

Abstract
Introduction
Cannabinoids are a group of terpenophenolic compounds derived from the Cannabis sativa L. plant. There is a growing body of evidence from cell culture and animal studies in support of cannabinoids possessing anticancer properties.
Method
A database search of peer reviewed articles published in English as full texts between January 1970 and April 2021 in Google Scholar, MEDLINE, PubMed and Web of Science was undertaken. References of relevant literature were searched to identify additional studies to construct a narrative literature review of oncological effects of cannabinoids in pre-clinical and clinical studies in various cancer types.
Results
Phyto-, endogenous and synthetic cannabinoids demonstrated antitumour effects both in vitro and in vivo. However, these effects are dependent on cancer type, the concentration and preparation of the cannabinoid and the abundance of receptor targets. The mechanism of action of synthetic cannabinoids, (−)-trans-Δ9-tetrahydrocannabinol (Δ9-THC) and cannabidiol (CBD) has mainly been described via the traditional cannabinoid receptors; CB1 and CB2, but reports have also indicated evidence of activity through GPR55, TRPM8 and other ion channels including TRPA1, TRPV1 and TRPV2.
Conclusion
Cannabinoids have shown to be efficacious both as a single agent and in combination with antineoplastic drugs. These effects have occurred through various receptors and ligands and modulation of signalling pathways involved in hallmarks of cancer pathology. There is a need for further studies to characterise its mode of action at the molecular level and to delineate efficacious dosage and route of administration in addition to synergistic regimes.
Keywords: Cannabinoids, Cancer, Cannabidiol, Tetrahydrocannabinol, Cannabinoid receptors, Endocannabinoid system
Introduction
Since time immemorial, the Cannabis plant has been used as a source of fibre, herbal remedy, medicinal and religious purposes (Kalant 2001; Goncalves et al. 2020). In the mid-nineteenth century, O’Shaughnessy and Moreau reported positive effects of cannabis on muscle spasms, vomiting, convulsions, rheumatism, tetanus, and rabies (O’Shaughnessy 1843; Zuardi 2006). However, during the twentieth century, its utilisation in Western medicine started to decline as a result of political prejudices and economic interests rather than scientific or medical reasons (Zuardi 2006). Over recent years, cannabis and its derivatives have been used for treating chemotherapy induced nausea and vomiting, epilepsy and multiple sclerosis amongst other indications (Parker et al. 2011; Kleckner 2019). Increasing data from and in vivo studies have started to show evidence of cannabis in modulating signalling pathways involved in cancer cell proliferation, autophagy, apoptosis and inhibition of angiogenesis and metastasis (Velasco et al. 2016). Emerging reports have also indicated synergistic effects of cannabinoids in combination with antineoplastic drugs (Moreno et al. 2019; Dariš et al. 2019; Fogli et al. 2006; Velasco et al. 2012).
The cannabis plant has been termed as a “storehouse” of several pharmacologically relevant compounds (Andre et al. 2016). The unique qualities of each cannabis variety or chemovar are the result of varying concentrations of numerous classes of bioactive molecules, most notably, cannabinoids as shown in Fig. 1, terpenoids and flavonoids (Chakravarti et al. 2014). Cannabinoids interact directly with cannabinoid receptors, which include G-protein coupled receptors (cannabinoid receptor 1, CB1 and cannabinoid receptor 2, CB2), ligand-gated ion channels (i.e. vanilloid cell surface channels) and nuclear receptors (i.e. peroxisome proliferator-activated receptor gamma, PPARγ) (Moreno et al. 2019; Śledziński et al. 2018) comprising the endogenous endocannabinoid system (ECS) (Zou and Kumar 2018). Three major classifications of cannabinoids include phytocannabinoids (plant-based), such as Δ9-tetrahydrocannabinol (Δ9-THC) and cannabidiol (CBD), endocannabinoids (or endogenous cannabinoids) which include anandamide (AEA) and 2-arachidonolyglycerol (2-AG) and synthetic cannabinoids that mimic the cannabinoid groups (1) and (2) (Pertwee 2006; Lu and Mackie 2016). Endocannabinoids play a crucial role in mediating physiological functions including metabolic, cardiovascular regulation, reproduction, inflammatory response, immune system and analgesia (Guindon and Hohmann 2012; Kaur et al. 2016). AEA and 2-AG are degraded by fatty acid amide hydrolase (FAAH) and monoacylglycerol lipase (MAGL) enzymes (Pisanti et al. 2013). Modulation of their activity may have potential therapeutic implications and inhibitors are under active investigation as pharmaceuticals. Synthetic cannabinoids have been studied extensively and some have been shown to be highly bioactive than their natural counterparts, some common ones include WIN55, 212–2 (potent CB1 receptor agonist), JWH-018, JWH-073, JWH-133 (CB receptor agonists) and SR141716 or Rimonabant (CB1 receptor antagonist) (Morales et al. 2017), overview shown in Fig. 2.
Fig. 1.

The chemical structures of Cannabigerol (CBG), Cannabidiol (CBD), Tetrahydrocannabinol (Δ9-THC), Cannabichromene (CBC) and Cannabinol (CBN)-type neutral, varinic and acidic phytocannabinoids. More than 120 phytocannabinoids have been isolated from Cannabis sativa L. which can be distinguished into eleven chemical subtypes (Gonçalves et al. 2020; ElSohly 2017). Their common chemical features include a dibenzopyran ring and a hydrophobic alkyl chain (Morales et al. 2017). Aside from Δ9-THC and CBD, there has been a current focus on the therapeutic properties of some minor, varinic and acidic cannabinoids (Andre et al. 2016; Franco et al. 2020). Created with BioRender.com
Fig. 2.
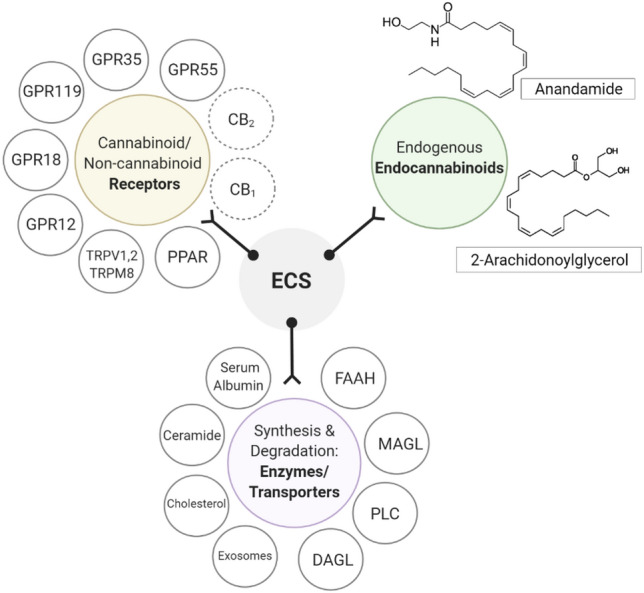
Overview of the components of the endocannabinoid system (ECS) which include endogenous endocannabinoids; Anandamide (AEA) and 2-Arachidonoylglycerol (2-AG), its major receptors classified into cannabinoid receptors 1 and 2, and non-cannabinoid receptors; GPR55, GPR35, GPR119, GPR18, GPR12, ion channels including transient receptor potential cation channel subfamily members; TRPM8, TRPV1, TRPV2, peroxisome-proliferator-activated receptors (PPAR). A third component of the system are its enzymes/transporters responsible for the synthesis and degradation of endocannabinoids including serum albumin, ceramide, cholesterol, diacylglycerol lipase (DAGL), phospholipase C (PLC), monoacylglycerol lipase (MAGL), fatty acid amide hydrolase (FAAH). Created with BioRender.com
Several studies have reported the varying affinities of phytocannabinoids for the classical CB1 and CB2 receptors with agonistic and antagonistic behaviours (Morales et al. 2017; Zhao and Abood 2013). However, it is now emerging that cannabinoids can interact with multiple orphan G-protein coupled receptors (GPCRs) including GPR12, GPR18, GPR35, GPR55, GPR119, opioid and serotonin receptors (Morales et al. 2017; Zhao and Abood 2013; Console-Bram et al. 2014; Brown et al. 2017; Soderstorm et al. 2017; Ferro et al. 2018; Guerrero-Alba 2019). The interaction of GPCRs is crucial for maintaining the ECS as it allows the production of endocannabinoids from cells through activation of Gq/11 or Gs proteins causing the activation of the cannabinoid receptor (Gyombolai et al. 2012). Furthermore, the downstream receptor-mediated effects of endocannabinoids also contribute to the plasticity of the ECS (Lu and Mackie 2016).
Since the first report of cannabinoids anticancer effects (Munson et al. 1975), there have been many studies investigating phytocannabinoids, endogenous and synthetic ones in multiple cancer models. Various signalling pathways and changes to internal conditions which favour antitumour activity by cannabinoids have been observed. CBD amongst other cannabinoids has shown to increase the de novo synthesis of ceramide through upregulation of a plethora of enzymes each catalysing specific biochemical steps. Ceramide synthases are one of the major group of enzymes involved and reports have revealed an upregulation of its six isoforms; CerS 1–6 (Ceramide Synthases 1–6) in cancer via cannabinoids (Gomez et al. 2002; Gustafsson et al. 2009; Schiffman et al. 2009). However, it is not clear whether specific isoform(s) upregulation correlates to the cancer type and whether this is also specific to the type of cannabinoid. An interesting finding from a report has shown siRNA-induced knockdown of ceramide synthase 1 (CerS1 isoform) prevented gemcitabine-induced caspase 9 activation (Senkal et al. 2007; Levy and Futerman 2010). This could be explored further when considering cannabinoids action synergistically with chemotherapy drugs as ceramide may have the ability to sensitize the cancer cells to chemotherapy agents. Another major area of cannabinoids action has been through modulating the cell cycle. In a recent report in gastric cancer cells, CBD-induced cell cycle arrest at the G0–G1 phase and retardation in this phase corresponded to a reduction in CDK2/cyclin E protein levels (Zhang et al. 2019). Apoptotic changes are prevalent in cannabinoids mechanism of action which include morphological changes to the cells and cytoplasmic vacuolization, an increase in cleaved caspase-3 and -9 levels and activation of the mitochondrial apoptotic pathway (Zhang et al. 2019; Schoeman et al. 2020). Endoplasmic reticulum (ER) stress which occurs following ceramide synthesis causes downstream apoptotic changes and increases in proapoptotic proteins, such as BAD and Bax, also resulting in an increase in reactive oxygen species (ROS) signalling (Zhang et al. 2019). Δ9-THC in glioma cells has shown to induce upregulation of the p8 protein (involved in ER stress and metastasis) via de novo synthesis of ceramide (Carracedo et al. 2006). From the literature available, it is evident that there is an interplay between cannabinoids downstream effects.
Overall cannabinoids induce apoptosis to inhibit proliferation, downregulate the vascular endothelial growth factor (VEGF) pathway affecting angiogenesis and dampen metastasis by inhibiting cell adhesion and migration through modifying matrix metalloproteinase 2, 9 (MMP2, 9), tissue inhibitor of matrix metalloproteinases 1 (TIMP1), inhibitor of DNA binding 1 (ID1) and inducing ER stress (Velasco et al. 2016). Cancer cells do not exist in isolation and the tumour microenvironment (TME) has also been an imperative target for cancer therapy as it can influence the propensity for tumour growth, metastasis and resistance to therapy. The TME is composed of a host of factors including cancer-associated fibroblasts (CAFs), immune and inflammatory cells, lymph and blood vasculature, neuroendocrine cells, and extracellular matrix (ECM) (Wang et al. 2017). Cancer stem cells (CSCs), a subpopulation of stem cells expressing CD44, CD24 and CD133, are tumorigenic with demonstrated resistance to certain chemotherapeutics and also play a role in metastasis (Yu et al. 2012). Reports have shown the involvement of cannabinoids in inhibiting CAFs and CSCs in prostate and breast cancer models (Sharma et al. 2014; Mohammadpour et al. 2017; Pietrovito et al. 2020). The aforementioned effects, however, occur at varying degrees which depend on the cancer cell line, the expression levels of cannabinoid receptors, the type of cannabinoid compound and dosage.
The aim of this review is to analyse pre-clinical work and outline previous and forthcoming clinical research studies exploring cannabinoids in cancer treatment. Below, we outline the research encompassing endogenous and non-endogenous cannabinoids in which we review the proposed mechanisms of action culminated from studies into various cancers and discuss the need for more clinical studies to explore the possible therapeutic efficacy of cannabinoids as a possible treatment for cancer.
Method
Research question
This narrative review was conducted of available literature reporting the treatment effects of all cannabinoids as either a single agent or co-administered with other antitumour therapies in all cancer types. The aim of this review is to analyse and evaluate pre-clinical and clinical research determining the use of cannabinoids as a potential anti-cancer therapy.
Search strategy and inclusion criteria
A broad electronic search was conducted on Google Scholar, MEDLINE, PubMed and Web of Science articles published in English between 1st January 1970 and 30th April 2021. Investigations of cannabinoids use in oncology clinical trials were searched using the database, clinicaltrials.gov.uk with the key words; “Cannabinoids and Cancer”, “Cannabis and Cancer”, “Tetrahydrocannabinol and Cancer”, “CBD and Cancer” and “THC and Cancer”. The literature search was performed by two independent researchers (N.M. and S.E.) and if any discrepancies were identified then these were resolved by a senior author (M.S.). The reference lists of all publications were screened for further relevant references. The free text search included articles citing both original research and literature reviews. Inclusion criteria encompassed all reports identifying cannabinoids use in pre-clinical cancer models which includes in vitro, in vivo and in ovo experimental models, as well as clinical research. In addition, reports of potential mechanisms of action and signalling pathways involved were also included. Where literature reviews were identified, the relevant cited studies were also identified and included for de novo analysis.
Data extraction and presentation
Two independent researchers (N.M. and S.E.) performed the data extraction. Primary research papers reporting half maximal inhibitory concentration (IC50) and concentrations where the described effects were observed in pre-clinical cancer models were included in separate tables for in vitro and in vivo investigations. Concentration values are presented as micro-molar concentrations (μM) with their standard deviation (S.D.), standard error (S.E), or range except when unreported in the original study.
Results
Mechanism of action and signalling pathways
The ECS is a complex system composed of different ligands, receptors and ion channels resulting in many signalling pathways subject to modulation from external cannabinoids as shown in Fig. 3. It is therefore no surprise that there remains ambiguity in its precise role within cancer pathophysiology (Wu 2019). Many pre-clinical studies and histological analysis of patient tumours, suggest that an upregulation in the CB1 and CB2 receptors, endogenous ligands and over-activation of the ECS correlates with more aggressive tumours (Dariš et al. 2019) although other reports have concluded the contrary (Jung et al. 2013; Tutino et al. 2019). Cancer is a heterogenous disease and current evidence should be interpreted on the basis that different tumour types have been shown to exhibit various levels of CB receptors as well as ECS components. The role of the endogenous endocannabinoids and CB receptors within each cancer system is specific to the underlying cancer, therefore conflicting data can be presented across different cancers. It has also been reported that some cannabinoids have shown oncological effects independent of known CB receptors (Moreno et al. 2019; Fogli et al. 2006) implying that there may be undiscovered cannabinoid receptors implicated in cancer pathophysiology.
Fig. 3.

Overview of the downstream activation and crosstalk of signalling pathways of cannabinoid and non-cannabinoid receptors. Activation of the cannabinoid receptors CB1 and CB2 (red arrows) via cannabinoids stimulate ERK1/2 signalling which activates p27 and p21 causing a decrease in cyclins D and E, cdc2 and cdk2 through an increase in pRb, leading to cell cycle arrest. Inhibition of the P13K pathway leads to a decrease in Akt which inhibits cell proliferation. Biosynthesis of ceramide takes place at the endoplasmic reticulum through a series of biochemical steps involving many enzymes which help to convert dihydroceramides (DhCers) into ceramide. An increase in ceramide level in turn increases the stress protein p8/Nupr1 and TRIB3 which activates upregulation of ATF4 and CHOP proteins. A decrease in Akt leads to a downregulation in mTORC1 signalling causing autophagy. Activation of TRPM8 (purple arrows) leads to an increase in ROS production which also induces ER stress. Stimulation of non-cannabinoid receptor GPR55 (blue arrows) through LPI via the subunit Gαq subunit stimulates the production of PLC to release Ca2+ and DAG which leads to the activation of MAPK/ERK signalling. This causes gene transcription by activation of transcription factors CREB and NF-κß. Gα12/13 subunit activates the RhoA/ROCK pathway which regulates PLC, actin cytoskeleton and p38/ATF2 activity. ATF2/p38 inhibits antiapoptotic proteins and enhances the interaction between Beclin-1 and Vps34 which is also inhibited by BCL-2 further enhancing ROS production by activation of the intrinsic apoptotic pathway (Velasco et al. 2012, 2016).Created with BioRender.com. TRPV1,2 transient receptor potential cation channel subfamily V member 1,2, TRPM8 transient receptor potential cation channel subfamily members (melastatin) 8, GPR55 orphan G-protein coupled receptor 55, ROS reactive oxygen species, ER endoplasmic reticulum, p8 protein p8 (Nuclear Protein 1, NUPR1), CHOP CCAAT/-enhancer-binding protein homologous protein, ATF4 activating transcription factor 4, TRIB3 tribbles pseudokinase 3, Akt protein kinase B, mTORC1 mammalian target of rapamycin C1, p21 cyclin-dependent kinase inhibitor 1, p27 cyclin-dependent kinase inhibitor 1B, CDK cyclin-dependent kinase, pRb retinoblastoma protein; Nuclear factor-kappaß (NF-κß), LPI Lysophosphatidylinositol, DAG diacylglycerol, BAD BCL2-associated agonist of cell death, ROCK rho-associated protein kinase, PLC phospholipase C
The characterisation of cannabinoids mechanism of action has been discerned from in vitro and in vivo studies. Reports of their oncological effects have been observed through modulating the hallmarks of cancer (Hanahan and Weinberg 2000, 2011) whilst ∆9-THC trends in inducing apoptosis and cytotoxicity through CB receptor-dependent pathways; CBD exhibits its activity via orphan GPCRs and non-GPRCs-mediated signalling (Velasco et al. 2012, 2016; Afrin et al. 2020).
