

Abstract
The spread of synthetic gene drives is often discussed in the context of panmictic populations connected by gene flow and described with simple deterministic models. Under such assumptions, an entire species could be altered by releasing a single individual carrying an invasive gene drive, such as a standard homing drive. While this remains a theoretical possibility, gene drive spread in natural populations is more complex and merits a more realistic assessment. The fate of any gene drive released in a population would be inextricably linked to the population’s ecology. Given the uncertainty often involved in ecological assessment of natural populations, understanding the sensitivity of gene drive spread to important ecological factors is critical. Here we review how different forms of density dependence, spatial heterogeneity, and mating behaviors can impact the spread of self-sustaining gene drives. We highlight specific aspects of gene drive dynamics and the target populations that need further research.
Keywords: genetic pest management, underdominance, CRISPR, spatial dynamics, density dependence, population alteration
INTRODUCTION
The past decade has seen an explosion of research on synthetic gene drives—genetic constructs that can spread through populations even when they decrease the viability and/or reproductive ability of individuals that carry them, much like naturally occurring selfish genetic elements (Burt & Trivers 2006). Recent interest in developing and assessing synthetic gene drive constructs has been fueled by their tremendous theoretical potential as tools for decreasing harm caused by vectors of diseases and other pests (Esvelt et al. 2014, Godfray et al. 2017, Gould 2008). For example, if genes that interfere with the pathogen vectoring ability of a mosquito could be linked to a gene drive, both the drive and antipathogen genes would spread and would result in entire populations of vectors unable to transmit the pathogen (Curtis 1968). Gene drives could also be designed to drastically reduce fecundity or viability of drive-homozygous pest individuals, or cause extremely biased sex ratios in the population (Craig et al. 1960), leading to suppression of the pest population as the drive spreads.
The idea of using genetic methods for controlling populations of pests has been around for at least eight decades (Curtis 1968; Serebrovsky 1940, 1969; Vanderplank 1947), but until recently, promising gene drive systems had eluded researchers. For much of the last century, genetic pest management research remained focused on using chromosomal translocations and transposable elements (reviewed in Gould & Schliekelman 2004). During the first decade of this century, research shifted to selfish genetic elements such as meiotic drivers, Medea, naturally occurring homing endonucleases (pre-CRISPR), and engineered underdominance (Macias et al. 2017, Sinkins & Gould 2006). Recent advances in genetic engineering technology, especially the advent of the CRISPR-Cas system, opened up the possibility of creating a host of new, highly potent gene drive systems (reviewed in Champer et al. 2016). While several new gene drive systems based on CRISPR-Cas remain simply as proposed designs (e.g., Champer et al. 2019a, 2020a; Dhole et al. 2019; Min et al. 2017; Noble et al. 2019; Prowse et al. 2017, 2019; Sudweeks et al. 2019), some have been built in yeast, insects, and mice and have been shown to be capable of driving through lab populations (Champer et al. 2018, 2019c, 2020b; DiCarlo et al. 2015; Grunwald et al. 2019; Hammond et al. 2016, 2018; Kyrou et al. 2018; Oberhofer et al. 2019; Pham et al. 2019). Inference about the potential performance of such gene drives in wild populations has so far relied on mathematical models, lab experiments, and empirical assessments of natural selfish genetic elements (e.g., Cash et al. 2019). The general agreement between empirical data and even very simple models (Akbari et al. 2014; Buchman et al. 2018a,b; Champer et al. 2019c, 2020b; Kyrou et al. 2018; Oberhofer et al. 2019; Schmidt et al. 2017; Windbichler et al. 2011) highlights the potential utility of models for anticipating the dynamics of these genetic elements.
Self-sustaining, synthetic gene drives work through one of two general mechanisms. The first type of gene drives biases the inheritance of the element in viable gametes from heterozygous parents. For example, meiotic drivers can propagate by disabling sperm of heterozygous males that do not contain the driver (Lindholm et al. 2016). Homing endonuclease gene (HEG) drives achieve a similar biased inheritance through conversion of wild-type alleles of heterozygous individuals into drive alleles (Godfray et al. 2017). The second general type of gene drives reduces the fitness of the wild-type alleles and thus gains an advantage by biasing selection instead of inheritance. For example, natural and synthetic Medea elements spread by causing the death of either gametes or zygotes that lack them (Akbari et al. 2014, Beeman et al. 1992, Chen et al. 2007). Another example is gene drives based on simple, 1-locus underdominance, which can cause reduced viability in heterozygotes. If an underdominant synthetic allele is more common in a population than the wild-type allele (e.g., due to a large release of transgenics), wild-type alleles suffer higher relative fitness loss than the synthetic alleles, as they are more likely to form unfit heterozygotes (Sinkins & Gould 2006).
Gene drives can further be categorized in terms of other important properties besides the mechanism used to gain an advantage. Detailed descriptions of the general properties of a wide variety of gene drive designs have been compiled by other researchers (Burt 2014, Champer et al. 2016). Two fundamental criteria are (a) the intended effect of the gene drive on the population and (b) the frequency-dependent dynamics of different drives. The intended goal for replacement drives (also known as modification drives) is to bring about a change in the genetic makeup of a natural population without significantly reducing the population size, as with a drive that spreads a pathogen-blocking gene in mosquitoes (e.g., Gantz et al. 2015, Pham et al. 2019). As an alternative to gene replacement, suppression drives aim to reduce the size of a pest population (e.g., Hammond et al. 2016, Kyrou et al. 2018). Gene drives that can cause very strong suppression may even be able to completely eradicate a population and are here subcategorized as eradication drives. As the level of suppression caused by a given gene drive can vary based on the ecological context of a particular population, the subcategorization of certain suppression drives as eradication drives is inexact. Similarly, the separation of gene drives into replacement or suppression categories is not absolute; some replacement drives could also cause some amount of population suppression. Even with these caveats, there is heuristic value in using these categories when discussing how different ecological factors affect the dynamics of gene drives.
Gene drives can also be categorized as threshold drives and non-threshold drives based on their frequency-dependent dynamics (Champer et al. 2016, Leftwich et al. 2018). Non-threshold gene drives exhibit a selective advantage over wild-type alleles irrespective of their frequency and can spread even when they are initially very rare. In contrast, threshold drives suffer a selective disadvantage unless they are present above a certain threshold frequency (i.e., they exhibit bistability). Successful population alteration with threshold drives therefore requires them to be released in large enough numbers to exceed this threshold release frequency or release threshold. Such release thresholds can result from fitness costs incurred by individuals due to carrying the drive alleles or can also be an intrinsic consequence of the drive mechanism. For instance, synthetic Medea constructs and some CRISPR-based toxin-antidote drives exhibit no threshold unless the drive constructs impose a fitness cost on the carriers (even on drive-homozygous individuals) that is independent of the drive mechanism (Champer et al. 2020a,b; Oberhofer et al. 2020; Ward et al. 2011). Even CRISPR-based homing drives can exhibit a release threshold under a limited set of conditions (Alphey & Bonsall 2014, Deredec et al. 2011, Tanaka et al. 2017), but it is unclear if such conditions can be met in natural environments. In contrast, underdominance mechanisms lead to a release threshold that has an intrinsic minimum level even if the drive construct bears no fitness cost (Sinkins & Gould 2006). The intrinsic minimum values for the release thresholds of different underdominance-based gene drives can differ widely based on the genetic structure of the drives (Altrock et al. 2010; Champer et al. 2019a, 2020a,b; Davis et al. 2001; Dhole et al. 2018; Leftwich et al. 2018; Marshall & Hay 2012b; Oberhofer et al. 2019; Ward et al. 2011). Irrespective of whether a gene drive exhibits a nonzero intrinsic minimum release threshold, increasing the fitness costs suffered by gene drive carriers generally substantially increases the actual release threshold of a drive (e.g., Alphey & Bonsall 2014, Altrock et al. 2010, Champer et al. 2020a, Dhole et al. 2018, Ward et al. 2011; but see Dhole et al. 2019) (Figure 1). Consequently, it can be difficult to categorize specific gene drives as non-threshold drives versus threshold drives because environment-dependent changes in fitness can cause some gene drives to exhibit a threshold in some environments and not in others. But even a context-specific categorization can be useful for discussing the behavior of different gene drives.
Figure 1.
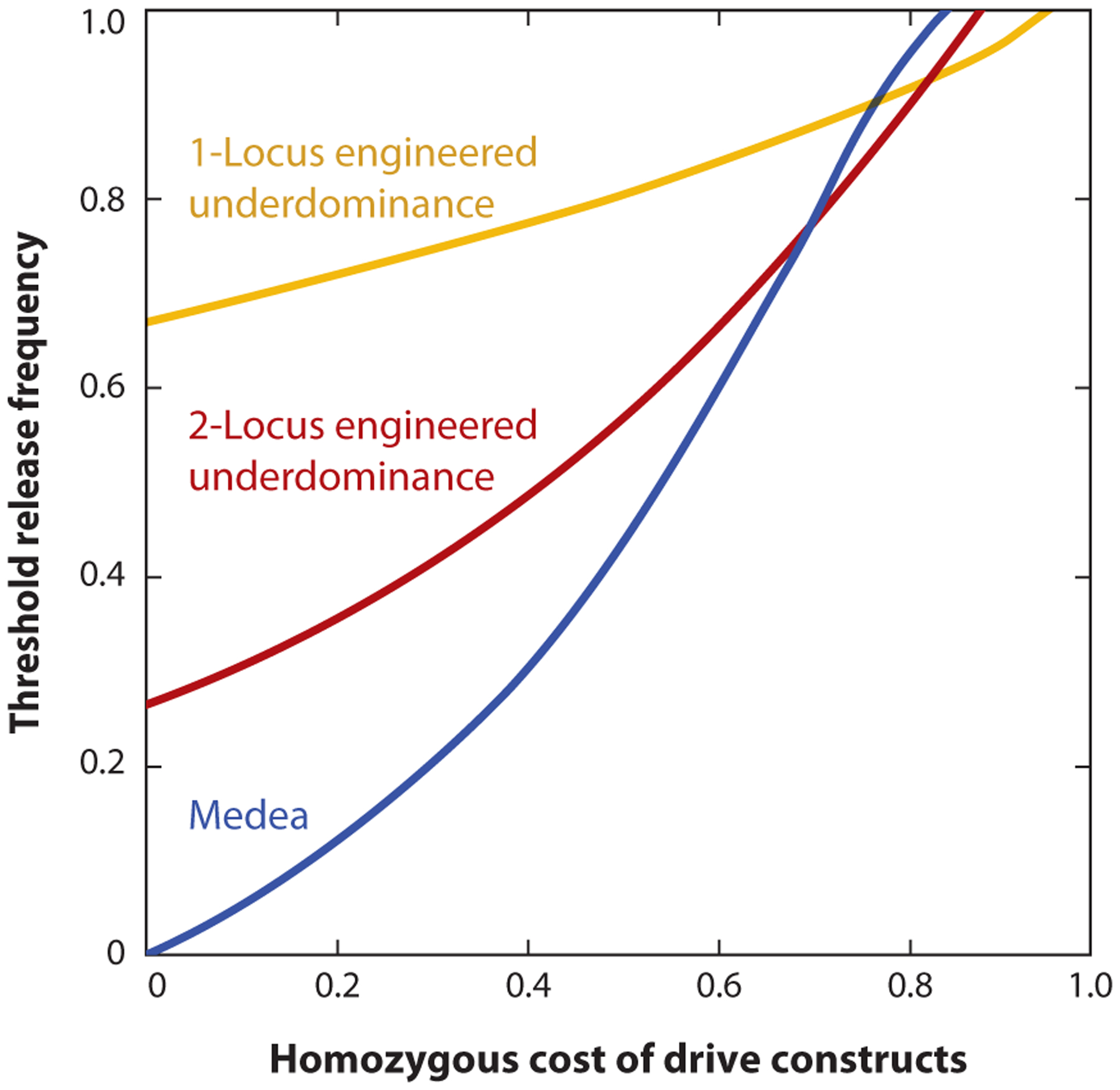
The threshold release frequency of threshold gene drives increases with the fitness cost imposed by the drive constructs. Shown here are the release thresholds for three gene drives—Medea (blue line), 2-locus engineered underdominance (red line), and 1-locus engineered underdominance (yellow line)—as they vary with the fitness cost incurred by individuals homozygous for the drive constructs.
In addition to requiring a larger release effort to exceed the threshold frequency during a release, the dynamics of threshold drives are qualitatively different from those of non-threshold drives, especially when spatial structure is considered (Barton 1979a). Moreover, threshold drives may also be less likely to invade nontarget populations relative to non-threshold drives because a larger cohort of migrants would be needed for surpassing the release threshold in a new population (Gould 2008). When confinement of a gene drive to specific populations is a concern, special caution must be exercised with efforts to use a gene drive that has a release threshold solely due to environment-dependent fitness effects, which may vary widely with time and between different environments (Backus & Delborne 2019).
A few gene drive approaches have been proposed that are temporally self-limiting (e.g., Killer-Rescue, Daisy-chain) (Gould et al. 2008, Noble et al. 2019). These drives are designed to initially spread before removing themselves from the population eventually and are not intended for permanent population alteration. Such drives may still be quite useful in some scenarios. For instance, if a pathogen requires a critical mass of vectors for persistence, a temporary suppression of the vector population below such critical mass may be sufficient to eliminate the pathogen. The long-term dynamics of these drives, especially across space, are significantly different from self-sustaining drives. We focus this review only on self-sustaining gene drives.
