

Abstract
The use of bacteriocins holds great promise in different areas such as health, food, nutrition, veterinary, nanotechnology, among others. Many research groups worldwide continue to advance the knowledge to unravel a novel range of therapeutic agents and food preservatives. This review addresses the advances of bacteriocins and their producer organisms as biocontrol agents for applications in the medical industry and agriculture. Furthermore, the bacteriocin mechanism of action and structural characteristics will be reviewed. Finally, the potential role of bacteriocins to modulate the signaling in host-associated microbial communities will be discussed.
Keywords: bacteriocin, bio-preservatives, agriculture, biomedical, microbial communities
Introduction
Bacteria living in microbial communities use several functions and strategies to survive or coexist with other microorganisms, competing to obtain nutrients and colonize space in their habitat (Hibbing et al. 2010). One of the strategies used by bacteria to guarantee their growth in communities is antagonism, which effectively limits the growth of other microorganisms (Russel et al. 2017). To accomplish antagonism, bacteria must produce inhibitory substances such as antibiotics, organic acids, siderophores, volatile organic compounds, antifungals, and bacteriocins (Riley 2009). In addition to inhibiting the growth of other microorganisms, bacteriocins have different traits that make them attractive for biotechnological applications. For example, while resistance against nisin exists, in general, the bacteriocin mechanism of action less often induces resistance as it happens with conventional antibiotics (Behrens et al. 2017). Furthermore, some bacteriocins are compounds produced by the natural host-associated microbiome; therefore, they are harmless to the host. Bacteriocins also show selective cytotoxicity toward cancer cells compared to normal cells (Kaur and Kaur 2015).
Classification, mechanism of action, and structural characteristics
Bacteriocins are antimicrobial peptides synthesized by the ribosome representing the most abundant and diverse group of bacterial defense systems (Silva et al. 2018). Bacteriocins were considered to have a narrow antimicrobial spectrum that could only inhibit bacterial strains closely related to produced bacteria; however, several studies have shown that there are bacteriocins able to kill different genera of bacteria and even certain yeasts, parasites, and cancer cells (Kaur and Kaur 2015; Baindara et al. 2018).
The success of bacteriocins in eliminating multidrug resistant pathogens (MDR) has led to medical applications to treat bacterial infections. In vivo tests have demonstrated the effectiveness of bacteriocins to treat infections in animal models, too (McCaughey et al. 2016; Van Staden et al. 2016). Lactic acid bacteria (LAB) produce bacteriocins, being nisin from Lactococcus lactis, the most well-known example (Silva et al. 2018). Nisin was approved for use as a food preservative for preventing the growth of Listeria monocytogenes and other Gram-positive pathogens (Price et al. 2018). The Bacillus genus also produces bacteriocins with attractive characteristics (Salazar-Marroquín et al. 2016), including subtilin (produced by Bacillus subtilis) and coagulin (produced by Bacillus coagulans). Bacillus thuringiensis produces bacteriocins with broad-spectrum activity, inhibiting various pathogens such as L. monocytogenes, Staphylococcus aureus, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Vibrio cholerae, in addition to the Aspergillus fungus (Salazar-Marroquín et al. 2016).
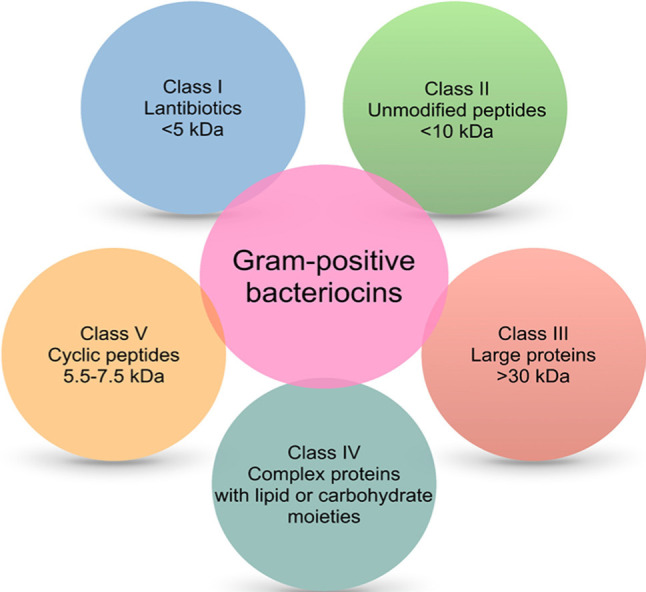
Bacteriocins of Gram-positive bacteria are cationic and amphiphilic molecules whose mass varies from < 5 to more than 30 kDa (Balciunas et al. 2013) (Fig. 1). Many classifications of bacteriocins are available, but their diverse chemical structures and inhibitory activities make their classification into a specific group quite difficult. Class I bacteriocins, also known as lantibiotics, contain in their primary structure uncommon amino acids like lanthionine, β-methyl lanthionine, and dehydroalanine. These unique amino acids formed by post-translational modifications can provide antimicrobial activity and peptide stability. For example, they can create covalent bridges that result in internal rings that give stability to the peptide structure. In addition, internal rings contribute to the formation of a secondary structure in water that favors antimicrobial activity (Almeida and Pokorni 2012). Around 30% of lantibiotics already identified have been purified from lactic acid bacteria, including the well-known nisin, mersacidin, and lacticin 3147 (Stoyanova et al. 2012). The class II bacteriocins are membrane-active and heat-stable peptides known as non-lantibiotics or pediocin-like antibiotics (Balandin et al. 2019). They do not harbor modified amino acids, and their molecular weights are lower than 10 kDa. Prototype bacteriocins of this group are pediocin PA-1, pentocin 31–1, enterocin P, sakacin G, enterocin A, two-peptide components (enterocin DD14, plantaracin E/F), sec-dependent secreted (acidocin B), and other not yet subclassified (bactofencin A peptides) (Liu et al. 2008; Balandin et al. 2019; Ladjouzi et al. 2020). The class III bacteriocins are large (> 30 kDa) heat-labile peptides composed of an N-terminal endopeptidase domain and a C-terminal substrate recognition domain. Bacteriocins of this group can lyse the cell wall of sensitive bacteria, although there are non-lytic bacteriocins in this group too, like helveticin J. Some examples of Class III bacteriocins are helveticin M, zoocin A and enterolysin A (bacteriolysins), and millericin B (murein hydrolase) (Alvarez-Sieiro et al. 2016; Sun et al. 2018). Class IV are complex peptide structures associated with lipid and carbohydrate moiety forming glycoproteins and lipoproteins. These structural characteristics make them sensitive to the action of glycolytic or lipolytic enzymes. Lactocin 27 and leuconocin S are prototype bacteriocins of this group and are recognized to disrupt bacterial cell membranes (Simons et al. 2020). Class V includes cyclic peptide structures like enterocin AS-48, pumilarin, lactocyclicin Q, and plantaricyclin A (Perez et al. 2018; Sánchez-Hidalgo et al. 2011). The circular nature of their structures provides Class V with superior stability against several stresses compared to most linear bacteriocins. Biosynthesis of circular bacteriocins involves cleavage of the leader peptide, circularization, and export to the extracellular space.
Fig. 1.
Structure-based classification of Gram-positive bacteriocins.
Gram-negative bacteria produce both high molecular weight (> 30 kDa) and low molecular weight (< 10 kDa) bacteriocins (Rebuffat 2016). The first bacteriocin identified from a Gram-negative bacterium was colicin, produced by Escherichia coli (Riley 2009). Bacteriocins of Gram-negative bacteria are classified into two main groups, colicins, and microcins (Fig. 2). Genes encoding colicins are found on plasmids whose products vary between 20 and 80 kDa. Colicins from E. coli inhibits closely related strains of the genus Salmonella and other E. coli strains. Colicins are organized in three different domains: the translocation domain (T) N-terminally located, the receptor binding (R) located in the central region, and the cytotoxic domain (C) located at C-terminus (Helbig and Braun 2011). Microcins are pH and heat-stable antimicrobial peptides ribosomally synthesized, hydrophobic, and low molecular weight. In some cases, microcins require post-translational modifications to be active, and they do not require a lysis process to be secreted (Baquero et al. 2019). Microcin production has been reported in several Enterobacteriaceae and some cyanobacteria (Rebuffat 2016; Parnasa et al. 2019).
Fig. 2.

Structural-based classification of Gram-negative bacteriocins.
Microcin mJ25 produced by E. coli was initially described as a circular peptide; now it is known that there is no union between the terminal residues, but a union through the lactamic link between the amino group (Gly1) and the carboxyl group (Glu8). These structures are known as “lasso-peptides” and are also described in organisms of the genus Streptomyces (Hegemann et al. 2015). Other types of high molecular weight bacteriocins of Gram-negative bacteria are pyocins (type R, F, and S), tailocins, and lectin-like bacteriocins. Genes encoding for pyocins are located on the bacterial chromosome, and their expression is induced by agents that damage DNA by activating the SOS response. R-type and F-type pyocins are non-flexible and flexible phage tail-like bacteriocins, respectively. The S-type pyocin is like the colicins and is formed by two proteins (a big one and a small one) that remain associated even during its purification process. The large protein is responsible for the antimicrobial activity, and the small one has an immune function for the producing bacteria (Michel-Briand and Baysse 2002; Atanaskovic and Kleanthous 2019; Oluyombo et al. 2019).
Tailocins are bacteriocins like phage tails and display a rigid or flexible structure, similar to R-type and F-type pyocins. Tailocins with contractile and flexible tail morphologies are designated as myotailocins and siphotailocins, respectively (Yao et al. 2017). These bacteriocins have been described in plant-associated Pseudomonas and Burkholderia strains, although similar bacteriocins are also produced by Clostridium difficile, Serratia plymithicum, and Serratia proteamaculans (Gebhart et al. 2015; Ghequire and De Mot 2015; Hurst et al. 2018).
Lectin-like bacteriocins (LlpAs) represent another type of antimicrobial protein secreted by members of the genus Pseudomonas. LlpAs are ~30 kDa proteins that resemble monocot mannose-binding lectins (MMBL) consisting of two B-lectin domains followed by a short carboxy-terminal extension and do not contain an immunity protein. They also include a preserved consensus sequence QxDxNxVx necessary for the activity of the bacteriocin. The best examples of LlpAs include LlpABW11M1 of Pseudomonas mosselii, LlpA1Pf-5 of Pseudomonas protegens Pf-5, and pyocin L1 of P. aeruginosa (Ghequire et al. 2018a). The production of LlpAs has also been reported in Burkholderia cepacia strains.
Bacteriocins exert several mechanisms of action towards Gram-positive and Gram-negative bacteria (Fig. 3). Class I bacteriocins produced by Gram-positive bacteria permeabilize bacterial membranes through pore-formation, leading to ion leakage and cell death. These include bacteriocins produced by Bacillus, Lactococcus, and Pediococcus genera. They cause pore-formation by recognizing lipid II or the mannose phosphotransferase system (Paiva et al. 2011). Class I bacteriocins of Gram-positive bacteria also inhibit cell wall synthesis (Abriouel et al. 2011; Sun et al. 2018) (Fig. 3a). Class II bacteriocins make pores as described by the barrel stave or carpet model. Some class III bacteriocins produced by Bacillus inhibit the activity of the phospholipase A2, responsible for membrane repair (Abriouel et al. 2011). Class III bacteriocins, like lysostaphin, act directly on the cell wall inhibiting peptidoglycan synthesis without permeabilizing the membrane (Mitkowski et al. 2019). The mannose phosphotransferase system is involved in recognition of some Gram-positive bacteriocins, such as lactococcin A and pediocin, leading to pore-formation and membrane permeabilization (Zhou et al. 2016).
Fig. 3.

Bacteriocin mechanism of action on a) Gram-positive and b) Gram-negative bacteria.