Studies have reported positive upregulation of ceramide sphingolipid metabolism, leading to the subsequent arrest of the cell cycle and apoptosis via downstream activation of signals through extracellular regulated kinase (ERK) upon cannabinoid action (Calvaruso et al. 2012). Additional studies have also concluded ∆9-THC’s role in regulating sphingolipid metabolism via serine palmitoyl transferase (SPT) (Śledziński et al. 2018) and recent reports have concluded other enzymes of the metabolism of sphingolipids to be regulated by cannabinoids (Shaw et al. 2018). Dihydroceramides which are metabolic intermediates of the de novo synthesis pathway have been involved in the mechanisms of promoting autophagy-mediated cancer cell death (Hernández-Tiedra et al. 2016). ∆9-THC increases the dihydroceramide:ceramide ratio in the endoplasmic reticulum of glioma cells causing pre-apoptotic changes (Hernández-Tiedra et al. 2016).
Activation of the CB receptors causes the induction of the ER stress-related response and promotes the upregulation of the transcription factor p8 (Nupr1), this further simulates the following transcription factors, activating transcription factor 4 (ATF-4), C/EBP-homologous protein (CHOP) and pseudokinase tribbles-homologue 3 (TRIB3) (Velasco et al. 2016). The inhibitory interaction of TRIB3 and a pro-survival kinase Akt is favoured which leads to the inhibition of the mammalian target of rapamycin target 1 (mTORC1) favouring cell autophagy. Autophagy is upstream of apoptosis in cannabinoid-induced cell death as shown in studies where blocking autophagy prevented cannabinoid-induced apoptosis (Salazar et al. 2009; Vara et al. 2011). An increase in ceramide level has also been associated with ER stress in cannabinoid-induced apoptosis in tumour cells (Salazar et al. 2009). In addition, other environmental stimuli may also promote ER stress which can lead to the activation of the apoptotic pathway. These include a decrease in intracellular Ca2+, viral infections, chemotherapy agents and oxidative stress (Schröder and Kaufman 2005; Śledziński et al. 2018).
The mitogen-activated protein kinase (MAPK) pathway has also been reported in numerous studies to be involved in cannabinoid response. Serine/threonine protein kinases are mainly involved in this pathway and act to convert extracellular stress into different cellular responses including, cell cycle arrest, apoptotic cell death and cytokine production via phosphorylation. The involvement of the MAPK pathway in cancer is complex as its response to different stimuli can produce conflicting outcomes. Brief activation of the ERK cascade leads to cell survival and proliferation, whilst chronic activation is pro-apoptotic (Howlett 2005; Javid et al. 2016).
CBD has been demonstrated to affect a diverse set of cellular targets. First, it inhibits FAAH and FABP (Fatty Acid-Binding Protein). FAAH is responsible for the breakdown of anandamide, whilst FABP aids the transport of anandamide to from extracellular spaces to intracellular targets, such FAAH or nuclear PPAR. Both effects result in indirect activation of CB1 and CB2 receptors through increased extracellular concentration of anandamide (Lee et al. 2007; Pistis and O’Sullivan 2017). Second, CBD activates the 5-HT1A serotonin receptor, PPARγ and the transient receptor potential cation subfamily channels; TRPV1, TRPV2 and TRPA1. CBD is also an antagonist of GPR55, transient receptor potential cation channel subfamily M member 8 (TRPM8) and T-type Ca2+ channels. Finally, CBD has also been reported to inhibit adenosine reuptake via multiple proposed mechanisms (Lee et al. 2007; Ibeas Bih et al. 2015; McPartland 2018). Antagonization of GPR55 via CBD has been reported to reduce proliferation of pancreatic tumour cells and its activation has been reported to lead to metastasis in triple-negative breast cancer when stimulated by LPI (Zhao and Abood 2013; Ferro et al. 2018; Andradas et al. 2016; Falasca and Ferro et al. 2018; Pellati et al. 2018). Below we summarise pre-clinical studies which include both in vitro and in vivo experimental results in various cancer models with summaries included in Tables 1 and 2.
Table 1.
Pre-clinical in vitro studies encompassing various cannabinoids in cancer models
| Cancer Cell line | Cannabinoid (s) | Inhibitory concentrations | In vitro actions | References | |
|---|---|---|---|---|---|
| PDAC | MIA PaCa2, PANC-1, Capan-2, BxPC-3 | Δ9-THC, SR141716, SR144528 | 0–5 µM | Apoptosis via CB2 and p8, ATF4 and TRIB3 and caspase-3 activation | Carracedo et al. (2006) |
| AsPC-I, HPFA-II, PANC-I, BxPC-3 | CBD | 0–10 µM | Antiproliferative effects via GPR55 | Ferro et al. (2018) | |
| BRAIN | Human Glioblastoma, U373-MG | Δ9-THC, AEA, HU-210, WIN 55,212–2 | 100 nM–10 µM | Accelerated cell proliferation via EGFR and MMP | McAllister et al. (2011) |
| Human Glioblastoma, U878MG, U373MG | CBD, SR141716, SR144528 | 5–40 µM | Antiproliferative effects correlated to induction of apoptosis | Singer et al. (2015) | |
|
Human Glioblastoma multiforme, SF126, U87-MG, U251, SF188, U373-MG, Human GBM cultures |
Δ9-THC, WIN 55,212–2 | 0.1 nM-2 µM | Antiproliferative effects and increase of apoptosis | Ellert-Miklaszewska et al. (2021) | |
| Rat C6 glioma cells | WIN 55,212–2, WIN 55,212–3 | 1–30 µM | Cell viability reduction, morphological changes to cells | Matas et al. (2007) | |
| Rat C6 glioma cells | Δ9-THC, CBD, CBD-A, CBG, CBC, AM251, JWH-133, AM630, SR141716A, SR144528 | 0–50 µM | CBD most potent. CBD, CBG and CBD-A activated TRPV1 | Ligresti et al. (2006) | |
| Murine Neuroblastoma, N18TG2 | AEA | 1–5 µM | Apoptosis and decrease in cleavage of PARP-1 | Marcu et al. (2010) | |
| Human Astrocytoma, U87MG | Δ9-THC, SR141716 | 1–10 µM | Apoptosis and autophagy via ER stress | Salazar et al. (2009) | |
| Human Glioma cancer, U251, SF126, U87 | Δ9-THC, CBD | 0.1–10 µM | Inhibition of cell proliferation, apoptosis | Qamri et al. (2009) | |
| Human Glioblastoma, U87-MG, T98G | CBD | 0–20 µM | Decrease in cell invasion via MMP-9, TIMP-1, TIMP-4, u-PA, PAI-1, VEGF | Solinas et al. (2013) | |
| Human Glioma, T98G, U87MG, Murine Glioma, GL261 | CBD, Δ9-THC (Pure and BDS) | 0–20 µM | Increase in radiosensitivity associated with increase in apoptosis and autophagy | Scott et al. (2014) | |
|
Human Glioblastoma, U251, 3832, 387 Primary glioma stem cells (GSC) lines |
CBD | 0–5 µM | Activation of p-p38 pathway, downregulation of key stem cell regulators; Sox2, Id1 and p-STAT3 | Singer et al. (2015) | |
| Human Neuroblastoma, SK-N-SH, IMR-32, NUB-6 and LAN-1 | Δ9-THC, CBD | 0–50 µg/mL | Cell viability reduction and apoptosis | Fisher et al. (2016) | |
| Human Glioblastoma, U87MG, Glioblastoma patient derived stem cell like cells (GIC) | Δ9-THC, CBD | 0–5 µM | Very significant reduction of the GIC population, induction of apoptosis | López-Valero et al. (2018) | |
| Human Glioma cells, U87MG (U87), A172, SW1783, U373MG (U373), T98G (T98), SW1088, and LN405 | Δ9-THC, CBD, SR141716, SR144528 | 0.9–3 µmol/L | Reduction in cell viability and induction of apoptosis and autophagy | Torres et al. (2011) | |
| Human Glioma cells, GOS3, U87 MG (U87), A172, SW1783, U118 MG (U118), U373 MG (U373), T98G (T98), SW1088, CCF-STTG1 (CCF) and LN405 | Δ9-THC, SR141716, SR144528 | 0–2.5 µM |
Sensitive and resistance cell line determined via reduction in cell viability Increased Mdk expression confers resistance of glioma cells to Δ9-THC pro-autophagic and antitumoural action |
Lorente et al. (2011) | |
| BREAST | Human Breast adenocarcinoma, MDA-MB-231, MCF-7, murine mammary carcinoma, 4T1 | Δ9-THC | 0- 20 µM | No decrease observed in cell viability for all cell lines and low level of cannabinoid receptors | McKallip et al. (2005) |
| Human Breast adenocarcinoma, EVSA-T | Δ9-THC | 3 and 5 µM | Antiproliferative effects rely on JunD activity and participation of p8 | Caffarel et al. (2008) | |
| Human Breast adenocarcinoma, MDA-MB-231, T47D, murine breast cancer, TSAE-1 | Met-F-AEA, SR141716A | 2.5–20 µM | Reduction in cell viability in dose-dependent manner and decrease of tyrosine phosphorylation of FAK and Src | Santoro et al. (2009) | |
| Human Breast adenocarcinoma, MDA-MB-231, T47D, MCF-7 | SR141716 | 0.1–1 µM |
Cell cycle arrest, decreased expression of cyclins D and E Antiproliferative effect requires lipid raft/caveolae integrity to occur |
Sarnataro et al. (2005) | |
| Human Breast adenocarcinoma, EVSA-T, MDA-MB-231, MDA-MB-468, SKBR3, MCF-7, T-47D | Δ9-THC, SR141716, SR144528 | 1–12 µmol/L | Reduction in cell proliferation via the CB2 receptor, cell cycle arrest, induction of apoptosis | Caffarel et al. (2006) | |
| Human Breast adenocarcinoma, MDA-MB-231, MCF-7 | Δ9-THC, CBD, CBG, CBC, AM251, JWH-133, AM630, SR141716A, SR144528 | 0–50 µM | CBD apoptotic effect via activation of the CB2 receptor and TRPV1 | Ligresti et al. (2006) | |
| Human Breast adenocarcinoma, MDA-MB-231, MDA-MB-231-Luc, MDA-MB-468 | WIN 55,212–2, JWH-133, AM251, SR144528 | 0–10 µM |
All cell lines express both CB1 and CB2 receptors Inhibition of cell proliferation and migration via COX-2 signalling and apoptosis |
Hirao-Suzuki et al. (2020) | |
|
Human Breast adenocarcinoma, MDA-MB231 Murine mammary carcinoma, 4T1.2 |
CBD | 1.5 µM | Inhibition of cell proliferation and invasion through modulation of ERK and ROS, downregulation of Id-1 expression and upregulation of Id-2 | Nallathambi et al. (2018) | |
| Human Breast adenocarcinoma, MDA-MB-231, SKBR3, MCF-7, ZR-75–1 | CBD, AM251, AM630, Capazepine | 0–10 µM | Decrease in cell viability, autophagy and apoptosis via ER stress, inhibition of Akt, mTOR signalling | Lin et al. (2019) | |
| Human Breast adenocarcinoma, SUM159, MDA-MB-231-SCP2, MVT-1, murine mammary carcinoma, 4T1.2 | CBD | 3–15 µM | Cell proliferation decreased, inhibition of the epidermal growth factor (EGF)-induced cell proliferation, migration, and invasion | Grimaldi et al. (2006) | |
| Human Breast adenocarcinoma, MCF-7, Murine mammary carcinoma, 4T1 | JWH-015, SR141716, SR144528 | 0–10 µM | Decrease in cell viability, apoptosis and reduced ERK1/2 levels, effects were dependent in a non-Gαi -mediated, calcium-dependency | McAllister et al. (2011) | |
| Human Breast adenocarcinoma, MDA-MB-231 | AEA, AM251 | 0–0.5 µM | Reduction in CD44+/CD24−/low/ESA+ cancer stem cell (CSC) invasiveness | Mohammadpour et al. (2017) | |
| Human Breast adenocarcinoma, MDA-MB-231 | CBDA, GSK0660, GW501516, ST-247 | 1–50 µM | CBDA inhibits PPARβ/δ mediated transcriptional activation and AP-1 | Gazzerro et al. (2010) | |
| Human Breast Cancer, MDA-MB-231, MCF-7 | CBD | 1–50 µM | Co-administration of CBDsol and paclitaxel or docetaxel showed a synergistic effect | Fraguas-Sánchez et al. (2020) | |
| GASTROINTESTINAL | Human Colon cancer, DLD-1, CaCo-2, SW620 | SR141716 | 0.1–20 µM | Reduction in cell proliferation and cell cycle arrest | Aviello et al. (2012) |
| Human Colon adenocarcinoma, Caco-2, HCT 116 | CBG, AM251, AM630, AMTB (TRPM8 antagonist), CBD, CBDV, CBC | 1–50 µM | Apoptosis, increase in ROS production and upregulation of CHOP expression | Borelli et al. (2014) | |
| Human Colorectal carcinoma, DLD-1, HCT116 | CBD BS (botanical substance), CBD, AM630, SR141716, SR144528 | 0.3–5 µM | Antiproliferative effects, no effect on cell viability | Romano et al. (2014) | |
| Human Colorectal cancer, Caco-2 | CBD | 0.1–10 µM | PhysO2 cells significantly more sensitive to antiproliferative effects of CBD than AtmosO2 | Macpherson et al. 2014 | |
| Human Colon cancer, DLD-1 | SR141716 | 0.1–10 µM | Inhibition of cell proliferation at higher concentrations | Gazzerro et al. (2010) | |
| Human Colon cancer, SW480 | CBD, WIN 55,212–2 | 0–15 µM | Induction of cellular ACPP, DUSP1, DUSP10, cleavage of PARP, Apoptosis | De Petrocellis et al. (2013) | |
| Human colorectal carcinoma, Caco-2, HCT116 | CBD, SR141716, AM251, SR144528, AM630, GW9662, Capsazepine | 0.01–10 µM | Reduction in cell viability and expression of phospho-Akt | Aviello et al. (2012) | |
| Human Colon cancer, HCT116, SW48 | SR141716 | 0–20 µM | Inhibition of cell growth, increase of caspase-3 and cleavage of PARP | Proto et al. (2017) | |
|
Human Colon cancer, HCT116 and DLD-1 Organoids |
SR141716 | 0.1–20 µM | Reduction in colon CSCs proliferation and tumour differentiated cells | Fiore et al. (2018) | |
| Human Hepatocellular carcinoma, HepG2, HuH-7 | Δ9-THC, JWH-015, SR141716, SR144528 | 1–8 µM | Reduction in cell viability occurred via CB2 receptor and autophagy | Vara et al. (2011) | |
| Human Hepatocellular carcinoma, BEL7402 | WIN 55, 212–2, AM630, JWH-015 | 0, 5 or 10 µM | CB2 mediated downregulation of phosphorylated ERK1/2 | Xu et al. (2016) | |
| Human Gastric adenocarcinoma, AGS | AEA, Meth-AEA (R- ( +)), CP 55,940 | 0.5–5 µM | Concentration-dependent effects in cell morphology and loss changes | Ortega et al. (2016) | |
| Gastric cancer, SGC7901, AGS cells | WIN 55,212–2 | 5 µM | Inhibition of cell migration, invasion and EMT | Xian et al. (2016) | |
| PROSTATE | Human Prostate Cancer, PC-3 | Δ9-THC, AM251, WIN55,212–2 | 0.5–10 µM | Reduction in cell viability and apoptosis | Ruiz et al. (1999) |
| Human Prostate Cancer, LNCaP, DU145, PC-3 | AEA | 1–10 µM | Decrease of EGFR levels in all cell lines via CB1 leading to an inhibition of EGF-stimulated growth | Mimeault et al. (2003) | |
| Human Prostate Cancer, LNCaP | MET-AEA, HU-210, JWH-015, SR141716, SR144528 | 0.05–5 µM | Involvement of PI3K pathway and modification of androgen receptor expression | Sanchez et al. (2003) | |
| Human Prostate carcinoma, LNCaP, PC3 | WIN-55,212–2, SR141716, SR144528 | 1–30 µM | Induction in p27/KIP1 and downregulation in cyclin and CDK levels. Upregulation of ERK1/2 and inhibition of PI3k/Akt pathways | Sarfaraz et al. (2006) | |
| Human Prostate cancer, LNCaP, 22RV1, DU-145, PC-3 | CBC, CBD, CBG, CBN, CBDA, CBGA, CBDV, CBGV, THC, THCA, THCV, THCVA | 1–10 µM | Decrease in cell viability and activation of the intrinsic apoptotic pathway | De Petrocellis et al. (2013) | |
| Human Prostate adenocarcinoma, PC-3, Primary cultures; BPH, LGG, HGG, PrC | AEA, 2-AG, Methanandamide (AM-356), SR141716 | 2.5, 5 and 10 µM | Cell cycle arrest and induction of apoptosis | Orellana-Serradell et al. (2015) | |
