

Abstract
The immune system plays a major role in the protection against cancer. Identifying and characterizing the pathways mediating this immune surveillance are thus critical for understanding how cancer cells are recognized and eliminated. Aneuploidy is a hallmark of cancer, and we previously found that untransformed cells that had undergone senescence due to highly abnormal karyotypes are eliminated by natural killer (NK) cells in vitro. However, the mechanisms underlying this process remained elusive. Here, using an in vitro NK cell killing system, we show that non‐cell‐autonomous mechanisms in aneuploid cells predominantly mediate their clearance by NK cells. Our data indicate that in untransformed aneuploid cells, NF‐κB signaling upregulation is central to elicit this immune response. Inactivating NF‐κB abolishes NK cell‐mediated clearance of untransformed aneuploid cells. In cancer cell lines, NF‐κB upregulation also correlates with the degree of aneuploidy. However, such upregulation in cancer cells is not sufficient to trigger NK cell‐mediated clearance, suggesting that additional mechanisms might be at play during cancer evolution to counteract NF‐κB‐mediated immunogenicity.
Keywords: aneuploidy, complex karyotypes, immune clearance, NF‐κB, senescence
Subject Categories: Cell Cycle, Immunology
This study shows that aneuploid cells upregulate NF‐κB pathway. This is crucial for promoting an inflammatory state, which in turn leads to NK cell‐mediated clearance of aneuploid senescent cells.

Introduction
Aneuploidy is defined as a state in which the chromosome number is not a multiple of the haploid complement (Pfau & Amon, 2012). In all organisms analyzed to date, an unbalanced karyotype has detrimental effects (Pfau & Amon, 2012; Santaguida & Amon, 2015). In yeast, aneuploidy leads to proliferative defects and proteotoxic stress (Torres et al, 2010). The impact of aneuploidy on higher eukaryotes is even more severe. Most single autosomal gains and all autosomal losses cause embryonic lethality. Aneuploidies that do survive embryonic development cause significant anatomical and physiological abnormalities (Lindsley et al, 1972; Lorke, 1994; Hassold & Hunt, 2001; Roper & Reeves, 2006). The severe impact of aneuploidy on mammalian physiology is also reflected at the cellular level. Trisomic mouse embryonic fibroblasts (MEFs) and aneuploid human cells proliferate more slowly than their euploid counterparts and experience a variety of cellular stresses (Williams et al, 2008; Stingele et al, 2012; Santaguida et al, 2015; Pfau et al, 2016). Among these, aneuploidy‐induced replication stress has been extensively studied. Upon chromosome mis‐segregation, cells exhibit slow replication fork progression rate and increased replication fork stalling during the following S phase. Replication stress triggers genomic instability and drives the evolution of highly abnormal karyotypes (Sheltzer et al, 2012; Ohashi et al, 2015; Lamm et al, 2016; Passerini et al, 2016; Santaguida et al, 2017).
Although aneuploidy is highly detrimental at both the cellular and organismal levels in untransformed cells, it is a hallmark of cancer, a disease characterized by uncontrolled cell proliferation (Gordon et al, 2012). About 90% of solid tumors and 75% of hematopoietic malignancies are characterized by whole chromosome gains and losses (Weaver & Cleveland, 2006). A high degree of aneuploidy is often associated with poor prognosis, immune evasion, and a reduced response to immunotherapy (Ben‐David & Amon, 2020). Given the negative effects of aneuploidy on primary cells, it remains unclear how cells with severe genomic imbalances could gain tumorigenic potential. Furthermore, which aneuploidy‐associated molecular features alter immune recognition during tumor evolution remains an active field of research.
By inducing high levels of chromosome mis‐segregation followed by continuous culturing, we previously generated cells with abnormal complex karyotypes that eventually cease to divide and enter a senescent‐like state. We have named such cells arrested with complex karyotypes (ArCK) cells (Santaguida et al, 2017; Wang et al, 2018). Prior work indicated that ArCK cells upregulate gene expression signatures related to an immune response that render them susceptible to elimination by natural killer (NK) cells in vitro (Santaguida et al, 2017). However, the molecular and functional bases for this immune recognition of ArCK cells remained unclear. Several pathways could be involved in this process. Nuclear factor‐kappaB (NF‐κB) is induced under several stress conditions to elicit a pro‐inflammatory response (Hayden & Ghosh, 2012; Liu et al, 2017). In the canonical NF‐κB pathway, stress induction causes IκB kinase complex (IKK) to phosphorylate IκB, thereby marking it for proteolytic degradation (Perkins, 2007). As a result of this degradation, RelA‐p50 translocates into the nucleus where it activates expression of pro‐inflammatory genes. In the non‐canonical NF‐κB pathway, phosphorylation and cleavage of p100 trigger the nuclear translocation of the RelB‐p52 complex to induce a pro‐inflammatory response (Perkins, 2007). Recent studies further suggest that cytosolic nucleic acids lead to cGAS/STING activation in senescent cells, which induces an interferon response via JAK‐STAT signaling pathway (Glück et al, 2017).
Here, we investigate which innate immune pathways contribute to NK cell‐mediated elimination of aneuploid cells and show that the NF‐κB pathway elicits pro‐inflammatory signals in ArCK cells. Inactivating both canonical and non‐canonical NF‐κB pathways in cells with an unbalanced karyotype prevents NK cell‐mediated killing in vitro. Furthermore, we find that the NF‐κB signature is upregulated in cancer cell lines possessing a higher degree of aneuploidy. However, this activation no longer enhanced NK cell‐mediated killing of cancer cells, raising the possibility that aneuploidy‐induced immunogenicity might be present only at the early stage of tumorigenesis and aneuploid cancer cells evolve mechanisms to evade immune clearance.
Results
An assay to assess elimination of ArCK cells by natural killer (NK) cells in vitro
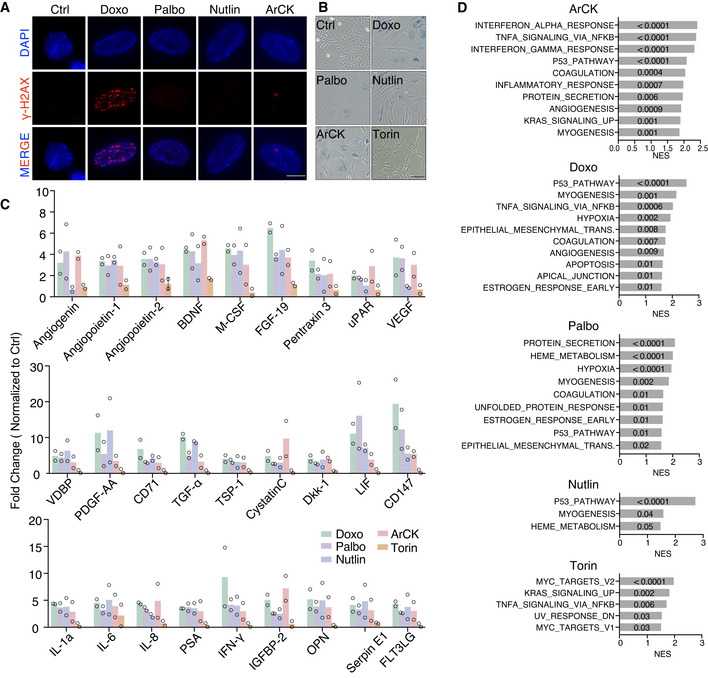
To address the molecular basis for immune recognition of ArCK cells, we established a co‐culture system to monitor the interactions between NK cells and ArCK cells. In this setup, we utilized human, untransformed RPE1‐hTERT cells in which chromosome segregation errors were forced by inhibiting the function of the spindle assembly checkpoint (SAC; Santaguida et al, 2010). To generate ArCK cells, we synchronized RPE1‐hTERT cells at the G1/S boundary and released them into the cell cycle in the presence of the SAC kinase Mps1 inhibitor reversine (Fig 1A). We removed the drug once the cells had undergone one round of aberrant mitosis due to SAC inhibition. 72 h after inducing chromosome mis‐segregation, we exposed cells to the spindle poison nocodazole for 12 h, which allowed us to remove dividing cells by mitotic shake‐off (Santaguida et al, 2017; Wang et al, 2018). We repeated the mitotic shake‐off 4 more times to ensure the removal of all cycling cells. Cells that remained on the tissue culture plate by the end of this procedure were highly enriched for the ArCK population (Fig 1A; Wang et al, 2018). Importantly, such cell cycle arrest was not due to the prolonged nocodazole treatment since euploid control cells were completely removed after two consecutive rounds of shake‐offs (Santaguida et al, 2017). We co‐cultured ArCK cells with an immortalized NK cell line activated by constitutive IL2 expression (NK92‐MI; Tam et al, 1999) and monitored their interactions by live cell imaging (Fig 1B).
Figure 1. ArCK cells are recognized by natural killer (NK) cells in vitro .
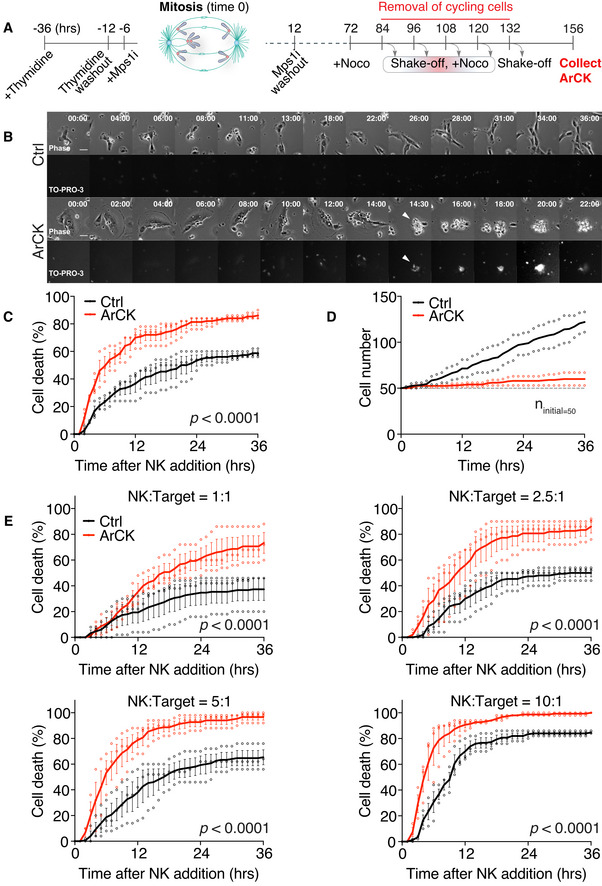
- Schematic representation for the generation of ArCK cells. Time 0 is defined by the estimated onset of Mps1 inhibitor‐induced chromosome mis‐segregation.
- Representative images of euploid control or ArCK cells interacting with NK cells. The NK cell‐mediated killing was measured at a 2.5:1 (NK cells: target cells) ratio and was recorded by live cell imaging for 36 h at a 30‐min interval. TO‐PRO‐3 (1 μM) was added to the medium at the same time of NK cell addition to measure cell membrane integrity. Phase contrast (top) and TO‐PRO‐3 signal (bottom) from the same field were presented. Arrowheads indicate ArCK cell death. All images were acquired at the same exposure time and light intensity. Scale bar 20 μM.
- Measurement of NK cell‐mediated killing of ArCK and euploid control cells (Ctrl) at a 2.5:1 (NK cells: target cells) ratio. 50 randomly chosen target cells were followed for 36 h by live cell imaging per condition per replicate. The cumulative cell death was calculated. n = 3 biological replicates; mean ± SEM. The statistical significance was determined using nonparametric Kolmogorov–Smirnov test (KS test) as described in the method section; P < 0.0001.
- Measurement of euploid control (Ctrl) and ArCK cell proliferation without NK cells. Live cell imaging of target cells without NK cells was performed using the same condition as described in the method section. For each condition, 50 cells were randomly chosen at the beginning of the movie as the initial population (indicated by the dashline, n initial = 50). The cumulative cell number was recorded. Dot plot of individual data points and mean was presented; n = 2 biological replicates.
- NK cell‐mediated cytotoxicity across various NK cell‐to‐target cell ratios. Either euploid control (Ctrl) or ArCK cells were co‐cultured with NK cells at the indicated NK cell: target cell ratios. The cumulative killing of target cells was measured. n = 3 biological replicates; mean ± SEM. P < 0.0001 for all four NK cell: target cell ratios, KS test.
To quantify the degree of NK cell killing of target cells, we first defined a killing event as a target cell that was (i) engaged by one or multiple NK cells, and (ii) lifted from the tissue culture plate (Fig 1B). We chose these criteria because they coincided with target cell membrane permeabilization as judged by the ability of the nucleic acid dye TO‐PRO3 to enter a cell (Figs 1B and EV1A). We tracked individual target cells and recorded the time when each of them was killed during the 36‐h live cell imaging. If a target cell divided during the time course, we followed only one of the resulting two cells for the remainder of the assay. We then calculated the cumulative cell death for each condition and generated killing curves at hourly resolution. We found that at a ratio of 2.5 NK cells to 1 target cell, ArCK cells were consistently killed twice as effectively as euploid control cells, during a 36‐h co‐culture experiment (Fig 1C).