The mechanism of action of Gram-negative bacteriocins, such as colicins, is through recognizing cell surface receptors of a target cell, through the Tol or TonB machinery, as shown in Fig. 3b. Colicins C domain (cytotoxicity domain) is responsible for eliminating other microorganisms through various mechanisms such as membrane permeabilization, nuclease activity, and inhibition of peptidoglycan or lipopolysaccharide O-antigen synthesis (Budič et al. 2011). Salmonella colicins (salmocins) display three mechanisms of action: SalE1a and SalE1b cause pore-formation in the membrane, SalE2 and SalE7 have DNase activity, and SalE3 have RNase activity (Schneider et al. 2018). Microcins are also membrane-pore formers, have DNase or RNase activity, and may inhibit protein synthesis (Yang et al. 2014).
The genus Pseudomonas produces high molecular weight bacteriocins such as R, F, and S type pyocins (Oluyombo et al. 2018). Besides the type B microcins (Ghequire et al. 2018a), tailocins (Ghequire and De Mot 2015), and LlpAs (Ghequire et al. 2018b). Pyocins and tailocins are characterized by having a complex structure that resembles phage tails (Ghequire and De Mot 2015; Patz et al. 2019), and the mechanism of action is based on the recognition of specific receptors on the cell surface causing pore formation, nonspecific degradation of nucleic acids or lipid II-degradation (Ghequire and De Mot 2018; Patz et al. 2019) (Fig. 3b).
Pyocins have a limited antimicrobial spectrum, mainly inhibiting competitors highly related to the producer strain (Redero et al. 2018). However, some R-type pyocins can inhibit other species such as Campylobacter sp., Neisseria gonorrhea, Neisseria meningitides, and Haemophilus ducreyi (Naz et al. 2015). Since the mechanism of action of pyocins depends on a cellular receptor, its use has been proposed to replace broad-spectrum antibiotics, to reduce the damage that antibiotics usually cause to the human microbiome (McCaughey et al. 2014).
LlpAs have a selective mechanism of action, different from other bacteriocins produced by Pseudomonas species. Probably because their structure does not consist of the classic three-domain model present in bacteriocins of similar size (T, R, and C). Instead, they contain two monocotyledonous mannose-binding lectin (MMBL) domains associate with the recognition of BamA (Ghequire et al. 2018a). This protein of the outer membrane of Gram-negative bacteria facilitates the insertion of other proteins into the cell membrane (Noinaj et al. 2014). Although the mechanism of action of LlpAs remains unknown, a “killing upon contact” mechanism has been suggested (Ghequire et al. 2018a).
Bacteriocins and natural DNA transformation
Bacteria can take up exogenous DNA and incorporate it into their genome through a process termed competence. Competent bacteria can use absorbed DNA as a source of nutrients, DNA reparation, or recombination with the genome. Natural DNA transformation happens when absorbed DNA is integrated into the genome (Veening and Blokesh 2017). This process is considered the primary mode of horizontal gene transfer (HGT) in bacteria, along with conjugation (direct cell to cell transfer of DNA via a specialized conjugal pilus) and phage transduction (DNA transfer mediated by viruses). Naturally competent bacteria couple the DNA-uptake process with other physiological responses, such as growth arrest and synthesis of antimicrobial polypeptides (bacteriocins) (Mignolet et al. 2018). Bacteria secrete bacteriocins upon entry into the competence state to kill surrounding competitors.
The competence pathway in Streptococcus pneumoniae is regulated by a secreted peptide pheromone, the competence-stimulating peptide (CSP). The precursor peptide of CSP, ComC, is processed by an ABC transporter/protease, ComAB, immediately after the double-glycine motif to yield the active CSP (Shanker and Federle 2017). Extracellular CSP activates the ComCDE two-component signal-transduction pathway, which turns on the sigma factor gene sigX/comX, to activate the expression of over 100 genes upon entering the competent state (reviewed by Shanker and Federle 2017). At least six CSP-responsive genes are involved in fratricide (killing/lytic factors directed against non-competent siblings). Among them, cibABC encodes a two-peptide bacteriocin responsible for lysis of cells lacking the corresponding immunity factor, CibC. The cbpD gene encodes a murein hydrolase containing a cytosine, histidine-dependent amidohydrolase peptidase. lytA encodes an effector of autolysis in S. pneumoniae. Interestingly, this predation mechanism appears to be restricted to isogenic or closely related strains, suggesting that competent cells target corresponding cells to acquire homologous DNA sequences to maintain genome integrity or acquire new gene alleles from siblings. This ability to tackle closely related strains would be discussed in the section “Bacteriocins as modulators of gastrointestinal microbiota and population diversity”. Streptococcus salivarius, on the other hand, modules competence and bacteriocin production through the ComRS complex, which serves as the connector that directly regulates both comX and bacteriocin genes (Mignolet et al. 2018). S. salivarius bacteriocins have a broad spectrum of bacterial prey including the closely related Streptococcus vestibularis, more distant streptococci (Streptococcus mutans and Streptococcus pyogenes), and opportunistic pathogens such as Enterococcus faecalis, L. monocytogenes, and S. aureus (Mignolet et al. 2018).
Bacteriocins as food antimicrobial and anticancer agents
Bacteriocin applications have been focused primarily on food preservation, either alone or in combination with other compounds. The long shelf life of food products relies on adding chemicals, sugars, salts, and other preservatives allowed by the regulation. The addition of these substances reduces water activity, inhibiting the growth of undesirable pathogenic microorganisms that can spoil food. However, the addition of these chemicals benefits the industry but not the consumer since the continuous consumption of chemical preservatives through packaged foods can affect consumers’ health. There is an association of these additives with chronic degenerative diseases, and the intake of these additives can prompt the development of some types of cancer (Monteiro et al. 2010; Moubarac et al. 2013). A more friendly strategy to preserve food products is the use of bacteriocins beneficial for both the food industry and consumers, helping to reduce the use of chemical preservatives in food (Sarika et al. 2019). The growth of pathogens in food can be controlled by the inoculation of bacteriocin-producing lactic acid bacteria or by the addition of purified bacteriocins (Silva et al. 2018). Bacteriocins have also been added to the coating of food packaging to reduce food spoilage (Salgado et al. 2015; Castellano et al. 2017).
The use of bacteriocins as food preservatives does not affect the organoleptic properties of foods. There are safe bacteriocins for human consumption, such as Enterocin AS-48 (Sánchez-Hidalgo et al. 2011), lacticin 3147 (Mills et al. 2017), and salmocins (Schneider et al. 2018) but only nisin (NisaplinTM, BiosafeTM), pediocin PA-1 (MicrogardTM, Alta 2431), sakacin (BactofermTM B-2, BactofermTM B-FM) and leucocin A (BactofermTM B-SF-43) are commercially used to improve shelf-life of food (Vijay Simha et al. 2012; Daba and Elkhateeb 2020).
The Food and Agriculture Organization (FAO) support the use of probiotics in food systems, since probiotics offer health benefits, especially for the gastrointestinal tract. Probiotics play an important role in modifying some metabolic pathways that, in turn, regulate cell proliferation, apoptosis, differentiation, angiogenesis, inflammation, and metastasis, which are relevant aspects to prevent the development of cancer (Bermudez-Brito et al. 2012).
Bacteriocins have shown cytotoxic activity against cancer cells, and therefore they could be considered tools to develop new anticancer drugs (Baindara et al. 2018). The charge of normal cell membranes is neutral, while cancer cells have a negative charge due to the high content of anionic phosphatidylserine, o-glycosylated mucins, sialylated gangliosides, and heparin sulfates. Bacteriocins, being cationic peptides, can preferentially bind to the negatively charged membrane of cancer cells compared to normal cells. Some bacteriocins with anticancer activities are colicins, which have shown cytotoxic activity against various human tumor cell lines such as breast cancer, colon cancer, and bone cancer (Kaur and Kaur 2015). Some examples of the potential applications of bacteriocins are shown in Table I.
Table I.
Bacteriocins with potential application as therapeutic and food preservatives.
| Bacteriocin | Producer bacteria | Target microorganism | Use | Reference |
|---|---|---|---|---|
| 1. Food preservation | ||||
| AMA-K, Leucocin K7 | L. plantarum AMA-K |
Enterococcus spp., E. coli, K. pneumoniae, Listeria spp. |
Amasi, fermented milk product | (Todorov 2008) |
| Aureocin A70 | S. aureus A70 | L. monocytogenes | Dairy product | (Carlin Fagundes et al. 2016) |
| Bacteriocin 32Y | L. curvatus | L. monocytogenes | Pork and beef | (Gálvez et al. 2007) |
| Bacteriocin GP1 | L. rhamnosus GP1 |
Staphylococcus sp., Aeromonas sp., Lactobacillus sp., Pseudomonas sp., Vibrio sp. |
Fish | (Sarika et al. 2019) |
| Bovicin HC5 + Nisin | Streptococcus bovis HC5 | L. monocytogenes, S. aureus | Fresh cheese | (Pimentel-Filho et al. 2014) |
| Divergicin M35 | Carnobacterium divergens M35 | L. monocytogenes | Smoked fish | (Benabbou et al. 2020) |
| Enterocin | E. faecium FAIR-E 198 | Listeria spp. | Feta cheese | (Sarantinopoulos et al. 2002) |
| Enterocin 416K1 | E. casseliflavus IM 416K1 | L. monocytogenes NCTC 10888 | Cottage cheese | (Iseppi et al. 2008) |
| Enterocin AS-48 | Enterococcus sp. | L. monocytogenes, B. cereus | Cheese, vegetable, purees, and soups | (Gálvez et al. 2007) |
| H1, H2, H3, H4 | Bacillus sp. | V. alginolyticus, Aeromonas hydrophilla, P. stutzeri | Antimicrobial used in fish | (Feliatra et al. 2018) |
| Lacticin 3147 | L. lactis | L. monocytogenes | Matured and cottage cheese | (Mills et al. 2017) |
| Lacticin NK24 | L. lactis |
Leuconostoc mesenteroides KCCM 11324 |
Seafood | (Lee and Paik 2001) |
| Leucocin K7 | L. mesenteroides K7 | L. monocytogenes | Dairy product | (Shi et al. 2016) |
| Mecedocin |
S.
macedonicus ACA-DC 198 |
C. tyrobutyricum LMG 1285T | Kasseri cheese | (Anastasiou et al. 2009) |
| NE |
L.
gasseri K7 (Rifr), L. gasseri LF221(Rifr) |
C. tyrobutyricum | Semi-mature cheese | (Bogovič Matijašić et al. 2007) |
| Nisin |
Lactococcus spp., Streptococcus spp. |
L. monocytogenes, Clostridium botulinum, S. mutans, L. innocua, S. aureus, S. pneumoniae, B. cereus |
Dairy products, meat, seafood | (Juturu and Wu 2018) |
| Pediocin PA1 | P. acidilactici | L. monocytogenes | Dairy products, meat | (Liu et al. 2008) |
| Plant-made salmocins | Salmonella spp. | S. enterica | Red meat | (Schneider et al. 2018) |
| Plant-made colicins (GRN 676, GRN 593) | E. coli |
E. coli, P. aeruginosa, Salmonella spp. |
Meat, fruits, or vegetables | (Hahn-Löbmann et al. 2019) |
| Psicolin 126, carnocyclin A | C. maltoaromaticum | L. monocytogenes | Ready-to-eat meat products | (Liu et al. 2014) |
| Reuterin | L. reuteri |
E.