In addition to gene drives, the cytoplasmic endosymbiont Wolbachia has been proposed as a tool for altering populations of certain insect disease vectors (Brownstein et al. 2003, Rasgon et al. 2003). These vertically transmitted endosymbionts can induce cytoplasmic incompatibility (CI) between gametes of infected males and uninfected females. While not a gene drive, CI-inducing Wolbachia can spread in an insect population with dynamics that are qualitatively identical to those of many threshold drives and can reduce the transmission of certain insect-borne pathogens (Barton & Turelli 2011, Brownstein et al. 2003, Rasgon et al. 2003). Indeed, some of the most relevant recent work on the spatial spread of threshold drives is rooted in understanding the spread of Wolbachia (e.g., Barton & Turelli 2011).
Over the past decade, many aspects of gene drive technology have been reviewed, including the mechanisms of different gene drives (Burt 2014, Champer et al. 2016), biosafety aspects of gene drives (Marshall & Akbari 2018), diverse gene drive applications (Esvelt et al. 2014, Flores & O’Neill 2018, Gould 2008, Moro et al. 2018, Rode et al. 2019), the history of gene drive research (Macias et al. 2017), gene drive design considerations (Akbari et al. 2019), and the current status of development of certain drives (Leftwich et al. 2018, Raban et al. 2020). A recent review by Hay et al. (2021) discusses progress toward new gene drive mechanisms that are being considered for achieving spatially restricted alteration, the long-term evolutionary stability of gene drives after release, and the hurdles that need to be navigated. Godfray et al. (2017) reviewed the use and dynamics of different endonuclease-based drives, addressing many factors that can affect drive spread in natural populations. David et al. (2013) discussed the downstream effects that gene drives could have on the ecology and evolution of altered populations. A number of committee and workshop reports have also focused on regulatory and practical considerations for gene drive applications (Delborne et al. 2018, Giese et al. 2019, James et al. 2018, Natl. Acad. Sci. Eng. Med. 2016). In this review, we focus on key ecological factors that will play a pivotal role in the application of gene drives to natural populations. We specifically address topics that have not recently been reviewed in detail. These are (a) the effects of different forms of density-dependent population dynamics, (b) spatial structure within and between populations, and (c) mating behavior and sexual selection. We place special emphasis on spatial structure for which there is a great deal of recent and older relevant literature that has not been reviewed in the context of diverse gene drives.
DENSITY-DEPENDENT POPULATION DYNAMICS
Population suppression gene drives can use two approaches for reducing population density (see Burt 2014, Champer et al. 2016, Sinkins & Gould 2006). The drive construct can be designed to disrupt an essential gene or include a harmful cargo gene so as to impose a direct fitness cost on individuals in the form of reduction in viability or fecundity (e.g., Hammond et al. 2018, Kyrou et al. 2018). Alternatively, but not mutually exclusively, the drive could be used to cause an extreme bias in the sex ratio of the offspring produced in the population (e.g., Galizi et al. 2014, 2016; Leitschuh et al. 2018; Prowse et al. 2019). A strongly female-biased sex ratio may reduce the number of progeny produced if many females are unable to find a mate and reproduce, while a male bias reduces the number of females, directly reducing the number of offspring that can be produced. Both approaches, fertility or viability reduction and sex ratio bias, are intended to impose a genetic load on the population, which is a measure of the extent to which the average fitness (reproductive capability) of a genetically altered population is reduced compared to that of a wild-type population, and ranges between 0 (no reduction in population fitness) and 1 (complete reduction).
Different types of gene drives differ in their ability to impose genetic load on a population (Burt 2003, Champer et al. 2016, Deredec et al. 2008, Dhole et al. 2018, Khamis et al. 2018). Gene drives with high intrinsic minimum thresholds due to the drive mechanism (in this review, gene drives with a release threshold greater than 0.5) require a large release effort even when individuals do not bear a fitness cost, and they may be unable to spread at all when they significantly reduce individual fitness. Gene drives with high intrinsic release thresholds are therefore thought to have limited potential for population suppression in most scenarios but may find use for suppression in specific contexts (Champer et al. 2016, Dhole et al. 2018; but for examples of temporary or specific-case population suppression, see Akbari et al. 2013, Khamis et al. 2018). Gene drives with potential for having low or no release thresholds are generally able to spread even when they lower fitness of individuals, so they can impose a higher maximum equilibrium genetic load (e.g., Dhole et al. 2018; but see Champer et al. 2019a, 2020a; Dhole et al. 2019), making low (or zero) threshold drives especially powerful for population suppression (Alphey 2014, Burt 2003, Champer et al. 2016, Lambert et al. 2018, Leftwich et al. 2018, Sinkins & Gould 2006). While it is clear that a higher genetic load would tend to cause a stronger population suppression, exactly how strongly a given amount of genetic load can suppress a population is a far more complex question.
How a given amount of genetic load translates to population suppression critically depends on, among other factors, the extent and form of density-dependent dynamics in the population (e.g., Alphey & Bonsall 2014, Deredec et al. 2011). When intraspecific competition for resources is strong, reduction in population density is usually accompanied by increasing growth rates as the per capita resource availability increases (May 1973). This increase in growth rate with decreasing density can partially compensate for the suppressing effect of the genetic load.
The exact nature of this density-dependent response (e.g., strength and shape of the response) plays a critical role in determining how the equilibrium size of a population would change after imposition of a genetic load through a gene drive. Imagine a population where growth is restricted only by the level of resource competition so that the per capita growth rate is highest at an extremely low density and decreases to zero at the equilibrium size of the population (Figure 2a). All three curves in this figure exhibit this property but differ in the rate at which the per capita growth rate changes at a given density. If a population exhibits simple logistic density-dependent dynamics, so that the per capita growth rate changes linearly with density (curve ii in Figure 2a), the size at equilibrium of that population would decrease linearly with the genetic load imposed by a gene drive at equilibrium (curve ii in Figure 2b). Alternatively, in a population where the per capita growth rate decreases only slightly with increasing numbers at low densities, but very strongly as density increases to the equilibrium value (curve i in Figure 2a), a small reduction in density from the equilibrium value would cause very strong compensation in the growth rate (May 1973). In a population that exhibits such density dependence, a much higher genetic load would be needed to achieve a similar level of suppression when compared to the logistic dynamics scenario (curve i in Figure 2b). Conversely, if the per capita growth rate responds strongly to competition at low densities and is affected less by further increase in density (curve iii in Figure 2a), even a small amount of genetic load may be sufficient to achieve a large amount of suppression (curve iii in Figure 2b). This is because there is little compensation in the per capita growth rate as the population is reduced from its equilibrium (curve iii in Figure 2a). For example, a model simulating a hypothetical mosquito population that exhibits density dependence similar to curve iii was used to predict that an engineered underdominance drive imposing an equilibrium genetic load around 20–30% can cause close to 80% suppression of the mosquito population (Khamis et al. 2018, figure 3), resulting in a 50% reduction in disease transmission (Khamis et al. 2018, figure 4). In a population that exhibits logistic density-dependent dynamics, a similar drive (when released above its release threshold) would cause only around 20–30% suppression at equilibrium (e.g., curve ii in Figure 2b). Factors that influence what form of density-dependent dynamics are exhibited by a given species (or population) are complex (Clutton-Brock et al. 1997, Herrando-Pérez et al. 2012, Lindström & Kokko 2002, Sinclair 2003). Understanding the density-dependent dynamics in the target population will be critical not only for accurate forecasting of the effect of a drive but also for choosing and designing the gene drive that can achieve the target level of suppression.
Figure 2.
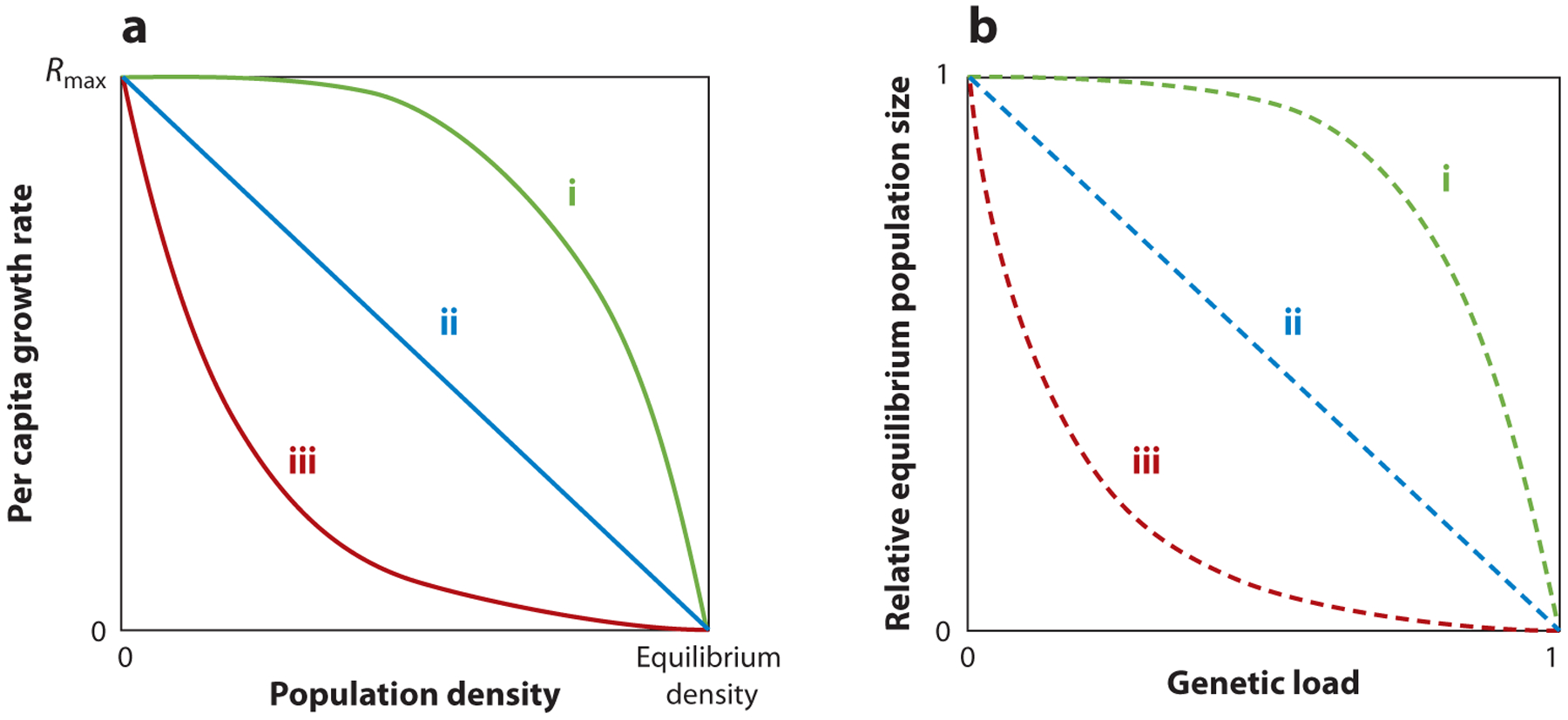
(a, solid curves) A highly simplified depiction of three forms of density-dependent dynamics is shown, along with (b, dashed curves) corresponding curves for the relative equilibrium population size reached with a given amount of genetic load. Rmax denotes the maximum per capita growth rate at a low density. Equilibrium density refers to the equilibrium population density of a fully wild-type population. In both panels, curve i is an overcompensatory response, curve ii is a linear response (e.g., simple logistic), and curve iii is an undercompensatory response to application of genetic load.
Figure 3.
