

Abstract
Anthelmintic resistance (AR) has thus far only rarely been reported for intestinal helminths of dogs and cats, in contrast to parasites of livestock and horses. We highlight possible reasons for this striking and important discrepancy, including ecological, biological and genetic factors and/or intervention regimens of key intestinal helminths concerning both host groups. In view of the current knowledge related to the genetics, mechanisms and principles of AR development, we point at issues which in our view contribute to a comparatively lower risk of AR development in intestinal helminths of dogs and cats. Finally, we specify research needs and provide recommendations by which, based on the available information about AR in ruminant and equine helminths, the development of AR in dog and cat helminths may best be documented, prevented or at least postponed.
Keywords: Drug resistance, Helminth, Development of AR, Canine, Feline, Worm
Graphical abstract
Highlights
-
•
Identifying causes of differences in anthelmintic resistance between livestock and dogs & cats.
-
•
Discussion of roles of AR mechanisms, genetics and husbandry in delayed AR evolution in dogs & cats.
-
•
Research needs for improved tools to monitor AR development in dogs & cats.
1. Introduction – anthelmintic resistance in intestinal helminths of dogs and cats
In dogs and cats helminth infections represent a significant health risk, especially in young animals; therefore strict and strategic anthelmintic parasite management and control is of relevance. Furthermore, some of the most frequent intestinal helminths of dogs (Toxocara canis, hookworms, Echinococcus spp.) or cats (Toxocara cati, hookworms) represent serious zoonotic risks, arising also from asymptomatic older infected pets (Beugnet et al., 2014; Deplazes et al., 2011; Drake and Carey, 2019; Little et al., 2009; Stafford et al., 2020). Similar to the situation in livestock, anthelmintic treatments are the most important component of systematic and effective prevention of clinical helminthoses. Notably, the prevention of patent infections in pets is fundamental in the context of veterinary public health.
Twenty years ago, Thompson and Roberts (2001) drew attention to the potential for drug resistance to emerge in intestinal helminths of dogs and cats, particularly nematodes: their concern was that a move away from infrequent applications targeting particular parasites (c.f. livestock) to a regular, metaphylactic use of broad-spectrum anthelmintic combinations could create an environment for AR to emerge. This potential risk may be considered to have been exacerbated by the subsequent proliferation of such anthelmintic combinations targeting different parasite groups simultaneously (defined as ‘Allwormers’) and the consequent increase of drug exposure.
There is evidence that AR has developed in intestinal nematodes of dogs in some geographical regions of the world. According to several reports (Jackson et al., 1987; Reynoldson et al., 1997; Kopp et al., 2007, 2008), resistance to pyrantel of the canine hookworm Ancylostoma caninum has been demonstrated in Australia. In a controlled study with dogs (n = 6) experimentally infected with a suspected pyrantel resistant A. caninum field population, an efficacy of only 25.7 %, based on reduction in worm burdens compared with the worm burden in untreated control dogs (n = 6), was observed (Kopp et al., 2007). In a subsequent smaller in vivo trial, involving two dogs, pyrantel efficacy of 71 % was found against another population of A. caninum from the Northern Territory of Australia (Kopp et al., 2008). In the absence of published findings on the current efficacy of pyrantel against A. caninum it is unknown how widespread pyrantel resistance is both within Australia and elsewhere at the moment. Pyrantel is the anthelmintic that has been used for longer than benzimidazoles or macrocyclic lactones for treating intestinal nematodes in dogs in Australia and might be therefore expected to be the first drug to fail as a result of AR. It is interesting that the demonstration of pyrantel resistance in A. caninum in dogs mirrors earlier evidence of AR in humans in Australia, where pyrantel had also been used frequently, and often suboptimally, to treat infections with A. duodenale (Reynoldson et al., 1997). The current use of drug combinations also including fenbendazole and/or macrocyclic lactones may mask the extent of AR to pyrantel. Future studies are highly needed to investigate anecdotal reports of treatment failures in dogs both within Australia and elsewhere. It needs to be pointed out that to the best knowledge of the authors there essentially is no routine surveillance concerning the anthelmintic efficacy established anywhere in the world. Furthermore, compared to the situation in livestock (Rose Vineer et al., 2020), anthelmintic efficacy field surveys in dogs and cats are extremely sparse. Only two examples we found for Europe with the study by (Miró et al., 2007) performed in stray dogs in Spain and a respective study from Germany (Becker et al., 2012), both showing no evidence of anthelmintic resistance concerning the employed benzimidazole drugs against the present intestinal nematodes, as well as for praziquantel against cestodes. However, clearly here and as always, ‘lack of evidence does not mean evidence of absence’, and certainly much more testing for drug efficacy is necessary in dogs and cats.
An alarming new issue in this respect has recently been documented in the USA, where high levels of AR involving multiple drug classes (benzimidazoles, pyrantel and macrocylic lactones) have been found in several A. caninum isolates either originating directly or presumably indirectly from intensively dewormed greyhounds (Jimenez Castro et al., 2019). The multi-drug resistance (against benzimidazoles, pyrantel and macrocyclic lactones) in one of these greyhound isolates was confirmed recently in a controlled trial, where eventually it was found that the cyclooctadepsipeptide emodepside was fully effective (Jimenez Castro et al., 2020). Similarly, another benzimidazole and macrocyclic lactone multi-drug resistant A. caninum isolate was described recently in the USA, also from a greyhound (Kitchen et al., 2019a). Not surprisingly, a long history of frequent (sometimes bi-weekly) anthelmintic treatments of dogs that were kept on confined ground with optimal environmental conditions for the development of hookworm larvae has led to very effective selection for AR (Jimenez Castro et al., 2019).
