Importantly, CD123 expression distinguishes a subset of leukemia-initiating cells in AML (39). Ruella and coworkers provided evidence that CD123 is ectopically expressed in B-ALL (40). Moreover, they showed that a subclone of B-ALL cells can undergo a transforming event giving rise to CD19−CD123+ cells with phenotypic and cytogenetic features of leukemia-initiating cells. Additionally, the selective expansion of CD19−CD123+ B-ALL cells under CD19-redirected CAR T immune pressure can give rise to malignant progeny. Other studies corroborated these findings showing aberrant CD123 expression in both pediatric and adult B-ALL (34,37,41–43).
Yu et al. provided further insights into the mechanisms of antigen escape following CD19-redirected CAR immunotherapy in a pediatric case of primary mediastinal large B-cell lymphoma (PMLBCL) (44). In a detailed genomic analysis, Yu et al. identified a missense mutation (G210D) in one allele of the CD19 gene. Computational modeling predicted that substitution of glycine with a negatively-charged aspartic acid in codon 210 would alter the physiochemical binding properties and subcellular distribution of CD19. Phenotypic analysis of G210D mutants revealed a cytoplasmic rather than a cell-surface localization of CD19. Of interest, Yu et al. attributed this mutation to a loss of DNA repair genes yielding transformants that expanded under CD19-redirected CAR T-mediated selective pressure.
CAR T cells targeting the B-cell lineage marker CD22 have been investigated as an alternative approach to overcome relapse from CD19 antigen loss. Durable clinical responses have been achieved in patients with CD19− B-ALL treated with CD22-redirected CAR T cells (45–48). Interestingly, a recent paper showed how CAR T cells could be redirected to a nucleophosmin neoepitope in AML (49). Future studies may identify similar novel targets for CAR T cells in B-ALL.
3.1. Novel antigens
3.1.1. BAFF-R
The B-cell activating factor receptor (BAFF-R), a transmembrane protein of the TNF receptor superfamily, is essential for B-cell maturation and survival (50–52). Importantly, BAFF-R is highly expressed in B-cell malignancies. The canonical ligand for BAFF-R is the B-cell activating factor (BAFF). Following activation, BAFF-R regulates tumor survival and proliferation in an NF-κB-dependent manner (53,54). BAFF-R upregulation is associated with disease progression in patients with B-cell lymphoma and pre-B-ALL (55–57). The critical role of BAFF-R in B-cells positions it as an ideal target for adoptive immunotherapies (58–61).
Qin et al. developed hCARTs against BAFF-R and provided evidence that BAFF-R-redirected CAR T cells have significant cytotoxicity against several B-ALL lines (62). Adoptively transferred BAFF-R-redirected CAR T cells successfully eradicated 10-day pre-established tumor xenografts after a single treatment and maintained efficacy against xenografts models lacking CD19 expression (62). Using patient-derived xenograft (PDX) models, tumors that evade CAR T cell detection by CD19 antigen loss are susceptible to BAFF-R-redirected CAR T cell cytolysis (62). Dong and coworkers also developed a high-affinity humanized monoclonal antibody against the natively expressed BAFF-R (63). By adapting the VH and VL chains to a scFv format and integrating into a CAR, they provided evidence that BAFF-R-redirected CAR T cells were highly effective in xenogeneic mouse models of B cell malignancies, including those with CD19 antigen loss. (64).
3.1.2. CSPG4
CSPG4 is a heavily glycosylated transmembrane protein overexpressed in several malignancies including melanoma, glioma, and triple-negative breast cancer. CSPG4 is also upregulated in mixed-lineage leukemia (MLL)-rearranged leukemia blasts, a form of leukemia with the unfortunate MLL 11q23 rearrangement which occurs in 10% of all leukemias (65–74). CSPG4 is an established anti-tumor target. Monoclonal antibodies that disrupt ligand access to CSPG4 have been developed, and shown therapeutic efficacy in cancer immunotherapies (75). Beyond competitive inhibition, conjugating CSPG4-specific antibodies with pro-apoptotic factors also has translational relevance (75). Fusion proteins linking the CSPG4 binding domain to soluble TRAIL (TNF-related apoptosis-inducing ligand) agonists can trigger cell death upon CSPG4 binding through the extrinsic apoptosis pathway (76). One inherent limitation in the use of CSPG4 as a target for adoptive immunotherapy is that its expression is not limited to tumor cells. Low levels of CSPG4 have been observed in activated pericytes and smooth muscle cells (77–79). This is concerning as a recent paper showed how mural cells, which are a subset of vascular smooth muscle cells surrounding capillary beds, are susceptible to CD19-redirected CAR T cell-induced neurotoxicity (80). Gene delivery approaches permissive for transient CAR expression are safer alternatives to lentiviral-mediated CAR expression whenever toxicities are a major concern. For this reason, mRNA-based CSPG4-redirected CAR T cells might be a preferable approach to limit the potential for “on-target off-tumor” toxicities, specifically with vascular smooth muscle cells found in various organs including the brain (81,82). It is encouraging to assert that mRNA-transfected CAR T cells can achieve tumoricidal reactions similar to those mediated by virally-transduced CAR T cells (83).
3.1.3. TSLPR
Thymic stromal lymphopoietin receptor (TSLPR) is a heterodimeric receptor complex activated by the thymic stromal lymphopoietin (TSLP) cytokine. TSLPR is overexpressed in 5% to 15% of B-ALL patients. CRLF2 translocations and deletions result in alternative promoter activation and TSLPR overexpression in B-ALL (84–89). CRLF2 gene rearrangements are associated with poor prognosis in B-ALL patients (90–95). TSLPR activation promotes JAK/STAT signal transduction in B-ALL blasts (96,97). Given its restricted expression in normal tissues, its role as a B-ALL oncoprotein, and its cell-surface overexpression and association with poor clinical outcomes, TSLPR is a potential target for CAR T cell therapies against B-ALL. Qin and colleagues evaluated the efficacy of TSLPR-targeting CAR T cells against CRLF2-overexpressing B-ALL in vitro and in vivo (98). TSLPR-targeting CAR T cells demonstrated potent cytotoxicity in vitro. Moreover, the anti-leukemic activity of TSLPR-targeting CAR T cells was demonstrated in mice engrafted with a TSLPR-expressing ALL cell line. TSLPR-redirected CAR T cells also eliminated human CRLF2-rearranged TSLPR-overexpressing ALL in PDX models. These findings implicate the cytokine receptor TSLPR as a promising therapeutic option for CRLF2-rearranged B-ALL patients.
3.1.4. Other Proposed Targets
CD20 is a non-glycosylated transmembrane phosphoprotein unique to B-cells. While CD20 is abundant on mature B-cells, it is absent on pro-B-cells, plasma cells, and hematopoietic stem cells (99). CD20 regulates cell cycle progression as well as differentiation, inhibits apoptosis, and facilitates calcium entry (100). Properties unique to CD20 make it an attractive target for CAR T cell therapies against cancer. In particular, CD20 is neither internalized nor secreted following ligand/antibody engagement. There are several monoclonal antibodies against CD20 including Rituximab (FDA-approved in 1997), Ofatumumab (FDA-approved in 2014), and Obinutuzumab (FDA-approved in 2013), which have been investigated or used for the treatment of B-ALL and other hematologic malignancies (101–103).
