

The neuroimmune system of the brain: Early studies (1990’s) on the neurological consequences of human immunodeficiency virus-1 (HIV-1) infection in the brain were instrumental in establishing that specific brain cell types can function as an innate immune system within the brain and in that role influence cognitive function (Kaul et al., 2005). It is now known that this system, referred to as the neuroimmune system, is an important signaling system that plays a key role in brain function under both physiological and pathophysiological conditions. The principal cellular components of the neuroimmune system are the glial cells, primarily astrocytes and microglia. Astrocytes are the most populous cell type in the brain, whereas microglia comprise about 10% of the brain cells. These cell types produce many of the same chemical signaling factors commonly associated with the peripheral immune system, including small proteins such as the cytokine interleukin-6 (IL-6) and the chemokines C-C motif chemokine 2 (CCL2) and C-X-C Motif Chemokine Ligand 10 (CXCl10), the focus of our studies and the neuroimmune factors that are used as examples in this short perspective.
Cytokines and chemokines are members of a large body of signaling factors with diverse and redundant actions that comprise the cytokine family of immune factors. An extensive body of research has documented that neuroimmune factors are normally produced at relatively low levels in the brain. However, when the brain is exposed to adverse conditions such as injury (e.g., traumatic brain injury), seizures (e.g., epilepsy), infection (e.g., HIV-1), drugs of abuse (e.g., alcohol) or processes involved in neurodegenerative (e.g., multiple sclerosis, Alzheimer’s disease) and psychiatric disorders (e.g., depression), expression can become dysregulated, resulting in chronically elevated brain levels. The elevated levels have been reported to function in repair processes, produce adaptive changes that alter brain function, and/or induce damage and toxicity. However, many of the mechanisms that underlie these actions have yet to be identified, a process that is challenged by the complexity and broad actions and interactions of the neuroimmune factors. Typically, multiple neuroimmune factors are produced under adverse conditions, different neuroimmune factors can have similar actions, interactions can occur between the actions of different neuroimmune factors, either at the signal transduction level or at common downstream targets, and neuroimmune actions can be covert in nature only to be revealed under specific conditions.
Considerable research has focused on the association of neuroimmune factors with various physiological or pathophysiological conditions. However, few studies have focused on specific neuroimmune factors as modulators/regulators of fundamental neurophysiological processes such as neuronal excitability, synaptic transmission, and synaptic plasticity, all of which are essential for normal brain function. Such actions are likely to play a key role in the cognitive dysfunction associated with many brain disorders. Our studies have focused on this issue, using the strategy classically used to identify neurotransmitters and their actions in the brain, identification of the actions of individual neuroimmune factors, information that can be used to understand more complex situations involving multiple neuroimmune factors.
Neuroimmune factors act through specific signal transduction pathways: A variety of studies have shown that specific cognate membrane receptors coupled to signal transduction pathways exist for cytokines and chemokines and mediate their biological effects. Receptors for the cytokine IL-6 partner with a signal transduction pathway involving tyrosine kinases (e.g., JAK2/STAT3), whereas receptors for chemokines such as CCL2 and CXCL10 are G-protein coupled receptors. Both pathways play a role in the regulation of intracellular biochemical processes and gene expression. Many cells in the brain including neurons and glial cells express membrane receptors for cytokines and chemokines and relevant signal transduction components, implicating them as potential downstream cellular targets of neuroimmune signaling.