With respect to ascarids, which are the most prevalent intestinal helminths in dogs and cats in Europe, there are no confirmed cases of AR so far. To the best of our knowledge, only one published study reports on the selection of AR in T. canis, again occurring on a greyhound breeding farm in the USA (Ridley et al., 1994). Without providing further details on the study data, e.g., concerning faecal egg counts or worm burden, this publication states that the efficacy of a 5 mg/kg pyrantel pamoate treatment in two experimentally infected animals was 81.6 %, whereas the efficacy of three day 50 mg fenbendazole/kg/day was 100 %. Naturally infected puppies (randomised in groups of 5 puppies) from two greyhound bitches were treated with either 5 mg pyrantel pamoate, 8.3 mg pyrantel pamoate or three days of 50 mg fenbendazole/kg/day, resulting in 83.8 %, 99.9 % and 100 % efficacy, respectively. However, the small group sizes, lack of statistical analyses and missing details concerning the parasitological data, do not allow an assessment of whether these findings confirm the presence of AR against pyrantel pamoate in T. canis.
Concerning tapeworms, to the best of the authors’ knowledge, resistance to the isoquinoline praziquantel as the cestocidal drug of choice has only recently been reported for the first time: a Dipylidium caninum worm population infecting dogs in the USA was apparently also resistant to epsiprantel, and could only be removed by treatment with nitroscanate or a combination of pyrantel/praziquantel/oxantel (Chelladurai et al., 2018). Earlier, there has been some concern raised that co-infection with the large pseudophyllidean tapeworm Spirometra erinacei might coincide with a reduced praziquantel efficacy against other cestodes such as Echinococcus granulosus (Jenkins, 1998). However, neither has this yet been reported again nor experimentally verified.
Despite the AR reports in dog parasites described above, it is evident that on a global scale the magnitude, intensity and quality (e.g., single or multi-drug class resistance) of the problem in dog/cat parasites is far less advanced than it is in livestock parasites (Box 1). Moreover, for cats the authors are not aware of any study describing AR in feline intestinal helminths. Nevertheless, we discuss the issue of potential AR development in cats jointly with that in dogs here because the spectrum of intestinal helminths infecting these two hosts is very similar, in part even overlapping, and dogs and cats share several other features such as equivalent anthelmintic drug classes and worm control approaches.
Box 1. AR in intestinal helminths of ruminants and horses.
-
•
AR occurring in numerous intestinal helminth species particularly in small ruminants, in horses and more recently also in cattle (Wolstenholme et al., 2004; Demeler et al., 2009; Gasbarre et al., 2009; Falzon et al., 2014; Cotter et al., 2015; Rose et al., 2015a) against all broad spectrum anthelmintics (Table 1).
-
•
AR is most advanced in small ruminant gastro-intestinal nematodes (GIN)s and therein most prevalent against benzimidazoles, but also frequent against imidazothiazoles/tetrahydropyrimidines and, increasingly, macrocyclic lactones (Kaplan and Vidyashankar, 2012; Rose et al., 2015b; Rose Vineer et al., 2020) leading to multiple drug class resistance
-
•
In horses AR evolved on a global scale in cyathostomins and Parascaris spp., where AR is involving mainly benzimidazoles and macrocyclic lactones, respectively (von Samson-Himmelstjerna, 2012).
-
•
Concerning trematode and cestode species AR is much less evolved: in trematodes, the liver fluke Fasciola hepatica developed resistance against the benzimidazole triclabendazole and also against some other flukicides. This occurred in many geographical regions during the recent decades (Fairweather et al., 2020; Hanna et al., 2015; Rose Vineer et al., 2020; Wolstenholme et al., 2004).
-
•
As shown for benzimidazole resistance in sheep (Silvestre et al., 2001) and cattle (Knapp-Lawitzke et al., 2015) nematodes, repeated underdosing or, as shown in horses (Drogemuller et al., 2004), gradual increasing dosaging can lead to selection/increase of AR.
Alt-text: Box 1
2. Principles of anthelmintic resistance development in livestock
The development of AR in parasitic helminths is currently best understood in ruminant trichostrongyles, where it has been shown to occur via selection with both hard and soft selective sweeps occurring on the individual farm level as revealed by selection of one or multiple resistance haplotypes in the population, respectively (Redman et al., 2015). The population size of these parasites is so huge (Gilleard, 2013) that multiple distinct haplotypes conferring resistance might be present in the population before the start of selection or arise spontaneously via recurrent mutation during the selection process, allowing selection of multiple haplotypes in soft sweeps (Redman et al., 2015). Thus, the initially susceptible, drug naïve parasite population either already includes a small number of resistant individuals or resistant individuals arise through new, recurrent mutation during repeated treatments. These resistant worms survive the first treatments and pass the resistance genotype/genotypes on to their progeny. Within a few generations, the initially small resistant subpopulation begins to expand, and treatment failure eventually ensues. The low sensitivity of most phenotypic AR assays means that AR is unlikely to be detectable until the resistant individuals comprise >25 % of the population (Martin et al., 1989). The speed of selection depends primarily on factors such as treatment frequency, the mode of inheritance of resistance and the size of the refugium. Several mathematical models support and field experiments, in which resistance is monitored closely, have validated this general view (Dobson et al., 2011; Learmount et al., 2012; Cornelius et al., 2016; Leathwick et al., 2019; Sauermann et al., 2019). They have elucidated the roles that anthelmintic efficacy, mode of resistance inheritance, features of parasite biology such as fecundity, lifespan and genetic diversity and the size of the refugia population all play in determining the rate at which selection occurs. Genetic diversity is a function of effective population size and mutation rate. While to the best of our knowledge there are no data available concerning the mutation rate in parasitic helminths, this is well studied in the model nematode Caenorhabditis elegans, where the mutation rate was approximately 3 × 10−10 mutations per site per cell division (Denver et al., 2009). This is similar to the one observed in other eukaryotes such as insects, but also humans and we anticipate that in parasitic helminths it will be similar. Accordingly, in parasitic nematodes it is assumed that mutation rate is not the main reason for the extreme genetic diversity in some species but that population size is the major factor (Gilleard and Redman, 2016).