coli, S. aureus, Candida albicans |
Food preservation | (Helal et al. 2016) |
| Sakacin P | L. sakei | L. monocytogenes | Beef and Salmon | (Teneva-Angelova et al. 2018) |
| Thuricin BtCspB | B. thuringiensis | B. cereus | Food preservation and disease associate to B. cereus | (Huang et al. 2016) |
| 2. Bacterial infections | ||||
| ABP118 | L. salivarius subsp. salivarius UCC118 | Bacteroides | Antimicrobial agent | (Riboulet-Bisson et al. 2012) |
| Colicins Js and Z | E. coli | Enteroinvasive, E. coli (EIEC) and Shigella | Gastrointestinal infections | Bosák et al. 2021 |
| Divercin V41 | C. divergens | L. monocytogenes | Antimicrobial agent | (Rihakova et al. 2010) |
| Duramycin | Streptomyces cinnamoneus | B. subtilis | Antimicrobial, anti-viral, immunomodulation, ion channel modulation, treatment of atherosclerosis and cystic fibrosis | (Huo et al. 2017) |
| Enterocin CRL35 | E. mundtii | L. monocytogenes | Gastrointestinal infections | (Salvucci et al. 2012) |
| Epidermin and mersacidin-like peptides | S. epidermidis | P. acnes | Acne, folliculitis. | (Gillor et al. 2008) |
| Gallidermin/epidermin | S. gallinarum | S. epidermidis, S. aureus | Skin infections or associated with implants and prostheses | (Bengtsson et al. 2018; Bonelli et al. 2006) |
| Gassericin E | L. gasseri EV1461 | Pathogens associated with vaginosis | Vaginal infections | (Maldonado-Barragán et al. 2016) |
| Haemocin type B |
Haemophilus haemolyticus |
H. influenza | Respiratory infections | (Latham et al. 2017) |
| Lactocin 160 | L. rhamnosus | G. vaginalis | Urogenital tract infections, bacterial vaginosis | (Turovskiy et al. 2009) |
| Laterosporulin10 | Brevibacillus sp. strain SKDU10 |
S. aureus, Mycobacterium tuberculosis (Mtb H37Rv), M. smegmatis MC2 155 |
Human microbial pathogens | (Baindara et al. 2016) |
| Mersacidin | B. amyloliquefaciens | Methicillin-resistant S. aureus (MRSA) |
Skin infection | (Kruszewska et al. 2004) |
| Microcin J25 (lasso-peptide) | E. coli | S. enterica, E. coli, S. flexnerii | Gastrointestinal infections | (Dobson et al. 2012) |
| Nisin A, Nisin Z, Nisaplin | L. lactis |
S.
mutans, S.
aureus, E.
faecalis, S. mastitis, C. albicans |
Gastrointestinal, respiratory, and skin infections, oral health | (Shin et al. 2016) |
| Oralpeace TM (encapsulated nisin) | L. lactis | S. mutans, P. gingivalis | Dental caries, gingivitis | (Perez et al. 2014) |
| Piscicolin 126 | Carnobacterium spp. | Listeria spp. | Antimicrobial agent | (Miller and McMullen 2014) |
| Plantaricin 423 | L. plantarum | Listeria spp. | Antimicrobial agent | (Guralp et al. 2013) |
| PLNC8 αβ | L. plantarum |
Staphylococcus sp., Porphyromonas gingivalis |
Antimicrobial agent | (Bengtsson et al. 2020) |
| R-pyocins | P. aeruginosa | P. aeruginosa | Antimicrobial agent | (Redero et al. 2018) |
| TOMM Streptolysin S (SLS) | S. pyogenes |
Clostridium sp., Listeria sp. |
Hemolytic and cytotoxic activity against macrophages and neutrophils | (Molloy et al. 2015) |
| 3. Anticancer drugs | ||||
| Cancer cell lines | ||||
| Azurin | P. aeruginosa | MCF-7, UISO-Mel-2, osteosarcoma (U2OS) | (Nguyen and Nguyen 2016) | |
| Bovicin HC5 | S. bovis HC5 | MCF-7, HepG2 | (Rodrigues et al. 2019) | |
| Colicin E3 | E. coli | P388, HeLa, HS913T | (Kohoutova et al. 2014 | |
| Duramycin | S. cinnamoneus | AsPC-1, Caco-2, Colo320, CT116, JJN3, Lovo, MCF-7, MDA-B-231, MIA PaCa-2 | (Rodrigues et al. 2019) | |
| Enterocin LNS18 | E. thailandicus | HepG2 | (Al-Madboly et al. 2020) | |
| Laterosporulin LS10 | Brevibacillus laterosporus SKDU10 | HeLa, MCF-7, H1299, HEK293T, HT1080 | (Baindara et al. 2016) | |
| sporus SKDU10 | ||||
| M2163, M2386 | L. casei ATCC 334 | SW480 | (Rodrigues et al. 2019) | |
| Microcin E492 | K. pneumoniae | HeLa, Burkitt lymphoma variant (RJ2.25) | (Kaur and Kaur 2015) | |
| Nisin A | L. lactis | Head and neck squamous cell carcinoma (HNSCC) | (Shin et al. 2016) | |
| Pediocin K2a2-3 | P. acidilactici K2a2-3 | HT2a, HeLa | (Villarante et al. 2011) | |
| Pediocin CP2 | P. acidilactici CP2 MTCC501 | HeLa, MCF-7, HepG2, murine myeloma (Sp2/0-Ag 14) | (Kumar et al. 2012) | |
| Pep27anal2 | S. pneumoniae | Jurkat, HL-60, AML-2, MCF-7, SNU-601 | (Rodrigues et al. 2019) | |
| Plantaricin A | L. plantarum C11 | GH4, Reh, Jurkat, PC12, N2A | (Sand et al. 2013) | |
| Plantaricin P1053 | L. plantarum PBS067 | E705 | (De Giani et al. 2019) | |
| Pyocin S2 | P. aeruginosa 42A | HepG2, Im9, murine tumor (mKS-A TU-7), human fetal foreskin fibroblast (HFFF) | (Abdi-Ali et al. 2004) | |
| Sungsanpin | Streptomyces spp. | A549 | (Um et al. 2013) | |
| Smegmatocin | M. smegmatis 14468 | HeLa, AS-II, HGC-27, mKS-A TU-7 | (Kaur and Kaur 2015) | |
NE – non specified
The potential therapeutic uses of bacteriocins produced by lactic acid bacteria have increased over time. López-Cuellar et al. (2016) found that 37% of the investigations on bacteriocins were focused on medical applications including cancer, systemic infections, stomatology, skincare, and contraceptives. 29% of studies focused on food preservation, 25% on bio-nanomaterials, and 9% within veterinary. The number of patents on bacteriocins has also increased. From 2004 to 2015, 245 bacteriocin patents were issued, 31% related to the biomedical field, 29% to food preservation, 5% to veterinary medicine, 13% to production and purification process, and 16% to molecular modifications in producer strains. The smallest proportion concerns bio-nanomaterials and industrial applications.
Bacteriocins in agriculture
The indiscriminate use of agrochemicals has caused severe damage to human health and the environment. This problem aims to find alternatives to fight pests and diseases in a more environmentally friendly way. Bacteria that produce inhibitory substances have been used as inoculants to indirectly stimulate the growth of crops, fighting the phytopathogens. Plant growth-promoting rhizobacteria (PGPR) are generally marketed in the form of mono or multi-inoculants that include bacteria such as Streptomyces venezuelae, Gluconacetobacter diazotrophicus, Burkholderia sp., Azospirillum brasilense, P. protegens, Pseudomonas putida, among others. Most of these formulations have been traded to promote plant growth and not fight plant pathogens (Cesa-Luna et al. 2020). Therefore, little efforts have been focused on applying of bacteriocins for plant disease biocontrol, and hence their production by PGPR is poorly understood.
Some examples of bacteriocins applied to agriculture are agrocin 84 and thuricin 17. Agrocin 84 is produced by Agrobacterium radiobacter K84 and is useful to kill Agrobacterium tumefaciens, the causal agent of crown gall disease in plants (Kim et al. 2006). Thuricin 17 is produced by B. thuringiensis NEB17, this bacteriocin is a plant biostimulant with no harmful effects on nodulating rhizobia or other PGPR (Nazari and Smith 2020). Pseudomonas syringae pv. ciccaronei strain NCPPB2355 produces an inhibitory bacteriocin against P. syringae subsp. savastanoi, the causal agent of olive knot disease. Other important bacteriocins are those produced by the genus of Pseudomonas and Bacillus (Table II). These bacteriocins inhibit one of the primary phytopathogenic fungi, Fusarium, which can infect different types of plants, including celery, onion, cabbage, banana, cucumber, tomato, eggplant, cantaloupe, watermelon, spinach, among others. Direct application of bacteriocin induces a resistance mechanism in plants against pathogens and abiotic stresses. Application of thuricin 17 on plants enhanced production of phenolics, phenylalanine ammonia-lyase activity, and antioxidant defense (Nazari and Smith 2020).
Table II.
Biocontrol potential of bacteriocin-producing microorganisms in agriculture.
| Bacteriocin | Producer bacterium | Phytopathogen | Reference |
|---|---|---|---|
| Amylocyclin | B. amyloliquefaciens | Ralstonia solanacearum and X. campestris | (Scholz et al. 2014) |
| Bacteriocin 32Y | P. aeruginosa RsB29 | Fusarium sp. | (Sindhu et al. 2016) |
| Carocin D | P. carotovorum subsp. carotovorum | P. carotovorum subsp. Carotovorum | (Grinter et al. 2012; Roh et al. 2010) |
| Enterocin UNAD 046 | E. faecalis | B. theobromae, A. niger, P. expansum, P. ultimum. | (David and Onifade, 2018) |
| Fluoricin BC8 | P. fluorescens BC8 | P. solanacearum | (Sindhu et al. 2016) |
| Gluconacin | G. diazotrophicus PAL5 | X. albilineans and X. vasicola pv. vasculorum. | (Oliveira et al. 2018) |
| LlpA | P. putida BW11M1 | P. syringae | (Parret et al. 2005) |
| Morricin 269, Kurstacin 287, Kenyacin 404, Entomocin 420, Tolworthcin 524 | B. thurigiensis | Trichoderma spp., A. nodulans, F. graminis, F. oxysporum, Rhizopus sp., Mucor rouxii | (De La Fuente-Salcido et al. 2008; Salazar-Marroquín et al. 2016) |
| NE | P. syringae pv. ciccaronei | P. syringae subsp. savastanoi | (Lavermicocca et al. 2002) |
| BLIS RC-2 | B. amyloliquefaciens RC-2 | R. necatrix, P. oryzae, A. tumefaciens, Xanthomonas campestris pv. campestris, C. dematium | (Abriouel et al. 2011) |
| NE | B. gladioli | Tatumella ptyseos | (Marín-Cevada et al. 2012) |
| BL8 | B. thuringiensis subsp. tochigiensis HD868 | A. niger, A. fumigatus, A. flavus, Cryphonectria parasitica, F. oxysporum, Penicillium digitatum. | (Subramanian and Smith 2015) |
| Plantazolicin | B. velezensis FZB42 (B. amyloliquefaciens subsp. plantarum) | B. anthracis and nematodes. | (Chowdhury et al. 2015) |
| Putidacin L1 | P. protegens, P. putida | P. syringae | (Rooney et al. 2020) |
| Rhizobiocin | Rhizobium spp. | P. savastanoi | (Kaur Maan and Garcha 2018) |
| SF4c tailocins | P. fluorescens SF4c | X. vesicatoria | (Príncipe et al. 2018) |
| Syringacin M | P. syringae pv. tomato DC3000 | P. syringae | (Li et al. 2020) |
NE – non specified
Bacteriocins as modulators of gastrointestinal microbiota and population diversity
The autochthonous bacteria that colonize the entire human gastrointestinal tract, from the mouth to the colon, confer various physiologic benefits to the host. The prokaryotic symbiont population in humans ranges from 103–105 CFU/ml in the jejunal lumen) of healthy individuals to 1011–1012 CFU/ml in the colon, gut microbiota, prevents pathogen growth in the gastrointestinal tract (Sundin et al. 2017). This regulation is given through various microbial mechanisms, one of them is the release of bacteriocins, which prevent dysbiosis and consolidate the homeostasis of the gastrointestinal microbiota. The homeostatic balance in the human gut microbiota has become a significant public health problem due to changes in eating habits, type of diet, and administration of broad-spectrum antibiotics (Cotter et al. 2013). Ultra-processed food intake has increased saturated fats, omega-6 fatty acids, trans-fatty acids, and simple carbohydrates in the human diet while it has decreased the intake of omega-3 fatty acids, fiber, and complex carbohydrates. This diet high in fat and carbohydrates and low in micronutrients can disturb the human microbiota with concomitant metabolic disorders (Miclotte and Van de Wiele 2020).
