

Abstract
The SARS-CoV-2 pandemic is a major global threat that sparked global research efforts. Pre-clinical and biochemical SARS-CoV-2 studies firstly rely on cell culture experiments where the importance of choosing an appropriate cell culture model is often underestimated. We here present a bottom-up approach to identify suitable permissive cancer cell lines for drug screening and virus research. Human cancer cell lines were screened for the SARS-CoV-2 cellular entry factors ACE2 and TMPRSS2 based on RNA-seq data of the Cancer Cell Line Encyclopedia (CCLE). However, experimentally testing permissiveness towards SARS-CoV-2 infection, we found limited correlation between receptor expression and permissiveness. This underlines that permissiveness of cells towards viral infection is determined not only by the presence of entry receptors but is defined by the availability of cellular resources, intrinsic immunity, and apoptosis. Aside from established cell culture infection models CACO-2 and CALU-3, three highly permissive human cell lines, colon cancer cell lines CL-14 and CL-40 and the breast cancer cell line CAL-51 and several low permissive cell lines were identified. Cell lines were characterised in more detail offering a broader choice of non-overexpression in vitro infection models to the scientific community. For some cell lines a truncated ACE2 mRNA and missense variants in TMPRSS2 might hint at disturbed host susceptibility towards viral entry.
Introduction
It is of no debate that the overcoming of the pandemic SARS-CoV-2 pathogen spreading across the continents urgently needs joint efforts in the scientific community. Within a few months in the midst of spring 2020 the pandemic was accompanied with a high case fatality rate of up to 20% for vulnerable risk groups [1] and high excess mortality in Europe monitored by EuroMOMO (https://www.euromomo.eu/graphs-and-maps) [2]. High transmissibility of SARS-CoV-2 with R0 of ∼2.5 exceeds that of SARS-CoV, the 2009 influenza A H1N1 virus and even the 1918 influenza pandemic [3], probably due to its airborne transmission via droplets and aerosol [4–6]. During summer time, the portion of asymptomatic cases in a small Italian village were quantified to 43% [7] and since pre-/asymptomatic virus carriers were observed to be contagious, these are thought to drive the pandemic [8]. Re-infection also has been reported to complicate the pandemic course [9].
While various vaccines are rolled out [10–12], options for antiviral treatment for severely diseased patients are still sparse. Neutralising antibodies are promising drugs [13], while small molecules like remdesivir [14] have not yet proven great benefits [15, 16]. Importantly, all future antivirals, when becoming available and broadly used, face the challenge of SARS-CoV-2 escape mutations [17, 18]. Thus, it is clear that drug screening and basic research in SARS-CoV-2 is and will be of continuous importance.
Current investigations are mainly limited to the VERO-E6 African green monkey cell line, human lung cancer-derived CALU-3 and human colon carcinoma cell line CACO-2. Using non-human cells to study SARS-CoV-2 bears the inherent risk to draw conclusions from findings that result from species-specific cell properties. The anti-SARS-CoV-2 properties of hydroxychloroquine were found using VERO-E6, while later studies detected no effect in human cells and patients [19, 20]. In addition, the virus mutates rapidly when replicating in VERO-E6 [21–23]. Overexpression of host factors, such as TMPRSS2, can overcome individual species-specific artefacts but comes with commonly observed difficulties of stable transgene expression, such as antibiotic selection, non-physiological protein level, or transgene silencing during passaging.
Human cancer cell lines are easily available from public collections, characterised in great detail, and offer a great range of tissue origins, expression patterns, growth rates, and functional properties to choose from. They serve as valuable and valid model systems to study different diseases [24, 25] and have been used specifically for SARS-CoV-2 viral entry [26, 27] or for antiviral studies against this virus [28]. Particularly the two surface proteins ACE2 and TMPRSS2 have been shown to contribute to viral binding and processing [26, 27] and are mainly expressed in bronchial transient secretory cells [29]. Interestingly, high gene expression of ACE2 is shown for other tissues such as myocardial cells, esophagus epithelial cells, and enterocytes, which might be related to non-respiratory symptoms observed in COVID-19 patients [30–33]. Many virus-host-interaction studies rely on ACE2/TMPRSS2-transfected cell lines such as 293T and HELA cells or non-human material such as African green monkey cells VERO-E6 [26, 34, 35]. A recent study validated HPAEpiC as a human AT2 cell-derived pulmonary alveolar epithelial cell line to be susceptible to SARS-CoV-2 [36].
In this study, we have pre-screened and ranked human cancer cell lines of the Leibniz Institute German Collection of Microorganisms and Cell Cultures (DSMZ) for ACE2 and TMPRSS2 expression within the rich resource Cancer Cell Line Encyclopedia (CCLE) publicly available RNA-seq data [37]. Selected cell lines were then validated for mRNA and protein levels of ACE2 and TMPRSS2 and tested in SARS-CoV-2 infection experiments, revealing a set of cell lines displaying high and low permissiveness. With these cell lines we devise appropriate, non-overexpressing cell model systems to facilitate experimental SARS-CoV-2 work for the understanding of viral entry, viral replication and to support drug development.
Results
Selection of cell model systems
Inspired by the publication of Hoffmann et al. pinpointing the host factors ACE2 as SARS-CoV-2 receptor and TMPRSS2 as viral S protein priming serine protease [26], we defined expression of these two factors as pre-selection criteria in our study. Since the Cancer Cell Line Encyclopedia (CCLE) as a comprehensive database provides large-scale sequencing data including mRNA-seq expression data for over 900 cell lines [37], we screened for gene expression of ACE2 and TMPRSS2 in this data set, which enabled us to identify potential susceptible cell lines prone to SARS-CoV-2 virus entry. At first, we selected for 301 human cell lines readily in stock at the DSMZ cell lines repository within this data set and visualised gene expression of ACE2, which was shown to be the cellular receptor for the viral S protein [38], and TMPRSS2 known to cleave SARS-CoV S protein [39] (Fig 1A). Gene expression for these two genes differed between and within the various tumour entities. In order to pre-select cell lines to be tested for SARS-CoV-2 permissiveness, the top 25 highest ACE2 gene expressing cell lines were selected, originating from diverse tumour origins (Fig 1B). Importantly, our short-list included cancer cell lines originating from tissues that are known for SARS-CoV-2 replication in vivo [40], e.g. the tongue squamous cell carcinoma cell lines CAL-27, CAL-33, and the esophageal squamous cell carcinoma cell line KYSE-510. Based on this evidence, we additionally selected the newly established oral squamous cell carcinoma cell lines UPCI-SCC-074 and UPCI-SCC-131, which are not contained in the CCLE dataset. Futhermore, we included the neuronal cells DBTRG-05MG and KELLY with high levels of neuropilin 1 (NRP1) mRNA expression (Fig 1B), which has been discussed as receptor for the viral spike protein [41, 42]. NRP1 expression was most prominent in DBTRG-06MG, CAL-51, CAL-85–1, CALU-3, SCC-25, CL-40, and KELLY (Fig 1B). Since the lung carcinoma cell line CALU-3, the colorectal adenocarcinoma CACO-2, and the green monkey kidney cell line subclones VERO-E6 and VERO-B4 are shown to be permissive for SARS-CoV-2 [26, 27], these were used as positive control cell lines. Altogether we selected 29 human cancer cell lines and four control cell lines.