The term refugium is of substantial relevance since it describes that part of a parasite population which is actually not exposed to the drug when anthelmintic treatments are being applied. For example, the free living larval stages of strongyles on the pasture/in soil, or the histotropic or migrating stages of strongyles or ascarids when treated with the tetrahydropyrimidine pyrantel, which only acts against stages in the intestinal lumen, are examples of parasite refugia with respect to selection for AR. The larger the part of the parasite population in refugia, the more the offspring of the resistant treatment survivors will be diluted in the next generation by the offspring of the refugia population that has escaped selection, and consequently the slower resistance will evolve. In horse and livestock nematodes, it has been demonstrated that maintaining a part of the worm population in refugia, e.g. by selectively treating only a fraction of the flock or herd, reduces or prevents the development of AR (Martin et al., 1981; Van Wyk, 2001; Kenyon and Jackson, 2012; Cornelius et al., 2016; Leathwick et al., 2017). However, there are several observations which suggest that a large refugium does not necessarily prevent the development of AR, one example being the widespread occurrence of pyrantel resistance in horse cyathostomins. In this context, it seems important to point out that, to generate an effective refugium, the unselected part of the respective worm population needs to be accessible for the host population in which the anthelmintic selection is being exerted.
More recently, careful genetic studies (primarily of known benzimidazole resistance alleles of isotype-1 β-tubulin, but also whole genome studies where the “resistance genes” per se are not identified) have demonstrated that soft selection of multiple pre-existing resistant worms is most likely, while genetic variability is largely maintained. However, hard selection of a single allele leading to strong loss of local variability may also contribute to AR development under some circumstances (Chaudhry et al., 2016; Redman et al., 2015). These general principles of soft sweeps that maintain overall high levels of genetic diversity within and between resistant isolates (presumably via selection from a pre-existing but small resistant sub-population within the drug naïve population) have been shown to apply not only in trichostrongyle parasites of livestock but also in at least two filarial nematodes with well documented AR (Dirofilaria immitis and Onchocerca volvulus) (Doyle et al., 2017; Sanchez et al., 2020), and are likely to extend to gastrointestinal helminths of dogs and cats.
Concerning the spatial spread of resistant parasite populations, mainly host and management factors are of relevance. Translocation of ruminant and equine hosts infected with resistant worms has repeatedly been shown to effectively lead to the spread of AR within and across countries (Artho et al., 2007; Chaudhry et al., 2015).
3. What is different in dogs and cats concerning occurrence, treatment and control of intestinal parasites compared with ruminants and horses?
Expert advice (e.g. European Scientific Counsel Companion Animal Parasites, ESCCAP or the Companion Animal Parasite Council, CAPC and the Tropical Council for Companion Animal Parasites, TroCCAP; (Dantas-Torres et al., 2020) and textbook recommendations (Bowman et al., 2014; Deplazes et al., 2016) concerning anthelmintic treatment frequency are based on the biology of the parasites (e.g. prepatent period and mode of transmission) and efficacy of the anthelmintics against the parasite stages involved. Animal age and evaluation of the individual risks of infection with clinically and/or zoonotically relevant helminths are also considered. Consequently, no strict treatment frequencies but rather a range between one to twelve treatments per year has been recommended. In comparison with the situation for sheep or horse parasites, however, the scientific evidence base for recommendations for controlling intestinal dog and cat helminths is weak and further research is needed. Reasons for this lack of evidence-based metaphylactic strategies include the difficulties of conducting large experimental trials (particularly ethical concerns) and limitations of studies with naturally infected dogs, as shown, for example, by Vienažindienė et al., 2018).
In an Australian questionnaire survey, approx. 50 % of the dog owners reported treating their dogs on a three-monthly basis against intestinal helminths (Palmer et al., 2010). More recent surveys in Germany, France and Spain indicated that the mean annual anthelmintic treatment frequency in dogs and cats was 2.1, 2.3, 3.1 and 1.7, 2.3, 2.6, respectively (Strube et al., 2019; Miró et al., 2020). These relatively low treatment frequencies are in contrast to treatment frequencies in some livestock species. In sheep, where generally only lambs are being repeatedly treated during the grazing season, globally treatment frequency ranges between only 1–2 treatments per season and bi-weekly treatments, depending on local epidemiology. In horses, all age classes usually receive anthelmintic treatments repeatedly during the year: a German study documented an average of 2.7 treatments per year in adult horses whereas foals experienced an average of 4.5 treatments annually, with 10 % being treated on a monthly basis (Fritzen et al., 2010). Accordingly, it seems probable that overall the treatment frequency against intestinal helminths in grazing animals such as sheep and horses is higher than in dogs and cats. This must be considered an assumption, however, due to the sparseness of relevant published data.
