

Summary
Bacillus cereus is a common food‐borne pathogen that is responsible for important outbreaks of food poisoning in humans. Diseases caused by B. cereus usually exhibit two major symptoms, emetic or diarrheic, depending on the toxins produced. It is assumed that after the ingestion of contaminated vegetables or processed food, spores of enterotoxigenic B. cereus reach the intestine, where they germinate and produce the enterotoxins that are responsible for food poisoning. In our study, we observed that sporulation is required for the survival of B. cereus in leaves but is dispensable in ready‐to‐eat vegetables, such as endives. We demonstrate that vegetative cells of B. cereus that are originally impaired in sporulation but not biofilm formation are able to reach the intestine and cause severe disorders in a murine model. Furthermore, our findings emphasise that the number of food poisoning cases associated with B. cereus is underestimated and suggest the need to revise the detection protocols, which are based primarily on spores and toxins.
Bacillus cereus is a common food‐borne pathogen Sporulation is not a determinant for the persistence of enterotoxigenic B. cereus on ready‐to‐eat vegetables.
Introduction
A major problem in food safety is the contamination of fresh and stored produce with human pathogens (Carlin et al., 2000; Burnett and Beuchat, 2001), which may provoke food poisoning outbreaks (Berget et al., 2010). Bacillus cereus is a natural inhabitant of soils and is able to live in association with plants. Members of this group of bacteria are also responsible for food poisoning outbreaks through consumption of raw vegetables and fruits or processed food (Bottone, 2010; Elhariry, 2011). Food poisoning caused by B. cereus is classified into two main categories, emetic and diarrheic, depending on the battery of toxins produced (Granum and Lund, 1997). An emetic poisoning is associated with the production of a dodecadepsipeptide called cereulide, (Agata et al., 1994; Agata et al., 1995b) a remarkably heat‐ and acid‐stable toxin typically detected in high concentrations in processed food contaminated with B. cereus (Ehling‐Schulz et al., 2004). Diarrheic poisoning is known to occur after the ingestion of food contaminated with enterotoxigenic B. cereus spores which, upon germination in the intestine, produce heat‐labile enterotoxins (McKillip, 2000; Berthold‐Pluta et al., 2015). A group of three primary enterotoxins are involved in diarrheic poisoning: haemolysin BL (Hbl), non‐haemolytic enterotoxin (Nhe) and cytotoxin K (CytK) (Lund et al., 2000; Stenfors Arnesen et al., 2008).
Bacillus cereus spores are highly adhesive and resistant to stress, which ensures their persistence in a variety of environments, under various sanitation measures and in ready‐to‐eat or processed food (Gao et al., 2015). Under more favourable environmental conditions, spores readily germinate to vegetative cells and begin new life cycles (Wiencek et al., 1990). In addition to spores, contamination with B. cereus in the food industry is attributable to the formation of biofilms, in which bacterial communities assemble in response to a variety of signals that enable microorganisms to adapt to different environments (Danhorn and Fuqua, 2007). In the proposed life cycle of B. cereus, these microbes live as saprophytes in soil, move to plants and later enter human‐related environments (hospitals, the food industry or human bodies). Spores or vegetative cells rotate depending on the life cycle stage, and they are proposed to remain as individual cells or become embedded in biofilms.
In this study, we investigated the relevance of spores and biofilms of B. cereus to the persistence of these species on infrequently investigated plants. The comparative analysis of a group of isolates (Table 1) involved in food poisoning enabled us to propose that sporulation is a major ecological trait in the phylloplane (i.e. on leaf surfaces) but that sporulation is dispensable in ready‐to‐eat vegetables. Additionally, we demonstrate that intrinsic genetic defects in sporulation do not diminish the pathogenicity of vegetative cells of B. cereus in a murine model.
Table 1.
Bacillus cereus strains used in this study.
| Strains | Origin | Toxins | Reference a |
|---|---|---|---|
| AH187 | Emetic outbreak (Ehling‐Schulz et al., 2005) | Nhe/Cereulide | BGSC |
| ATCC 14579 | Cowshed (Ceuppens et al., 2013) | Hbl/Nhe/CytK | CECT |
| DSM 4282 | Diarrheal outbreak (Ehling‐Schulz et al., 2005) | Hbl/Nhe/CytK | DSMZ |
| DSM 4313 | Diarrheal outbreak (Ehling‐Schulz et al., 2005) | Hbl/Nhe/CytK | DSMZ |
| DSM 14729 | Unknown | Hbl/Nhe/CytK | DSMZ |
| UMAF8564 | Melon rhizosphere (García‐Gutiérrez et al., 2012) | Hbl/Nhe | UMA |
| 6A27 | Diarrheal outbreak (Ehling‐Schulz et al., 2005) | Hbl/Nhe | BGSC |
| DSM 8438 | Diarrheal outbreak (Ehling‐Schulz et al., 2005) | Nhe/CytK | DSMZ |
| DSM 2302 | Foodstuff (Crovadore et al., 2016) | Hbl/Nhe/CytK | DSMZ |
Origin indicates the source where the strains were isolated. Toxins are the main enterotoxin or emetic toxin produced by each stain. Reference corresponds to the culture collection where the strains were obtained.
CECT (Spanish Type Culture Collection). BGSC (Bacillus Genetic Stock Center). DSMZ (German Collection of Microorganisms and Cell Cultures). UMA (University of Malaga).
Results
Sporulation permits B. cereus to persist on melon leaves but is dispensable on endive
We investigated the ability of a group of B. cereus strains isolated from different outbreaks of food poisoning (Table 1) to colonize and persist on leave of two different plant species, melon and endive. Two differentiated behaviours of B. cereus strains were observed on and in melon leaves: seven strains (AH187, ATCC 14579, DSM 4282, DSM 4313, DSM 14729, UMAF8564 and 6A27) were capable of persisting at 104–106 CFUs per gram of leaf and had a spore content of 20–40% (Fig. S1), and two strains (DSM 8438 and DSM 2302) which the total CFU and spores counts per gram of leaf were below the detection threshold (Fig. S1). In endive, however, all the strains persisted at 104–106 CFU per gram of leaf. The percentages of sporulation were similar to those recorded on melon leaves with the exception of DSM 2302, which did not show any sign of sporulation. With these findings, we compared the population dynamics of strain AH187 (a representative of the group of persister and sporulating strains), DSM 8438 and DSM 2302 (non‐persister on melon leaf and non‐sporulating strains) on both plant leaves (Fig. 1). The population size of AH187 progressively decreased and stabilized at 106 CFU per gram of melon leaf 8 days after inoculation; however, DSM 8438 and DSM 2302 showed a sharp decrease within the first 4 days and became undetectable after 8 days of inoculation (Fig. 1). The AH187 population dynamics on endive leaves were similar to those on melon leaves, and DSM 8438 and DSM 2302 stabilized at 104‐105 CFU per gram of endive leaves after 2 DAI (Fig. 1). The sporulation rate of AH187 and DSM 8438 increased starting at 4–6 HAI, and no spores were detected for DSM 2302. Similar results for the persister group were recorded on cucumber leaves and fruit, and DSM 8438 and DSM 2302 did not sporulate in any of the studied cucumber organs (Fig. S1).
Fig. 1.
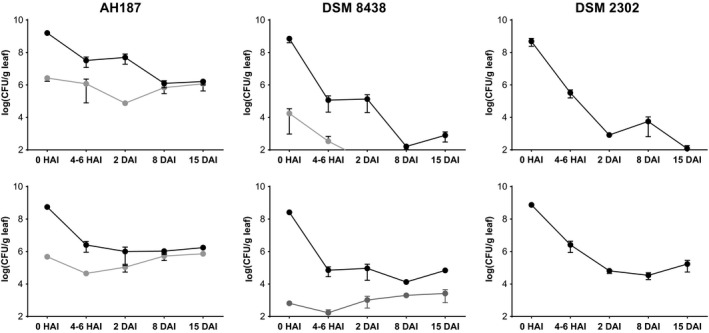
Sporulation is essential for the persistence of B. cereus on melon leaves but not on endive leaves. CFU counts per gram of melon and endive leaf after inoculation (0 h (HAI) 4–6 h (HAI), 2 days, 8 days and 15 days (DAI)) with B. cereus strains AH187, DSM 8438 or DSM 2302 strains. The top plots correspond to melon leaf counts, and the bottom plots correspond to endive counts. The black line shows the counts of total CFUs on either melon or endive leaves. The grey line shows the CFU counts corresponding to spores on melon and endive leaves. The detection limit of the technique is 102 CFU g‐1 leaf. Error bars are the standard error of three or more replicates.
B. cereus DSM 2302 is genetically impaired in sporulation but capable of biofilm formation
We sequenced and compared the genomes of all the strains used in this study to define the genetic defects for the inability of DSM 2302 to sporulate, and we searched for genetic traits defining a plant‐associated lifestyle. Multilocus sequence analysis using 4 conserved genes (16S rRNA, gyrB, rpoB and rpoD) demonstrated that AH187, DSM 2302 and DSM 8438 belonged to completely separate phylogenetic branches (Fig. S2). This finding enabled us to surmise that the strains that do not share specific niches employ different evolutionary strategies and that their evolutionary distance might be related to their different lifestyles. A deeper study of the pangenome of the nine B. cereus strains (Fig. S3A) identified a total of 48 844 coding DNA sequences (CDSs) in the nine B. cereus strains. Of these total CDSs, 30 746 were highly conserved across all nine genomes and comprised the core genome of these B. cereus strains. The core genes were clustered into 3 326 orthologous groups (OGs), the additional genes (see material and methods for details) were clustered into 3 325 Ogs, and the unique genes of the nine strains comprised 3050; therefore, all CDSs were clustered into 9 702 Ogs. Notably, AH187 and DSM 2302 had the largest number of unique genes, exhibiting a total of 444 and 414 exclusive sequences, respectively, which supported our hypothesis on the specialization of these strains to different ecological niches (Table S3). Further analysis of the core genome by 10 different combinations of the sequences of the nine studied strains demonstrated that the number of genes did not decrease with the increase in the number of genomes added to the study (Fig. S3B). In addition, the pangenome of the studied strains was open, indicating that each sequenced genome contributed new genes to the pangenome but never reached an exact number of genes (Fig. S3B). A more specific comparison of DSM 2302 with AH187 showed an uneven distribution of the genes or the absence of several complete regions, including operons (Fig. S3C), as well as widely conserved regions in both strains.