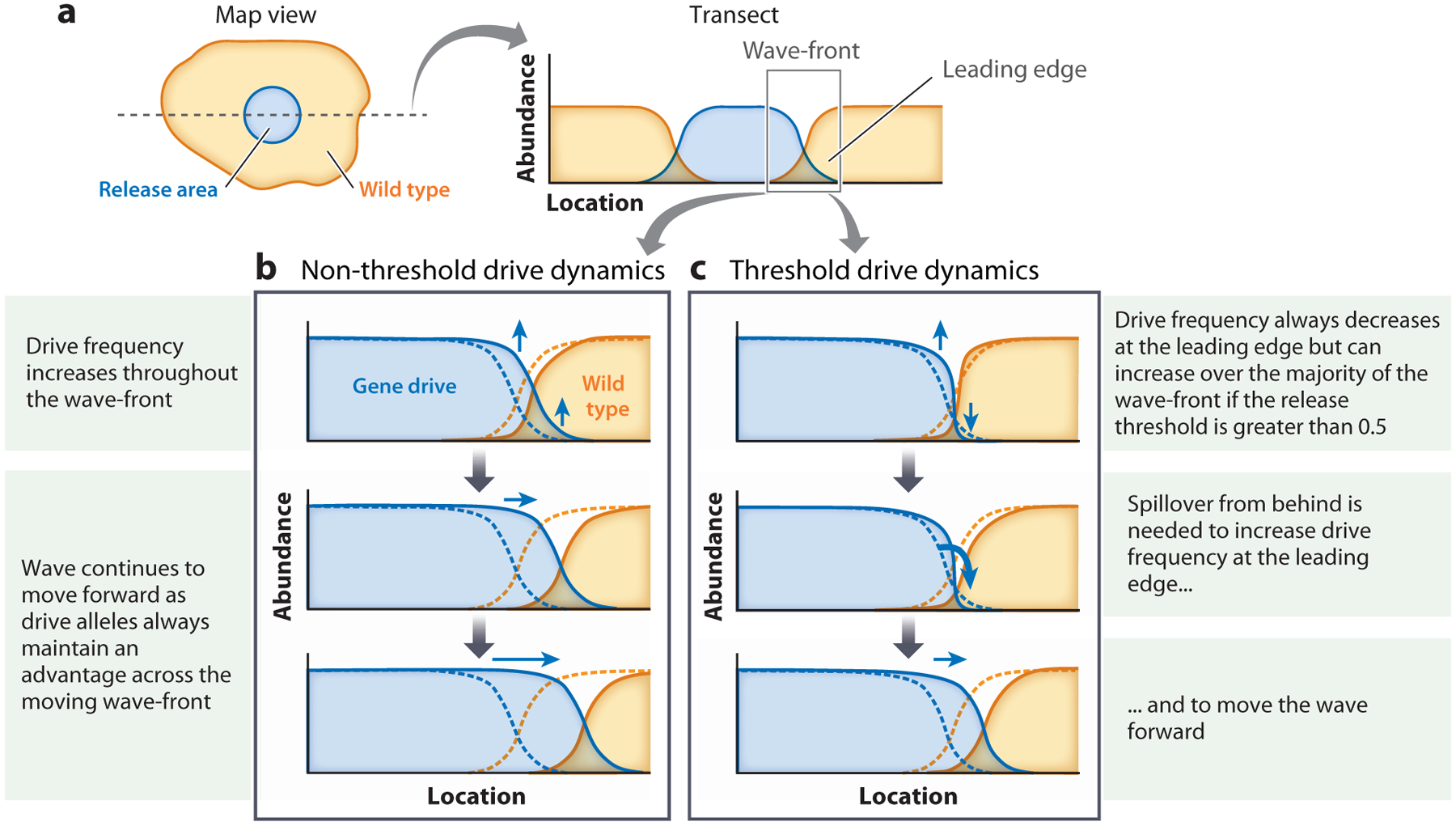
Fisherian versus Bartonian wave spread. Dynamics are shown after a gene drive (blue) is introduced in a central release area within a wild-type (orange) population. (a) Abundance of wild-type and gene-drive alleles across a transect through the release area is shown after local drive establishment within the release area. Dynamics across a wave-front (region shown by gray box) are shown for two types of gene drive. Original wave positions are shown with dashed curves. (b) As a non-threshold gene drive has a fitness advantage over wild-type genes even at very low frequencies, its abundance increases throughout the wave-front, allowing the drive to spread. (c) The fitness disadvantage faced by a threshold drive when at a low frequency causes a decrease in its local abundance at the leading edge; an influx of drive-carrying individuals into the leading edge from behind is needed to overcompensate for this decrease in order for the gene drive to spread.
Figure 4.
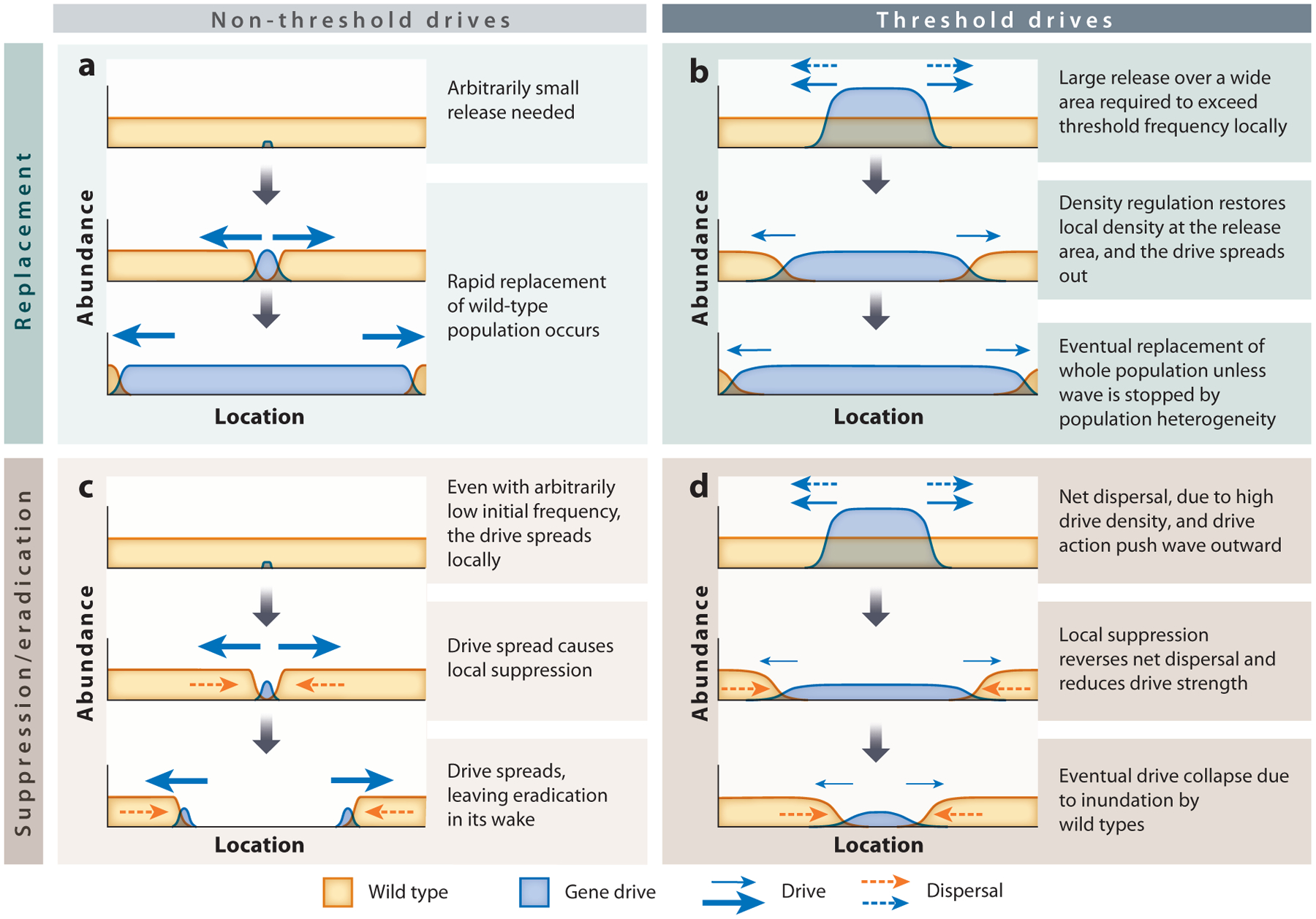
The dynamics of replacement (top row) and suppression or eradication (bottom row) are shown for non-threshold (left column) and threshold (right column) drives. Within each panel, gray downward arrows indicate the progression of time. Horizontal arrows show the direction and relative strengths (indicated by arrow size) of factors that favor, in particular contexts, the spread of the gene drive (blue arrows) or of the wild-type alleles (orange arrows).
While we have focused here on the functional types of density-dependent dynamics and their effect on suppression, other aspects of density dependence can influence gene drive dynamics. A recent review by Godfray et al. (2017) discusses some of these in great detail, allowing us to cover them only briefly here. If density-dependent competition slows down development (e.g., Gimnig et al. 2002, Walsh et al. 2012), it can increase the generation time in a population and slow the spread of population replacement strategies (Hancock et al. 2016a). Strong Allee effects (Stephens et al. 1999), where a population exhibits negative per capita growth rate at very low densities, would also play an important role in population suppression. Strong Allee effects can aid in population eradication with gene drives by contributing to the reduction of replacement rate below 1 (Wilkins et al. 2018). Such Allee effects may also prevent subsequent recolonization of the emptied habitat by wild-type individuals. Another important consideration is the timing of gene drive effects. Suppression drives can potentially be designed to reduce fitness at different time points during an individual’s development. If this fitness reduction occurs at a developmental stage before density-dependent competition occurs, the compensating increase in fitness of the remaining individuals can counter some of the drive’s suppression effect. Timing fitness reduction to occur after the life stage where density-dependent competition is strongest (e.g., after larval stage in many insects) can produce greater suppression because this also allows competition to reduce the population’s reproductive output (Alphey & Bonsall 2014, Edgington & Alphey 2018, Yakob & Bonsall 2009).
Different species can certainly exhibit different forms of density-dependent dynamics (Bellows 1981, May 1973), but the relationship between per capita growth rate and density can vary widely even between populations of a single species or within a population with season, local genetic makeup, presence of other species, etc. (e.g., Rajagopalan et al. 1977, Walsh et al. 2013). Even different localities within a population could exhibit differences in density dependence in heterogeneous habitats.
Unfortunately, density dependence is poorly understood for natural populations of most pest species. Some empirical data are available for certain model organisms, some important disease vectors, and a few species of mammals (Bellows 1981; Chambers et al. 1999; Díaz et al. 2010; Gimnig et al. 2002; Hancock et al. 2016b; Lord 1998; Marlow et al. 2016; Muriu et al. 2013; Twigg & Williams 1999; Walsh et al. 2011, 2012, 2013; Yoshioka et al. 2012). These data have been valuable for designing and parameterizing theoretical studies, but even in these taxa we know little about population- or environment-specific variation in density-dependent dynamics. Theoretical studies of gene drives have made different assumptions about density-dependent dynamics based on simplicity or on the biology of the specific taxon addressed. But different assumptions have been made by different models even when describing the same taxon (e.g., Akbari et al. 2013; Champer et al. 2019b; Eckhoff et al. 2017; Khamis et al. 2018; North et al. 2013, 2019; Sánchez et al. 2019). Given the critical role played by density dependence in determining the effect of gene drives on populations, we reiterate the call made by others before us for more empirical research on this topic (Beaghton et al. 2016, Godfray et al. 2017, Legros et al. 2009, Moro et al. 2018, Walsh et al. 2011).
We recognize the difficulty in gathering data that can allow determination of the functional form of density dependence. While a number examples of each form of dynamics have been suggested (e.g., Bull & Bonsall 2008, Sibly et al. 2005), much debate even exists about what form of density-dependent dynamics should be inferred from the same data (see Doncaster 2006, Getz & Lloyd-Smith 2006, Peacock & Garshelis 2006, Ross 2006). Factors such as stochasticity and seasonality in resource availability can also influence the apparent functional form exhibited by a population (Bull & Bonsall 2008, Lande et al. 1997, Lindström & Kokko 2002, Sæther 1997). In many cases, it may not be possible to determine the exact functional form of such dynamics with confidence. In these scenarios, mathematical models intended for forecasting population dynamics and for risk assessment would need to incorporate such uncertainties in the analyses.
SPATIAL STRUCTURE WITHIN AND BETWEEN POPULATIONS
Few natural populations, if any, exist as a single well-mixed (i.e., panmictic and ecologically interacting) collection of individuals living across a uniform space. Most species occur in multiple, partially isolated populations in a heterogeneous landscape. Even within populations with a generally uniform distribution of individuals over a homogeneous landscape (continuous populations), individuals are more likely to interact with others that are in closer proximity. Therefore, in most populations, individual fitness is determined more by properties such as local density or local genotypic frequencies rather than by the population-level averages for such quantities. Moreover, the fate of a gene drive released into a spatially structured natural population depends disproportionately on the fitness of the individuals near the boundary of the release area (Barton 1979a, Barton & Turelli 2011). Whether the released transgenic individuals can spread the gene drive outward from the release area or are inundated and removed by incoming wild types is determined largely by the interactions in the zone where the cohorts of the wild types and transgenics meet. These details are lost in models that assume a well-mixed population. Many properties of a drive that affect these fine-scale, within-population dynamics are also important in determining how, and if, the drive spreads across a network of populations connected by migration. Understanding these spatial dynamics is crucial not only for optimal drive design and deployment but also for understanding the potential risks of unintended spread.
A great deal of theoretical work has addressed how Mendelian and selfish genetic elements behave in spatially explicit landscapes. These studies range from simple 2-deme simulations through analytical diffusion models to biologically complex and highly detailed simulations of real geographical areas. Some of the theoretical underpinnings of the spatial dynamics of gene drives were revealed by the work of evolutionary biologists studying the spread of naturally occurring genetic elements (e.g., Barton 1979a,b; Barton & Hewitt 1985, 1989; Lande 1985; Piálek & Barton 1997; Rouhani & Barton 1987). Many recent studies have extended this body of work to Wolbachia and synthetic gene drives and have begun to incorporate more biological complexity (Barton & Turelli 2011; Beaghton et al. 2016, 2017a,b; Bull et al. 2019a,b; Champer et al. 2019b, 2020c; Eckhoff et al. 2017; Girardin et al. 2019; Hancock & Godfray 2012; Hancock et al. 2019; Huang et al. 2011; Legros et al. 2013; North et al. 2013, 2019; Tanaka et al. 2017). Below, we first discuss the work describing within-population spatial dynamics and then address the effects of spatial structure on gene drive spread across a network of populations.
