

ABSTRACT
Bee orchids have long been an excellent example of how dishonest signal works in plant-animal interaction. Many studies compared the flower structures that resemble female bees, leading toward pseudo-copulation of the male bees on the flower. Using Machine Learning, we tested whether nature is capable of besting artificial intelligence. A total of 2000 images of related bees, wasps, and Ophrys sp. were collected from the Google Image Repository. Unsuitable images were later filtered out manually, leaving a total of 995 images in the final selection. 80% of these images were used to build a supervised model using Logistic Regression, while the model accuracy was tested using 20% of the remaining images. Based on our results using Wolfram Mathematica, the Ophrys is not capable of fooling artificial intelligence. The accuracy, accuracy baseline, mean cross-entropy, Area Under ROC (receiver operating characteristic curve) curve (AUC) and the confusion matrix gave excellent image classification. However, we can now show the key points and highlights of the images and how the structures closely resemble actual bees using the SURF method. Rather than just a descriptive method, ML learning has enabled a more quantitative approach. Since this is a simple test, we encourage other scientists to adopt our approach using a larger dataset and better database samples.
KEYWORDS: Bee orchids, bees, mimicry, pseudo-copulation, machine learning, artificial intelligence
Introduction
Plants use multiple strategies to promote the propagation of their seed/pollen. Some of the most common methods are by an animal or by the wind in the natural world. While most strategies rely on direct or indirect signaling, few species of plants rely on dishonest signals. One of the most prominent examples is the bee orchids (genus Ophrys).1 Flowers of each Ophrys species mimic sexually receptive females (bees) and fool actual amorous male bees into mating with them via pseudo-copulation. This form of mimicry is called Pouyannian mimicry.2 These flowers emit pseudo-pheromones (allomone) that mimic the sexual pheromones of bees females counterparts in addition to the visual and tactile cues.3,4 They would attempt to copulate with the flower, which transfers pollen on their head or abdomen.5 The pollen is then transferred to another flower of the same orchid species during another pseudo-copulation attempt by the bees.
Besides the allomone, males are also attracted by the morphological cues like color, shape and hair structures of the labellum that resemble an actual female bee. Although floral scents remain as the most prominent key features in successful pollination in most literature, however, it is studied that the color of the perianth or labellum can play an important role to increase the contrast of the flower against the background, and thus increase the rate of pollination.6–14 Moreover, wide-ranging studies on how the evolution of flower color is shaped by bee vision in the North Hemisphere further reinforce the significance of the flowers’ morphology.15,16,postulated that such deception by the Ophrys might have a secondary effect, in which the pseudo-bee design might be able to deter herbivores from eating the orchids. While many studies are conducted on the co-evolution between these orchids and bees, we would like to review the matter from another perspective.
Evolution in nature typically takes millions of years, while computing technology has only emerged in less than a century. Machine learning (ML) is increasingly popular over the recent decades with the booming of the digital revolution that powers everyday tools from speech recognition in Siri to autonomous vehicles in Tesla. Machine learning is a subset of artificial intelligence (AI) that improves computational algorithms automatically through experience. It is built based on training data to make predictive analysis and generate outcome without being programmed directly. Machine learning typically creates a model based on the training data attested by a separate test dataset. One of the most significant applications of machine learning is undoubtedly computer vision. Computer vision is a multidisciplinary science that aims to automate tasks primarily based on visual input such as images and videos.17 The components of computer vision functions require steps for visual images to be obtained, interpreted, evaluated, and understood, and high-dimensional data derived from the physical world to provide numerical or symbolic information in decision-making and analysis.18 In other words, visual inputs are disentangled and transformed into binary information that can be processed and understood by computers easily. Computers then can use this information to perform various types of tasks without human’s help, such as pattern recognition and categorization. These capabilities allow scientists, especially wildlife researchers, to conduct their studies efficiently when it comes to data collection.
In this short communication, we aim to test two objectives on the Pouyannian strategy of the Bee Orchids. For the first objective, we wanted to test whether the deception by the orchids would be able to fool Machine Learning (ML). We wanted to study the key characteristics between the Ophrys orchids and the bees using ML for the second objective.