Consistent with the apparent phenotypic defect in sporulation exhibited by DSM 2302, 36 genes involved in the sporulation programme were absent (Table 2, Fig. 2D) (Illing and Errington, 1991; Smith and Youngman, 1993; Tovar‐Rojo et al., 2002). To confirm this genotype, we analysed the intrinsic ability of the strains to sporulate in the spore‐promoting medium DSM (Fig. 2B). All the strains that could persist over melon leaves showed high sporulation rates in this medium (Fig. 2B, grey bars), but no sign of sporulation was observed in cultures of DSM 2302. Contrast‐phase microscopy analysis of single cells confirmed the presence of refringent endospores within AH187 cells but neither endospores nor mother cell‐released spores in cultures of DSM 2302 (Fig. 2B).
Table 2.
Genes absent in DSM 2302 strain compared with AH187 strain.
| Gene | Product |
|---|---|
| BCAH187_A0483 | SpoIIM; stage II sporulation protein M a , b |
| BCAH187_A0633 | Sporulation sigma factor‐processing peptidase b |
| BCAH187_A0925 | Spore germination protein AC |
| BCAH187_A1089 | Acid‐soluble spore protein b |
| BCAH187_A1120 | YjcZ family sporulation protein a , b |
| BCAH187_A1125 | YjcZ family sporulation protein a , b |
| BCAH187_A1216 | YjcZ family sporulation protein a , b |
| BCAH187_A1409 | Spore‐specific protein b |
| BCAH187_A2125 | Spore permease b |
| BCAH187_A2217 | AbrB family transcriptional regulator |
| BCAH187_A2515 | Spo0E; stage 0 sporulation regulatory protein a , b |
| BCAH187_A2546 | Exosporium protein H |
| BCAH187_A2674 | Spore germination protein gerPA/gerPF b |
| BCAH187_A2711 | YjcZ family sporulation protein a , b |
| BCAH187_A2919 | CotX; spore coat protein X |
| BCAH187_A2920 | CotW; spore coat protein W b |
| BCAH187_A2921 | Spore coat protein X |
| BCAH187_A3146 | CotF; spore coat protein F |
| BCAH187_A3147 | YqcI/YcgG family protein a , b |
| BCAH187_A3151 | Small acid‐soluble spore protein A (major alpha‐type SASP) |
| BCAH187_A3165 | Spore germination protein GerAC |
| BCAH187_A3166 | Spore germination protein A2 |
| BCAH187_A3167 | Spore germination protein GerAA a , b |
| BCAH187_A3201 | Putative spore germination B3 GerAC family protein |
| BCAH187_A3200 | GerKB; spore germination protein KB a . Spore permease b |
| BCAH187_A3202 | Spore germination protein KA |
| BCAH187_A3600 | GerSB; Spore germination protein |
| BCAH187_A4093 | YjcZ family sporulation protein a , b |
| BCAH187_A4323 | SpoIIIAA; stage III sporulation protein AA |
| BCAH187_A4629 | Germination protein gerE |
| BCAH187_A5258 | Exosporium leader peptide‐containing protein b |
| BCAH187_A5308 | SpoVAD; stage V sporulation protein AD |
| BCAH187_A5309 | SpoVAE; stage V sporulation protein AE |
| BCAH187_C0228 | SpoVAD; stage V sporulation protein AD |
| BCAH187_C0229 | SpoVAE; stage V sporulation protein AE |
| BCAH187_E0003 | Sporulation protein a , b |
Gene corresponds to AH187 locus absent in DSM 2302 strain. Product indicates the protein associated to each gene.
The annotation of the product corresponds to orthologous genes in other Bacillus species.
The annotation of the product corresponds to GO/KO term or Pfam domain function.
Fig. 2.
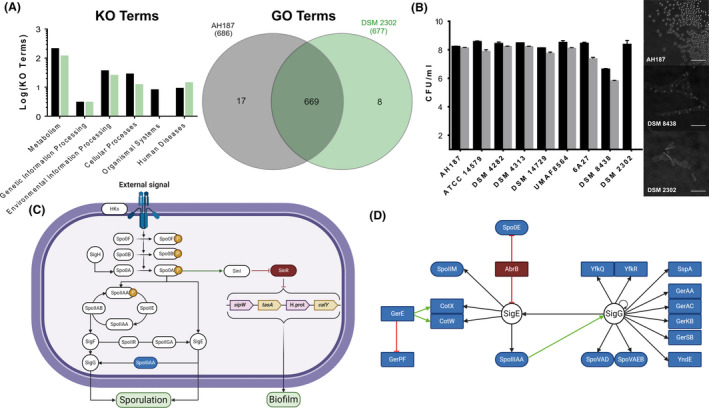
B. cereus DSM 2302 is unable to sporulate, but it retains the machinery to form biofilms and cause human diseases.
A. KO and GO term comparison between AH187 and DSM 2302 strains. Black bars and grey circles correspond to the AH187 strain, while green bars and circles are the DSM 2302 strain. The KO plot shows unique KO terms of each strain compared to the other. Pie plot showing GO terms shared and non‐shared between AH187 and DSM 2302 strains. See also Table S3, which shows KO and GO terms present in DSM 2302 and not in the AH187 strain.
B. Sporulation of B. cereus strains in DSM medium after 12 h incubation at 30°C with shaking. The plot shows the counts of total CFU ml‐1 (black bars) and spores (grey bars) plated in LB agar after incubation in DSM medium. The detection limit of the technique is 101 CFU ml‐1. Error bars are the standard error of three or more replicates. Statistical analysis is the result of the comparison of all strains against AH187. *(P < 0,05), **(P < 0,01), ***(P < 0,001). The pictures correspond to microscopy images showing the different sporulation behaviours of the AH187, DSM 8438 and DSM 2302 strains after 12 h of incubation at 30°C with shaking in DSM medium. Scale bar corresponds to 10 µm.
C. Sporulation and biofilm pathway in B. cereus group. Blue proteins are absent in DSM 2302 strain. Created with BioRender.com.
D. Sporulation pathway of genes absent in B. cereus DSM 2302. Blue proteins refer to genes present in AH187 and absent in strain DSM 2302 in addition to AbrB regulator. Green arrows indicate an activation process, and red arrows indicate repression of the target gene. SigE and SigG are present in DSM 2302 genome. Created with BioRender.com.
In Bacillus, sporulation and biofilms are known to contribute to the persistence of Bacillus and other species in different habitats, and these two cellular destinies are under the control of the master regulator spo0A (Fig. 2C). We disregarded a hypothetical collateral interruption of the biofilm regulatory pathway in DSM 2302, given that most of the lacking genes related to sporulation were structural and belonged to stages of the sporulation programme downstream of phosphorelay (Table 2). In addition, we identified the two major regulators of the expression of biofilm‐related genes sinR and sinI along with the gene cluster sipW‐to‐calY, known to be essential for the assembly of the extracellular matrix (ECM) (Fig. S4B). Another indispensable component of the ECM is exopolysaccharides (EPSs). Two genomic regions containing EPS‐related genes were found in DSM 2302: (i) one similar to the so‐called eps1 of the type strain ATCC 14579 and orthologous to the epsA‐O of B. subtilis. This region was also found in strain AH187 which, notably, varied in the genes of the central core of the region (Fig. S4C). (ii) One region homologous to the so‐called eps2 region of ATCC 14579, which was absent in strain AH187 (Fig. S4D). According to this genetic information, DSM 2302 was able to form biofilms in static cultures, although phenotypically differentiated from AH187. Colonies of DSM 2302 stained with Congo Red (known to be related to the product of the eps2 region in the type strain ATCC 14579) and adhered to the walls and bottom of the wells but failed to form pellicles in the air–liquid interphase of static liquid cultures. Strain AH187 formed colonies that stained slightly with Congo Red, formed pellicles in the air–liquid interphase and adhered to the wells in static liquid cultures. (Fig. S4A). The pangenomic analysis of DSM 2302 additionally showed several genes involved in the assembly of the ECM, polysaccharide and putrescine biosynthesis, all of which are putative structural integrant of the biofilm programme (Table S4).
Further genomic analysis demonstrated the presence of genes related to a plant‐associated lifestyle in the genome of AH187 but absent in DSM 2302. These genes included several response regulators; ABC transporters hypothetically involved in exoprotein production, competence or sporulation; a regulator inducible by reactive oxygen species (ROS); a putative endoglucanase involved in plant cell wall degradation; diverse surface proteins and important enzymes related to plant nitrogen sources, such as asparagine, glutamine or proline (Table S5). In contrast, DSM 2302 had more genes related to human disorders, bacterial invasion of epithelial cells, bacterial extracellular infection, cell wall surface anchoring, cell adhesion and internalization, and antimicrobial resistance in the genome (Fig. 2A), most of which were observed in the related food‐borne bacteria (Roux et al., 2005; Franciosa et al., 2009).
In summary, the defect of sporulation, the lack of genes related to the metabolism of nitrogen sources related to plants and the overrepresentation of genes associated with pathogenic interactions with humans are consistent with the low survival of DSM 2302 on plants.
Metabolic nature of endive enables the persistence of vegetative cells of B cereus
In addition to the lack of sporulation, the different fitness levels of DSM 2302 on melon or endives leaves raised the question of the major influence of the host. With in‐depth metabolomic analysis of leaf tissue, the chemical profiles of both plant species demonstrated significant differences in metabolic families and metabolite concentrations (Fig. S5). Carbohydrates, amino acids, lipids and nucleotides were largely observed in the endive (Fig. 3A). More specifically, chlorogenate, allantoic acid, quinate, galactarate and glucarate were clearly the most representative endive metabolites. However, secondary metabolites of flavonoids (such as isovitexin), glutathione, pinoresinol and siderophores were the most frequently observed metabolites in melon leaves (Fig. 3B).
Fig. 3.
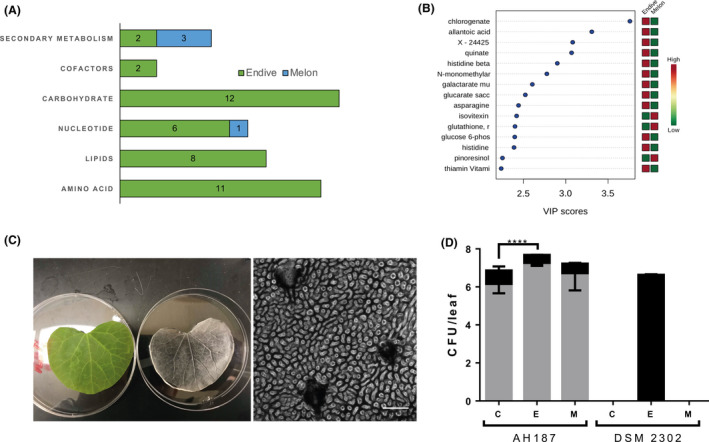
Differences in the nutrient content of the plant host impact the survival of non‐sporulating B. cereus DSM 2302.