The cell surface glycoprotein CD52 is another potential target that has not received much attention. Whereas CD52 is expressed on differentiating B-cells and T cells, it is absent on terminally differentiated, circulating, plasma cells. Alemtuzumab is a humanized CD52-specific monoclonal antibody that has shown promising results in chronic lymphocytic leukemia (CLL) patients leading to its FDA approval in 2014 (104). Although, its clinical benefit in the treatment of ALL has been poor (105), repurposing the scFv in a CAR construct for CAR T cell therapy may restore its potential as a therapeutic target for ALL.
3.2. Multi-targeting CARs
3.2.1. Dual-targeting strategies
To overcome the adaptive response of B-ALL cells to CD19-redirected CAR T cells, and the aberrant expression of CD123, Ruella et al., devised a dual receptor approach targeting both CD19 and CD123 simultaneously and set out to test this construct using a novel preclinical model of relapse. In this xenograft model, NSG mice were injected with primary ALL blasts (CD19+ CD123+) isolated from a patient before CD19-redirected CAR T treatment, as well as after relapse (CD19− CD123+). As CD123-redirected CAR T cells demonstrated efficacy against B-ALL cells exhibiting resistance to CD19-redirected CAR T cells, their findings highlight the therapeutic potential of dual receptor CARs in preventing relapse due to antigen loss. Complementing the findings from Ruella and coworkers, a more recent study demonstrated that CAR T cells with two discrete scFv domains, one against CD19 and the other against CD123, demonstrated cytotoxic effects against their corresponding targets both in vitro and in vivo. This further supports the idea that a dual CAR approach is effective in augmenting the response against leukemic blasts and reducing overall rates of disease relapse (106).
Simultaneous multi-specific targeting can enhance the durability of B-ALL remission while minimizing the risk of antigen escape-associated relapse (45,107). A bispecific CAR platform is one approach to effectively overcome escape variants. As surface receptor engagement is an important determinant of efficacy, simultaneously targeting two distinct antigens also raises the overall T cell anti-tumor response. Beneficial attributes of bispecific CAR platforms in advanced B-cell malignancies and glioblastoma have been established (45,108–110). In one innovative design, CD19- and CD22-specific scFvs were combined to form a single bivalent receptor (45). These bispecific CAR T cells, targeting both CD19 and CD22, demonstrated potent cytotoxic effects against CD19+CD22+, CD19−CD22+, and CD19−CD22+ cells in B-ALL (45).
In one pilot study (including 51 ALL patients) the efficacy and safety of a sequential CAR T cell infusion strategy (targeting CD19 followed by CD22) were established (111). The authors reported a minimal residual disease-negative response rate of 96.0% among the 51 ALL patients and a median progression-free survival of 13.6 months (with a median follow-up of 16.7 months) (111). The benefits of a sequential CAR T cell infusion strategy including an increased leukemia-free survival rate were corroborated by other studies (112,113). Fig. 3A illustrates different approaches for having dual-targeting CAR T cells.
Figure 3.
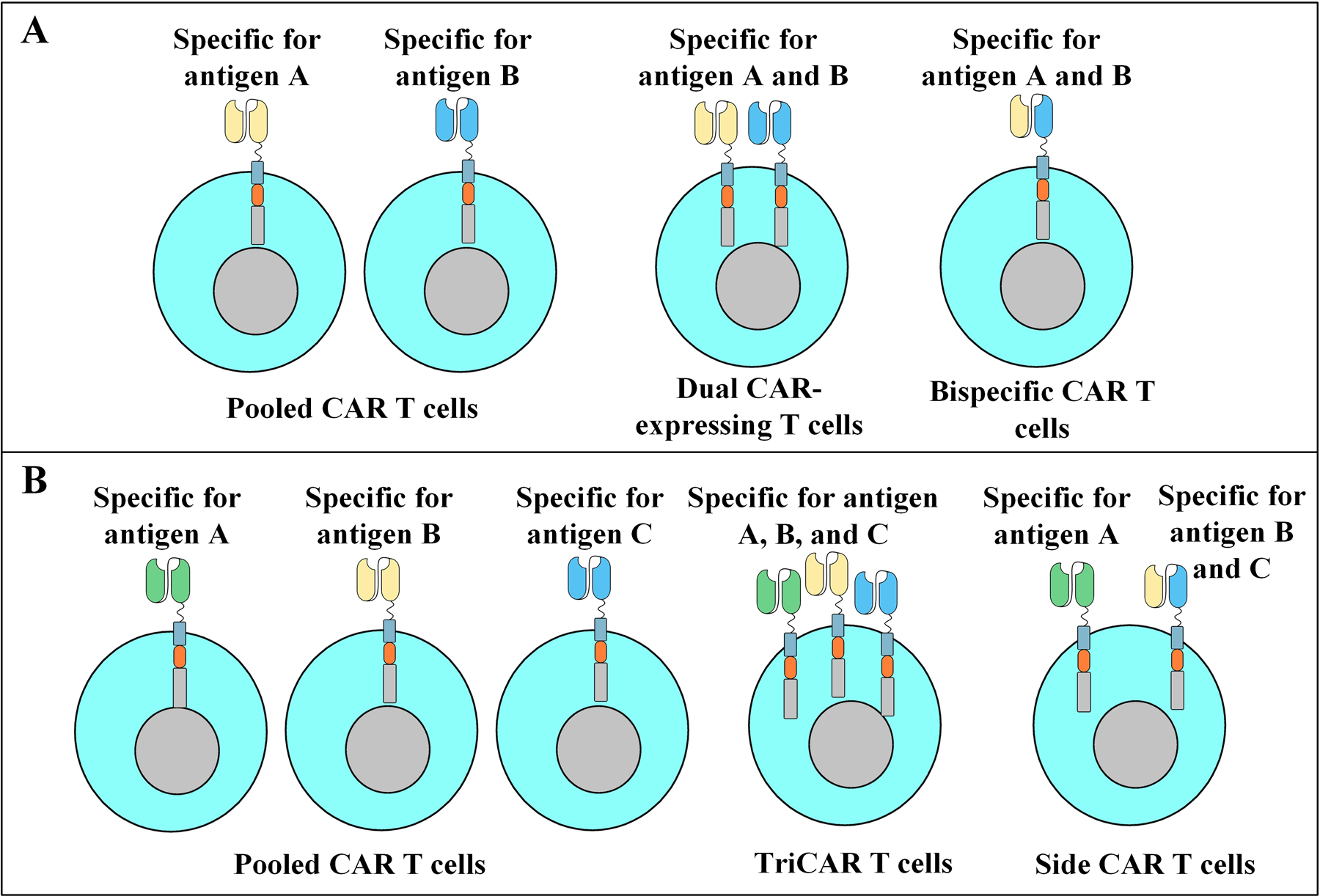
Different strategies for having multi-targeting CAR T cells. A: Dual-targeting CAR T cells. Pooled CAR T cells are a combination of two populations of distinct CAR T cells each targeting a different antigen of interest (e.g. CD19 and CD20). Dual CAR-expressing CAR T cells express two different CAR constructs on a single cell with each CAR targeting a different antigen of interest. Bispecific CAR T cells are equipped with a bispecific targeting domain thus enabling them to target two antigens of interest without the need for the expression of two distinct CARs. B: Trivalent CAR T cells. Pooled CAR T cells are a combination of three populations of distinct CAR T cells each capable of targeting a different antigen of interest (e.g. CD19, CD20, and CD22). Triple CAR-expressing CAR T cells express three different CAR constructs on a single cell with each CAR targeting a different antigen of interest. SideCAR T cells co-express a conventional CAR specific for antigen A alongside a bispecific CAR specific for antigen B and C.