Figure EV1. Characterization of the NK cell killing assay.
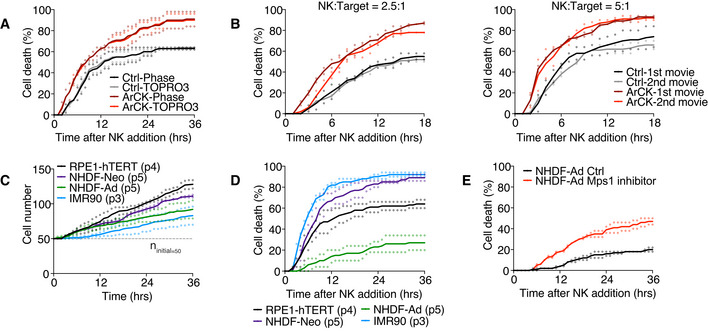
- Side‐by‐side comparison analyzing NK cell‐mediated killing on euploid control or ArCK cells by phase contrast image (Phase) or TO‐PRO3 signal. Cells were cultured as described in Fig 1A. Statistical analyses were performed as in Fig 1C. Individual points and mean were presented; n = 2 biological replicates. Ctrl‐Phase vs. Ctrl‐TOPRO3, P = 0.89, n.s.; ArCK‐Phase vs. ArCK‐TOPRO3, P = 0.89, n.s.; KS test.
- Measurement of NK cell‐mediated killing of ArCK cells in two consecutive 18‐h time lapse experiments. After the first 18 h of the analysis, the cell suspension was collected and co‐cultured with a second set of target cells. NK cell‐mediated killing was measured in the first (black and dark red curves) and the second (gray and light red curves) 18‐h time lapse and plotted on the same graph. The killing assay was performed at a NK cell‐to‐target cell ratio of 2.5:1 (left panel) and 5:1 (right panel); n = 2 biological replicates. NK:Target = 2.5:1, Ctrl‐1st movie vs. Ctrl‐2nd movie, P = 1.00, n.s.; ArCK‐1st movie vs. ArCK‐2nd movie, P = 0.21, n.s.; NK:Target = 5:1, Ctrl‐1st movie vs. Ctrl‐2nd movie, P = 0.28, n.s.; ArCK‐1st movie vs. ArCK‐2nd movie, P = 0.97, n.s.; KS test.
- Cell proliferation measurements in the absence of NK cells. RPE1‐hTERT (passage 4), human normal neonatal or adult human dermal fibroblasts (NHDF‐Neo, passage 5 or NHDF‐Ad, passage 5), and human embryonic lung fibroblast (IMR90, passage 3) were plated side by side in NK cell medium, and cell proliferation rate was recorded using live cell imaging as described in Fig 1D. The dashed line indicates the starting cell number (n initial = 50). Dot plot of individual data points and mean is shown; n = 2 biological replicates.
- NK cell‐mediated cytotoxicity across different cell types. The killing of RPE1‐hTERT, human normal neonatal or adult human dermal fibroblasts (NHDF‐Neo or NHDF‐Ad), and human embryonic lung fibroblast (IMR90) were measured as described in Fig 1 using a NK cell‐to‐target cell ratio of 2.5–1; n = 2 biological replicates. Individual data and mean are shown.
- Human normal adult dermal fibroblasts (NHDF‐Ad) were treated with either DMSO or the Mps1 inhibitor reversine (500 nM) for 24 h. Drugs were washed out, and NK cell‐mediated killing was compared between DMSO‐treated (NHDF‐Ad Ctrl) and Mps1 inhibitor‐treated (NHDF‐Ad Mps1 inhibitor) cells as described in Fig 1C; n = 2 biological replicates. NHDF‐Ad Ctrl vs. NHDF‐Ad Mps1 inhibitor, P = 0.001; KS test.
Arrested with complex karyotypes cells hardly divided during the 36‐hour time lapse employed in our NK cell killing assay whereas euploid control cells continued to divide (Fig 1D). It was thus possible that the difference in NK cell‐mediated cytotoxicity toward euploid and ArCK cells was affected by the fact that NK cells became limiting when co‐cultured with euploid cells but not aneuploid cells. To test this possibility, we analyzed the effect of changing the NK cell‐to‐target cell ratio. We found that even at high NK cell‐to‐target cell ratio (5:1 and 10:1), ArCK cells were still more effectively killed than euploid controls (Fig 1E). We conclude that NK cells were not limited in our assay. We further note that when a cell divided during observation, we followed only one of the two cells after cell division, which corrected for the bias in target cell number. To address the possibility that NK cells became exhausted during the course of the co‐culture experiment, we divided the 36‐h assay into two time courses, where the same population of NK cells was consecutively co‐cultured with target cells for 18 h each. NK cells were equally effective in killing the target cells (Fig EV1B) in this experimental setup, indicating that NK cell exhaustion did not occur within the time course of the analysis. We propose that the eventual plateauing of the killing curve as the assay proceeds is likely due to NK cells taking longer to find their targets.
We next set out to test why euploid control cells are readily killed by NK cells in our in vitro assay. One possible explanation was that RPE1‐hTERT cells express human telomerase reverse transcriptase (hTERT) and harbor a KRAS mutation (Nicolantonio et al, 2008), which could generate oncogenic transformation‐associated NK cell stimulatory signals (Chiossone et al, 2018; Shimasaki et al, 2020). To test this possibility, we assessed NK cell‐mediated cytotoxicity across three different types of early passage euploid primary fibroblasts derived from normal donors (human embryonic lung fibroblast, IMR90, and normal neonatal or adult human dermal fibroblasts, NHDF‐Neo or NHDF‐Ad). The analysis of these primary cells revealed large variations in both cell proliferation and NK cell‐mediated killing (Fig EV1C and D). Adult human dermal fibroblasts were not readily eliminated by NK cells, whereas both neonatal human dermal fibroblasts and IMR90 cells were highly immunogenic. Thus, it appears that NK cell‐mediated killing differs significantly between primary cultured cells. Importantly, we also observed a consistent twofold increase in killing on the Mps1 inhibitor reversine‐induced aneuploid NHDF‐Ad cells compared with their euploid controls (Fig EV1E), indicating that NK cell‐mediated immune clearance of aneuploid cells is not a cell type‐specific phenotype. We conclude that in the assay we developed here, highly aneuploid RPE1‐hTERT cells are more effectively recognized and eliminated by NK92‐MI cells in vitro than their euploid counterparts. Since we had developed robust protocols to generate the aneuploid cell population using RPE1‐hTERT cells (Santaguida et al, 2017; Wang et al, 2018), we decided to focus on this cell line to investigate the effects of karyotype alterations on NK cell‐mediated immune clearance.
Prolonged cell cycle arrest associated with features of senescence elicits NK cell‐mediated cytotoxicity
Arrested with complex karyotypes cells are largely arrested in G1 and exhibit features of senescence (Santaguida et al, 2017; Wang et al, 2018). Permanent cell cycle arrest has been shown to elicit an immune response (Gorgoulis et al, 2019). To determine whether G1 arrest per se is sufficient to cause immune recognition, we assessed NK cell‐mediated cytotoxicity toward G1‐arrested cells induced by three different methods. We treated RPE1‐hTERT cells for 7 days with (i) the topoisomerase II inhibitor, doxorubicin, to induce high levels of DNA damage (Pommier et al, 2010); (ii) the cyclin‐dependent kinases CDK4/6 inhibitor, palbociclib; or (iii) the imidazoline analog, nutlin3, to disrupt the interaction between p53 and its ubiquitin ligase Mdm2, thereby stabilizing p53. All three conditions have been shown to cause features associated with cellular senescence (Sliwinska et al, 2009; Oliveira & Bernards, 2018; Wiley et al, 2018).
DNA content analysis by flow cytometry and EdU incorporation showed that after 7 days, all 3 treatments caused the cells to arrest in G1 (Fig 2A–C). With the exception of nutlin3 treatment, these G1 arrests were irreversible: Most cells did not resume proliferation following drug washout as judged by cell proliferation assays (Fig 2D). Co‐culturing these G1‐arrested cells with NK cells revealed that irrespective of the means by which the arrest was induced, NK cells exhibited a twofold increase in killing on these G1‐arrested cells compared with the untreated proliferating control cells (Fig 2E).
Figure 2. Prolonged cell cycle arrest associated with features of senescence elicits NK cell‐mediated cytotoxicity.
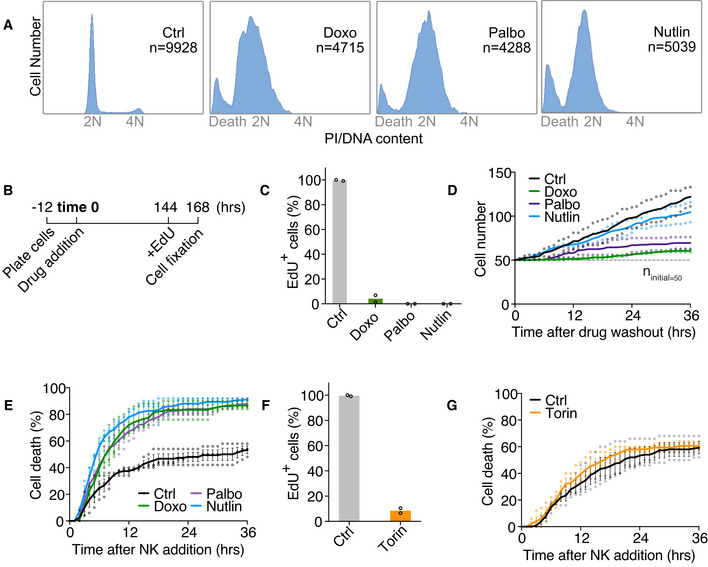
- DNA content analysis of various G1 arrests. RPE1‐hTERT cells were treated for 7 days with doxorubicin (Doxo; 100 ng/ml), palbociclib (Palbo; 5 μM), or nutlin3 (Nutlin; 10 μM). Total number of cells analyzed is indicated by n in each condition. Results were comparable between 2 biological replicates.
- Schematics of EdU incorporation assay. Drugs were applied to RPE1‐hTERT cells 12 h after initial cell plating. 6 days later (144 h), cells were switched to drug medium containing 5‐ethynyl‐2’‐deoxyuridine (EdU; 10 μM) for 24 h before fixation and analysis.
- The percentage of EdU‐positive cells after doxorubicin (Doxo), palbociclib (Palbo), or nutlin3 (Nutlin) treatment. EdU incorporation was performed as described in (B). At least 100 cells were analyzed per condition per replicate. Individual data points and mean are shown; n = 2 biological replicates.
- Cell proliferation (in the absence of NK cells) after 7 days of doxorubicin (Doxo), palbociclib (Palbo), or nutlin3 (Nutlin) treatment. The drugs were washed out after 7 days, and the cells were re‐plated for live cell imaging. Cell proliferation was measured as described in Fig 1D. The dashed line indicates the starting cell number (n initial = 50). Dot plot of individual data points and mean are shown; n = 2 biological replicates.
- NK cell‐mediated killing for doxorubicin (Doxo), palbociclib (Palbo), and nutlin3 (Nutlin) treated samples (NK cell: target cell = 2.5:1). n = 3 biological replicates; mean ± SEM. Ctrl vs. Doxo, P < 0.0001; Ctrl vs. Palbo, P < 0.0001; Ctrl vs. Nutlin, P < 0.0001; KS test.
- The percentage of EdU‐positive cells after 7 days of torin1 treatment was determined as described in (B) and (C). Individual data points and mean are shown; n = 2 biological replicates.
- NK cell‐mediated cytotoxicity toward torin1‐treated cells. Torin1‐treated (Torin) cells were generated as described in (F), and the NK cell killing assay was performed as described in Fig 1. n = 3 biological replicates; mean ± SEM. Ctrl vs. Torin, P = 0.79, not significant (n.s.); KS test.
Inactivation of the TORC1 pathway also causes cell cycle arrest (Sousa‐Victor et al, 2015), but cells enter a quiescent state instead of senescence (Sousa‐Victor et al, 2015; Kucheryavenko et al, 2019). RPE1‐hTERT cells were mostly arrested in cell cycle upon treatment with 1 μM of the mTOR kinase inhibitor torin1 after 7 days (Fig 2F). Yet NK cell recognition and killing were not enhanced in cells treated with torin1 (Fig 2G). We conclude that G1 arrest in target cells contributes to NK cell engagement, but only when accompanied by features of senescence.