A. Major metabolites grouped by superfamilies found in melon and endive leaves.
B. Important metabolites identified by PLS‐DA. The coloured boxes on the right indicate the relative concentration of the corresponding metabolite in each studied surface (red indicates a higher relative concentration, and green indicates a lower relative concentration of the corresponding metabolite).
C. The picture on the left shows a melon leaf and its corresponding replica of PDMS (artificial leaf surface). The image on the right is a micrograph of the surface of an artificial melon leaf. Scale bar corresponds to 100 µm.
D. Total CFU (black bars) and spore counts (grey bars) per artificial leaf surface: without metabolites (C) with the majority endive metabolites (quinate, galactarate and glucarate) (E) and the majority flavonoid (isovitexin) found in melon leaves (M) 15 days after inoculation with B. cereus AH187 and DSM 2302 strains. The black bar corresponds to the total CFUs per artificial leaf surface, and the grey bars are the counts of CFUs corresponding to spores. The detection limit of the technique is 102 CFU/g leaf. Error bars are the standard error of three replicates. Statistical analysis is the result of the comparison of the DSM 2302 strain against AH187. *(P < 0.05), **(P < 0.01), ***(P < 0.001), ****(P < 0.0001).
These findings along with the bacterial population dynamics showed above suggested that endive leaves were more conducive to bacterial growth, while melon leaves represented a more aggressive and stressful environment for bacteria. To confirm this hypothesis, we studied the bacterial behaviour in PDMS artificial surfaces that mirrored the topology of melon leaves (Fig. 3C) but were supplemented with a solution of quinate, galactarate and saccharate (5:1:1), the major components detected in endive. AH187 persisted on melon artificial leaves alone or supplemented with endive metabolites; however, DSM 2302 only persisted in the presence of endive‐related metabolites (Fig. 3D). The addition of isovitexin, the main flavonoid detected in melon leaves, permitted an insignificant slight increase in the AH187 population but this flavonoid did not prop any growth of DSM 2302 (Fig. 3D). The spore content of AH187 remained unchanged at 30‐40% under experimental conditions (Fig. 3D) According to these findings, a null sigE (a mother cell‐specific transcription factor of early sporulation in Bacillus spp.) mutant of AH187, unable to sporulate but capable of forming biofilms, could not sporulate or persist on melon leaves or on artificial leaf surfaces lacking the major endive‐related metabolites.
Vegetative cells of DSM 2302 can reach the intestine in mice and cause illness
The strain DSM 2302 was isolated in a food poisoning outbreak, and we have demonstrated that vegetative cells can persist on endive, a produce that is directly consumed by humans. This evidence suggests that vegetative cells of this strain may reach humans and provoke food poisoning. To test this idea, we simulated a stomach passage with basal pH conditions (pH 2) and after meal ingestion conditions (pH 4.5) using a nutritive broth (JB) as a food source. Only AH187 survived at pH 2, and this strain was observed at very low levels and largely existing as spores (Fig. 4A). The population densities of AH187 or DSM 2302 were comparable and remained unchanged at a pH value of 4.5 for 5 h. Thus, vegetative cells of DSM 2302 should be able to overcome the acidic condition of the stomach after food ingestion and reach the intestine. To confirm this possibility, we inoculated 108 CFU of DSM 2302 into 3‐month‐old mice. One‐third of mice inoculated with DSM 2302 cells died after 6 h of bacterial inoculation. We evaluated the histological damage of the intestine of the mice as indirect evidence of the presence of this strain in the guts. Haematoxylin–eosin staining showed continuity of the epithelium in the villi and crypts of the intestinal microvilli in untreated mice (Fig. 4B). However, the mice inoculated with DSM 2302 showed disruption of the epithelium. Similar results were obtained in immunohistochemical analyses using anti‐Ck or anti‐pan‐cadherin antibodies and confocal microscopy (Fig. 4B).
Fig. 4.
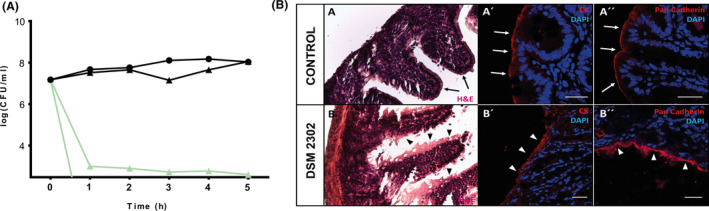
Vegetative cells of DSM 2302 can survive simulating stomach conditions and cause symptoms in mouse intestines. A Counts of total CFU/ml on gastric media (GM) supplemented with JB broth of AH187 (▲) and DSM 2302 (●) strains at 37°C for 5 h. Black lines correspond to pH 4.5 simulating conditions after food intake. Green lines correspond to pH 2 (basal stomach conditions). The detection limit of the technique is 101 CFU ml‐1. Error bars are the standard error of three replicates. B Comparative histological caecum analysis of intestine sections from control and infected mice. Haematoxylin–eosin staining showed (A) epithelium continuity in the villi and crypts of the intestinal microvilli in control intestine (PBS) and (B) epithelium disruption of intestines of infected mice. Immunohistochemistry and confocal microscopy showed epithelial continuity in the villi and crypts of the intestinal microvilli with CK (A′; arrows) and pan‐cadherin (A′′, arrows) markers of the control intestine and epithelial disruption in the infected caecums using CK (B′; arrowheads) or pan‐cadherin (B′′, arrowheads). All nuclei were stained with DAPI for immunofluorescence. Scale Bars (A′, A′′, B′, B′′) = 25 mm.
These results enable us to conclude that vegetative cells of DSM 2302 (which lack of the acid resistance cereulide toxin) are capable of passing through the stomach and reaching the gut, subsequently causing illness in the host.
Discussion
Various studies have previously reported the relevance of B. cereus spores in food poisoning caused by the consumption of contaminated raw food or vegetables (Turnbull, 1976; Schoeni and Wong, 2005). It is assumed that both vegetative cells and spores can reach the intestine, but the survival percentage of spores is considerably higher due to their resistance to stomach conditions (Wijnands et al., 2009; Ceuppens et al., 2012a). Therefore, based on this presumption, the few vegetative cells that survive are insufficient to reach the minimum infective dose required to cause disease (105–108 CFU) (Stenfors Arnesen et al., 2008). In keeping with this model, it is believed that diarrheic diseases provoked by enterotoxigenic B. cereus are associated with spore‐contaminating foodstuffs, which germinate in the gut and produce the enterotoxins responsible for the symptoms (Clavel et al., 2007; Ceuppens et al., 2012a; Ceuppens et al., 2012b). In our study, we demonstrate that sporulation is not a determinant ecological trait for the persistence of enterotoxigenic B. cereus on ready‐to‐eat vegetables.
Sporulation and biofilm formation are two ecological traits contributing to the completion of the life cycle of B. cereus. Our genomic analysis demonstrated that strain DSM 2302, which is naturally unable to sporulate, lacks sporulation genes belonging to mid‐late stages of sporulation. However, the first stages related to the phosphorelay and thus sporulation decision‐making, as well as other cellular destinies, such as biofilms, are preserved (Hamon and Lazazzera, 2001; Parashar et al., 2013; Fagerlund et al., 2014; Majed et al., 2016). The inability to persist on/in melon leaves which was observed for both DSM 2302 or a sigE mutant of the emetic strain (AH187) that was affected in sporulation but not biofilm formation on melon leaves, indicated a major contribution of sporulation over biofilm formation in facilitating long‐term bacterial survival in this ecological niche. Consistent with this, metabolomic analysis demonstrated that melon leaves are scarce in carbon and nitrogen sources and dominated by secondary metabolites, especially flavonoids, known as antibacterials and antibiofilms (Cushnie and Lamb, 2005; Awolola et al., 2014; Matilla‐Cuenca et al., 2020). Contrary to this scenario, the endive is rich in nutrients and thus supportive of vegetative growth (Fig. 3B), most likely favouring biofilm formation as the preferred long‐term bacterial survival strategy. We hypothesize that similar to the findings that have been reported for B. subtilis in the interaction with the rhizosphere, the collection of sensor kinases exposed on bacterial cell surfaces may discriminate the variety of signals and thus the final cellular decision‐making (Chen et al., 2012). Therefore, it remains to be determined how different metabolites found in our analysis are sensed by these Bacillus cells and integrated in the sporulation or biofilm pathways.
In addition to biofilm formation and sporulation, our genomic analysis showed a group of genes related to the use of plant carbon and nitrogen sources in the emetic strain AH187 (representative of Bacillus strain survival on plants), and other persisters strains, but not in DSM 2302. Even though the absence of these genes would be considered an adaptive disadvantage a priori, mutational analysis of these genes in strain AH187 did not indicate that they affect bacterial fitness on leaves (Fig. S6). In alignment with our findings, a previous study also found that genes related to the use of plant carbon sources do not impart an ecological advantage; in fact, the presence of amino acids in food, such as vegetables, could improve bacterial adaptability (Duport et al., 2016). However, considering that only viable single mutants could be tested, we could not determine whether this genetic background may collectively contribute to the survival of bacterial cells on plants, either as single vegetative cells or spores or as communities within biofilms.
It is generally assumed that in B. cereus life cycle spores reach the stomach and germinate, giving rise to metabolically active vegetative cells capable of colonizing the gut and producing the toxin responsible for damage to the host (Clavel et al., 2004; Stenfors Arnesen et al., 2008). We have, however, demonstrated that vegetative cells of DSM 2302 are able to overcome acid conditions typically found in the stomach after food ingestion (pH 4.5) (Fig. 4A) (Clavel et al., 2004) for 5 h, which is sufficient time for the food to move forward from the stomach to the intestine (Camilleri et al., 1989; Dressman et al., 1990). At this stage, two possible but non‐exclusive bacterial behaviours are the formation of biofilms associated with the gut and the production of enterotoxins. Genomic analysis of DSM 2302 showed the presence of well‐known genes of the biofilm pathway (regulatory and structural elements) and other genes whose products are related to such traits as bacterial cell adhesion to the host tissue, ECM assembly, swarming motility (Sturgill and Rather, 2004; Patel et al., 2006; Flemming and Wingender, 2010; Nwodo et al., 2012) or bacterial invasion of epithelial cells, as previously reported for the human food poisoning pathogens L. monocytogenes and E. coli (Roux et al., 2005; Franciosa et al., 2009). In addition, DSM 2302 adheres to the bottom of the wells of static cultures, a low oxygen concentration environment that mimics the gut (Fig. S4A). DSM 2302 is genetically determined to produce the three primary known enterotoxins (Hbl, Nhe and Cytk) and the enterotoxins BceT, haemolysin I (cerolysin O), haemolysin II (HlyII), haemolysin III (HlyIII) and enterotoxin CwpFM (EntFM). BceT is lethal against mice, haemolysins are involved in lysing and permeabilizing host cells or intracellular organelles, and these toxins also have haemolytic activity against blood cells. EntFM is involved in bacterial virulence, motility, adhesion to epithelial cells and biofilm formation (Agata et al., 1995a; Tran et al., 2010; Ramarao and Sanchis, 2013). All these findings indicate that DSM 2302 has the necessary arsenal to adhere to epithelial cells of the gut and produce the toxins responsible for food poisoning in humans.