Methodology
To test the first objective, we decided to use Wolfram Mathematica as the primary Machine Learning (ML) platform. There are numerous other ML platforms such as Google Teachable Machine, Keras, and Tensorflow with Python; however, we found Wolfram Mathematica is more user-friendly. The most suitable algorithm is calculated automatically by Mathematica. Furthermore, much of the backend calculation is handled by Wolfram Server, reducing the workload and learning time from the actual machine. We began with collating O. apifera, O. Insectifera, O. bombyliflora and O. speculum using Google Images. As for the bee pollinators, we searched the images of common bees and wasps commonly linked to the pseudo-copulation with the orchids. To simplify the categorization, we did not discriminate between bees and wasps and we group together all the images under the bees category. This is appropriate enough as the main goal is to test the ML on the differences between flowers and bees.
We downloaded the images using Fatkun Batch Download for Google Chrome. For each of the categories, we started with about 400 images, with 2000 images altogether. However, we vetted the images to select the best images for ML. We outlined a few criteria’s for the orchid and bees. The images must be clear and focused on the organism only. Multiple species on a single image were also discarded to prevent double classification by the machine. The images must be anterior or lateral view. Images from a far distance (more than 1 meter of the object) were also discarded. Images that have watermarks were also discarded. We also made sure the correct species were selected and arranged in their respective categories for the orchids.
For the final selection for the images used, we had 113 for O. apifera, 243 for O. bombyliflora, 264 for O. insectifera, 253 for O. speculum and 122 for the bees. In Mathematica, we used Classify function (refer to https://reference.wolfram.com/language/ref/Classify.html for more information) to train and test the model simultaneously. This method of ML is considered supervised learning. We used random sample and complement sample to create a 80:20 ratio. Eighty percent is used for training, and 20% is used for testing the model accuracy. Method of classification is automatically selected by Mathematica, as the backend calculation ran several methods simultaneously and suggested the most accurate to be used. In all of our training sessions, all the methods suggested was Logistic Regression, the most common method.
We performed an accuracy check for our classify model. We compared the accuracy, accuracy baseline, mean cross-entropy, Area Under ROC (receiver operating characteristic curve) curve [AUC) and the confusion matrix. For the best range of values of these descriptors, we refer to 19. Accuracy is based on percentages; a 100% accuracy means the model is perfect. Accuracy baseline is the accuracy based on the simplest model of assumption. In our example, the accuracy baseline is the method that uses heuristics, summary statistics, randomness, or machine learning to create predictions for a dataset. The accuracy value is compared against the baseline accuracy. Mean cross-entropy measures the difference between two probability distributions for a given random variable or set of events. A perfect prediction is 0.00, and any values that are more than two is considered flawed. For the AUC, 0.9 to 1.0 is considered very good, and anything less than 0.5 can be considered a fail. The confusion matrix is used to explain the Type I and Type II error.
For the second objective, we wanted to see the key characteristics that the machine uses to distinguish between bees and orchids. We used ImageKeypoints and ImageHighlights (refer to https://reference.wolfram.com/language/ref/ImageKeypoints.html for more information]. In simple term, ML locates the most prominent aspect of the objects contained in a picture. In our example, the feature might be the edges of the wings or the leaf. It could also be the hair structure on the abdomen of certain bees. In a typical picture, the key points range from hundreds to thousands depending on the complexity of the objects. In this research, we limit the number of key points to the 100th strongest points. Several methods derive the key points, and these points are highlighted using SURF (Speeded Up Robust Features). SURF is a fast and robust algorithm used to compare images. It uses a circular sampling of the pixels, and it also accounts for the direction of the points. Since the objective is to detect the similarity key points, we selected the best picture for each of the orchids and two images for the bees.
Results
Table 1 shows the overall results from the classification method. Overall, the accuracy, accuracy baseline, mean cross-entropy and AUC all gave excellent results. The confusion matrix table showed that ML could perfectly distinguish O. apifera. However, ML did misclassify one O. insectifera, three O. bombyliflora and four O. speculum.
Table 1.