However, it has to be acknowledged that much about how anthelmintic products are used is driven by factors not related with intestinal helminth infection associated concerns. For instance, in the USA, there is rightful concern about infections with the potentially deadly and zoonotic Dirofilaria immitis (McCall et al., 2008). Thus, much of parasite control has been bundled into heartworm prevention products. Since monthly administration of low dose macrocyclic lactones can prevent heartworm infections, monthly treatment supplanted the prior utilization of daily diethylcarbamazine. The available products have since morphed into all-round monthly parasite control: historically, first, pyrantel pamoate was added to the tablets for internal parasite control. Next, macrocyclic lactones, e.g., milbemycin oxime, were supplied in products that prevented heartworms and killed internal parasites. Today, upon this monthly orally administered platform, more and more has been added, e.g., flea control with lufenuron (a chitin inhibitor), spinosad (an insecticide); and to topical applied and absorbed heartworm preventive/dewormers have been added also topical insecticides, e.g., imidacloprid (Noack et al., 2021). In addition, for tapeworm control, praziquantel has been added to the monthly heartworm preventives. With the development of the isoxazolines, these are now being added for tick and mite control as well, with one oral monthly product containing an isoxazoline, a macrocyclic lactone, and pyrantel pamoate. Therefore, in the USA, it is remarkably difficult to recommend only treating dogs for roundworms and tapeworms 2 to 4 times a year. The reason is that many are already being treated for these parasites monthly. Thus, products have been designed that kill worms, fleas, and ticks that are part of the veterinary pharmacopeia, without enough consideration being given to what overuse might mean. As this heartworm-control driven treatment strategy certainly bears the risk of AR selection, in areas where the former is being employed anthelmintic drug efficacy testing should become a routine procedure in all dog worm management programs.
In the context of treatment practices, the combination of anthelmintics with different modes of action but targeting the same parasite species (such as the benzimidazoles and pyrantel, both targeting intestinal helminths) is considered the most effective strategy to prevent or delay the development of AR in ruminants and horses (Bartram et al., 2012; Geary et al., 2012). This strategy is most effective when the target worm populations have not yet developed resistance to either compound class in the drug combination product but has also shown significant benefits even when moderate levels of resistance to both compounds alone is present. For dogs and cats, a product combining febantel and pyrantel that exhibits a synergistic efficacy (Mehlhorn et al., 2003) has been used frequently for decades, unintentionally establishing a propitious situation that conserves susceptibility to these compounds. In livestock, products containing compounds with different mode of action targeting the same helminths are also very often employed in Australia, New Zealand and South Africa but were introduced only in response to the widespread occurrence of AR (Geary et al., 2012). To the best of our knowledge such products have so far not been licensed in the European Union nor in North America.
Trichostrongylids in sheep and goats and cyathostomins in horses share basic biology and ecology with hookworms of companion animals: like trichostrongylids of livestock, hookworms of companion animals can be transmitted by the faecal oral route from contaminated environments with direct life cycles. The key differences are the extent of environmental contamination and contact of companion animals with those contaminated environments. Livestock are kept as grazing animals in constant contact with contaminated pasture, with the result that parasitism can occur at very high prevalences of often over 80 %, even in adult animals. Populations of parasite species with high prevalences, high infection intensity and a high reproduction potential can be expected to also have a high genetic diversity. For the canine hookworm A. caninum, regionally high prevalence, infection intensity and reproductive potential also occur (at least in young animals), but because puppies and young dogs are usually not kept in big herds, the effective population size of A. caninum populations will certainly in most situations be considerably smaller than that of most trichostrongyle or cyathostomin populations (Box 2, Box 3). Nevertheless, for canine hookworms a relative high risk for AR development may be anticipated in situations where the dogs are constantly exposed to reinfection, e.g. when being kept in the same confined space, as it is the case for the above mentioned greyhound cases (Jimenez Castro et al., 2019; Kitchen et al., 2019b); Jimenez Castro et al., 2020).
Box 2. Key differences between grazing animals and dogs/cats concerning AR development.
-
•
While in livestock and horses gastro-intestinal helminths with a high prevalence of anthelmintic resistance (AR) exhibit a direct life cycle (e.g. Haemonchus contortus in sheep and cyathostomins in horses), major intestinal helminths of dogs/cats also employ intermediate and/or paratenic hosts (e.g. Toxocara canis, T. cati, hookworms and tapeworms). Furthermore, large fox, stray dog and cat populations regularly display higher prevalences than domestic animals and therefore represent reservoirs of the worm populations without exposure to anthelmintics. In addition, overlapping areas between wild and domestic ruminants are limited, contrasting with large overlapping roaming areas between wild and domestic carnivores (see Fig. 1). These factors might considerably extend the refugia of unselected fractions of the respective total worm populations.
-
•
Effective population size (Ne) of a trichostrongyle population is assumed to be several orders of magnitude larger than in pet animal worm populations (Box 3), and thus the chances for spontaneous AR-associated mutation are much lower.
-
•
Pet animals are mostly being kept, surveyed and treated as individual animals and not as flocks or herds, which impacts the population size of the parasites they host.
-
•
Treatment frequency: In intensively reared lambs and in previous years also in foals on many stud farms, anthelmintic treatments are/were routinely applied often at monthly, sometimes even more frequent intervals during the course of their first year of life. Though this is not the case for calves, which usually receive much less frequent anthelmintic treatments. In puppies and kittens it is recommended to treat at biweekly intervals but only until weaning. Thus the total number of treatments may be considered to be overall similar to that in intensively reared lambs and foals.