3.2.2. Trivalent CARs
Given the incidence of CD19− disease relapse, approaches to broaden the specificity of CAR T cells against additional tumor antigens have been developed (114,115). One such study targeted three distinct leukemia antigens (CD19, CD20, and CD22) to evaluate whether a trivalent CAR platform could control disease progression during CD19− relapse (114,115). Two second-generation trivalent CAR T cell products targeting 1) CD19 (using the FMC63 scFv), 2) CD20 (using a Rituximab-derived scFv), and 3) CD22 (using the m971 scFv) were developed (114). The first CAR T cell product expressed three individual CARs, with each CAR directing T cell cytotoxicity against a unique leukemia antigen (hereafter referred to as TriCAR); the second CAR T cell product (hereafter referred to as SideCAR) expressed a conventional CD19-targeting CAR as well as a CD20- and CD22-targeting bispecific CAR achieved through a tandem arrangement (114). In vitro cytolytic activity was enhanced in T cells expressing either TriCAR or SideCAR relative to CD19-redirected CAR T cells (114). Trivalent CAR T cells were also functionally effective against CD19− CD20+CD22+ target cells highlighting their translational relevance against CD19 escape variants (114). In Fig. 3B, we outline various trivalent CAR designs. Future studies will likely provide more insights into their therapeutic potential and clinical impact.
It can be an arduous task to find one TSA, let alone two or three antigens that are strictly restricted to only tumor cell. In this regard, one of the main downsides of multitargeted CAR T cells might be the increased risk of on-target off-tumor toxicities delivered to healthy tissues. In the past years, researchers have meticulously devised several counterstrategies to overcome such limitations which include inhibitory CARs (iCARs); in this context, the absence of a specific antigen on tumor cells permits downstream signaling cascades inducing tumoricidal responses, or logic-gated CAR T cells that enforce antitumor reactions only towards tumor cells that simultaneously express two target antigens recognized by the CAR constructs (2,6). While these countertactics sound reasonable, their practicality can only be determined in clinical trials in years to come. Another potential strategy for reversing adverse events attributed to CAR T cells is to equip T cells with safety switches that enable their rapid elimination from the circulation upon the introduction of a safety switch-activating agent (2,6). Aside from these, transient CAR expression seems like one of the most reliable strategies and have been well-tolerated by a number of breast cancer, melanoma, and pancreatic cancer patients enrolled in clinical trials NCT03060356 and NCT01897415. According to one study with six pancreatic ductal adenocarcinoma patients, mRNA-based mesothelin-redirected CAR T cells showed no sign of CRS, neurotoxicity, or any other serious dose-limiting toxicities (DLT) (116). However, there are inherent limitations in the use of mRNA-based CAR T cells and they require frequent infusion to sustain antitumor responses.
4. Future Opportunities and Challenges
Rapid advances in the development of CAR T cells for B-ALL paved the way for a number of FDA approvals against a broadening range of hematologic malignancies. However, the therapeutic potential of CAR T cells can be further expanded if infallible countertactics are undertaken, including advanced CAR design and engineering to enhance T cell persistence and extend the limited durable efficacy displayed by senescent T cells, or dual targeting approaches to eradicate tumor cells that evade immunosurveillance by antigen escape and antigen loss. Accumulating evidence suggests that optimizing CAR design: by including of fully-human or humanized targeting domains in the CAR construct, selecting a suitable costimulatory domain, and identifying alternate target antigens will enhance the therapeutic promise of CAR T cells even further. The discovery of novel target antigens and the development of multi-targeted CAR T cells can substantially diminish the risk of disease relapse. However, multitargeting strategies might result in life-threatening side effects if the targeted antigen sets are not cancer-specific. Moreover, since finding TSAs is challenging, identifying and targeting aberrantly-glycosylated forms of antigens, known as Tn and Sialyl Tn glycoform, that are only expressed by tumor cells might be an alternative worth considering. Combining other treatment modalities, such as oncolytic virotherapy or bispecific T-cell engagers (BiTEs), may also enhance the clinical efficacy of adoptively transferred CAR T cells. We are at the nexus of important breakthroughs that will propel the field of CAR T cell therapies to even greater heights.
Acknowledgments
R.OC is supported by an NIH grant RO1CA226983-03.
Financial support:
Pouya Safarzadeh Kozani: Not applicable; Pooria Safarzadeh Kozani: Not applicable; R.S. O’Connor: R.OC is supported by a National Institutes of Health (NIH) grant RO1CA226983-03.
Abbreviations:
- B-ALL
B-cell acute lymphoblastic leukemia
- R/R
relapsed or refractory
- CAR
chimeric antigen receptor
- FDA
US Food and Drug Administration
- DLBCL
diffuse large B-cell lymphoma
- MCL
mantle cell lymphoma
- ACT
adoptive T cell therapies
- TAA
tumor-associated antigens
- TSA
tumor-specific antigens
- scFv
single-chain variable fragment
- TCR
T cell receptor
- APCs
antigen-presenting cells
- MHC
major histocompatibility complexes
- hCARTs
CAR T cells expressing a humanized scFv
- CR
complete remission
- CRS
cytokine release syndrome
- CSPG4
chondroitin sulphate proteoglycan 4
- AML
acute myeloid leukemia
- PMLBCL
primary mediastinal large B-cell lymphoma
- BAFF-R
B-cell activating factor receptor
- BAFF
B-cell activating factor
- PDX
patient-derived xenograft
- MLL
mixed-lineage leukemia
- TRAIL
TNF-related apoptosis-inducing ligand
- TSLPR
thymic stromal lymphopoietin receptor
- TSLP
thymic stromal lymphopoietin
- CLL
chronic lymphocytic leukemia
- iCARs
inhibitory CARs
- DLT
dose-limiting toxicities
- BiTEs
bispecific T-cell engagers
Footnotes
Conflict of interest:
Pouya Safarzadeh Kozani: No conflict of interest to declare; Pooria Safarzadeh Kozani: No conflict of interest to declare; Roddy S. O’Connor: R.OC is an inventor on patents related to CAR T cell therapy, the rights of which are assigned to the University of Pennsylvania and licensed to Novartis AG.