Within-Population Dynamics
Consider the release of a large number of gene drive-carrying individuals in a certain area within an otherwise wild-type population. In real populations that are not perfectly mixed, spatial heterogeneity arises in several ways. First, the release of a large number of transgenics causes an increase in the local density (even if temporary), in turn affecting related factors such as density-dependent dispersal or competition within that area (e.g., Backus & Gross 2016). Second, the genetic composition of potential mates available to individuals near the edge of such a release area or of an area where the gene drive has already been established is very different than that available to individuals at the center of or completely outside that area. The gene drive would spread out if the frequency of drive alleles increases at the forefront of the boundary of the release area or of an area where the drive is already established. If instead the wild-type frequency keeps increasing at the boundary, the boundary would be pushed inward, causing drive failure. The change in allelic frequencies at the boundary is determined by two factors: (a) the frequency-dependent relative fitness of the alleles (transgenic and wild-type) at the boundary, and (b) the addition of alleles through dispersal from either side of the boundary. Fisher (1937) used a reaction-diffusion framework to describe these dynamics at the boundary for beneficial mutations as a wave of genotypic frequencies. Barton (1979a) extended this work to describe the spread of natural genetic elements that exhibit a threshold behavior similar to threshold gene drives. While the reaction-diffusion framework requires many simplifying assumptions, it has been very useful for understanding the spatial behavior of genetic elements without a frequency threshold for their fitness relative to wild types (Fisherian waves) and those with a frequency threshold (Bartonian waves). The spatial dynamics of these two types of genetic elements are significantly and qualitatively different. Below we first highlight how the Fisherian and Bartonian dynamics differ and then separately consider the dynamics of threshold and non-threshold drives under the influence of different ecological factors.
Fisherian versus Bartonian waves.
At the forefront of the boundary of a release area, the leading edge of the wave (Figure 3a), drive alleles are by definition going to be rare initially. Non-threshold drive alleles have an evolutionary fitness advantage over wild-type alleles irrespective of their frequency and can increase in frequency even at the leading edge of the wave. Thus, regardless of the fate of the drive-carrying individuals behind the leading edge of the wave, just the evolutionary fitness advantage at the forefront of the boundary can pull the wave of a non-threshold drive forward (Figure 3b). In contrast to these Fisherian wave or pulled wave (Stokes 1976) dynamics of non-threshold drives, threshold drives cannot increase in frequency through an evolutionary fitness advantage at the leading edge where they are rare. Threshold drives therefore critically rely on dispersal (spillover) of drive-carrying individuals from areas where they are at high frequency into the leading edge for the wave to move forward (Figure 3c). Movement of this Bartonian or pushed wave-front of threshold drives is intimately tied to the fate and dispersal of drive-carrying individuals behind the leading edge (Barton 1979a, Beaghton et al. 2016, Godfray et al. 2017, Stokes 1976).
Non-threshold drives.
Gene drives that exhibit Fisherian dynamics can establish a spreading wave even in the absence of any additional transgenics dispersing in from behind the wave (e.g., Beaghton et al. 2016, Tanaka et al. 2017) (Figure 4a). While any spreading wave requires forward dispersal from the leading edge, the lack of need of dispersal into the leading edge from behind has important implications for population suppression or eradication with non-threshold drives. A non-threshold eradication drive can spread across a continuously distributed but non-panmictic population even if it leaves empty space in the wake of the wave, similar to a spreading ring of forest fire (Beaghton et al. 2016, Tanaka et al. 2017) (Figure 4c). This could result in eradication of an entire uniform, continuous population, assuming there is no long-range dispersal that could allow wild types to recolonize the empty space. While reaction-diffusion models assume only very short-range dispersal, stochastic, individual-based, spatially explicit models of eradication drives show that if wild types can disperse to occupy the empty space created by an eradication drive, perpetual local cycles of drive invasion, eradication, and wild-type recolonization can occur in continuous but non-panmictic populations (Champer et al. 2019b) or in structured metapopulations (Bull et al. 2019b, Eckhoff et al. 2017, North et al. 2019). Similar cyclical spatiotemporal dynamics are also predicted for certain reversal drives released to eliminate a previously established homing gene drive, between the alternative states of local occupation by wild types, homing drive, and reversal drive in spatial and nonspatial contexts (Girardin et al. 2019, Vella et al. 2017). Such dynamics can interfere with complete eradication of the population. However, if strong Allee effects prevent or slow down the establishment of a viable population in the eradicated area by a small number of dispersing wild types, complete eradication may still be feasible. Repeated introductions of the gene drive may also help to reduce such perpetual dynamics (Eckhoff et al. 2017, North et al. 2019). Gene drives that suppress a population, but do not cause eradication, may avoid such cycles because low but persistent presence of individuals carrying such a drive in the suppressed areas could resist recolonization by a small number of incoming wild types. If the epidemiological, conservation, or agricultural purpose behind population alteration does not require eradication, a less powerful suppression drive could offer an efficient solution.
In contrast to suppression drives, non-threshold, replacement drives are expected to be pulled forward with no impact of wild types dispersing into areas that have already been occupied by individuals with the gene drive and altered phenotype. Of course, in more realistic scenarios with spatial heterogeneity, seasonality, evolution of resistance alleles, molecular breakdown of gene drive components, and other complexities, even non-threshold replacement drives can fail to spread or achieve the desired outcome (e.g., Beaghton et al. 2017b; Bull 2017; Bull et al. 2019a,b; Girardin et al. 2019).
Threshold drives.
As mentioned in the section titled Fisherian Versus Bartonian Waves, an increase in the frequency of a threshold drive at the leading edge of the boundary of the release area depends critically on addition of transgenes from behind the leading edge through dispersal (Barton 1979a) (Figure 3c). The movement of such pushed waves over continuous space is therefore affected by all the factors that influence net dispersal across the wave-front, in addition to the relative evolutionary fitness of the drive and wild-type alleles (Barton 1979a; Barton & Hewitt 1985, 1989; Barton & Rouhani 1991; Barton & Turelli 2011; Michalakis & Olivieri 1993; Piálek & Barton 1997; Rouhani & Barton 1987).
The threshold release frequency of a gene drive determines the net selective advantage (or disadvantage) experienced by drive alleles over the width of the wave-front in continuous populations. Assuming equal local densities in the area behind a wave (where the drive has established) and the area occupied by wild types in front of the wave, and no differences in dispersal between transgenics and wild types, a gene drive with a release threshold much greater than 0.5 will always face a selective disadvantage across a wave-front. Reaction-diffusion models therefore lead to the conclusion that, all else being equal, gene drives with a release threshold much greater than 0.5 cannot spread out, even when the initial drive frequency in the release area is far greater than the release threshold (Barton & Turelli 2011). However, a wave of a gene drive with a release threshold lower than 0.5 (e.g., Champer et al. 2020b, Dhole et al. 2019, Oberhofer et al. 2019, Ward et al. 2011) may be able to move forward (outward from the release area) across a uniform landscape (Barton & Turelli 2011) (Figure 4b). In the section titled Critical Bubbles and Release Strategies for Threshold Drives we discuss possible deviations from and caveats for this conclusion. This conclusion is also our motivation for using a release threshold of 0.5 to distinguish between high-threshold and low-threshold (see below) drives in this review. This terminology is used solely for the purpose of the discussion in this review and is not intended to contend with other definitions of these terms (James et al. 2018, Natl. Acad. Sci. Eng. Med. 2016).
Even for gene drives with a release threshold less than 0.5 (here referred to as low-threshold drives), their frequency at the leading edge can increase only through dispersal of drive alleles from behind. Therefore, a reduction in the net dispersal of drive alleles into the leading edge can halt the spread of threshold drives (Barton 1979a, Barton & Turelli 2011, Piálek & Barton 1997). A difference in density of individuals between two regions can give rise to a bias in net dispersal of alleles from high- to low-density areas simply because there are more individuals that can disperse from the high-density region. Such density gradients can occur due to heterogeneity in the distribution of resources that determine local carrying capacity or due to the fitness effects of the gene drive itself. If a traveling Bartonian wave of a threshold drive encounters a density gradient, with density increasing in the direction of wave spread, the spillover of drive alleles into the leading edge of the wave from behind may get outnumbered by inundation of wild types from the more dense area in front of the wave (Barton & Turelli 2011). Conversely, a density gradient with density decreasing in the direction of gene drive spread can favor the spread of the drive, for example, in a scenario where the drive release occurs in an area of high carrying capacity (e.g., Champer et al. 2018). A wave of a spreading threshold drive could also be halted by a zone of low density because of the unfavorable density gradient on the opposite side of the zone (Barton 1979a, Piálek & Barton 1997). Such effects of density gradients can have an especially strong influence on threshold drives that cause population suppression. Consider a threshold drive such that transgenic homozygotes have lower viability (or reproductive ability) relative to wild-type homozygotes. If this disadvantage, which is frequency-independent, results in a lower local density in the area where the drive is present (i.e., if the drive causes local population suppression), net dispersal would act against drive spread (Barton 1979a, Barton & Turelli 2011) (Figure 4d). Such a dispersal disadvantage would certainly further reduce the ability of spatial spread for high-threshold drives, but if the density reduction due to the gene drive is strong enough, even low-threshold drives could fail to spread.
Theoretical studies that have examined the effects of density gradients on the spatial dynamics of threshold drives (or natural elements that exhibit bistability) have largely focused on density gradients as a property of the environment, rather than as resulting from fitness effects of different genotypes (e.g., Barton 1979a, Barton & Hewitt 1985, Barton & Rouhani 1991, Barton & Turelli 2011, Champer et al. 2020c). If genotypic fitness can affect local density, the form of density-dependent dynamics could have a strong influence on the formation of density gradients and, in turn, on the spatial dynamics of gene drives. For instance, in a population with density dependence described by curve iii in Figure 2 (e.g., Sibly et al. 2005), even small fitness costs of the drive constructs can cause a large difference in the densities on either side of the wave, limiting the spread of the drive even if the threshold release frequency of the drive remains the same. We are not aware of any analytical diffusion model of Bartonian wave dynamics that allows a strong effect on local density of a spreading genetic element of interest (see results for a Fisherian wave in Beaghton et al. 2016), but some recent individual-based simulation studies suggest that genotype-mediated changes in local density play an important role in the spread of threshold drives (Champer et al. 2019b, 2020c; Huang et al. 2011). Theoretical studies that can incorporate different forms of density-dependent dynamics into models of spatial spread of gene drives would be valuable for understanding how threshold (and non-threshold) gene drives would behave in natural populations of different species or in different environmental conditions. Such models will also be useful for shedding light on the potential applicability of threshold drives for suppression of spatially structured populations, which thus far has been addressed only in panmictic scenarios (e.g., Akbari et al. 2013; Dhole et al. 2018, 2019; Khamis et al. 2018).
Much like density gradients, an unfavorable gradient in dispersal or a barrier to dispersal can similarly halt the spread of a threshold drive (Barton 1979a). In the case of urban dwelling mosquitoes such as Aedes aegypti, the landscape between cities or towns is expected to be such a barrier. Even features such as roads and rivers may provide strong enough dispersal barriers to halt the spread of certain threshold drives. In general, environmental heterogeneity is likely to limit the spread of threshold drives across a landscape.
Another factor in many pest species that adds complexity to spatial dynamics of gene drives is a separation between the life stages that face density-dependent regulation and the life stages that disperse and reproduce. In a model of an eradication driving-Y gene drive, which exhibits Fisherian wave dynamics, Beaghton et al. (2016) showed that the speed of wave spread for the drive decreases in proportion with increasing fraction of total lifetime spent in immobile juvenile stages. The spread of threshold drives, which relies critically on dispersal of transgenics from behind the leading edge of the wave, may be especially sensitive to a lag between density regulation and dispersal. Spatially explicit models that can incorporate such developmental delays will be useful for understanding how rapidly threshold drives can spread through such stage-structured populations, such as those of disease-vectoring mosquitoes.
Critical bubbles and release strategies for threshold drives.
The release of a threshold drive certainly needs to be large enough to exceed the release threshold within the release area, but how much should the threshold be exceeded by, and how large should the release area be? The answer, of course, is not straightforward. Barton and colleagues (Barton 1979a, Barton & Turelli 2011, Rouhani & Barton 1987) have derived expressions that describe the critical frequency distribution over space, termed the critical bubble, that needs to be exceeded to establish a spreading wave for simple underdominance elements and for Wolbachia that can induce CI. While these expressions rely on some simplifying assumptions that may not be realistic for many scenarios of gene drive releases, they have high heuristic value. They show that for threshold drives, the threshold release frequency needs to be exceeded over a release area with a minimum critical radius. Initial establishment of the drive over a large area helps the wave spread in two ways. First, release over a larger area reduces the dilution effect of dispersal; a large area is less likely to be inundated by wild types from the surroundings than a small one. Second, the geography of the boundary of the release area matters. A strong curvature of the boundary results in a bias in the net dispersal from outside the curve to inside (Barton 1979a, Champer et al. 2020c). Increasing the release area can reduce the curvature of the boundary, aiding the spread of the drive to some extent.
The analysis of a reaction-diffusion model by Barton & Turelli (2011) suggests three interesting things about the critical radius. First, the critical radius increases very slowly with increasing release threshold of a gene drive until the threshold begins to exceed ~0.3, after which the critical radius increases very rapidly. Second, while the initial release frequency needs to exceed the threshold release frequency, the magnitude of this excess has only a small effect on the critical radius. Third, the absolute critical radius certainly increases with the dispersal rate of individuals. Similar expressions are not yet available for other types of gene drives that exert strong selection (e.g., suppression drives or drives that cause strong reduction in heterozygote fitness) or have a more complex genetic structure than simple 1-locus underdominance. One caveat to these results regarding the critical radius is that this analysis does not account for the increase in local density, even if temporary, that must come with a gene drive release. Increasing the initial release frequency of a gene drive, for instance from 0.5 (1:1 drive:wild type) to 0.8 (4:1 drive:wild type), can correspond to a significant increase in the density gradient at the boundary of the release area. Even if the gradient remains short-lived due to local density regulation, it may have a significant impact on the critical radius required to get a spreading wave established. It remains to be seen how large an effect such density changes can have on the critical radius. For application to gene drive release in natural populations, estimating the critical bubble size would need much extended and more case-specific analyses that can account for factors such as sustained differences in density due to repeated releases of the drive, the density-dependent dynamics that can remove density gradients, heterogeneity in the carrying capacity across space and time, etc. (e.g., see Champer et al. 2018).