Probiotics can colonize, at least temporally, the human gastrointestinal tract due to the efficient competition mediate by bacteriocin production. Thus, the intake of Lactobacillus species in probiotherapy has shown health-promoting effects on treating inflammatory gastrointestinal diseases like constipation, diarrhea, irritable bowel syndrome, gastritis, gastroesophageal reflux, ulcerative colitis syndrome, Crohn’s disease, among others (Kumar et al. 2016). Bacteriocins can play an essential role in the homeostasis of different subpopulations of microbial communities. For example, in the relationship of certain bacteriocin-producing, sensitive, and resistant bacterial populations bacteria can interact with each other in a set of incessant battles without a clear winner (Kerr et al. 2002).
In some cases, the growth rate of a resistant population can be higher than that of the bacteriocin-producing population (P), which generally possess a plasmid with genes encoding the bacteriocin and bacteriocin-specific immunity protein that make the bacteriocin-producing population immune to its bacteriocin. Still, at the same time, the resistant population (R) has a slower growth rate than that of the sensitive population (S). The susceptible population has an advantage over the resistant population because sensitive bacteria have a higher growth rate. The resistant population has an advantage over the bacteriocin-producing population because of its higher growth rate. And the bacteriocin-producing population can displace susceptible populations because bacteriocin-producing bacteria can kill sensitive bacteria making the three types of bacterial populations coexist in a balance of subpopulations preserving the diversity of the community (Kerr et al. 2002).
The bioinformatic analysis of bacteriocins encoded within 317 microbial genomes found in the human intestine revealed 175 bacteriocins in Firmicutes (which includes LAB), 79 in Proteobacteria, 34 in Bacteroidetes, and 25 in Actinobacteria (Drissi et al. 2015). The analysis showed that bacteriocins produced by the intestinal bacteria display wide differences, in the size and amino acid composition, compared to other bacteriocins. These bacteriocins contain less aspartic acid, leucine, arginine, and glutamic acid but more lysine and methionine. Depending on their α-helical structure, charge, and hydrophobicity, they may have a broader spectrum of activity (Zelezetsky and Tossi 2006) but, in turn, lower antimicrobial activity and, therefore, they can better modulate microbial populations (Drissi et al. 2015). The microbial community that inhabits the human gut appears to impart specific functions to human metabolism and health by interconnecting signals from the brain, the immune system, the endocrine system, and the gut microbiota itself (Vivarelli et al. 2019). So, depending on the type of bacteria colonizing the gastrointestinal tract will determine the type of signaling molecules released and, therefore, the impact on host health and disease. That is why the microbial diversity of microbiota is tightly regulated. An example of this type of regulation exerted by bacteriocins is the effect of plantaricin P1053 produced by Lactobacillus plantarum strain PBS067; which exhibited a broad-spectrum of antimicrobial activity against Gram-positive and Gram-negative bacteria. Furthermore, plantaricin P1053 showed an improvement in the viability of healthy cells and a proliferation reduction of cancerogenic human intestinal cells. The mechanism involved in this case was through the epidermal growth factor receptor (EGFR) pathways (De Giani et al. 2019). Bifidobacterium longum subsp. longum NCC2705 produces the bacteriocin serpin, which is a protease inhibitor that interacts directly with the host factors. Serpin inhibits pancreatic and neutrophil elastases by mediating some gastrointestinal anti-inflammatory effects (Ivanov et al. 2006). The production of bacteriocins by the microbiota that inhabits the human gut affects the individual’s metabolic processes, whether it improves health or causes dysbiosis and disease Therefore, bacteriocins production by the microbiota is tightly regulated. One way of exploiting the bacteriocin potential of prevailing bacterial commensals to cure multiresistant infections is to stimulate the endogenous bacteriocin producers at specific times and locations. S. salivarius population, for example, produces bacteriocins of high potency against infectious pathogens and is dominant and genetically diverse in the human digestive tract (Hols et al. 2019). Bacteriocin-related genes of S. salivarius can be activated upon addition of short ComS pheromone into the culture medium (Mignolet et al. 2018). Thus pheromone-based mobilization of bacteriocins in the commensal microbiota could be achieved in vivo by the addition of ComS pheromone which complexes with the ComR sensor activating the master regulator of competence (ComX), and coupling competence and predation response in S. salivarius (Hols et al. 2019). Nevertheless, oral administration of signaling pheromones remains elusive. To minimize environmental influences (i.e. resist most digestive proteases, the stomach barrier, and low solubility of signaling pheromones) and ensure the activating pheromone efficiency in vivo, more advanced enabling formulations to improve oral bioavailability is required.
A multidrug-resistant E. faecalis strain was actively killed by commensal enterococci. A heptapeptide pheromone, cOB1, produced by native E. faecalis; was involved in the killing of multidrug-resistant E. faecalis strain V583, the killing of V583, resulted from lethal cross-talk between accumulated mobile elements (Gilmore et al. 2015). Since multidrug-resistant Enterococcus possessed the limited ability to grow in the presence of commensal Enterococcus strains due to the production of peptide pheromones. We could hypothesize that infections caused by MDR strains can be fought by the same genera commensal strains using the suitable pheromone to activate the killing response. MDR enterococci colonize the patient after perturbating the native flora by antibiotic treatment when commensal enterococci strains are excluded. Therefore, a potential therapy could be the formulation of enterococci native strain along with the signaling pheromone. Currently, there is controversy over the adequate use of probiotherapy, more research must be done about whether probiotics are helpful and safe for various health conditions. We still do not know the concentrations necessary to benefit healthy and sick individuals and the time of probiotics intake to improve individual health.
Bacteriocins commercially available: a patent perspective
According to the World Intellectual Property Organization (WIPO), over the last 30 years, more than 800 patent applications with the term “bacteriocin” in title or abstract were published, while Espacenet website reports more than 8900. Fig. 4 shows the patents published between January 1, 2000, and August 7, 2020, using the Patent Inspiration search engine with the term “bacteriocin”. Over the last 20 years, China has published 234 patents, followed by the United States with 132, while Mexico only published 17 patents (Fig. 4). Among these patents, 312 (36.4%) are associated with nisin and lactic acid bacteria.
Fig. 4.

Timeline of bacteriocin patents reported worldwide from January 1, 2000 to August 7, 2020. Countries with the highest number of reported patents per year are shown. The figure was generated with the Patent Inspiration search engine (https://www.patentinspiration.com).
CN – China, US – United States, KR – Korea, RU – Russian Federation, UA – Ukraine, CA – Canada, JP – Japan, AU – Australia, NZ – New Zealand, GE – Germany, FR – France, TW – Taiwan, AT – Austria, PL – Poland, ES – Spain, AR – Argentina, MX – Mexico, BR – Brazil, DK – Denmark, CZ – Czech Republic, HU – Hungary, ZA – South Africa, SE – Sweden, IL – Israel, GB – United Kingdom.
Bacteriocins have fascinating properties concerning their size, structure, mechanism of action, inhibitory spectrum, and immunity mechanisms that endorse them with market potential. However, just four bacteriocin formulations are commercially available: nisin (NisaplinTM, BiosafeTM, OralpeaceTM), pediocin PA-1 (MicrogardTM, Alta 2341), sakacin (BactofermTM B-2, BactofermTM B-FM) and leucocin A (BactofermTM B-SF-43) are mainly used as food preservatives in the United States and Canada (Daba and Elkhateeb 2020; Radaic et al. 2020). Other FDA-approved bacteriocins, with the intended use as an antibacterial for food, are colicins, salmocins, and Clostridium phage lysins, but they are not in the market yet (Hahn-Löbmann et al. 2019). One limitation of using purified bacteriocins in the food industry could be the high cost of production and purification compared to the price of food additives. It is more feasible to produce formulations of whole bacteria with their metabolites and use them as “protective cultures” on foods. Thus, several bacteria that produce bacteriocin have obtained the GRAS status and are used commercially as a preservative in a wide range of food products or as probiotics. In the list is Carnobacterium divergens M35, Bacillus coagulans GBI-30, Bacillus subtilis strain SG 188, Lactobacillus plantarum Lp-115, Lactobacillus fermentum CECT5716, Lactobacillus paracasei strain F19, Lactobacillus plantarum strain 299v, Bacillus coagulans SNZ1969, Lactobacillus acidophilus DDS-1, Bifidobacterium animalis subsp. lactic UABla-12, Bifidobacterium longum BB536, Bifidobacterium bifidum Rosell®-71, Bifidobacterium longum ssp. infantis Rosell®-33, Lactobacillus helveticus Rosell®-52, Lactobacillus rhamnosus LGG®, Lactobacillus curvatus DSM 18775, and Streptococcus salivarius K12. An alternative to the costly fermentation production and purification of bacteriocins from a natural producer strain is chemical synthesis. Advances in solid-phase peptide chemical synthesis, lower price for reagents and building blocks, has made the chemical synthesis of bacteriocins more attractive and competitive. Furthermore, through chemical approaches, it is possible to perform amino acid substitution, use non-natural or modified residues, and make backbone and side-chain modifications to improve potency or stability of bacteriocins (Bédard and Biron 2018). Those advanced chemical methods will surely enable the screening and identification of more potent or stable bacteriocins.
Bacteriocin formulations can be used as nutritional supplementation. Few in vivo experiments on bacteriocin dietary formulations have described the effects of bacteriocins on the gastrointestinal microbiota of mice, rats, rabbits, ruminants, fish, and poultry. A multispecies probiotic combination (Lactobacillus reuteri, Enterococcus faecium, B. animalis, Pediococcus acidilactici, and Lactobacillus salivarius) increased nutrient digestibility, digestive enzyme activities, and anti-inflammatory effect in broilers (Palamidi et al. 2016). The efficacy of L. acidophilus, B. subtilis, and Clostridium butyricum supplementation in broilers improved growth performance, ileal amino acids digestibility, and humoral immunity (Zhang and Kim 2014). The addition of nisin (alone or in combination with salinomycin or monensin) to broilers’ diet was associated with an apparent nutrient digestibility (Kierończyk et al. 2017). Dietary supplementation with Paenibacillus ehimensis NPUST1 (bacteriocin-like activities against Aeromonas hydrophila) improved the growth performance, immunity, and disease resistance in Nile tilapia (Chen et al. 2019). Altogether, these reports indicate the potential of bacteriocins as nutritional supplementation.