| Human Prostate cancer, LNCaP, PC-3 | WIN 55,212–2, SR141716, SR144528 | 0- 10 µM | WIN prevents neuroendocrine differentiation by downregulation of PI3K/Akt/mTOR signalling | Morell et al. (2016) | |
| LUNG | Human Lung carcinoma, NCI-H292 | Δ9-THC, AEA, HU-210, WIN 55,212–2 | 0.1–10 µM | Increase in cell proliferation dependent on EGFR and MMP | Hart et al. (2004) |
| Human NSCLC, EGF-induced, A549, SW-1573 | Δ9-THC | 1–20 µM | Apoptosis and inhibition of proliferation via EGF-induced phosphorylation of ERK1/2, JNK1/1 and Akt | Preet et al. (2008) | |
|
Human Lung adenocarcinoma, A549, H460 Primary non-small-cell lung carcinoma cells |
CBD, AM251, AM630, Capsazepine, NS-398 | 0–10 µM | Decrease in the viability of the cells and upregulation of COX-2 and PPAR-γ expression, PGE2, PGD2, and 15d-PGJ2 | Ramer et al. (2013) | |
| Human NSCLC; A549 (epithelial), CALU1 (mesenchymal) | JWH-015, SR144528 | 0–5 µM | Decreased migratory and invasive abilities via reduction in FAK, VCAM1, MMP2 | Ravi et al. (2016) | |
| Human Lung cancer; A549 | WIN 55,212–2 | 5–20 µM | Decline in cell viability due to apoptosis | Müller et al. (2017) | |
| BLOOD | Human Leukaemia; CEM (acute lymphoblastic), HEL-92 (erythroblastic), HL60 (acute promyelocytic), MOLT-4 (acute lymphoblastic) and PBMCs | Δ9-THC | 0–100 µM | Cell death via activation of MAPK | Powles et al. (2005) |
| Human Leukaemia, Jurkat, MOLT-4 and murine lymphoma, EL-4 | CBD, SR141716A, SR144528, CAPZ | 0- 10 µM | Significant reduction in cell viability and apoptosis through the CB2 receptor | McKallip et al. (2006) | |
| Human Myeloma, U266, U266-LR7, RPMI, RPMI-LR5, MM1.S, MM1.R | WIN 55,212–2 | 5–50 µM | Apoptosis | Barbado et al. (2017) | |
| Human T acute lymphoblastic leukaemia, Jurkat | CBD | 0.01–10 µM | Reduction in cell viability and cell cycle changes | Kalenderoglu et al. (2017) | |
| SKIN | Melanoma, A375, MelJuso and murine melanoma, B16 | Δ9-THC, WIN-55,212–2, SR141716, SR144528 | 0.5–1 µM | Reduction in cell viability, angiogenesis, and metastasis via CB receptors | Blázquez et al. (2006) |
| Human Melanoma, CHL-1, A375, SK-MEL-28BD | Δ9-THC, CBD | 0–10 µM | Decrease in cell viability | Armstrong et al. (2015) | |
| Murine squamous, non-melanoma skin cancer; JWF2 | AEA, AMG9810, AM251, AM630 | 2.5- 40 µM | Reduction in cell viability and apoptosis via ER stress | Soliman et al. (2016) | |
| Human renal carcinoma, 786-O, SMKT-R2, SMKT-R3, Caki-2, RCC-6, 769-P, Caki-1, ACHN | WIN 55,212–2, JWH-133, SR141716A, AM630 | 0–25 µM | Reduction in cell proliferation and induction of apoptosis | Khan et al. (2018) | |
| Human ovarian cancer, SKOV-3 | CBD | 10–50 µM | Inhibition of cell proliferation | Fraguas-Sánchez et al. (2020) | |
| Rat Adrenal Gland; PC12 cells | DHA-DA, AEA | 0–80 µM | NOS activation, increased Ca2+ signalling leading to apoptosis via GPR55 activation | Akimov et al. (2021) |
MET-AEA (methanandamide, non-hydrolyzable analogue of AEA), AEA (anandamide), DHA-DA (N-docosahexaenoyl dopamine), AM251 (CB1 antagonist), HU-210 (CB1 agonist), JWH-015 (CB2 agonist), JWH-133 (CB2 agonist), WIN 55,212–2 (CB1 agonist), SR141716 (CB1 inverse agonist), SR144528 (CB2 inverse agonist), N-oleoylethanolamine (NOE) (acidic ceramidase inhibitor), LY294002 (PI3K inhibitor), PD98059 (ERK inhibitor), PBMCs (peripheral blood mononuclear cells), AM630 (CB2 antagonist), GW9662 (PPAR-γ antagonist), GSK066 (PPARβ/δ antagonist), GSK501516 (PPARδ antagonist), AMG9810 (TRPV1 antagonist)
Table 2.
Pre-clinical in vivo studies encompassing various cannabinoids in cancer models
| In vivo model | Cannabinoid (s) | Observed changes | References | |
|---|---|---|---|---|
| PDAC |
Immunodeficient nude mice and human PDAC cell lines MIA PaCa-2, PANC-1, Capan-2, BxPC-3 |
Δ9-THC, JWH-133, WIN-55,212–2 | Reduction in growth of tumour and induction of apoptosis via activation of the p8-ATF-4TRB3 proapoptotic pathway | Carracedo et al. (2006) |
| KPC PDAC mouse model (mutations in KRAS, PanIN, TP53) mice with homozygous deletion of GPR55 created KPCG strain | CBD | KPC mice treated with combination of CBD and GEM survived longer than vehicle or GEM alone | Ferro et al. (2018) | |
| BRAIN | Athymic female CD-1 nude mice and human glioma U87, U373 cell lines | CBD, SR141716, SR144528 | CBD significantly inhibited the growth of tumours | Massi et al. (2004) |
| Nude mice and human astrocytoma U87MG | Δ9-THC | Autophagy and apoptosis | Salazar et al. (2009) | |
| Female C57BL/6 and murine glioma GL261 | CBD, Δ9-THC (Pure and BDS) | Triple combination of CBD, Δ9-THC and irradiation significantly reduced tumour growth | Scott et al. (2004) | |
| Female Athymic (nu/nu) mice and human glioblastoma U251 and primary glioma stem cells 3832, 387 | CBD | Increase in the survival rate of mice bearing GSC xenografts | Singer et al. (2015) | |
| Immunodeficient (NOD/SCID) mice and human neuroblastoma SK-N-SH cell lines | Δ9-THC, CBD | Reduction in the growth of tumours and increase in activated caspase-3 | Fisher et al. (2016) | |
| Nude mice and human glioblastoma U87MG cell line | Δ9-THC, CBD | Reduction in tumour growth | López-Valero et al. (2018) | |
| Nude mice and U87, T98 cell lines | Δ9-THC, CBD | Reduction in tumour growth more significant when combined with temozolomide (TMZ) | Torres et al. (2011) | |
| Nude mice and U87, T98 cell lines | Δ9-THC, SR141716, SR144528 | Silencing of Mdk sensitizes cannabinoid resistant tumours to Δ9-THC anticancer action, although no effect on tumour growth | Lorente et al. (2011) | |
| BREAST | Female adult BALB/c and SCID-NOD mice and murine mammary carcinoma 4T1 | Δ9-THC | Increase in tumour growth and metastasis due to inhibition of specific antitumor immune response | McKallip et al. (2005) |
|
Male athymic mice and human breast adenocarcinoma, MDA-MB-231, MCF-7 |
Δ9-THC, CBD, CBG, CBC, SR141716A, SR144528 | CBD inhibited tumour growth and reduced lung metastasis | Ligresti et al. (2006) | |
| Male C57BL/6 N mice and murine mammary carcinoma, TSAE-1 | Met-F-AEA, SR141716A | Reduction of metastatic nodes in mice | Santoro et al. (2009) | |
| Female adult CD1 nude mice and human breast adenocarcinoma, MDA-MB-231 | SR141716 | Reduction in tumour volume | Sarnataro et al. (2005) | |
| Severe combined immunodeficient CB-17 mice and human breast adenocarcinoma, MDA-MB-231/luc/486 | WIN 55,212–2, JWH-133 | 40–50% reduction in tumour burden, 65–80% reduction in lung metastases | Hirao-Suzuki et al. (2020) | |
| Female BALB/cfC3H mice and murine mammary carcinoma 4T1 | CBD | Significant reduction of primary tumour mass and size and lung metastatic foci | Shrivastava et al. (2011) | |
| Female BALB/c and FVB mice and murine mammary carcinoma 4T1 | CBD | Reduction in the growth of tumours and vascularity and inhibition of lung metastasis | Grimaldi et al. (2006) | |
| Female BALB/cfC3H mice and murine mammary carcinoma 4T1 | JWH-015 | Significant reduction in primary tumour burden and metastasis | McAllister et al. (2011) | |
| COLORECTAL | Male C57BL/6 N mice, chemically induced colon cancer | SR141716 | Inhibition of tumour growth and reduction in ACF induced colon cancer | Santoro et al. (2009) |
| Male athymic (nu/nu) mice and human hepatocellular carcinoma, HepG2 and HuH-7 | Δ9-THC, JWH-015, SR-141716, SR144258 | Reduction of tumour growth and ascites | Vara et al. (2011) | |
| Male ICR mice and human colorectal carcinoma, Caco-2 and HCT116 | CBD | Reduction in ACF, polyps and tumour formation in AOM model | Aviello et al. (2012) | |
| Male ICR and athymic nude female mice and human colon adenocarcinoma, Caco-2 and HCT 116 | CBG, AM-251, AM-630 | CBG inhibited colon cancer growth | Borelli et al. (2014) | |
| Male ICR and athymic nude mice and human colorectal carcinoma, DLD-1 and HCT 116 | CBD, CBD BS | Reduction of AOM induced preneoplastic lesions and overall tumour growth | Romano et al. (2014) | |
| Female SCID mice and human colon cancer, HCT116 and SW48 | SR141716 |
Significant reduction in tumour growth Destabilization of the nuclear localization of β-Catenin |
Proto et al. (2017) | |
| PROSTATE | Male MR-1 nude mice and prostate carcinoma, LNCaP, 22RV1, DU-145 and PC-3 | CBC, CBD, CBG, CBN, CBDA, CBGA, CBDV, CBGV, THC, THCA, THCV, THCVA BDS | Reduction of the LNCaP xenograft growth | De Petrocellis et al. (2013) |
| Male athymic nude-FOxn1 (nu/nu) mice and human prostate cancer LNCaP | WIN 55,212–2, SR-141716, SR-144528 | Reduction in rate of growth and size of tumours | Morell et al. (2016) | |
| LUNG | Male C57BL/6 (H-2b) and BALB/c mice (H-2d) and murine Lewis/alveolar cell lung carcinoma | Δ9-THC, SR141716, SR144528 | Increase in the growth of the 3LL and L1C2 tumors in vivo | Zhu et al. (2000) |
| SCID CB-17 mice and human NSCLC, EGF-induced, A549, SW-1573 | Δ9-THC, WIN 55,212–2, JWH-133 | Inhibition of tumour growth and reduction in lung metastasis | Preet et al. (2008) | |
| Female NMRI (nu/nu) mice and human Lung adenocarcinoma, A549, H460 | CBD, AM-251, AM-630 | Reduction in tumour growth | Ramer et al. (2013) | |
| FVB mice and human Non-small cell lung cancer (NSCLC); A549, CALU1. Murine ED1 | JWH-015, SR144528 | Reduction in tumour growth and metastatic lesions | Ravi et al. (2016) | |
| BLOOD | Female adult mice C57BL/6 | CBD, SR141716A, SR144528 | Reduction in tumour growth | McKallip et al. (2006) |
| SKIN | C57BL/6 nude mice and murine melanoma, B16 cell line | Δ9-THC, WIN-55,212–2, SR141716, SR144528 | Decrease in tumour growth, proliferation, angiogenesis, and metastasis | Blázquez et al. (2006) |
| Male athymic nude (nu/nu) mice and human melanoma, CHL-1, A375, SK-MEL-28BD cell lines | Δ9-THC, CBD | Reduction in tumour growth | Ramer et al. (2013) | |
| NOD/scid/IL-2R gammae null (NSG) mice and human myeloma, U266, U266-LR7, RPMI, RPMI-LR5, MM1.S, MM1.R cells | WIN 55,212–2 | Reduction in tumour growth | Barbado et al. (2017) | |
| Female C57B6 mice and human rhabdomyosarcoma, RD, JR1, RH6, RH2 (ERMS) and RH30, RH4, RH41, RH3, and RH28 (ARMS) | AM251 | Abrogates lung metastasis formation | Marshall et al. (2011) |
Pancreatic adenocarcinoma
In vitro
A study analysing the in vitro effects of synthetic receptor agonists of CB1 and CB2, WIN55, 212–2, ACEA and JWH-015 found they each induced a high level of apoptosis of MIA PaCa-2 cells (Console-Bram et al. 2014). The same study showed that a CB1 antagonist, N-(piperidin-1-1yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide (AM251), induced apoptosis and transcriptional changes of the genes involved in the janus kinase/signal transducers, activators of transcription signalling network (JAK/STAT) and MAPK signalling pathways in the MIA PaCa-2 pancreatic cancer cell line through activation independent of the CB1 receptor-independent pathways (Fogli et al. 2006). AM251, which expresses molecular similarities with cyclo-oxygenase-2 (COX-2) inhibitor celecoxib, demonstrated a synergistic interaction with 5-fluorouracil (5-FU) increasing their anti-cancer activity when administered in appropriate ratios as demonstrated by a combination index of 0.52 (Fogli et al. 2006).
Dando et al. report arachidonoyl cyclopropylamide (ACPA) and GW, CB1 and CB2 selective agonists, respectively, inhibited proliferation and invasion of PANC-1 cells (Dando et al. 2013). Activation of the receptors via cannabinoid receptor agonists showed an elevation in 5′ adenosine monophosphate-activated protein kinase (APMK) activation via a ROS-dependent increase of AMP/ATP ratio promoting cell autophagy and subsequent inhibition of cell growth (Dando et al. 2013; Brandi et al. 2013). ∆9-THC has been shown to induce a reduction in cell viability via apoptosis in a dose-dependent manner, specifically via the de novo synthesized ceramide up-regulation of the p8 and ATF-4, TRIB3 ER stress genes in MIA PaCa-2 and PANC-1 cells (Carracedo et al. 2006). The p8 protein has been shown to increase with ceramide treatment and potentiates anticancer effects (Javid et al. 2016). In support of this, MIA PaCa-2 cells treated with ∆9-THC caused an increase in p8 mRNA levels in vitro. Knockdown of the p8 gene prevented apoptosis by ∆9-THC in these cells (Carracedo et al. 2006). In addition to p8 and TRIB3 stress-related genes, further ER stress-inducing genes have been identified and associated with apoptosis, such as CHOP and ATF-4, where mRNA levels were elevated following ∆9-THC treatment (Ohoko et al. 2005).
Cannabinoids in combination with chemotherapy agents have shown promising results in pancreatic cancer cell line studies. One study reported the increase in gemcitabine activity by synergism with CB1 and CB2 receptor ligands by a NF-κß-dependent mechanism (Donadelli et al. 2011). This synergistic inhibition of tumour growth was most marked in gemcitabine-resistant cell lines (Donadelli et al. 2011). Gemcitabine increased cannabinoid-induced autophagy through a ROS-mediated mechanism and cannabinoids enhanced the apoptotic effect of gemcitabine (Donadelli et al. 2011). Ferro and co-workers reported the anticancer effects of blocking the putative GPR55 receptor in pancreatic cancer cells via CBD. A cross between GPR55 homozygous knockout and mice which do not harbour the TP53 mutation did not reveal any statistical difference in survival. Investigators analysed the possible role that p53 may play in regulating GPR55. In pancreatic ductal adenocarcinoma cell lines, they report a negative regulation of GPR55 with TP53 status, where overexpression of wild-type p53 in the AsPC-1 cell line (harbouring a TP53 mutation) caused a reduction in GPR55 expression. Further analysis revealed the negative regulation was through modulation of the micro-RNA miR34b-3p. Pharmacological inhibition of GPR55 via CBD in various pancreatic cell lines, inhibited anchorage-dependent growth. Treatment with CID16020046 (CID), an antagonist of GPR55, revealed similar results in AsPC-1 and HPFA-II and cell cycle arrest at the G1–S phase in PANC-1 and HPFA-II in a dose-dependent manner. Cyclin D1, activation of tumour-suppressor protein (RB) was also reduced in CBD treatment and an inhibition of MEK/ERK and ERK-dependent pathways was also observed. The study demonstrates a novel pathway by which gemcitabine may be potentiating anticancer effects through inhibiting GPR55 via CBD antagonization (Ferro et al. 2018).