The discussion above is restricted to the analysis of a single release in one area. However, the spatial patterns of release can play a large role in establishment and spread of threshold drives. Huang et al. (2011) explored a model of an age-structured population in a 32×32 grid of discrete but highly connected patches. Individuals with a 2-locus engineered underdominance drive (intrinsic minimum release threshold ~0.27) were released either in a central core of patches (central release) or into an equivalent area of patches that were uniformly distributed over the entire grid (distributed patches). They found that total number of released transgenic individuals needed to establish the drive depended on many factors including age of released individuals, fitness costs, and migration rates. An interaction between dispersal rate and fitness differences between drive and wild-type individuals determined whether a smaller number of total released individuals was needed in the central releases or the distributed patch releases. Detailed simulation models of mosquito populations also suggest an important role of multiple releases distributed across the landscape (Eckhoff et al. 2017, Legros et al. 2013, North et al. 2019). These models demonstrate that while the general principles from simple reaction-diffusion models are upheld, biological properties of the target species will be critical in determining optimal gene drive design and release strategies.
Empirical data on these biological properties are not available for most of the species proposed as targets for gene drives. However, several recent empirical studies have focused on elucidating the spatial ecology of important disease vector species (e.g., Dao et al. 2014; Epopa et al. 2017; Goubert et al. 2016; Guagliardo et al. 2019; Huestis et al. 2019; Kotsakiozi et al. 2018; Lehmann et al. 2017; Rašić et al. 2015, 2014). More studies on the ecology of pests are critically needed.
Between-population spread.
Migration rates are expected to impact the spread of both threshold and non-threshold gene drives between populations with finite size. However, the migration rates needed for any establishment of threshold drives in a new population are much more substantial (Dhole et al. 2018, Láruson & Reed 2016, Marshall & Hay 2012a, Noble et al. 2018). Modeling efforts on between-population spread have been mostly focused on threshold drives because the goal of threshold drive developers is often prevention of cross-population spread, and models that can provide insights into the risk of a threshold drive spreading will be of special interest to regulatory authorities (Backus & Delborne 2019). There are similarities between factors that impact the spread of gene drives in single non-panmictic populations and those that impact spread between discrete populations. But there are also qualitative differences in the dynamics because the speed of local population dynamics can be much faster than the speed of dispersal dynamics between discrete populations.
As a first approximation of interpopulation spatial structure, many theoretical studies have used a simple scenario where two (or very few) large, well-mixed populations exchange migrants at some specified rate every generation (e.g., Akbari et al. 2013; Altrock et al. 2010; Dhole et al. 2018, 2019; Láruson & Reed 2016; Marshall & Hay 2012a; Noble et al. 2019; Sudweeks et al. 2019). While highly simplified, this scenario provides a very convenient method for studying basic properties of gene drive spread between populations. Such models are not intended to forecast the exact dynamics of a gene drive in natural conditions but rather are used as tools to describe basic gene drive properties and, importantly, give comparative measures of the tendencies of different gene drives to spread to new populations through migration (e.g., Dhole et al. 2018, Marshall & Hay 2012a). They can also help in determining how different properties of the gene drive or of the two populations can affect the drive’s ability to spread across population boundaries (e.g., relative fitness of transgenics, the level of haploinsufficiency of target genes, level of resistance in neighboring populations, etc.) (Dhole et al. 2019, Noble et al. 2019, Oberhofer et al. 2019, Sudweeks et al. 2019).
As mentioned above, the within-population spread of non-threshold gene drives is largely independent of the dynamics behind the wave-front, making even complete eradication feasible (with caveats regarding recolonization by wild types) (Beaghton et al. 2016, Champer et al. 2019b, Okamoto et al. 2013). Spread across different populations, however, can be difficult for even non-threshold eradication drives because if the source population is eradicated very rapidly, the drive may not have a chance to spread to a new population (Eckhoff et al. 2017, North et al. 2013). The fate of an eradication drive, and also of strong suppression drives, is highly dependent on the relative rates of local eradication, dispersal across populations, and recolonization by wild types (Eckhoff et al. 2017; Lande 1985; North et al. 2013, 2019). As in the case of a population uniformly distributed over a continuous landscape (Champer et al. 2019b, Girardin et al. 2019), for a range of realistic dispersal rates, perpetual cycles of drive invasion, local extinction, and wild-type recolonization can occur across a large network of connected populations (Eckhoff et al. 2017; North et al. 2013, 2019). If individual populations are sufficiently small to make stochastic local extinctions common, the spread of replacement drives across multiple populations also becomes less certain. Moreover, if populations experience extreme seasonal fluctuations, stochastic local extinctions may become more likely during seasonal lows (Eckhoff et al. 2017). Seasonal changes in dispersal may also influence how gene drives spread between populations (North et al. 2013, 2019). Temporal variation in dispersal and fitness has not yet received substantial formal investigation with regard to gene drives, but it will certainly play an important role in spread between populations.
Threshold drives need a much larger cohort of transgenics to establish a drive in a new population, so much higher dispersal rates would be required for them to invade new populations than are needed for non-threshold drives. Very high dispersal rates can also cause the source population to be inundated by incoming wild types and cause the drive to be lost from all populations (e.g., Dhole et al. 2018, Láruson & Reed 2016). Threshold drives are therefore unlikely to be suitable for alteration (even replacement) of multiple populations, unless large releases are carried out in each targeted population (but see Dhole et al. 2018, Láruson & Reed 2016, Marshall & Hay 2012a). If threshold drives were to be used for eradication or suppression, they would face an even stronger challenge when spreading across multiple populations, assuming they can spread within a population in the first place. This theoretical inability of threshold drives to spread across populations, or in other words, the ability to remain localized to a population, is attractive when spread across multiple populations is undesirable (Buchman et al. 2018a, Curtis & Robinson 1971, Davis et al. 2001, Dhole et al. 2018, Magori & Gould 2006, Marshall 2010, Marshall & Hay 2012a, Natl. Acad. Sci. Eng. Med. 2016).
The theoretical studies discussed above that analyze the likelihood of gene drive spread across multiple populations assume panmictic populations or patches that exchange migrants (e.g., Dhole et al. 2018, 2019; Huang et al. 2011; Láruson & Reed 2016; Marshall & Hay 2012a; Noble et al. 2018, 2019). In such simple models, drive-carrying individuals migrating into a new population are expected to interact largely with wild types that have a much higher overall frequency. In real scenarios, however, migrating individuals are more likely to arrive in a certain area of the new population. For example, transgenics migrating via ships would arrive at a port, or populations connected by narrow corridors (e.g., highways between towns) would arrive at the end of the corridor. A recent study by Champer et al. (2020c) modeled a 2-deme scenario with the populations within each deme spread over a continuous space. The study shows that when migrants are more likely to arrive in close proximity, the likelihood of a threshold drive invading the new population is much higher than expected from panmictic deme models. This is an important result and clearly calls for empirical research to assess spatial patterns of migration into new populations. Of course, real populations are not continuously distributed. Discrete barriers such as rivers, roads, human habitations, or even the tendency of individuals to be located near discretely dispersed resources such as ponds can result in small patches of individuals that are panmictic within the patch and somewhat isolated from others. Gene drive spread between such heterogeneous populations might be more realistically simulated by population network models where each population is modeled as a closely connected group of panmictic demes (e.g., Huang et al. 2011, North & Godfray 2018, North et al. 2019, Sánchez et al. 2019).
As discussed above, strong population structure is likely to make gene drive spread more difficult, especially for eradication drives, which may not be able to spread to all populations before going extinct (Eckhoff et al. 2017, North et al. 2019). One possible solution to reduce the likelihood of local extinction may be to repeatedly introduce the drive in multiple changing locations (e.g., North et al. 2019). Long-term and large-scale eradication of pest species will require more effort in spatially structured populations than estimated by panmictic models. However, spatial structure could make it easier to effect changes in local populations, when global spread is undesirable. Any genetic differentiation between highly spatially structured populations may also allow the use of population-specific molecular confinement mechanisms (see below).
Localized population alteration.
Non-threshold drives may provide a mechanism for altering natural populations on a large scale, but they pose a challenge for spatial containment when only local population alteration is desired (Natl. Acad. Sci. Eng. Med. 2016). As we have mentioned before, threshold drives could provide a solution in such cases, especially low-threshold drives. Several threshold drive designs fit into the category of low-threshold drives, especially if the drive constructs themselves impose only a small cost on the carriers (e.g., Akbari et al. 2013, 2014; Champer et al. 2020a,b; Davis et al. 2001; Marshall & Hay 2011; Marshall et al. 2011; Oberhofer et al. 2019; Ward et al. 2011). Such low-threshold drives may be able to achieve local population alteration without requiring very large releases in the target populations. However, if a gene drive construct imposes a significant cost on the carrier, this cost is likely to shift the release threshold to a high value (e.g., Dhole et al. 2018, 2019; Leftwich et al. 2018; Ward et al. 2011), which would raise the amount of release effort required and also increase the likelihood of inundation by incoming wild-type migrants. Therefore, while a number of mechanisms may be available for localized population replacement, localized population suppression remains a challenge.
A few gene drives that exhibit a release threshold solely due to fitness costs incurred by individuals carrying the gene drive have been proposed for localized population suppression (Champer et al. 2020a, James et al. 2018). If the fitness costs to the carriers remain stable across time and environment, such gene drives would remain spatially restricted. However, environment-dependent or evolved changes in fitness of gene drive carriers after the release may unpredictably change or even remove the release threshold and thus the localization ability of such gene drives. As we stated before, special care would need to be taken when using such gene drives for spatially restricted use, for instance, to ensure that the fitness costs incurred by different genotypes remain within bounds that allow maintenance of desired release thresholds for the entire period that the gene drive remains active in the population. Gene drives that exhibit intrinsic minimum release thresholds may provide a form of added insurance against such changes in fitness. We recently proposed two designs for new gene drives that may provide a solution: the Tethered Homing (TH) drive and a homing drive targeting locally fixed alleles (Dhole et al. 2019, Sudweeks et al. 2019).
The TH design is based on driving a fitness-reducing homing construct that is anchored to a threshold component (Dhole et al. 2019). The homing construct is essentially a fitness-reducing suppression HEG drive with its Cas endonuclease removed, disabling its ability to spread rapidly on its own. The threshold component is any threshold drive that can be linked with genes for germline-specific expression of a Cas endonuclease. Because of its release threshold, the threshold component can spread only in the target population where it is released in sufficient numbers, thus establishing a localized population of individuals that express the Cas endonuclease in their germline. In absence of the homing construct, the Cas endonuclease has no activity because it lacks any guide RNAs. The homing construct, with its Cas endonuclease removed, can spread through gene conversion only when it co-occurs with the threshold component. Any homing constructs that reach a nontarget population cannot spread there in absence of the Cas-bearing threshold component and will be removed by natural selection due to their fitness costs. This dependency of the homing construct on a component that is able to remain localized can allow it to spread rapidly in the target pest population but can prevent it from invading nontarget populations. A major benefit of the TH design over other threshold drives is the separation of the threshold component from the high fitness costs required for population suppression, which come from the homing construct in a TH drive system. Thus, unlike other threshold drives, the release threshold of the TH drive remains largely unaffected by the fitness costs imposed by the TH drive, allowing the possibility of strong localized suppression without the need for a very large release effort. The intrinsic minimum release threshold of the threshold component can provide the insurance against environment-dependent changes in fitness mentioned above. Another benefit of this system is that the threshold component provides a persistent, spatially restricted source of Cas endonuclease that can be used to spread additional homing components targeting other genes for stronger population suppression or population alteration. Because the threshold component could be any threshold drive that can be linked with Cas endonuclease genes, this system may provide additional flexibility for creating a drive in different species.
Taking a very different approach to local containment, one that is not based on a release threshold, Sudweeks et al. (2019) have proposed a method for localizing non-threshold CRISPR-based homing drives to islands where invasive pests are targeted for eradication. The method relies on the fact that on isolated islands, drift and localized selection are likely to cause fixation of alleles that are polymorphic on the mainland. If these locally fixed alleles are used as targets for a suppression homing drive, the drive is expected to rapidly spread on the island by targeting the locally fixed allele, but it will not permanently establish in mainland populations where homologs of the fixed island allele are present and cannot be targeted by the homing drive’s guide RNAs. While this approach may work for islands, it is much less likely that two large mainland populations will have differential fixation of many targetable alleles, making the method less suitable for locally altering mainland populations.
Both these strategies have very specific requirements for achieving successful localized spread (Dhole et al. 2019, Sudweeks et al. 2019), but they provide a rare set of solutions for safer locally restricted, yet efficient population suppression (Hay et al. 2021). It is our hope that ongoing progress in genetic engineering techniques will facilitate creation of these systems in pest species important in epidemiological and conservation applications.