Mechanisms triggering senescence contribute to NK cell recognition in ArCK cells
The observation that senescence triggered by multiple mechanisms led to NK cell recognition begged the question of what features in aneuploid cells elicit NK cell‐mediated clearance. To address this, we compared a collection of cellular markers contributing to senescence in ArCK cells to those of cells treated with doxorubicin, palbociclib, or nutlin3 for 7 days. First, we assessed DNA damage levels across all conditions by measuring nuclear γ‐H2AX foci (Figs 3A and EV2A). DNA damage can increase the expression of NK cell‐activating receptor (NKG2D) ligands such as MICA and ULBP2, thereby triggering NK cell‐mediated clearance (Raulet & Guerra, 2009). In untreated proliferating control cells, more than 80% of the cells harbored fewer than 10 γ‐H2AX foci per nucleus. As expected, doxorubicin caused significantly higher levels of DNA damage in the euploid cells, such that approximately 90% of the cells displayed more than 20 foci and ˜50% of this population had 50 foci or more (Fig 3A, panel 2). In contrast, the DNA damage levels in palbociclib‐ and nutlin3‐treated cells were comparable to untreated control cells (Fig 3A, panels 3 and 4). About one third of the ArCK cells harbored more than 10 foci (Fig 3A, panel 5), likely caused by replication stress and/or endogenous reactive oxygen species (ROS) associated with aneuploidy (Li et al, 2010; Passerini et al, 2016; Santaguida et al, 2017).
Figure 3. Mechanisms triggering senescence contribute to NK cell recognition in ArCK cells.
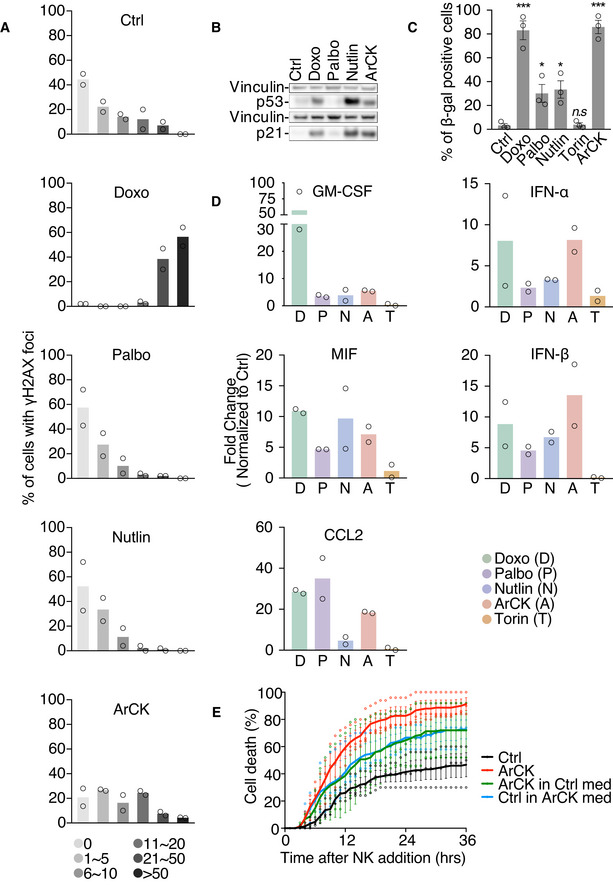
- γ‐H2AX foci were analyzed in ArCK‐ and G1‐arrested cells (generated as described in Fig 2). At least 50 cells were analyzed per condition per replicate. n = 2 biological replicates; individual values and mean are shown. The distribution of the foci number in each treated condition was compared to that of control using Kolmogorov–Smirnov test. Ctrl vs. Doxo, P < 0.0001; Ctrl vs. Palbo, P = 0.07, n.s.; Ctrl vs. Nutlin, P = 0.11, n.s.; Ctrl vs. ArCK, P = 0.0007.
- p53 and p21 levels were determined by Western blot analysis. Vinculin was used as loading control. Results were comparable between 2 biological replicates.
- The degree of senescence was measured by senescence‐associated β‐galactosidase (β‐gal) activity. The graph shows the percentage of β‐gal‐positive cells. At least 100 cells were analyzed per condition per replicate. n = 3 biological replicates; mean ± SEM. Ctrl vs. Doxo, ***P = 0.0006; Ctrl vs. Palbo, *P = 0.025; Ctrl vs. Nutlin, *P = 0.016; Ctrl vs. Torin, P = 0.806, n.s.; Ctrl vs. ArCK, ***P = 0.0001; unpaired t‐test.
- Secreted cytokine and interferon levels were determined in cell supernatants. Media were collected after 36 h of incubation with cells grown as described in Figs 1 and 2. Cytokine and interferon levels were shown as fold change normalized to euploid control cells. Individual values and mean are shown; n = 2 biological replicates.
- NK cell medium was incubated with either euploid control or ArCK cells for 12 h. At the time of NK cell addition, media were switched between ArCK and euploid control cells (Ctrl). NK cell killing was measured as described in Fig 1C. For reference, NK cell killing of ArCK and euploid control cells (Ctrl) without medium switch were performed side by side and plotted on the graph. Black, euploid control cells without medium switch; red, ArCK cells without medium switch; blue, euploid control cells in ArCK cell condition medium; green, ArCK cells in euploid control cell condition medium. n = 3 biological replicates; mean ± SEM. Ctrl vs. ArCK, P < 0.0001; Ctrl vs. Ctrl in ArCK med, P < 0.0001; Ctrl vs. ArCK in Ctrl med, P < 0.0001; ArCK vs. ArCK in Ctrl med, P = 0.0002; KS test.
Figure EV2. Characterization of aneuploid‐ and G1‐arrested cells.
- Representative images of γ‐H2AX staining in the indicated samples. γ‐H2AX is in red and DNA in blue. Scale bar 10 μm.
- Representative image of senescence‐associated β‐galactosidase staining in the indicated samples. Scale bar 100 μm.
- Analysis of all cytokines secreted by the indicated cells. Cytokine levels are shown as fold change in euploid control cells; n = 2 biological replicates. Individual values and mean are plotted.
- Gene set enrichment analysis (GSEA) for doxorubicin‐treated (Doxo), palbociclib‐treated (Palbo), nutlin3‐treated (Nutlin), torin1‐treated (Torin) and ArCK cells relative to euploid proliferating control cells. Only the top 10 ranked hallmarks are presented in Doxo, Palbo, and ArCK conditions. The normalized enrichment score (NES) is plotted. The numbers on the NES score bar indicate the corresponding p‐values for each hallmark (FDR q value ≤ 0.05).
Induction of the DNA damage response genes p53 and p21 agreed with the presence of γ‐H2AX foci with the obvious exception of nutlin3‐treated cells (as nutlin3 inhibits Mdm2 to stabilize p53 but does not cause endogenous DNA damage; Fig 3B). We also assessed senescence‐associated beta galactosidase activity (SA‐beta‐gal), a biomarker frequently used to assess degree of senescence. Over 80% of beta‐gal‐positive cells were observed in doxorubicin‐treated and ArCK cells. Palbociclib‐ or nutlin3‐treated cells exhibited a milder increase in the levels of SA‐beta‐gal‐positive cells (˜30%), whereas torin1‐treated quiescent cells did not show a significant increase in the proportion of SA‐beta‐gal compared with the untreated control cells (Figs 3C and EV2B). We then examined the senescence‐associated secretory phenotype (SASP), which plays a critical role in immune cell recruitment (Gorgoulis et al, 2019). We found the composition of SASP varied between different G1 arrests (Figs 3D and EV2C). Nevertheless, arrested cells that did elicit NK cell‐mediated killing all secreted a plethora of chemokines and cytokines including factors contributing to NK cell recognition [e.g., CCL2 (Robertson, 2002)], whereas the secretion in torin1‐treated cells remained low. We conclude that ArCK cells attract NK cells, at least in part, by expressing a canonical senescence immune recognition program.
To determine the role of secreted factors in NK cell‐mediated killing on aneuploid cells, we collected the culture medium from aneuploid cells and applied it to euploid control cells (Fig 3E). Medium previously used to culture ArCK cells for 12 h increased NK cell‐mediated cytotoxicity toward euploid cells by ˜1.5‐fold. However, this conditioned medium switch did not completely abolish the differences in NK cell‐mediated killing between aneuploid cells and their euploid counter parts. NK cells were still more efficient at killing ArCK cells than euploid cells that have been cultured in pre‐conditioned medium from aneuploid cells (Fig 3E). This could be due to the fact that secreted factors accumulated to higher levels during the 36‐hour live cell imaging, and/or the possibility that ArCK cell surface features enable their elimination by NK cells. We conclude that conditioned medium provides one or more factors that upregulate NK cell‐mediated killing and that aneuploid cells could generate both cell autonomous and non‐cell‐autonomous signals that render them susceptible to NK cell‐mediated cytotoxicity.
NF‐κB and interferon‐mediated pathways are upregulated in ArCK cells
Based on our notion that both secreted factors and cell surface features of aneuploid cells contribute to NK cell‐mediated killing, we next aimed to determine how NK cell recognition is induced in aneuploid cells. For this, we profiled the gene expression signature of ArCK cells and compared it to those of doxorubicin‐, palbociclib‐, nutlin3‐, and torin1‐treated cells by RNA sequencing. Based on gene set enrichment analysis (GSEA), we found there were common significantly upregulated hallmarks shared by G1 arrests that were associated with features of senescence. For example, the p53 pathway is highly upregulated in ArCK‐, doxorubicin‐, palbociclib‐, and nutlin3‐treated cells, but is absent in the torin1‐treated quiescent cells (Fig 4A). The hallmark gene set “TNFalpha via NF‐κB signaling” was among the most upregulated pathways in aneuploid‐ and doxorubicin‐treated samples (Figs 4A and EV2D). ArCK cells also exhibited increased expression of the interferon alpha and gamma response, immune complement, JAK‐STAT, and interleukin (IL)‐related pathways (Fig 4A and B). In palbociclib‐ and nutlin3‐treated samples, we did observe a mild upregulation of immune‐related signatures, but none of them reached significance (FDR q value ≤ 0.05; Figs 4A and EV2D). Interestingly, even though NF‐κB signaling upregulation was also significant in torin1‐treated cells, they were not recognized by NK cells, suggesting NF‐κB activation alone in quiescent cells is not sufficient to cause NK cell‐mediated cytotoxicity (Figs 4A and EV2D).
Figure 4. NF‐κB pathway is activated in ArCK cells.
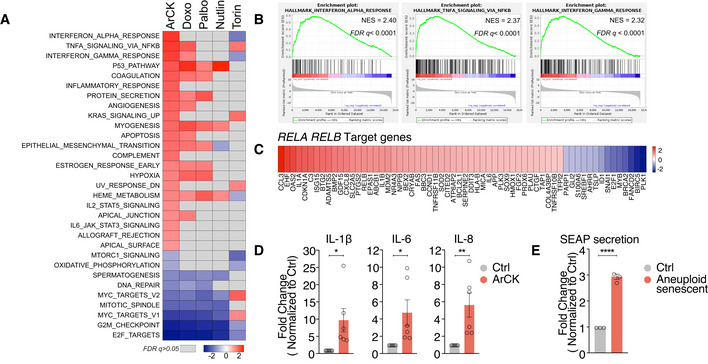
- Significantly differentially expressed hallmarks in ArCK cells compared with euploid control cells are shown in the first column in the heatmap (FDR q value ≤ 0.05). Normalized enrichment scores are plotted. The corresponding NES for these hallmarks in doxorubicin‐treated (Doxo), palbociclib‐treated (Palbo), nutlin3‐treated (Nutlin), and torin1‐treated (Torin) cells is also plotted. Hallmarks did not reach statistical significance (FDR q value > 0.05) are shown in gray.
- Enrichment plots for “Interferon alpha response”, “TNFalpha signaling via NF‐κB”, and “Interferon gamma response” hallmark signatures in ArCK cells compared with the euploid control cells.
- Significantly differentially expressed RELA and RELB target genes in ArCK cells compared with euploid control cells were identified by ingenuity pathway analysis based on RNA sequencing data (Log2 fold change, p‐value 0.05).
- RT–qPCR quantifying NF‐κB downstream target gene expression in ArCK and euploid control (Ctrl) cells. n = 6 biological replicates; mean ± SEM. IL‐1b, *P = 0.029; IL‐6, *P = 0.032; IL‐8, **P = 0.008; unpaired t‐test.
- Measurement of NF‐κB activity with NF‐κB alkaline phosphatase (SEAP) reporter assay. The reporter was expressed in RPE1‐hTERT cells by transient transfection. 10 h after transfection, cells were treated with either DMSO (Ctrl) or reversine (500 nM; Aneuploid senescent) for 96 h and the secretion of alkaline phosphatase in the culture supernatant was measured. The secretion level was normalized to cell number for each condition. n = 3 biological replicates; mean ± SEM; unpaired t‐test, ****P < 0.0001.