In summary, we identified a strain of B. cereus (DSM 2302) naturally impaired in sporulation but still able to survive in certain vegetables and caused illness in mammal hosts (Fig. 5). Based on our findings, we propose that the persistence of this strain on plants relies on the metabolic nature of the host and the ability of the bacteria to form biofilms. Vegetative cells bearing structural elements related to biofilm formation (adhesins or EPSs) and protected by food can cross the stomach of humans and reach the intestine. The biofilm‐related structural factors would further ensure either the adhesion of cells to the intestine or the formation of biofilms, as well as the production of enterotoxins that ensure the release of nutrients from the human host. From a clinical perspective, our findings reinforce the presumed underestimation of food poisoning cases caused by B. cereus and the need to improve the detection protocols that largely rely on the presence of spores or toxins in food (Soni et al., 2016; Vidic et al., 2020).
Fig. 5.
Reconsidering the life cycle of B. cereus from ready‐to‐eat vegetables to humans. Non‐sporulating strains could survive over foodstuff products such as vegetables because of the large amount of nutrients compared to plant leaves, which are a more hostile niche. Furthermore, the ingestion of these contaminated foods increases stomach pH, and these conditions allow vegetative cells to survive stomach passage and enter the intestine to provoke human food poisoning. Created with BioRender.com.
Experimental procedures
Bacterial strains and growth conditions
Nine B. cereus isolates from food poisoning outbreaks were used in this study (Table 1). Bacteria were routinely pre‐cultured in LB broth (10 g l‐1 tryptone, 5 g l‐1 yeast extract and 5 g l‐1 NaCl) or in 1.5% LB agar. B. cereus strains were grown overnight with shaking at 28°C. To study biofilm formation, B. cereus strains were cultured for 72 h at 28°C without shaking in TY broth (10 g l‐1 tryptone, 5 g l‐1 yeast extract, 5 g l‐1 NaCl, 10 mM MgSO4 and 1 mM MnSO4) or 1.5% TY agar supplemented with 20 and 10 µg ml‐1 Congo Red and Coomassie Brilliant Blue filtered dyes, respectively (Caro‐Astorga et al., 2014). For swarming motility assays, 0.7% TY agar was used (Caro‐Astorga et al., 2020). For sporulation experiments, cells were cultured in Difco Sporulation Medium (DSM) (Schaeffer et al., 1965) for 24 h with shaking at 28°C. To simulate in vitro gastric passage, the cells were incubated in J broth (JB) (5 g l‐1 peptone, 15 g l‐1 yeast extract, 3 g l‐1 K2HPO4 and 2 g l‐1 filtered glucose) (Clavel et al., 2004).
Genome sequencing, assembly and annotation
Bacterial DNA was isolated using the JetFlex™ Genomic DNA Purification Kit (Thermo Fisher Scientific, Bremen, Germany) following the kit protocol for Gram‐positive bacteria. A DNA library was generated by NextSeq 550 (Illumina, United Kingdom) sequencing. The genome assembly was performed using the B. cereus type strain (ATCC 14579) as the reference strain of the group with the workflow of Pipeline A5‐miseq (Coil et al., 2015). The assembled genomes were annotated with the NCBI Prokaryotic Genome Annotation Pipeline (PGAP) version 4.3 algorithm (Angiuoli et al., 2008) and deposited in NCBI GenBank (Table S1). Genomes of strains used in this work (ATCC 14579, AH187 and DSM 2302) were sequenced, and their genomes were obtained from the NCBI complete bacterial genome repository (http://www.ncbi.nlm.nih.gov/genome/). This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession PHKO00000000. The version described in this paper is version PHKO0000000.
Comparative genome, pangenome and phylogenetic analysis
Genomic analyses for comparison of the strains listed in Table 1 were performed as follows: amino acid sequences for each strain were obtained from the GenBank files using the Perl programming language. Pfam domains (Finn et al., 2016) were identified inside the proteins using a Perl script and an InterProScan algorithm in the InterPro database (Jones et al., 2014). Protein–protein comparisons among the strains of the analysis were performed to obtain the bidirectional best hits (BBH), and orthologous and paralogous proteins were identified using the NCBI BLASTP algorithm. The protein sequences were clustered according to a percentage of identity > 80%, which retained the same Pfam domain with the OrthoMCL algorithm (Li et al., 2003). The protein clusters were classified into three different groups: core (proteins shared among all the studied strains), accessory (proteins shared among all study strains except one or less) and unique (proteins only present in one of the studied strains and not in the others). To compare the function of B. cereus strains, GO terms (The Gene Ontology Consortium, 2015) and KO terms (KEGG Orthology) were obtained and analysed using InterProScan. The WEGO (Web Gene Ontology Annotation Plot) tool (http://wego.genomics.org.cn/) was used for GO term annotation, and the KAAS server (https://www.genome.jp/kegg/kaas/) (Kanehisa et al., 2016a) and BlastKOALA (https://www.kegg.jp/blastkoala/) (Kanehisa et al., 2016b) were used for KO term annotation. GO and KO term comparisons and visualization were performed using GraphPad Prism version 6.0 (GraphPad Software, San Diego, CA, USA) and InteractiVenn (Heberle et al., 2015). Multilocus sequence phylogenetic analyses of the nine strains used for the pangenomic analysis, employing Bacillus subtilis subsp. subtilis 168 as an outgroup reference organism, was performed using the nucleotide sequences of the 16S rRNA, DNA gyrase B subunit gyrB, RNA polymerase sigma 70 factor rpoD and RNA polymerase beta 70 factor rpoB genes (Das et al., 2014). The MUSCLE algorithm included in MEGA7 software was utilized to align the concatenated gene sequences. Next, a maximum likelihood tree was built with the MEGA7 program using the neighbour‐joining substitution model, and bootstrap values of each branch were calculated 1000 times. To study the presence/absence of sporulation genes, a comparison between the B. cereus AH187 and B. cereus DSM 2302 genomes was performed with the Gview server tool (https://server.gview.ca). The B. cereus AH187 gff file was obtained from NCBI (ASM2122v1), and the B. cereus DSM 2302 fasta file was generated in the pangenome study described elsewhere in this publication. Unique genes were used to identify the Gene Ontology functional categories using comparativeGo (https://www.comparativego.com). Sporulation genes from these two strains were selected and compared by blast2 sequencing to obtain the final list of sporulation genes of B. cereus AH187 with no matches in B. cereus DSM 2302. To determine the possible function of genes annotated as hypothetical proteins or with unknown function, their amino acid sequences were compared with proteins from other organism using the HHpred tool of the Max Planck Institute for Developmental Biology (Tübingen, Germany) (Söding, 2005; Zimmermann et al., 2018). We also searched for orthologous genes in other Bacillus species, or we determined the hypothetical function according to the corresponding GO/KO term of the Pfam domain.
Sporulation and image acquisition
The sporulation assays were performed in DSM medium. One colony of each strain was picked from an LB agar plate and resuspended in DSM broth. Each strain was incubated at 28°C with shaking for 24 h. Bacterial concentrations in CFU ml‐1 were determined by plating 100 µl aliquots of 10‐fold serial dilutions onto LB agar. To estimate the proportion of spores, each dilution was heated at 80°C for 10 min and subsequently plated. Microscopy images from DSM cultures were taken on a Nikon Eclipse Ti fluorescence microscope with a Super Plan Fluor objective 100x equipped with a Hamamatsu Orca R2 monochrome 1.3 MP CCD camera. Image processing (for proportion of spores) was performed using ImageJ software.
Construction of a B. cereus sporulation mutants
B. cereus mutants were obtained by electroporation using the pMAD plasmid (Arnaud et al., 2004). To generate mutagenesis constructs, NEBuilder HiFi DNA Assembly Master Mix (New England Biolabs, MA, USA) was utilized following the manufacturer’s instructions, and specific primers (Table S2) were employed to amplify the upstream and downstream regions of the target gene. The pMAD vector was digested with SmaI and incubated at 50°C for 1 h in a thermocycler with the upstream and downstream fragments. The total amount of fragments was 0.2 pmol with a 1:2 DNA ratio (vector:insert). The constructions were utilized to transform electrocompetent B. cereus cells by electroporation as described previously with several modifications (Silo‐Suh et al., 1994; Caro‐Astorga et al., 2014). One microgram of plasmid was incubated with 100 µl of electrocompetent cell suspension on ice for 5–10 min. The mixture of bacteria and DNA was electroporated in a 0.2‐cm chilled cuvette using the following electroporation parameters: 1.4 kV voltage, 25 µF capacitance and 200 Ω resistance. The electroporated cells were recovered in LB at 30°C for 5 h and then plated onto LB plates supplemented with erythromycin to select bacterial transformants. To obtain mutants, B. cereus transformants were incubated at 40°C without antibiotics to block plasmid replication and force the shuttle vector to integrate into the chromosome via homologous recombination.
Dynamics of bacterial cells on plants
To evaluate the persistence of B. cereus strains on plants, melon (Cucumis melo cv. Rochet Panal) (Fitó, Barcelona, Spain) leaves, cucumber (Cucumis sativus cv. Marketer) (Fitó, Barcelona, Spain) leaves, cucumber fruit and endive (Cichorium endivia) were analysed. Melon and cucumber plants were grown under greenhouse conditions, and fresh cucumber fruit and endive were purchased in a grocery store. Cucumber fruit and endive were rinsed with 1% (w/v) sodium hypochlorite and 96% ethanol in distilled water for 4 min before being inoculated with B. cereus cell suspensions. A bacterial suspension (1 ml) at a concentration of 108 CFU ml‐1 in distilled water was spread onto the adaxial axis of second and third leaves of melon or cucumber leaves and around cucumber fruit and endive leaves. After bacterial inoculation, the melon and cucumber plants were incubated in a growth chamber at 25°C with a 16‐h light/8‐h dark photoperiod and 85% relative humidity. Endive and cucumber fruit were incubated at room temperature. Bacterial persistence and sporulation were evaluated 15 days after inoculation. Leaves and cucumber fruit skin were placed individually into sterile plastic stomacher bags with 10 ml of distilled water and homogenized for 3 min in a stomacher homogenizer (Interscience, France). One hundred microlitres of 10‐fold serial dilutions of the homogenates were plated onto LB agar plates and incubated at 28°C for 24 h. To estimate the level of spores, serial dilutions were incubated at 80°C for 10 min to kill vegetative cells, and the spores were subsequently plated and incubated overnight at 28°C.