Results from the ML output
| Species | O. apifera | O. bombyliflora | O. insectifera | O. speculum |
| Number of test examples | 47 | 73 | 77 | 75 |
| Accuracy | 100.00% | 95.92.3% | 98.7 1.3% | 94.7 2.6% |
| Accuracy baseline | 53 7% | 64 6% | 79 5% | 60 6% |
| Mean cross entropy | 0.009 0.004 | 0.09 0.040 | 0.009 0.09 | 0.200 0.09 |
| Area under ROC curve | Apifera -> 1 Bees ->1 |
Bombylifora ->1 Bees ->1 |
Insectifera->0.99 Bees-0.99 |
Speculum->0.99 Bees -> 0.99 |
| Confusion matrix | 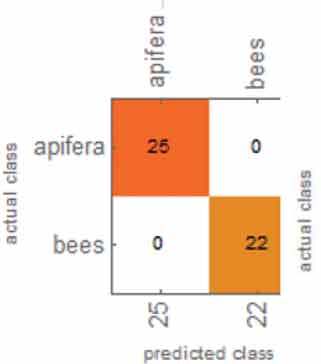 |
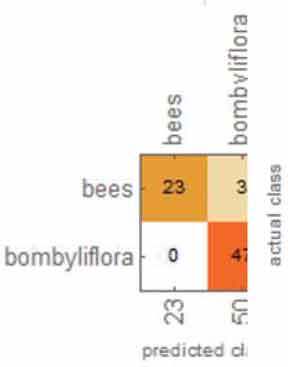 |
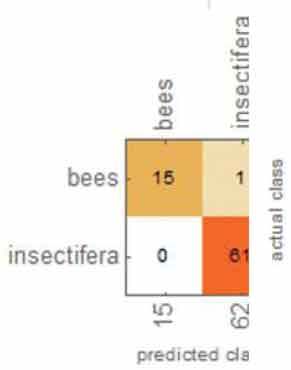 |
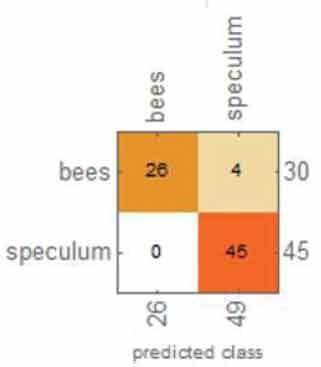 |
Figure 1 (A to F) shows the analysis of the key characteristics using the SURF method. Figure 1(c), and d are all pictures of the samples that ML misclassified. Picture 1 F shows Dasyscolia ciliata in pseudo-copulation with O. speculum. ML correctly identifies this image.
Figure 1.

A: Image of Dasyscolia ciliata against a normal background. Notice that the ML wrongly targeted an open white background (bottom left) as the key feature. Image is from Karim Haddad ©. (https://www.inaturalist.org/photos/66075769). (b): Image of Ophrys apifera. Image taken from T.Voekler, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons (https://commons.wikimedia.org/wiki/File:Flower_of_Bee_Orchid_Ophrys_apifera.png). (c): Image of Ophrys insectifera. Image taken from Bouba at French Wikipedia, CC BY-SA 3.0 <http://creativecommons.org/licenses/by-sa/3.0/>, via Wikimedia Commons. (https://commons.wikimedia.org/wiki/File:Insectifera.jpg). (d): Image of Ophrys bombyliflora. Image taken from Orchi, CC BY-SA 3.0 <http://creativecommons.org/licenses/by-sa/3.0/>, via Wikimedia Commons (https://commons.wikimedia.org/wiki/File:Ophrys_bombyliflora_Mallorca_01.jpg). (e): Image of Ophyrys speculum. Image taken from Carsten Niehaus CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons. (https://upload.wikimedia.org/wikipedia/commons/e/e8/Ophrys_speculum_d.JPG). Notice that ML mistakenly highlighted an out of focus flower at the bottom right of the main flower. (f): Image of Dasyscolia ciliata in a pseudo-copulation with O. speculum. Image taken from Pietro Niolu CC BY-SA 3.0 <http://creativecommons.org/licenses/by-sa/3.0/>, via Wikimedia Commons (https://pt.wikipedia.org/wiki/Dasyscolia_ciliata) Additional note: the circles represent the SURF highlights. The yellow circle refers to points in which the direction matters whilst the red circle disregards the direction. In terms of calculation there is not much of a difference in the selection and this is done automatically by the ML
Discussion
Our results have shown that the Pouyannian mimicry of the Ophrys species cannot fool ML. The overall recognition values show an almost perfect classification. Computer vision is not new to ecology and wildlife studies. In recent years, machine learning utilizes computer vision has been used extensively by ecologists to cope with large volumes of scientific data.20 Some tools also encourage citizen scientists to capture and upload the pictures of animals around them into a cloud-based database using computer vision. The most prominent examples are probably iNaturalist, PlantSnap, LeafSnap Seek, Merlin Bird ID, Picture Bird/Insect/Fish applications that include all species worldwide.21–23
This paper does not aim for the intricacies and complexity of ML (as most technical works are done by using Convolutional Neural Networks (CNN)) but instead using the most basic logistic regression image analysis. Furthermore, our classification analysis is quite simple as there are only five classes (four species of Bee orchids and one bee category). In comparison, the iNaturalist compares images from thousands of species in its database and most of the backend calculations are done in the cloud server. ML is becoming more accessible with the inclusion of more user-friendly Graphical User Input (GUI). Google Teachable Machine and even Apple ML Xcode adopt a click and drag interface with little coding knowledge needed. Wolfram Mathematica is also a good example, where the calculations are done within 13 lines of simple codes (see supplemental material for the Mathematica Notebook file). We wish to iterate that this paper does not compare how computer vision fares against insect vision. These are a very different paradigm of information processing.