Given the importance of NF‐κB pathway in mediating immune recognition, we further characterized this pathway in aneuploid cells. RNA‐seq analysis revealed that both RelA and RelB target genes were significantly upregulated in ArCK cells and this was further confirmed by RT–qPCR analysis (Fig 4C and D). This suggested a possible role of both canonical and non‐canonical NF‐κB pathways. To substantiate NF‐κB activation in aneuploid cells, we employed an NF‐κB reporter assay in which the expression and secretion of alkaline phosphatase (AP) is controlled by an NF‐κB regulatory element (Signorino et al, 2014). Using this assay, we observed a threefold increase in AP secretion 96 h post‐reversine‐induced chromosome mis‐segregation (this would allow for the collection of growth medium at a time point reachable from chromosome mis‐segregation without cell splitting or the loss of the AP conditioned medium, but also reasonably close to the time of ArCK collection; Fig 4E). Furthermore, we also observed a significant increase in the nuclear translocation of RelA in ArCK cells (Fig EV3A and B), in agreement with previous reports showing NF‐κB activation following chromosome mis‐segregation (Vasudevan et al, 2020). Altogether, these results indicate that NF‐κB pathway is indeed upregulated in ArCK cells.
Figure EV3. NF‐κB pathway contributes to NK cell‐mediated killing in ArCK cells.
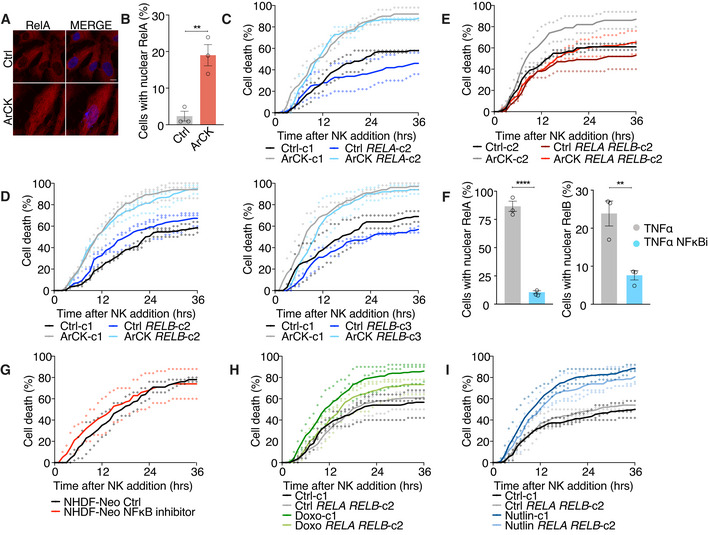
-
A, BRepresentative images (A) and quantification (B) of RelA nuclear translocation signal in ArCK and euploid control cells. At least 86 cells were analyzed per condition per replicate. n = 3 biological replicates; mean ± SEM. **P = 0.0063; unpaired t‐test. Scale bar 10 μm.
-
C–EThe effect of inactivating RELA (C), RELB (D), and both RELA and RELB (E) on NK cell‐mediated cytotoxicity in ArCK cells. The experiment was performed as described in Fig 5, except a different single cell clone was used. ArCK‐c1 vs. ArCK RELA‐c2, P = 0.28, n.s.; ArCK‐c1 vs. ArCK RELB‐c2, P = 0.72, n.s.; ArCK‐c1 vs. ArCK RELB‐c3, P = 0.02; ArCK‐c2 vs. ArCK RELA RELB‐c2, P = 0.0004; KS test. The same controls are plotted in (E) and Fig 5E since all three replicates for both RELA RELB KO clones were performed side by side at the same time.
-
FRPE1‐hTERT cells were treated with either DMSO or the NF‐κB inhibitor BMS‐345541 (5 μM) for 48 h. TNF‐α (100 ng/ml) was added to cells in both conditions for 1 h prior to cell fixation. RelA and RelB nuclear translocation was quantified and shown. n = 3 biological replicates; mean ± SEM. RelA, TNF‐α vs. TNF‐α NF‐κB inhibitor, ****P < 0.0001; RelB, TNF‐α vs. TNF‐α NF‐κB inhibitor, **P = 0.0097; unpaired t‐test.
-
GHuman normal neonatal dermal fibroblasts (NHDF‐Neo) were treated with either DMSO or the NF‐κB inhibitor BMS‐345541 (5 μM) for 48 h. The drug was washed out, and NK cell‐mediated killing was assessed as described in Fig 1C. Dot plot of individual data points and mean is presented in all of the NK cell killing assays; n = 2 biological replicates. NHDF‐Neo Ctrl vs. NHDF‐Neo NF‐κB inhibitor, P = 0.28, n.s.; KS test.
-
H, IThe effect of inactivating both RELA and RELB on NK cell‐mediated cytotoxicity in 7‐day doxorubicin (H) and nutlin3‐treated (I) cells. The experiment was performed as described in Fig 5, except a different single cell clone was used. n = 3 biological replicates; Doxo‐c1 vs. Doxo RELA RELB‐c2, P = 0.04. Nutlin‐c1 vs. Nutlin RELA RELB‐c2, P = 0.18, n.s.; KS test.
Both canonical and non‐canonical NF‐κB pathways are required for NK cell‐mediated killing of ArCK cells
What is the relevance of the NF‐κB pathway activation in NK cell‐mediated elimination of aneuploid cells? To address this question, we generated RELA and RELB single and double KO cell lines using CRISPR‐Cas9 method. In most of the RELA or RELB single knockout clones, we did not observe a significant decrease in NK cell‐mediated cytotoxicity toward ArCK cells (Figs 5A–D and EV3C and D). However, when we knocked out both RELA and RELB, NK cell‐mediated killing in ArCK cells was significantly reduced to a level comparable to the killing of proliferating euploid controls (Figs 5E and F and EV3E). Similar results were observed when ArCK cells were treated with a NF‐κB inhibitor BMS‐345541 that interferes with both catalytic subunits of IKK [albeit with different binding affinities (Burke et al, 2003; Yang et al, 2006)] to block NF‐κB activation (Figs 5G and EV3F). Importantly, the NF‐κB inhibitor treatment in other high immunogenic primary cells (NHDF‐Neo) did not lead to a reduction in NK cell‐mediated killing (Fig EV3G), suggesting that NF‐κB pathway activation in ArCK cells is caused by features associated with aneuploidy induction. We further examined the effect of RELA RELB depletion on NK cell‐mediated killing in senescent cells triggered by other mechanisms. Based on GSEA analysis, doxorubicin‐treated cells also elicit a NF‐κB signature (Fig 4A). Indeed, inactivating NF‐κB pathway in doxorubicin‐treated cells led to a modest but significant decrease in NK cell‐mediated killing (Figs 5H and EV3H). We conclude that DNA damage downstream of aneuploidy could be involved, at least in part, in eliciting the immune response in ArCK cells. On the other hand, RELA and RELB depletion did not rescue NK cell‐mediated killing in nutlin3‐treated cells, which did not exhibit NF‐κB signature (Figs 4A, 5I, and EV3I). This suggests that multiple pathways could be involved in eliciting the immune response, most likely depending on the mechanism leading to cellular senescence. Together, we conclude that aneuploidy triggers NF‐κB activation in primary untransformed cells, which contributes to their recognition and elimination by NK cells.
Figure 5. Both canonical and non‐canonical NF‐κB pathways are required for NK cell‐mediated killing of ArCK cells.
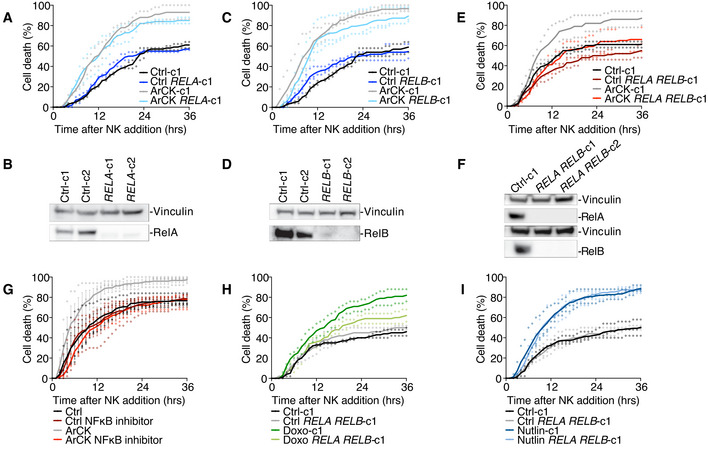
-
ANK cell‐mediated killing of RELA ArCK cells was compared with clones harboring an empty vector. ArCK RELA knockout cells were generated, and the NK cell‐mediated cytotoxicity was measured as described in the Method section. Dot plot of individual data points and mean was presented. n = 2 biological replicates; ArCK‐c1 vs. ArCK RELA‐c1, P = 0.70, n.s.; KS test.
-
BMeasurement of RelA protein levels in RELA KO single cell clones generated in RPE1‐hTERT cells.
-
CThe effect of inactivating RELB on NK cell‐mediated cytotoxicity in ArCK cells. The same experimental methods were used as described in (A). n = 3 biological replicates; ArCK‐c1 vs. ArCK RELB‐c1, P = 0.47, n.s.; KS test.
-
DMeasurement of RelB protein levels in RELB KO single cell clones generated in RPE1‐hTERT cells.
-
EThe effect of inactivating both RELA and RELB on NK cell‐mediated cytotoxicity in ArCK cells. n = 2 biological replicates; ArCK‐c1 vs. ArCK RELA RELB‐ c1, P = 0.0014; KS test.
-
FMeasurement of RelA and RelB protein levels in RELA RELB double KO single cell clones generated in RPE1‐hTERT cells.
-
GArCK or euploid proliferating control cells were treated with either DMSO or the NF‐κB inhibitor BMS‐345541 (5 μM) for 48 h before assessing NK cell‐mediated cytotoxicity. The drug was washed out during the NK cell co‐culture assay. n = 3 biological replicates; mean ± SEM. ArCK vs. ArCK NF‐κB inhibitor, P < 0.0001; KS test.
-
H, IThe effect of inactivating both RELA and RELB on NK cell‐mediated cytotoxicity in 7‐day doxorubicin (H) and nutlin3‐treated (I) cells. n ≥ 2 biological replicates; Doxo‐c1 vs. Doxo RELA RELB‐c1, P = 0.02. Nutlin‐c1 vs. Nutlin RELA RELB‐c1, P = 0.44, n.s.; KS test.
Retrotransposon activation is involved in triggering immune clearance of ArCK cells
ArCK cells also induced interferon alpha and gamma responses as judged by RNA‐seq and RT–qPCR analysis (Fig EV4A and B). Alpha and gamma interferon responses are primarily mediated by the JAK‐STAT pathway (Villarino et al, 2017). We confirmed that the activation of these two gene expression signatures was indeed mediated by the JAK‐STAT pathway as inactivation of STAT1 by CRISPR‐Cas9 reduced both the interferon alpha and interferon gamma responses in ArCK cells (Fig EV4C and D). To determine the biological relevance of the JAK‐STAT response, we depleted STAT1 in ArCK cells. Our data indicate that deletion of STAT1 alone is not sufficient to significantly affect NK cell‐mediated immune clearance in ArCK cells (Fig EV4E). We speculate that JAK‐STAT pathway activation is not essential, but perhaps, potentiates aneuploid cells for immune recognition by NK cells. Remarkably, our data suggest that, although multiple pathways might be activated in aneuploid cells, the NF‐κB pathway plays a major role in NK cell‐mediated immunogenicity of ArCK cells.
Figure EV4. STAT1‐response facilitates immune recognition of ArCK cells.
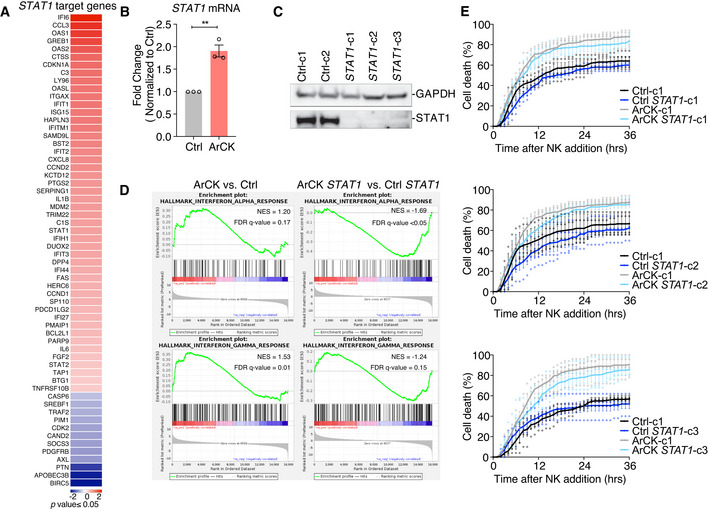
- Significantly differentially expressed STAT1 target genes in ArCK cells compared with euploid control cells were identified by ingenuity pathway analysis based on RNA sequencing data. (Log2 fold change, p‐value ≤ 0.05).
- Measurement of STAT1 mRNA levels in ArCK cells normalized to euploid control cells. n = 3 biological replicates; mean ± SEM. **P = 0.0025; unpaired t‐test.
- Measurement of STAT1 protein levels in STAT1 KO single cell clones generated in RPE1‐hTERT cells.