Fig 1. ACE2 and TMPRSS2 gene expression data for 301 DSMZ human cell lines in the CCLE RNA-seq data set.
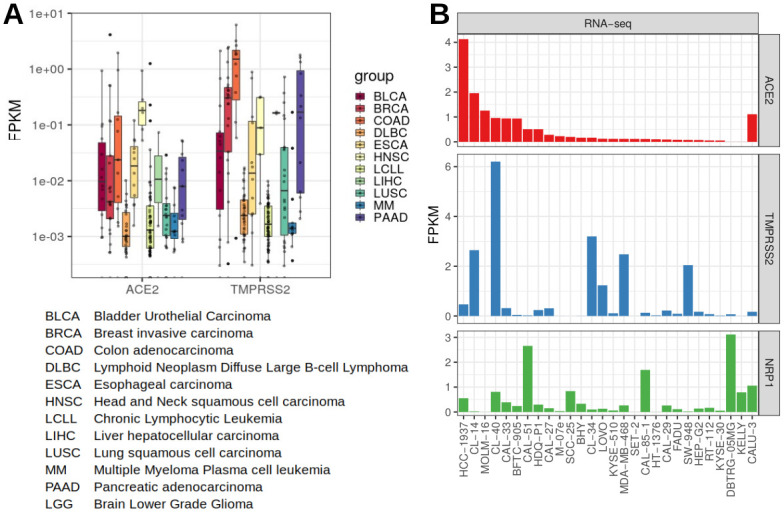
A: Expression of ACE2 and TMPRSS2 for selected disease entities. RNA-seq data were normalised and calculated to FPKM values. Gene expression for ACE2 and TMPRSS2 varies between and within the different tumour species. B: Expression of ACE2, TMPRSS2 and NRP1, cell lines sorted to ACE2 expression levels. Top 25 ACE2 expressing cell lines were selected for further studies, plus CALU-3 and two further neuronal cell lines with high NRP1 levels. FPKM—fragments per kilobase million: normalised gene expression data.
In order to validate ACE2 and TMPRSS2 expression levels, we performed qRT-PCR on the short-listed cell lines. All selected cell lines exhibited considerable ACE2 mRNA expression according to the RNA-seq data and/or qRT-PCR except for DBTRG-05MG and KELLY, whereas TMPRSS2 mRNA levels were particularly higher in CL-40, CL-34, CL-14, and SW-948 (Figs 1B and 2A). Apart from qualitative verification for mRNA expression via RNA-seq data and qRT-PCR (Fig 2A), protein expression of ACE2 and TMPRSS2 was confirmed via western blot for specific cell lines (Fig 2B, S1 Fig).
Fig 2. Verified presence of ACE2 and TMPRSS2 in carcinoma cell lines determined by qRT-PCR and western blots.
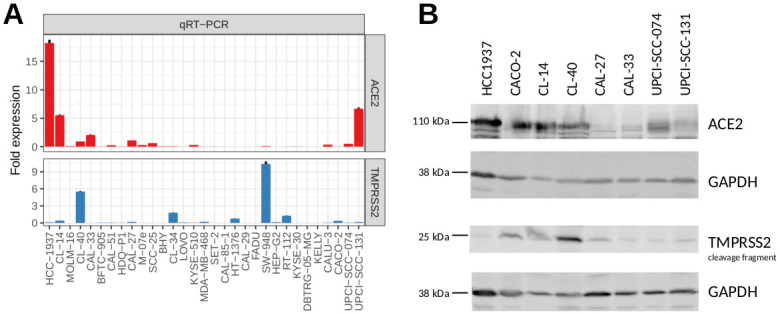
A: Quantification of ACE2 and TMPRSS2 transcripts in the selected candidate cell lines by qRT-PCR (normalised to CAL-27 and CL-40). B: Western blot analysis was performed to assess the expression of proteins associated with SARS-CoV-2 cell entry.
For expanding our cell line search spectrum, we interrogated the LL-100 cell line data set, comprising RNA-seq and whole-exome sequencing (WES) data of 100 human leukemia and lymphoma cell lines [24]. However, none of these cell lines showed considerable expression of ACE2 and TMPRSS2. This is reflected in the CCLE results, in which cell lines of the hematopoetic lineages exhibited lower gene expression of both genes compared to other cell lines of different origin (see Fig 1A DLBC, LCLL and MM tumour entities).
The presence of the entry factors ACE2 and TMPRSS2 in cancer cell lines of various origins is in accordance with their expression in corresponding primary tissue, where ACE2 and TMPRSS2 were detected in cells from multiple tissues including respiratory tract, esophagus and colon [31, 33, 43–45] and virus entry was evidenced in gastro-intestinal tissues [46], organs of the respiratory tract and various other tissues [40].
Testing for SARS-CoV-2 infectivity and permissiveness
Receptor ACE2 and surface proteinase TMPRSS2 have been shown to play a key role for SARS-CoV-2 cellular entry, however, whether their co-expression is sufficient to allow entry and whether alternative receptors may exist, is still under debate [45]. In order to shed light on infectivity and permissiveness of the appointed cell lines in context of their ACE2/TMPRSS2 expression, we subjected cells to experimental infection with wildtype SARS-CoV-2. Permissiveness to viral infection is defined as the ability of a given host cell to permit the entire viral replication cycle and to eventually release mature virions. Thus, we sampled the supernatant of the short-listed cell lines at four consecutive days following infection with a low amount of inoculum to sensitively detect viral replication.