Biofilm and swarming motility assays
In vitro biofilm assays of B. cereus strains were performed in TY broth and in TY 1.5% agar supplemented with 20 and 10 µg ml‐1 of Congo Red and Coomassie Brilliant Blue filtered dyes, respectively (Romero et al., 2010; Caro‐Astorga et al., 2014). For biofilm assays in TY broth, a colony from a pre‐culture in LB agar at 28°C for each strain was previously resuspended in 1 ml of TY medium; 10 µl of this culture was inoculated into 1 ml of TY broth in the centre wells of a 24‐well plate, and the plate was incubated without shaking at 28°C for 72 h. To evaluate bacterial adhesion to abiotic surfaces (i.e. adhesion to the well wall), the biofilm of each well was stained with 1% crystal violet solution (O’Toole et al., 2000) for 3 min and subsequently rinsed several times with distilled water until the excess dye was removed. Finally, the plate was dried on filter paper on a bench. To study biofilm formation on agar plates (colony morphology and staining), 2 µl of the pre‐culture described above was inoculated separately on 1.5% TY agar plates with Congo Red and Coomassie Brilliant Blue, and the plates were incubated at 28°C for 72 h. To study swarming motility, 0.7% TY agar plates were utilized. Two microlitres of the pre‐culture described above were inoculated on TY plates and incubated at 28°C for 72 h before measuring the diameter of colonies.
Leaf and fruit metabolome analysis
Metabolomic analysis of melon and endive leaves was performed by Metabolon® (Durham, NC, USA). One hundred milligrams of each tissue (from a pool of leaves) was homogenized and lyophilized. Samples were prepared using the automated MicroLab STAR® system from Hamilton Company (Franklin, MA, USA). Samples were extracted with methanol under vigorous shaking for 2 min to precipitate protein and dissociate small molecules bound to protein or trapped in the precipitated protein matrix followed by centrifugation to recover chemically diverse metabolites. The resulting extract was divided into four fractions: two for analysis by two separate reversed‐phase (RP)/UPLC‐MS/MS methods using positive ion mode electrospray ionization (ESI), one for analysis by RP/UPLC‐MS/MS using negative ion mode ESI, and one for analysis by HILIC/UPLC‐MS/MS using negative ion mode ESI. Samples were placed briefly on a TurboVap® (Zymark) to remove the organic solvent. The metabolic analysis was performed by ultra‐performance liquid chromatography (UPLC) coupled to a mass spectrometer with a heated electrospray ionization (HESI‐II) source and Orbitrap mass analyser operated at 35,000 mass resolution. The four extracts were dried and subsequently reconstituted in methanol. One aliquot was analysed using acidic positive ion conditions, chromatographically optimized for more hydrophilic compounds and subsequently gradient‐eluted from a C18 column (Waters UPLC BEH C18‐2.1 × 100 mm, 1.7 µm) using water and methanol containing 0.05% perfluoropentanoic acid (PFPA) and 0.1% formic acid (FA). A second aliquot was also analysed using acidic positive ion conditions but was chromatographically optimized for more hydrophobic compounds. The extract was gradient‐eluted from the aforementioned C18 column using methanol, acetonitrile, water, 0.05% PFPA and 0.01% FA and was operated at an organic content that was higher overall. A third aliquot was analysed using basic negative ion optimized conditions using a separate dedicated C18 column. The basic extracts were gradient‐eluted from the column using methanol and water but with 6.5 mM ammonium bicarbonate at pH 8. The fourth aliquot was analysed via negative ionization following elution from a HILIC column (Waters UPLC BEH Amide 2.1 × 150 mm, 1.7 µm) using a gradient consisting of water and acetonitrile with 10 mM ammonium formate, pH 10.8. The MS analysis alternated between MS and data‐dependent MSn scans using dynamic exclusion. The scan range varied slightly between methods but covered approximately 70–1000 m/z. Raw data were extracted, peaks were identified, and data were submitted to QC processing using Metabolon’s hardware and software.
To visualize the differences between melon and endive leaves, MetaboAnalyst (https://www.metaboanalyst.ca/) was used to generate the heatmap with hierarchical clustering and to determine the most important metabolites through PLS‐DA analysis.
Construction of artificial leaf surfaces, bacterial inoculation and incubation
To produce artificial leaf surfaces, melon leaves were grown as described above. The second and third leaves were taken to mould the leaf surface replica. The process of making artificial leaves was undertaken as previously described (Doan et al., 2020a) and summarized as follows: first, a negative stamp of the adaxial part of the leaf was created. The negative stamp was made with a mixture of 10:1 polydimethylsiloxane (PDMS) and curing agent from a Sylgard 184 PDMS elastomer kit (Dow Chemical, Midland, TX, USA). The mixture was poured over the real leaf previously joined with double‐sided adhesive tape on a Petri dish and incubated at 30°C for 17–24 h. When the negative stamp was solidified, it was carefully separated from the real leaf and subsequently exposed to UV light (400 F, 400 W and λ = 187–254 nm) for 1 h to fix the PDMS. After UV exposure, the negative stamp was treated with a mixture of 1000:1 toluene and octadecyltrichlorosilane (ODTS) for 5 min to create a soft cover that facilitated the later separation of the negative stamp from the positive stamp. Next, the negative stamp was rinsed with absolute ethanol for 2 min and dried for at least 10 h at room temperature. To make the positive stamp (artificial leaf), the PDMS mixture was made as described above, and it was poured onto the negative stamp, which was attached to a Petri dish. When the PDMS mixture was solidified, the positive stamp was carefully separated from the negative stamp. The fabricated artificial leaves were autoclaved and rinsed with a mixture of 1:1000 Triton X‐100 in distilled water. Artificial leaves were cut into coupons measuring 25.75 cm in diameter and inoculated with 20‐µl drops (5 drops per coupon) of a 108 CFU ml‐1 bacterial suspension in distilled water (Elbing and Brent, 2002; Doan et al., 2020b) or in distilled water supplemented with 0.5% quinate, 0.1% galactarate and 0.1% saccharate or with 1 mM isovitexin to mimic endive and melon leaf metabolite profiles, respectively (obtained from the metabolomic analysis). Inoculated coupons were incubated over moistened filter paper with 10 ml of distilled water into a Petri dish plate or without humidity conditions (without distilled water), allowing the drops to dry on an open Petri dish. All the plates with coupons were incubated for 15 days in a growth chamber as described above for real leaves. After 15 days, the coupons were vortexed for 15 s and sonicated for 10 min in an ultrasonic bath in 10 ml of distilled water in a 50‐ml tube (Doan et al., 2020b). Tenfold serial dilutions were made, and 100 µl of each dilution was plated on LB agar. To estimate the level of spores, the dilutions were heated at 80°C for 10 min and plated and incubated at 28°C for 24 h to obtain CFU counts.
Gastric passage simulation
To analyse gastric passage simulation after food ingestion, we employed the protocol described previously (Clavel et al., 2004; Clavel et al., 2007): B. cereus strains were incubated in LB agar at 30°C for 24 h. One colony was transferred into a 250‐ml flask with 40 ml of J broth (JB) and incubated for 18 h at 30°C and 130 rpm. B. cereus cells were added to a 1:1 mixture of JB and gastric media (GM) (4.8 g l‐1 NaCl, 1.56 g l‐1 NaHCO3, 2.2 g l‐1 KCl and 0.22 g l‐1 CaCl2) at pH 2 and 4.5 plus 500 U l‐1 pepsin to an initial concentration of 106–107 CFU ml‐1 bacteria and incubated at 37°C for 5 h and 130 rpm to simulate human stomach conditions. Samples were taken hourly, 10‐fold serial dilutions were made in 0.2 M sodium phosphate buffer at pH 7 to neutralize the stomach pH, and 100 µl of each dilution were plated onto LB agar. For the determination of spores, the dilutions were heated at 80°C for 10 min and plated onto LB agar.
Inoculation of mice with B. cereus and histological analysis of intestines
CD1 female mice (3 months old) were purchased from the Andalusian Centre for Nanomedicine and Biotechnology (University of Malaga). Each experimental group consisted of six mice housed in cages that were maintained at a controlled temperature and light cycle; the mice were fed an autoclaved standard balanced conventional diet and a gelatine‐only diet 24 h before the infection. Animals were handled in accordance with international regulations for animal welfare. Bacteria were administered by intragastric gavage using a blunt‐end needle. Six randomly allocated mice were challenged with 100 µl of 108 CFU of B. cereus DSM 2302 in 1x PBS or with 100 µl of 1× PBS (Rolny et al., 2014). After 6 h of bacterial administration, mice were anaesthetized with an injection of 200 ml of a mix of 10% ketamine, 6% xylazine and 84% water each and sacrificed by cervical dislocation. Samples from the caecum region were aseptically collected and washed in PBS and immediately fixed in 4% formalin overnight (4°C) for histological preparation. To obtain frozen sections, tissues were cryoprotected in 15% and 30% sucrose:PBS solutions, embedded in OCT, snap‐frozen in liquid N2‐cooled isopentane and sectioned in a cryostat. OCT blocks were cut into 10‐mm sections, and some of them were stained with haematoxylin–eosin. At least three caeca were analysed for each treatment.
Immunohistochemistry and confocal microscopy
Immunofluorescence analyses were performed by blocking non‐specific binding sites with 16% sheep serum, 1% bovine serum albumin and 0.5% Triton X‐100 in PBS (SBT) and incubating either tissue section slide with primary antibodies (anti‐cytokeratin ‘DAKO Z0622, 1:100 dilution’ and anti‐PanCadherin ‘Sigma C3678, 1:100 dilution’) overnight at 4°C. Next, the samples were washed in PBS (3 × 5 min) and incubated with the secondary antibody (Alexa Fluor® 647 ‘InmunoJackson 711‐605‐152’, 1:200 dilution) + DAPI (Sigma D9542, 1:2000 dilution) for 2 h at room temperature. All samples were finally washed in PBS (3 × 5 min), mounted in a 1:1 PBS/glycerol solution and analysed using an SP5 laser confocal microscope (LEICA, Wetzlar, Germany).
Ethics approval and informed consent for mouse experiments
Experiments were carried out in accordance with the Spanish Institutional Animal Use and Care Committee regulations and with the approval of the Regional Andalusian Government (code A/ES/14/43). The methods were carried out in accordance with the relevant guidelines and regulations.