In vivo
Administration of ∆9-THC at 15 mg/kg/day into a xenograft model of MIA PaCa-2 pancreatic tumour growth showed a reduction in the tumour burden (Carracedo et al. 2006). A synthetic cannabinoid, WIN55, 212–2 was found to increase the expression of downstream targets of the ER stress-related pathway involved in apoptosis in pancreatic cancer in comparison to healthy controls, demonstrating apoptotic selectivity effect of cannabinoids to cancer cells (Carracedo et al. 2006).
The role of other cannabinoid receptors including GPR55 has been speculated to be involved in regulating many cancer types including pancreatic cancer. A study by Ferro et al. revealed genetic ablation of GPR55 in a KPC mouse model of pancreatic ductal adenocarcinoma (PDAC) significantly prolonged survival and KPC mice treated with CBD and gemcitabine as a combination treatment survived three times longer than control or gemcitabine single treatment (Ferro et al. 2018). Immunohistochemistry analysis of the tumours revealed CBD inhibition of GPR55 affected signalling pathways involved in gemcitabine resistance. CBD was able to counteract the effect of gemcitabine on ERK phosphorylation and downregulated the enzyme’s ribonucleotide reductases 1 and 2 (RRM1/2), a marker for gemcitabine resistance (Ferro et al. 2018). In line with this, gemcitabine-treated tumours from KPC mice expressed high levels of RRM1 and reduced levels were observed in KPCG mice upon treatment with CBD (Ferro et al. 2018). The counteractions of CBD on gemcitabine only occurred when both drugs were administered together, suggesting synergistic effects of CBD on gemcitabine’s mode of action in vivo (Ferro et al. 2018). Donadelli et al. also reported an enhanced effect with combination therapy. CB1 antagonist, Rimonabant, combined with gemcitabine reduced tumour growth when compared to single therapy in vivo (Donadelli et al. 2011). An increase in ROS and autophagy pathways were observed which may explain the synergistic effects they observed (Donadelli et al. 2011).
The translation of preclinical data to the clinic remains to be somewhat unclear as many factors in cannabinoids pharmacokinetics, bioactivity and efficacy remain undetermined (Ladin et al. 2016; Millar et al. 2018). In addition, their low aqueous solubility and poor stability (sensitivity to light, temperature and oxidation) make developing effective formulations a problem (Fraguas-Sánchez et al. 2020). The route of cannabinoid administration remains uncertain as the oral bioavailability is very low and is subject to a significant first-pass effect in the body (Millar et al. 2018). Therefore, alternative routes of administration are required, although it has been reported that intratumour (IT) administration of low doses of cannabinoids has improved efficacy of the drug as well as survival (Ngwa et al. 2017, 2018; Yasmin-Karim et al. 2018). Successful administration has been reported when cannabinoids were combined with radiotherapy in treating pancreatic cancer (Yasmin-Karim et al. 2018).
A recent study has reported the use of CBD and ∆9-THC inhibited proliferation of pancreatic cancer and stellate cells. PDL-1 (a key target for immune checkpoint blockade) expression was reduced in mice tumours via the PAK-1-dependent pathway (p-21 activated kinase 1) activated by Kirsten rat sarcoma (KRAS). Their findings suggest a novelty for the cannabinoids in which KRAS, an undruggable target expressed in many lethal cancers can be supressed through targeting PAK1 and the suppression of PDL-1 could be enhanced for immune checkpoint blockade therapy in pancreatic cancers (Yang et al. 2020).
Brain cancer
In vitro
Investigation into human glioma cell lines U87 and U373 administered with CBD led to a decrease in mitochondrial oxidative metabolism, cell viability and antiproliferative effects correlated to induction of apoptosis (Massi et al. 2004). Solinas et al. investigated CBD in U87-MG and T98G glioma cell lines and reported inhibition of cell proliferation and invasiveness, a downregulation of ERK and Akt signalling and a decrease in the hypoxia-inducible factor HIF-1α expression (Solinas et al. 2013). In the following neuroblastoma cell lines, SK-N-SH, IMR-32, NUB-6 and LAN-1, CBD and ∆9-THC treatment induced antitumorigenic activity by decreasing cell viability and invasiveness, arrest of the cell cycle at the G1/G0 phase and an increase in activated caspase-3, albeit CBD was more potent in these effects when compared to ∆9-THC (Fisher et al. 2016). Salazar et al. investigated ∆9-THC in the astrocytoma cell line U87MG and in vivo where they report autophagy induction via the upregulation of p8 leading to apoptosis and inhibition of Akt and mTORC1 (Salazar et al. 2009).
A recent study has reported in the following human glioma cell lines A172, U251, U87 MG, U118 MG and LN18, CBD induced autophagic rather than apoptotic cell death. Specifically, CBD caused mitochondrial dysfunction and lethal mitophagy arrest mechanistically via TRPV4 with an influx of calcium (Huang et al. 2021). Further analysis revealed ER stress and in particular the ATF4-DDIT3-TRIB3-AKT-MTOR axis downstream of TRPV4 was involved in CBD’s mitophagy effect. Combination of CBD and temozolomide (TMZ) in neurosphere cultures and mouse models conveyed synergistic effects in reducing tumour burden and improving survival rates (Torres et al. 2011). Their findings suggest a novel TRPV4-CBD-mitophagy pathway in glioma and combination of CBD and TMZ as a potential to explore in future clinical studies. Additionally, Vrechi and colleagues show CBD stimulates autophagy signal transduction via crosstalk of ERK1/2 and AKT kinases and that CBD-induced autophagy was reduced in presence of CB receptors and TRPV1 receptor antagonists, AM251, AM630 and capsazepine in neuroblastoma and murine astrocyte cell lines (Vrechi et al. 2021).
Kolbe et al. recently investigated the effects of cannabinoids in glioblastoma multiforme (GBM) cells derived from primary human tumour samples and to identify possible receptors involved. Their findings revealed ∆9-THC reduced the number of Ki67 immuno-reactive nuclei, through GPR55. Their findings suggest that the sensitivity of cannabinoids and receptor-dependent signalling pathways should be considered to reflect the heterogeneity amongst GBM forms which is critical for when evaluating this translationally to clinic (Kolbe et al. 2021). Mutation-driven cancers are important to take into account when tailoring specific treatments. In a recent paper, Ellert-Miklaszewska et al. investigated the use of synthetic cannabinoids in GBM which have frequent TP53 or PTEN genetic defects rendering it from chemotherapy treatments. Their experimental work showed synthetic cannabinoids not only reduce tumour cells but that p53 could also act as an activator or inhibitor of autophagy and apoptosis and this depends on subcellular localisation and the mutant variant of p53 (Ellert-Miklaszewska et al. 2021).
In vivo
In a glioma mouse model treated with CBD daily at 0.5 mg/mouse, Massi and colleagues reported a significant reduction in xenografted human U87 tumour growth in vivo (Massi et al. 2004). A further study investigating CBD’s action in tumours from derived glioma stem cells (GSCs) which known to be resistant to therapies, reported in vivoan increase in the production of ROS leading to the inhibition of cell survival and an increase in the survival rate of mice bearing the GSC xenograft (Singer et al. 2015). They also observed activation of the p-p38 pathway and a downregulation of stem cell regulators including Sox2, Id-1 (a transcription factor involved in cell growth, senescence and differentiation) and p-STAT3 which inhibited the self-renewal of the cells (Singer et al. 2015). Although CBD inhibited glioma progression, a fraction of therapeutic resistance to CBD in a subset of glioma cells was due to the upregulation of antioxidant response genes (Singer et al. 2015). SK-N-SH neuroblastoma cell line induced in nude mice treated with CBD and ∆9-THC led to a reduction in tumour burden and an observed increase in activated caspase-3 (Fisher et al. 2016). Various forms of cannabinoids have been trialled and tested to measure the most efficacious form for oncological effects and these include a pure (P) form versus a botanical drug substance (BDS) which is an active form of the drug that has been cultivated usually available as a powder, tablet or elixir. In a study by Scott et al. using P and BDS forms for both CBD and ∆9-THC, they report efficacious activity for CBD-P in comparison to CBD-BDS and vice versa for ∆9-THC (Scott et al. 2014). As discussed earlier in their in vitrofindings, they report similar outcomes in their orthotopic murine model of glioma and in particular they observed a significant decrease in tumour volumes when both cannabinoids were administered with irradiation, p < 0.001 (Scott et al. 2014). These findings support the anticancer effects of cannabinoid treatment in glioma as a single therapy and also as an addition in combination treatment.
Cannabinoids share the common anticancer effect of apoptosis in their mode of action; however, it has also become apparent that autophagy is also involved. The process of apoptosis and autophagy interplay, where the survival function of autophagy negatively regulates apoptosis and inhibition of apoptosis blocks autophagy (Marino et al. 2014). Salazar and co-workers investigated ∆9-THC in a murine model of astrocytoma and found that autophagy is upstream of apoptosis in cannabinoid-induced cell death as shown by blocking autophagy, prevented cannabinoid-induced apoptosis (Salazar et al. 2009). ∆9-THC induced the effects of stimulation of ceramide synthesis de novo, ER stress, upregulation of p8 and TRIB3, phosphorylation of eIF2α on Ser51 via the activation of the CB1 receptor (Salazar et al. 2009). A human glioblastoma-induced murine model investigating GICs (glioma initiating cells; a subpopulation of cells responsible for the aggressiveness of GBM) was treated with ∆9-THC, CBD and TMZ in varying combinations. They reported an effective tumour reduction when CBD and ∆9-THC with TMZ were co-administered and that treatment with a high ratio of CBD was most efficacious (López-Valero et al. 2018).
Breast cancer
In vitro
McKallip et al. investigated the effects of ∆9-THC in human breast cancer cell lines MDA-MB-231, MCF-7 and mouse mammary carcinoma 4T-1. They reported a low expression of cannabinoid receptors; CB1 and CB2 in these cell lines. ∆9-THC did not affect cell viability in MCF-7 and 4T-1 cell lines but increased the size of a 4T1 primary tumour and enhanced metastasis in vivo. ∆9-THC exposure caused an increase in IL-4 and IL-10 cytokines and suppression of cell-mediated Th1 response by enhancement of Th2 cytokines due to upregulation in Th2-related genes. These findings suggest exposure to ∆9-THC may increase susceptibility to breast cancer which does not express cannabinoid receptors and is resistant to ∆9-THC-induced apoptosis (McKallip et al. 2005). In another study by Caffarel and colleagues ∆9-THC was investigated in the following human breast cancer cell lines; MCF-7, EVSA-T, MDA-MB-231, MDA-MB468, T-47D and SKBr3. They reported a reduction in human breast cancer cell proliferation by arrest of the cell cycle at the G2–M phase via down-regulation of the cyclin-dependent kinase (CDK1 or Cdc2) protein and an induction of apoptosis via the CB2 cannabinoid receptor which was highly expressed in the EVSA-T cell line. CB2 expression was also found to be correlated with tumours that had a low response to conventional therapies and that were also positive for certain prognostic markers including oestrogen, progesterone receptors and the presence of ERBB2/HER-2 oncogene. The psychotropic effects of cannabinoids are mediated via the CB1 rather than CB2, suggesting a cannabinoid therapy that would target the CB2 receptor would be beneficial (Caffarel et al. 2006). In a follow-up study investigating the ∆9-THC antiproliferative mechanism, exposure to ∆9-THC upregulated JunD expression, a proto-oncogene which belongs to the AP-1 transcription factor family, in the tumour cells. In addition, they also identified the involvement of the cyclin-dependent kinase inhibitor p27 and testin (a tumour-suppressor gene) as candidate targets of JunD. Stress protein p8, however was involved in ∆9-THC antiproliferative action in a JunD-independent manner, suggesting a multimodal mechanism of action (Caffarel et al. 2008).
In an interesting report by Blasco-Benito et al., they found ∆9-THC was able to disrupt the HER2–CB2R complex by selective binding to CB2R. Additionally, they concluded the antitumour efficacy of a botanical drug preparation to be more potent than pure ∆9-THC for both cell lines and animal studies (Blasco-Benito et al. 2019). Ligresti et al. investigated the anticancer properties of plant-based cannabinoids including CBD, CBG, CBC, CBDA and ∆9-THCA in addition to assessing the use of enriched CBD or ∆9-THC cannabis extracts over pure cannabinoids (Ligresti et al. 2006). Within the breast cancer cell lines, MDA-MB-231 and MCF-7, treated with the above cannabinoids, CBD was the most potent in its antiproliferative activity (Ligresti et al. 2006). They also report CBD mediated its apoptotic effects via the following routes: the direct or indirect activation of the receptors CB2 and TRPV1, receptor-independent elevation of intracellular Ca2+ and ROS generation (Ligresti et al. 2006).
Synthetic agonists or antagonists of cannabinoid receptors have been used to study the role of the ECS in cancer signalling and growth. Sarnataro and co-workers investigated the effects of Rimonabant, a CB1 antagonist, in the invasive human breast cancer line MDA-MB-231 and in the less-invasive lines, T47D and MCF-7 (Sarnataro et al. 2006). Treatment with Rimonabant caused antiproliferative effects characteristic of G1–S-phase cell cycle arrest accompanied by a downregulation in cyclins D and E with associated upregulation of cyclin-dependent kinase inhibitor p27KIP1. No observed apoptosis or necrosis occurred in vitro (Sarnataro et al. 2006). Additionally, within the invasive cells, these effects were found to be associated with lipid raft/caveolae as previously shown by the group (Sarnataro et al. 2005). Rimonabant caused complete displacement of the CB1 receptor from lipid rafts and the depletion of cholesterol by methyl-β-cyclodextrin (MCD) prevented these effects (Sarnataro et al. 2006). In cells overexpressing the CB1 receptor, Rimonabant inhibited MAPK signalling and decreased ERK1/2 activity (Sarnataro et al. 2006). Pre-treatment with MCD before Rimonabant administration caused a depletion in cholesterol and this reverted the inhibitory effects on ERK1/2 via Rimonabant, suggesting an interplay between the CB1 receptor and lipid raft motility in breast tumour growth (Sarnataro et al. 2006). JWH-015, an agonist of the CB2 receptor, in human MCF-7 mammary carcinoma cells reduced viability by inducing apoptosis independent of Gαi signalling or by pharmacological blockade of CB1, GPR55, TRPV1 or TRPA1 receptors and instead these effects were calcium-dependent and caused changes in MAPK/ERK signalling (Hanlon et al. 2016).
CBD has also been shown to downregulate Id-1 in the aggressive human breast cancer line MDA-MB-231 through modulation of ERK and ROS pathways leading to a decrease in Id-1 expression and also upregulated Id-2 (a transcriptional regulator) (McAllister et al. 2011). Shrivastava et al. observed a complex interplay between apoptosis and autophagy in CBD-treated invasive breast cancer cells, MDA-MB-231 (Shrivastava et al. 2011). In particular, CBD induced ER stress which led to the inhibition of AKT and mTOR signalling in vitroindicated by low levels of phosphorylated cyclin D1, mTOR and 4EBP1 (Shrivastava et al. 2011). Further analysis revealed CBD inhibited the association between beclin1 (central role in autophagy) and BCL-2 known to inhibit autophagy through cleavage of Beclin-1 and enhanced the interaction between Beclin-1 and Vps34 favouring autophagy (Shrivastava et al. 2011). Electron microscopy revealed morphological changes to MDA-MB-231 CBD-treated cells which included nuclear condensation, margination, increased vacuolization, decrease in intracellular organelles and enlarged mitochondria evident of apoptotic activity (Shrivastava et al. 2011). They hypothesized that the event changes in inducing autophagy may also cause apoptosis as the cleavage product from Beclin-1 translocates to the mitochondria and induces cytochrome C (Shrivastava et al. 2011). These observations and hypothesis suggest CBD may be able to control the complex interplay between autophagy and apoptosis in these breast cancer cells (Shrivastava et al. 2011). CBD also increased ROS levels and blockage of ROS inhibited apoptotic and autophagy pathways (Shrivastava et al. 2011). These effects were independent of cannabinoid and vanilloid receptor activation (Shrivastava et al. 2011).
Many drugs have failed in clinics for many of the aggressive cancers due to the recalcitrant TME. The TME plays a major role in contributing to the growth and invasion of cancer and in particular tumour-associated macrophages (TAMs) which are a class of immune cells contributing to the immunosuppressive TME through interchange of its two forms: M1 (anti-tumorigenic) and M2 (pro-tumorigenic) (Lin et al. 2019). Elbaz and colleagues investigated CBD in triple-negative breast cancer (TNBC) cell lines SUM159, MDA-MB-231-SCP2, MVT-1, 4T1.2 and in murine leukaemia RAW264.7. They observed CBD inhibited EGF-induced proliferation and chemotaxis in the cell lines, activated EGFR, ERK, Akt, and NF-κß pathways in addition to inhibition of matrix metallopeptidase 2 and 9 (MMP2 and MMP9) secretion (Elbaz et al. 2015). A cancer education experiment (conditioned media from CBD-treated cancer cells) showed a significant reduction in the number of migrated RAW 264.7 cells towards this medium which also contained lower levels of granulocyte–macrophage colony-stimulating factor (GM-CSF) and chemokine ligand 3 (CCL3) cytokines, crucial for macrophage recruitment and activation (Elbaz et al. 2015). They observed a reduction in tumour growth and metastasis and inhibition of the recruitment of total and M2 macrophages to the stroma of the primary tumour and secondary lung metastasis (Elbaz et al. 2015).