References
- 1.Sotillo E, Barrett DM, Black KL, Bagashev A, Oldridge D, Wu G, et al. Convergence of acquired mutations and alternative splicing of CD19 enables resistance to CART-19 immunotherapy. Cancer discovery 2015;5(12):1282–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hashem Boroojerdi M, Rahbarizadeh F, Safarzadeh Kozani P, Kamali E, Safarzadeh Kozani P. Strategies for having a more effective and less toxic CAR T-cell therapy for acute lymphoblastic leukemia. Med Oncol 2020;37(11):100 doi 10.1007/s12032-020-01416-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mullard A FDA approves fourth CAR-T cell therapy. Nat Rev Drug Discov 2021. doi 10.1038/d41573-021-00031-9. [DOI] [PubMed] [Google Scholar]
- 4.Kalos M, June CH. Adoptive T cell transfer for cancer immunotherapy in the era of synthetic biology. Immunity 2013;39(1):49–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rosenberg SA, Restifo NP. Adoptive cell transfer as personalized immunotherapy for human cancer. Science 2015;348(6230):62–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Safarzadeh Kozani P, Safarzadeh Kozani P, Rahbarizadeh F, Khoshtinat Nikkhoi S. Strategies for Dodging the Obstacles in CAR T Cell Therapy. Frontiers in Oncology 2021;11(924) doi 10.3389/fonc.2021.627549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sadelain M, Brentjens R, Rivière I. The basic principles of chimeric antigen receptor design. Cancer discovery 2013;3(4):388–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sommermeyer D, Hill T, Shamah SM, Salter AI, Chen Y, Mohler KM, et al. Fully human CD19-specific chimeric antigen receptors for T-cell therapy. Leukemia 2017;31(10):2191–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lamers CH, Willemsen R, van Elzakker P, van Steenbergen-Langeveld S, Broertjes M, Oosterwijk-Wakka J, et al. Immune responses to transgene and retroviral vector in patients treated with ex vivo–engineered T cells. Blood 2011;117(1):72–82. [DOI] [PubMed] [Google Scholar]
- 10.Turtle CJ, Hanafi L-A, Berger C, Gooley TA, Cherian S, Hudecek M, et al. CD19 CAR–T cells of defined CD4+: CD8+ composition in adult B cell ALL patients. The Journal of clinical investigation 2016;126(6):2123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Haas AR, Tanyi JL, O’Hara MH, Gladney WL, Lacey SF, Torigian DA, et al. Phase I Study of Lentiviral-Transduced Chimeric Antigen Receptor-Modified T Cells Recognizing Mesothelin in Advanced Solid Cancers. Mol Ther 2019;27(11):1919–29 doi 10.1016/j.ymthe.2019.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Qian L, Li D, Ma L, He T, Qi F, Shen J, et al. The novel anti-CD19 chimeric antigen receptors with humanized scFv (single-chain variable fragment) trigger leukemia cell killing. Cellular immunology 2016;304:49–54. [DOI] [PubMed] [Google Scholar]
- 13.Cao J, Wang G, Cheng H, Wei C, Qi K, Sang W, et al. Potent anti-leukemia activities of humanized CD19-targeted Chimeric antigen receptor T (CAR-T) cells in patients with relapsed/refractory acute lymphoblastic leukemia. American journal of hematology 2018;93(7):851–8. [DOI] [PubMed] [Google Scholar]
- 14.Hu Y, Wu Z, Luo Y, Shi J, Yu J, Pu C, et al. Potent anti-leukemia activities of chimeric antigen receptor–Modified T cells against CD19 in Chinese patients with relapsed/refractory acute lymphocytic leukemia. Clinical Cancer Research 2017;23(13):3297–306. [DOI] [PubMed] [Google Scholar]
- 15.Heng G, Jia J, Li S, Fu G, Wang M, Qin D, et al. Sustained therapeutic efficacy of humanized anti-CD19 chimeric antigen receptor T cells in relapsed/refractory acute lymphoblastic leukemia. Clinical Cancer Research 2020;26(7):1606–15. [DOI] [PubMed] [Google Scholar]
- 16.Brudno JN, Kochenderfer JN. Toxicities of chimeric antigen receptor T cells: recognition and management. Blood 2016;127(26):3321–30 doi 10.1182/blood-2016-04-703751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cao J, Wang G, Cheng H, Wei C, Qi K, Sang W, et al. Potent anti-leukemia activities of humanized CD19-targeted Chimeric antigen receptor T (CAR-T) cells in patients with relapsed/refractory acute lymphoblastic leukemia. Am J Hematol 2018;93(7):851–8 doi 10.1002/ajh.25108. [DOI] [PubMed] [Google Scholar]
- 18.Cao J, Cheng H, Shi M, Wang G, Chen W, Qi K, et al. Humanized CD19-specific chimeric antigen-receptor T-cells in 2 adults with newly diagnosed B-cell acute lymphoblastic leukemia. Leukemia 2019;33(11):2751–3 doi 10.1038/s41375-019-0516-7. [DOI] [PubMed] [Google Scholar]
- 19.Zhao Y, Liu Z, Wang X, Wu H, Zhang J, Yang J, et al. Treatment with Humanized Selective CD19CAR-T Cells Shows Efficacy in Highly Treated B-ALL Patients Who Have Relapsed after Receiving Murine-Based CD19CAR-T Therapies. Clin Cancer Res 2019;25(18):5595–607 doi 10.1158/1078-0432.Ccr-19-0916. [DOI] [PubMed] [Google Scholar]
- 20.Long AH, Haso WM, Shern JF, Wanhainen KM, Murgai M, Ingaramo M, et al. 4-1BB costimulation ameliorates T cell exhaustion induced by tonic signaling of chimeric antigen receptors. Nat Med 2015;21(6):581–90 doi 10.1038/nm.3838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Roth M, Linkowski M, Tarim J, Piperdi S, Sowers R, Geller D, et al. Ganglioside GD2 as a therapeutic target for antibody-mediated therapy in patients with osteosarcoma. Cancer 2014;120(4):548–54 doi 10.1002/cncr.28461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tsao CY, Sabbatino F, Cheung NV, Hsu JC, Villani V, Wang X, et al. Anti-proliferative and pro-apoptotic activity of GD2 ganglioside-specific monoclonal antibody 3F8 in human melanoma cells. Oncoimmunology 2015;4(8):e1023975 doi 10.1080/2162402X.2015.1023975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cheung NK, Dyer MA. Neuroblastoma: developmental biology, cancer genomics and immunotherapy. Nat Rev Cancer 2013;13(6):397–411 doi 10.1038/nrc3526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nieba L, Honegger A, Krebber C, Plückthun A. Disrupting the hydrophobic patches at the antibody variable/constant domain interface: improved in vivo folding and physical characterization of an engineered scFv fragment. Protein Eng 1997;10(4):435–44 doi 10.1093/protein/10.4.435. [DOI] [PubMed] [Google Scholar]
- 25.Whitlow M, Filpula D, Rollence ML, Feng SL, Wood JF. Multivalent Fvs: characterization of single-chain Fv oligomers and preparation of a bispecific Fv. Protein Eng 1994;7(8):1017–26 doi 10.1093/protein/7.8.1017. [DOI] [PubMed] [Google Scholar]