Bees and wasps are attracted to the shape of the Bee orchids and the pseudo-pheromones produced by the orchids to simulate female insect counterpart pheromones.24 To attract a male, three fundamental stimuli, which are olfactory, visual and tactile, are keys to successful pollination. There is plenty of scholarly literature detailing the pollination of Ophrys species through olfactory signals.5,25–27 Ophrys flowers release chemical compounds that are similar to those released by pollinator females. They make a variety of other compounds and the basic ones, which gives them their specificity. When a male is close to a female, it can visually detect a female’s species-specific optical signals. At this point, visual signals play a crucial role in informing males of the exact location of females (or Ophrys flowers) and reaffirming whether they belong to the same species. The males then avoid visited flowers after realizing they are being ‘fooled’.28 Although our results show excellent reliability in terms of the classification, further analysis by looking at the confusion matrix reveals that ML is not 100% perfect for most classification. Although the numbers are pretty low, several images manage to confuse the ML. We would also like to state that, using a more advanced method such as Deep CNN and additional data input, the ML might achieve perfect classification.
Building upon these misclassification images, we can further analyze the characteristics, similarities and differences between these two organisms. 13,have outlined several Ophrys species’ physical characteristics through a detailed micromorphological study of the labella. 29,has described the similarities between Dasyscolia ciliata and O. speculum. However, most comparisons are descriptive, and we believe that our paper might provide a more quantitative approach using ML. Our ML results on the key characteristics using the SURF method clearly outline points that depict the structure of an insect. ML could pick out the antennae, head, thorax and abdomen. The wings are also highlighted but not as the strongest key points in the image. The sepal and the lateral sepal of the O. apifera are highlighted as the main characteristics. While this feature might be helpful in a Pouyannian mimicry strategy to confuse bees, ML uses this as a prominent distinguishing feature for classification. This perhaps has led to a perfect classification and identification of O. apifera by ML.
The basal field is also highlighted as the primary key points in all the Ophrys species tested. Another feature commonly highlighted amongst the tested species is the pseudo-eyes. Our ML output concurs with 30. They stated that the first main characteristics that mimic bees for Ophrys are the shape of the stigmatic cavity, the size of the basal field, the volume, color and position of the pseudo-eyes. Another two key points that ML highlighted is the speculum (for O. insectifera, O. bombyliflora and O. speculum) and hair margin [for O. insectifera and O. speculum). 30,stated that the second set of characteristics is the shape and arrangement of humps (speculum], location, and hair length. The speculum is one of the most interesting key points as it is the most reflective part of the plant with an iridescent color that changes depending on viewing angles.13
The key characteristics of our ML are related to the evolutionary adaptations special to the Orchid family. The median petal in orchids, termed the labellum, differs in size from the two lateral petals. The labellum is usually more prominent and more elaborate in both shape and markings; it is frequently deeply lobed and, in some species, more frequent in family Orchidoideae. The invaginated form toward the proximal end forms a spur-like structure.31 The labellum/lip is the modified petal that is usually involved in false signals. The modification of the labellum in orchids allows several tactics such as pseudo-copulation, pseudo hormone chemicals, rotten smells, and even kettle traps (which falsely offers food) in the Phalaenopsis [slipper orchids). 32,reported how the labellum patterns could confuse the bees, to the extent that after the failed copulation, the bees would actually hover and observe the pattern up to a minute afterward.