Compared to the food industry, the medical field could represent a higher profit for the use of bacteriocins. However, to exploit the full potential of bacteriocins in the medical industry, they must overcome some drawbacks such as sensitivity to proteases, immunogenicity issues, and the development of bacteriocin resistance by pathogenic bacteria. In this regard, advanced chemical approaches can be used to make disulfide bridges, head-to-tail macrocyclization, N-terminus formylation, amino acid substitutions, and other modifications; to make bacteriocins more potent and stable, enabling them to surpass their current drawbacks (Bédard and Biron 2018). Another factor that prevents the commercial use of bacteriocins in medical applications might be attributed to the low approval of the regulatory process. Over the last decade, the number of in vivo trials has increased, but clinical application of bacteriocins requires more investigation to determine their efficacy, stability, and kinetic properties in/on the human body. For example, nisin ZP and nisin AP, significantly reduced the tumor volume in mouse-induced oral cancer. Lacticin 3147 reduced S. aureus Xen 29 growth and prevented dissemination of the pathogen in the spleen, liver, and kidney of a murine model. Salivaricin prevented Candida albicans colonization in the oral cavity of a mouse model. ESL5 has been applied as a lotion in a patient with inflammatory acne lesions caused by Propionibacterium acnes (López-Cuellar et al. 2016; Soltani et al. 2021). Lantibiotics such as nisin, clausin, and amyloliquecidin (AmyA) are effective in treating S. aureus-induced skin infection in mice (van Staden et al. 2016). AS-48 prevents and treats skin diseases, even with multi-drug resistant microorganisms, and has the potential as a leishmanicidal agent (Cebrian et al. 2019). Despite their therapeutic possibilities, bacteriocins have not yet entered into clinical use, and only a limited number have been selected for tests in clinical trials. NAI-107 (Microbispora corallina) and mutacin 1140 (S. mutans JH1000) are at the late preclinical phase; NVB302 and Moli1901 (Actinoplanes liguriae NCIMB41362) have completed phase I and phase II clinical trials (for clinical studies, see Ongey et al. 2017; Soltani et al. 2021). Finally, apart from those technical limitations mentioned, several factors not covered in this review preclude most patented products make it to market.
Conclusions
Bacteriocins have become an attractive tool to preserve food and improve human health. Bacteriocins can eliminate specific pathogen microorganisms while favoring the preservation of other populations. Since the impact of bacteriocins on each microbial community is not well understood yet, there are limitations to exploit all their potential. It is necessary to continue performing rigorous research focused on developing antimicrobials, anticancer agents, and microbiota modulators before bacteriocins can be available to consumers.
Acknowledgments
Funding was provided by the intramural program of the Vicerrectoría de Investigación y Estudios de Posgrado, BUAP through grants PV17-ID00337, 100524554-VIEP2019. Catherine Cesa-Luna received a scholarship (number 294272) from the National Council of Science and Technology (CONACyT).
Footnotes
Conflict of interest
The authors do not report any financial or personal connections with other persons or organizations, which might negatively affect the contents of this publication and/or claim authorship rights to this publication.
Literature
- Abdi-Ali A, Worobec EA, Deezagi A, Malekzadeh F. Cytotoxic effects of pyocin S2 produced by Pseudomonas aeruginosa on the growth of three human cell lines. Can J Microbiol. 2004. May 01;50(5):375–381. 10.1139/w04-019 [DOI] [PubMed] [Google Scholar]
- Abriouel H, Franz CMAP, Omar NB, Gálvez A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol Rev. 2011. January;35(1):201–232. 10.1111/j.1574-6976.2010.00244.x [DOI] [PubMed] [Google Scholar]
- Al-Madboly LA, El-Deeb NM, Kabbash A, Nael MA, Kenawy AM, Ragab AE. Purification, characterization, identification, and anticancer activity of a circular bacteriocin from Enterococcus thailandicus. Front Bioeng Biotechnol. 2020. June 23;8:450. 10.3389/fbioe.2020.00450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida PF, Pokorny A. Interactions of antimicrobial peptides with lipid bilayers. In: Egelman EH, editor. Comprehensive Biophysics. Amsterdam (Netherlands): Elsevier; 2012. p. 189–222. 10.1016/B978-0-12-374920-8.00515-4 [DOI] [Google Scholar]
- Alvarez-Sieiro P, Montalbán-López M, Mu D, Kuipers OP. Bacteriocins of lactic acid bacteria: extending the family. Appl Microbiol Biotechnol. 2016. April;100(7):2939–2951. 10.1007/s00253-016-7343-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anastasiou R, Aktypis A, Georgalaki M, Papadelli M, De Vuyst L, Tsakalidou E. Inhibition of Clostridium tyrobutyricum by Streptococcus macedonicus ACA-DC 198 under conditions mimicking Kasseri cheese production and ripening. Int Dairy J. 2009. May;19(5): 330–335. 10.1016/j.idairyj.2008.12.001 [DOI] [Google Scholar]
- Atanaskovic I, Kleanthous C. Tools and approaches for dissecting protein bacteriocin import in Gram-Negative bacteria. Front Microbiol. 2019. March 28;10:646. 10.3389/fmicb.2019.00646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baindara P, Korpole S, Grover V. Bacteriocins: perspective for the development of novel anticancer drugs. Appl Microbiol Biotechnol. 2018. December;102(24):10393–10408. 10.1007/s00253-018-9420-8 [DOI] [PubMed] [Google Scholar]
- Baindara P, Singh N, Ranjan M, Nallabelli N, Chaudhry V, Pathania GL, Sharma N, Kumar A, Patil PB, Korpole S. Laterosporulin10: a novel defensin like Class IId bacteriocin from Brevibacillus sp. strain SKDU10 with inhibitory activity against microbial pathogens. Microbiology. 2016. August 01;162(8):1286–1299. 10.1099/mic.0.000316 [DOI] [PubMed] [Google Scholar]
- Balandin SV, Sheremeteva EV, Ovchinnikova TV. Pediocin-like antimicrobial peptides of bacteria. Biochemistry (Mosc). 2019. May; 84(5):464–478. 10.1134/S000629791905002X [DOI] [PubMed] [Google Scholar]
- Balciunas EM, Castillo Martinez FA, Todorov SD, Franco BDGM, Converti A, Oliveira RPS. Novel biotechnological applications of bacteriocins: a review. Food Control. 2013. July;32(1):134–142. 10.1016/j.foodcont.2012.11.025 [DOI] [Google Scholar]
- Baquero F, Lanza VF, Baquero MR, del Campo R, Bravo-Vázquez DA. Microcins in Enterobacteriaceae: peptide antimicrobials in the eco-active intestinal chemosphere. Front Microbiol. 2019. October 9;10:2261. 10.3389/fmicb.2019.02261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bédard F, Biron E. Recent progress in the chemical synthesis of class II and S-glycosylated bacteriocins. Front Microbiol. 2018. May 23;9:1048. 10.3389/fmicb.2018.01048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrens HM, Six A, Walker D, Kleanthous C. The therapeutic potential of bacteriocins as protein antibiotics. Emerging Top Life Sci. 2017. April 21;1(1):65–74. 10.1042/ETLS20160016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benabbou R, Subirade M, Desbiens M, Fliss I. Divergicin M35-chitosan film: development and characterization. Probiotics Antimicrob Proteins. 2020. December;12(4):1562–1570. 10.1007/s12602-020-09660-9 [DOI] [PubMed] [Google Scholar]
- Bengtsson T, Lönn J, Khalaf H, Palm E. The lantibiotic gallidermin acts bactericidal against Staphylococcus epidermidis and Staphylococcus aureus and antagonizes the bacteria‐induced proinflammatory responses in dermal fibroblasts. MicrobiologyOpen. 2018. December; 7(6):e00606. 10.1002/mbo3.606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bengtsson T, Selegård R, Musa A, Hultenby K, Utterström J, Sivlér P, Skog M, Nayeri F, Hellmark B, Söderquist B, et al. Plantaricin NC8 αβ exerts potent antimicrobial activity against Staphylococcus spp. and enhances the effects of antibiotics. Sci Rep. 2020. December; 10(1):3580. 10.1038/s41598-020-60570-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermudez-Brito M, Plaza-Díaz J, Muñoz-Quezada S, Gómez-Llorente C, Gil A. Probiotic mechanisms of action. Ann Nutr Metab. 2012;61(2):160–174. 10.1159/000342079 [DOI] [PubMed] [Google Scholar]
- Bogovič Matijašić B, Koman Rajšp M, Perko B, Rogelj I. Inhibition of Clostridium tyrobutyricum in cheese by Lactobacillus gasseri. Int Dairy J. 2007. February;17(2):157–166. 10.1016/j.idairyj.2006.01.011 [DOI] [Google Scholar]
- Bonelli RR, Schneider T, Sahl HG, Wiedemann I. Insights into in vivo activities of lantibiotics from gallidermin and epidermin mode-of-action studies. Antimicrob Agents Chemother. 2006. April;50(4): 1449–1457. 10.1128/AAC.50.4.1449-1457.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosák J, Hrala M, Micenková L, Šmajs D. Non-antibiotic antibacterial peptides and proteins of Escherichia coli: efficacy and potency of bacteriocins. Expert Rev Anti Infect Ther. 2021. March;19(3):309–322. 10.1080/14787210.2020.1816824 [DOI] [PubMed] [Google Scholar]
- Budič M, Rijavec M, Petkovšek Ž, Žgur-Bertok D. Escherichia coli bacteriocins: antimicrobial efficacy and prevalence among isolates from patients with bacteraemia. PLoS One. 2011. December 19;6(12): e28769. 10.1371/journal.pone.0028769 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlin Fagundes P, Miceli de Farias F, Cabral da Silva Santos O, Souza da Paz JA, Ceotto-Vigoder H, Sales Alviano D, Villela Romanos MT, de Freire Bastos MC. The four-component aureocin A70 as a promising agent for food biopreservation. Int J Food Microbiol. 2016. November;237:39–46. 10.1016/j.ijfoodmicro.2016.08.017 [DOI] [PubMed] [Google Scholar]
- Castellano P, Pérez Ibarreche M, Blanco Massani M, Fontana C, Vignolo G. Strategies for pathogen biocontrol using lactic acid bacteria and their metabolites: A focus on meat ecosystems and industrial environments. Microorganisms. 2017. July 11;5(3):38. 10.3390/microorganisms5030038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cebrián R, Rodríguez-Cabezas ME, Martín-Escolano R, Rubiño S, Garrido-Barros M, Montalbán-López M, Rosales MJ, Sánchez-Moreno M, Valdivia E, Martínez-Bueno M, et al. Preclinical studies of toxicity and safety of the AS-48 bacteriocin. J Adv Res. 2019. November;20:129–139. 10.1016/j.jare.2019.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cesa-Luna C, Baez A, Quintero-Hernández V, De la Cruz-Enríquez J, Castañeda-Antonio MD, Muñoz-Rojas J. The importance of antimicrobial compounds produced by beneficial bacteria on the biocontrol of phytopathogens. Acta Biol Colomb. 2020. January 01;25(1):140–154. 10.15446/abc.v25n1.76867 [DOI] [Google Scholar]
- Chen SW, Liu CH, Hu SY. Dietary administration of probiotic Paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019. January;84:695–703. 10.1016/j.fsi.2018.10.059 [DOI] [PubMed] [Google Scholar]
- Chowdhury SP, Hartmann A, Gao X, Borriss R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42 – a review. Front Microbiol. 2015. July 28;6:780. 10.3389/fmicb.2015.00780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotter PD, Ross RP, Hill C. Bacteriocins – a viable alternative to antibiotics? Nat Rev Microbiol. 2013. February;11(2):95–105. 10.1038/nrmicro2937 [DOI] [PubMed] [Google Scholar]