Neuroimmune factors alter neuronal function: The ability of neuroimmune factors to act as modulators/regulators of neuronal physiology was revealed in early physiological experiments involving electrical recordings of neuronal activity or imaging of intracellular Ca2+ levels from live cells in vivo or in vitro. For example, our studies of cultured hippocampal or cerebellar neurons demonstrated that acute exposure to the chemokines CCL2 or CXCL10 altered basic neuronal properties such as resting intracellular Ca2+ levels, Ca2+ signaling evoked by activation of metabotropic glutamate receptors (mGluR), and action potential generation (Nelson and Gruol, 2004; van Gassen et al., 2005). Ca2+ is an important second messenger that controls numerous neuronal functions, action potential generation and responses to transmitters are essential for synaptic transmission, which mediates brain function and, consequently behavior. In contrast, acute exposure to the cytokine IL-6 did produce changes in intracellular Ca2+ or electrical activity of cultured hippocampal or cerebellar neurons. However, chronic treatment with IL-6 increased resting Ca2+ levels, enhanced the Ca2+ response evoked by activation of mGluR, and depressed action potential generation in the cultured neurons (Nelson et al., 2002, 2004). Thus, both chemokines and cytokines can alter important neuronal properties, and often the same properties, although the conditions for these actions (e.g., acute vs. chronic exposure) and likely the behavior consequences differ.
Studies of transgenic mice that chronically express elevated brain levels of CCL2 (CCL2-tg) or IL-6 (IL-6 tg) further demonstrated that these neuroimmune factors can alter neuronal and synaptic function under conditions of long-term exposure, such as occurs in neurodegenerative and psychiatric disorders or chronic brain infection (Gruol, 2013, 2015). In both transgenic lines, genetic manipulation targeted the increased expression to astrocytes, which are closely associated with synaptic networks in the brain and are regulators of synaptic function. These studies were carried out in acutely isolated hippocampal slices from the IL-6 tg or CCL2-tg mice at the excitatory glutamatergic Schaffer collateral (SC) to CA1 pyramidal neuron (SC-CA1) synapse, the most highly studied synapses in the brain (Figure 1A). Synaptic responses were recorded extracellularly (field potential recordings) and were evoked by brief electrical stimulation of the SC. Results showed that excitatory synaptic responses at the SC-CA1 synapse evoked by SC stimulation were enhanced in hippocampus from IL-6 tg mice (Nelson et al., 2012), whereas excitatory synaptic responses at the SC-CA1 synapse evoked by SC stimulation were not altered in hippocampus from CCL2-tg mice (Bray et al., 2013). However, in hippocampus from CCL2-tg mice an enhancement of action potential firing evoked in the soma by the synaptic response was observed, indicative of increased excitability of the pyramidal neurons (Bray et al., 2013).
Figure 1.
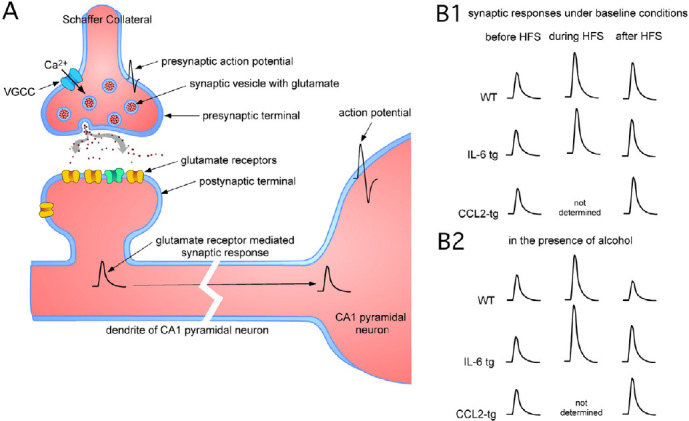
Excitatory synaptic transmission at the Schaffer collateral to CA1 pyramidal neuron synapse.