- 26.Vezys V, Penaloza-MacMaster P, Barber DL, Ha SJ, Konieczny B, Freeman GJ, et al. 4–1BB signaling synergizes with programmed death ligand 1 blockade to augment CD8 T cell responses during chronic viral infection. J Immunol 2011;187(4):1634–42 doi 10.4049/jimmunol.1100077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang C, Wen T, Routy JP, Bernard NF, Sekaly RP, Watts TH. 4-1BBL induces TNF receptor-associated factor 1-dependent Bim modulation in human T cells and is a critical component in the costimulation-dependent rescue of functionally impaired HIV-specific CD8 T cells. J Immunol 2007;179(12):8252–63 doi 10.4049/jimmunol.179.12.8252. [DOI] [PubMed] [Google Scholar]
- 28.Frigault MJ, Lee J, Basil MC, Carpenito C, Motohashi S, Scholler J, et al. Identification of chimeric antigen receptors that mediate constitutive or inducible proliferation of T cells. Cancer Immunol Res 2015;3(4):356–67 doi 10.1158/2326-6066.CIR-14-0186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhao JX, Yang L, Gu ZN, Chen HQ, Tian FW, Chen YQ, et al. Stabilization of the single-chain fragment variable by an interdomain disulfide bond and its effect on antibody affinity. Int J Mol Sci 2010;12(1):1–11 doi 10.3390/ijms12010001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Landoni E, Fucá G, Wang J, Chirasani VR, Yao Z, Dukhovlinova E, et al. Modifications to the Framework Regions Eliminate Chimeric Antigen Receptor Tonic Signaling. Cancer Immunol Res 2021. doi 10.1158/2326-6066.Cir-20-0451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Shah NN, Fry TJ. Mechanisms of resistance to CAR T cell therapy. Nat Rev Clin Oncol 2019;16(6):372–85 doi 10.1038/s41571-019-0184-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Park JH, Geyer MB, Brentjens RJ. CD19-targeted CAR T-cell therapeutics for hematologic malignancies: interpreting clinical outcomes to date. Blood, The Journal of the American Society of Hematology 2016;127(26):3312–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Turtle CJ, Riddell SR. Artificial antigen presenting cells for use in adoptive immunotherapy. Cancer journal (Sudbury, Mass) 2010;16(4):374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Muñoz L, Nomdedéu JF, López O, Carnicer MJ, Bellido M, Aventín A, et al. Interleukin-3 receptor alpha chain (CD123) is widely expressed in hematologic malignancies. Haematologica 2001;86(12):1261–9. [PubMed] [Google Scholar]
- 35.Del Giudice I, Matutes E, Morilla R, Morilla A, Owusu-Ankomah K, Rafiq F, et al. The diagnostic value of CD123 in B-cell disorders with hairy or villous lymphocytes. Haematologica 2004;89(3):303–8. [PubMed] [Google Scholar]
- 36.Angelot-Delettre F, Roggy A, Frankel AE, Lamarthee B, Seilles E, Biichle S, et al. In vivo and in vitro sensitivity of blastic plasmacytoid dendritic cell neoplasm to SL-401, an interleukin-3 receptor targeted biologic agent. Haematologica 2015;100(2):223–30 doi 10.3324/haematol.2014.111740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Testa U, Pelosi E, Frankel A. CD 123 is a membrane biomarker and a therapeutic target in hematologic malignancies. Biomark Res 2014;2(1):4 doi 10.1186/2050-7771-2-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pardanani A, Reichard KK, Zblewski D, Abdelrahman RA, Wassie EA, Morice Ii WG, et al. CD123 immunostaining patterns in systemic mastocytosis: differential expression in disease subgroups and potential prognostic value. Leukemia 2016;30(4):914–8 doi 10.1038/leu.2015.348. [DOI] [PubMed] [Google Scholar]
- 39.Ehninger A, Kramer M, Röllig C, Thiede C, Bornhäuser M, von Bonin M, et al. Distribution and levels of cell surface expression of CD33 and CD123 in acute myeloid leukemia. Blood Cancer J 2014;4(6):e218 doi 10.1038/bcj.2014.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ruella M, Barrett DM, Kenderian SS, Shestova O, Hofmann TJ, Perazzelli J, et al. Dual CD19 and CD123 targeting prevents antigen-loss relapses after CD19-directed immunotherapies. The Journal of clinical investigation 2016;126(10):3814–26 doi 10.1172/JCI87366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Djokic M, Björklund E, Blennow E, Mazur J, Söderhäll S, Porwit A. Overexpression of CD123 correlates with the hyperdiploid genotype in acute lymphoblastic leukemia. Haematologica 2009;94(7):1016–9 doi 10.3324/haematol.2008.000299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jordan CT, Upchurch D, Szilvassy SJ, Guzman ML, Howard DS, Pettigrew AL, et al. The interleukin-3 receptor alpha chain is a unique marker for human acute myelogenous leukemia stem cells. Leukemia 2000;14(10):1777–84 doi 10.1038/sj.leu.2401903. [DOI] [PubMed] [Google Scholar]
- 43.Riccioni R, Rossini A, Calabrò L, Diverio D, Pasquini L, Lococo F, et al. Immunophenotypic features of acute myeloid leukemias overexpressing the interleukin 3 receptor alpha chain. Leuk Lymphoma 2004;45(8):1511–7 doi 10.1080/104281090310001646031. [DOI] [PubMed] [Google Scholar]
- 44.Yu AL, Gilman AL, Ozkaynak MF, London WB, Kreissman SG, Chen HX, et al. Anti-GD2 antibody with GM-CSF, interleukin-2, and isotretinoin for neuroblastoma. The New England journal of medicine 2010;363(14):1324–34 doi 10.1056/NEJMoa0911123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Fry TJ, Shah NN, Orentas RJ, Stetler-Stevenson M, Yuan CM, Ramakrishna S, et al. CD22-targeted CAR T cells induce remission in B-ALL that is naive or resistant to CD19-targeted CAR immunotherapy. Nature medicine 2018;24(1):20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shah NN, Stevenson MS, Yuan CM, Richards K, Delbrook C, Kreitman RJ, et al. Characterization of CD22 expression in acute lymphoblastic leukemia. Pediatric blood & cancer 2015;62(6):964–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rosenthal J, Naqvi AS, Wertheim G, Paessler M, Rheingold SR, Thomas-Tikhonenko A, et al. Semi-Quantitative Analysis of CD19 and CD22 Expression in B-Lymphoblastic Leukemia and Implications for Targerted Immunotherapy. Blood 2017;130(Supplement 1):1331.-. [Google Scholar]