Within our short-listed panel, we identified large variation in the degree of permissiveness and virus replication kinetics. Surprisingly, the majority of tested cell lines (15 of 29) showed no detectable virus replication despite the presence of ACE2 and TMPRSS2 (Fig 3A, S2 Fig). Further 11 cell lines showed detectable SARS-CoV-2 infectious titres that did not exceed the amount of the inoculum. Only three cell lines were categorized highly permissive, amplifying the initial virus inoculum by factors of ∼2 to ∼500 between 1 and 4 dpi. These were the colon carcinoma cell lines CL-14 and CL-40, as well as breast carcinoma cell line CAL-51. Infectious titres of SARS-CoV-2 grown on CL-14 and on CAL-51 were concurrent with the widely used CALU-3 (lung carcinoma), CACO-2 (colon carcinoma), VERO-E6 and VERO-B4 (normal African green monkey kidney cells) (Fig 3B). Whereas CL-14 already has been described to be permissive in a preprint [47], the breast carcinoma cell line CAL-51 with high levels of NRP1 beside ACE2 and TMPRSS2 expression and the colon carcinoma cell line CL-40 have not been identified before. Despite comparable peak titres, virus replication kinetics differed largely between cell lines, where CAL-51 and CACO-2 showed increase in titres throughout day three postinfection while VERO-E6, VERO-B4, and CL-14 produced highest levels within 24h, while the viral titer for CL-40 remained close to the level of the initial inoculum of SARS-CoV-2 (Fig 3, Table 1).
Fig 3. Susceptibility of tumour cell lines toward SARS-CoV-2 infection.

Virus titers in the supernatant of infected cell lines at 1–4 days postinfection (dpi) determined by plaque assay for the top 10 ACE2 expressing cell lines (A) and controls (B). The red dotted line indicates the initial inoculum (2000 pfu/ml); the blue dashed line specifies the minimal limit of detection LOD 33.3 pfu/ml. Number of replicates varied between 3–8. pfu: plaque-forming units; mean values with SEM. (C) Predominant productive virus production in highly permissive cell lines seen in the correlation between qRT-PCR measured viral nucleoprotein N mRNA and releasing virions tested by viral plaques. Eight replicates served for the means of the viral titers and one representative expression fold change of two biological replicates qRT-PCR with three technical replicates. Note the logarithmic scaling for both x and y-axis.
Table 1. Permissive cell line characteristics.
| cell line | Species | Sex | Tissue | Peak viral load, dpi | Maximal peak, pfu/ml | Doubling time |
|---|---|---|---|---|---|---|
| CL-14 | human | male | colon | 1 | 1.1 x 105 | ∼1w |
| CL-40 | human | female | colon | 2 | 5.8 x 103 | ∼3–4d |
| CAL-51 | human | female | breast | 3 | 1.6 x 106 | ∼30h* |
| CALU-3 | human | male | lung | 3 | 1.6 x 106 | ∼84h* |
| CACO-2 | human | male | colon | 4 | 3.0 x 105 | 80h |
| VERO-B4 | monkey | female | kidney | 2 | 1.4 x 105 | ∼25h |
| VERO-E6 | monkey | female | kidney | 4 | 1.1 x 106 | 22h+ |
Peak viral load and maximal peak were determined during this study.
Species, sex, tissue and doubling time were taken from https://www.dsmz.de/collection/catalogue/human-and-animal-cell-lines/catalogue.
*own observation;
+ATCC.
Therapeutic neutralising antibodies bind to the viral Spike protein and prevent the interaction with ACE2 [13]. However, it is unclear whether different levels of ACE2 expression on the surface of cells would interfere with the ability to block virus entry. Thus, we used three different permissive cell lines of our panel (CL-14, CAL-51, and VERO-E6) with varying ACE2 levels and performed in vitro neutralisation assays using the therapeutic anti-Spike antibody COR-101 currently used in clinical trials (see S3 Fig) [48]. While the comparability among these cell lines is limited due to several variables (cell density, virus growth kinetic, cell medium, etc.), we observed ≥93% inhibition of virus entry at 100 ng/ml in all three cell lines, compared to the lowest nAb concentration. The inhibition curves suggest a reduced neutralisation efficiency in the CAL-51 breast cancer cell line, which might be attributed to NRP-1-mediated or NRP-1-assisted virus entry [41, 42]. NRP-1 is an alternative or assisting SARS-CoV-2 entry factor that shows elevated levels in CAL-51 cells (see Fig 1A). However, the complete virus neutralisation at sufficient antibody levels suggests either the absence of ACE2-independent entry or the additional inhibition of a Spike/NRP-1 interaction.
In contrast, low-permissive cell lines, such as the breast cancer HCC-1937, the colon adenocarcinoma CL-34, and esophageal squamous cell carcinoma KYSE-30 cell lines showed slow replication kinetics, which was also true for the oral squamous cell carcinoma cell lines UPCI-SCC-074 and UPCI-SCC-131 but not for the neuronal cells DBTRG-05MG and KELLY despite high levels of NRP1 (Fig 3A, S2 Fig). Non-permissive cell lines showed no detectable SARS-CoV-2 titres at any time after infection.
In order to test for abortive viral cycles particularly in the non-productive virus releasing cell lines, we compared mRNA levels of the nucleoprotein in each three control, high-permissive, low-permissive, and non-permissive cell lines to viral plaques after 24 hpi (Fig 3C). We found a high correlation (R2 = 0.7) between viral expression and infective virus titers in the tested cell lines, probably reduced by high variation in viral infection of the cell lines at detection limit. Hence, these data argue for the absence of abortive virus replication or defects in releasing mature virions in the low-permissive cell lines.
Genomic analyses of ACE2 and TMPRSS2
Since we found limited concordance between ACE2/TMPRSS2 expression and virus permissiveness of the cell lines, we seeked for genomic differences in the receptors. Of note, gene expression of different splice variants of the ACE2 mRNA became evident comparing the read numbers of the exons as seen in the CCLE RNA-seq data for MOLM-16, CAL-33, BFTC-905, and CAL-27 (Fig 4A, S4 Fig, S1 Table). The truncated ACE2 lacking exon 1–9 has been reported recently [49, 50].
Fig 4. ACE2 and TMPRSS2 transcript variants for the cell lines.