SEX AND MATING
All gene drives depend on mating and fertilization between transgenic and wild-type individuals. Mate choice by wild-type females against drive-carrying males can strongly hamper the spread of gene drives (Huang et al. 2009, 2011; Khamis et al. 2018; Leftwich et al. 2016; Manser et al. 2015, 2017; Wedell 2013). While females of some species can discriminate against males that carry certain natural meiotic drivers (Wedell & Price 2015), it is not clear if females could quickly evolve to directly discriminate against synthetic gene drives that can spread rapidly through a population. However, many existing behaviors can create a mating bias. For instance, if adult females mate only during limited periods in their lifetimes, the number of wild-type females that are available to mate at a given time could be low. For example, Aedes mosquitoes mate soon after eclosing and usually mate only once during their lifetimes in the field (Foster & Lea 1975, Williams & Berger 1980, Young & Downe 1982). Therefore, the only wild-type females available for mating with transgenic males would be females that eclose after the release. Even if adult females mate many times and do not have second-male sperm precedence, it must be remembered that the majority of individuals in an insect population are typically found in the immature stages and are therefore unavailable for mating. When calculating an appropriate release threshold, it is important to consider not just the number of individuals released but also their reproductive value (Fisher 1930) and the reproductive value of the individuals with whom they mate (Huang et al. 2009, Legros et al. 2013). In a model of gene drive spread in an age-structured population, Huang et al. (2009) showed that low levels of polyandry can significantly hamper the spread of a gene drive, especially in cases of male-only releases. Even if only females instead of males are released, there can be limitations to mate availability. This limitation can be partially mitigated by bi-sex releases (Huang et al. 2009, Khamis et al. 2018, Legros et al. 2013, Robert et al. 2013). In many species, however, the release of females may not be feasible due to the disease-transmitting capabilities of females (e.g., in mosquitoes) or due to the potential increase in population growth (even temporary) as a result of the addition of females. Another example of factors that can reduce matings between drive and wild-type individuals is inbreeding, caused either by strong population structure or by active mate choice. Two models by Bull (Bull 2017, Bull et al. 2019b) show that even low levels of inbreeding (or especially selfing in plants) can strongly hamper the spread of gene drives.
In polyandrous species, where females mate with multiple males within a breeding cycle, sperm competition can hamper the reproductive ability of drive-carrying males even in the absence of mate choice (Lindholm et al. 2016, Wedell 2013). There is considerable evidence that certain sex-ratio distorting drives can reduce males’ sperm-competitive ability in a number of taxa (Price & Wedell 2008). The t-haplotype in mice is a large cluster of genes that biases its transmission through heterozygous males by rendering sperm without the t-haplotype dysfunctional (Herrmann & Bauer 2012). In an experimentally evolving population of mice, Manser et al. (2017) showed that the spread of a t-haplotype is significantly hindered in populations with strong sperm competition. A gene drive that links a sex-ratio distorter to the t-haplotype is being considered for eradication of invasive mice on islands (Godwin et al. 2019, Leitschuh et al. 2018). A sperm-competitive disadvantage could enforce a much larger release effort for successful alteration with such a gene drive.
Sex-ratio biasing drives that cause a scarcity of males can suppress a population if a large number of females are unable to find a mate. The level of polygyny, i.e., the mean number of mates per male, can severely affect the efficacy of such gene drives. For example, a Y-shredder drive that reduces the number of males would require a much stronger bias in the sex ratio in a polygynous population than in a monogamous population to achieve the same level of suppression (Prowse et al. 2019). If the female-biased sex ratio is caused through reduction in male-forming sperm, most of the offspring are born as females, resulting in an increase in the absolute number of females. In populations where a small number of males can fertilize a large number of females, such sex-ratio bias could actually increase population size because it increases the number of fertilized females (Prowse et al. 2019). Male-reducing gene drives that do not increase the absolute number of females (e.g., by high mortality of male zygotes or offspring) are much more likely to produce the desired population suppression. As described previously, late-stage juvenile mortality can also aid suppression by maintaining strong density-dependent competition at juvenile stages.
The effect of gene drives on the reproductive interactions between individuals and its implications for gene drive spread are generally poorly understood. While viability of transgenic individuals under natural conditions has begun to receive some much-needed attention, more research will be required on the reproductive abilities of transgenics in natural populations.
CONCLUSIONS
A large number of ecological factors will affect the fate of gene drives in natural populations. Our ability to accurately forecast the impact of gene drives on populations will depend upon how well we understand these ecological effects. The modeling work reviewed here shows that variation in ecological factors is likely to have an especially strong impact on the success of suppression drives compared to that of replacement drives.
As has been highlighted by other authors, self-sustaining, non-threshold gene drives should be developed only for applications where any consequences of unrestricted spread are viewed by society as acceptable because of the positive gains expected. In these cases, a better understanding of the pest population’s ecology could assist in the development of gene drives most suited to a project’s goals and could guide efficient strategies for spatial and temporal field releases of the drives.
In cases where unrestricted spread of a gene drive is deemed societally unacceptable, gene drives with substantial release thresholds or other strong localizing mechanisms may provide a solution for a contained management of pest populations. However, the modeling work done to date indicates that it can sometimes be difficult to achieve any spatial spread for threshold drives, especially those with high release thresholds. Creating gene drives that can achieve the balance between ability to spread and ability to remain locally confined is a challenge that will require a better understanding of the match between the properties of the specific gene drives and the pest population’s ecology in the targeted environment. Sensitivity analyses based on detailed and locally relevant models could be useful in guiding the direction and intensity of empirical research for measuring the relevant ecological parameters. This research could, in turn, help to build more robust models that are useful for regulatory decision-making.
Breakthroughs in molecular biology are resulting in rapid progress toward genetically stable gene drives that could be released into natural populations. It is imperative that the ecological and population genetic research keep pace with the molecular work if we are to make informed decisions about release of gene drives to address the myriad conservation, agricultural, and human health issues for which their use has been proposed.
ACKNOWLEDGMENTS
We are grateful to Austin Burt, Brandon Hollingsworth, Bruce Hay, Jackson Champer, Jennifer Baltzegar, Michael Vella, and Thomas Prowse for providing valuable feedback on a previous version of this manuscript. All authors were funded by NIH-NIAID 1R01AI139085-01.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings that might be perceived as affecting the objectivity of this review.
LITERATURE CITED
- Akbari OS, Chen C-H, Marshall JM, Huang H, Antoshechkin I, Hay BA. 2014. Novel synthetic Medea selfish genetic elements drive population replacement in Drosophila; a theoretical exploration of Medea-dependent population suppression. ACS Synth. Biol 3(12):915–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akbari OS, Marshall JM, Raban R, Kandul NP, Edula JR, Leon T. 2019. Winning the tug-of-war between effector gene design and pathogen evolution in vector population replacement strategies. Front. Genet 10:1072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akbari OS, Matzen KD, Marshall JM, Huang H, Ward CM, Hay BA. 2013. A synthetic gene drive system for local, reversible modification and suppression of insect populations. Curr. Biol 23(8):671–77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alphey L. 2014. Genetic control of mosquitoes. Annu. Rev. Entomol 59:205–24 [DOI] [PubMed] [Google Scholar]
- Alphey N, Bonsall MB. 2014. Interplay of population genetics and dynamics in the genetic control of mosquitoes. J. R. Soc. Interface 11(93):20131071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altrock PM, Traulsen A, Reeves RG, Reed FA. 2010. Using underdominance to bi-stably transform local populations. J. Theor. Biol 267(1):62–75 [DOI] [PubMed] [Google Scholar]
- Backus GA, Delborne JA. 2019. Threshold-dependent gene drives in the wild: spread, controllability, and ecological uncertainty. Bioscience 69(11):900–7 [Google Scholar]
- Backus GA, Gross K. 2016. Genetic engineering to eradicate invasive mice on islands: modeling the efficiency and ecological impacts. Ecosphere 7(12):e01589 [Google Scholar]
- Barton NH. 1979a. The dynamics of hybrid zones. Heredity (Edinb.) 43(3):341–59 [Google Scholar]
- Barton NH. 1979b. Gene flow past a cline. Heredity (Edinb.) 43(3):333–39 [Google Scholar]
- Barton NH, Hewitt GM. 1985. Analysis of hybrid zones. Annu. Rev. Ecol. Syst 16:113–48 [Google Scholar]
- Barton NH, Hewitt GM. 1989. Adaptation, speciation and hybrid zones. Nature 341(6242):497–503 [DOI] [PubMed] [Google Scholar]
- Barton NH, Rouhani S. 1991. The probability of fixation of a new karyotype in a continuous population. Evolution 45(3):499–517 [DOI] [PubMed] [Google Scholar]
- Barton NH, Turelli M. 2011. Spatial waves of advance with bistable dynamics: cytoplasmic and genetic analogues of Allee effects. Am. Nat 178(3):E48–75 [DOI] [PubMed] [Google Scholar]
- Beaghton A, Beaghton PJ, Burt A. 2016. Gene drive through a landscape: reaction-diffusion models of population suppression and elimination by a sex ratio distorter. Theor. Popul. Biol 108:51–69 [DOI] [PubMed] [Google Scholar]
- Beaghton A, Beaghton PJ, Burt A. 2017a. Vector control with driving Y chromosomes: modelling the evolution of resistance. Malar. J 16(1):286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beaghton A, Hammond A, Nolan T, Crisanti A, Godfray HC, Burt A. 2017b. Requirements for driving antipathogen effector genes into populations of disease vectors by homing. Genetics 205(4):1587–96 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beeman RW, Friesen KS, Denell RE. 1992. Maternal-effect selfish genes in flour beetles. Science 256(5053):89–92 [DOI] [PubMed] [Google Scholar]
- Bellows TS. 1981. The descriptive properties of some models for density dependence. J. Anim. Ecol 50:139–56 [Google Scholar]
- Brownstein JS, Hett E, O’Neill SL. 2003. The potential of virulent Wolbachia to modulate disease transmission by insects. J. Invertebr. Pathol 84(1):24–29 [DOI] [PubMed] [Google Scholar]
- Buchman AB, Ivy T, Marshall JM, Akbari OS, Hay BA. 2018a. Engineered reciprocal chromosome translocations drive high threshold, reversible population replacement in Drosophila. ACS Synth. Biol 7(5):1359–70 [DOI] [PubMed] [Google Scholar]
- Buchman AB, Marshall JM, Ostrovski D, Yang T, Akbari OS. 2018b. Synthetically engineered Medea gene drive system in the worldwide crop pest Drosophila suzukii. PNAS 115(18):4725–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bull JC, Bonsall MB. 2008. Overcompensatory population dynamic responses to environmental stochasticity. J. Anim. Ecol 77:1296–305 [DOI] [PubMed] [Google Scholar]
- Bull JJ. 2017. Lethal gene drive selects inbreeding. Evol. Med. Public Health 2017(1):1–16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bull JJ, Remien CH, Gomulkiewicz R, Krone SM. 2019a. Spatial structure undermines parasite suppression by gene drive cargo. PeerJ. 7:e7921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bull JJ, Remien CH, Krone SM. 2019b. Gene-drive-mediated extinction is thwarted by population structure and evolution of sib mating. Evol. Med. Public Health 2019(1):66–81 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt A. 2003. Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc. R. Soc. B Biol. Sci 270(1518):921–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt A. 2014. Heritable strategies for controlling insect vectors of disease. Philos. Trans. R. Soc. B Biol. Sci 369(1645):20130432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt A, Trivers R. 2006. Genes in Conflict: The Biology of Selfish Genetic Elements. Cambridge, MA: Harvard Univ. Press [Google Scholar]
- Cash SA, Lorenzen MD, Gould F. 2019. The distribution and spread of naturally occurring Medea selfish genetic elements in the United States. Ecol. Evol 9(24):14407–16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers LK, Singleton GR, Hinds LA. 1999. Fertility control of wild mouse populations: the effects of hormonal competence and an imposed level of sterility. Wildl. Res 26(5):579–91 [Google Scholar]
- Champer J, Buchman A, Akbari OS. 2016. Cheating evolution: engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet 17(3):146. [DOI] [PubMed] [Google Scholar]