-
•
Accumulation of AR-associated alleles occurs in livestock parasite populations on pasture/in the environment while this does not occur or at least to a much lesser extent concerning dog/cat helminths, also due to hygienic measures (e.g. collection and disposal of faeces by pet owners).
Alt-text: Box 2
Box 3. Estimation of population sizes for sheep and canine parasites.
The effective population size (Ne) is based on the reproduction rate of an idealized population with random allele distribution. The following formula is used to calculate the effective population size:
| Ne = 4NfNm/(Nf + Nm) |
Where Nf stands for the number of females and Nm for the number of males.
Ne will always only represent a fraction of the total population size as there are many factors which exclude individuals from the reproductive process.
Gilleard has estimated the population size for H. contortus in a single sheep flock (Gilleard, 2013) and based on his numbers we estimate:
-
•
500 sheep in a flock
-
•
500 female worms per host
-
•
2000 eggs per female and day
-
•
5 × 108 eggs on pasture per day
-
•
Lambs remain infected/reinfected for at least a year
-
•
~1.8 × 1013 eggs on pasture per year
For A. caninum, data on worm and egg counts in naturally infected dog puppies have been published (Sowemimo and Asaolu, 2008). On average approximately 6.5 female worms were found per infected dog and the female egg production was estimated to be 2074 (range 821–25,160). In the following calculation we will use slightly higher numbers,
-
•
10 puppies in a typical breeding kennel with two pregnant bitches per year
-
•
20 female worms per puppy
-
•
5000 eggs per female and day
-
•
4 × 106 eggs per kennel per day
-
•
Since worms can reach an age of up to six month, puppies are assumed to shed eggs for at most 10 month (Deplazes et al., 2016)
-
•
3 × 108 eggs per kennel and year
For T. canis, naturally infected puppies have been shown to show highly variable egg shedding in the range of 1000 to 34,000 epg with an average of about 5000 epg (Fisher et al., 1994). For the following estimation, we will use slightly higher values:
-
•
10 puppies in a typical breeding kennel with two pregnant bitches per year
-
•
10,000 epg
-
•
40 g faeces per puppy (of course highly dependent on the breed/size)
-
•
4 × 106 eggs per kennel per day
-
•
After prenatal infection of puppies, prepatency was reported to be at least 3 weeks post partum and 70 % of the adult worms were expelled around 9–10 weeks after birth (Schnieder et al., 2011b). However, according to Deplazes et al. (2016) the adult worms can live on average for 4 month. Thus we assumed 100 days of egg shedding if puppies were left untreated. Since reinfections rarely lead to patent infections, contribution of older dogs to contamination of the environment can be neglected.
-
•
4 × 108 eggs per kennel per year
This means the number of parasites produced by a typical group of lambs infected with H. contortus and dog puppies infected with A. caninum or T. canis is estimated to differ by about 60,000 and 45,000-fold, respectively.
The effective population size is of course much smaller for all three parasite species. However, it can be assumed that survival of H. contortus larvae on pasture is much higher than survival of A. caninum or T. canis eggs in a kennel, where most of the eggs can be expected to be removed together with the faeces of the puppies or bitches even before infectious stages have developed.
Alt-text: Box 3
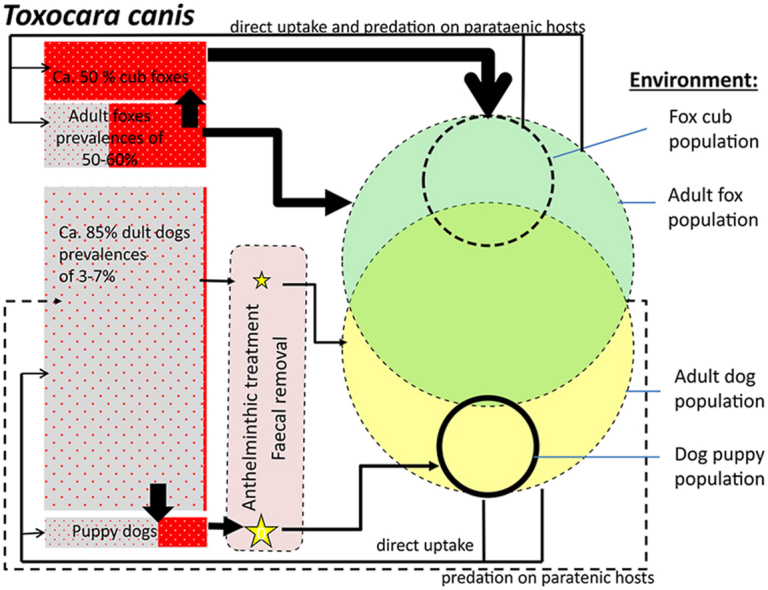
With respect to ascarids, widespread AR has been documented in Parascaris sp. of horses but so far no relevant AR occurrence has been found in canine T. canis despite decades of anthelmintic treatments, especially of puppies. Fig. 1 visualises the differences in parasite transmission for these two key ascarid species. Both species occur with low egg shedding prevalences in adult horses and dogs, respectively, but high prevalences are seen in foals and young horses and in puppies less than 6 months of age. As pointed out above, contact with contaminated environments is likely the most significant epidemiological factor determining prevalence and population size of horse ascarids. Parascaris transmission in horses is mainly dominated by high rates of reproduction of the parasite in foals and young horses. They intensively contaminate their environment with Parascaris eggs, where these can remain infective for months, including between grazing seasons (Nielsen, 2016). In Parascaris, AR selection is favoured by the fact that these heavily infected foals and young horses are generally under the highest anthelmintic pressure (Fritzen et al., 2010) and thus contaminating their environment with the eggs of AR survivors of treatment. This is exacerbated by the fact that hygienic measures to minimise environmental contamination and contact are mostly not employed. In addition, there are comparatively limited effective refugia due to very low egg shedding prevalences in untreated older horses, lack of paratenic hosts or wild animal