Cannabinoid’s effect on ER has been evident in many cancer studies, however, the exact mechanism by which this occurs remain elusive. In a recent study by de la Harpe et al., they found CBD selectively targeted MCF7 cells via oxidative stress-induced ER stress and UPR (unfolded protein response) activation, and these effects were caused by calcium influx via the TRPV1 receptor as opposed to MDA-MB-231 cells. This suggests the difference in CBD treatment was dependent on localization of TRPV1 (de la Harpe et al., 2021).
In vivo
One of the factors to consider in cannabinoid treatment is the abundance of cannabinoid receptors in the tissue of interest. In a study investigating the effects of ∆9-THC in a murine model of mammary carcinoma, it was found that the murine mammary carcinoma cell line 4T1 first did not express detectable levels of the cannabinoid receptors CB1 and CB2 and second, these cells were resistant to the cytotoxicity of ∆9-THC. They also show treatment with ∆9-THC led to an increase in tumour growth and metastasis due to an increase in production of IL-4 and IL-10 which suppressed the cell-mediated Th1 response by enhancing Th2-associated cytokines (McKallip et al. 2005). This finding was supported by the injection of anti-IL-4 and anti-IL-10 monoclonal antibodies which partially reversed the immune suppression of ∆9-THC in 4T1 cells (McKallip et al. 2005).
A study investigating the effects of the endogenous cannabinoid, Met-F-AEA (a metabolically stable anandamide analogue) in a highly invasive murine breast cancer model reported a significantly reduced amount and size of metastatic nodes and this effect was antagonized by the selective CB1 antagonist Rimonabant (Grimaldi et al. 2006). Molecular interrogation in treated cells with the endogenous cannabinoid caused a decrease in tyrosine phosphorylation of focal adhesion kinase (FAK) and steroid receptor coactivator (Src) and these effects were mitigated by Rimonabant (Grimaldi et al. 2006). They concluded CB1 receptor agonists by modulating FAK phosphorylation inhibited tumour cell invasion and metastasis and therefore CB1 receptor activation may represent a novel therapeutic target for the treatment of breast carcinoma and metastasis (Grimaldi et al. 2006). Rimonabant has also been reported to significantly reduce tumour volume in vivo in the invasive human MDA-MD-231 murine model and this effect occurred via the CB1R lipid raft/caveolae-mediated mechanism (Sarnataro et al. 2006).
In a human MDA-MB-231 breast carcinoma xenografted tumour model, both CBD and CBD enriched extract treatment induced apoptosis, inhibited the growth of tumours and metastasis in vivo (Ligresti et al. 2006). CBD has also been shown to modulate transcriptional activity downstream in breast cancer. A study by McAllister and colleagues investigated CBD treatment of a murine model of metastatic breast cancer and found CBD inhibited Id-1 gene expression in the primary tumour and lung metastasis in vivo through modulation of the ERK and ROS pathways (McAllister et al. 2011). Caffarel et al. have shown using a genetically engineered animal model of ErbB2-driven metastatic breast cancer (MMTV-neu mice), ∆9-THC and JWH-133 (selective CB2 agonist) reduce metastatic progression via AKT pathway inhibition (Caffarel et al. 2010).
Cannabinoids mechanistic actions have been reported to be CB-independent with studies reporting other channels through which they may activate their oncological effects, such as GPR55 or vanilloid channels. Hanlon and co-workers report using JWH-015, a CB2 agonist, significantly reduced tumour burden and metastasis of murine mammary carcinoma 4T1 cells in immunocompetent mice and these effects were dependent on calcium and induced changes to MAPK/ERK signalling which were independent of G-protein-coupled signalling, CB or vanilloid receptors (McAllister et al. 2011).
Other gastrointestinal (GI) cancers
In vitro
In a study investigating human colorectal cancer cells using the lines DLD-1, CaCo-2 and SW620, treatment with Rimonabant significantly reduced cell proliferation and induced death. In DLD-1 cells, treatment resulted in G2–M-phase cell cycle arrest without inducing apoptosis or necrosis (Aviello et al. 2012). Further investigation revealed an increase in mitotic catastrophe characterized by changes in the following, cyclin B1, PARP-1 (involved in DNA repair) Aurora B (involved in the attachment of the mitotic spindle in prophase), phosphorylated p38/MAPK and Chk1 (checkpoint kinase 1) in a time-dependent manner (Aviello et al. 2012). Rimonabant, can therefore mediate cancer tumour growth via mitotic catastrophe inducing cell-cycle arrest during spindle assembly and DNA-damage checkpoints (Aviello et al. 2012).
In hepatocellular carcinoma cell lines, HepG2 and Huh-7, treatment with ∆9-THC and JWH-015 (synthetic CB2 receptor agonist) reduced cell viability through activation of the CB2 receptor. Autophagy was subsequently induced by the upregulation of TRIB3, stimulation of adenosine monophosphate-activated kinase (AMPK) and Akt/mTORC1 inhibition (Vara et al. 2011).
In human colorectal cell lines, Caco-2 and HCT116, CBD treatment protected DNA from oxidative damage, reduced cell proliferation and increased endocannabinoid levels via CB1, TRPV1 and PPARγ (Romano et al. 2014). In addition, CBD treatment of colorectal carcinoma cell line DLD-1, reduced cell proliferation (Macpherson et al. 2014). An interesting study investigated the antiproliferative effects of CBD in Caco-2 cell line in various oxygen environments and found the antitumour effects of CBD to be greater in PhysO2 than AtmosO2. They conclude that CBD induced a mitochondrial production of ROS in PhysO2 cells, suggesting that the cellular redox environment can influence how CBD induced antiproliferative effects in PhysO2 to AtmosO2 cells (Nallathambi et al. 2018). This study demonstrates the important role microenvironments play in cell cultures when studying the pharmacokinetics and mechanism of drugs. Macpherson and colleagues report the increase in sensitivity to CBD-induced antiproliferative effects through changes to cell energetics, from a drop in oxygen and a loss in mitochondrial membrane integrity in cells under the atmospheric condition to the increase in ROS in mitochondria under low oxygen conditions (Nallathambi et al. 2018).
Purified cannabinoids have been mainly reported in inducing apoptosis, inhibiting proliferation and metastasis in many cancer types, however, other forms such as unheated extracts of the plants have been less studied. Nallathambi and colleagues identified unheated extract fractions (F7: THCA, F3: CBGA) from C. sativa which displayed cytotoxic effects in colorectal cancer cell lines, HCT116 and CCD-18Co and adenomatous polyps but reduced activity on normal colon cell lines (Nallathambi et al. 2018). Combination treatment analysed by the Bliss independence model, exhibited more potent cytotoxic effects which included cell cycle arrest, cell death and a reduction in genes involved in the Wnt signalling pathway (Proto et al. 2017).
In vivo
Rimonabant treatment in a mouse model of azoxymethane-induced colon carcinogenesis caused a significant reduction in aberrant crypt foci formation, which is a neoplastic precursor to colorectal cancer and additionally observed inhibitory effects with changes to mitotic and DNA-damage checkpoints in their cell lines as mentioned previously (Aviello et al. 2012). Another study investigated the synthetic cannabinoids effects on the Wnt/β-catenin pathway, a signalling pathway involved in the formation of colorectal cancer (Borelli et al. 2014). The administration of rimonabant in HCT116 xenografts caused a significant reduction in tumour growth and destabilized the nuclear localization of β-catenin in vivo by inhibiting the canonical Wnt pathway (Borelli et al. 2014). This study suggests a novel use for cannabinoids in treating colorectal cancer harbouring mutations in β-catenin.
In a murine model of hepatocellular carcinoma, treatment with JWH-015 and ∆9-THC, both cannabinoids reduced subcutaneous xenograft growth; however, this effect was not observed when autophagy was pharmacologically inhibited (Vara et al. 2011) indicating the importance of cell death in both cannabinoids reducing tumour burden in vivo. Furthermore, administration of the cannabinoids also led to a reduction in ascites (abnormal build-up of fluid in the abdomen) formation (Vara et al. 2011). In support of the mechanisms observed in the HCC cell lines, Salazar et al. investigated ∆9-THC in the astrocytoma cell line U87MG and in vivo where they report autophagy induction via the upregulation of p8 leading to apoptosis and inhibition of Akt and mTORC1 (Salazar et al. 2009).
The effect of CBD in gastrointestinal cancers has also been studied. In a study by Aviello et al., CBD treatment in an azoxymethane (AOM)-induced murine model of colon cancer, reduced aberrant crypt foci, polyps, tumour growth and led to a decrease in expression of inducible nitric oxide synthase (iNOS) and phosphorylated Akt with an upregulation in caspase-3 (Aviello et al. 2012). CBG’s anticancer effect has been observed in colon cancer models. Borelli et al. evaluated the antineoplastic effects in xenograft models of colon cancer and observed a reduction tumour growth, however due to the limitation in the model, they further tested CBG in an AOM colon murine model which mimics the tumour in situ and found CBG completely abolished the formation of aberrant crypt foci and reduced the number of tumours (Borelli et al. 2014). In addition, Romano et al. tested the effects of the BDS form of CBD, which contains a high content of CBD on colorectal cancer growth in both xenograft and AOM models. They also observed a reduction in tumour growth, preneoplastic lesions and polyps (Macpherson et al. 2014).
Prostate cancer
In vitro
∆9-THC induced apoptosis in a PC-3 prostate cancer cell line in a dose-dependent manner (Sreevalsan et al. 2011). CBD’s pro-apoptotic nature has been shown to be phosphate-dependent in prostate and colon cancer cells (De Petrocellis et al. 2013). In LNCaP (prostate) and SW480 (colon) cancer cell lines, the growth and mRNA expression of several phosphatases inhibited cannabinoid-induced PARP cleavage (De Petrocellis et al. 2013). De Petrocellis et al. investigated CBD’s effect in prostate carcinoma cell lines; LNCaP, 22RV1 (positive for androgen receptor), DU-145 and PC-3 (negative for androgen receptor). CBD treatment significantly decreased cell viability and potentiated the effects of bicalutamide and docetaxel (standard drugs for treatment of prostate carcinoma) against LNCaP and DU-145 xenograft tumours and when given alone reduced LNCaP xenograft size. CBD administered between 1 and 10 µM induced apoptosis and markers of intrinsic apoptotic pathways (PUMA, CHOP expression and intracellular Ca2+). In LNCaP cells, the pro-apoptotic effect of CBD was only partly due to TRPM8 antagonism and was accompanied by down-regulation of AR, p53 activation and elevation of ROS. LNCaP cells differentiated to androgen-insensitive neuroendocrine-like cells were more sensitive to CBD-induced apoptosis (De Petrocellis et al. 2013).
Gynaecological cancers
In vitro
The effects of ∆9-THC were also investigated in aggressive endometrial cancer. Zhang et al. report in HEC-1B and An3ca aggressive endometrial cancer cell lines a high level of cannabinoid receptor expression and treatment with ∆9-THC inhibited cell viability and motility by inhibiting epithelial-mesenchymal transition (EMT) in addition to down-regulation of the MMP9 gene in inhibiting metastasis. These findings suggest regulation and targeting of the MMP9-related pathways via ∆9-THC treatment may inhibit metastasis in this aggressive cancer type (Zhang et al. 2018). A recent study investigated the oncological effects of CBD as a monotherapy and in combination with chemotherapy drugs in ovarian cancer, administered as Poly lactic-co-glycolytic acid (PGLA)-microparticles (Fraguas-Sánchez et al. 2020). Their results show the combination of paclitaxel (PTX) with CBD to be effective in vitro and in ovo (Fraguas-Sánchez et al. 2020). CBD administered as microparticles was more efficacious than in single solution and in ovo, PTX resulted in a 1.5-fold tumour growth inhibition whereas in combination with CBD this increased to a twofold decrease, suggesting a promising therapy to explore in treating ovarian cancer as it provides the advantageous effect of using a lower dose of the antineoplastic drug whilst maintaining the same efficacy (Fraguas-Sánchez et al. 2020).
Clinical studies
The anticancer effects of cannabinoids have so far been limited to preclinical studies and translation to the clinic has remained stagnant. The first report of the use of cannabinoids on cancer patients was a pilot study that investigated ∆9-THC on nine terminal patients with recurrent glioblastoma where standard therapy remained unhopeful as a curative (Guzmán et al. 2006). These patients underwent intracranial administration of ∆9-THC, as this route was deemed the safest and patients did not exhibit any of the associated psychoactive effects (Guzmán et al. 2006). In-depth analysis of two patients’ tumours revealed molecular effects associated with cannabinoids antitumour action, which included decreased cell proliferation, stimulation of apoptosis and autophagy (Guzmán et al. 2006). Although positive effects were observed, the small case number hinders any statistically significant conclusions to be drawn from this study.
A recently published completed clinical study investigated the safety and preliminary efficacy of nabiximols oromucosal cannabinoid spray and dose intense (DIT) TMZ in patients with first recurrence glioblastoma (Twelves et al. 2021). The study included an open label arm where patients received nabiximols (n = 6) and a randomised, double-blind, and placebo-controlled arm (n = 12 and n = 9). Up to 12 sprays/days with DIT for 12 months were administered and the safety, efficacy and pharmacokinetics of TMZ were observed. Study reports a 33% of nabiximols and placebo-treated patients were progression free for 6 months and survival at 1 year for nabiximols was 83% and 44% for placebo patients and no effects of nabiximols on TMZ were reported. Although nabiximols spray was tolerable in GBM patients, a major limitation to the study was the small size of enrolled patients, specifically 21 across 9 sites and there was no predetermined power calculation to the study to define the minimum number of patients for statistical power (Twelves et al. 2021). Nevertheless, the observations warrant the need for further clinical trials to help establish safe and efficacious routes of administration, patient sub-stratification and to explore its possible synergistic effects with other antitumour agents as shown in pre-clinical data. Table 3 summarises clinical trials investigating cannabinoids including synthetic versions, CBD and ∆9-THC in cancer treatment.
Table 3.