- Champer J, Champer SE, Kim IK, Clark AG, Messer PW. 2019a. Design and analysis of CRISPR-based underdominance toxin-antidote gene drives. bioRxiv 861435. 10.1101/861435 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champer J, Kim IK, Champer SE, Clark AG, Messer PW. 2019b. Suppression gene drive in continuous space can result in unstable persistence of both drive and wild-type alleles. bioRxiv 769810. 10.1101/769810 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champer J, Kim IK, Champer SE, Clark AG, Messer PW. 2020a. Performance analysis of novel toxin-antidote CRISPR gene drive systems. BMC Biol. 18(1):1–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champer J, Lee E, Yang E, Liu C, Clark AG, Messer PW. 2020b. A toxin-antidote CRISPR gene drive system for regional population modification. Nat. Commun 11(1):1–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champer J, Liu J, Oh SY, Reeves R, Luthra A, et al. 2018. Reducing resistance allele formation in CRISPR gene drive. PNAS 115(21):5522–27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champer J, Yang E, Lee YL, Liu J, Clark AG, Messer PW. 2019c. Resistance is futile: a CRISPR homing gene drive targeting a haplolethal gene. bioRxiv 651737. 10.1101/65173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champer J, Zhao J, Champer SE, Liu J, Messer PW. 2020c. Population dynamics of underdominance gene drive systems in continuous space. ACS Synth. Biol 9:779–92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C-H, Huang H, Ward CM, Su JT, Schaeffer LV, et al. 2007. A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science 316(5824):597–600 [DOI] [PubMed] [Google Scholar]
- Clutton-Brock TH, Illius AW, Wilson K, Grenfell BT, MacColl ADC, Albon SD. 1997. Stability and instability in ungulate populations: an empirical analysis. Am. Nat 149(2):195–219 [Google Scholar]
- Craig GB, Hickey WA, VandeHey RC. 1960. An inherited male-producing factor in Aedes aegypti. Science 132(3443):1887–89 [DOI] [PubMed] [Google Scholar]
- Curtis CF. 1968. Possible use of translocations to fix desirable genes in insect pest populations. Nature 218(5139):368–69 [DOI] [PubMed] [Google Scholar]
- Curtis CF, Robinson AS. 1971. Computer simulation of the use of double translocations for pest control. Genetics 69(1):97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dao A, Yaro AS, Diallo M, Timbine S, Huestis DL, et al. 2014. Signatures of aestivation and migration in Sahelian malaria mosquito populations. Nature 516(7531):387–90 [DOI] [PMC free article] [PubMed] [Google Scholar]
- David AS, Kaser JM, Morey AC, Roth AM, Andow DA. 2013. Release of genetically engineered insects: a framework to identify potential ecological effects. Ecol. Evol 3(11):4000–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis S, Bax N, Grewe P. 2001. Engineered underdominance allows efficient and economical introgression of traits into pest populations. J. Theor. Biol 212(1):83–98 [DOI] [PubMed] [Google Scholar]
- Delborne J, Kuzma J, Gould F, Frow E, Leitschuh C, Sudweeks J. 2018. Mapping research and governance needs for gene drives. J. Responsible Innov 5(Suppl. 1):S4–12 [Google Scholar]
- Deredec A, Burt A, Godfray HC. 2008. The population genetics of using homing endonuclease genes in vector and pest management. Genetics 179(4):2013–26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deredec A, Godfray HC, Burt A. 2011. Requirements for effective malaria control with homing endonuclease genes. PNAS 108(43):E874–80 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhole S, Lloyd AL, Gould F. 2019. Tethered homing gene drives: a new design for spatially restricted population replacement and suppression. Evol. Appl 12(8):1688–1702 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhole S, Vella MR, Lloyd AL, Gould F. 2018. Invasion and migration of spatially self-limiting gene drives: a comparative analysis. Evol. Appl 11(5):794–808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Díaz M, Torre I, Arrizabalaga A. 2010. Relative roles of density and rainfall on the short-term regulation of Mediterraneanwood mouse Apodemus sylvaticus populations. Acta Theriol. 55(3):251–60 [Google Scholar]
- DiCarlo JE, Chavez A, Dietz SL, Esvelt KM, Church GM. 2015. Safeguarding CRISPR-Cas9 gene drives in yeast. Nat. Biotechnol 33(12):1250–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doncaster CP. 2006. Comment on “On the regulation of populations of mammals, birds, fish, and insects” III. Science 311(5764):1100. [DOI] [PubMed] [Google Scholar]
- Eckhoff PA, Wenger EA, Godfray HC, Burt A. 2017. Impact of mosquito gene drive on malaria elimination in a computational model with explicit spatial and temporal dynamics. PNAS 114(2):E255–64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edgington MP, Alphey LS. 2018. Population dynamics of engineered underdominance and killer-rescue gene drives in the control of disease vectors. PLOS Comput. Biol 14(3):e1006059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epopa PS, Millogo AA, Collins CM, North A, Tripet F, et al. 2017. The use of sequential mark-release-recapture experiments to estimate population size, survival and dispersal of male mosquitoes of the Anopheles gambiae complex in Bana, a west African humid savannah village. Parasites Vectors 10(1):376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esvelt KM, Smidler AL, Catteruccia F, Church GM. 2014. Concerning RNA-guided gene drives for the alteration of wild populations. eLife 3:e03401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher RA. 1930. The Genetical Theory of Natural Selection. Oxford, UK: Clarendon [Google Scholar]
- Fisher RA. 1937. The wave of advance of advantageous genes. Ann. Eugen 7(4):355–69 [Google Scholar]
- Flores HA, O’Neill LS. 2018. Controlling vector-borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol 16(8):508–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foster WA, Lea AO. 1975. Renewable fecundity of male Aedes aegypti following replenishment of seminal vesicles and accessory glands. J. Insect Physiol 21(5):1085–90 [DOI] [PubMed] [Google Scholar]
- Galizi R, Doyle LA, Menichelli M, Bernardini F, Deredec A, et al. 2014. A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nat. Commun 5:3977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galizi R, Hammond A, Kyrou K, Taxiarchi C, Bernardini F, et al. 2016. A CRISPR-Cas9 sex-ratio distortion system for genetic control. Sci. Rep 6:31139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gantz VM, Jasinskiene N, Tatarenkova O, Fazekas A, Macias VM, et al. 2015. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. PNAS 112(49):E6736–43 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Getz WM, Lloyd-Smith JO. 2006. Comment on “On the regulation of populations of mammals, birds, fish, and insects” I. Science 311(5764):1100. [DOI] [PubMed] [Google Scholar]
- Giese B, Frieß J, Barton N, Messer P, Débarre F, et al. 2019. Gene drives: dynamics and regulatory matters—a report from the workshop “Evaluation of Spatial and Temporal Control of Gene Drives,” April 4–5, 2019, Vienna. BioEssays 10(9):1900151. [DOI] [PubMed] [Google Scholar]
- Gimnig JE, Ombok M, Otieno S, Kaufman MG, Vulule JM, Walker ED. 2002. Density-dependent development of Anopheles gambiae (Diptera: Culicidae) larvae in artificial habitats. J. Med. Entomol 39(1):162–72 [DOI] [PubMed] [Google Scholar]
- Girardin L, Calvez V, Débarre F. 2019. Catch me if you can: a spatial model for a brake-driven gene drive reversal. Bull. Math. Biol 81(12):5054–88 [DOI] [PubMed] [Google Scholar]
- Godfray HC, North A, Burt A. 2017. How driving endonuclease genes can be used to combat pests and disease vectors. BMC Biol. 15(1):81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godwin J, Serr M, Barnhill-Dilling SK, Blondel DV, Brown PR, et al. 2019. Rodent gene drives for conservation: opportunities and data needs. Proc. R. Soc. B 286(1914):20191606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goubert C, Minard G, Vieira C, Boulesteix M. 2016. Population genetics of the Asian tiger mosquito Aedes albopictus, an invasive vector of human diseases. Heredity (Edinb.) 117(3):125–34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould F. 2008. Broadening the application of evolutionarily based genetic pest management. Evol. Int. J. Org. Evol 62(2):500–10 [DOI] [PubMed] [Google Scholar]
- Gould F, Huang Y, Legros M, Lloyd AL. 2008. A Killer–Rescue system for self-limiting gene drive of antipathogen constructs. Proc. R. Soc. B Biol. Sci 275(1653):2823–29 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould F, Schliekelman P. 2004. Population genetics of autocidal control and strain replacement. Annu. Rev. Entomol 49:193–217 [DOI] [PubMed] [Google Scholar]
- Grunwald HA, Gantz VM, Poplawski G, Xu XS, Bier E, Cooper KL. 2019. Super-Mendelian inheritance mediated by CRISPR-Cas9 in the female mouse germline. Nature 566(7742):105–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guagliardo SAJ, Lee Y, Pierce AA, Wong J, Chu YY, et al. 2019. The genetic structure of Aedes aegypti populations is driven by boat traffic in the Peruvian Amazon. PLOS Negl. Trop. Dis 13(9):e0007552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond A, Galizi R, Kyrou K, Simoni A, Siniscalchi C, et al. 2016. A CRISPR-Cas9 gene drive system targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol 34(1):78–83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond A, Kyrou K, Gribble M, Karlsson X, Morianou I, et al. 2018. Improved CRISPR-based suppression gene drives mitigate resistance and impose a large reproductive load on laboratory-contained mosquito populations. bioRxiv 360339. 10.1101/360339 [DOI] [Google Scholar]
- Hancock PA, Godfray HC. 2012. Modelling the spread of Wolbachia in spatially heterogeneous environments. J. R. Soc. Interface 9(76):3045–54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hancock PA, Ritchie SA, Koenraadt CJM, Scott TW, Hoffmann AA, Godfray HC. 2019. Predicting the spatial dynamics of Wolbachia infections in Aedes aegypti arbovirus vector populations in heterogeneous landscapes. J. Appl. Ecol 56(7):1674–86 [Google Scholar]
- Hancock PA, White VL, Callahan AG, Godfray CHJ, Hoffmann AA, Ritchie SA. 2016a. Density-dependent population dynamics in Aedes aegypti slow the spread of wMel Wolbachia. J. Appl. Ecol 53(3):785–93 [Google Scholar]
- Hancock PA, White VL, Ritchie SA, Hoffmann AA, Godfray HC. 2016b. Predicting Wolbachia invasion dynamics in Aedes aegypti populations using models of density-dependent demographic traits. BMC Biol. 14(1):96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hay BA, Oberhofer G, Guo M. 2021. Engineering the composition and fate of wild populations with gene drive. Annu. Rev. Entomol 66:In press. 10.1146/annurev-ento-020117-043154 [DOI] [PubMed] [Google Scholar]
- Herrando-Pérez S, Delean S, Brook BW, Bradshaw CJA. 2012. Density dependence: an ecological Tower of Babel. Oecologia 170(3):585–603 [DOI] [PubMed] [Google Scholar]
- Herrmann BG, Bauer H. 2012. The mouse t-haplotype: a selfish chromosome—genetics, molecular mechanism, and evolution. In Evolution of the House Mouse, ed. Macholán M, Baird SJE, Munclinger P, Piálek J, p. 297. Cambridge, UK: Cambridge Univ. Press [Google Scholar]
- Huang Y, Lloyd AL, Legros M, Gould F. 2009. Gene-drive in age-structured insect populations. Evol. Appl 2(2):143–59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang Y, Lloyd AL, Legros M, Gould F. 2011. Gene-drive into insect populations with age and spatial structure: a theoretical assessment. Evol. Appl 4(3):415–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huestis DL, Dao A, Diallo M, Sanogo ZL, Samake D, et al. 2019. Windborne long-distance migration of malaria mosquitoes in the Sahel. Nature 574(7778):404–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- James S, Collins FH, Welkhoff PA, Emerson C, Godfray HCJ, et al. 2018. Pathway to deployment of gene drive mosquitoes as a potential biocontrol tool for elimination of malaria in sub-Saharan Africa: recommendations of a scientific working group. Am. J. Trop. Med. Hyg 98(6_Suppl.):1–49 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khamis D, El Mouden C, Kura K, Bonsall MB. 2018. Ecological effects on underdominance threshold drives for vector control. J. Theor. Biol 456:1–15 [DOI] [PubMed] [Google Scholar]
- Kotsakiozi P, Evans BR, Gloria-Soria A, Kamgang B, Mayanja M, et al. 2018. Population structure of a vector of human diseases: Aedes aegypti in its ancestral range, Africa. Ecol. Evol 8(16):7835–48 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyrou K, Hammond AM, Galizi R, Kranjc N, Burt A, et al. 2018. A CRISPR-Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol 36(11):1062–66 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambert B, North A, Burt A, Godfray HC. 2018. The use of driving endonuclease genes to suppress mosquito vectors of malaria in temporally variable environments. Malar. J 17(1):154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lande R. 1985. The fixation of chromosomal rearrangements in a subdivided population with local extinction and colonization. Heredity (Edinb.) 54(3):323–32 [DOI] [PubMed] [Google Scholar]
- Lande R, Sæther B-E, Engen S. 1997. Threshold harvesting for sustainability of fluctuating resources. Ecology 78(5):1341–50 [Google Scholar]
- Láruson JÁ, Reed FA. 2016. Stability of underdominant genetic polymorphisms in population networks. J. Theor. Biol 390:156–63 [DOI] [PubMed] [Google Scholar]
- Leftwich PT, Bolton M, Chapman T. 2016. Evolutionary biology and genetic techniques for insect control. Evol. Appl 9(1):212–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leftwich PT, Edgington MP, Harvey-Samuel T, Carabajal Paladino ZL, Norman VC, Alphey L. 2018. Recent advances in threshold-dependent gene drives for mosquitoes. Biochem. Soc. Trans 46(5):1203–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legros M, Lloyd AL, Huang Y, Gould F. 2009. Density-dependent intraspecific competition in the larval stage of Aedes aegypti (Diptera: Culicidae): revisiting the current paradigm. J. Med. Entomol 46(3):409–19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legros M, Xu C, Morrison A, Scott TW, Lloyd AL, Gould F. 2013. Modeling the dynamics of a non-limited and a self-limited gene drive system in structured Aedes aegypti populations. PLOS ONE 8(12):e83354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehmann T, Weetman D, Huestis DL, Yaro AS, Kassogue Y, et al. 2017. Tracing the origin of the early wet-season Anopheles coluzzii in the Sahel. Evol. Appl 10(7):704–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitschuh CM, Kanavy D, Backus GA, Valdez RX, Serr M, et al. 2018. Developing gene drive technologies to eradicate invasive rodents from islands. J. Responsible Innov 5(Suppl. 1):S121–38 [Google Scholar]