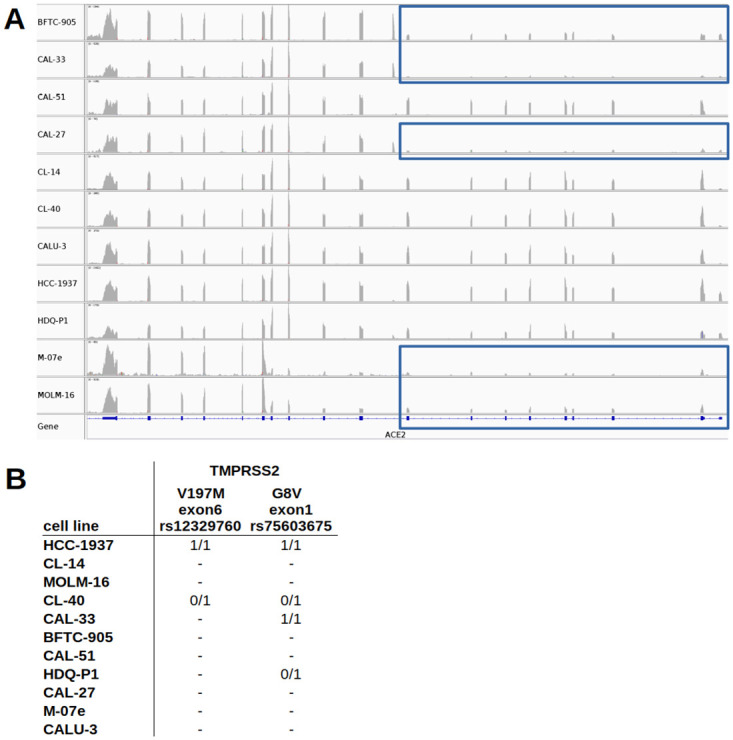
A: Different exon usage of ACE2 for the top 10 ACE2 expressing cell lines plus CALU-3 (see S2 Fig for the top 25 cells). BFTC-905, CAL-33, CAL-27, M-07e, and MOLM-16 show reduced exon expression from exon 1–9 (blue rectangles). Cell lines are ordered alphabetically. ACE2 is reversely oriented. B: Missense variants in the coding regions of TMPRSS2 (V197M = rs12329760, G6V = rs75603675) for the selected cell lines. No mutations were found in the coding regions of ACE2. 0/1 denotes heterozygous and 1/1 homozygous mutations.
Furthermore, missense variants were detected in TMPRSS2 for HCC-1937 (homozygous), CL-40 (heterozygous), CAL-33 (homozygous), and HDQ-P1 (heterozygous) (Fig 4B), namely V197M (V160M) in exon 6 (rs12329760) and/or G8V in exon 1 (rs75603675). We assume that these variations may have a considerable effect on the virus-host interaction.
Discussion
In this study, we have systematically identified and characterised SARS-CoV-2 permissive cancer cell lines that (except one) have not been published as such before. Of 29 selected human cell lines three tested high-level positive permissive for SARS-CoV-2. These selected cell lines are available from the German repository DSMZ. Cancer cell lines have been an integral tool for basic research and drug screening for decades and still are the preferred choice for the vast majority of virus research labs. For SARS-CoV-2 infection experiments, different replication kinetics as well as different degrees of permissiveness can aid answering specific questions and allow different experimental setups, e.g. in compound screening assays. We found permissiveness to SARS-CoV-2 in only 14 out of 29 cell lines pre-selected for the expression of the known entry factor ACE2 (Figs 1B and 2). Three of them replicated the virus to high titres (CL-14, CL-40, and CAL-51) and further 11 showed low levels with mainly slow virus kinetics compared to commonly used VERO-E6, CALU-3, and CACO-2 cells (Fig 3, S2 Fig, Table 1).
In order to understand the non-permissiveness of the majority of top 25 ACE2 expressing cell lines, we closely examined splice and single nucleotide variants. ACE2 splice variants at the N-terminus, containing eight amino acids critical for binding to the receptor binding domain (RBD) of the spike protein of SARS-CoV-2, are predicted to disturb SARS-CoV-2 attachment [51]. One splice variant lacking exon1-exon9 was observed in e.g. MOLM-16, CAL-33, and CAL-27 (Fig 4A, S4 Fig, S1 Table), cell lines that indeed were not shown to be non-permissive despite their pronounced ACE2 gene expression (Fig 1B). This truncated ACE2 isoform has been reported to be interferon inducible [49] and fails to bind to SARS-CoV-2 [50].
Whereas the lacking permissiveness for SARS-CoV-2 of the above mentioned cell lines is likely to be explained by a truncated ACE2 transcript (Fig 4A), it is more challenging to elucidate, why the candidate cell line CL-40 tested less permissive than CL-14 and CAL-51 despite high TMPRSS2 transcript levels. CL-40 as well as HCC-1937 hold a missense variation in exon 6 of TMRPSS2 (Fig 4B) causing an amino acid exchange at position 197 (V197M/V160M) within the Scavenger receptor cysteine-rich (SRCR) domain responsible for protein-protein interaction and ligand binding (ExPASy) and is described for various cancer types as well as cell lines (COSMIC). Hence, we speculate that interaction with SARS-CoV-2 might be disturbed by this variant at V197M, which is known for differential allelic frequencies in populations [52].
Importantly, the expression levels of ACE2, TMPRSS2 and NRP1 are not predictive of SARS-CoV-2 permissiveness per se. While ACE2 seems to be an essential component, however, it is not sufficient on its own. Permissiveness includes the ability of the virus to enter the cell, replicate, and release infectious progeny. A number of cellular processes can effectively interfere with this replication cycle, including intrinsic immune response, type-I and type-III interferon responses and apoptosis induction [53] and thus determine permissiveness. For SARS-CoV-2, several essential host factors apart from surface receptors were recently discovered [54].
Meanwhile, several organs and cell types have been reported to be infected by SARS-CoV-2, even for those with no or low basal ACE2 expression [30, 45]. Hence, alternate receptors, proteases and antagonists for SARS-CoV-2 infection could widen the options for viral entry [45] as shown for cathepsin L [55] and CD147 [56]. Besides genetic and epigenetic variations, variable activation of ACE2 could also be triggered by interferon pathways [57, 58].
Seeking for SARS-CoV-2 permissive cell lines by screening ACE2 RNA-seq-derived expression levels, the human native breast carcinoma cell line CAL-51 and the colon adenocarcinoma cell lines CL-40 and CL-14 have been identified to produce high levels of infectious SARS-CoV-2 virus particles. CAL-51 in particular could enrich SARS-CoV-2 studies trifold: low doubling time of this cell line, viral stock production comparable to commonly used VERO-E6, and a further organ tissue as infection model. Its doubling time is lower than that of the colon carcinoma cell lines CACO-2/CL-14 and the lung carcinoma cell line CALU-3 (Table 1), avoiding the mutation selection pressure to adapt to another species host as seen for the African green monkey cell line VERO-E6 [21–23]. Different replication kinetics as well as different degrees of permissiveness can aid answering specific scientific questions and allow different experimental setups, e.g. in compound screening assays.