- GSEA enrichment plot for interferon alpha and interferon gamma hallmarks in ArCK cells generated in either control or STAT1 KO RPE1‐hTERT cells. Two single cell cloned control cell lines and three single cell cloned STAT1 KO cell lines (shown in C) were used in the RNA‐seq analysis.
- ArCK cells lacking STAT1 were generated as described in the Method section. NK cell‐mediated cell death in STAT1 ArCK cells was compared with control cells which harbored an empty vector. n = 4 biological replicates; mean ± SEM. ArCK‐c1 vs. ArCK STAT1‐c1, P = 0.7, n.s.; ArCK‐c1 vs. ArCK STAT1‐c2, P = 0.05, n.s.; ArCK‐c1 vs. ArCK STAT1‐c3, P = 0.14, n.s.; KS test.
Chromosome mis‐segregation also generates micronuclei (Janssen et al, 2011; Crasta et al, 2012; Liu et al, 2018; Martin & Santaguida, 2020). The nuclear envelope of these micronuclei is unstable, leading to their frequent rupture. This in turn causes DNA to spill into the cytoplasm, which activates the cGAS‐STING pathway (Dou et al, 2017; Harding et al, 2017; Mackenzie et al, 2017; Bakhoum et al, 2018). cGAS‐STING could lead to the induction of the NF‐κB and interferon response (Dunphy et al, 2018). However, in our experimental setup, we did not observe a significant cGAS‐STING pathway activation in reversine‐induced aneuploid cells as judged by the low degree of IRF3 phosphorylation (Fig EV5A). Furthermore, STING knockout did not affect NK cell‐mediated killing in ArCK cells (Fig EV5B). Together, these data suggest that, at least in our in vitro assay, NF‐κB is the predominant pathway by which aneuploidy induces NK cell‐mediated cytotoxicity.
Figure EV5. Analysis of cGAS‐STING and retrotransposon activity in ArCK cells and NF‐κB activation in cancer cells following chromosome mis‐segregation.
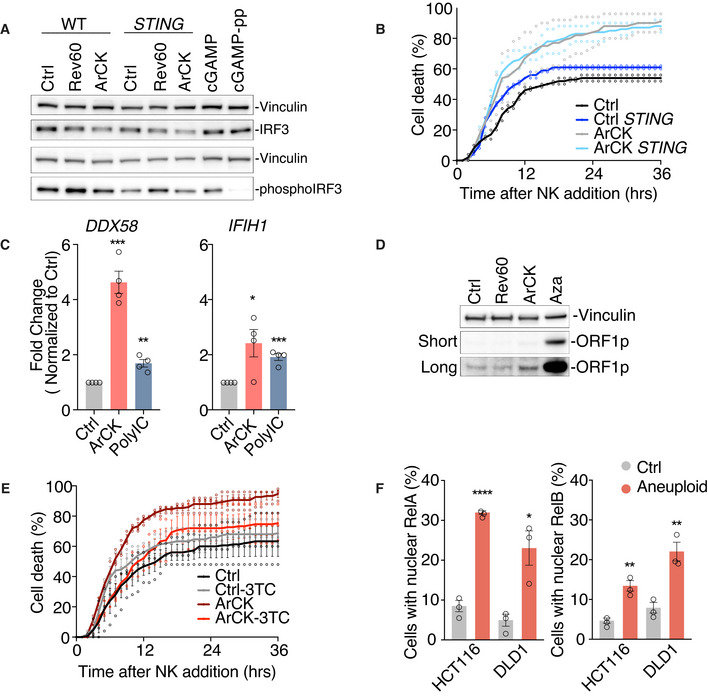
- IRF3 and phospho‐IRF3 levels were analyzed by Western blot in euploid proliferating cells (Ctrl), cells 60 h post‐reversine treatment (Rev60), or ArCK cells that are either functional for STING (lanes 1‐3) or lacking the gene (lanes 4–6). RPE1‐hTERT cells treated with cGAMP (10 μg/ml) for 24 h were used as a positive control (lanes 7 and 8). To confirm the specificity of the phospho‐IRF3‐antibody, protein lysates from cGAMP‐treated cells were incubated with lambda phosphatase (lane 8).
- ArCK STING KO cells were generated, and NK cell‐mediated cytotoxicity was compared with control cell lines that harbor an empty vector as described in Fig 1C; n = 2 biological replicates. ArCK vs. STING ArCK, P = 0.91, n.s.; KS test.
- Measurement of DDX58 and IFIH1 mRNA levels in ArCK cells by RT–qPCR shown as fold change compared with euploid control cells. RPE1‐hTERT cells treated with PolyIC (10 μg/ml) for 24 h were used as a positive control. n = 4 biological replicates; mean ± SEM. DDX58, Ctrl vs. ArCK, ***P = 0.0001; Ctrl vs. PolyIC, **P = 0.0023; IFIH1, Ctrl vs. ArCK, *P = 0.029; Ctrl vs. PolyIC, ***P = 0.0003; unpaired t‐test.
- ArCK cells were grown as described in Fig 1A to determine ORF1 protein levels. RPE1‐hTERT cells treated with azacitidine (Aza, 5 μM) for 5 days (lane 4) were used as a positive control. ORF1p levels under both long exposure and short exposure were presented. Results were comparable between 2 biological replicates.
- The effect of inhibiting reverse transcriptase activity on NK cell‐mediated cytotoxicity in aneuploid cells. ArCK cells were generated as described in Fig 1A and were continuously treated with reverse transcriptase inhibitor 3TC (7.5 μM) following chromosome mis‐segregation. Control RPE1‐hTERT cells were treated with 3TC for 3 days. 3TC was washed out during the NK cell co‐culture assay. n = 3 biological replicates; mean ± SEM. ArCK vs. ArCK‐3TC, P < 0.0001; KS test.
- HCT116 and DLD1 cells were treated with either DMSO (Ctrl) or the Mps1 inhibitor reversine (500 nM) for 48 h (Aneuploid). The percentage of cells with nuclear RelA (left) and nuclear RelB (right) signals were quantified. n = 3 biological replicates; mean ± SEM. RelA, HCT116 Ctrl vs. HCT116 Aneuploid, ****P < 0.0001; DLD1 Ctrl vs. DLD1 Aneuploid, *P = 0.017. RelB, HCT116 Ctrl vs. HCT116 Aneuploid, **P = 0.004; DLD1 Ctrl vs. DLD1 Aneuploid, **P = 0.006; unpaired t‐test.
Senescence is also able to induce retrotransposon activation (Cecco et al, 2019). This could generate dsRNA intermediates to induce both NF‐κB and interferon signatures (Zamanian‐Daryoush et al, 2000; Alexopoulou et al, 2001). Since ArCK cells are senescent (Santaguida et al, 2017; Wang et al, 2018), we tested whether they might be able to trigger retrotransposon activation. We found that in aneuploid cells, two dsRNA sensors, Rig‐I (DDX58) and Mda5 (IFIH1), were upregulated at the mRNA level compared with their euploid control cells (Fig EV5C). Furthermore, ORF1p, one of the proteins encoded by the LINE‐1 retrotransposon, was mildly induced in ArCK cells (Fig EV5D). Thus, upregulation of retrotransposons could be relevant to NK cell recognition of aneuploid cells. To suppress retrotransposition, we inhibited reverse transcription by treating cells with the cytosine analog 2’,3’‐dideoxy‐3’‐thiacytidine (3TC). We found suppressing retrotransposition led to a partial reduction in NK cell‐mediated cytotoxicity toward ArCK cells (Fig EV5E). We conclude that retrotransposon activation in aneuploid cells might be involved, at least partially, in NF‐κB upregulation for NK cell‐mediated immune clearance in ArCK cells.
NF‐κB pathway is upregulated in highly aneuploid cancer cell lines
Our data indicate that in untransformed cells, aneuploidy induction upregulates NF‐κB pathway, which contributes to NK cell‐mediated immune clearance in vitro. Interestingly, in tumor cells high levels of aneuploidy correlate with immune evasion (Davoli et al, 2017; Taylor et al, 2018). It is thus possible that the transformed state of cancer cells suppressed aneuploidy‐induced NF‐κB signaling. To test this hypothesis, we interrogated the association between NF‐κB activation and the degree of aneuploidy in full‐blown tumors using the cancer cell line encyclopedia [CCLE; (Barretina et al, 2012; Ghandi et al, 2019)]. The degree of aneuploidy was scored in almost 1,000 cell lines in the CCLE as recently described (Cohen‐Sharir et al, 2021). We then created two groups of cell lines, a highly aneuploid and a near‐euploid group, defined as the top and bottom quartiles of the number of arm‐level chromosome gains and losses, respectively. To assess NF‐κB activity in these two cell line groups, we created a ssGSEA signature score (Subramanian et al, 2005) for the Hallmark_TNFA_signaling_via_NF‐κB gene set and computed the association between this signature and the degree of aneuploidy by linear regression analysis (see Methods). We found that highly aneuploid cancer cell lines exhibit significantly higher transcriptional signature of NF‐κB activity compared with the near‐diploid lines (Fig 6A and B). This suggests that aneuploidy could also contribute to NF‐κB upregulation in transformed cells. To test whether NF‐κB activation in cancer cell lines also contributes to NK cell‐mediated killing, we induced aneuploidy in the pseudo‐diploid colon cancer cell lines HCT116 and DLD1 using the Mps1 inhibitor reversine. In agreement with previous reports, we observed NF‐κB pathway upregulation following chromosome mis‐segregation in both cancer cell lines indicated by a significant increase in both RelA and RelB nuclear translocation frequencies (Fig EV5F; Vasudevan et al, 2020). However, we did not see an increase in NK cell‐mediated killing in reversine‐induced aneuploid HCT116 and DLD1 cancer cells (Fig 6C). Our data suggest that although an aneuploidy‐associated NF‐κB response may still be evident in transformed cell lines, it is not sufficient to enhance the NK cell‐mediated immune response.
Figure 6. NF‐κB is active in highly aneuploid cancer cell lines.
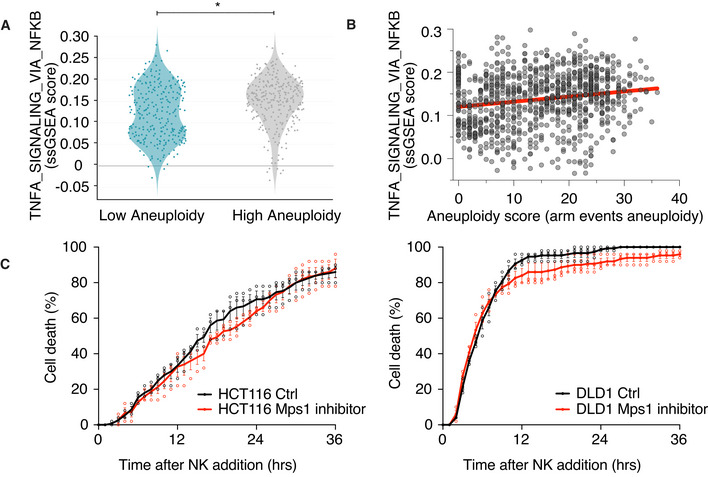
- Comparison of the Hallmark gene expression signature “TNFA_signaling via NF‐κB” in near‐diploid (low aneuploidy) and highly aneuploid (high aneuploidy) human cancer cell lines from the CCLE. *P = 5e‐08, empirical Bayes‐moderated t‐statistics. The y‐axis represents the ssGSEA expression score. The gray line marks an ssGSEA score of 0, indicating the genes in the hallmark “TNFA_signaling via NF‐κB” geneset are not differentially regulated.
- Correlation plot between ssGSEA expression score of “TNFA_signaling via NF‐κB” and the aneuploid score for human cancer cell lines from the CCLE. The trend line for Spearman’s correlation plot is indicated in red. Spearman’s ρ = 0.181, P‐value = 2.2e‐08.
- HCT116 (left) or DLD1 cells (right) were treated with either DMSO or the Mps1 inhibitor reversine (500 nM) for 48 h before assessing NK cell‐mediated cytotoxicity. The drug was washed out during the NK cell co‐culture assay. n = 3 biological replicates; mean ± SEM. HCT116 vs. HCT116 Mps1 inhibitor, P = 0.44, n.s.; DLD1 vs. DLD1 Mps1 inhibitor, P = 0.44, n.s.; KS test.
Discussion
We previously found that untransformed cells that underwent senescence due to highly abnormal karyotypes are recognized by NK cells in vitro. Here, we investigated the molecular mechanism contributing to NK cell‐mediated immune clearance and identified NF‐κB signaling to be central to the interaction between aneuploid cells and NK cells.