Statistical analysis
The statistical analyses were performed using the statistical software GraphPad Prism version 6.0 (GraphPad Software, CA, USA). For melon and endive leaf experiments, one‐way non‐parametric ANOVA (Kruskal–Wallis test) was performed with Dunn’s multiple comparison tests. For artificial surface experiments, two‐way ANOVA (with the Bonferroni test) was applied. The results with P‐values < 0.05 were considered to be significant. Metabolome data were normalized for internal consistency by processing a constant weight of sample per volume of extraction solvent. No post hoc mathematical normalization was imposed on the data. Data were scaled to the median value for each compound, and missing values were imputed with the minimum detected value for that compound. Statistical calculations were performed using natural log‐transformed scaled imputed data (table not included).
Funding Information
This work was supported by grants from European Research Council Starting Grant (BacBio 637971), and Plan Nacional de I + D+i of Ministerio de Economía y Competitividad, Ministerio de Ciencia e Innovación (AGL2016‐78662‐R and PID2019‐107724GB‐I00). Johan HJ Leveau was financed by USDA‐NIFA grant number 2014‐67017‐21695. Juan Antonio Guadix acknowledges financial support by grant (PIER‐0084‐2019) from Proyectos de Investigación en Salud de Junta de Andalucía.
Conflict of interests
The authors declare no competing interests.
Author contributions
D.R. conceived the study. D.R. and M.L.A.G. designed the experiments. M.L.A.G. performed the main experimental work. M.L.A.G. and L.D.M analysed and processed the whole genomes. M.L.A.G. and J.H. designed the metabolite assays over artificial leaf surfaces. M.L.A.G., H.K.D and J.H.J.L. made and design the artificial leaf surfaces protocol. S.S.T. processed the real surfaces for metabolome analysis. A.M.S.T. and J.A.G. performed the mouse assays and analysis. D.R. and M.L.A.G. wrote the manuscript. D.R., M.L.A.G., L.D.M., J.H., J.A.G., J.H.J.L. and A.V. contributed critically to writing the final version of the manuscript.
Supporting information
Fig. S1. Persistence and sporulation of B. cereus strains on melon and cucumber leaves, cucumber fruit or endive. CFU counts per gram of leaf fruit 15 days after inoculation with B. cereus AH187, ATCC 14579, DSM 4282, DSM 4313, DSM 14729, UMAF8565, 6A27, DSM 8438 and DSM 2302. The black bar plots at the top show the total CFU count per gram of leaf or fruit. The grey bar graphs at the bottom correspond to the spore count per gram of leaf or fruit. The detection limit of the technique is 102 CFU/g of leaf. Error bars are the standard error of three or more replicates. Statistical analysis is the result of the comparison of all strains against AH187. *(P<0,05), **(P<0,01), ***(P<0,001) (Dunn’s test). No counts of the DSM 2302 strain appeared in any sporulation assay or on any surface, except endive. No significant counts of DSM 8438 strain appeared over all the surfaces (total CFUs and spores), except endive.
Fig. S2. Phylogenetic relation of B. cereus AH187, ATCC 14579, DSM 4282, DSM 4313, DSM 14729, UMAF8564, 6A27, DSM 8438 and DSM 2302 strains. Red points indicate the strains that were not able to persist on the melon leaf surface (DSM 8438 and DSM 2302). B. subtilis subsp. subtilis 168 was used as an out‐group. In the analysis, four concatenated genes (16S rRNA, gyrB, rpoD and rpoB) were handled to build a neighbour‐joining tree using the MUSCLE algorithm of MEGA7.
Fig. S3. Pangenome analysis of the strains AH187, ATCC 14579, DSM 4282, DSM 4313, DSM 14729, UMAF8564, 6A27, DSM 8438 and DSM 2302. A Schematic view of the pangenome analysis of the nine studied strains. The inner layer represents the pangenome (all the analysed genes). Each colour layer corresponds to the genome of one B. cereus strain. B Statistical estimation with Willenbock (blue) and Tettelin (red) (Tettelin et al., 2005; Willenbrock et al., 2007). The plot on the top points corresponds to pangenome size according to the number of genomes. In the plot on the bottom, the points correspond to the calculation of the core genome for each genome. C Schematic view of the genomic comparison between strains AH187 and DSM 2302. Genes present in strain AH187 are represented as brown lines throughout the genome, while genes absent in strain DSM 2302 are represented as red lines just above previous. The percentage of CG is represented in black, and the GC skew is represented in purple to locate the intergenic zones and the origins of replication of these strains, respectively.
Fig. S4. Comparison of biofilm formation capacity of AH187 and DSM 2302. 72 h in vitro biofilm and swarming assays of AH187 and DSM 2302 strains on TY 1.5% agar supplemented with 20 µg/ml and 10 µg/ml Congo Red and Coomassie Brilliant Blue dyes (A1), TY 0.7% agar (A2) and TY broth (A3). A4 shows bacterial adhesion to an abiotic surface after staining with crystal violet. B sipW‐calY locus implicated in extracellular matrix synthesis of AH187 and DSM 2302 compared to the B. cereus type strain (ATCC 14579). C Region homologous to the eps1 region of ATCC 14579 involved in exopolysaccharide biosynthesis of AH187 and DSM 2302 strains. Middle grey genes correspond to different genes compared to each other. D Region homologous to the eps2 region of ATCC 14579 involved in exopolysaccharide biosynthesis found in AH187 and DSM 2302 strains. Grey genes correspond to different genes located in the eps2 genome region.
Fig. S5. Metabolomic profile of melon and endive leaf. Clustering result shown as a heatmap of three replicates of leaves of endive (orange) and melon leaves (green). In the heatmap, red indicates a higher concentration of a metabolite; in contrast, blue indicates a lower concentration of the metabolite.
Fig. S6. Persistence of AH187 mutants on melon leaves. CFU (black bar plot at the left) and spore (grey bar plot at the right) counts of AH187 mutants of the genes listed in Table S5 on melon leaves 15 days after bacteria inoculation. The detection limit of the technique is 102 CFU/g of leaf. Error bars are the standard error of three or more replicates. Statistical analysis is the result of the comparison of all the mutants against AH187 wild‐type. *(P<0.05), **(P<0.01), ***(P<0.001) (Dunn’s test). sigE mutant conts felt below the detection limit.
Table S1. Sequenced B. cereus strains deposited in Genbank. Sequenced B. cereus strains deposited in Genbank.
Table S2. Primers used to construct mutants of B. cereus AH187.
Table S3. Pangenome analysis of B. cereus strains. Core indicates the proteins shared among all the studied strains, accessory shows the proteins shared among all studied strains except one or less, and unique enumerates the proteins only present in one of the studied strains and not in the others. Differences in core proteins correspond to paralogous genes.
Table S4. GO and KO terms, obtained from the pangenome analysis that are present in B. cereus DSM 2302 strains and absent in AH187 strain. GO and terms associated with DSM 2302 strain with their corresponding function and DSM 2302 genes related with each GO/KO term.
Table S5. Genes absent in B. cereus DSM 2302 and DSM 8438. Genomic comparison among persister and non persister strains reveal genes non present in DSM 8438 and DSM 2302 strains.
Acknowledgements
We would like to thank Josefa Gómez from the Ultrasequencing Unit of the SCBI‐UMA for DNA sequencing, Zulema Udaondo from Bio‐Iliberis and Rocío Bautista from Bioinformatic Unit of the SCBI‐UMA for pangenome and genome analysis support, John Pearson from Bionand for guidance and assistance with microscopy images, María Isabel Castillo from Bionand‐UMA for managing and feeding mice and Dr. Atul Parikh from UC Davis for access to his laboratory for the fabrication of PDMS leaf replicants used in this study. This work was supported by grants from European Research Council Starting Grant (BacBio 637971), and Plan Nacional de I + D+i of Ministerio de Economía y Competitividad, Ministerio de Ciencia e Innovación (AGL2016‐78662‐R and PID2019‐107724GB‐I00). Johan HJ Leveau was financed by USDA‐NIFA grant number 2014‐67017‐21695. Juan Antonio Guadix acknowledges financial support by grant (PIER‐0084‐2019) from Proyectos de Investigación en Salud de Junta de Andalucía.