reservoirs, and a lack of hypobiotic stages and vertical transmission (Fig. 1). In contrast, T. canis eggs in the environment are not the major source of infection for canid puppies: without anthelmintic prevention, these are regularly infected in utero by reactivated third stage larvae from the bitches (Schnieder et al., 2011). Anthelmintic treatment for T. canis is most intense in puppies under four months of age to prevent clinical signs of toxocarosis. Therefore, such frequently treated puppies theoretically represent the most important risk group for AR selection in Toxocara. However, puppies have a restricted habitat range as compared with adult dogs and foxes and may only marginally contribute to the general environmental contamination with eggs (Nijsse et al., 2015). Furthermore, effective hygienic measures will usually be employed in dog breeding kennels and tend to correspond with intensive treatment, thus avoiding the build-up of an AR selected parasite population in the environment. Additionally, depending on the endemic area and involvement of wild animals in the transmission, potential refugia populations of T. canis can establish at distinct levels, namely as adult worms in canid definitive hosts (e.g., domestic dogs, foxes and coyotes), in the environment in the form of eggs with high tenacity and in paratenic hosts. In particular, the potential role of foxes in Europe or coyotes in North America in the context of limiting AR development in T. canis by providing substantial refugia has to be considered. Noteworthy, T. canis is frequently found in fox cubs with for example prevalences of patent infections in Europe of 73%–87 % and, to a slightly lesser degree, also in adult foxes (prevalences of 37–53 %) (Saeed et al., 2006; Reperant et al., 2007), all not exposed to anthelmintic treatments. In general, dog population densities are much higher (around 10 dogs/km2 in rural and 115/km2 in urban areas) as compared with densities of foxes (around 1–5/km2 in rural, 6–32/km2 in urban areas) (Hegglin and Deplazes, 2013). However, the proportion of highly susceptible cubs (=puppies), is much higher in the fox population (around 30 % of the total population in spring as compared with around 5–15 % puppies <6 month of age in the dog population in industrialized societies). Toxocara canis prevalence in wild carnivores other than foxes appears to be generally lower, with study findings ranging between 19 % (Bridger et al., 2009) and 24 % (Redman et al., 2016) in coyotes, between 0.2 and 2% in wolves in Canada (Wapenaar et al., 2013) and up to 13 % in Europe (Bindke et al., 2019; Bryan et al., 2012; Szafrańska et al., 2010), and up to 14 % in golden jackals in Asia (Siyadatpanah et al., 2019). Thus, overlapping wildlife and domestic parasite transmission, the combination of diverse refugia and other factors mentioned above may explain why AR evolution in T. canis has essentially not been observed.
Fig. 1.

Ascarid transmission ecology, host population structure and anthelmintic exposure. Red areas show the relative proportion of patent infections, size of arrows the significance of the transmission and the size of the yellow stars represents the relative anthelmintic exposure. (A) Parascaris transmission in adult horses and foals (<1 year) and young horses (under 3 years): foals and young horses present highest prevalences (up to >60 % (Fritzen et al., 2010; Lyons et al., 2006)) and contribute to the major environmental contamination with eggs. Infective larvae persist over years in mostly small and defined environmental habitats (pastures/stables). Pasture and stable management (faecal removal) and anthelmintic exposure can reduce environmental contamination. (B) Toxocara canis transmission in adult pet dogs and foxes and their puppies/cubs (under 6 month of age). Parasite transmission ecology is more complex than in example A, based on the presence of wild canids (i.e. red fox dominating in Europe, coyotes in Northern America, dingoes in Australia, raccoon dogs in Eastern-Central Europe) or stray, not owned, domestic dogs. The example represents a typical European urban situation with a high dog population (110 dogs/km2 (95 % > 6 month of age) and 30 foxes/km2 (50% > 6 month of age in late summer). Toxocara canis prevalences with hypobiotic larval stages is approximately 100 % (as indicated by small dots) in both hosts; intestinal infections in adult dogs (approximately 85 % of the total dog population) 1–3%, their puppies 20 %, and in adult foxes 50–60 %, their cubs nearly 100 %. Besides transmission in the environment (black arrows), intrauterine and lactogenic vertical transmission to the offspring is indicated by dashed black arrows. The environmental contamination with Toxocara eggs is strongly dependent on the use of effective anthelmintics and hygiene measures (removal of faeces). As compared with example A, canid overlapping environments are larger and little determined (dashed lines) for adult populations; puppies use restricted and smaller areas (fox dens, kennels, private gardens).