- 48.Shah NN, Highfill SL, Shalabi H, Yates B, Jin J, Wolters PL, et al. CD4/CD8 T-cell selection affects chimeric antigen receptor (CAR) T-cell potency and toxicity: updated results from a phase I anti-CD22 CAR T-cell trial. Journal of Clinical Oncology 2020;38(17):1938–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Xie G, Ivica NA, Jia B, Li Y, Dong H, Liang Y, et al. CAR-T cells targeting a nucleophosmin neoepitope exhibit potent specific activity in mouse models of acute myeloid leukaemia. Nat Biomed Eng 2020. doi 10.1038/s41551-020-00625-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Treml JF, Hao Y, Stadanlick JE, Cancro MP. The BLyS family: toward a molecular understanding of B cell homeostasis. Cell Biochem Biophys 2009;53(1):1–16 doi 10.1007/s12013-008-9036-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Rodig SJ, Shahsafaei A, Li B, Mackay CR, Dorfman DM. BAFF-R, the major B cell-activating factor receptor, is expressed on most mature B cells and B-cell lymphoproliferative disorders. Hum Pathol 2005;36(10):1113–9 doi 10.1016/j.humpath.2005.08.005. [DOI] [PubMed] [Google Scholar]
- 52.Novak AJ, Grote DM, Stenson M, Ziesmer SC, Witzig TE, Habermann TM, et al. Expression of BLyS and its receptors in B-cell non-Hodgkin lymphoma: correlation with disease activity and patient outcome. Blood 2004;104(8):2247–53 doi 10.1182/blood-2004-02-0762. [DOI] [PubMed] [Google Scholar]
- 53.Pham LV, Fu L, Tamayo AT, Bueso-Ramos C, Drakos E, Vega F, et al. Constitutive BR3 receptor signaling in diffuse, large B-cell lymphomas stabilizes nuclear factor-κB–inducing kinase while activating both canonical and alternative nuclear factor-κB pathways. Blood, The Journal of the American Society of Hematology 2011;117(1):200–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fu L, Lin-Lee Y-C, Pham LV, Tamayo AT, Yoshimura LC, Ford RJ. BAFF-R promotes cell proliferation and survival through interaction with IKKβ and NF-κB/c-Rel in the nucleus of normal and neoplastic B-lymphoid cells. Blood, The Journal of the American Society of Hematology 2009;113(19):4627–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Li Y-J, Jiang W-Q, Rao H-L, Huang J-J, Xia Y, Huang H-Q, et al. Expression of BAFF and BAFF-R in follicular lymphoma: correlation with clinicopathologic characteristics and survival outcomes. PloS one 2012;7(12):e50936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Shen X, Wang M, Guo Y, Ju S. The Correlation Between Non-Hodgkin Lymphoma and Expression Levels of B-Cell Activating Factor and Its Receptors. Advances in clinical and experimental medicine: official organ Wroclaw Medical University 2016;25(5):837–44. [DOI] [PubMed] [Google Scholar]
- 57.Parameswaran R, Müschen M, Kim Y-m, Groffen J, Heisterkamp N. A functional receptor for B-cell–activating factor is expressed on human acute lymphoblastic leukemias. Cancer research 2010;70(11):4346–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Novak AJ, Grote DM, Stenson M, Ziesmer SC, Witzig TE, Habermann TM, et al. Expression of BLyS and its receptors in B-cell non-Hodgkin lymphoma: correlation with disease activity and patient outcome. Blood 2004;104(8):2247–53. [DOI] [PubMed] [Google Scholar]
- 59.Rodig SJ, Shahsafaei A, Li B, Mackay CR, Dorfman DM. BAFF-R, the major B cell–activating factor receptor, is expressed on most mature B cells and B-cell lymphoproliferative disorders. Human pathology 2005;36(10):1113–9. [DOI] [PubMed] [Google Scholar]
- 60.Nakamura N, Hase H, Sakurai D, Yoshida S, Abe M, Tsukada N, et al. Expression of BAFF-R (BR3) in normal and neoplastic lymphoid tissues characterized with a newly developed monoclonal antibody. Virchows Archiv 2005;447(1):53–60. [DOI] [PubMed] [Google Scholar]
- 61.Paterson JC, Tedoldi S, Craxton A, Jones M, Hansmann M-L, Collins G, et al. The differential expression of LCK and BAFF-receptor and their role in apoptosis in human lymphomas. Haematologica 2006;91(6):772–80. [PubMed] [Google Scholar]
- 62.Qin H, Dong Z, Wang X, Cheng WA, Wen F, Xue W, et al. CAR T cells targeting BAFF-R can overcome CD19 antigen loss in B cell malignancies. Science translational medicine 2019;11(511):eaaw9414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Qin H, Wei G, Sakamaki I, Dong Z, Cheng WA, Smith DL, et al. Novel BAFF-Receptor Antibody to Natively Folded Recombinant Protein Eliminates Drug-Resistant Human B-cell Malignancies In Vivo. Clin Cancer Res 2018;24(5):1114–23 doi 10.1158/1078-0432.Ccr-17-1193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Dong Z, Cheng WA, Smith DL, Huang B, Zhang T, Chang WC, et al. Antitumor efficacy of BAFF-R targeting CAR T cells manufactured under clinic-ready conditions. Cancer Immunology, Immunotherapy: CII 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Campoli MR, Chang C-C, Kageshita T, Wang X, McCarthy JB, Ferrone S. Human high molecular weight-melanoma-associated antigen (HMW-MAA): a melanoma cell surface chondroitin sulfate proteoglycan (MSCP) with biological and clinical significance. Critical Reviews™ in Immunology 2004;24(4). [DOI] [PubMed] [Google Scholar]
- 66.Hilden JM, Smith FO, Frestedt JL, McGlennen R, Howells WB, Sorensen PH, et al. MLL gene rearrangement, cytogenetic 11q23 abnormalities, and expression of the NG2 molecule in infant acute myeloid leukemia. Blood, The Journal of the American Society of Hematology 1997;89(10):3801–5. [PubMed] [Google Scholar]
- 67.Ilieva KM, Cheung A, Mele S, Chiaruttini G, Crescioli S, Griffin M, et al. Chondroitin sulfate proteoglycan 4 and its potential as an antibody immunotherapy target across different tumor types. Frontiers in immunology 2018;8:1911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Schwartz S, Rieder H, Schläger B, Burmeister T, Fischer L, Thiel E. Expression of the human homologue of rat NG2 in adult acute lymphoblastic leukemia: close association with MLL rearrangement and a CD10−/CD24−/CD65s+/CD15+ B-cell phenotype. Leukemia 2003;17(8):1589–95. [DOI] [PubMed] [Google Scholar]
- 69.Wang X, Osada T, Wang Y, Yu L, Sakakura K, Katayama A, et al. CSPG4 protein as a new target for the antibody-based immunotherapy of triple-negative breast cancer. Journal of the National Cancer Institute 2010;102(19):1496–512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Winters AC, Bernt KM. MLL-rearranged leukemias—an update on science and clinical approaches. Frontiers in pediatrics 2017;5:4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wuchter C, Harbott J, Schoch C, Schnittger S, Borkhardt A, Karawajew L, et al. Detection of acute leukemia cells with mixed lineage leukemia (MLL) gene rearrangements by flow cytometry using monoclonal antibody 7.1. Leukemia 2000;14(7):1232–8. [DOI] [PubMed] [Google Scholar]
- 72.Yadavilli S, Hwang EI, Packer RJ, Nazarian J. The role of NG2 proteoglycan in glioma. Translational Oncology 2016;9(1):57–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Behm FG, Smith FO, Raimondi SC, Pui CH, Bernstein ID. Human homologue of the rat chondroitin sulfate proteoglycan, NG2, detected by monoclonal antibody 7.1, identifies childhood acute lymphoblastic leukemias with t(4;11)(q21;q23) or t(11;19)(q23;p13) and MLL gene rearrangements. Blood 1996;87(3):1134–9. [PubMed] [Google Scholar]