In an e-mail conversation with Farah Alia Nordin (personal communication, 20th May 2021], a leading orchid taxonomist in Malaysia, we raised the question of why Ophrys (several hundred species have developed this particular Pouyannian strategy). In contrast, with the vast diversity of orchids in the tropics, none have developed such a strategy (one particular species in Malaysia mimics a slipper (Phalaenopsis), although this is no fault of the orchid, but rather the person who described it). Even though the diversity of orchids in the temperate (more specifically, European countries) is only 1/3 of the orchid species diversity in the tropics, it’s the plant’s specific biomes and habitat that plays a role. Ophrys are terrestrial and are often in the open environment, whereas most orchids in the tropics are composed of mixed between terrestrial and epiphytic. Being in the open requires specific adaptation to attract the pollinators toward them. In the tropics, referring to the slipper orchids, these orchids have kettle traps that offer no rewards to the insects that fall into them. Other genera such as Bulbophyllum produced a rotten meat smell that attracts flies and wasps as their pollinators. Different orchids use different strategies depending on the habitat and origins.
Nonetheless, pollination of Ophrys is not always species-specific; hence hybridization and gene flow between Ophrys species are allowed through the imperfection of ethological and even mechanical barriers.27 The labellum of some species of orchid flowers, such as Ophrys helenae, resembles a dark hole that allows males to rest overnight, which increase the rate of pollination. Since the pollinating interaction is species-specific, pollinators serve as a pre-pollinating differentiation factor. The flowers benefit from the pollinator species’ pre-mating isolation factors to ensure genetic differentiation from similar Ophrys plants.28
In conclusion, using ML is an efficient method to study the key points and highlights of plants morphological study. However, there are several caveats to our methods. First, our sample is only limited to pictures gathered from online databases and are prone to misclassification if careful vetting was not conducted. We believe that if anyone were to replicate this method using actual pictures of both orchids and bees (taken from a set angle and distance), the classification method and the key points and highlights would have produced a much accurate result. Secondly, our sample number is minimal; there are various species of Ophrys with a multitude of designs and colors. Third, we did not distinguish specific bees with their own specific Ophrys species. We would like to encourage other researchers to use our suggested method on a larger scale dataset. We are also working on analyzing the key characteristics of the orchids in the tropical country and conducting a thorough analysis comparison between the labellum structures of epiphytic and terrestrial orchids. We also believe that using a more advanced ML technique such as Keras, Tensorflow, will provide a better accuracy since our method uses the most basic ML using Wolfram Mathematica. In future studies, we are hoping to adopt Computer Vision (CV) to observe the movement pattern of the insects that are being deceived by these orchids.
Supplementary Material
Acknowledgments
We wish to thank the students of BZT 403 2020/2021 for helping with the preliminary testing. We would also like to thank OEM Intipakar Limited and Universiti Sains Malaysia tech services for the help in procuring and using Wolfram Mathematica. This study is funded by USM RUI grant PBIOLOGI/1001/ 8011124.
Funding Statement
We wish to thank the students of BZT 403 2020/2021 for helping with the data collection. We would also like to thank OEM Intipakar Limited and Universiti Sains Malaysia tech services for the help in procuring and using Wolfram Mathematica. This study is funded by USM RUI grant PBIOLOGI/1001/ 8011124 and USM Industry grant 304 /PBIOLOGI /6501144 /I145.
Disclosure Statement
All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript.
Declarations of interest
None.
Data availability statement
The data used in this study is not available at any open-source data deposition.
Supplementary material
Supplemental data for this article can be accessed on the publisher’s website.
Author contributions
Jenny Wong and Nik Fadzly conducted the data collection and experiments. Jenny Wong, Wan Fatma Zuharah and Nik Fadzly co-wrote the manuscript. All authors have agreed to the final submission.