In summary, we find that the expression of ACE2 and TMPRSS2 are indicative but not sufficient for SARS-CoV-2 productive infection in human cell lines, probably partly due to a truncated ACE2 splice variant and TMPRSS2 variants. Out of 29 tested cancer cell lines, we have identified three highly and 11 lowly permissive cell lines that vastly broaden the choice for SARS-CoV-2 cell culture models. The breast carcinoma cell line CAL-51 in particular may serve as a suitable cell line for SARS-CoV-2 studies for viral propagation because of its easy management.
Methods
CCLE RNA-seq data set analysis
RNA-seq data were retrieved and pre-processed as described [59]. Briefly, alignment files were downloaded from CGHub via genetorrent, sorted (samtools 0.1.19) and converted to fastq files (bedtools v2.21.0). Reads were trimmed via fastq-mcf (ea-utils 1.1.2–686). STAR (2.5.3a) served as read mapper to the human genome Gencode 26, HTSeq (0.11.3) was used as count tool. Raw counts were normalised and transformed to FPKM (fragments per kilobase million) via R/Bioconductor (3.6.9) loading DESeq2 (1.26.0) and visualised via ggplot2 (3.1.1). Different exon usage was discovered by visualising the alignment files via IGV (2.8.0) [60]. For detecting mutations, mapped reads were assigned to read groups (picard, 2.9.2 [www.broadinstitute.github.io/picard]); reads were split, trimmed and reassigned (GenomeAnalysisTK, 3.7–0, SplitNCigarReads [61]); mutations were called by the HaplotypeCaller (GenomeAnalysisTK, 3.7–0,) [61]; and mutation effects were predicted via the Ensembl VEP (GRCh38, v90) [62]. Mutations were filtered to ≥10 depth.
Cultivation of human cell lines
The continuous cell lines are part of the cell culture collection at DSMZ (https://www.dsmz.de/collection/catalogue/human-and-animal-cell-lines). Tissue origin, doubling-time, growth properties, cytogenetics and more information on these cell lines can be retrieved from the DSMZ website. Cell lines were grown at 37°C in a humidified atmosphere of air containing 5% CO2. The basic growth media (Life Technologies, Darmstadt, Germany) were supplemented with 10–20% fetal bovine serum (Sigma Aldrich, Taufkirchen, Germany). No antibiotics were added to the cultures. All cell lines were free of mycoplasma and other bacterial, yeast and fungi contaminations as tested by PCR and microbiological growth assays [63]. The authenticity of the cell lines was determined by DNA typing [64]. The cells were first cultured in cell culture flasks to confluence, detached with trypsin/EDTA, washed with PBS and pelleted for RNA and protein preparation. For virus infection experiments the cells were seeded in 96-well plates at different densities starting with 0.25—1.5 x 105 cells per well as highest cell number depending on the cell line. Optimal cell densities were determined by observing the growth progression with an IncuCyte instrument (Essen Bioscience, Hertfordshire, UK). Cells were then seeded at semi-confluence or complete confluence and three further dilutions each with half of the previous cell densities. The cells were grown for one day before the virus infection experiments were started.
qRT-PCR analysis
Quantitative Real-time polymerase chain reaction (qRT-PCR) analysis for human mRNA was performed to quantify specific transcripts in the cells. Total RNA was extracted from cell line samples using TRIzol reagent (Fisher Scientific, Schwerte, Germany). cDNA was generated from 1 μg RNA by random priming using the Biozym cDNA Synthesis Kit (Biozym, Hessisch Oldendorf, Germany). qRT-PCR analysis was performed with the 7500 Real-time System, using commercial buffer and primer sets for the amplification of ACE2 (Hs01085333_m1) and TMPRSS2 (Hs01122322_m1) transcripts (Thermo Fisher Scientific, Darmstadt, Germany).
Viral RNA of infected cell cultures was extracted using spin filter membranes (innuPREP Virus TS RNA Kit from Analytik Jena) according to the manufacturer’s recommendations. Briefly, cells were lysed in the presence of carrier RNA and Proteinase K at 70°C for 10 min. Nucleic acids were then bound to the surface of the spin filter membranes, washed and subsequently eluted with RNase-free water. One to five μg RNA were then reverse transcribed to cDNA applying random hexamers. The primers for the qRT-PCR for the amplification of the SARS-CoV-2 N gene were N_Sarbeco_F (5´-CACATTGGCACCCGCAATC-3’) and N_Sarbeco_R (5’-GAGGAACGAGAAGAGGCTTG-3’) [65].
We analysed the transcript of TATA box binding protein (TBP) for normalisation of expression levels using SYBR Green for quantification. Quantitative analyses were performed in triplicate, if not indicated differently, analysed and visualised with R/Bioconductor packages ddCt and ggplot2.
Western blots
Antibodies against ACE2 (ab15348), TMPRSS2 (ab109131), and GAPDH (ab8245) were obtained from Abcam (Cambridge, UK). Western blot samples were prepared as described previously [66]. Proteins on nitrocellulose membranes were visualized with the biotin/streptavidin-horseradish peroxidase system (GE Healthcare; Little Chalfont, UK) in combination with the “Renaissance Western Blot Chemoluminescence Reagent” (Perkin Elmer; Waltham, MA, USA). Documentation was performed using the digital system ChemoStar Imager (INTAS, Göttingen, Germany).
Viral infection of cell lines
Infection with SARS-CoV-2 was performed with virus strain Zagreb (SARS-CoV2/ZG/297–20, University Hospital for Infectious Diseases, Zagreb, Croatia). If not indicated otherwise, cells were seeded onto 96-well cell culture plates and infected 24–48 hours later with 250 pfu/well using a 10 μl inoculum and 100 μl total well volume.
Virus plaque and neutralisation assays
Quantification of SARS-CoV-2 infectious units was done via a virus plaque assay. Supernatants of infected cells were taken 1–4 dpi, serially diluted, and used to infect confluent VERO-E6 cells on 96-well cell culture plates for one hour. Then, the inoculum was removed and cells were overlaid with cell culture medium (MEM, 10% FCS, 2 mM glutamine) containing 1.5% methyl-cellulose. After 3 days, virus plaques were counted from phase contrast microscopic images taken with a Sartorius IncuCyte S3 at 4x magnification.
For neutralisation assays antibody COR-101 and virus were mixed and incubated for 1h at 37°C and applied to cells for 24 hpi. COR-101 for was a kind gift of Corat Therapeutics GmbH (Braunschweig, Germany).
Supporting information
Original western blots to cropped western blot images in Fig 2B.