Overview of clinical trials for investigation of cannabinoids in cancer. Key search terms included: “Cancer and Cannabinoids, Cannabis, Cannabidiol, Tetrahydrocannabinol” (www.clinicaltrials.gov)
| Trial name | Conditions | Phase | n | Study type | Drug | Location | Status | NCT no. |
|---|---|---|---|---|---|---|---|---|
| A Pilot Study of Dronabinol for Adult Patients With Primary Gliomas | Brain Neoplasms|Nausea|Vomiting | I | 33 | Interventional | Dronabinol | North Carlina, USA | Completed | NCT00314808 |
| A Phase 1 Study of Dexanabinol in Patients With Advanced Solid Tumours | Solid Tumour | I | 40 | Interventional | Dexanabinol|Other: Cremophor | Leeds/Newcastle/Glasgow | Completed | NCT01489826 |
| A Safety Study of Sativex in Combination With Dose-intense Temozolomide in Patients With Recurrent Glioblastoma | Cancer | I/II | 6 | Interventional | Sativex | Leeds/Bristol/London | Completed | NCT01812603 |
| A Safety Study of Sativex Compared With Placebo (Both With Dose-intense Temozolomide) in Recurrent Glioblastoma Patients | Cancer | I/II | 21 | Interventional | Sativex|Placebo | Germany | Completed | NCT01812616 |
| A Pharmacokinetic Study of Single Doses of Sativex in Treatment-induced Mucositis | Head and Neck Squamous Cell Carcinoma | I | 10 | Interventional | Sativex | London | Terminated | NCT01975688 |
| Assessment of Single Doses of Oral Dexanabinol in Healthy Subjects | Safety|Tolerability|Pharmacokinetics|Cancer | I | 40 | Interventional | Dexanabinol, Placebo | Nottingham | Completed | NCT02054754 |
| A Study: Pure CBD as Single-agent for Solid Tumor | Solid Tumor | II | 60 | Interventional | Cannabidiol | Israel | Unknown status | NCT02255292 |
| A Study of Dexanabinol in Combination With Chemotherapy in Patients With Advanced Tumours | Hepatocellular Carcinoma|Pancreatic Cancer | I | 112 | Interventional | Dexanabinol| Sorafenib| Nab-paclitaxel| Gemcitabine | Switzerland/Germany/Spain | Unknown status | NCT02423239 |
| A Study to Assess the Pharmacokinetic (PK) Properties of Sativex® in Patients With Advanced Cancer | Advanced Cancer | I | 0 | Interventional | Sativex | United Kingdom | Withdrawn | NCT02432612 |
| Study on Cannabinoid Receptor Expression in Gastrointestinal Diseases | Ulcerative Colitis|Crohn's Disease|Colon Cancer | – | 31 | Observational | N/A | Austria | Completed | NCT02735941 |
| The Effect of Cannabis in Pancreatic Cancer | Neoplasms Pancreatic|Cachexia; Cancer|Cannabis|Appetite Loss|Palliative Medicine|Morbidity|Mortality | II | 32 | Interventional | THC and CBD Mixture | Denmark | Unknown status | NCT03245658 |
| Tolerability of Cannabis in Patients Receiving Concurrent Chemoradiation for Glioblastoma | Glioblastoma | I | 1 | Interventional | Cannabis| Temozolomide| Radiation Therapy | New York, USA | Terminated | NCT03246113 |
| Medical Cannabis During Chemoradiation for Head and Neck Cancer | Head and Neck Cancer | – | 30 | Observational | Cannabis | New York, USA | Recruiting | NCT03431363 |
| TN-TC11G (THC + CBD) Combination With Temozolomide and Radiotherapy in Patients With Newly-diagnosed Glioblastoma | Glioblastoma | I/II | 30 | Interventional | TN-TC11G| Temozolomide Oral Product| Radiotherapy | Spain | Not yet recruiting | NCT03529448 |
| A Study of the Efficacy of Cannabidiol in Patients With Multiple Myeloma, Glioblastoma Multiforme, and GI Malignancies | Cancer of Pancreas|Cancer of Liver|Cancer of Rectum|Cancer of Colon|Cancer, Gall Bladder|Myeloma Multiple|Glioblastoma Multiforme | I/ II | 160 | Interventional | Cannabidiol| Bortezomib| Leucovorin| 5-FU| Oxaliplatin| Bevacizumab| Irinotecan| Gemcitabine| Temozolomide | Orlando/Florida, USA | Not yet recruiting | NCT03607643 |
| Cannabis Use in Cancer Patients | Solid Tumor, Adult | – | 30 | Observational | Cannabis | Colorado, USA | Recruiting | NCT03617692 |
| Pilot, Syndros, Decreasing Use of Opioids in Breast Cancer Subjects With Bone Mets | Bone Metastases|Breast Cancer|Pain | Early I | 20 | Interventional | Syndros | Arizona, USA | Recruiting | NCT03661892 |
| Pharmacokinetic (PK) and Pharmacodynamics (PD) Study of Ilera Specific Products | Cancer and other ailments | – | 10 | Observational | Registry|Other: PK microsampling of blood | Philadelphia/ Pennsylvania, USA | Terminated | NCT03886753 |
| Efficacy and Safety of Dronabinol in the Improvement of Chemotherapy-induced and Tumor-related Symptoms in Advanced Pancreatic Cancer | Pancreatic Cancer Non-resectable|Chemotherapy-induced Nausea and Vomiting|Pancreatic Cancer Metastatic | III | 140 | Interventional | Dronabinol in Oral Dosage Form| Placebo in Oral Dosage Form | Austria | Recruiting | NCT03984214 |
| Dibenzyl Trisulphide (GUINEAHEN WEED) for Stage IV Cancer | Stage IV Prostate Cancer|Stage IV Colon Cancer|Stage IV Breast Cancer|Stage IV Cancer of the Cervix | Early I | 104 | Interventional | Dibenzyl trisulphide capsules| Placebo | Jamaica | Unknown status | NCT04113096 |
| Effect of Hemp-CBD on Patients With CIPN | Chemotherapy-induced Peripheral Neuropathy|Colorectal Cancer Stage II|Colorectal Cancer Stage III|Breast Cancer|Ovarian Cancer|Uterine Cancer | II | 100 | Interventional | Hemp-based CBD| Placebo oral tablet | Pennsylvania, USA | Recruiting | NCT04398446 |
| Epidiolex (CBD) in Patients With Biochemically Recurrent Prostate Cancer | Prostate Cancer Recurrent|Prostate Cancer|Prostate Adenocarcinoma | I | 18 | Interventional | Epidiolex Oral Liquid | Kentucky, USA | Recruiting | NCT04428203 |
Conclusion
Plant-based, endogenous and synthetic cannabinoid compounds have shown merits in not only alleviating the unwanted side effects of antineoplastic drug regiments, but have also shown promising evidence in decreasing tumour burden, and one in vivo study so far concludes increasing survival rates in mice. The antitumour effects of cannabinoids trend in modulating processes which include apoptosis and autophagy through first stimulating de novo synthesis of ceramide which induces activation of ER stress-related signalling proteins further leading to the inhibition of the AKT/mTORC1 axis promoting cell cycle arrest and additional mechanisms, such as cell death and aging. Other pathways involved mechanistically are activation of MAPK/ERK signalling through calcium induction. Strategies that would optimize the anticancer effects of cannabinoids through interference of these signalling cross-talks may prove useful for therapeutic intervention. Nevertheless, we found that these effects were reached differently downstream depending on the type of cancer, the dosage of the compound and which receptor/ligands were activated. We also found the co-administration of cannabinoids with chemotherapy drugs enhanced the potency of these effects. These synergistic effects should be targeted for translation to clinical application, especially in cancers which are refractory to chemotherapy. Various extracted forms of cannabinoids from C. sativa have shown varying cytotoxic effects which should be explored in more detail in future studies as majority of the evidence originates from studies investigating mainly ∆9-THC and CBD’s actions. Whilst the emerging evidence of phytocannabinoid anticancer effects are promising, there remains a paucity of clinical evaluation which must be overcome.
Funding
There were no sources of funding associated with this manuscript.
Declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Code of availability
Not applicable.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Afrin F, Chi M, Eamens AL, Duchatel RJ, Douglas AM, Schneider J, Gedye C, Woldu AS, Dun MD. Can hemp help? Low-THC cannabis and non-THC cannabinoids for the treatment of cancer. Cancers (basel) 2020;12(4):1033. doi: 10.3390/cancers12041033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akimov MG, Gamisonia AM, Dudina PV, Gretskaya NM, Gaydaryova AA, Kuznetsov AS, Zinchenko GN, Bezuglov VV. GPR55 receptor activation by the N-acyl dopamine family lipids induces apoptosis in cancer cells via the nitric oxide synthase (nNOS) over-stimulation. Int J Mol Sci. 2021;22(2):622. doi: 10.3390/ijms22020622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andradas C, Blasco-Benito S, Castillo-Lluva S, Dillenburg-Pilla P, Diez-Alarcia R, Juanes-García A, García-Taboada E, Hernando-Llorente R, Soriano J, Hamann S, Wenners A, Alkatout I, Klapper W, Rocken C, Bauer M, Arnold N, Quintanilla M, Megías D, Vicente-Manzanares M, Urigüen L, Gutkind JS, Guzmán M, Pérez-Gómez E, Sánchez C. Activation of the orphan receptor GPR55 by lysophosphatidylinositol promotes metastasis in triple-negative breast cancer. Oncotarget. 2016;7(30):47565–47575. doi: 10.18632/oncotarget.10206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andre CM, Hausman J-F, Guerriero G. Cannabis sativa: the plant of the thousand and one molecules. Front Plant Sci. 2016;7:19. doi: 10.3389/fpls.2016.00019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong JL, Hill DS, McKee CS, Hernandez-Tiedra S, Lorente M, Lopez-Valero I, Eleni Anagnostou M, Babatunde F, Corazzari M, Redfern CPF, Velasco G, Lovat PE. Exploiting cannabinoid-induced cytotoxic autophagy to drive melanoma cell death. J Invest Dermatol. 2015;135(6):1629–1637. doi: 10.1038/jid.2015.45. [DOI] [PubMed] [Google Scholar]
- Aviello G, Romano B, Borrelli F, Capasso R, Gallo L, Piscitelli F, Di Marzo V, Izzo AA. Chemopreventive effect of the non-psychotropic phytocannabinoid cannabidiol on experimental colon cancer. J Mol Med (berl) 2012;90(8):925–934. doi: 10.1007/s00109-011-0856-x. [DOI] [PubMed] [Google Scholar]
- Barbado MV, Medrano M, Caballero-Velázquez T, Álvarez-Laderas I, Sánchez-Abarca LI, García-Guerrero E, Martín-Sánchez J, Rosado IV, Piruat JI, Gonzalez-Naranjo P, Campillo NE, Páez JA, Pérez-Simón JA. Cannabinoid derivatives exert a potent anti-myeloma activity both in vitro and in vivo. Int J Cancer. 2017;140(3):674–685. doi: 10.1002/ijc.30483. [DOI] [PubMed] [Google Scholar]
- Blasco-Benito S, Moreno E, Seijo-Vila M, Tundidor I, Andradas C, Caffarel MM, Caro-Villalobos M, Urigüen L, Diez-Alarcia R, Moreno-Bueno G, Hernández L, Manso L, Homar-Ruano P, McCormick PJ, Bibic L, Bernadó-Morales C, Arribas J, Canals M, Casadó V, Canela EI, Guzmán M, Pérez-Gómez E, Sánchez C. Therapeutic targeting of HER2-CB2R heteromers in HER2-positive breast cancer. Proc Natl Acad Sci USA. 2019;116(9):3863–3872. doi: 10.1073/pnas.1815034116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blázquez C, Carracedo A, Barrado L, Real PJ, Fernández-Luna JL, Velasco G, Malumbres M, Guzmán M. Cannabinoid receptors as novel targets for the treatment of melanoma. FASEB J. 2006;20(14):2633–2635. doi: 10.1096/fj.06-6638fje. [DOI] [PubMed] [Google Scholar]
- Borrelli F, Pagano E, Romano B, Panzera S, Maiello F, Coppola D, De Petrocellis L, Buono L, Orlando P, Izzo AA. Colon carcinogenesis is inhibited by the TRPM8 antagonist cannabigerol, a Cannabis-derived non-psychotropic cannabinoid. Carcinogenesis. 2014;35(12):2787–2797. doi: 10.1093/carcin/bgu205. [DOI] [PubMed] [Google Scholar]
- Brandi J, Dando I, Palmieri M, Donadelli M, Cecconi D. Comparative proteomic and phosphoproteomic profiling of pancreatic adenocarcinoma cells treated with CB1 or CB2 agonists. Electrophoresis. 2013;34(9–10):1359–1368. doi: 10.1002/elps.201200402. [DOI] [PubMed] [Google Scholar]
- Brown KJ, Laun AS, Song Z-H. Cannabidiol, a novel inverse agonist for GPR12. Biochem Biophys Res Commun. 2017;493(1):451–454. doi: 10.1016/j.bbrc.2017.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caffarel MM, Sarrió D, Palacios J, Guzmán M, Sánchez C. Delta9-tetrahydrocannabinol inhibits cell cycle progression in human breast cancer cells through Cdc2 regulation. Cancer Res. 2006;66(13):6615–6621. doi: 10.1158/0008-5472.CAN-05-4566. [DOI] [PubMed] [Google Scholar]
- Caffarel M, Andradas C, Mira E, Pérez-Gómez E, Cerutti C, Moreno-Bueno G, Flores JM, García-Real I, Palacios J, Mañes S, Guzmán M, Sánchez C (2010) Cannabinoids reduce ErbB2-driven breast cancer progression through Akt inhibition. Mol Cancer 9:196. 10.1186/1476-4598-9-196 [DOI] [PMC free article] [PubMed]
- Caffarel M, Moreno-Bueno G, Cerutti C, et al. JunD is involved in the antiproliferative effect of Δ9-tetrahydrocannabinol on human breast cancer cells. Oncogene. 2008;27:5033–5044. doi: 10.1038/onc.2008.145. [DOI] [PubMed] [Google Scholar]
- Calvaruso G, Pellerito O, Notaro A, Giuliano M. Cannabinoid-associated cell death mechanisms in tumor models (review) Int J Oncol. 2012;41(2):407–413. doi: 10.3892/ijo.2012.1476. [DOI] [PubMed] [Google Scholar]
- Carracedo A, Lorente M, Egia A, Blázquez C, García S, Giroux V, Malicet C, Villuendas R, Gironella M, González-Feria L, Piris MA, Iovanna JL, Guzmán M, Velasco G. The stress-regulated protein p8 mediates cannabinoid-induced apoptosis of tumor cells. Cancer Cell. 2006;9(4):301–312. doi: 10.1016/j.ccr.2006.03.005. [DOI] [PubMed] [Google Scholar]
- Chakravarti B, Ravi J, Ganju RK. Cannabinoids as therapeutic agents in cancer: current status and future implications. Oncotarget. 2014;5(15):5852–5872. doi: 10.18632/oncotarget.v5i15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Console-Bram L, Brailoiu E, Brailoiu GC, Sharir H, Abood ME. Activation of GPR18 by cannabinoid compounds: a tale of biased agonism. Br J Pharmacol. 2014;171(16):3908–3917. doi: 10.1111/bph.12746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dando I, Donadelli M, Costanzo C, Dalla Pozza E, D'Alessandro A, Zolla L, Palmieri M. Cannabinoids inhibit energetic metabolism and induce AMPK-dependent autophagy in pancreatic cancer cells. Cell Death Dis. 2013;4(6):e664. doi: 10.1038/cddis.2013.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dariš B, Tancer Verboten M, Knez Ž, Ferk P. Cannabinoids in cancer treatment: therapeutic potential and legislation. Bosn J Basic Med Sci. 2019;19(1):14–23. doi: 10.17305/bjbms.2018.3532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Harpe A, Beukes N, Frost CL. CBD activation of TRPV1 induces oxidative signaling and subsequent ER stress in breast cancer cell lines. Biotechnol Appl Biochem. 2021 doi: 10.1002/bab.2119. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L, Ligresti A, Schiano Moriello A, Iappelli M, Verde R, Stott CG, Cristino L, Orlando P, Di Marzo V. Non-THC cannabinoids inhibit prostate carcinoma growth in vitro and in vivo: pro-apoptotic effects and underlying mechanisms. Br J Pharmacol. 2013;168(1):79–102. doi: 10.1111/j.1476-5381.2012.02027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donadelli M, Dando I, Zaniboni T, Costanzo C, Dalla Pozza E, Scupoli MT, Scarpa A, Zappavigna S, Marra M, Abbruzzese A, Bifulco M, Caraglia M, Palmieri M. Gemcitabine/cannabinoid combination triggers autophagy in pancreatic cancer cells through a ROS-mediated mechanism. Cell Death Dis. 2011;2(4):e152. doi: 10.1038/cddis.2011.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elbaz M, et al. Modulation of the tumor microenvironment and inhibition of EGF/EGFR pathway: novel anti-tumor mechanisms of Cannabidiol in breast cancer. Mol Oncol. 2015;9(4):906–919. doi: 10.1016/j.molonc.2014.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellert-Miklaszewska A, Ciechomska IA, Kaminska B. Synthetic cannabinoids induce autophagy and mitochondrial apoptotic pathways in human glioblastoma cells independently of deficiency in TP53 or PTEN tumor suppressors. Cancers. 2021;13(3):419. doi: 10.3390/cancers13030419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellert-Miklaszewska A, Ciechomska IA, Kaminska B. Synthetic cannabinoids induce autophagy and mitochondrial apoptotic pathways in human glioblastoma cells independently of deficiency in TP53 or PTEN tumor suppressors. Cancers (basel) 2021;13(3):419. doi: 10.3390/cancers13030419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ElSohly MA, et al. Phytochemistry of Cannabis sativa L. Prog Chem Org Nat Prod. 2017 doi: 10.1007/978-3-319-45541-9_1. [DOI] [PubMed] [Google Scholar]