- 74.Smith FO, Rauch C, Williams DE, March CJ, Arthur D, Hilden J, et al. The human homologue of rat NG2, a chondroitin sulfate proteoglycan, is not expressed on the cell surface of normal hematopoietic cells but is expressed by acute myeloid leukemia blasts from poor-prognosis patients with abnormalities of chromosome band 11q23. Blood 1996;87(3):1123–33. [PubMed] [Google Scholar]
- 75.Jordaan S, Chetty S, Mungra N, Koopmans I, Van Bommel PE, Helfrich W, et al. CSPG4: a target for selective delivery of human cytolytic fusion proteins and TRAIL. Biomedicines 2017;5(3):37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.de Bruyn M, Rybczynska AA, Wei Y, Schwenkert M, Fey GH, Dierckx RA, et al. Melanoma-associated Chondroitin Sulfate Proteoglycan (MCSP)-targeted delivery of soluble TRAIL potently inhibits melanoma outgrowth in vitro and in vivo. Molecular cancer 2010;9(1):1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ozerdem U Targeting pericytes diminishes neovascularization in orthotopic uveal melanoma in nerve/glial antigen 2 proteoglycan knockout mouse. Ophthalmic research 2006;38(5):251–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ozerdem U Targeting of pericytes diminishes neovascularization and lymphangiogenesis in prostate cancer. The Prostate 2006;66(3):294–304. [DOI] [PubMed] [Google Scholar]
- 79.Tordsson JM, Ohlsson LG, Abrahmsén LB, Karlström PJ, Lando PA, Brodin TN. Phage-selected primate antibodies fused to superantigens for immunotherapy of malignant melanoma. Cancer Immunology, Immunotherapy 2000;48(12):691–702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Parker KR, Migliorini D, Perkey E, Yost KE, Bhaduri A, Bagga P, et al. Single-Cell Analyses Identify Brain Mural Cells Expressing CD19 as Potential Off-Tumor Targets for CAR-T Immunotherapies. Cell 2020;183(1):126–42.e17 doi 10.1016/j.cell.2020.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Harrer DC, Simon B, Fujii S-i, Shimizu K, Uslu U, Schule G, et al. RNA-transfection of γ/δ T cells with a chimeric antigen receptor or an α/β T-cell receptor: a safer alternative to genetically engineered α/β T cells for the immunotherapy of melanoma. BMC cancer 2017;17(1):1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Krug C, Birkholz K, Paulus A, Schwenkert M, Schmidt P, Hoffmann N, et al. Stability and activity of MCSP-specific chimeric antigen receptors (CARs) depend on the scFv antigen-binding domain and the protein backbone. Cancer Immunology, Immunotherapy 2015;64(12):1623–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Birkholz K, Hombach A, Krug C, Reuter S, Kershaw M, Kämpgen E, et al. Transfer of mRNA encoding recombinant immunoreceptors reprograms CD4+ and CD8+ T cells for use in the adoptive immunotherapy of cancer. Gene Ther 2009;16(5):596–604 doi 10.1038/gt.2008.189. [DOI] [PubMed] [Google Scholar]
- 84.Hertzberg L, Vendramini E, Ganmore I, Cazzaniga G, Schmitz M, Chalker J, et al. Down syndrome acute lymphoblastic leukemia, a highly heterogeneous disease in which aberrant expression of CRLF2 is associated with mutated JAK2: a report from the International BFM Study Group. Blood, The Journal of the American Society of Hematology 2010;115(5):1006–17. [DOI] [PubMed] [Google Scholar]
- 85.Loh ML, Zhang J, Harvey RC, Roberts K, Payne-Turner D, Kang H, et al. Tyrosine kinome sequencing of pediatric acute lymphoblastic leukemia: a report from the Children’s Oncology Group TARGET Project. Blood 2013;121(3):485–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Mullighan CG, Collins-Underwood JR, Phillips LA, Loudin MG, Liu W, Zhang J, et al. Rearrangement of CRLF2 in B-progenitor–and Down syndrome–associated acute lymphoblastic leukemia. Nature genetics 2009;41(11):1243–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Roberts KG, Li Y, Payne-Turner D, Harvey RC, Yang Y-L, Pei D, et al. Targetable kinase-activating lesions in Ph-like acute lymphoblastic leukemia. New England journal of medicine 2014;371(11):1005–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Russell LJ, Capasso M, Vater I, Akasaka T, Bernard OA, Calasanz MJ, et al. Deregulated expression of cytokine receptor gene, CRLF2, is involved in lymphoid transformation in B-cell precursor acute lymphoblastic leukemia. Blood, The Journal of the American Society of Hematology 2009;114(13):2688–98. [DOI] [PubMed] [Google Scholar]
- 89.Yoda A, Yoda Y, Chiaretti S, Bar-Natan M, Mani K, Rodig SJ, et al. Functional screening identifies CRLF2 in precursor B-cell acute lymphoblastic leukemia. Proceedings of the National Academy of Sciences 2010;107(1):252–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Cario G, Zimmermann M, Romey R, Gesk S, Vater I, Harbott J, et al. Presence of the P2RY8-CRLF2 rearrangement is associated with a poor prognosis in non–high-risk precursor B-cell acute lymphoblastic leukemia in children treated according to the ALL-BFM 2000 protocol. Blood, The Journal of the American Society of Hematology 2010;115(26):5393–7. [DOI] [PubMed] [Google Scholar]
- 91.Chen I, Harvey RC, Mullighan CG, Gastier-Foster J, Wharton W, Kang H, et al. Outcome modeling with CRLF2, IKZF1, JAK, and minimal residual disease in pediatric acute lymphoblastic leukemia: a Children’s Oncology Group study. Blood 2012;119(15):3512–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Harvey RC, Mullighan CG, Chen I-M, Wharton W, Mikhail FM, Carroll AJ, et al. Rearrangement of CRLF2 is associated with mutation of JAK kinases, alteration of IKZF1, Hispanic/Latino ethnicity, and a poor outcome in pediatric B-progenitor acute lymphoblastic leukemia. Blood, The Journal of the American Society of Hematology 2010;115(26):5312–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Moorman AV, Schwab C, Ensor HM, Russell LJ, Morrison H, Jones L, et al. IGH@ translocations, CRLF2 deregulation, and microdeletions in adolescents and adults with acute lymphoblastic leukemia. J Clin Oncol 2012;30(25):3100–8. [DOI] [PubMed] [Google Scholar]
- 94.Palmi C, Vendramini E, Silvestri D, Longinotti G, Frison D, Cario G, et al. Poor prognosis for P2RY8-CRLF2 fusion but not for CRLF2 over-expression in children with intermediate risk B-cell precursor acute lymphoblastic leukemia. Leukemia 2012;26(10):2245–53. [DOI] [PubMed] [Google Scholar]