The NF‐κB pathway contributes to the immunogenicity of ArCK cells
Multiple studies have shown that senescent cells are recognized and eliminated by NK cells in vitro (Soriani et al, 2009, 2014; Iannello et al, 2013; Sagiv et al, 2016). In this study, we investigated how aneuploidy‐induced senescence causes NK cell recognition. We first tested the hypothesis that G1 arrest elicits an NK cell response by comparing NK cell killing kinetics between various G1 arrests. Our analysis revealed that G1 arrest per se is not sufficient to cause recognition by NK cells because mTOR inhibition, which caused a quiescence‐like G1 arrest, did not elicit an NK cell‐mediated killing. Instead, in accordance with other reported NK cell–senescent cell interactions (Iannello et al, 2013), we found that the senescence‐associated secretory program is primarily responsible for the NK cell‐mediated killing of ArCK cells. Medium harvested from aneuploid cell cultures increased the ability of NK cells to kill euploid cells, suggesting that aneuploid cells can establish a pro‐inflammatory environment where immune clearance takes place.
Our data revealed that NF‐κB pathway upregulation could be one of the major causes of NK cell‐mediated immunogenicity of aneuploid cells in vitro. We observed upregulation of both the canonical and non‐canonical NF‐κB pathways in ArCK cells, and inactivation of both pathways, but not of either one of these pathways alone, is sufficient to protect them from NK cell‐mediated killing in vitro.
Could other immune response inducing pathways contribute to aneuploid cell recognition by NK cells? Chromosome mis‐segregation has been linked to induction of an interferon response (Mackenzie et al, 2017; Vasudevan et al, 2020). Although we observed an upregulation of the alpha and gamma interferon response in ArCK cells, our data indicate that inactivation of STAT1, the major transcription factor mediating the interferon response, did not significantly affect the elimination of ArCK cells by NK cells. Interestingly, STAT1 activation has been observed in untransformed and cancer cell lines where aneuploidy was induced by continuous exposure to reversine (F. Foijer, personal communication). We speculate that in such experimental setup, persistent DNA damage, which accompanies CIN rather than aneuploidy per se, is likely to be the primary cause for JAK/STAT1 pathway activation.
NF‐κB activation in ArCK cells relies on multiple signals
A key question arising from our findings is what causes NF‐κB activation in aneuploid cells. Micronuclei—a well‐known byproduct of chromosome segregation errors and aneuploidy (Crasta et al, 2012)—do not appear to be a major source of immune pathway activation in ArCK cells. On the other hand, while increased retrotransposition could contribute to NF‐κB activation in aneuploid cells, the aneuploidy‐associated stresses are likely to be the major activators of this immune response. Proteotoxic, oxidative, and genotoxic stresses are defining features associated with the aneuploid state [reviewed in (Santaguida & Amon, 2015)]. We previously found that the surface molecules MICA, MICB, CD155, CD112, ULBP1, and ULBP2—that mediate NK cell recognition—are subtly (about 2‐fold) upregulated in ArCK cells (Santaguida et al, 2017). MICA and MICB are activated in response to proteotoxic stress, CD112 (also known as Nectin‐2) and CD155 (also known as PVR) are expressed in response to DNA damage, and ULBP1 and ULBP2 are the product of both cellular stresses and DNA damage (Raulet & Guerra, 2009). This suggested that multiple features of the aneuploid state contribute to the upregulated immunogenicity. We propose that instead of a unique NK cell‐activating feature, the upregulated NK cell‐mediated clearance of aneuploid cells is likely mediated by a combination of stresses elicited by the aneuploid state.
Immune recognition of aneuploidy in cancer
Aneuploidy is a hallmark of cancer that correlates with aggressive disease and immune evasion. Yet in primary cells there is ample evidence that chromosome instability and aneuploidy both elicit a variety of immune responses. For example, micronuclei associated with chromosome mis‐segregation activate the interferon response via the cGAS/STING pathway (Harding et al, 2017; Mackenzie et al, 2017). Chromosome instability upregulates stress‐activated protein kinase (SAPK) and c‐Jun N‐terminal kinase (JNK) pathways, which contribute to inflammatory response (Clemente‐Ruiz et al, 2016; Benhra et al, 2018). MHC complex and antigen processing gene signatures have also been shown to be associated with aneuploidy (Dürrbaum et al, 2014). Hence, a crucial question in the field is how malignant transformation dampens aneuploidy and CIN‐induced immunogenicity. We investigated whether the aneuploidy‐induced NF‐κB signature could be down‐regulated in highly aneuploid cancer cell lines. Our data argue that this was not the case; indeed, the more aneuploid the cancer cell lines, the higher the NF‐κB signaling levels, suggesting that aneuploidy also contributes to NF‐κB upregulation in transformed cells. However, we found that the induction of aneuploidy in pseudo‐diploid cancer cells did not further enhance the NK cell‐mediated killing. We speculate that in transformed cells, other events occurring during tumorigenesis likely serve to counteract the NF‐κB‐mediated immunogenicity and render the cancer cells insensitive for NK cell‐mediated killing. Interestingly, in a tumor environment, the degree of NF‐κB activation inversely correlates with the degree of aneuploidy (Taylor et al, 2018), raising the possibility that silencing of the aneuploidy‐induced immunogenicity could be a non‐cell autonomous event in cancer, perhaps induced by cells in the tumor microenvironment. Understanding which aspects of aneuploidy activate NF‐κB signaling and how the activity of the pathway is modulated during tumor evolution will be the critical next steps in understanding the role of aneuploidy during tumorigenesis.
Materials and Methods
Cell culture
RPE1‐hTERT cells (ATCC Cat# CRL‐4000), HCT116 cells (ATCC Cat# CCL‐247), and HeLa cells (ATCC Cat# CCL‐2) were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen) supplied with 10% FBS (Atlanta Biologicals of South America origin), penicillin/streptomycin (100 U/ml) and L‐Glutamine (2 mM). Human primary IMR90 (ATCC Cat# CCL‐186), adult, and neonatal normal human dermal fibroblasts (NHDF‐Ad and NHDF‐Neo; Lonza Cat# CC‐2511 and Cat# CC‐2509, respectively) were cultured in Eagle’s minimum essential medium (EMEM, ATCC) supplied with 10% FBS, penicillin/streptomycin (100 U/ml) and L‐Glutamine (2 mM). DLD‐1 cells (ATCC Cat# CCL‐221) were cultured in RPMI‐1640 (Invitrogen) supplied with 10% FBS, penicillin/streptomycin (100 U/ml) and L‐Glutamine (2 mM). NK92‐MI cells (ATCC Cat# CRL‐2408) were cultured in MyeloCult H5100 medium (STEMCELL Technologies). All cells were grown at 37°C with 5% CO2 in a humidified environment.
Generation of cells arrested with complex karyotypes (ArCK)
To generate ArCK cells, 2.5 × 105 RPE1‐hTERT cells were plated on a 10‐cm culture dish and synchronized with thymidine (5 mM) for 24 h. Cells were then released into complete medium. 6 h after thymidine release, cells were switched into medium containing reversine (500 nM). Reversine was washed out 18 h later. 60 h after drug washout, nocodazole (100 ng/ml, Sigma‐Aldrich) was added to the culture. 12 h after nocodazole addition, mitotic cells were eliminated from the cell population by shake‐off. The shake‐off process was performed five times in total at a 12‐h interval. The cells left on the plate after five shake‐offs were called the ArCK population.
Generation of cell cycle‐arrested cells
To generate G1 arrested samples, 5 × 105 RPE1‐hTERT cells were plated on a 10‐cm culture dish and treated with the following drugs: doxorubicin (100 ng/ml, Sigma‐Aldrich), palbociclib (5 μM, LC Laboratories), nutlin3 (10 μM, Cayman Chemical), and torin1 (1 μM, LC Laboratories). Cells were collected after 7 days of drug treatment.
Video microscopy
All live cell imaging was performed using a spinning disk microscope (10× objective) with the environmental chamber maintained at 37°C and 5% CO2 level. Target cells were plated onto 12‐well glass‐bottom plates in complete normal growth medium at a density 4–6 × 104 cells/well overnight to allow attachment. Target cells were switched into NK cell growth medium MyeloCult H5100 and incubated for 10 h before starting the live cell imaging. To assess NK cell‐mediated cytotoxicity, NK92‐MI cells were resuspended into target cell condition medium at the indicated NK cell‐to‐target cell ratio immediately before the start of the NK cell killing assay. The cell mixture was filmed for 36 h at a 30‐min time interval. To assess target cell growth without NK cells, target cells were filmed using the same imaging setting and time scale except no NK cells were added.
Cell cycle analysis by flow cytometry
Cells were trypsinized and resuspended into 10 ml complete growth medium. After washing twice in cold phosphate‐buffered saline (PBS), cells were fixed and permeablized in 70% ethanol at −20°C overnight. Cells were then pelleted and incubated with RNAse (100 μg/ml, Thermo Fisher) and DAPI (1 μg/ml, Thermo Fisher) for 1 h before flow cytometry analysis.
Immunofluorescence
RPE1‐hTERT cells were seeded onto fibronectin (10 μg/ml, Sigma‐Aldrich)‐coated coverslips at 50‐70% confluency and allowed to attach overnight. For RelA and anti‐Phospho‐histone H2A.X staining, cells were fixed at room temperature with 4% paraformaldehyde in PBS for 15 min and permeabilized with 0.1% Triton X‐100 in PBS for 10 min. Cells were then blocked in 3% bovine serum albumin (BSA) in PBS for 40 min. Cells were incubated with primary antibodies for 90 min at room temperature. The RelB staining protocol was adapted from (Vasudevan et al, 2020). The following primary antibodies were used as follows: anti‐phospho‐histone H2A.X (Cell Signaling Technology #9718, 1:500), anti‐RelA (Santa Cruz Biotechnology sc‐8008 or Abcam ab16502, 1:50), and anti‐RelB (Abcam ab180127 or ab33907, 1:1,000). The following secondary antibodies were used as follows: Donkey anti‐mouse IgG Cy3 (EuroClone, 1:10,000), Alexa Fluor 488 goat anti‐Rabbit IgG (Thermo Fisher, 1:1,000). Either Hoechest or DAPI was used to stain DNA. Images were acquired using a DeltaVision (60×) or Leica SP8 Confocal (AOBS, 63× oil objective) microscope. Acquired images were analyzed with Fiji software.
Western blot analysis
To prepare protein samples, protease inhibitor cocktail (Roche) and phosphatase inhibitor cocktail (Roche) were added to RIPA lysis buffer (Thermo Fisher Scientific) immediately before use. Cells were lysed in cold RIPA buffer, and the lysate concentration was measured by Bradford assay. The lysate was then diluted with loading buffer and heated at 98°C for 5 min. Proteins were resolved on NuPAGE 4‐12% Bis‐Tris gels (Thermo Fisher Scientific) based on the manufacturer’s instructions and transferred onto 0.2‐μm PVDF membranes. Blots were blocked for 1 h at room temperature in OneBlock blocking buffer (Genesee Scientific). Primary antibodies were incubated over night at 4°C. The following primary antibodies were used as follows: anti‐GAPDH (Santa Cruz sc‐365062, 1:1,000), anti‐Vinculin (Sigma‐Aldrich V9131, 1:5,000), anti‐p53 (Santa Cruz sc‐126, 1:200), anti‐p21 (Cell Signaling Technology #2947, 1:1,000), anti‐p65/RelA (Cell Signaling Technology #8242, 1:1,000), anti‐RelB (Abcam #180127, 1:1,000), anti‐Stat1 (Cell Signaling Technology #9175, 1:500), anti‐IRF3 (Cell Signaling Technology, #11904, 1:1,000), anti‐Phospho‐IRF3 (Cell Signaling Technology, #4947, 1:1,000), and anti‐ORF1p (Cell Signaling Technology #88701, 1:1,000).
Beta galactosidase staining
ArCK, doxorubicin, palbociclib, nutlin3, torin1, and untreated euploid proliferating RPE1‐hTERT cells were plated with normal complete growth medium into 6‐well plates at a density of 4 × 105 cells/well. 24 h later, cells were fixed and stained for the β‐Galactosidase activity using a senescence β‐Galactosidase staining kit (Cell signaling technology #9860) following manufacturer’s instructions.
Cytokine measurement
ArCK, doxorubicin, palbociclib, nutlin3, torin1, and untreated euploid proliferating RPE1‐hTERT cells were generated as described above. 8 ml of complete normal growth medium was placed onto cells and incubated for 36 h. The conditioned medium was harvested, and cell debris was eliminated by centrifugation. To determine the levels of secreted cytokines, condition medium was incubated with proteome profiler human XL cytokine array (R&D Systems, ARY022B) and cytokine levels were measured based on manufacturer’s instructions. The levels of interferon alpha and interferon beta in the condition medium were determined using IFN alpha and IFN beta human ELISA kit (Thermo Fisher Scientific, 411001 and 414101, respectively). The total cell number from each sample was measured using a cellometer (Nexcelom). All cytokines, IFN alpha, and IFN beta readings were normalized to cell number.