- Lindholm AK, Dyer KA, Firman RC, Fishman L, Forstmeier W, et al. 2016. The ecology and evolutionary dynamics of meiotic drive. Trends Ecol. Evol 31(4):315–26 [DOI] [PubMed] [Google Scholar]
- Lindström J, Kokko H. 2002. Cohort effects and population dynamics. Ecol. Lett 5(3):338–44 [Google Scholar]
- Lord CC. 1998. Density dependence in larval Aedes albopictus (Diptera: Culicidae). J. Med. Entomol 35(5):825–29 [DOI] [PubMed] [Google Scholar]
- Macias VM, Ohm JR, Rasgon JL. 2017. Gene drive for mosquito control: Where did it come from and where are we headed? Int. J. Environ. Res. Public Health 14(9):1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magori K, Gould F. 2006. Genetically engineered underdominance for manipulation of pest populations: a deterministic model. Genetics 172(4):2613–20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manser A, König B, Lindholm AK. 2015. Female house mice avoid fertilization by t haplotype incompatible males in a mate choice experiment. J. Evol. Biol 28(1):54–64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manser A, Lindholm AK, Simmons LW, Firman RC. 2017. Sperm competition suppresses gene drive among experimentally evolving populations of house mice. Mol. Ecol 26(20):5784–92 [DOI] [PubMed] [Google Scholar]
- Marlow NJ, Thompson PC, Rose K, Kok NE. 2016. Compensatory responses by a fox population to artificial density reduction in a rangeland area in Western Australia. Conserv. Sci. West. Aust 10(3):1–10 [Google Scholar]
- Marshall JM. 2010. The Cartagena Protocol and genetically modified mosquitoes. Nat. Biotechnol 28(9):896–97 [DOI] [PubMed] [Google Scholar]
- Marshall JM, Akbari OS. 2018. Can CRISPR-based gene drive be confined in the wild? A question for molecular and population biology. ACS Chem. Biol 13(2):424–30 [DOI] [PubMed] [Google Scholar]
- Marshall JM, Hay BA. 2011. Inverse Medea as a novel gene drive system for local population replacement: a theoretical analysis. J. Hered 102(3):336–41 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall JM, Hay BA. 2012a. Confinement of gene drive systems to local populations: a comparative analysis. J. Theor. Biol 294:153–71 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall JM, Hay BA. 2012b. General principles of single-construct chromosomal gene drive. Evol. Int. J. Org. Evol 66(7):2150–66 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall JM, Pittman GW, Buchman AB, Hay BA. 2011. Semele: a killer-male, rescue-female system for suppression and replacement of insect disease vector populations. Genetics 187(2):535–51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- May RM. 1973. Stability and Complexity in Model Ecosystems, Vol. 1. Princeton, NJ: Princeton Univ. Press; [PubMed] [Google Scholar]
- Michalakis Y, Olivieri I. 1993. The influence of local extinctions on the probability of fixation of chromosomal rearrangements. J. Evol. Biol 6(2):153–70 [Google Scholar]
- Min J, Noble C, Najjar D, Esvelt K. 2017. Daisy quorum drives for the genetic restoration of wild populations. bioRxiv 115618. 10.1101/115618 [DOI] [Google Scholar]
- Moro D, Byrne M, Kennedy M, Campbell S, Tizard M. 2018. Identifying knowledge gaps for gene drive research to control invasive animal species: the next CRISPR step. Glob. Ecol. Conserv 13:e00363 [Google Scholar]
- Muriu SM, Coulson T, Mbogo CM, Godfray HC. 2013. Larval density dependence in Anopheles gambiae ss, the major African vector of malaria. J. Anim. Ecol 82(1):166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Natl. Acad. Sci. Eng. Med. 2016. Gene Drives on the Horizon: Advancing Science, Navigating Uncertainty, and Aligning Research with Public Values. Washington, DC: Natl. Acad. Press; [PubMed] [Google Scholar]
- Noble C, Adlam B, Church GM, Esvelt KM, Nowak MA. 2018. Current CRISPR gene drive systems are likely to be highly invasive in wild populations. eLife 7:e33423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noble C, Min J, Olejarz J, Buchthal J, Chavez A, et al. 2019. Daisy-chain gene drives for the alteration of local populations. PNAS 116(17):8275–82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- North AR, Burt A, Godfray HC. 2013. Modelling the spatial spread of a homing endonuclease gene in a mosquito population. J. Appl. Ecol 50(5):1216–25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- North AR, Burt A, Godfray HC. 2019. Modelling the potential of genetic control of malaria mosquitoes at national scale. BMC Biol. 17(1):26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- North AR, Godfray HC. 2018. Modelling the persistence of mosquito vectors of malaria in Burkina Faso. Malar. J 17(1):140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oberhofer G, Ivy T, Hay BA. 2019. Cleave and Rescue, a novel selfish genetic element and general strategy for gene drive. PNAS 116(13):6250–59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oberhofer G, Ivy T, Hay BA. 2020. Gene drive and resilience through renewal with next generation Cleave and Rescue selfish genetic elements. PNAS 117(16):9013–21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okamoto KW, Robert MA, Lloyd AL, Gould F. 2013. A reduce and replace strategy for suppressing vector-borne diseases: insights from a stochastic, spatial model. PLOS ONE 8(12):e81860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peacock E, Garshelis DL. 2006. Comment on “On the regulation of populations of mammals, birds, fish, and insects” IV. Science 313(5783):45. [DOI] [PubMed] [Google Scholar]
- Pham TB, Phong CH, Bennett JB, Hwang K, Jasinskiene N, et al. 2019. Experimental population modification of the malaria vector mosquito, Anopheles stephensi. PLOS Genet. 15(12):e1008440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piálek J, Barton NH. 1997. The spread of an advantageous allele across a barrier: the effects of random drift and selection against heterozygotes. Genetics 145(2):493–504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price TAR, Wedell N. 2008. Selfish genetic elements and sexual selection: their impact on male fertility. Genetica 132(3):295. [DOI] [PubMed] [Google Scholar]
- Prowse TAA, Adikusuma F, Cassey P, Thomas P, Ross JV. 2019. A Y-chromosome shredding gene drive for controlling pest vertebrate populations. eLife 8:e41873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prowse TAA, Cassey P, Ross JV, Pfitzner C, Wittmann TA, Thomas P. 2017. Dodging silver bullets: Good CRISPR gene-drive design is critical for eradicating exotic vertebrates. Proc. R. Soc. B Biol. Sci 284(1860):20170799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raban RR, Marshall JM, Akbari OS. 2020. Progress towards engineering gene drives for population control. J. Exp. Biol 223:jeb208181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajagopalan PK, Curtis CF, Brooks GD, Menon PK. 1977. The density dependence of larval mortality of Culex pipiens fatigans in an urban situation and prediction of its effects on genetic control operations. Indian J. Med. Res 65:77–85 [PubMed] [Google Scholar]
- Rasgon JL, Styer LM, Scott TW. 2003. Wolbachia-induced mortality as a mechanism to modulate pathogen transmission by vector arthropods. J. Med. Entomol 40(2):125–32 [DOI] [PubMed] [Google Scholar]
- Rašić G, Endersby-Harshman N, Tantowijoyo W, Goundar A, White V, et al. 2015. Aedes aegypti has spatially structured and seasonally stable populations in Yogyakarta, Indonesia. Parasites Vectors 8(1):610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rašić G, Filipović I, Weeks AR, Hoffmann AA. 2014. Genome-wide SNPs lead to strong signals of geographic structure and relatedness patterns in the major arbovirus vector, Aedes aegypti. BMC Genom. 15(1):275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robert MA, Okamoto K, Lloyd AL, Gould F. 2013. A reduce and replace strategy for suppressing vector-borne diseases: insights from a deterministic model. PLOS ONE 8(9):e73233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rode NO, Estoup A, Bourguet D, Courtier-Orgogozo V, Débarre F. 2019. Population management using gene drive: molecular design, models of spread dynamics and assessment of ecological risks. Conserv. Genet 20:671–90 [Google Scholar]
- Ross JV. 2006. Comment on “On the regulation of populations of mammals, birds, fish, and insects” II. Science 311(5764):1100. [DOI] [PubMed] [Google Scholar]
- Rouhani S, Barton N. 1987. Speciation and the “shifting balance” in a continuous population. Theor. Popul. Biol 31(3):465–92 [Google Scholar]
- Sæther B-E. 1997. Environmental stochasticity and population dynamics of large herbivores: a search for mechanisms. Trends Ecol. Evol 12(4):143–49 [DOI] [PubMed] [Google Scholar]
- Sánchez CMH, Wu SL, Bennett JB, Marshall JM. 2019. MGDriv E: a modular simulation framework for the spread of gene drives through spatially explicit mosquito populations. Methods Ecol. Evol 11:229–39 [Google Scholar]
- Schmidt TL, Barton NH, Rašić G, Turley AP, Montgomery BL, et al. 2017. Local introduction and heterogeneous spatial spread of dengue-suppressing Wolbachia through an urban population of Aedes aegypti. PLOS Biol. 15(5):e2001894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serebrovsky AS. 1940. On the possibility of a new method for the control of insect pests. Zool. Zhurnal 19:618–90 [Google Scholar]
- Serebrovsky AS. 1969. On the possibility of a new method for the control of insect pests. In Proceedings of a Panel on Application of the Sterile-Male Technique for the Eradication or Control of Harmful Species of Insects, Organised by the Joint FAO/IAEA Division of Atomic Energy in Food and Agriculture and Held in Vienna, 27–31 May 1968, pp. 123–237. Vienna: Int. Atomic Energy Agency [Google Scholar]
- Sibly RM, Barker D, Denham MC, Hone J, Pagel M. 2005. On the regulation of populations of mammals, birds, fish, and insects. Science 309(5734):607–10 [DOI] [PubMed] [Google Scholar]
- Sinclair ARE. 2003. Mammal population regulation, keystone processes and ecosystem dynamics. Philos. Trans. R. Soc. B Biol. Sci 358(1438):1729–40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinkins SP, Gould F. 2006. Gene drive systems for insect disease vectors. Nat. Rev. Genet 7(6):427–35 [DOI] [PubMed] [Google Scholar]
- Stephens PA, Sutherland WJ, Freckleton RP. 1999. What is the Allee effect? Oikos 87:185–90 [Google Scholar]
- Stokes AN. 1976. On two types of moving front in quasilinear diffusion. Math. Biosci 31(3–4):307–15 [Google Scholar]
- Sudweeks J, Hollingsworth B, Blondel DV, Campbell KJ, Dhole S, et al. 2019. Locally fixed alleles: a method to localize gene drive to island populations. Sci. Rep 9(1):1–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka H, Stone HA, Nelson DR. 2017. Spatial gene drives and pushed genetic waves. PNAS 114(32):8452–57 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twigg LE, Williams CK. 1999. Fertility control of overabundant species; can it work for feral rabbits? Ecol. Lett 2(5):281–85 [DOI] [PubMed] [Google Scholar]
- Vanderplank FL. 1947. Experiments in the hybridisation of tsetse-flies (Glossina, Diptera) and the possibility of a new method of control. Trans. R. Entomol. Soc. Lond 98:1–18 [Google Scholar]
- Vella MR, Gunning CE, Lloyd AL, Gould F. 2017. Evaluating strategies for reversing CRISPR-Cas9 gene drives. Sci. Rep 7(1):1–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walsh RK, Aguilar CL, Facchinelli L, Valerio L, Ramsey JM, et al. 2013. Regulation of Aedes aegypti population dynamics in field systems: quantifying direct and delayed density dependence. Am. J. Trop. Med. Hyg 89(1):68–77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walsh RK, Bradley C, Apperson CS, Gould F. 2012. An experimental field study of delayed density dependence in natural populations of Aedes albopictus. PLOS ONE 7(4):e35959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walsh RK, Facchinelli L, Ramsey JM, Bond JG, Gould F. 2011. Assessing the impact of density dependence in field populations of Aedes aegypti. J. Vector Ecol 36(2):300–7 [DOI] [PubMed] [Google Scholar]
- Ward CM, Su JT, Huang Y, Lloyd AL, Gould F, Hay BA. 2011. Medea selfish genetic elements as tools for altering traits of wild populations: a theoretical analysis. Evol. Int. J. Org. Evol 65(4):1149–62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wedell N. 2013. The dynamic relationship between polyandry and selfish genetic elements. Philos. Trans. R. Soc. B Biol. Sci 368(1613):20120049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wedell N, Price TAR. 2015. Selfish genetic elements and sexual selection. In Current Perspectives on Sexual Selection, ed. Hoquet T, pp. 165–90. Dordrecht, Neth.: Springer [Google Scholar]
- Wilkins KE, Prowse TAA, Cassey P, Thomas PQ, Ross JV. 2018. Pest demography critically determines the viability of synthetic gene drives for population control. Math. Biosci 305:160–69 [DOI] [PubMed] [Google Scholar]
- Williams RW, Berger A. 1980. The relation of female polygamy to gonotrophic activity in the ROCK strain of Aedes aegypti. Mosq. News 40(4):597–604 [Google Scholar]
- Windbichler N, Menichelli M, Papathanos PA, Thyme SB, Li H, et al. 2011. A synthetic homing endonuclease-based gene drive system in the human malaria mosquito. Nature 473(7346):212–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yakob L, Bonsall MB. 2009. Importance of space and competition in optimizing genetic control strategies. J. Econ. Entomol 102(1):50–57 [DOI] [PubMed] [Google Scholar]
- Yoshioka M, Couret J, Kim F, McMillan J, Burkot TR, et al. 2012. Diet and density dependent competition affect larval performance and oviposition site selection in the mosquito species Aedes albopictus (Diptera: Culicidae). Parasites Vectors 5(1):225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young ADM, Downe AER. 1982. Renewal of sexual receptivity in mated female mosquitoes, Aedes aegypti. Physiol. Entomol 7(4):467–71 [Google Scholar]