- Falasca M, Ferro R. Role of the lysophosphatidylinositol/GPR55 axis in cancer. Adv Biol Regul. 2016 doi: 10.1016/j.jbior.2015.10.003. [DOI] [PubMed] [Google Scholar]
- Ferro R, et al. GPR55 signalling promotes proliferation of pancreatic cancer cells and tumour growth in mice, and its inhibition increases effects of gemcitabine. Oncogene. 2018;37(49):6368–6382. doi: 10.1038/s41388-018-0390-1. [DOI] [PubMed] [Google Scholar]
- Fiore D, Ramesh P, Proto MC, Piscopo C, Franceschelli S, Anzelmo S, Medema JP, Bifulco M, Gazzerro P. Rimonabant kills colon cancer stem cells without inducing toxicity in normal colon organoids. Front Pharmacol. 2018;8:949. doi: 10.3389/fphar.2017.00949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher T, Golan H, Schiby G, PriChen S, Smoum R, Moshe I, Peshes-Yaloz N, Castiel A, Waldman D, Gallily R, Mechoulam R, Toren A (2016) In vitro and in vivo efficacy of non-psychoactive cannabidiol in neuroblastoma. Curr Oncol 23(2):S15-22. 10.3747/co.23.2893 [DOI] [PMC free article] [PubMed]
- Fogli S, Nieri P, Chicca A, Adinolfi B, Mariotti V, Iacopetti P, Breschi MC, Pellegrini S. Cannabinoid derivatives induce cell death in pancreatic MIA PaCa-2 cells via a receptor-independent mechanism. FEBS Lett. 2006;580(7):1733–1739. doi: 10.1016/j.febslet.2006.02.024. [DOI] [PubMed] [Google Scholar]
- Fraguas-Sánchez AI, Fernández-Carballido A, Simancas-Herbada R, Martin-Sabroso C, Torres-Suárez AI. CBD loaded microparticles as a potential formulation to improve paclitaxel and doxorubicin-based chemotherapy in breast cancer. Int J Pharm. 2020;25(574):118916. doi: 10.1016/j.ijpharm.2019.118916. [DOI] [PubMed] [Google Scholar]
- Franco R, Rivas-Santisteban R, Reyes-Resina I, Casanovas M, Pérez-Olives C, Ferreiro-Vera C, Navarro G, Sánchez de Medina V, Nadal X. Pharmacological potential of varinic-, minor-, and acidic phytocannabinoids. Pharmacol Res. 2020;158:104801. doi: 10.1016/j.phrs.2020.104801. [DOI] [PubMed] [Google Scholar]
- Gazzerro P, Malfitano AM, Proto MC, Santoro A, Pisanti S, Caruso MG, Notarnicola M, Messa C, Laezza C, Misso G, Caraglia M, Bifulco M. Synergistic inhibition of human colon cancer cell growth by the cannabinoid CB1 receptor antagonist rimonabant and oxaliplatin. Oncol Rep. 2010;23(1):171–175. [PubMed] [Google Scholar]
- Gómez del Pulgar T, Velasco G, Sánchez C, Haro A, Guzmán M (2002) De novo-synthesized ceramide is involved in cannabinoid-induced apoptosis. Biochem J 363(Pt 1):183–8. 10.1042/0264-6021:3630183 [DOI] [PMC free article] [PubMed]
- Gonçalves ECD, Baldasso GM, Bicca MA, Paes RS, Capasso R, Dutra RC. Terpenoids, cannabimimetic ligands, beyond the Cannabis plant. Molecules. 2020;25(7):1567. doi: 10.3390/molecules25071567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grimaldi C, Pisanti S, Laezza C, Malfitano AM, Santoro A, Vitale M, Caruso MG, Notarnicola M, Iacuzzo I, Portella G, Di Marzo V, Bifulco M. Anandamide inhibits adhesion and migration of breast cancer cells. Exp Cell Res. 2006;312(4):363–373. doi: 10.1016/j.yexcr.2005.10.024. [DOI] [PubMed] [Google Scholar]
- Guerrero-Alba R, et al. Some prospective alternatives for treating pain: the endocannabinoid system and its putative receptors GPR18 and GPR55. Front Pharmacol. 2019;9:1496. doi: 10.3389/fphar.2018.01496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guindon J, Hohmann A. The endocannabinoid system and pain. CNS Neurol Disord Drug Targets. 2012 doi: 10.2174/187152709789824660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gustafsson K, Sander B, Bielawski J, Hannun YA, Flygare J. Potentiation of cannabinoid-induced cytotoxicity in mantle cell lymphoma through modulation of ceramide metabolism. Mol Cancer Res MCR. 2009;7(7):1086–1098. doi: 10.1158/1541-7786.MCR-08-0361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guzmán M, Duarte MJ, Blázquez C, Ravina J, Rosa MC, Galve-Roperh I, Sánchez C, Velasco G, González-Feria L. A pilot clinical study of Delta9-tetrahydrocannabinol in patients with recurrent glioblastoma multiforme. Br J Cancer. 2006;95(2):197–203. doi: 10.1038/sj.bjc.6603236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gyombolai P, et al. Regulation of endocannabinoid release by G proteins: a paracrine mechanism of G protein-coupled receptor action. Mol Cell Endocrinol. 2012 doi: 10.1016/j.mce.2011.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000 doi: 10.1016/S0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011 doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- Hanlon KE, Lozano-Ondoua AN, Umaretiya PJ, Symons-Liguori AM, Chandramouli A, Moy JK, Kwass WK, Mantyh PW, Nelson MA, Vanderah TW. Modulation of breast cancer cell viability by a cannabinoid receptor 2 agonist, JWH-015, is calcium dependent. Breast Cancer (dove Med Press) 2016;15(8):59–71. doi: 10.2147/BCTT.S100393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hart S, Fischer OM, Axel U. Cannabinoids induce cancer cell proliferation via tumor necrosis factor α-converting enzyme (TACE/ADAM17)-mediated transactivation of the epidermal growth factor receptor. Cancer Res. 2004;64(6):1943–1950. doi: 10.1158/0008-5472.CAN-03-3720. [DOI] [PubMed] [Google Scholar]
- Hernández-Tiedra S, Fabriàs G, Dávila D, Salanueva ÍJ, Casas J, Montes LR, Antón Z, García-Taboada E, Salazar-Roa M, Lorente M, Nylandsted J, Armstrong J, López-Valero I, McKee CS, Serrano-Puebla A, García-López R, González-Martínez J, Abad JL, Hanada K, Boya P, Goñi F, Guzmán M, Lovat P, Jäättelä M, Alonso A, Velasco G. Dihydroceramide accumulation mediates cytotoxic autophagy of cancer cells via autolysosome destabilization. Autophagy. 2016;12(11):2213–2229. doi: 10.1080/15548627.2016.1213927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirao-Suzuki M, Takeda S, Koga T, Takiguchi M, Toda A. Cannabidiolic acid dampens the expression of cyclooxygenase-2 in MDA-MB-231 breast cancer cells: possible implication of the peroxisome proliferator-activated receptor β/δ abrogation. J Toxicol Sci. 2020;45(4):227–236. doi: 10.2131/jts.45.227. [DOI] [PubMed] [Google Scholar]
- Howlett AC. Cannabinoid receptor signaling. Handb Exp Pharmacol. 2005;168:53–79. doi: 10.1007/3-540-26573-2_2. [DOI] [PubMed] [Google Scholar]
- Huang T, Xu T, Wang Y, Zhou Y, Yu D, Wang Z, He L, Chen Z, Zhang Y, Davidson D, Dai Y, Hang C, Liu X, Yan C. Cannabidiol inhibits human glioma by induction of lethal mitophagy through activating TRPV4. Autophagy. 2021;25:1–15. doi: 10.1080/15548627.2021.1885203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibeas Bih C, Chen T, Nunn AV, Bazelot M, Dallas M, Whalley BJ. Molecular targets of cannabidiol in neurological disorders. Neurotherapeutics. 2015;12(4):699–730. doi: 10.1007/s13311-015-0377-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Javid FA, Phillips RM, Afshinjavid S, Verde R, Ligresti A. Cannabinoid pharmacology in cancer research: a new hope for cancer patients? Eur J Pharmacol. 2016;775:1–14. doi: 10.1016/j.ejphar.2016.02.010. [DOI] [PubMed] [Google Scholar]
- Jung CK, Kang WK, Park JM, Ahn HJ, Kim SW, Taek OS, Choi KY. Expression of the cannabinoid type I receptor and prognosis following surgery in colorectal cancer. Oncol Lett. 2013;5(3):870–876. doi: 10.3892/ol.2012.1081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalant H. Medicinal use of cannabis: history and current status. Pain Res Manage. 2001;6(2):80–91. doi: 10.1155/2001/469629. [DOI] [PubMed] [Google Scholar]
- Kalenderoglou N, Macpherson T, Wright KL. Cannabidiol reduces leukemic cell size—but is it important? Front Pharmacol. 2017;8:144. doi: 10.3389/fphar.2017.00144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaur R, Ambwani RS, Singh S. Endocannabinoid system: a multi-facet therapeutic target. Curr Clin Pharmacol. 2016 doi: 10.2174/1574884711666160418105339. [DOI] [PubMed] [Google Scholar]
- Khan MI, Sobocińska AA, Brodaczewska KK, Zielniok K, Gajewska M, Kieda C, Czarnecka AM, Szczylik C. Involvement of the CB2 cannabinoid receptor in cell growth inhibition and G0/G1 cell cycle arrest via the cannabinoid agonist WIN 55,212–2 in renal cell carcinoma. BMC Cancer. 2018;18(1):583. doi: 10.1186/s12885-018-4496-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleckner AS, et al. Opportunities for cannabis in supportive care in cancer. Ther Adv Med Oncol. 2019 doi: 10.1177/1758835919866362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolbe MR, Hohmann T, Hohmann U, Ghadban C, Mackie K, Zöller C, Prell J, Illert J, Strauss C, Dehghani F (2021) THC Reduces Ki67-Immunoreactive Cells Derived from Human Primary Glioblastoma in a GPR55-Dependent Manner. Cancers (basel) 13(5):1064. 10.3390/cancers13051064 [DOI] [PMC free article] [PubMed]
- Ladin DA, Soliman E, Griffin L, Van Dross R. Preclinical and clinical assessment of cannabinoids as anti-cancer agents. Front Pharmacol. 2016;7(7):361. doi: 10.3389/fphar.2016.00361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JLC, Bertoglio LJ, Guimarães FS, Stevenson CW. Cannabidiol regulation of emotion and emotional memory processing: relevance for treating anxiety-related and substance abuse disorders. Br J Pharmacol. 2017;174(19):3242–3256. doi: 10.1111/bph.13724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levy M, Futerman AH. Mammalian ceramide synthases. IUBMB Life. 2010;62(5):347–356. doi: 10.1002/iub.319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ligresti A, Moriello AS, Starowicz K, Matias I, Pisanti S, De Petrocellis L, Laezza C, Portella G, Bifulco M, Di Marzo V. Antitumor activity of plant cannabinoids with emphasis on the effect of cannabidiol on human breast carcinoma. J Pharmacol Exp Ther. 2006;318(3):1375–1387. doi: 10.1124/jpet.106.105247. [DOI] [PubMed] [Google Scholar]
- Lin Y, Xu J, Lan H. Tumor-associated macrophages in tumor metastasis: biological roles and clinical therapeutic applications. J Hematol Oncol. 2019;12:76. doi: 10.1186/s13045-019-0760-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- López-Valero I, Saiz-Ladera C, Torres S, Hernández-Tiedra S, García-Taboada E, Rodríguez-Fornés F, Barba M, Dávila D, Salvador-Tormo N, Guzmán M, Sepúlveda JM, Sánchez-Gómez P, Lorente M, Velasco G. Targeting glioma initiating cells with A combined therapy of cannabinoids and temozolomide. Biochem Pharmacol. 2018;157:266–274. doi: 10.1016/j.bcp.2018.09.007. [DOI] [PubMed] [Google Scholar]
- Lorente M, Torres S, Salazar M, Carracedo A, Hernández-Tiedra S, Rodríguez-Fornés F, García-Taboada E, Meléndez B, Mollejo M, Campos-Martín Y, Lakatosh SA, Barcia J, Guzmán M, Velasco G. Stimulation of the midkine/ALK axis renders glioma cells resistant to cannabinoid antitumoral action. Cell Death Differ. 2011;18(6):959–973. doi: 10.1038/cdd.2010.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu H-C, Mackie K. An introduction to the endogenous cannabinoid system. Biol Psychiat. 2016;79(7):516–525. doi: 10.1016/j.biopsych.2015.07.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macpherson T, Armstrong JA, Criddle DN, Wright KL. Physiological intestinal oxygen modulates the Caco-2 cell model and increases sensitivity to the phytocannabinoid cannabidiol. Vitro Cell Dev Biol Anim. 2014;50(5):417–426. doi: 10.1007/s11626-013-9719-9. [DOI] [PubMed] [Google Scholar]
- Marcu JP, Christian RT, Lau D, Zielinski AJ, Horowitz MP, Lee J, Pakdel A, Allison J, Limbad C, Moore DH, Yount GL, Desprez PY, McAllister SD. Cannabidiol enhances the inhibitory effects of delta9-tetrahydrocannabinol on human glioblastoma cell proliferation and survival. Mol Cancer Ther. 2010;9(1):180–189. doi: 10.1158/1535-7163.MCT-09-0407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mariño G, Niso-Santano M, Baehrecke EH, Kroemer G. Self-consumption: the interplay of autophagy and apoptosis. Nat Rev Mol Cell Biol. 2014;15(2):81–94. doi: 10.1038/nrm3735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall AD, Lagutina I, Grosveld GC. PAX3-FOXO1 induces cannabinoid receptor 1 to enhance cell invasion and metastasis. Cancer Res. 2011;71(24):7471–7481. doi: 10.1158/0008-5472.CAN-11-0924. [DOI] [PubMed] [Google Scholar]
- Massi P, Vaccani A, Ceruti S, Colombo A, Abbracchio MP, Parolaro D. Antitumor effects of cannabidiol, a nonpsychoactive cannabinoid, on human glioma cell lines. J Pharmacol Exp Ther. 2004;308(3):838–845. doi: 10.1124/jpet.103.061002. [DOI] [PubMed] [Google Scholar]
- Matas D, Juknat A, Pietr M, Klin Y, Vogel Z. Anandamide protects from low serum-induced apoptosis via its degradation to ethanolamine. J Biol Chem. 2007;282(11):7885–7892. doi: 10.1074/jbc.M608646200. [DOI] [PubMed] [Google Scholar]
- McAllister SD, Murase R, Christian RT, Lau D, Zielinski AJ, Allison J, Almanza C, Pakdel A, Lee J, Limbad C, Liu Y, Debs RJ, Moore DH, Desprez PY. Pathways mediating the effects of cannabidiol on the reduction of breast cancer cell proliferation, invasion, and metastasis. Breast Cancer Res Treat. 2011;129(1):37–47. doi: 10.1007/s10549-010-1177-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKallip RJ, Nagarkatti M, Nagarkatti PS. Δ-9-tetrahydrocannabinol enhances breast cancer growth and metastasis by suppression of the antitumor immune response. J Immunol. 2005;174(6):3281–3289. doi: 10.4049/jimmunol.174.6.3281. [DOI] [PubMed] [Google Scholar]
- McKallip RJ, Jia W, Schlomer J, Warren JW, Nagarkatti PS, Nagarkatti M. Cannabidiol-induced apoptosis in human leukemia cells: a novel role of cannabidiol in the regulation of p22phox and Nox4 expression. Mol Pharmacol. 2006;70(3):897–908. doi: 10.1124/mol.106.023937. [DOI] [PubMed] [Google Scholar]
- McPartland JM. ‘Cannabis systematics at the levels of family, genus, and species. Cannabis Cannabinoid Res. 2018 doi: 10.1089/can.2018.0039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millar SA, Stone NL, Yates AS, O'Sullivan SE. A systematic review on the pharmacokinetics of cannabidiol in humans. Front Pharmacol. 2018;26(9):1365. doi: 10.3389/fphar.2018.01365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mimeault M, Pommery N, Wattez N, Bailly C, Hénichart JP. Anti-proliferative and apoptotic effects of anandamide in human prostatic cancer cell lines: implication of epidermal growth factor receptor down-regulation and ceramide production. Prostate. 2003;56:1–12. doi: 10.1002/pros.10190. [DOI] [PubMed] [Google Scholar]
- Mohammadpour F, et al. Anti-invasion effects of cannabinoids agonist and antagonist on human breast cancer stem cells. Iran J Pharm Res. 2017 doi: 10.22037/ijpr.2017.2143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morales P, Reggio PH, Jagerovic N. An overview on medicinal chemistry of synthetic and natural derivatives of cannabidiol. Front Pharmacol. 2017;8:422. doi: 10.3389/fphar.2017.00422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morell C, Bort A, Vara D, Ramos-Torres A, Rodríguez-Henche N, Díaz-Laviada I. The cannabinoid WIN 55,212–2 prevents neuroendocrine differentiation of LNCaP prostate cancer cells. Prostate Cancer Prostatic Dis. 2016;19(3):248–257. doi: 10.1038/pcan.2016.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno E, Cavic M, Krivokuca A, Casadó V, Canela E. The endocannabinoid system as a target in cancer diseases: are we there yet? Front Pharmacol. 2019;10:339. doi: 10.3389/fphar.2019.00339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller L, Radtke A, Decker J, Koch M, Belge G. The synthetic cannabinoid WIN 55,212–2 elicits death in human cancer cell lines. Anticancer Res. 2017;37(11):6341–6345. doi: 10.21873/anticanres.12086. [DOI] [PubMed] [Google Scholar]
- Munson AE, Harris LS, Friedman MA, Dewey WL, Carchman RA (1975) Antineoplastic activity of cannabinoids. J Natl Cancer Inst 55(3):597–602. 10.1093/jnci/55.3.597 [DOI] [PubMed]