RNA sequencing and data analysis
Total RNA was purified using RNeasy Mini Kits (QIAGEN) and sequenced on Illumina HiSeq2000. The RNA‐seq data were aligned to a transcriptome derived from the human hg38 primary assembly and an ensembl version 89 annotation with STAR version 2.5.3a (Dobin et al, 2013). Gene expression was summarized using RSEM version 1.3.0 (Li & Dewey, 2011) and SAMtools/1.3 (Li et al, 2009). An integer count table for differential expression analysis and log2 transcripts per million (TPM) with a plus 1 offset for data visualization was prepared with TIBCO Spotfire Analyst (version 7.11.1). Differential expression analysis was done with DESeq2 [version 1.24.0 or version 1.30.0 (Love et al, 2014) running under R (version 3.6.0 or version 4.0.3). Pre‐ranked Gene Set Enrichment Analysis [versions 2‐3.0_beta2, 4.0.3 or 4.1.0, (Subramanian et al, 2005)] was run using the DESeq2 Wald statistic as a ranking metric and gene set collections from ΜsigDB [versions 6.2, 7.0, or 7.2 (Liberzon et al, 2015)]. Three biological replicates were included per condition in the RNA sequencing. The treated samples were compared with the euploid control cells processed and sequenced from the same experiment to avoid batch effect.
Generation of knockout cell lines using the CRISPR‐Cas9 system
Lentiviral CRISPR‐Cas9 plasmid LentiCRISPR_v2‐Puro (Brett Stringer’s Lab) cloned with guide RNA (gRNA) designed by Feng Zhang’s lab from the Broad Institute targeting exons of human RELA (AGCGCCCCTCGCACTTGTAG), RELB (TCGCCGCGTCGCCAGACCGC), and STAT1 (ATTGATCATCCAGCTGTGAC) was purchased from GenScript. CRISPRv2 constructs along with packaging plasmids pMD2.G (Addgene 12259) and psPAX2 (Addgene 12260) were transfected into 293FT cells (Thermo Fisher, Cat# R70007) using TransIT‐LT1 transfection reagent (Mirus). Virus was collected, and the lentiviral titer was estimated by Lenti‐X GoStix Plus (TaKaRa). RPE1‐hTERT cells were plated at ˜60% confluency and infected at a MOI of 1 for RELA, RELB, or STAT1 single knockouts, and at a MOI of 2 for RELA RELB double knockout. Virus was washed out 20 h post‐infection, and the non‐infected cells were selected against by puromycin treatment. LentiCRISPR_v2‐Puro vector without gRNAs was integrated into RPE1‐hTERT cells to generate the control cell line. 2 d after puromycin selection, single cells were sorted into 96 wells and expanded as individual clones. The successful knockout of RELA, RELB, or STAT1 was verified by Western blot analysis. All experiments described above were performed in at least two single cell knockout clones.
RT–qPCR analysis
Total RNA was purified using an RNeasy Mini Kit (QIAGEN). RNA concentration was determined using a NanoDrop. 750 ng of RNA was used for reverse transcription reactions using SuperScript III First‐Strand Synthesis SuperMix (Invitrogen). The mRNA levels were then quantified by qPCR using SYBR Premix Ex Taq (TaKaRa) on Roche Light Cycler. Primer sequences: STAT1‐Forward GATGTTTCATTTGCCACCATCCGTTTTC. STAT1‐Reverse GGCGTTTTCCAGAATTTTCCTTTCTTCC. GAPDH‐Forward CCATGTTCGTCATGGGTGTGAACCATG. GAPDH‐Reverse CCACAGCCTTGGCAGCGCCAGTAGAGG. DDX58_Forward CGGCACAGAAGTGTATATTGGATGCATTC. DDX58_Reverse GGAAGCACTTGCTACCTCTTGCTCTTC.
IFIH1_Forward CTGGGACTAACAGCTTCACCTGGTGTTG. IFIH1_Reverse GCATCTGCAATGGCAAACTTCTTGCATG. To measure NF‐κB target expressions, 500 ng of RNA was retro‐transcribed using OneScript Plus cDNA Synthesis Kit (abm, G236) and the following TaqMan assays were used (Thermo Fisher Scientific): IL‐1b, hs00174097_m1; IL‐6, Hs00985639_m1; IL‐8, hs00174103_m1.
NF‐κB Secreted Alkaline Phosphatase (SEAP) Reporter Assay
A NF‐κB Secreted Alkaline Phosphatase Reporter Assay Kit (Novus biologicals, NBP2‐25286) was used to measure the secretion of the SEAP protein under the control of the NF‐κB promoter. To generate NF‐κB secreted alkaline phosphatase reporter cell line, RPE1‐hTERT cells were plated in a six‐well plate in 2 ml DMEM, containing 10% FBS, L‐Glutamine, and nonessential amino acids. 12 h later, cells were transfected with 1 μg pNF‐κB /SEAP plasmid using 6 μl Lipofectamine 3000 (Invitrogen, L3000008) per well. 10 h after transfection, cells were replaced with medium containing either reversine (500 nM) or DMSO for 96 h. The supernatant was then collected to measure the levels of SEAP according to the manufacturer’s instructions. The SEAP assay standard curve used to calculate the sample SEAP concentration was generated by loading a serial dilution of SEAP standard on the same plate. The absorbance was measured after 1 h of incubation with the PNPP substrate using a PHERAStar FSX Microplate Reader (BMG LABTECH). SEAP secretion levels were then normalized to cell number for each condition.
CCLE data analysis
Gene expression data for the CCLE lines were obtained from DepMap 19Q1 DepMap release (www. DepMap.org). A ssGSEA signature (Subramanian et al, 2005) score was calculated for the Hallmark_TNFA_signaling_via_NF‐κB gene set. Aneuploidy scores were obtained from Ref.(Cohen‐Sharir et al.). The association between the signature score and the aneuploidy groups was assessed by linear regression analysis, using the R package limma (Ritchie et al, 2015). Significance was calculated by empirical Bayes‐moderated t‐statistics.
Statistical analysis
To test statistical significance on the killing assay, the raw data corresponding to the time points when a target cell was killed by NK cells were pooled from individual biological replicates for each condition. The cells that were not killed at the end of the 36‐h killing assay were assigned to 36 h as the killing time. The distribution of the killing time in each condition was compared, and the significance was determined using nonparametric Kolmogorov–Smirnov test (KS test). Significance was called at P < 0.05. All statistical analysis was performed using GraphPad Prism or R software. Details of the statistical tests on data besides the killing assay were stated in the associated figure legends.
Author contributions
RWW, AA, and SS conceptualized the study; RWW, SV, UB‐D, and SS investigated the study; RWW, UB‐D, AA, and SS wrote the manuscript; UB‐D, AA, and SS contributed to funding acquisition and supervised the study. All authors discussed the results and commented on the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Supporting information
Expanded View Figures PDF
Review Process File
Acknowledgements
We thank Floris Foijer for insightful discussions and for sharing data prior to publication. We thank Jacqueline Lees, Iain Cheeseman, Federica Facciotti, and members of the Amon and Santaguida labs for their helpful comments and discussions regarding this project and manuscript. We thank Charlie Whittaker, Dikshant Pradhan, and the Swanson Biotechnology Center for help with the gene expression analysis. This work was supported by grants to S. S. from the Italian Association for Cancer Research (MFAG 2018—ID. 21665 project), Fondazione Cariplo, Ricerca Finalizzata (GR‐2018‐12367077), the Rita‐Levi Montalcini program from MIUR and by the Italian Ministry of Health with Ricerca Corrente and 5 × 1,000 funds and by NIH grant CA206157 to A.A., who was an investigator of the Howard Hughes Medical Institute, the Paul F. Glenn Center for Biology of Aging Research at MIT and the Ludwig Center at MIT. Work in the Ben‐David lab is supported by the Azrieli Foundation, the Richard Eimert Research Fund on Solid Tumors, the Tel Aviv University Cancer Biology Research Center, and the Israel Cancer Association. This work is dedicated to the memory of Angelika Amon.
EMBO reports (2021) 22: e52032.
Data availability
The RNA‐seq data sets generated for this study can be accessed at Gene Expression Omnibus (GEO) database with the accession number GEO: GSE154919 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE154919).
References
- Alexopoulou L, Holt AC, Medzhitov R, Flavell RA (2001) Recognition of double‐stranded RNA and activation of NF‐κB by Toll‐like receptor 3. Nature 413: 732–738 [DOI] [PubMed] [Google Scholar]
- Bakhoum SF, Ngo B, Laughney AM, Cavallo J‐A, Murphy CJ, Ly P, Shah P, Sriram RK, Watkins TBK, Taunk NK et al (2018) Chromosomal instability drives metastasis through a cytosolic DNA response. Nature 553: 467–472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barretina J, Caponigro G, Stransky N, Venkatesan K, Margolin AA, Kim S, Wilson CJ, Lehár J, Kryukov GV, Sonkin D et al (2012) The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 483: 603–607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben‐David U, Amon A (2020) Context is everything: aneuploidy in cancer. Nat Rev Genet 21: 44–62 [DOI] [PubMed] [Google Scholar]
- Benhra N, Barrio L, Muzzopappa M, Milán M (2018) Chromosomal instability induces cellular invasion in epithelial tissues. Dev Cell 47: 161–174 [DOI] [PubMed] [Google Scholar]
- Burke JR, Pattoli MA, Gregor KR, Brassil PJ, MacMaster JF, McIntyre KW, Yang X, Iotzova VS, Clarke W, Strnad J et al (2003) BMS‐345541 is a highly selective inhibitor of IκB kinase that binds at an allosteric site of the enzyme and blocks NF‐κB‐dependent transcription in Mice*. J Biol Chem 278: 1450–1456 [DOI] [PubMed] [Google Scholar]
- Chiossone L, Dumas P‐Y, Vienne M, Vivier E (2018) Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol 18: 671–688 [DOI] [PubMed] [Google Scholar]
- Clemente‐Ruiz M, Murillo‐Maldonado JM, Benhra N, Barrio L, Pérez L, Quiroga G, Nebreda AR, Milán M (2016) Gene dosage imbalance contributes to chromosomal instability‐induced tumorigenesis. Dev Cell 36: 290–302 [DOI] [PubMed] [Google Scholar]
- Cohen‐Sharir Y, McFarland JM, Abdusamad M, Marquis C, Bernhard SV, Kazachkova M, Tang H, Ippolito MR, Laue K, Zerbib J et al (2021) Aneuploidy renders cancer cells vulnerable to mitotic checkpoint inhibition. Nature 590: 486–491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crasta K, Ganem NJ, Dagher R, Lantermann AB, Ivanova EV, Pan Y, Nezi L, Protopopov A, Chowdhury D, Pellman D (2012) DNA breaks and chromosome pulverization from errors in mitosis. Nature 482: 53–58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davoli T, Uno H, Wooten EC, Elledge SJ (2017) Tumor aneuploidy correlates with markers of immune evasion and with reduced response to immunotherapy. Science 355: eaaf8399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Cecco M, Ito T, Petrashen AP, Elias AE, Skvir NJ, Criscione SW, Caligiana A, Brocculi G, Adney EM, Boeke JD et al (2019) L1 drives IFN in senescent cells and promotes age‐associated inflammation. Nature 566: 73–78 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Nicolantonio F, Arena S, Gallicchio M, Zecchin D, Martini M, Flonta SE, Stella GM, Lamba S, Cancelliere C, Russo M et al (2008) Replacement of normal with mutant alleles in the genome of normal human cells unveils mutation‐specific drug responses. Proc Natl Acad Sci USA 105: 20864–20869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dobin A, Davis CA, Schlesinger F, Drenkow J, Zaleski C, Jha S, Batut P, Chaisson M, Gingeras TR (2013) STAR: ultrafast universal RNA‐seq aligner. Bioinformatics 29: 15–21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dou Z, Ghosh K, Vizioli MG, Zhu J, Sen P, Wangensteen KJ, Simithy J, Lan Y, Lin Y, Zhou Z et al (2017) Cytoplasmic chromatin triggers inflammation in senescence and cancer. Nature 550: 402–406 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunphy G, Flannery SM, Almine JF, Connolly DJ, Paulus C, Jønsson KL, Jakobsen MR, Nevels MM, Bowie AG, Unterholzner L (2018) Non‐canonical activation of the DNA sensing adaptor STING by ATM and IFI16 mediates NF‐κB signaling after nuclear DNA damage. Mol Cell 71: 745–760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dürrbaum M, Kuznetsova AY, Passerini V, Stingele S, Stoehr G, Storchová Z (2014) Unique features of the transcriptional response to model aneuploidy in human cells. BMC Genom 15: 139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghandi M, Huang FW, Jané‐Valbuena J, Kryukov GV, Lo CC, McDonald ER, Barretina J, Gelfand ET, Bielski CM, Li H et al (2019) Next‐generation characterization of the Cancer Cell Line Encyclopedia. Nature 569: 503–508 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glück S, Guey B, Gulen MF, Wolter K, Kang T‐W, Schmacke NA, Bridgeman A, Rehwinkel J, Zender L, Ablasser A (2017) Innate immune sensing of cytosolic chromatin fragments through cGAS promotes senescence. Nat Cell Biol 19: 1061–1070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon DJ, Resio B, Pellman D (2012) Causes and consequences of aneuploidy in cancer. Nat Rev Genet 13: 189–203 [DOI] [PubMed] [Google Scholar]