Microb. Biotechnol. (2021) 14(4), 1550–1565
References
- Agata, N. , Mori, M. , Ohta, M. , Suwan, S. , Ohtani, I. , and Isobe, M. (1994) A novel dodecadepsipeptide, cereulide, isolated from Bacillus cereus causes vacuole formation in hEp‐2 cells. FEMS Microbiol Lett 121: 31–34. [DOI] [PubMed] [Google Scholar]
- Agata, N. , Ohta, M. , Arakawa, Y. , and Mori, M. (1995a) The bceT gene of Bacillus cereus encodes an enterotoxic protein. Microbiology 141: 983–988. [DOI] [PubMed] [Google Scholar]
- Agata, N. , Ohta, M. , Mori, M. , and Isobe, M. (1995b) A novel dodecadepsipeptide, cereulide, is an emetic toxin of Bacillus cereus . FEMS Microbiol Lett 129: 17–19. [DOI] [PubMed] [Google Scholar]
- Angiuoli, S.V. , Gussman, A. , Klimke, W. , Cochrane, G. , Field, D. , Garrity, G.M. , et al. (2008) Toward an online repository of standard operating procedures (SOPs) for (meta)genomic annotation. Omi A J Integr Biol 12: 137–141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnaud, M. , Chastanet, A. , and Débarbouillé, M. (2004) New vector for efficient allelic replacement in naturally gram‐positive bacteria. Appl Environ Microbiol 70: 6887–6891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Awolola, G.V. , Koorbanally, N.A. , Chenia, H. , Shode, F.O. , and Baijnath, H. (2014) Antibacterial and anti‐biofilm activity of flavonoids and triterpenes isolated from the extracts of Ficus sansibarica Warb. subsp. sansibarica (Moraceae) extracts. Afr J Tradit Complement Altern Med 11: 124–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger, C.N. , Sodha, S.V. , Shaw, R.K. , Griffin, P.M. , Pink, D. , Hand, P. , and Frankel, G. (2010) Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ Microbiol 12: 2385–2397. [DOI] [PubMed] [Google Scholar]
- Berthold‐Pluta, A. , Pluta, A. , and Garbowska, M. (2015) The effect of selected factors on the survival of Bacillus cereus in the human gastrointestinal tract. Microb Pathog 82: 7–14. [DOI] [PubMed] [Google Scholar]
- Bottone, E.J. (2010) Bacillus cereus, a volatile human pathogen. Clin Microbiol Rev 23: 382–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnett, S.L. , and Beuchat, L.R. (2001) Human pathogens associated with raw produce and unpasteurized juices, and difficulties in decontamination. J Ind Microbiol Biotechnol 27: 104–110. [DOI] [PubMed] [Google Scholar]
- Camilleri, M. , Colemont, L.j. , Phillips, S.f. , Brown, M.l. , Thomforde, G.m. , Chapman, N. , and Zinsmeister, A.r. (1989) Human gastric emptying and colonic filling of solids characterized by a new method. Am J Physiol. – Gastrointest Liver Physiol 257: G284–G290. [DOI] [PubMed] [Google Scholar]
- Carlin, F. , Girardin, H. , Peck, M.W. , Stringer, S.C. , Barker, G.C. , Martinez, A. , et al. (2000) Research on factors allowing a risk assessment of spore‐forming pathogenic bacteria in cooked chilled foods containing vegetables: a FAIR collaborative project. Int J Food Microbiol 60: 117–135. [DOI] [PubMed] [Google Scholar]
- Caro‐Astorga, J. , Álvarez‐Mena, A. , Hierrezuelo, J. , Guadix, J.A. , Heredia‐Ponce, Z. , Arboleda‐Estudillo, Y. , et al. (2020) Two genomic regions encoding exopolysaccharide production systems have complementary functions in B. cereus multicellularity and host interaction. Sci Rep 10: 1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caro‐Astorga, J. , Pérez‐García, A. , de Vicente, A. , and Romero, D. (2014) A genomic region involved in the formation of adhesin fibers in Bacillus cereus biofilms. Front Microbiol 5: 745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceuppens, S. , Timmery, S. , Mahillon, J. , Uyttendaele, M. , and Boon, N. (2013) Small Bacillus cereus ATCC 14579 subpopulations are responsible for cytotoxin K production. J Appl Microbiol 114: 899–906. [DOI] [PubMed] [Google Scholar]
- Ceuppens, S. , Uyttendaele, M. , Hamelink, S. , Boon, N. , and Van De Wiele, T. (2012a) Inactivation of Bacillus cereus vegetative cells by gastric acid and bile during in vitro gastrointestinal transit. Gut Pathog 4: 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceuppens, S. , Uyttendaele, M. , Drieskens, K. , Heyndrickx, M. , Rajkovic, A. , Boon, N. , et al. (2012b) Survival and germination of Bacillus cereus spores without outgrowth or enterotoxin production during in vitro simulation of gastrointestinal transit. Appl Environ Microbiol 78: 7698–7705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, Y. , Cao, S. , Chai, Y. , Clardy, J. , Kolter, R. , Guo, J.‐H. , and Losick, R. (2012) A Bacillus subtilis sensor kinase involved in triggering biofilm formation on the roots of tomato plants. Mol Microbiol 85: 418–430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clavel, T. , Carlin, F. , Lairon, D. , Nguyen‐The, C. , and Schmitt, P. (2004) Survival of Bacillus cereus spores and vegetative cells in acid media simulating human stomach. J Appl Microbiol 97: 214–219. [DOI] [PubMed] [Google Scholar]
- Clavel, T. , Carlin, F. , Dargaignaratz, C. , Lairon, D. , Nguyen‐The, C. , and Schmitt, P. (2007) Effects of porcine bile on survival of Bacillus cereus vegetative cells and Haemolysin BL enterotoxin production in reconstituted human small intestine media. J Appl Microbiol 103: 1568–1575. [DOI] [PubMed] [Google Scholar]
- Coil, D. , Jospin, G. , and Darling, A.E. (2015) A5‐miseq: an updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 31: 587–589. [DOI] [PubMed] [Google Scholar]
- Crovadore, J. , Calmin, G. , Tonacini, J. , Chablais, R. , Schnyder, B. , Messelhäußer, U. , et al. (2016) Whole‐genome sequences of seven strains of Bacillus cereus isolated from foodstuff or poisoning incidents. Genome Announc 4: 4–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cushnie, T.P.T. , and Lamb, A.J. (2005) Antimicrobial activity of flavonoids. Int J Antimicrob Agents 26: 343–356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danhorn, T. , and Fuqua, C. (2007) Biofilm formation by plant‐associated bacteria. Annu Rev Microbiol 61: 401–422. [DOI] [PubMed] [Google Scholar]
- Das, S. , Dash, H.R. , Mangwani, N. , Chakraborty, J. , and Kumari, S. (2014) Understanding molecular identification and polyphasic taxonomic approaches for genetic relatedness and phylogenetic relationships of microorganisms. J Microbiol Methods 103: 80–100. [DOI] [PubMed] [Google Scholar]
- Doan, H.K. , Antequera‐Gómez, M.L. , Parikh, A.N. , and Leveau, J.H.J. (2020a) Leaf surface topography contributes to the ability of Escherichia coli on leafy greens to resist removal by washing, escape disinfection with chlorine, and disperse through splash. Front Microbiol 11: 1485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doan, H.K. , Ngassam, V.N. , Gilmore, S.F. , Tecon, R. , Parikh, A.N. , and Leveau, J.H.J. (2020b) Topography‐driven shape, spread, and retention of leaf surface water impacts microbial dispersion and activity in the phyllosphere. Phytobiomes J 4: 268–280. [Google Scholar]
- Dressman, J.B. , Berardi, R.R. , Dermentzoglou, L.C. , Russell, T.L. , Schmaltz, S.P. , Barnett, J.L. , et al. (1990) Upper gastrointestinal (GI) pH in young, healthy men and women. Pharm Res An Off J Am Assoc Pharm Sci 7: 756–761. [DOI] [PubMed] [Google Scholar]
- Duport, C. , Jobin, M. , and Schmitt, P. (2016) Adaptation in Bacillus cereus: from stress to disease. Front Microbiol 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehling‐Schulz, M. , Fricker, M. , and Scherer, S. (2004) Bacillus cereus, the causative agent of an emetic type of food‐borne illness. Mol Nutr Food Res 48: 479–487. [DOI] [PubMed] [Google Scholar]
- Ehling‐Schulz, M. , Svensson, B. , Guinebretiere, M.‐H. , Lindbäck, T. , Andersson, M. , Schulz, A. , et al. (2005) Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 151: 183–197. [DOI] [PubMed] [Google Scholar]
- Elbing, K. , and Brent, R. (2002) Media preparation and bacteriological tools. Curr Protoc Mol Biol Chapter 1: 1.1.1‐1.1.7. [DOI] [PubMed] [Google Scholar]
- Elhariry, H.M. (2011) Attachment strength and biofilm forming ability of Bacillus cereus on green‐leafy vegetables: cabbage and lettuce. Food Microbiol. 28: 1266–1274. [DOI] [PubMed] [Google Scholar]
- Fagerlund, A. , Dubois, T. , Økstad, O. A. , Verplaetse, E. , Gilois, N. , Bennaceur, I. , et al. (2014) SinR controls enterotoxin expression in bacillus thuringiensis biofilms. PLoS One 9: e87532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finn, R.D. , Coggill, P. , Eberhardt, R.Y. , Eddy, S.R. , Mistry, J. , Mitchell, A.L. , et al. (2016) The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res 44: D279–D285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flemming, H.C. , and Wingender, J. (2010) The biofilm matrix. Nat Rev Microbiol 8: 623–633. [DOI] [PubMed] [Google Scholar]
- Franciosa, G. , Maugliani, A. , Scalfaro, C. , Floridi, F. , and Aureli, P. (2009) Expression of internalin a and biofilm formation among Listeria monocytogenes clinical isolates. Int J Immunopathol Pharmacol 22: 183–193. [DOI] [PubMed] [Google Scholar]
- Gao, T. , Foulston, L. , Chai, Y. , Wang, Q. , and Losick, R. (2015) Alternative modes of biofilm formation by plant‐associated Bacillus cereus . Microbiologyopen 4: 452–464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- García‐Gutiérrez, L. , Romero, D. , Zeriouh, H. , Cazorla, F.M. , Torés, J.A. , de Vicente, A. , and Pérez‐García, A. (2012) Isolation and selection of plant growth‐promoting rhizobacteria as inducers of systemic resistance in melon. Plant Soil 358: 201–212. [Google Scholar]
- Granum, P.E. , and Lund, T. (1997) Bacillus cereus and its food poisoning toxins. FEMS Microbiol Lett 157: 223–228. [DOI] [PubMed] [Google Scholar]
- Hamon, M.A. , and Lazazzera, B.A. (2001) The sporulation transcription factor Spo0A is required for biofilm development in Bacillus subtilis . Mol Microbiol 42: 1199–1209. [DOI] [PubMed] [Google Scholar]
- Heberle, H. , Meirelles, V.G. , da Silva, F.R. , Telles, G.P. , and Minghim, R. (2015) InteractiVenn: a web‐based tool for the analysis of sets through Venn diagrams. BMC Bioinform 16: 169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Illing, N. , and Errington, J. (1991) The spoIIIA operon of Bacillus subtilis defines a new temporal class of mother‐cell‐specific sporulation genes under the control of the σE form of RNA polymerase. Mol Microbiol 5: 1927–1940. [DOI] [PubMed] [Google Scholar]
- Jones, P. , Binns, D. , Chang, H.‐y. , Fraser, M. , Li, W. , McAnulla, C. , et al. (2014) InterProScan 5: genome‐scale protein function classification. Bioinformatics 30: 1236–1240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanehisa, M. , Sato, Y. , Kawashima, M. , Furumichi, M. , and Tanabe, M. (2016a) KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res 44: D457–D462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanehisa, M. , Sato, Y. , and Morishima, K. (2016b) BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J Mol Biol 428: 726–731. [DOI] [PubMed] [Google Scholar]
- Li, L. , Stoeckert, C.J. , and Roos, D.S. (2003) OrthoMCL: IDENTIFICATION of ortholog groups for eukaryotic genomes. Genome Res 13: 2178–2189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lund, T. , De Buyser, M.L. , and Granum, P.E. (2000) A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol Microbiol 38: 254–261. [DOI] [PubMed] [Google Scholar]