(A) Simplified diagram of components involved in synaptic function at the Schaffer collateral to CA1 pyramidal neuron synapse [Adapted from Gruol (2013)] . During experiments the Schaffer collaterals are electrically stimulated causing a depolarization of the axon (presynaptic) terminal and a presynaptic action potential. The depolarization activates voltage-gated Ca2+ channels (VGCCs) located at the presynaptic terminal resulting in Ca2+ influx, which initiates transmitter (glutamate) release from the terminal. Glutamate interacts with postsynaptic glutamate receptors, which translate the chemical signal into an electrical response (the synaptic response). The synaptic response is conducted to the cell soma where, if it is large is large enough, it elicits one or more action potentials. The action potentials are conducted to the axon terminal of the pyramidal neuron where they activate synaptic transmission to postsynaptic neurons of the synaptic network. A synaptic network involves both excitatory and inhibitory neurons that interact and define the final output of the network and how it affects brain function. Changes to synaptic network activity will change brain function. Not shown are glial cells that are in close contact with pre-and postsynaptic components of the synapse. (B) Diagram showing the relative amplitude of synaptic responses before HFS, during HFS and after HFS (during LTP). Responses are of similar amplitude for the three genotypes under baseline conditions (B1), during HFS for the WT and IL-6 tg (CCL2-tg not determined) and after HSF. In the presence of acute alcohol (60 mM) (B2), synaptic responses are of similar amplitude before HFS. The synaptic response during HFS is significantly larger in the hippocampus from the IL-6 tg mice than in the hippocampus from the WT mice (CCL2-tg not determined). After HFS (during LTP) the synaptic response is reduced in the hippocampus from the WT mice, whereas in the IL-6 tg and CCL2 tg mice, the magnitude of the synaptic response is similar to that observed before alcohol exposure. HFS: High-frequency electrical stimulation; IL-6 tg and CCL2-tg: transgenic mice that chronically express elevated brain levels of C-C motif chemokine 2 (CCL2) or interleukin-6 (IL); LTP: long-term potentiation; WT: wild-type.
Neuroimmune factors alter synaptic plasticity: Emerging research also indicates that neuroimmune factors can modulate synaptic plasticity, a fundamental brain process that is considered to be a cellular mechanism of memory and learning. Synaptic plasticity is a short- or long-term change in the efficacy of synaptic transmission induced by repetitive activity of synaptic networks. Both presynaptic and postsynaptic mechanisms contribute to synaptic plasticity. Experimentally, synaptic plasticity is induced at the SC-CA1 synapse by high-frequency electrical stimulation (HFS) of the SCs. Two types of synaptic plasticity are produced, a short-term (sec-min) enhancement of synaptic responses termed post-tetanic potentiation (PTP) followed by a long-term (hours to days or longer) enhancement of synaptic responses termed long-term potentiation (LTP). Both forms of synaptic plasticity play a role in memory and learning. Synaptic plasticity (PTP and LTP) induced by HFS was not altered by the elevated levels of IL-6 or CCL2 in hippocampal slices from the IL-6-tg and CCL2-tg mice, respectively. In contrast, studies in hippocampal slices from wildtype rats showed that acute application of IL-6 produced a depression of PTP and LTP at the SC-CA1 synapse (Tancredi et al., 2000). Building on this work are studies showing that HFS results in IL-6 production by glial cells and serves as a negative regulator of LTP, IL-6 can alter another form of synaptic plasticity termed long-term synaptic depression (LTD), IL-6 reduces presynaptic glutamate release at cortical synapses, and in behavioral studies, that IL-6 is involved in cognitive function (Gruol, 2015).
Interactions with alcohol: Although synaptic plasticity was not altered in hippocampus from the IL-6-tg and CCL2-tg mice, our recent studies on the effects of alcohol (ethanol) revealed that covert neuroadaptive changes had been produced by chronic exposure to IL-6 or CCL2. Alcohol/neuroimmune interactions were of interest because alcohol significantly alters brain function, transiently or persistently depending on the pattern of use, recent studies indicate that alcohol (and other drugs of abuse) can induce glial cells to produce elevated levels of neuroimmune factors in the brain (Lacagnina et al., 2017), and alcohol use disorders are often co-morbid with other detrimental conditions associated with elevated levels of neuroimmune factors in the brain (e.g., major depression). Using the IL-6 tg and CCL2-tg mice to model long-term exposure of the brain to these neuroimmune factors, as would occur with chronic alcohol abuse, we tested the possibility that IL-6 or CCL2 can produce neuroadaptive changes that alter the effects of alcohol, an interaction that could play a role in the development of alcohol dependence.