(PDF)
Virus titers in the supernatant of infected cell lines at 1-4 dpi determined by plaque assay for the top 11-25 ACE2 expressing, two neuronal (DBTRG-05MG, KELLY), and two oral squamous cell carcinoma (UPCI-SCC-074, UPCI-SCC-131) cell lines. The red dotted line indicates the initial inoculum (2000 pfu/ml); the blue dashed line specifies the minimal limit of detection LOD 33.3 pfu/ml. Number of replicates varied between 3-8. pfu: plaque-forming units; mean values with SEM.
(TIFF)
Neutralising SARS-CoV-2 antibody was applied to the high permissive cell lines CAL-51 and CL-14 and the control cell line VERO-E6. With increasing antibody concentration viral production is decreasing in all three cell lines. The red dotted line indicates the initial inoculum (2000 pfu/ml); the blue dashed line specifies the minimal limit of detection LOD 33.3 pfu/ml. Note the logarithmic scaling for the x-axis.
(TIF)
Different exon usage of ACE2 for the top 25 ACE2 expressing cell lines plus CALU-3. RNA-seq data were visualised via IGV and cell lines ordered alphabetically.
(TIFF)
Gene expression, splice and single nucleotide variants of ACE2 and TMPRSS2 in the cell lines used in this study. These were identified on the basis of CCLE RNA-seq data (FPKM, see Material and methods). +truncated splice variant observed; 0/1 heterozygous, 1/1 homozygous mutation.
(ODS)
Acknowledgments
We thank Hilmar Quentmeier for the initial coordination of activities for this project, Klaus Steube for reviewing the manuscript, and Sonja Eberth for useful reference hints.
Data Availability
All relevant data are within the paper and its Supporting information files.
Funding Statement
This work was supported by grants from the Helmholtz Gemeinschaft (EU partnership grant PIE-0008) and the German Ministry of Science and Education (BMBF grant 01KI20140A) to Luka Cicin-Sain.
References
- 1. Onder G, Rezza G, Brusaferro S. Case-fatality rate and characteristics of patients dying in relation to COVID-19 in Italy. JAMA. 2020;323(18):1775–1776. [DOI] [PubMed] [Google Scholar]
- 2. Kanieff M, Rago G, Minelli G, Lamagni T, Sadicova O, Selb J, et al. The potential for a concerted system for the rapid monitoring of excess mortality throughout Europe. Euro Surveill. 2010;15(43):19697. doi: 10.2807/ese.15.43.19697-en [DOI] [PubMed] [Google Scholar]
- 3. Petersen E, Koopmans M, Go U, Hamer DH, Petrosillo N, Castelli F, et al. Comparing SARS-CoV-2 with SARS-CoV and influenza pandemics. Lancet Infect Dis. 2020;20(9):e238–e244. doi: 10.1016/S1473-3099(20)30484-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. van Doremalen N, Bushmaker T, Morris DH, Holbrook MG, Gamble A, Williamson BN, et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N Engl J Med. 2020;382(16):1564–1567. doi: 10.1056/NEJMc2004973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Tang S, Mao Y, Jones RM, Tan Q, Ji JS, Li N, et al. Aerosol transmission of SARS-CoV-2? Evidence, prevention and control. Environ Int. 2020;144:106039. doi: 10.1016/j.envint.2020.106039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Fears AC, Klimstra WB, Duprex P, Hartman A, Weaver SC, Plante KS, et al. Persistence of Severe Acute Respiratory Syndrome Coronavirus 2 in Aerosol Suspensions. Emerg Infect Dis. 2020;26(9):2168–2171. doi: 10.3201/eid2609.201806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Lavezzo E, Franchin E, Ciavarella C, Cuomo-Dannenburg G, Barzon L, Del Vecchio C, et al. Suppression of a SARS-CoV-2 outbreak in the Italian municipality of Vo’. Nature. 2020;584(7821):425–429. doi: 10.1038/s41586-020-2488-1 [DOI] [PubMed] [Google Scholar]
- 8. Nikolai LA, Meyer CG, Kremsner PG, Velavan TP. Asymptomatic SARS Coronavirus 2 infection: Invisible yet invincible. Int J Infect Dis. 2020;100:112–116. doi: 10.1016/j.ijid.2020.08.076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Cohen JI, Burbelo PD. Reinfection with SARS-CoV-2: Implications for Vaccines. Clin Infect Dis. 2020. doi: 10.1093/cid/ciaa1866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Tu YF, Chien CS, Yarmishyn AA, Lin YY, Luo YH, Lin YT, et al. A review of SARS-CoV-2 and the ongoing clinical trials. Int J Mol Sci. 2020;21(7):2657. doi: 10.3390/ijms21072657 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Jackson LA, Anderson EJ, Rouphael NG, Roberts PC, Makhene M, Coler RN, et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N Engl J Med. 2020;383(20):1920–1931. doi: 10.1056/NEJMoa2022483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Mulligan MJ, Lyke KE, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature. 2020;586(7830):589–593. doi: 10.1038/s41586-020-2639-4 [DOI] [PubMed] [Google Scholar]
- 13. Zhou G, Zhao Q. Perspectives on therapeutic neutralizing antibodies against the Novel Coronavirus SARS-CoV-2. Int J Biol Sci. 2020;16(10):1718–1723. doi: 10.7150/ijbs.45123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Gordon CJ, Tchesnokov EP, Woolner E, Perry JK, Feng JY, Porter DP, et al. Remdesivir is a direct-acting antiviral that inhibits RNA-dependent RNA polymerase from severe acute respiratory syndrome coronavirus 2 with high potency. J Biol Chem. 2020;295(20):6785–6797. doi: 10.1074/jbc.RA120.013679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Wang Y, Zhang D, Du G, Du R, Zhao J, Jin Y, et al. Remdesivir in adults with severe COVID-19: a randomised, double-blind, placebo-controlled, multicentre trial. Lancet. 2020;395(10236):1569–1578. doi: 10.1016/S0140-6736(20)31022-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Goldman JD, Lye DCB, Hui DS, Marks KM, Bruno R, Montejano R, et al. Remdesivir for 5 or 10 Days in Patients with Severe Covid-19. N Engl J Med. 2020;383(19):1827–1837. doi: 10.1056/NEJMoa2015301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. McCallum M, De Marco A, Lempp F, Tortorici MA, Pinto D, Walls AC, et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. bioRxiv 2021.01.14.426475. Available from: https://www.biorxiv.org/content/10.1101/2021.01.14.426475v1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Wang Z, Schmidt F, Weisblum Y, Muecksch F, Barnes CO, Finkin S, et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature. 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Hoffmann M, Mösbauer K, Hofmann-Winkler H, Kaul A, Kleine-Weber H, Krüger N, et al. Chloroquine does not inhibit infection of human lung cells with SARS-CoV-2. Nature. 2020;585(7826):588–590. doi: 10.1038/s41586-020-2575-3 [DOI] [PubMed] [Google Scholar]