- Daba GM, Elkhateeb WA. Bacteriocins of lactic acid bacteria as biotechnological tools in food and pharmaceuticals: current applications and future prospects. Biocatal Agric Biotechnol. 2020. September;28(28):101750. 10.1016/j.bcab.2020.101750 [DOI] [Google Scholar]
- David OM, Onifade OE. Effects of partially purified enterocins from Enterococcus faecalis strains on the growth of some phytopathogenic fungi. Ruhuna J Sci. 2018. December 31;9(2):160–168. 10.4038/rjs.v9i2.44 [DOI] [Google Scholar]
- De Giani A, Bovio F, Forcella M, Fusi P, Sello G, Di Gennaro P. Identification of a bacteriocin-like compound from Lactobacillus plantarum with antimicrobial activity and effects on normal and cancerogenic human intestinal cells. AMB Express. 2019. December;9(1):88. 10.1186/s13568-019-0813-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Fuente-Salcido N, Guadalupe Alanís-Guzmán M, Bideshi DK, Salcedo-Hernández R, Bautista-Justo M, Barboza-Corona JE. Enhanced synthesis and antimicrobial activities of bacteriocins produced by Mexican strains of Bacillus thuringiensis. Arch Microbiol. 2008. December;190(6):633–640. 10.1007/s00203-008-0414-2 [DOI] [PubMed] [Google Scholar]
- Dobson A, Cotter PD, Ross RP, Hill C. Bacteriocin production: a probiotic trait? Appl Environ Microbiol. 2012. January 01;78(1):1–6. 10.1128/AEM.05576-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drissi F, Buffet S, Raoult D, Merhej V. Common occurrence of antibacterial agents in human intestinal microbiota. Front Microbiol. 2015. May 07;6:441. 10.3389/fmicb.2015.00441 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feliatra F, Muchlisin ZA, Teruna HY, Utamy WR, Nursyirwani N, Dahliaty A. Potential of bacteriocins produced by probiotic bacteria isolated from tiger shrimp and prawns as antibacterial to Vibrio, Pseudomonas, and Aeromonas species on fish. F1000 Res. 2018;7:415. 10.12688/f1000research.13958.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabrielsen C, Brede DA, Nes IF, Diep DB. Circular bacteriocins: biosynthesis and mode of action. Appl Environ Microbiol. 2014. November 15;80(22):6854–6862. 10.1128/AEM.02284-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gálvez A, Abriouel H, López RL, Omar NB. Bacteriocin-based strategies for food biopreservation. Int J Food Microbiol. 2007. November; 120(1–2):51–70. 10.1016/j.ijfoodmicro.2007.06.001 [DOI] [PubMed] [Google Scholar]
- Gebhart D, Lok S, Clare S, Tomas M, Stares M, Scholl D, Donskey CJ, Lawley TD, Govoni GR. A modified R-type bacteriocin specifically targeting Clostridium difficile prevents colonization of mice without affecting gut microbiota diversity. MBio. 2015. May 01;6(2):e02368-14. 10.1128/mBio.02368-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghequire MGK, De Mot R. The tailocin tale: peeling off phage tails. Trends Microbiol. 2015. October;23(10):587–590. 10.1016/j.tim.2015.07.011 [DOI] [PubMed] [Google Scholar]
- Ghequire MGK, De Mot R. Turning over a new leaf: bacteriocins going green. Trends Microbiol. 2018. January;26(1):1–2. 10.1016/j.tim.2017.11.001 [DOI] [PubMed] [Google Scholar]
- Ghequire MGK, Öztürk B, De Mot R. Lectin-like bacteriocins. Front Microbiol. 2018a. November 12;9:2706. 10.3389/fmicb.2018.02706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghequire MGK, Swings T, Michiels J, Buchanan SK, De Mot R. Hitting with a BAM: selective killing by lectin-like bacteriocins. MBio. 2018b. March 20;9(2):e02138-17. 10.1128/mBio.02138-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillor O, Etzion A, Riley MA. The dual role of bacteriocins as anti- and probiotics. Appl Microbiol Biotechnol. 2008. December;81(4):591–606. 10.1007/s00253-008-1726-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilmore MS, Rauch M, Ramsey MM, Himes PR, Varahan S, Manson JM, Lebreton F, Hancock LE. Pheromone killing of multidrug-resistant Enterococcus faecalis V583 by native commensal strains. Proc Natl Acad Sci USA. 2015. June 09;112(23):7273–7278. 10.1073/pnas.1500553112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grinter R, Milner J, Walker D. Bacteriocins active against plant pathogenic bacteria. Biochem Soc Trans. 2012. December 01;40(6):1498–1502. 10.1042/BST20120206 [DOI] [PubMed] [Google Scholar]
- Guralp SA, Murgha YE, Rouillard JM, Gulari E. From design to screening: a new antimicrobial peptide discovery pipeline. PLoS One. 2013. March 19;8(3):e59305. 10.1371/journal.pone.0059305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahn-Löbmann S, Stephan A, Schulz S, Schneider T, Shaverskyi A, Tusé D, Giritch A, Gleba Y. Colicins and salmocins – New classes of plant-made non-antibiotic food antibacterials. Front Plant Sci. 2019. April 9;10:437. 10.3389/fpls.2019.00437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hegemann JD, Zimmermann M, Xie X, Marahiel MA. Lasso peptides: an intriguing class of bacterial natural products. Acc Chem Res. 2015. July 21;48(7):1909–1919. 10.1021/acs.accounts.5b00156 [DOI] [PubMed] [Google Scholar]
- Helal MMI, Hashem AM, Ghobashy MOI, Shalaby SG. Some physiological and biological studies on reuterin production from Lactobacillus reuteri. J Prob Health. 2016;04(03):1–8. 10.4172/2329-8901.1000156 [DOI] [Google Scholar]
- Helbig S, Braun V. Mapping functional domains of colicin M. J Bacteriol. 2011. February 15;193(4):815–821. 10.1128/JB.01206-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hibbing ME, Fuqua C, Parsek MR, Peterson SB. Bacterial competition: surviving and thriving in the microbial jungle. Nat Rev Microbiol. 2010. January;8(1):15–25. 10.1038/nrmicro2259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hols P, Ledesma-García L, Gabant P, Mignolet J. Mobilization of microbiota commensals and their bacteriocins for therapeutics. Trends Microbiol. 2019. August;27(8):690–702. 10.1016/j.tim.2019.03.007 [DOI] [PubMed] [Google Scholar]
- Huang T, Zhang X, Pan J, Su X, Jin X, Guan X. Purification and characterization of a novel cold shock protein-like bacteriocin synthesized by Bacillus thuringiensis. Sci Rep. 2016. December 16;6(1):35560. 10.1038/srep35560 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huo L, Ökesli A, Zhao M, van der Donk WA. Insights into the biosynthesis of duramycin. Appl Environ Microbiol. 2017. February 01; 83(3):e02698-16. 10.1128/AEM.02698-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurst MRH, Beattie A, Jones SA, Laugraud A, van Koten C, Harper L. Serratia proteamaculans strain AGR96X encodes an antifeeding prophage (tailocin) with activity against grass grub (Costelytra giveni) and manuka beetle (Pyronota species) larvae. Appl Environ Microbiol. 2018. March 16;84(10):e02739-17. 10.1128/AEM.02739-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iseppi R, Pilati F, Marini M, Toselli M, de Niederhäusern S, Guerrieri E, Messi P, Sabia C, Manicardi G, Anacarso I, et al. Antilisterial activity of a polymeric film coated with hybrid coatings doped with Enterocin 416K1 for use as bioactive food packaging. Int J Food Microbiol. 2008. April;123(3):281–287. 10.1016/j.ijfoodmicro.2007.12.015 [DOI] [PubMed] [Google Scholar]
- Ivanov D, Emonet C, Foata F, Affolter M, Delley M, Fisseha M, Blum-Sperisen S, Kochhar S, Arigoni F. A serpin from the gut bacterium Bifidobacterium longum inhibits eukaryotic elastase-like serine proteases. J Biol Chem. 2006. June;281(25):17246–17252. 10.1074/jbc.M601678200 [DOI] [PubMed] [Google Scholar]
- Juturu V, Wu JC. Microbial production of bacteriocins: latest research development and applications. Biotechnol Adv. 2018. December; 36(8): 2187–2200. 10.1016/j.biotechadv.2018.10.007 [DOI] [PubMed] [Google Scholar]
- Kaur Maan P, Garcha S. Bacteriocins from Gram-negative Rhizobium spp. Adv Biores. January 2018;9(1):36–43. 10.15515/abr.0976-4585.9.1.3643 [DOI] [Google Scholar]
- Kaur S, Kaur S. Bacteriocins as potential anticancer agents. Front Pharmacol. 2015. November 10;6:272. 10.3389/fphar.2015.00272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerr B, Riley MA, Feldman MW, Bohannan BJM. Local dispersal promotes biodiversity in a real-life game of rock-paper-scissors. Nature. 2002. July;418(6894):171–174. 10.1038/nature00823 [DOI] [PubMed] [Google Scholar]
- Kierończyk B, Sassek M, Pruszyńska-Oszmałek E, Kołodziejski P, Rawski M, Świątkiewicz S, Józefiak D. The physiological response of broiler chickens to the dietary supplementation of the bacteriocin nisin and ionophore coccidiostats. Poult Sci. 2017. November;96(11):4026–4037. 10.3382/ps/pex234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim JG, Park BK, Kim SU, Choi D, Nahm BH, Moon JS, Reader JS, Farrand SK, Hwang I. Bases of biocontrol: sequence predicts synthesis and mode of action of agrocin 84, the Trojan Horse antibiotic that controls crown gall. Proc Natl Acad Sci USA. 2006. June 06;103(23):8846–8851. 10.1073/pnas.0602965103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kjos M, Nes IF, Diep DB. Mechanisms of resistance to bacteriocins targeting the mannose phosphotransferase system. Appl Environ Microbiol. 2011. May 15;77(10):3335–3342. 10.1128/AEM.02602-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohoutova D, Smajs D, Moravkova P, Cyrany J, Moravkova M, Forstlova M, Cihak M, Rejchrt S, Bures J. Escherichia colistrains of phylogenetic group B2 and D and bacteriocin production are associated with advanced colorectal neoplasia. BMC Infect Dis. 2014. December;14(1):733. 10.1186/s12879-014-0733-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruszewska D, Sahl HG, Bierbaum G, Pag U, Hynes SO, Ljungh Å. Mersacidin eradicates methicillin-resistant Staphylococcus aureus (MRSA) in a mouse rhinitis model. J Antimicrob Chemother. 2004. September 01;54(3):648–653. 10.1093/jac/dkh387 [DOI] [PubMed] [Google Scholar]
- Kumar B, Balgir PP, Kaur B, Mittu B, Chauhan A. In vitro cytotoxicity of native and rec-Pediocin CP2 against cancer cell lines: a comparative study. Pharm Anal Acta. 2012;03(08):1–4. 10.4172/2153-2435.1000183 [DOI] [Google Scholar]
- Kumar M, Dhaka P, Vijay D, Vergis J, Mohan V, Kumar A, Kurkure NV, Barbuddhe SB, Malik SVS, Rawool DB. Antimicrobial effects of Lactobacillus plantarum and Lactobacillus acidophilus against multidrug-resistant enteroaggregative Escherichia coli. Int J Antimicrob Agents. 2016. September;48(3):265–270. 10.1016/j.ijantimicag.2016.05.014 [DOI] [PubMed] [Google Scholar]
- Ladjouzi R, Lucau-Danila A, Benachour A, Drider D. A leaderless two-peptide bacteriocin, enterocin DD14, is involved in its own self-immunity: evidence and insights. Front Bioeng Biotechnol. 2020. June 26;8:644. 10.3389/fbioe.2020.00644 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Latham RD, Gell DA, Fairbairn RL, Lyons AB, Shukla SD, Cho KY, Jones DA, Harkness NM, Tristram SG. An isolate of Haemophilus haemolyticus produces a bacteriocin-like substance that inhibits the growth of nontypeable Haemophilus influenzae. Int J Antimicrob Agents. 2017. April;49(4):503–506. 10.1016/j.ijantimicag.2016.12.010 [DOI] [PubMed] [Google Scholar]