- Majed, R. , Faille, C. , Kallassy, M. , and Gohar, M. (2016) Bacillus cereus biofilms‐same, only different. Front Microbiol 7: 1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matilla‐Cuenca, L. , Gil, C. , Cuesta, S. , Rapún‐Araiz, B. , Žiemyt≐, M. , Mira, A. , et al. (2020) Antibiofilm activity of flavonoids on staphylococcal biofilms through targeting BAP amyloids. Sci Rep 10: 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKillip, J.L. (2000) Prevalence and expression of enterotoxins in Bacillus cereus and other Bacillus spp., a literature review. Antonie Van Leeuwenhoek 77: 393–399. [DOI] [PubMed] [Google Scholar]
- Nwodo, U.U. , Green, E. , and Okoh, A.I. (2012) Bacterial exopolysaccharides: functionality and prospects. Int J Mol Sci 13: 14002–14015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Toole, G. , Kaplan, H.B. , and Kolter, R. (2000) Biofilm formation as microbial development. Annu Rev Microbiol 54: 49–79. [DOI] [PubMed] [Google Scholar]
- Parashar, V. , Konkol, M.A. , Kearns, D.B. , and Neiditch, M.B. (2013) A plasmid‐encoded phosphatase regulates Bacillus subtilis biofilm architecture, sporulation, and genetic competence. J Bacteriol 195: 2437–2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel, C.N. , Wortham, B.W. , Lines, J.L. , Fetherston, J.D. , Perry, R.D. , and Oliveira, M.A. (2006) Polyamines are essential for the formation of plague biofilm. J Bacteriol 188: 2355–2363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramarao, N. , and Sanchis, V. (2013) The pore‐forming haemolysins of Bacillus cereus: a review. Toxins 5: 7492–7502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolny, I.S. , Minnaard, J. , Racedo, S.M. , and Pérez, P.F. (2014) Murine model of Bacillus cereus gastrointestinal infection. J Med Microbiol 63: 1741–1749. [DOI] [PubMed] [Google Scholar]
- Romero, D. , Aguilar, C. , Losick, R. , and Kolter, R. (2010) Amyloid fibers provide structural integrity to Bacillus subtilis biofilms. Proc Natl Acad Sci USA 107: 2230–2234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roux, A. , Beloin, C. , and Ghigo, J.M. (2005) Combined inactivation and expression strategy to study gene function under physiological conditions: application to identification of new Escherichia coli adhesins. J Bacteriol 187: 1001–1013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaeffer, P. , Millet, J. , and Aubert, J.P. (1965) Catabolic repression of bacterial sporulation. Proc Natl Acad Sci USA 54: 704–711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoeni, J.L. , and Wong, A.C. (2005) Bacillus cereus food poisoning and its toxins. J Food Prot 68: 636–648. [DOI] [PubMed] [Google Scholar]
- Silo‐Suh, L.A. , Lethbridge, B.J. , Raffel, S.J. , He, H. , Clardy, J. , and Handelsman, J. (1994) Biological activities of two fungistatic antibiotics produced by Bacillus cereus UW85. Appl Environ Microbiol 60: 2023–2030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, K. , and Youngman, P. (1993) Evidence that the spoIIM gene of Bacillus subtilis is transcribed by RNA polymerase associated with σE . J Bacteriol 175: 3618–3627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Söding, J. (2005) Protein homology detection by HMM‐HMM comparison. Bioinformatics 21: 951–960. [DOI] [PubMed] [Google Scholar]
- Soni, A. , Oey, I. , Silcock, P. , and Bremer, P. (2016) Bacillus spores in the food industry: a review on resistance and response to novel inactivation technologies. Compr Rev Food Sci Food Saf 15: 1139–1148. [DOI] [PubMed] [Google Scholar]
- Stenfors Arnesen, L.P. , Fagerlund, A. , and Granum, P.E. (2008) From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol Rev 32: 579–606. [DOI] [PubMed] [Google Scholar]
- Sturgill, G. , and Rather, P.N. (2004) Evidence that putrescine acts as an extracellular signal required for swarming in Proteus mirabilis . Mol Microbiol 51: 437–446. [DOI] [PubMed] [Google Scholar]
- Tettelin, H. , Masignani, V. , Cieslewicz, M.j. , Donati, C. , Medini, D. , Ward, N.l. , et al. (2005) Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: implications for the microbial “pan‐genome”. Proc Natl Acad Sci USA 102: 13950–13955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- The Gene Ontology Consortium . (2015) Gene ontology consortium: Going forward. Nucleic Acids Res. 43: D1049–D1056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tovar‐Rojo, F. , Chander, M. , Setlow, B. , and Setlow, P. (2002) The products of the spoVA operon are involved in dipicolinic acid uptake into developing spores of Bacillus subtilis . J Bacteriol 184: 584–587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran, S.L. , Guillemet, E. , Gohar, M. , Lereclus, D. , and Ramarao, N. (2010) CwpFM (EntFM) is a Bacillus cereus potential cell wall peptidase implicated in adhesion, biofilm formation, and virulence. J Bacteriol 192: 2638–2642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turnbull, P.C.B. (1976) Studies on the production of enterotoxins by Bacillus cereus . J Clin Pathol 29: 941–948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vidic, J. , Chaix, C. , Manzano, M. , and Heyndrickx, M. (2020) Food sensing: detection of Bacillus cereus Spores In Dairy Products. Biosensors 10: 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiencek, K.M. , Klapes, N.A. , and Foegeding, P.M. (1990) Hydrophobicity of Bacillus and Clostridium spores. Appl Environ Microbiol 56: 2600–2605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wijnands, L.M. , Pielaat, A. , Dufrenne, J.B. , Zwietering, M.H. , and Van Leusden, F.M. (2009) Modelling the number of viable vegetative cells of Bacillus cereus passing through the stomach. J Appl Microbiol 106: 258–267. [DOI] [PubMed] [Google Scholar]
- Willenbrock, H. , Hallin, P.F. , Wassenaar, T.M. , and Ussery, D.W. (2007) Characterization of probiotic Escherichia coli isolates with a novel pan‐genome microarray. Genome Biol 8: 267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmermann, L. , Stephens, A. , Nam, S.‐Z. , Rau, D. , Kübler, J. , Lozajic, M. , et al. (2018) A completely reimplemented MPI bioinformatics toolkit with a new hHpred Server at its Core. J Mol Biol 430: 2237–2243. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Persistence and sporulation of B. cereus strains on melon and cucumber leaves, cucumber fruit or endive. CFU counts per gram of leaf fruit 15 days after inoculation with B. cereus AH187, ATCC 14579, DSM 4282, DSM 4313, DSM 14729, UMAF8565, 6A27, DSM 8438 and DSM 2302. The black bar plots at the top show the total CFU count per gram of leaf or fruit. The grey bar graphs at the bottom correspond to the spore count per gram of leaf or fruit. The detection limit of the technique is 102 CFU/g of leaf. Error bars are the standard error of three or more replicates. Statistical analysis is the result of the comparison of all strains against AH187. *(P<0,05), **(P<0,01), ***(P<0,001) (Dunn’s test). No counts of the DSM 2302 strain appeared in any sporulation assay or on any surface, except endive. No significant counts of DSM 8438 strain appeared over all the surfaces (total CFUs and spores), except endive.
Fig. S2. Phylogenetic relation of B. cereus AH187, ATCC 14579, DSM 4282, DSM 4313, DSM 14729, UMAF8564, 6A27, DSM 8438 and DSM 2302 strains. Red points indicate the strains that were not able to persist on the melon leaf surface (DSM 8438 and DSM 2302). B. subtilis subsp. subtilis 168 was used as an out‐group. In the analysis, four concatenated genes (16S rRNA, gyrB, rpoD and rpoB) were handled to build a neighbour‐joining tree using the MUSCLE algorithm of MEGA7.
Fig. S3. Pangenome analysis of the strains AH187, ATCC 14579, DSM 4282, DSM 4313, DSM 14729, UMAF8564, 6A27, DSM 8438 and DSM 2302. A Schematic view of the pangenome analysis of the nine studied strains. The inner layer represents the pangenome (all the analysed genes). Each colour layer corresponds to the genome of one B. cereus strain. B Statistical estimation with Willenbock (blue) and Tettelin (red) (Tettelin et al., 2005; Willenbrock et al., 2007). The plot on the top points corresponds to pangenome size according to the number of genomes. In the plot on the bottom, the points correspond to the calculation of the core genome for each genome. C Schematic view of the genomic comparison between strains AH187 and DSM 2302. Genes present in strain AH187 are represented as brown lines throughout the genome, while genes absent in strain DSM 2302 are represented as red lines just above previous. The percentage of CG is represented in black, and the GC skew is represented in purple to locate the intergenic zones and the origins of replication of these strains, respectively.
Fig. S4. Comparison of biofilm formation capacity of AH187 and DSM 2302. 72 h in vitro biofilm and swarming assays of AH187 and DSM 2302 strains on TY 1.5% agar supplemented with 20 µg/ml and 10 µg/ml Congo Red and Coomassie Brilliant Blue dyes (A1), TY 0.7% agar (A2) and TY broth (A3). A4 shows bacterial adhesion to an abiotic surface after staining with crystal violet. B sipW‐calY locus implicated in extracellular matrix synthesis of AH187 and DSM 2302 compared to the B. cereus type strain (ATCC 14579). C Region homologous to the eps1 region of ATCC 14579 involved in exopolysaccharide biosynthesis of AH187 and DSM 2302 strains. Middle grey genes correspond to different genes compared to each other. D Region homologous to the eps2 region of ATCC 14579 involved in exopolysaccharide biosynthesis found in AH187 and DSM 2302 strains. Grey genes correspond to different genes located in the eps2 genome region.
Fig. S5. Metabolomic profile of melon and endive leaf. Clustering result shown as a heatmap of three replicates of leaves of endive (orange) and melon leaves (green). In the heatmap, red indicates a higher concentration of a metabolite; in contrast, blue indicates a lower concentration of the metabolite.
Fig. S6. Persistence of AH187 mutants on melon leaves. CFU (black bar plot at the left) and spore (grey bar plot at the right) counts of AH187 mutants of the genes listed in Table S5 on melon leaves 15 days after bacteria inoculation. The detection limit of the technique is 102 CFU/g of leaf. Error bars are the standard error of three or more replicates. Statistical analysis is the result of the comparison of all the mutants against AH187 wild‐type. *(P<0.05), **(P<0.01), ***(P<0.001) (Dunn’s test). sigE mutant conts felt below the detection limit.
Table S1. Sequenced B. cereus strains deposited in Genbank. Sequenced B. cereus strains deposited in Genbank.
Table S2. Primers used to construct mutants of B. cereus AH187.
Table S3. Pangenome analysis of B. cereus strains. Core indicates the proteins shared among all the studied strains, accessory shows the proteins shared among all studied strains except one or less, and unique enumerates the proteins only present in one of the studied strains and not in the others. Differences in core proteins correspond to paralogous genes.
Table S4. GO and KO terms, obtained from the pangenome analysis that are present in B. cereus DSM 2302 strains and absent in AH187 strain. GO and terms associated with DSM 2302 strain with their corresponding function and DSM 2302 genes related with each GO/KO term.
Table S5. Genes absent in B. cereus DSM 2302 and DSM 8438. Genomic comparison among persister and non persister strains reveal genes non present in DSM 8438 and DSM 2302 strains.