- 20. Boulware DR, Pullen MF, Bangdiwala AS, Pastick KA, Lofgren SM, Okafor EC, et al. A Randomized Trial of Hydroxychloroquine as Postexposure Prophylaxis for Covid-19. N Engl J Med. 2020;383(6):517–525. doi: 10.1056/NEJMoa2016638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Ogando NS, Dalebout TJ, Zevenhoven-Dobbe JC, Limpens RWAL, van der Meer Y, Caly L, et al. SARS-coronavirus-2 replication in Vero E6 cells: replication kinetics, rapid adaptation and cytopathology. J Gen Virol. 2020;101:925–940. doi: 10.1099/jgv.0.001453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Sasaki M, Uemura K, Sato A, Toba S, Sanaki T, Maenaka K, et al. SARS-CoV-2 variants with mutations at the S1/S2 cleavage site are generated in vitro during propagation in TMPRSS2-deficient cells. PLoS Pathog. 2021;17(1):e1009233. doi: 10.1371/journal.ppat.1009233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Chaudhry MZ, Eschke K, Grashoff M, Abassi L, Kim Y, Brunotte L, et al. SARS-CoV-2 Quasispecies Mediate Rapid Virus Evolution and Adaptation. bioRxiv 2020.08.10.241414. Available from: https://www.biorxiv.org/content/10.1101/2020.08.10.241414v1 [Google Scholar]
- 24. Quentmeier H, Pommerenke C, Dirks WG, Eberth S, Koeppel M, MacLeod RAF, et al. The LL-100 panel: 100 cell lines for blood cancer studies. Sci Rep. 2019;9(1):8218. doi: 10.1038/s41598-019-44491-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. MacLeod RA, Nagel S, Scherr M, Pommerenke C, Koeppel M, Meyer C, et al. Human leukemia and lymphoma cell lines as models and resources. Curr Med Chem. 2008;15(4):339–359. doi: 10.2174/092986708783497319 [DOI] [PubMed] [Google Scholar]
- 26. Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181(2):271–280. doi: 10.1016/j.cell.2020.02.052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, Veesler D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell. 2020;181(2):281–292. doi: 10.1016/j.cell.2020.02.058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Sheahan TP, Sims AC, Zhou S, Graham RL, Pruijssers AJ, Agostini ML, et al. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci Transl Med. 2020;12(541):eabb5883. doi: 10.1126/scitranslmed.abb5883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Lukassen S, Chua RL, Trefzer T, Kahn NC, Schneider MA, Muley T, et al. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020;39(10):e105114. doi: 10.15252/embj.20105114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Zou X, Chen K, Zou J, Han P, Hao J, Han Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front Med. 2020;10(2):185–192. doi: 10.1007/s11684-020-0754-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Qi F, Qian S, Zhang S, Zhang Z. Single cell RNA sequencing of 13 human tissues identify cell types and receptors of human coronaviruses. Biochem Biophys Res Commun. 2020;526(1):135–140. doi: 10.1016/j.bbrc.2020.03.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Cao Y, Liu X, Xiong L, Cai K. Imaging and clinical features of patients with 2019 novel coronavirus SARS-CoV-2: A systematic review and meta-analysis. J Med Virol. 2020;92(9):1449–1459. doi: 10.1002/jmv.25822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Lamers MM, Beumer J, van der Vaart J, Knoops K, Puschhof J, Breugem TI, et al. SARS-CoV-2 productively infects human gut enterocytes. Science. 2020;369(6499):50–54. doi: 10.1126/science.abc1669 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Shulla A, Heald-Sargent T, Subramanya G, Zhao J, Perlman S, Gallagher T. A transmembrane serine protease is linked to the severe acute respiratory syndrome coronavirus receptor and activates virus entry. J Virol. 2011;85(2):873–882. doi: 10.1128/JVI.02062-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270–273. doi: 10.1038/s41586-020-2012-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Wang P, Luo R, Zhang M, Wang Y, Song T, Tao T, et al. A cross-talk between epithelium and endothelium mediates human alveolar–capillary injury during SARS-CoV-2 infection. Cell Death Dis. 2020;11(12):1042. doi: 10.1038/s41419-020-03252-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Barretina J, Caponigro G, Stransky N, Venkatesan K, Margolin AA, Kim S, et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature. 2012;483(7391):603–607. doi: 10.1038/nature11003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, Berne MA, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426:450–454. doi: 10.1038/nature02145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Belouzard S, Chu VC, Whittaker GR. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proc Natl Acad Sci USA. 2009;106(14):5871–5876. doi: 10.1073/pnas.0809524106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Zou L, Ruan F, Huang M, Liang L, Huang H, Hong Z, et al. SARS-CoV-2 viral load in upper respiratory specimens of infected patients. N Engl J Med. 2020;382(12):1177–1179. doi: 10.1056/NEJMc2001737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Daly JL, Simonetti B, Klein K, Chen KE, Williamson MK, Antón-Plágaro C, et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science. 2020;370:861–865. doi: 10.1126/science.abd3072 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Cantuti-Castelvetri L, Ojha R, Pedro LD, Djannatian M, Franz J, Kuivanen S, et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science. 2020;370:856–860. doi: 10.1126/science.abd2985 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Puelles VG, Lütgehetmann M, Lindenmeyer MT, Sperhake JP, Wong MN, Allweiss L, et al. Multiorgan and renal tropism of SARS-CoV-2. N Engl J Med. 2020;383(6):590–592. doi: 10.1056/NEJMc2011400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Sungnak W, Huang N, Bécavin C, Berg M, Queen R, Litvinukova M, et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat Med. 2020;26(5):681–687. doi: 10.1038/s41591-020-0868-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Singh M, Bansal V, Feschotte C. A Single-Cell RNA Expression Map of Human Coronavirus Entry Factors. Cell Rep. 2020;32:108175. doi: 10.1016/j.celrep.2020.108175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Cheung KS, Hung IF, Chan PP, Lung KC, Tso E, Liu R, et al. Gastrointestinal manifestations of SARS-CoV-2 infection and virus load in fecal samples from the Hong Kong cohort and systematic review and meta-analysis. Gastroenterology. 2020;159(1):81–95. doi: 10.1053/j.gastro.2020.03.065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Bojkova D, McGreig JE, McLaughlin KM, Masterson SG, Widera M, Krähling V, et al. SARS-CoV-2 and SARS-CoV differ in their cell tropism and drug sensitivity profiles. bioRxiv, 2020.04.03.024257. Available from: https://www.biorxiv.org/content/10.1101/2020.04.03.024257v1 [Google Scholar]