References
- 1.Baguette M, Bertrand JAM, Stevens VM, Schatz B.. Why are there so many bee-orchid species? Adaptive radiation by intra-specific competition for mnesic pollinators. Biol Rev. 2020;95(6):1–6. doi: 10.1111/brv.12633. [DOI] [PubMed] [Google Scholar]
- 2.Pasteur G. A classificatory review of mimicry systems. Annu Rev Ecol Syst. 1982;13(1):169–199. doi: 10.1146/annurev.es.13.110182.001125. [DOI] [Google Scholar]
- 3.Ayasse M, Schiestl FP, Paulus HF, Ibarra F, Francke W. Pollinator attraction in a sexually deceptive orchid by means of unconventional chemicals. Proc R Soc B. 2003;270(1514):517–522. doi: 10.1098/rspb.2002.2271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Schiestl FP, Ayasse M, Paulus HF, Erdmann D, Francke W. Variation of floral scent emission and postpollination changes in individual flowers of Ophrys sphegodes subsp. sphegodes. J Chem Ecol. 1997;23(12):2881–2895. doi: 10.1023/A:1022527430163. [DOI] [Google Scholar]
- 5.STÖKLStoekl J, Schlüter PM, Stuessy TF, Paulus HF, Fraberger R, Erdmann D, Schulz C, Francke W, Assum G, Ayasse M. Speciation in sexually deceptive orchids: pollinator-driven selection maintains discrete odour phenotypes in hybridizing species. Biol J Linn Soc. 2009;98(2):439–451. doi: 10.1111/j.1095-8312.2009.01279.x. [DOI] [Google Scholar]
- 6.Kullenberg B. On the scents and colours of Ophrys flowers and their specific pollinators among the aculeate Hymenoptera. Svensk Bot Tidskr. 1956;50:25–46. [Google Scholar]
- 7.Kullenberg B. Studies in Ophrys pollination. Zool Bidrag Fran Uppsala. 1961;34:1–340. [Google Scholar]
- 8.Spaethe J, Moser WH, Paulus HF. Increase of pollinator attraction by means of a visual signal in the sexually deceptive orchid, Ophrys heldreichii (Orchidaceae). Plant Syst Evol. 2007;264(1–2):31–40. doi: 10.1007/s00606-006-0503-0. [DOI] [Google Scholar]
- 9.Spaethe J, Streinzer M, Paulus HF. Why sexually deceptive orchids have colored flowers. Commun Integr Biol. 2010;3(2):139–141. doi: 10.4161/cib.3.2.10333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Streinzer M, Ellis T, Paulus HF, Spaethe J. Visual discrimination between two sexually deceptive Ophrys species by a bee pollinator. Arthropod Plant Interact. 2010;4(3):141–148. doi: 10.1007/s11829-010-9093-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Streinzer M, Paulus HF, Spaethe J. Floral colour signal increases short-range detectability of a sexually deceptive orchid to its bee pollinator. J Exp Biol. 2009;212(9):1365–1370. doi: 10.1242/jeb.027482. [DOI] [PubMed] [Google Scholar]
- 12.Vereecken NJ, Schiestl FP. On the roles of colour and scent in a specialized floral mimicry system. Ann Bot. 2009;104(6):1077–1084. doi: 10.1093/aob/mcp208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bradshaw E, Rudall P, Devey D, ThomasWestwood M, Glover BJ, Bateman R. Comparative labellum micromorphology of the sexually deceptive temperate orchid genus Ophrys : diverse epidermal cell types and multiple origins of structural colour. Bot J Linn Soc. 2010;162(3):504–540. doi: 10.1111/j.1095-8339.2010.01033.x. [DOI] [Google Scholar]
- 14.Dyer AG, Paulk AC, Reser DH. Colour processing in complex environments: insights from the visual system of bees. Proc R Soc B. 2011;278(1707):952–959. doi: 10.1098/rspb.2010.2412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dyer AG, Garcia JE, Shrestha M, Lunau K. Seeing in colour: a hundred years of studies on bee vision since the work of the Nobel laureate Karl von Frisch. Proc R Soc Vic. 2015;127(1):66–72. doi: 10.1071/RS15006. [DOI] [Google Scholar]