- Nallathambi R, Mazuz M, Namdar D, Shik M, Namintzer D, Vinayaka AC, Ion A, Faigenboim A, Nasser A, Laish I, Konikoff FM, Koltai H. Identification of synergistic interaction between cannabis-derived compounds for cytotoxic activity in colorectal cancer cell lines and colon polyps that induces apoptosis-related cell death and distinct gene expression. Cannabis Cannabinoid Res. 2018;3(1):120–135. doi: 10.1089/can.2018.0010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ngwa W, Kumar R, Moreau M, Dabney R, Herman A. Nanoparticle drones to target lung cancer with radiosensitizers and cannabinoids. Front Oncol. 2017;7:208. doi: 10.3389/fonc.2017.00208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ngwa W, Irabor OC, Schoenfeld JD, Hesser J, Demaria S, Formenti SC. Using immunotherapy to boost the abscopal effect. Nat Rev Cancer. 2018;18(5):313–322. doi: 10.1038/nrc.2018.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Shaughnessy WB. On the preparations of the Indian Hemp, or gunjah*. Prov Med Surg J. 1843 doi: 10.1136/bmj.1.2629.1171. [DOI] [Google Scholar]
- Ohoka N, Yoshii S, Hattori T, Onozaki K, Hayashi H. TRB3, a novel ER stress-inducible gene, is induced via ATF4-CHOP pathway and is involved in cell death. EMBO J. 2005;24(6):1243–1255. doi: 10.1038/sj.emboj.7600596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orellana-Serradell O, Poblete CE, Sanchez C, Castellón EA, Gallegos I, Huidobro C, Llanos MN, Contreras HR. Proapoptotic effect of endocannabinoids in prostate cancer cells. Oncol Rep. 2015;33(4):1599–1608. doi: 10.3892/or.2015.3746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ortega A, García-Hernández VM, Ruiz-García E, Meneses-García A, Herrera-Gómez A, Aguilar-Ponce JL, Montes-Servín E, Prospero-García O, Del Angel SA. Comparing the effects of endogenous and synthetic cannabinoid receptor agonists on survival of gastric cancer cells. Life Sci. 2016;15(165):56–62. doi: 10.1016/j.lfs.2016.09.010. [DOI] [PubMed] [Google Scholar]
- Parker LA, Rock EM, Limebeer CL. Regulation of nausea and vomiting by cannabinoids. Br J Pharmacol. 2011 doi: 10.1111/j.1476-5381.2010.01176.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pellati F, Borgonetti V, Brighenti V, Biagi M, Benvenuti S, Corsi L. Cannabis sativa L. and nonpsychoactive cannabinoids: their chemistry and role against oxidative stress, inflammation, and cancer. Biomed Res Int. 2018;2018:1691428. doi: 10.1155/2018/1691428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RG. Cannabinoid pharmacology: the first 66 years. Br J Pharmacol. 2006 doi: 10.1038/sj.bjp.0706406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pietrovito L, et al. Treatment with cannabinoids as a promising approach for impairing fibroblast activation and prostate cancer progression. Int J Mol Sci. 2020 doi: 10.3390/ijms21030787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pisanti S, et al. The endocannabinoid signaling system in cancer. Trends Pharmacol Sci. 2013 doi: 10.1016/j.tips.2013.03.003. [DOI] [PubMed] [Google Scholar]
- Pistis M, O’Sullivan SE. The role of nuclear hormone receptors in cannabinoid function. Adv Pharmacol. 2017 doi: 10.1016/bs.apha.2017.03.008. [DOI] [PubMed] [Google Scholar]
- Powles T, te Poele R, Shamash J, Chaplin T, Propper D, Joel S, Oliver T, Liu WM. Cannabis-induced cytotoxicity in leukemic cell lines: the role of the cannabinoid receptors and the MAPK pathway. Blood. 2005;105(3):1214–1221. doi: 10.1182/blood-2004-03-1182. [DOI] [PubMed] [Google Scholar]
- Preet A, Ganju R, Groopman J. Δ9-Tetrahydrocannabinol inhibits epithelial growth factor-induced lung cancer cell migration in vitro as well as its growth and metastasis in vivo. Oncogene. 2008;27:339–346. doi: 10.1038/sj.onc.1210641. [DOI] [PubMed] [Google Scholar]
- Proto MC, Fiore D, Piscopo C, et al. Inhibition of Wnt/β-Catenin pathway and Histone acetyltransferase activity by Rimonabant: a therapeutic target for colon cancer. Sci Rep. 2017;7:11678. doi: 10.1038/s41598-017-11688-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qamri Z, Preet A, Nasser MW, Bass CE, Leone G, Barsky SH, Ganju RK. Synthetic cannabinoid receptor agonists inhibit tumor growth and metastasis of breast cancer. Mol Cancer Ther. 2009;8(11):3117–3129. doi: 10.1158/1535-7163.MCT-09-0448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramer R, Heinemann K, Merkord J, Rohde H, Salamon A, Linnebacher M, Hinz B. COX-2 and PPAR-γ confer cannabidiol-induced apoptosis of human lung cancer cells. Mol Cancer Ther. 2013;12(1):69–82. doi: 10.1158/1535-7163.MCT-12-0335. [DOI] [PubMed] [Google Scholar]
- Ravi J, Elbaz M, Wani NA, Nasser MW, Ganju RK. Cannabinoid receptor-2 agonist inhibits macrophage induced EMT in non-small cell lung cancer by downregulation of EGFR pathway. Mol Carcinog. 2016;55(12):2063–2076. doi: 10.1002/mc.22451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romano B, Borrelli F, Pagano E, Cascio MG, Pertwee RG, Izzo AA. Inhibition of colon carcinogenesis by a standardized Cannabis sativa extract with high content of cannabidiol. Phytomedicine. 2014;21(5):631–639. doi: 10.1016/j.phymed.2013.11.006. [DOI] [PubMed] [Google Scholar]
- Ruiz L, Miguel A, Diaz-Laviada I. v9-Tetrahydrocannabinol induces apoptosis in human prostate PC-3 cells via a receptor-independent mechanism. FEBS Lett. 1999 doi: 10.1016/S0014-5793(99)01073-X. [DOI] [PubMed] [Google Scholar]
- Salazar M, Carracedo A, Salanueva ÍJ, Hernández-Tiedra S, Lorente M, et al. Cannabinoid action induces autophagy-mediated cell death through stimulation of ER stress in human glioma cells. J Clin Investig. 2009;119(5):1359–1372. doi: 10.1172/jci37948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez MG, Sanchez AM, Ruiz-Llorente L, Diaz-Laviada I. Enhancement of androgen receptor expression induced by (R)-methanandamide in prostate LNCaP cells. FEBS Lett. 2003;555:561–566. doi: 10.1016/s0014-5793(03)01349-8. [DOI] [PubMed] [Google Scholar]
- Santoro A, Pisanti S, Grimaldi C, Izzo AA, Borrelli F, Proto MC, Malfitano AM, Gazzerro P, Laezza C, Bifulco M. Rimonabant inhibits human colon cancer cell growth and reduces the formation of precancerous lesions in the mouse colon. Int J Cancer. 2009;125(5):996–1003. doi: 10.1002/ijc.24483. [DOI] [PubMed] [Google Scholar]
- Sarfaraz S, Afaq F, Adhami VM, Malik A, Mukhtar H. Cannabinoid receptor agonist-induced apoptosis of human prostate cancer cells LNCaP proceeds through sustained activation of ERK1/2 leading to G1 cell cycle arrest. J Biol Chem. 2006;281:39480–39491. doi: 10.1074/jbc.M603495200. [DOI] [PubMed] [Google Scholar]
- Sarnataro D, Grimaldi C, Pisanti S, Gazzerro P, Laezza C, Zurzolo C, Bifulco M. Plasma membrane and lysosomal localization of CB1 cannabinoid receptor are dependent on lipid rafts and regulated by anandamide in human breast cancer cells. FEBS Lett. 2005;579(28):6343–6349. doi: 10.1016/j.febslet.2005.10.016. [DOI] [PubMed] [Google Scholar]
- Sarnataro D, Pisanti S, Santoro A, Gazzerro P, Malfitano AM, Laezza C, Bifulco M. The cannabinoid CB1 receptor antagonist rimonabant (SR141716) inhibits human breast cancer cell proliferation through a lipid raft-mediated mechanism. Mol Pharmacol. 2006;70(4):1298–1306. doi: 10.1124/mol.106.025601. [DOI] [PubMed] [Google Scholar]
- Schiffmann S, Sandner J, Birod K, Wobst I, Angioni C, Ruckhäberle E, Kaufmann M, Ackermann H, Lötsch J, Schmidt H, Geisslinger G, Grösch S. Ceramide synthases and ceramide levels are increased in breast cancer tissue. Carcinogenesis. 2009;30(5):745–752. doi: 10.1093/carcin/bgp061. [DOI] [PubMed] [Google Scholar]
- Schoeman R, Beukes N, Frost C. Cannabinoid combination induces cytoplasmic vacuolation in MCF-7 breast cancer cells. Molecules (basel, Switzerland) 2020;25(20):4682. doi: 10.3390/molecules25204682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schröder M, Kaufman RJ. The mammalian unfolded protein response. Annu Rev Biochem. 2005;74(1):739–789. doi: 10.1146/annurev.biochem.73.011303.074134. [DOI] [PubMed] [Google Scholar]
- Scott KA, Dalgleish AG, Liu WM. The combination of cannabidiol and D 9-tetrahydrocannabinol enhances the anticancer effects of radiation in an orthotopic murine glioma model. Mol Cancer Ther. 2014 doi: 10.1158/1535-7163. [DOI] [PubMed] [Google Scholar]
- Senkal CE, Ponnusamy S, Rossi MJ, Bialewski J, Sinha D, Jiang JC, Jazwinski SM, Hannun YA, Ogretmen B. Role of human longevity assurance gene 1 and C18-ceramide in chemotherapy-induced cell death in human head and neck squamous cell carcinomas. Mol Cancer Ther. 2007;6(2):712–722. doi: 10.1158/1535-7163.MCT-06-0558. [DOI] [PubMed] [Google Scholar]
- Sharma M, et al. In vitro anticancer activity of plant-derived cannabidiol on prostate cancer cell lines. Pharmacol Pharm. 2014 doi: 10.4236/pp.2014.58091. [DOI] [Google Scholar]
- Shaw J, Costa-Pinheiro P, Patterson L, Drews K, Spiegel S, Kester M. Novel sphingolipid-based cancer therapeutics in the personalized medicine era. Adv Cancer Res. 2018;140:327–366. doi: 10.1016/bs.acr.2018.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrivastava A, Kuzontkoski PM, Groopman JE, Prasad A. Cannabidiol induces programmed cell death in breast cancer cells by coordinating the cross-talk between apoptosis and autophagy. Mol Cancer Ther. 2011;10(7):1161–1172. doi: 10.1158/1535-7163. [DOI] [PubMed] [Google Scholar]
- Singer E, Judkins J, Salomonis N, Matlaf L, Soteropoulos P, McAllister S, Soroceanu L. Reactive oxygen species-mediated therapeutic response and resistance in glioblastoma. Cell Death Dis. 2015;6(1):e1601. doi: 10.1038/cddis.2014.566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Śledziński P, Zeyland J, Słomski R, Nowak A. The current state and future perspectives of cannabinoids in cancer biology. Cancer Med. 2018;7(3):765–775. doi: 10.1002/cam4.1312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soderstrom K, Soliman E, Dross RV. Cannabinoids modulate neuronal activity and cancer by CB1 and CB2 receptor-independent mechanisms. Front Pharmacol. 2017;8:720. doi: 10.3389/fphar.2017.00720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soliman E, Henderson KL, Danell AS, Van Dross R. Arachidonoyl-ethanolamide activates endoplasmic reticulum stress-apoptosis in tumorigenic keratinocytes: Role of cyclooxygenase-2 and novel J-series prostamides. Mol Carcinog. 2016;55(2):117–130. doi: 10.1002/mc.22257. [DOI] [PubMed] [Google Scholar]
- Solinas M, Massi P, Cinquina V, Valenti M, Bolognini D, Gariboldi M, Monti E, Rubino T, Parolaro D (2013) Cannabidiol, a non-psychoactive cannabinoid compound, inhibits proliferation and invasion in U87-MG and T98G glioma cells through a multitarget effect. PLoS One. 8(10):e76918. 10.1371/journal.pone.0076918 [DOI] [PMC free article] [PubMed]
- Sreevalsan S, Joseph S, Jutooru I, Chadalapaka G, Safe SH. Induction of apoptosis by cannabinoids in prostate and colon cancer cells is phosphatase dependent. Anticancer Res. 2011;31(11):3799–3807. [PMC free article] [PubMed] [Google Scholar]
- Torres S, Lorente M, Rodríguez-Fornés F, Hernández-Tiedra S, Salazar M, García-Taboada E, Barcia J, Guzmán M, Velasco G. A combined preclinical therapy of cannabinoids and temozolomide against glioma. Mol Cancer Ther. 2011;10(1):90–103. doi: 10.1158/1535-7163.MCT-10-0688. [DOI] [PubMed] [Google Scholar]
- Tutino V, Caruso MG, De Nunzio V, Lorusso D, Veronese N, Gigante I, Notarnicola M, Giannelli G. Down-regulation of cannabinoid type 1 (CB1) receptor and its downstream signaling pathways in metastatic colorectal cancer. Cancers. 2019;11(5):708. doi: 10.3390/cancers11050708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twelves C, Sabel M, Checketts D, et al. A phase 1b randomised, placebo-controlled trial of nabiximols cannabinoid oromucosal spray with temozolomide in patients with recurrent glioblastoma. Br J Cancer. 2021;124:1379–1387. doi: 10.1038/s41416-021-01259-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vara D, Salazar M, Olea-Herrero N, et al. Anti-tumoral action of cannabinoids on hepatocellular carcinoma: role of AMPK-dependent activation of autophagy. Cell Death Differ. 2011;18:1099–1111. doi: 10.1038/cdd.2011.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Velasco G, Sánchez C, Guzmán M. Towards the use of cannabinoids as antitumour agents. Nat Rev Cancer. 2012;12(6):436–444. doi: 10.1038/nrc3247. [DOI] [PubMed] [Google Scholar]
- Velasco G, Sánchez C, Guzmán M. Anticancer mechanisms of cannabinoids. Curr Oncol. 2016;23:23–32. doi: 10.3747/co.23.3080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vrechi TAM, Leão AHFF, Morais IBM, et al. Cannabidiol induces autophagy via ERK1/2 activation in neural cells. Sci Rep. 2021;11:5434. doi: 10.1038/s41598-021-84879-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang M, Zhao J, Zhang L, Wei F, Lian Y, Wu Y, Gong Z, Zhang S, Zhou J, Cao K, Li X, Xiong W, Li G, Zeng Z, Guo C. Role of tumor microenvironment in tumorigenesis. J Cancer. 2017;8(5):761–773. doi: 10.7150/jca.17648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J. Cannabis, cannabinoid receptors, and endocannabinoid system: yesterday, today, and tomorrow. Acta Pharmacol Sin. 2019;40(3):297–299. doi: 10.1038/s41401-019-0210-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xian X, Huang L, Zhang B, Wu C, Cui J, Wang Z. WIN 55,212–2 inhibits the epithelial mesenchymal transition of gastric cancer cells via COX-2 signals. Cell Physiol Biochem. 2016;39(6):2149–2157. doi: 10.1159/000447910. [DOI] [PubMed] [Google Scholar]
- Xu D, Wang J, Zhou Z, He Z, Zhao Q. Cannabinoid WIN55, 212–2 induces cell cycle arrest and inhibits the proliferation and migration of human BEL7402 hepatocellular carcinoma cells. Mol Med Rep. 2015;12(6):7963–7970. doi: 10.3892/mmr.2015.4477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Huynh N, Dumesny C, Wang K, He H, Nikfarjam M. Cannabinoids inhibited pancreatic cancer via P-21 activated kinase 1 mediated pathway. Int J Mol Sci. 2020;21(21):8035. doi: 10.3390/ijms21218035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasmin-Karim S, Moreau M, Mueller R, Sinha N, Dabney R, Herman A, Ngwa W. Enhancing the therapeutic efficacy of cancer treatment with cannabinoids. Front Oncol. 2018;24(8):114. doi: 10.3389/fonc.2018.00114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu Z, Pestell TG, Lisanti MP, Pestell RG. Cancer stem cells. Int J Biochem Cell Biol. 2012;44(12):2144–2151. doi: 10.1016/j.biocel.2012.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Zheng W, Shen K, Shen W. ∆9-tetrahydrocannabinol inhibits epithelial-mesenchymal transition and metastasis by targeting matrix metalloproteinase-9 in endometrial cancer. Oncol Lett. 2018;15(6):8527–8535. doi: 10.3892/ol.2018.8407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Qin Y, Pan Z, Li M, Liu X, Chen X, Qu G, Zhou L, Xu M, Zheng Q, Li D. Cannabidiol induces cell cycle arrest and cell apoptosis in human gastric cancer SGC-7901 cells. Biomolecules. 2019;9(8):302. doi: 10.3390/biom9080302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao P, Abood ME. GPR55 and GPR35 and their relationship to cannabinoid and lysophospholipid receptors. Life Sci. 2013 doi: 10.1016/j.lfs.2012.06.039. [DOI] [PubMed] [Google Scholar]
- Zhu LX, Sharma S, Stolina M, Gardner B, Roth MD, Tashkin DP, Dubinett SM. Delta-9-tetrahydrocannabinol inhibits antitumor immunity by a CB2 receptor-mediated, cytokine-dependent pathway. J Immunol. 2000;165(1):373–380. doi: 10.4049/jimmunol.165.1.373. [DOI] [PubMed] [Google Scholar]
- Zou S, Kumar U. Cannabinoid receptors and the endocannabinoid system: signaling and function in the central nervous system. Int J Mol Sci. 2018;19(3):833. doi: 10.3390/ijms19030833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuardi AW. History of cannabis as a medicine: a review. Rev Bras Psiquiatr. 2006 doi: 10.1590/S1516-44462006000200015. [DOI] [PubMed] [Google Scholar]