- Lavermicocca P, Lonigro SL, Valerio F, Evidente A, Visconti A. Reduction of olive knot disease by a bacteriocin from Pseudomonas syringae pv. ciccaronei. Appl Environ Microbiol. 2002. March;68(3): 1403–1407. 10.1128/AEM.68.3.1403-1407.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee NK, Paik HD. Partial characterization of lacticin NK24, a newly identified bacteriocin of Lactococcus lactis NK24 isolated from Jeot-gal. Food Microbiol. 2001. February;18(1):17–24. 10.1006/fmic.2000.0368 [DOI] [Google Scholar]
- Li JZ, Zhou LY, Peng YL, Fan J. Pseudomonas bacteriocin syringacin M released upon desiccation suppresses the growth of sensitive bacteria in plant necrotic lesions. Microb Biotechnol. 2020. January;13(1):134–147. 10.1111/1751-7915.13367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu G, Lv Y, Li P, Zhou K, Zhang J. Pentocin 31–1, an anti-Listeria bacteriocin produced by Lactobacillus pentosus 31–1 isolated from Xuan-Wei Ham, a traditional China fermented meat product. Food Control. 2008. April;19(4):353–359. 10.1016/j.foodcont.2007.04.010 [DOI] [Google Scholar]
- Liu X, Basu U, Miller P, McMullen LM. Stress response and adaptation of Listeria monocytogenes 08-5923 exposed to a sublethal dose of carnocyclin A. Appl Environ Microbiol. 2014. July 01;80(13):3835–3841. 10.1128/AEM.00350-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- López-Cuellar MR, Rodríguez-Hernández AI, Chavarría-Hernández N. LAB bacteriocin applications in the last decade. Biotechnol Biotechnol Equip. 2016. November 01;30(6):1039–1050. 10.1080/13102818.2016.1232605 [DOI] [Google Scholar]
- Maldonado-Barragán A, Caballero-Guerrero B, Martín V, Ruiz-Barba JL, Rodríguez JM. Purification and genetic characterization of gassericin E, a novel co-culture inducible bacteriocin from Lactobacillus gasseri EV1461 isolated from the vagina of a healthy woman. BMC Microbiol. 2016. December;16(1):37. 10.1186/s12866-016-0663-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marín-Cevada V, Muñoz-Rojas J, Caballero-Mellado J, Mascarúa-Esparza MA, Castañeda-Lucio M, Carreño-López R, Estrada-de los Santos P, Fuentes-Ramírez LE. Antagonistic interactions among bacteria inhabiting pineapple. Appl Soil Ecol. 2012. October;61:230–235. 10.1016/j.apsoil.2011.11.014 [DOI] [Google Scholar]
- McCaughey LC, Grinter R, Josts I, Roszak AW, Waløen KI, Cogdell RJ, Milner J, Evans T, Kelly S, Tucker NP, et al. Lectin-like bacteriocins from Pseudomonas spp. utilise D-rhamnose containing lipopolysaccharide as a cellular receptor. PLoS Pathog. 2014. February 6; 10(2):e1003898. 10.1371/journal.ppat.1003898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCaughey LC, Ritchie ND, Douce GR, Evans TJ, Walker D. Efficacy of species-specific protein antibiotics in a murine model of acute Pseudomonas aeruginosa lung infection. Sci Rep. 2016. September; 6(1):30201. 10.1038/srep30201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michel-Briand Y, Baysse C. The pyocins of Pseudomonas aeruginosa. Biochimie. 2002. May;84(5–6):499–510. 10.1016/S0300-9084(02)01422-0 [DOI] [PubMed] [Google Scholar]
- Miclotte L, Van de Wiele T. Food processing, gut microbiota and the globesity problem. Crit Rev Food Sci Nutr. 2020. June 16;60(11): 1769–1782. 10.1080/10408398.2019.1596878 [DOI] [PubMed] [Google Scholar]
- Mignolet J, Fontaine L, Sass A, Nannan C, Mahillon J, Coenye T, Hols P. Circuitry rewiring directly couples competence to predation in the gut dweller Streptococcus salivarius. Cell Rep. 2018. February; 22(7):1627–1638. 10.1016/j.celrep.2018.01.055 [DOI] [PubMed] [Google Scholar]
- Miller P, McMullen LM. Mechanism for temperature-dependent production of piscicolin 126. Microbiology. 2014. August 01;160(8): 1670–1678. 10.1099/mic.0.078030-0 [DOI] [PubMed] [Google Scholar]
- Mills S, Griffin C, O’Connor PM, Serrano LM, Meijer WC, Hill C, Ross RP. A multibacteriocin cheese starter system, comprising nisin and lacticin 3147 in Lactococcus lactis, in combination with plantaricin from Lactobacillus plantarum. Appl Environ Microbiol. 2017. July 15;83(14):e00799-17. 10.1128/AEM.00799-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitkowski P, Jagielska E, Nowak E, Bujnicki JM, Stefaniak F, Niedziałek D, Bochtler M, Sabała I. Structural bases of peptidoglycan recognition by lysostaphin SH3b domain. Sci Rep. 2019. December; 9(1):5965. 10.1038/s41598-019-42435-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molloy EM, Casjens SR, Cox CL, Maxson T, Ethridge NA, Margos G, Fingerle V, Mitchell DA. Identification of the minimal cytolytic unit for streptolysin S and an expansion of the toxin family. BMC Microbiol. 2015. December;15(1):141. 10.1186/s12866-015-0464-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monteiro CA, Levy RB, Claro RM, de Castro IRR, Cannon G. Increasing consumption of ultra-processed foods and likely impact on human health: evidence from Brazil. Public Health Nutr. 2010. December 20;14(1):5–13. 10.1017/S1368980010003241 [DOI] [PubMed] [Google Scholar]
- Moubarac JC, Martins APB, Claro RM, Levy RB, Cannon G, Monteiro CA. Consumption of ultra-processed foods and likely impact on human health. Evidence from Canada. Public Health Nutr. 2013. December;16(12):2240–2248. 10.1017/S1368980012005009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naz SA, Jabeen N, Sohail M, Rasool SA. Biophysicochemical characterization of Pyocin SA189 produced by Pseudomonas aeruginosa SA189. Braz J Microbiol. 2015. December;46(4):1147–1154. 10.1590/S1517-838246420140737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nazari M, Smith DL. A PGPR-produced bacteriocin for sustainable agriculture: a review of thuricin 17 characteristics and applications. Front Plant Sci. 2020. July 7;11:916. 10.3389/fpls.2020.00916 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neish AS. Microbes in gastrointestinal health and disease. Gastroenterology. 2009. January;136(1):65–80. 10.1053/j.gastro.2008.10.080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen C, Nguyen VD. Discovery of azurin-like anticancer bacteriocins from human gut microbiome through homology modeling and molecular docking against the tumor suppressor p53. Biomed Res Int. 2016;2016:1–12. 10.1155/2016/8490482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noinaj N, Kuszak AJ, Balusek C, Gumbart JC, Buchanan SK. Lateral opening and exit pore formation are required for BamA function. Structure. 2014. July;22(7):1055–1062. 10.1016/j.str.2014.05.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oliveira MM, Ramos ETA, Drechsel MM, Vidal MS, Schwab S, Baldani JI. Gluconacin from Gluconacetobacter diazotrophicus PAL5 is an active bacteriocin against phytopathogenic and beneficial sugarcane bacteria. J Appl Microbiol. 2018. December;125(6):1812–1826. 10.1111/jam.14074 [DOI] [PubMed] [Google Scholar]
- Oluyombo O, Penfold CN, Diggle SP. Competition in biofilms between cystic fibrosis isolates of Pseudomonas aeruginosa is shaped by R-pyocins. MBio. 2019. January 29;10(1):e01828-18. 10.1128/mBio.01828-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ongey EL, Yassi H, Pflugmacher S, Neubauer P. Pharmacological and pharmacokinetic properties of lanthipeptides undergoing clinical studies. Biotechnol Lett. 2017. April;39(4):473–482. 10.1007/s10529-016-2279-9 [DOI] [PubMed] [Google Scholar]
- Paiva AD, Breukink E, Mantovani HC. Role of lipid II and membrane thickness in the mechanism of action of the lantibiotic bovicin HC5. Antimicrob Agents Chemother. 2011. November;55(11):5284–5293. 10.1128/AAC.00638-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palamidi I, Fegeros K, Mohnl M, Abdelrahman WHA, Schatzmayr G, Theodoropoulos G, Mountzouris KC. Probiotic form effects on growth performance, digestive function, and immune related biomarkers in broilers. Poult Sci. 2016. July;95(7):1598–1608. 10.3382/ps/pew052 [DOI] [PubMed] [Google Scholar]
- Parnasa R, Sendersky E, Simkovsky R, Waldman Ben-Asher H, Golden SS, Schwarz R. A microcin processing peptidase‐like protein of the cyanobacterium Synechococcus elongatus is essential for secretion of biofilm‐promoting proteins. Environ Microbiol Rep. 2019. June;11(3):456–463. 10.1111/1758-2229.12751 [DOI] [PubMed] [Google Scholar]
- Parret AHA, Temmerman K, De Mot R. Novel lectin-like bacteriocins of biocontrol strain Pseudomonas fluorescens Pf-5. Appl Environ Microbiol. 2005. September;71(9):5197–5207. 10.1128/AEM.71.9.5197-5207.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patz S, Becker Y, Richert-Pöggeler KR, Berger B, Ruppel S, Huson DH, Becker M. Phage tail-like particles are versatile bacterial nanomachines – a mini-review. J Adv Res. 2019. September;19:75–84. 10.1016/j.jare.2019.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez RH, Zendo T, Sonomoto K. Circular and leaderless bacteriocins: Biosynthesis, mode of action, applications, and prospects. Front Microbiol. 2018. September 4;9:2085. 10.3389/fmicb.2018.02085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez RH, Zendo T, Sonomoto K. Novel bacteriocins from lactic acid bacteria (LAB): various structures and applications. Microb Cell Fact. 2014;13(Suppl 1):S3. 10.1186/1475-2859-13-S1-S3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pimentel-Filho NJ, Mantovani HC, de Carvalho AF, Dias RS, Vanetti MCD. Efficacy of bovicin HC5 and nisin combination against Listeria monocytogenes and Staphylococcus aureus in fresh cheese. Int J Food Sci Technol. 2014. February;49(2):416–422. 10.1111/ijfs.12316 [DOI] [Google Scholar]
- Pogány Simonová M, Chrastinová Ľ, Lauková A. Autochtonous strain Enterococcus faecium EF2019(CCM7420), its bacteriocin and their beneficial effects in broiler rabbits – a review. Animals (Basel). 2020. July 14;10(7):1188. 10.3390/ani10071188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price R, Jayeola V, Niedermeyer J, Parsons C, Kathariou S. The Listeria monocytogenes key virulence determinants hly and prfA are involved in biofilm formation and aggregation but not colonization of fresh produce. Pathogens. 2018. February 01;7(1):18. 10.3390/pathogens7010018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Príncipe A, Fernandez M, Torasso M, Godino A, Fischer S. Effectiveness of tailocins produced by Pseudomonas fluorescens SF4c in controlling the bacterial-spot disease in tomatoes caused by Xanthomonas vesicatoria. Microbiol Res. 2018. July;212-213:94–102. 10.1016/j.micres.2018.05.010 [DOI] [PubMed] [Google Scholar]
- Rebuffat S. Microcins and other bacteriocins: Bridging the gaps between killing strategies, ecology and applications. In: Dorit RL, Roy SM, Riley MA, editors. The Bacteriocins: Current knowledge and future prospects. Norfolk (UK): Caister Academic Press; 2016. p. 11–34. 10.21775/9781910190371.02 [DOI] [Google Scholar]
- Redero M, López-Causapé C, Aznar J, Oliver A, Blázquez J, Prieto AI. Susceptibility to R-pyocins of Pseudomonas aeruginosa clinical isolates from cystic fibrosis patients. J Antimicrob Chemother. 2018. October 01;73(10):2770–2776. 10.1093/jac/dky261 [DOI] [PubMed] [Google Scholar]