- 16.Lev-Yadun S, Ne’eman G. Does bee or wasp mimicry by orchid flowers also deter herbivores? Arthropod Plant Interact. 2012;6(3):327–332. doi: 10.1007/s11829-012-9199-y. [DOI] [Google Scholar]
- 17.Sonka M, Hlavac V, Boyle R. Image Processing, Analysis, and Machine Vision. Thomson; 2008. UK: Chapman. ISBN 978-0-495-08252-1. [Google Scholar]
- 18.Klette R. Concise computer vision. New York: Springer; 2014. [Google Scholar]
- 19.Bishop C. Pattern Recognition and Machine Learning. New York: Springer; 2006. https://www.springer.com/gp/book/9780387310732. [Google Scholar]
- 20.Weinstein BG, Prugh L. A computer vision for animal ecology. J Anim Ecol. 2018;87(3):533–545. doi: 10.1111/1365-2656.12780. [DOI] [PubMed] [Google Scholar]
- 21.Gogul I, Kumar VS (2017). Flower species recognition system using convolution neural networks and transfer learning. 2017 4th International Conference on Signal Processing, Communications and Networking (ICSCN -2017), March 16 – 18, 2017, Chennai, INDIA. [Google Scholar]
- 22.Hafemann LG, Oliveira LS, Cavalin P (2014). Forest species recognition using deep convolutional neural networks. 2014 22nd International Conference on Pattern Recognition (ICPR 2014): Stockholm, Sweden. [Google Scholar]
- 23.Turkoglu M, Hanbay D. Leaf-based plant species recognition based on improved local binary pattern and extreme learning machine. Physica A. 2019;527:121297. doi: 10.1016/j.physa.2019.121297. [DOI] [Google Scholar]
- 24.Ayasse M. Chemical mimicry in sexually deceptive orchids of the genus Ophrys. Phyton. 2007;46:221–223. [Google Scholar]
- 25.Gögler J, Stökl J, Sramkova A, Twele R, Francke W, Cozzolino S, Cortis P, Scrugli A, Ayasse M. MÉNAGE À TROIS-TWO ENDEMIC SPECIES OF DECEPTIVE ORCHIDS AND ONE POLLINATOR SPECIES. Evolution. 2009;63(9):2222–2234. doi: 10.1111/j.1558-5646.2009.00712.x. [DOI] [PubMed] [Google Scholar]
- 26.Schiestl FP. On the success of a swindle: pollination by deception in orchids. Naturwissenschaften. 2005;92(6):255–264. doi: 10.1007/s00114-005-0636-y. [DOI] [PubMed] [Google Scholar]
- 27.SCHLÜTERSchlueter PM, Ruas PM, Kohl G, Ruas CF, Stuessy TF, Paulus HF. Genetic patterns and pollination in Ophrys iricolor and O. mesaritica (Orchidaceae): sympatric evolution by pollinator shift. Bot J Linn Soc. 2009;159(4):583–598. doi: 10.1111/j.1095-8339.2009.00957.x. [DOI] [Google Scholar]
- 28.Paulus HF. Deceived males–pollination biology of the Mediterranean orchid genus Ophrys (Orchidaceae). J Eur Orch. 2006;38:303–353. [Google Scholar]
- 29.Paulus HF. Speciation, pattern recognition and the maximization of pollination: general questions and answers given by the reproductive biology of the orchid genus Ophrys. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2019;205(3):285–300. doi: 10.1007/s00359-019-01350-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Romolini R, Soca R. New species in Ophrys (Orchidaceae) to the Italian and French florae. J Eur Orch. 2011;43:759–784. [Google Scholar]
- 31.Bell AK, Roberts DL, Hawkins JA, Rudall PJ, Box MS, Bateman R. Comparative micromorphology of nectariferous and nectarless labellar spurs in selected clades of subtribe Orchidinae (Orchidaceae). Bot J Linn Soc. 2009;160(4):369–387. doi: 10.1111/j.1095-8339.2009.00985.x. [DOI] [Google Scholar]
- 32.Stejskal K, Streinzer M, Dyer A, Paulus HF, Spaethe J, Borges RM. Functional significance of labellum pattern variation in a sexually deceptive orchid (Ophrys heldreichii): evidence of individual signature learning effects. PLoS One. 2015;10(11):e0142971. doi: 10.1371/journal.pone.0142971. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data used in this study is not available at any open-source data deposition.