Tapeworms of canids (especially Echinococcus and Taenia spp.) are the only intestinal parasites that have so far been controlled, with variable success, by adopting intensive anthelmintic treatment programs with praziquantel. Praziquantel is highly efficient, eliminating 99–100 % of the Echinococcus burdens (Gemmell et al., 1980). Mass praziquantel treatment of dogs, combined with an intensive education program, was a major successful control strategy against cystic echinococcosis in New Zealand, Tasmania, and parts of Cyprus (Craig et al., 2017). In other parts of the world, especially under continental situations, control programs successfully reduced the prevalences of E. granulosus s.l. in dogs, but elimination was not achieved (Craig and Larrieu, 2006). Mass treatment of foxes with praziquantel baits distributed monthly over several years also resulted in a strong and significant reduction of the environmental E. multilocularis egg contamination (Hegglin and Deplazes, 2013). Importantly, despite decades of praziquantel (mass) treatments, no indications of AR have been observed in canine taeniid species so far. In the particular case of E. multilocularis in Europe, pet dogs are only marginally contributing to the contamination of vole habitats with worm eggs, and are therefore considered relevant for zoonotic transmission but less significant for maintaining the typical fox-vole cycle of the parasite, which represents the main source of infection for dogs/cats (Romig et al., 2017). Therefore, monthly metaphylactic treatment of dogs with a high risk of being sporadically infected with E. multilocularis is justified because it prevents the excretion of directly infective eggs shed with the faeces and their dispersion in the immediate surroundings of their human caregivers. Basically, as the prevalences of patent E. multilocularis infections are relatively low in pet dogs (e.g. under 1%; Dyachenko et al., 2008; Deplazes et al., 2011; Bružinskaitè-Schmidhalter et al., 2012), most metaphylactic treatments will not be encountered by the parasite. Therefore, despite high treatment frequencies, such treatments are less likely to contribute to AR development. However, the alarming report of epsiprantel and/or praziquantel resistance in D. caninum in single dogs in the United States (Chelladurai et al., 2018) has to be considered seriously, especially in the context of future Echinococcus control programs in areas with simultaneous high prevalences of D. caninum. In contrast to the taeniid species, the life cycle of Dipylidium involves dogs and cats as definitive and fleas and lice as intermediate hosts. The high reproduction rate in definitive hosts and the relatively short development time in intermediate hosts plus the lack of paratenic hosts and other potential refugia, additionally combined with the possible small scale transmission (e.g. within a kennel or apartment) (Bowman et al., 2014; Deplazes et al., 2016) could favor AR development in D. caninum. Further analyses are needed to elucidate AR development and occurrence in D. caninum in the United States. This may hold true in particular if praziquantel is administered over years at the recommended high frequency of 4–12 times per year in “allwormers” or as a single compound in control programs against Echinococcus species. Therefore, suspected cases of AR should be deeper investigated using appropriate tests, and post-treatment controls may be suitable to differentiate between treatment failures due to AR versus underdosing due to various reasons.
In relation to dogs, household cats generally experience lower anthelmintic treatment frequencies, although cats assessed to have a high worm infection risk are considered to be treated more often (Strube et al., 2019). Moreover, in many industrialized countries (where the risk for AR development might be considered the highest) a considerable number of stray cats but only comparatively few stray dogs exist and the former do not receive anthelmintic treatments at all, thus presumably contributing to the maintenance of large refugia. Thus, the risk for development of AR in cat intestinal helminths can probably be considered as similar as or even lower than the one in dogs.
4. Concluding remarks
Potential reasons for the still comparatively rare finding, to date, of AR development in dog and cat intestinal helminths include major epidemiological and biological differences (e.g. larger refugia in dog/cat intestinal helminths), significantly different husbandry settings (e.g. individual rather than herd keepings, better hygiene options), overall lower anthelmintic treatment frequency at least in non-heartworm areas, plus a smaller parasite population size leading to a comparatively low genetic diversity in the parasite populations. These all are factors that normally prevent the build-up and spread of AR-selected infective worm stages in the environment of dogs and cats (Box 2, Box 3). However, the most recent findings of multiple drug class AR in A. caninum and D. caninum in dogs in the USA are a clear warning signal which should alarm veterinarians and pet owners as to the potential risk of AR development. Looking at the situation in the USA from a drug-history/parasitological perspective may provide some insight into normative facts contributing to AR development. Noteworthy, AR in intestinal helminths of dogs and cats has not yet been documented in most countries and it appears that there are practical and effective means to avoid AR development in dog/cat intestinal helminths in the future by applying lessons learnt from intestinal helminths of livestock. A key aspect to achieve this goal is the practical application of an integrated set of worm control procedures mitigating the risk of AR development (see Box 4). Despite the current absence of widespread AR in canine/feline intestinal helminths, there certainly is no reason for complacency and there is an obvious urgent need for more scientific data (see Outstanding Questions, Box 5).
Box 4. Prevention of AR development in dogs and cats.
-
•
Worm control recommendations should be scientifically based and performed under veterinary supervision with correct dosing based on careful weighing of the animals and using products according to manufacturer's instructions.
-
•
Restrict treatment frequency and intensity to the least required to ensure animal health and prevention of human exposure to zoonotic helminths based on individual risk analysis. For instance, regarding animal health, ESCCAP deworming frequency recommendations are based on the seasonal occurrence of Dirofilaria immitis in endemic regions and on risk behavior of the single dog regarding ingestion of gastropods to prevent canine angiostrongylosis. Regarding the prevention for zoonotic helminths, the assessment is based again on the individual risk behavior of dogs concerning the ingestion of intermediate and/or paratenic hosts of Toxocara canis and Echinococcus multilocularis.
-
•
Particularly in kennels/catteries, animal shelters or other places where dogs are kept in larger numbers (e.g. greyhound race tracks) and often treated with anthelmintics, regular coprological testing (including novel tools such as copro-DNA and -antigen testing) to monitor post treatment status is advisable. Before switching to another product check that drugs have been used appropriately, at the correct doses. In any case of putative reduced efficiency of the anthelmintic used, a professional analysis of the situation by veterinary parasitologists should be initiated to diagnose and control such developments in an early stage.