- 48. Bertoglio F, Fühner V, Ruschig M, Heine PA, Abasi L, Klünemann T, et al. A SARS-CoV-2 neutralizing antibody selected from COVID-19 patients by phage display is binding to the ACE2-RBD interface and is tolerant to most known recently emerging RBD mutations. bioRxiv 2020.12.03.409318. Available from https://www.biorxiv.org/content/10.1101/2020.12.03.409318v2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Ng KW, Attig J, Bolland W, Young GR, Major J, Wrobel AG, et al. Tissue-specific and interferon-inducible expression of nonfunctional ACE2 through endogenous retroelement co-option. Nat Genet. 2020;52(12):1294–1302. doi: 10.1038/s41588-020-00732-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Onabajo OO, Banday AR, Stanifer ML, Yan W, Obajemu A, Santer DM, et al. Interferons and viruses induce a novel truncated ACE2 isoform and not the full-length SARS-CoV-2 receptor. Nat Genet. 2020;52(12):1283–1293. doi: 10.1038/s41588-020-00731-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Rehman SU, Tabish M. Alternative splicing of ACE2 possibly generates variants that may limit the entry of SARS-CoV-2: a potential therapeutic approach using SSOs. Clin Sci. 2020;134:1143–1150. doi: 10.1042/CS20200419 [DOI] [PubMed] [Google Scholar]
- 52. Hou Y, Zhao J, Martin W, Kallianpur A, Chung MK, Jehi L, et al. New insights into genetic susceptibility of COVID-19: an ACE2 and TMPRSS2 polymorphism analysis. BMC Med. 2020;18:216. doi: 10.1186/s12916-020-01673-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Howley PM, Knipe, David M. Fields Virology: Emerging Viruses. Wolters Kluwer Health; 2020. ISBN 978-1-975112-55-4
- 54. Wei J, Alfajaro MM, DeWeirdt PC, Hanna RE, Lu-Culligan WJ, Cai WL, et al. Genome-wide CRISPR Screens Reveal Host Factors Critical for SARS-CoV-2 Infection. Cell. 2021;184(1):76–91. doi: 10.1016/j.cell.2020.10.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun. 2020;11(1):1620. doi: 10.1038/s41467-020-15562-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Wang K, Chen W, Zhang Z, Deng Y, Lian JQ, Du P, et al. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct Target Ther. 2020;5(1):283. doi: 10.1038/s41392-020-00426-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Ziegler CGK, Allon SJ, Nyquist SK, Mbano IM, Miao VN, Tzouanas CN, et al. HCA Lung Biological Network. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell. 2020;181(5):1016–1035. doi: 10.1016/j.cell.2020.04.035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Smith JC, Sausville EL, Girish V, Yuan ML, Vasudevan A, John KM, et al. Cigarette smoke exposure and inflammatory signaling increase the expression of the SARS-CoV-2 receptor ACE2 in the respiratory tract. Dev Cell. 2020;53(5):514–529. doi: 10.1016/j.devcel.2020.05.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Uphoff CC, Pommerenke C, Denkmann SA, Drexler HG. Screening human cell lines for viral infections applying RNA-Seq data analysis. PLoS One. 2019;14(1):e0210404. doi: 10.1371/journal.pone.0210404 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Robinson JT, Thorvaldsdóttir H, Wenger AM, Zehir A, Mesirov JP. Variant review with the integrative genomics iewer. Cancer Res. 2017;77(21):e31–e34. doi: 10.1158/0008-5472.CAN-17-0337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K, Kernytsky A., et al. The genome analysis toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010;20:1297–1303. doi: 10.1101/gr.107524.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. McLaren W, Pritchard B, Rios D, Chen Y, Flicek P, Cunningham F. Deriving the consequences of genomic variants with the Ensembl API and SNP effect predictor. Bioinformatics. 2010;26:2069–2070. doi: 10.1093/bioinformatics/btq330 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Uphoff CC, Drexler HG. Comparative PCR analysis for detection of mycoplasma infections in continuous cell lines. In Vitro Cell Dev Biol Anim. 2002;38:79–85. [DOI] [PubMed] [Google Scholar]
- 64. Dirks WG, Drexler HG. STR DNA typing of human cell lines: detection of intra- and interspecies cross-contamination. Methods Mol Biol. 2013;946:27–38. doi: 10.1007/978-1-62703-128-8_3 [DOI] [PubMed] [Google Scholar]
- 65. Corman VM, Landt O, Kaiser M, Molenkamp R, Meijer A, Chu DKW, et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. 2020;25:2000045. doi: 10.2807/1560-7917.ES.2020.25.3.2000045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Quentmeier H, Schneider B, Röhrs S, Romani J, Zaborski M, Macleod RA, et al. SET-NUP214 fusion in acute myeloid leukemia—and T-cell acute lymphoblastic leukemia-derived cell lines. J Hematol Oncol. 2009;2:3. doi: 10.1186/1756-8722-2-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Original western blots to cropped western blot images in Fig 2B.
(PDF)
Virus titers in the supernatant of infected cell lines at 1-4 dpi determined by plaque assay for the top 11-25 ACE2 expressing, two neuronal (DBTRG-05MG, KELLY), and two oral squamous cell carcinoma (UPCI-SCC-074, UPCI-SCC-131) cell lines. The red dotted line indicates the initial inoculum (2000 pfu/ml); the blue dashed line specifies the minimal limit of detection LOD 33.3 pfu/ml. Number of replicates varied between 3-8. pfu: plaque-forming units; mean values with SEM.
(TIFF)
Neutralising SARS-CoV-2 antibody was applied to the high permissive cell lines CAL-51 and CL-14 and the control cell line VERO-E6. With increasing antibody concentration viral production is decreasing in all three cell lines. The red dotted line indicates the initial inoculum (2000 pfu/ml); the blue dashed line specifies the minimal limit of detection LOD 33.3 pfu/ml. Note the logarithmic scaling for the x-axis.
(TIF)
Different exon usage of ACE2 for the top 25 ACE2 expressing cell lines plus CALU-3. RNA-seq data were visualised via IGV and cell lines ordered alphabetically.
(TIFF)
Gene expression, splice and single nucleotide variants of ACE2 and TMPRSS2 in the cell lines used in this study. These were identified on the basis of CCLE RNA-seq data (FPKM, see Material and methods). +truncated splice variant observed; 0/1 heterozygous, 1/1 homozygous mutation.
(ODS)
Data Availability Statement
All relevant data are within the paper and its Supporting information files.