- Gorgoulis V, Adams PD, Alimonti A, Bennett DC, Bischof O, Bishop C, Campisi J, Collado M, Evangelou K, Ferbeyre G et al (2019) Cellular senescence: defining a path forward. Cell 179: 813–827 [DOI] [PubMed] [Google Scholar]
- Harding SM, Benci JL, Irianto J, Discher DE, Minn AJ, Greenberg RA (2017) Mitotic progression following DNA damage enables pattern recognition within micronuclei. Nature 548: 466–470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hassold T, Hunt P (2001) To err (meiotically) is human: the genesis of human aneuploidy. Nat Rev Genet 2: 280–291 [DOI] [PubMed] [Google Scholar]
- Hayden MS, Ghosh S (2012) NF‐κB, the first quarter‐century: remarkable progress and outstanding questions. Gene Dev 26: 203–234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iannello A, Thompson TW, Ardolino M, Lowe SW, Raulet DH (2013) p53‐dependent chemokine production by senescent tumor cells supports NKG2D‐dependent tumor elimination by natural killer cells. J Exp Medicine 210: 2057–2069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janssen A, van der Burg M, Szuhai K, Kops GJPL, Medema RH (2011) Chromosome segregation errors as a cause of DNA damage and structural chromosome aberrations. Science 333: 1895–1898 [DOI] [PubMed] [Google Scholar]
- Kucheryavenko O, Nelson G, von Zglinicki T, Korolchuk VI, Carroll B (2019) The mTORC1‐autophagy pathway is a target for senescent cell elimination. Biogerontology 20: 331–335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamm N, Ben‐David U, Golan‐Lev T, Storchová Z, Benvenisty N, Kerem B (2016) Genomic instability in human pluripotent stem cells arises from replicative stress and chromosome condensation defects. Cell Stem Cell 18: 253–261 [DOI] [PubMed] [Google Scholar]
- Li B, Dewey CN (2011) RSEM: accurate transcript quantification from RNA‐Seq data with or without a reference genome. BMC Bioinformatics 12: 323 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R (2009) The sequence alignment/Map format and SAMtools. Bioinformatics 25: 2078–2079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li M, Fang X, Baker DJ, Guo L, Gao X, Wei Z, Han S, van Deursen JM, Zhang P (2010) The ATM–p53 pathway suppresses aneuploidy‐induced tumorigenesis. Proc Natl Acad Sci USA 107: 14188–14193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liberzon A, Birger C, Thorvaldsdóttir H, Ghandi M, Mesirov JP, Tamayo P (2015) The molecular signatures database hallmark gene set collection. Cell Syst 1: 417–425 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindsley DL, Sandler L, Baker BS, Carpenter ATC, Denell RE, Hall JC, Jacobs PA, Miklos GLG, Davis BK, Gethmann RC et al (1972) Segmental aneuploidy and the genetic gross structure of the Drosophila genome. Genetics 71: 157–184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu S, Kwon M, Mannino M, Yang N, Renda F, Khodjakov A, Pellman D (2018) Nuclear envelope assembly defects link mitotic errors to chromothripsis. Nature 561: 551–555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu T, Zhang L, Joo D, Sun S‐C (2017) NF‐κB signaling in inflammation. Signal Transduct Target Ther 2: 17023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lorke DE (1994) Developmental characteristics of trisomy 19 mice. Cells Tissues Organs 150: 159–169 [DOI] [PubMed] [Google Scholar]
- Love MI, Huber W, Anders S (2014) Moderated estimation of fold change and dispersion for RNA‐seq data with DESeq2. Genome Biol 15: 550 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackenzie KJ, Carroll P, Martin C‐A, Murina O, Fluteau A, Simpson DJ, Olova N, Sutcliffe H, Rainger JK, Leitch A et al (2017) cGAS surveillance of micronuclei links genome instability to innate immunity. Nature 548: 461–465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin S, Santaguida S (2020) Understanding complexity of cancer genomes: lessons from errors. Dev Cell 53: 500–502 [DOI] [PubMed] [Google Scholar]
- Ohashi A, Ohori M, Iwai K, Nakayama Y, Nambu T, Morishita D, Kawamoto T, Miyamoto M, Hirayama T, Okaniwa M et al (2015) Aneuploidy generates proteotoxic stress and DNA damage concurrently with p53‐mediated post‐mitotic apoptosis in SAC‐impaired cells. Nat Commun 6: 7668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Oliveira RL, Bernards R (2018) Anti‐cancer therapy: senescence is the new black. Embo J 37: e99386 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Passerini V, Ozeri‐Galai E, de Pagter MS, Donnelly N, Schmalbrock S, Kloosterman WP, Kerem B, Storchová Z (2016) The presence of extra chromosomes leads to genomic instability. Nat Commun 7: 10754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkins ND (2007) Integrating cell‐signalling pathways with NF‐κB and IKK function. Nat Rev Mol Cell Biol 8: 49–62 [DOI] [PubMed] [Google Scholar]
- Pfau SJ, Amon A (2012) Chromosomal instability and aneuploidy in cancer: from yeast to man. Embo Rep 13: 515–527 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfau SJ, Silberman RE, Knouse KA, Amon A (2016) Aneuploidy impairs hematopoietic stem cell fitness and is selected against in regenerating tissues in vivo. Gene Dev 30: 1395–1408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pommier Y, Leo E, Zhang H, Marchand C (2010) DNA topoisomerases and their poisoning by anticancer and antibacterial drugs. Chem Biol 17: 421–433 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raulet DH, Guerra N (2009) Oncogenic stress sensed by the immune system: role of natural killer cell receptors. Nat Rev Immunol 9: 568–580 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, Smyth GK (2015) limma powers differential expression analyses for RNA‐sequencing and microarray studies. Nucleic Acids Res 43: e47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson MJ (2002) Role of chemokines in the biology of natural killer cells. J Leukocyte Biol 71: 173–183 [PubMed] [Google Scholar]
- Roper RJ, Reeves RH (2006) Understanding the basis for down syndrome phenotypes. Plos Genet 2: e50 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sagiv A, Burton DGA, Moshayev Z, Vadai E, Wensveen F, Ben‐Dor S, Golani O, Polic B, Krizhanovsky V (2016) NKG2D ligands mediate immunosurveillance of senescent cells. Aging 8: 328–344 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santaguida S, Amon A (2015) Short‐ and long‐term effects of chromosome mis‐segregation and aneuploidy. Nat Rev Mol Cell Bio 16: 473–485 [DOI] [PubMed] [Google Scholar]
- Santaguida S, Richardson A, Iyer DR, M’Saad O, Zasadil L, Knouse KA, Wong YL, Rhind N, Desai A, Amon A (2017) Chromosome Mis‐segregation generates cell‐cycle‐arrested cells with complex karyotypes that are eliminated by the immune system. Dev Cell 41: 638–651 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santaguida S, Tighe A, D’Alise AM, Taylor SS, Musacchio A (2010) Dissecting the role of MPS1 in chromosome biorientation and the spindle checkpoint through the small molecule inhibitor reversine. J Cell Biol 190: 73–87 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santaguida S, Vasile E, White E, Amon A (2015) Aneuploidy‐induced cellular stresses limit autophagic degradation. Gene Dev 29: 2010–2021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheltzer JM, Torres EM, Dunham MJ, Amon A (2012) Transcriptional consequences of aneuploidy. Proc Natl Acad Sci USA 109: 12644–12649 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimasaki N, Jain A, Campana D (2020) NK cells for cancer immunotherapy. Nat Rev Drug Discov 19: 200–218 [DOI] [PubMed] [Google Scholar]
- Signorino G, Mohammadi N, Patanè F, Buscetta M, Venza M, Venza I, Mancuso G, Midiri A, Alexopoulou L, Teti G et al (2014) Role of toll‐like receptor 13 in innate immune recognition of group B streptococci. Infect Immun 82: 5013–5022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sliwinska MA, Mosieniak G, Wolanin K, Babik A, Piwocka K, Magalska A, Szczepanowska J, Fronk J, Sikora E (2009) Induction of senescence with doxorubicin leads to increased genomic instability of HCT116 cells. Mech Ageing Dev 130: 24–32 [DOI] [PubMed] [Google Scholar]
- Soriani A, Iannitto ML, Ricci B, Fionda C, Malgarini G, Morrone S, Peruzzi G, Ricciardi MR, Petrucci MT, Cippitelli M et al (2014) Reactive oxygen species– and DNA damage response‐dependent NK Cell activating ligand upregulation occurs at transcriptional levels and requires the transcriptional factor E2F1. J Immunol 193: 950–960 [DOI] [PubMed] [Google Scholar]
- Soriani A, Zingoni A, Cerboni C, Iannitto ML, Ricciardi MR, Di Gialleonardo V, Cippitelli M, Fionda C, Petrucci MT, Guarini A et al (2009) ATM‐ATR–dependent up‐regulation of DNAM‐1 and NKG2D ligands on multiple myeloma cells by therapeutic agents results in enhanced NK‐cell susceptibility and is associated with a senescent phenotype. Blood 113: 3503–3511 [DOI] [PubMed] [Google Scholar]
- Sousa‐Victor P, García‐Prat L, Muñoz‐Cánoves P (2015) Dual mTORC1/C2 inhibitors: gerosuppressors with potential anti‐aging effect. Oncotarget 6: 23052–23054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stingele S, Stoehr G, Peplowska K, Cox J, Mann M, Storchova Z (2012) Global analysis of genome, transcriptome and proteome reveals the response to aneuploidy in human cells. Mol Syst Biol 8: 608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian A, Tamayo P, Mootha Vk, Mukherjee S, Ebert Bl, Gillette Ma, Paulovich A, Pomeroy Sl, Golub Tr, Lander Es et al (2005) Gene set enrichment analysis: A knowledge‐based approach for interpreting genome‐wide expression profiles. Proc Natl Acad Sci USA 102: 15545–15550 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tam YK, Maki G, Miyagawa B, Hennemann B, Tonn T, Klingemann H‐G (1999) Characterization of genetically altered, interleukin 2‐independent natural killer cell lines suitable for adoptive cellular immunotherapy. Hum Gene Ther 10: 1359–1373 [DOI] [PubMed] [Google Scholar]
- Taylor AM, Shih J, Ha G, Gao GF, Zhang X, Berger AC, Schumacher SE, Wang C, Hu H, Liu J et al (2018) Genomic and functional approaches to understanding cancer aneuploidy. Cancer Cell 33: 676–689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres EM, Dephoure N, Panneerselvam A, Tucker CM, Whittaker CA, Gygi SP, Dunham MJ, Amon A (2010) Identification of aneuploidy‐tolerating mutations. Cell 143: 71–83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasudevan A, Baruah PS, Smith JC, Wang Z, Sayles NM, Andrews P, Kendall J, Leu J, Chunduri NK, Levy D et al (2020) Single‐chromosomal gains can function as metastasis suppressors and promoters in colon cancer. Dev Cell 52: 413–428 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villarino AV, Kanno Y, O’Shea JJ (2017) Mechanisms and consequences of Jak–STAT signaling in the immune system. Nat Immunol 18: 374–384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang RW, MacDuffie E, Santaguida S (2018) Generation and isolation of cell cycle‐arrested cells with complex karyotypes. J Vis Exp Jove 134: 57215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weaver BA, Cleveland DW (2006) Does aneuploidy cause cancer? Curr Opin Cell Biol 18: 658–667 [DOI] [PubMed] [Google Scholar]
- Wiley CD, Schaum N, Alimirah F, Lopez‐Dominguez JA, Orjalo AV, Scott G, Desprez P‐Y, Benz C, Davalos AR, Campisi J (2018) Small‐molecule MDM2 antagonists attenuate the senescence‐associated secretory phenotype. Sci Rep 8: 2410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams BR, Prabhu VR, Hunter KE, Glazier CM, Whittaker CA, Housman DE, Amon A (2008) Aneuploidy affects proliferation and spontaneous immortalization in mammalian cells. Science 322: 703–709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang J, Amiri KI, Burke JR, Schmid JA, Richmond A (2006) BMS‐345541 targets inhibitor of κB kinase and induces apoptosis in melanoma: involvement of nuclear factor κB and mitochondria pathways. Clin Cancer Res 12: 950–960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zamanian‐Daryoush M, Mogensen TH, DiDonato JA, Williams BRG (2000) NF‐κB activation by double‐stranded‐RNA‐activated protein kinase (PKR) is mediated through NF‐κB‐inducing kinase and IκB kinase. Mol Cell Biol 20: 1278–1290 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Expanded View Figures PDF
Review Process File
Data Availability Statement
The RNA‐seq data sets generated for this study can be accessed at Gene Expression Omnibus (GEO) database with the accession number GEO: GSE154919 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE154919).