In the absence of alcohol, LTP and PTP in hippocampus from IL-6 tg, CCL2-tg and their respective control mice were of similar magnitude. Exposure to pharmacologically relevant doses of alcohol, 20 mM (considered a moderate dose) and 60 mM (considered a high dose), produced a dose-dependent depression of both LTP and PTP in hippocampus from control mice. However, in hippocampus from IL-6 tg mice acute exposure to 20 mM or 60 mM alcohol had no apparent effect on LTP or PTP, whereas in hippocampus from CCL2-tg mice, 20 mM acute alcohol enhanced PTP and LTP while 60 mM had no apparent effect. Thus, alcohol revealed covert neuroadaptive changes produced by the neuroimmune factors that altered the effects of alcohol on cellular processes involved in memory and learning. Consistent with this effect, in a behavioral test of learning, cued and contextual fear conditioning, acute alcohol depressed cued and contextual learning in control mice but not in CCL2 mice (Bray et al., 2013). The contextual conditioning task in this test is considered to be hippocampal-dependent, whereas the cued conditioning task is thought to be hippocampal-independent. These interactions between IL-6 or CCL2 and alcohol suggest that there are common synaptic targets between these neuroimmune factors and alcohol.
The mechanisms mediating the covert neuroadaptive effects of IL-6 and CCL2 and interactions with alcohol have yet to be identified. However, our recent study points to the involvement of presynaptic mechanisms that regulate transmitter release (Gruol et al., 2020). In these studies, synaptic responses elicited during HFS showed an initial facilitation followed by a depression, a pattern of changes known to be dependent on presynaptic mechanisms underlying transmitter release. The facilitated synaptic responses were of similar magnitude in hippocampus from IL-6 tg and control mice (Figure 1B1). Acute alcohol exposure (60 mM) significantly enhanced the facilitatory synaptic responses in the hippocampus from the IL-6 tg mice with no effect on the facilitatory synaptic responses in hippocampus from the control mice (Figure 1B2). Regression analysis showed that in the presence of alcohol, the magnitude of the facilitatory synaptic responses in the hippocampus from the IL-6 tg mice predicted the magnitude of LTP at a 27% level, whereas without alcohol the predicted value was only 15%. In control mice, in the presence of alcohol the magnitude of the facilitatory synaptic response predicted the magnitude of LTP at a 3% level, whereas without alcohol the predicted value was 30%. These results are consistent with neuroadaptive effects of IL-6 on presynaptic machinery involved in transmitter release. Studies by others have shown that alcohol alters presynaptic mechanisms of transmitter release, resulting in decreased release (Gioia and McCool, 2017). Thus, the lack of effect of alcohol on LTP in the IL-6 tg mice may result from IL-6-induced neuroadaptive effects that blocked the ability of alcohol to reduce transmitter release. Consistent with this possibility, synaptic responses elicited by single SC stimulations were also increased by alcohol exposure in the hippocampus of the IL-6 tg mice.
Taken together, these studies provide representative examples of how individual neuroimmune factors can affect neuronal processes that underlie brain function. Identification of the cellular targets and neuronal consequences of neuroimmune actions is basic to an understanding of the role of the neuroimmune system in the brain. This area is understudied, critical to an understanding of the mechanisms underlying the effects of the neuroimmune system on cognitive function, and ripe for investigation.
This work was supported by National Institutes of Health Grant AA024484 (to DLG).