- 95.Yamashita Y, Shimada A, Yamada T, Yamaji K, Hori T, Tsurusawa M, et al. IKZF1 and CRLF2 gene alterations correlate with poor prognosis in Japanese BCR-ABL1-negative high-risk B-cell precursor acute lymphoblastic leukemia. Pediatric Blood & Cancer 2013;60(10):1587–92. [DOI] [PubMed] [Google Scholar]
- 96.Tasian SK, Doral MY, Borowitz MJ, Wood BL, Chen I-M, Harvey RC, et al. Aberrant STAT5 and PI3K/mTOR pathway signaling occurs in human CRLF2-rearranged B-precursor acute lymphoblastic leukemia. Blood, The Journal of the American Society of Hematology 2012;120(4):833–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Van Bodegom D, Zhong J, Kopp N, Dutta C, Kim M-S, Bird L, et al. Differences in signaling through the B-cell leukemia oncoprotein CRLF2 in response to TSLP and through mutant JAK2. Blood 2012;120(14):2853–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Qin H, Cho M, Haso W, Zhang L, Tasian SK, Oo HZ, et al. Eradication of B-ALL using chimeric antigen receptor–expressing T cells targeting the TSLPR oncoprotein. Blood, The Journal of the American Society of Hematology 2015;126(5):629–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Nadler LM, Ritz J, Hardy R, Pesando JM, Schlossman SF, Stashenko P. A unique cell surface antigen identifying lymphoid malignancies of B cell origin. J Clin Invest 1981;67(1):134–40 doi 10.1172/jci110005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Czuczman MS, Olejniczak S, Gowda A, Kotowski A, Binder A, Kaur H, et al. Acquirement of rituximab resistance in lymphoma cell lines is associated with both global CD20 gene and protein down-regulation regulated at the pretranscriptional and posttranscriptional levels. Clin Cancer Res 2008;14(5):1561–70 doi 10.1158/1078-0432.Ccr-07-1254. [DOI] [PubMed] [Google Scholar]
- 101.Awasthi A, Ayello J, van de Ven C, Elmacken M, Reggio C, Barth MJ, et al. Comparative Study of Obinutuzumab (GA101) Vs. Rituximab Against CD20+ rituximab-Sensitive and -Resistant Burkitt (BL) and Acute Lymphoblastic Leukemia (B-ALL): Potential Targeted Therapy in Patients with High Risk BL and Pre-B-ALL. Blood 2014;124(21):2251- doi 10.1182/blood.V124.21.2251.2251. [DOI] [Google Scholar]
- 102.Jabbour E, Richard-Carpentier G, Sasaki Y, Konopleva M, Patel K, Roberts K, et al. Hyper-CVAD regimen in combination with ofatumumab as frontline therapy for adults with Philadelphia chromosome-negative B-cell acute lymphoblastic leukaemia: a single-arm, phase 2 trial. Lancet Haematol 2020;7(7):e523–e33 doi 10.1016/s2352-3026(20)30144-7. [DOI] [PubMed] [Google Scholar]
- 103.Saini KS, Azim HA Jr, Cocorocchio E, Vanazzi A, Saini ML, Raviele PR, et al. Rituximab in Hodgkin lymphoma: is the target always a hit? Cancer Treat Rev 2011;37(5):385–90 doi 10.1016/j.ctrv.2010.11.005. [DOI] [PubMed] [Google Scholar]
- 104.Hu Y, Turner MJ, Shields J, Gale MS, Hutto E, Roberts BL, et al. Investigation of the mechanism of action of alemtuzumab in a human CD52 transgenic mouse model. Immunology 2009;128(2):260–70 doi 10.1111/j.1365-2567.2009.03115.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Angiolillo AL, Yu AL, Reaman G, Ingle AM, Secola R, Adamson PC. A phase II study of Campath-1H in children with relapsed or refractory acute lymphoblastic leukemia: a Children’s Oncology Group report. Pediatr Blood Cancer 2009;53(6):978–83 doi 10.1002/pbc.22209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Yang CG, Lang MF, Fu X, Lin H, Zhang LC, Ge GS, et al. Application of short hydrophobic elastin-like polypeptides for expression and purification of active proteins. 3 Biotech 2020;10(4):156 doi 10.1007/s13205-020-2139-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Qin H, Ramakrishna S, Nguyen S, Fountaine TJ, Ponduri A, Stetler-Stevenson M, et al. Preclinical Development of Bivalent Chimeric Antigen Receptors Targeting Both CD19 and CD22. Mol Ther Oncolytics 2018;11:127–37 doi 10.1016/j.omto.2018.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Grada Z, Hegde M, Byrd T, Shaffer DR, Ghazi A, Brawley VS, et al. TanCAR: a novel bispecific chimeric antigen receptor for cancer immunotherapy. Molecular Therapy-Nucleic Acids 2013;2:e105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Ahmed N, Brawley V, Hegde M, Bielamowicz K, Wakefield A, Ghazi A, et al. Autologous HER2 CMV bispecific CAR T cells are safe and demonstrate clinical benefit for glioblastoma in a Phase I trial. Journal for immunotherapy of cancer 2015;3(S2):O11. [Google Scholar]
- 110.Hegde M, Corder A, Chow KK, Mukherjee M, Ashoori A, Kew Y, et al. Combinational targeting offsets antigen escape and enhances effector functions of adoptively transferred T cells in glioblastoma. Molecular Therapy 2013;21(11):2087–101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Wang N, Hu X, Cao W, Li C, Xiao Y, Cao Y, et al. Efficacy and safety of CAR19/22 T-cell cocktail therapy in patients with refractory/relapsed B-cell malignancies. Blood 2020;135(1):17–27 doi 10.1182/blood.2019000017. [DOI] [PubMed] [Google Scholar]
- 112.Pan J, Zuo S, Deng B, Xu X, Li C, Zheng Q, et al. Sequential CD19–22 CAR T therapy induces sustained remission in children with r/r B-ALL. Blood 2020;135(5):387–91 doi 10.1182/blood.2019003293. [DOI] [PubMed] [Google Scholar]
- 113.Shah NN. The one-two punch (of CAR T cells). Blood 2020;135(5):303–4 doi 10.1182/blood.2019004272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Fousek K, Watanabe J, George A, An X, Samaha H, Navai SA, et al. Targeting Primary Pre-B Cell Acute Lymphoblastic Leukemia and CD19-Negative Relapses Using Trivalent CAR T Cells. Blood 2017;130(Supplement 1):4614.- doi 10.1182/blood.V130.Suppl_1.4614.4614. [DOI] [Google Scholar]
- 115.Fousek K, Watanabe J, Joseph SK, George A, An X, Byrd TT, et al. CAR T-cells that target acute B-lineage leukemia irrespective of CD19 expression. Leukemia 2020:1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Beatty GL, O’Hara MH, Lacey SF, Torigian DA, Nazimuddin F, Chen F, et al. Activity of Mesothelin-Specific Chimeric Antigen Receptor T Cells Against Pancreatic Carcinoma Metastases in a Phase 1 Trial. Gastroenterology 2018;155(1):29–32 doi 10.1053/j.gastro.2018.03.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Qian L, Li D, Ma L, He T, Qi F, Shen J, et al. The novel anti-CD19 chimeric antigen receptors with humanized scFv (single-chain variable fragment) trigger leukemia cell killing. Cell Immunol 2016;304–305:49–54 doi 10.1016/j.cellimm.2016.03.003. [DOI] [PubMed] [Google Scholar]