- Riboulet-Bisson E, Sturme MHJ, Jeffery IB, O’Donnell MM, Neville BA, Forde BM, Claesson MJ, Harris H, Gardiner GE, Casey PG, et al. Effect of Lactobacillus salivarius bacteriocin Abp118 on the mouse and pig intestinal microbiota. PLoS One. 2012. February 17;7(2):e31113. 10.1371/journal.pone.0031113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rihakova J, Cappelier JM, Hue I, Demnerova K, Fédérighi M, Prévost H, Drider D. In vivo activities of recombinant divercin V41 and its structural variants against Listeria monocytogenes. Antimicrob Agents Chemother. 2010. January;54(1):563–564. 10.1128/AAC.00765-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riley MA. Bacteriocins, biology, ecology, and evolution. In: Schaechter M, editor. Encyclopedia of Microbiology. Cambridge (USA): Academic Press; 2009. p. 32–44. 10.1016/B978-012373944-5.00065-1 [DOI] [Google Scholar]
- Rodrigues G, Silva GGO, Buccini DF, Duque HM, Dias SC, Franco OL. Bacterial proteinaceous compounds with multiple activities toward cancers and microbial infection. Front Microbiol. 2019. August 6;10:1690. 10.3389/fmicb.2019.01690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roh E, Park TH, Kim M, Lee S, Ryu S, Oh CS, Rhee S, Kim DH, Park BS, Heu S. Characterization of a new bacteriocin, Carocin D, from Pectobacterium carotovorum subsp. carotovorum Pcc21. Appl Environ Microbiol. 2010. November 15;76(22):7541–7549. 10.1128/AEM.03103-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rooney WM, Grinter RW, Correia A, Parkhill J, Walker DC, Milner JJ. Engineering bacteriocin‐mediated resistance against the plant pathogen Pseudomonas syringae. Plant Biotechnol J. 2020. May; 18(5):1296–1306. 10.1111/pbi.13294 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russel J, Røder HL, Madsen JS, Burmølle M, Sørensen SJ. Antagonism correlates with metabolic similarity in diverse bacteria. Proc Natl Acad Sci USA. 2017. October 03;114(40):10684–10688. 10.1073/pnas.1706016114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salazar-Marroquín EL, Galán-Wong LJ, Moreno-Medina VR, Reyes-López MÁ, Pereyra-Alférez B. Bacteriocins synthesized by Bacillus thuringiensis: generalities and potential applications. Rev Med Microbiol. 2016. July;27(3):95–101. 10.1097/MRM.0000000000000076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salgado PR, Ortiz CM, Musso YS, Di Giorgio L, Mauri AN. Edible films and coatings containing bioactives. Curr Opin Food Sci. 2015. October;5:86–92. 10.1016/j.cofs.2015.09.004 [DOI] [Google Scholar]
- Salvucci E, Saavedra L, Hebert EM, Haro C, Sesma F. Enterocin CRL35 inhibits Listeria monocytogenes in a murine model. Foodborne Pathog Dis. 2012. January;9(1):68–74. 10.1089/fpd.2011.0972 [DOI] [PubMed] [Google Scholar]
- Sánchez-Hidalgo M, Montalbán-López M, Cebrián R, Valdivia E, Martínez-Bueno M, Maqueda M. AS-48 bacteriocin: close to perfection. Cell Mol Life Sci. 2011. September;68(17):2845–2857. 10.1007/s00018-011-0724-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sand SL, Nissen-Meyer J, Sand O, Haug TM. Plantaricin A, a cationic peptide produced by Lactobacillus plantarum, permeabilizes eukaryotic cell membranes by a mechanism dependent on negative surface charge linked to glycosylated membrane proteins. Biochim Biophys Acta (BBA) – Biomembranes. 2013. February;1828(2):249–259. 10.1016/j.bbamem.2012.11.001 [DOI] [PubMed] [Google Scholar]
- Sarantinopoulos P, Leroy F, Leontopoulou E, Georgalaki MD, Kalantzopoulos G, Tsakalidou E, Vuyst LD. Bacteriocin production by Enterococcus faecium FAIR-E 198 in view of its application as adjunct starter in Greek Feta cheese making. Int J Food Microbiol. 2002. January;72(1–2):125–136. 10.1016/S0168-1605(01)00633-X [DOI] [PubMed] [Google Scholar]
- Sarika AR, Lipton AP, Aishwarya MS. Biopreservative efficacy of bacteriocin gp1 of Lactobacillus rhamnosus gp1 on stored fish filets. Front Nutr. 2019. March 22;6:29. 10.3389/fnut.2019.00029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider T, Hahn-Löbmann S, Stephan A, Schulz S, Giritch A, Naumann M, Kleinschmidt M, Tusé D, Gleba Y. Plant-made Salmonella bacteriocins salmocins for control of Salmonella pathovars. Sci Rep. 2018. December;8(1):4078. 10.1038/s41598-018-22465-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholz R, Vater J, Budiharjo A, Wang Z, He Y, Dietel K, Schwecke T, Herfort S, Lasch P, Borriss R. Amylocyclicin, a novel circular bacteriocin produced by Bacillus amyloliquefaciens FZB42. J Bacteriol. 2014. May 15;196(10):1842–1852. 10.1128/JB.01474-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shanker E, Federle M. Quorum sensing regulation of competence and bacteriocins in Streptococcus pneumoniae and mutans. Genes (Basel). 2017. January 05;8(1):15. 10.3390/genes8010015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi F, Wang Y, Li Y, Wang X. Mode of action of leucocin K7 produced by Leuconostoc mesenteroides K7 against Listeria monocytogenes and its potential in milk preservation. Biotechnol Lett. 2016. September;38(9):1551–1557. 10.1007/s10529-016-2127-y [DOI] [PubMed] [Google Scholar]
- Shin JM, Gwak JW, Kamarajan P, Fenno JC, Rickard AH, Kapila YL. Biomedical applications of nisin. J Appl Microbiol. 2016. June;120(6):1449–1465. 10.1111/jam.13033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva CCG, Silva SPM, Ribeiro SC. Application of bacteriocins and protective cultures in dairy food preservation. Front Microbiol. 2018. April 9;9:594. 10.3389/fmicb.2018.00594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simons A, Alhanout K, Duval RE. Bacteriocins, antimicrobial peptides from bacterial origin: overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms. 2020. April 27;8(5):639. 10.3390/microorganisms8050639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sindhu SS, Sehrawat A, Sharma R, Dahiya A. Biopesticides: use of rhizosphere bacteria for biological control of plant pathogens. Def Life Sci J. 2016. October 07;1(2):135–148. 10.14429/dlsj.1.10747 [DOI] [Google Scholar]
- Soltani S, Hammami R, Cotter PD, Rebuffat S, Said LB, Gaudreau H, Bédard F, Biron E, Drider D, Fliss I. 2021. Bacteriocins as a new generation of antimicrobials: toxicity aspects and regulations. EMS Microbiol Rev. 2021. January 8;45(1):fuaa039. 10.1093/femsre/fuaa039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoyanova LG, Ustyugova EA, Netrusov AI. Antibacterial metabolites of lactic acid bacteria: their diversity and properties. Appl Biochem Microbiol. 2012. May;48(3):229–243. 10.1134/S0003683812030143 [DOI] [PubMed] [Google Scholar]
- Subramanian S, Smith DL. Bacteriocins from the rhizosphere microbiome – from an agriculture perspective. Front Plant Sci. 2015. October 30;6:909. 10.3389/fpls.2015.00909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Z, Wang X, Zhang X, Wu H, Zou Y, Li P, Sun C, Xu W, Liu F, Wang D. Class III bacteriocin Helveticin-M causes sublethal damage on target cells through impairment of cell wall and membrane. J Ind Microbiol Biotechnol. 2018. March 01;45(3):213–227. 10.1007/s10295-018-2008-6 [DOI] [PubMed] [Google Scholar]
- Teneva-Angelova T, Hristova I, Pavlov A, Beshkova D. Chapter 4 – Lactic acid bacteria – From nature through food to health. In: Holban AM, Grumezescu AM, editors. Handbook of Food Bioengineering, Advances in Biotechnology for Food Industry. Cambridge (USA): Academic Press; 2018. p. 91–133. 10.1016/B978-0-12-811443-8.00004-9 [DOI] [Google Scholar]
- Todorov SD. Bacteriocin production by Lactobacillus plantarum AMA-K isolated from Amasi, a Zimbabwean fermented milk product and study of the adsorption of bacteriocin AMA-K to Listeria sp. Braz J Microbiol. 2008. March;39(1):178–187. 10.1590/S1517-83822008000100035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turovskiy Y, Ludescher RD, Aroutcheva AA, Faro S, Chikindas ML. Lactocin 160, a bacteriocin produced by vaginal Lactobacillus rhamnosus, targets cytoplasmic membranes of the vaginal pathogen, Gardnerella vaginalis. Probiotics Antimicrob Proteins. 2009. June;1(1):67–74. 10.1007/s12602-008-9003-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Um S, Kim YJ, Kwon H, Wen H, Kim SH, Kwon HC, Park S, Shin J, Oh DC. Sungsanpin, a lasso peptide from a deep-sea streptomycete. J Nat Prod. 2013. May 24;76(5):873–879. 10.1021/np300902g [DOI] [PubMed] [Google Scholar]
- van Staden ADP, Heunis T, Smith C, Deane S, Dicks LMT. Efficacy of lantibiotic treatment of Staphylococcus aureus-induced skin infections, monitored by in vivo bioluminescent imaging. Antimicrob Agents Chemother. 2016. July;60(7):3948–3955. 10.1128/AAC.02938-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veening JW, Blokesch M. Interbacterial predation as a strategy for DNA acquisition in naturally competent bacteria. Nat Rev Microbiol. 2017. October;15(10):621–629. 10.1038/nrmicro.2017.66 [DOI] [PubMed] [Google Scholar]
- Vijay Simha B, Sood SK, Kumariya R, Garsa AK. Simple and rapid purification of pediocin PA-1 from Pediococcus pentosaceous NCDC 273 suitable for industrial application. Microbiol Res. 2012. October;167(9):544–549. 10.1016/j.micres.2012.01.001 [DOI] [PubMed] [Google Scholar]
- Villarante KI, Elegado FB, Iwatani S, Zendo T, Sonomoto K, de Guzman EE. Purification, characterization and in vitro cytotoxicity of the bacteriocin from Pediococcus acidilactici K2a2–3 against human colon adenocarcinoma (HT29) and human cervical carcinoma (HeLa) cells. World J Microbiol Biotechnol. 2011. April; 27(4):975–980. 10.1007/s11274-010-0541-1 [DOI] [Google Scholar]
- Vivarelli S, Salemi R, Candido S, Falzone L, Santagati M, Stefani S, Torino F, Banna GL, Tonini G, Libra M. Gut microbiota and cancer: from pathogenesis to therapy. Cancers (Basel). 2019. January 03;11(1):38. 10.3390/cancers11010038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang SC, Lin CH, Sung CT, Fang JY. Antibacterial activities of bacteriocins: application in foods and pharmaceuticals. Front Microbiol. 2014. May 26;5:241. 10.3389/fmicb.2014.00241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao GW, Duarte I, Le TT, Carmody L, LiPuma JJ, Young R, Gonzalez CF. A broad-host-range tailocin from Burkholderia cenocepacia. Appl Environ Microbiol. 2017. May 15;83(10):e03414-16. 10.1128/AEM.03414-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zelezetsky I, Tossi A. Alpha-helical antimicrobial peptides – using a sequence template to guide structure-activity relationship studies. Biochim Biophys Acta (BBA) – Biomembranes. 2006. September;1758(9): 1436–1449. 10.1016/j.bbamem.2006.03.021 [DOI] [PubMed] [Google Scholar]
- Zhang ZF, Kim IH. Effects of multistrain probiotics on growth performance, apparent ileal nutrient digestibility, blood characteristics, cecal microbial shedding, and excreta odor contents in broilers. Poult Sci. 2014. February;93(2):364–370. 10.3382/ps.2013-03314 [DOI] [PubMed] [Google Scholar]
- Zhou W, Wang G, Wang C, Ren F, Hao Y. Both IIC and IID components of mannose phosphotransferase system are involved in the specific recognition between immunity protein PedB and bacteriocin-receptor complex. PLoS One. 2016. October 24;11(10):e0164973. 10.1371/journal.pone.0164973 [DOI] [PMC free article] [PubMed] [Google Scholar]