Footnotes
Copyright license agreement: The Copyright License Agreement has been signed by the author before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
C-Editors: Zhao M, Wang L; T-Editor: Jia Y
References
- 1.Bray JG, Reyes KC, Roberts AJ, Ransohoff RM, Gruol DL. Synaptic plasticity in the hippocampus shows resistance to acute ethanol exposure in transgenic mice with astrocyte-targeted enhanced CCL2 expression. Neuropharmacology. 2013;67:115–125. doi: 10.1016/j.neuropharm.2012.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gioia DA, McCool B. Strain-dependent effects of acute alcohol on synaptic vesicle recycling and post-tetanic potentiation in medial glutamate inputs to the mouse basolateral amygdala. Alcohol Clin Exp Res. 2017;41:735–746. doi: 10.1111/acer.13343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gruol DL. Alcohol-chemokine interaction and neurotransmission. In: Cui C, Grandison L, Noronha A, editors. Neural-Immune Interactions in Brain Function and Alcohol Related Disorders. New York: Springer; 2013. pp. 387–424. [Google Scholar]
- 4.Gruol DL. IL-6 regulation of synaptic function in the CNS. Neuropharmacology. 2015;96:42–54. doi: 10.1016/j.neuropharm.2014.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gruol DL, Hernandez RV, Roberts A. Alcohol enhances responses to high frequency stimulation in hippocampus from transgenic mice with increased astrocyte expression of IL-6. Cell Mol Neurobiol. 2020 doi: 10.1007/s10571-020-00902-6. doi: 101007/s10571-020-00902-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kaul M, Zheng J, Okamoto S, Gendelman HE, Lipton SA. HIV-1 infection and AIDS: consequences for the central nervous system. Cell Death Differ 12 Suppl. 2005;1:878–892. doi: 10.1038/sj.cdd.4401623. [DOI] [PubMed] [Google Scholar]
- 7.Lacagnina MJ, Rivera PD, Bilbo SD. Glial and neuroimmune mechanisms as critical modulators of drug use and abuse. Neuropsychopharmacology. 2017;42:156–177. doi: 10.1038/npp.2016.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nelson TE, Gruol DL. The chemokine CXCL10 modulates excitatory activity and intracellular calcium signaling in cultured hippocampal neurons. J Neuroimmunol. 2004;156:74–87. doi: 10.1016/j.jneuroim.2004.07.009. [DOI] [PubMed] [Google Scholar]
- 9.Nelson TE, Ur CL, Gruol DL. Chronic interleukin-6 exposure alters electrophysiological properties and calcium signaling in developing cerebellar purkinje neurons in culture. J Neurophysiol. 2002;88:475–486. doi: 10.1152/jn.2002.88.1.475. [DOI] [PubMed] [Google Scholar]
- 10.Nelson TE, Netzeband JG, Gruol DL. Chronic interleukin-6 exposure alters metabotropic glutamate receptor-activated calcium signalling in cerebellar Purkinje neurons. Eur J Neurosci. 2004;20:2387–2400. doi: 10.1111/j.1460-9568.2004.03706.x. [DOI] [PubMed] [Google Scholar]
- 11.Nelson TE, Olde Engberink A, Hernandez R, Puro A, Huitron-Resendiz S, Hao C, De Graan PN, Gruol DL. Altered synaptic transmission in the hippocampus of transgenic mice with enhanced central nervous systems expression of interleukin-6. Brain Behav Immun. 2012;26:959–971. doi: 10.1016/j.bbi.2012.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tancredi V, D’Antuono M, Cafe C, Giovedi S, Bue MC, D’Arcangelo G, Onofri F, Benfenati F. The inhibitory effects of interleukin-6 on synaptic plasticity in the rat hippocampus are associated with an inhibition of mitogen-activated protein kinase ERK. J Neurochem. 2000;75:634–643. doi: 10.1046/j.1471-4159.2000.0750634.x. [DOI] [PubMed] [Google Scholar]
- 13.van Gassen KL, Netzeband JG, de Graan PN, Gruol DL. The chemokine CCL2 modulates Ca2+ dynamics and electrophysiological properties of cultured cerebellar Purkinje neurons. Eur J Neurosci. 2005;21:2949–2957. doi: 10.1111/j.1460-9568.2005.04113.x. [DOI] [PubMed] [Google Scholar]