-
•
Remain vigilant concerning potential development of anthelmintic resistance and regularly (e.g. once per year) perform post treatment coprological check-ups.
-
•
If post treatment worm eggs are still found and coprophagia can be excluded, immediate treatment with a drug for which no resistance can be expected (e.g. emodepside as described by Jimenez Castro and Kaplan (2020), in order to prevent further shedding of eggs from resistant worms.
-
•
All animals newly obtained or to be introduced into a group of animals should be put under quarantine containment, examined for worm infections and, if positive, effectively treated.
-
•
Increase awareness of resistance problem by dog owners and motivate them to remove dog faeces from the environment to avoidcontamination. This is particularly relevant in cases of continued egg shedding post treatment as well as generally in kennels and breeding farms to prevent potential build-up of selected stages in the environment.
-
•
Selective treatment of only a fraction of the host population to avoid selection of resistance is not advisable for zoonotic canine/feline helminths: this is not a practical approach for pets since it would allow temporary excretion of potentially zoonotic stages in the immediate surroundings of humans.
-
•
Exclude that dogs with resistant parasites can roam freely until they have been tested negative for helminth at least twice.
Alt-text: Box 4
Box 5. Outstanding questions.
-
•
How can monitoring of drug susceptibility effectively be employed in the field?
-
•
Do the same mechanisms of AR as found in livestock parasitic nematodes occur in intestinal helminths (IH) of dogs and cats and if not what are potential mechanism of AR in these parasites?
-
•
What is the frequency of AR associated alleles in phenotypically susceptible populations of dog/cat IH?
-
•
How can modelling of AR evolution help to examine the effect of treatment frequency, refugium size or hygiene on the selection of AR in dog/cat IH?
-
•
Can the combination of drugs with different mode of action prevent/postpone AR development in dog/cat IH?
-
•
Which strategies can be developed to prevent or postpone the development of AR in IH of dogs/cats?
Alt-text: Box 5
The "good" often comes with potential unintended consequences, and thus, the hardest thing to do in much of the world is to assess how are we going to achieve the objectives outlined in Box 4, within a world with many different rules and attitudes. In the case of companion animals, it is important that AR mitigating procedures also take into account the requirements of veterinary public health. In the field, increased monitoring of drug efficacy in dogs/cats e.g. by annual FECRT or at least regular post treatment coproscopic testing is needed. The aim being to detect changes of the current situation at the earliest possible stage, and thus to enable appropriate preventive measures before resistant populations are spreading. This monitoring should be compulsory for those worm populations under highest risk for AR development, i.e. in kennels/catteries/shelters/racing dogs but also in other dogs post treatment check-ups are recommended. Coproscopic testing either using conventional methods such as flotation methods or recently marketed copro-antigen-tests (Elsemore et al., 2017) are easy to employ tools which should be more often used than is currently the case in the field. Additionally, improved and less laborious methods for drug susceptibility testing need to be developed to enable routine monitoring. Practical advice as to how to perform anthelmintic drug efficacy testing specifically for hookworm infections in US dogs has recently been provided by Jimenez Castro and Kaplan (2020). This can in principle be taken advantage of also in other geographical regions as well as for other IH species although some aspects which differ depending on the respective site (e.g. availability of drugs such as emodepside in Europe for dogs) need to be taken into consideration.
Not least and crucially, a key component in AR mitigation is hygiene, comprising removal and appropriate disposal of faeces. These basic measures, which are easier to accomplish in pets compared to grazing animals, not only reduce infection intensity and thus the effective helminth population size, but also remove the progeny of any treatment survivors.
Table 1.
Anthelmintic resistance, drug classes currently available and their mode of action, spectrum of efficacy and host availability.
| Drug class | Mechanism of action | Spectrum of efficacy | Hosts for which registered products are available | ARa situationb |
|---|---|---|---|---|
| Piperazines | Opening of chloride ion channels – paralysis | Intestinal nematodes | Human, livestock animals, horses | – |
| Benzimidazoles | Inhibition of microtubule polymerization – a.o. energy depletion | Nematodes, selected trematodes and cestodes | Human, pet and livestock animals | Livestock: ++++ |
| Dogs: + | ||||
| Cats: - | ||||
| Humans: - | ||||
| Imidazothiazoles | Agonists of nicotinergic acetyl choline-receptors – paralysis | Nematodes | Human, pet and livestock animals | Livestock: +++ |
| Dogs: - | ||||
| Cats: - | ||||
| Humans: - | ||||
| Tetrahydropyrimidines | Agonists of nicotinergic acetyl choline-receptors – paralysis | Nematodes | Human, pet and livestock animals | Livestock: ++++ |
| Dogs: + | ||||
| Cats: - | ||||
| Humans: - | ||||
| Macrocyclic lactones | Glutamate and GABA-gated chloride channels – paralysis | Nematodes, arthropods | Human, pet and livestock animals | Livestock: ++++ |
| Dogs: + | ||||
| Cats: - | ||||
| Humans: - | ||||
| Cyclooctadepsipeptides | Multiple mechanisms, a.o. calcium- and voltage-gated potassium channels – paralysis | Nematodes | Dogs, cats | – |
| Amino-acetonitril-derivatives | Allosteric activator of nicotinergic acetyl choline-receptor – paralysis | Nematodes | Small ruminants | Livestock: + |
| Dogs: - | ||||
| Cats: - | ||||
| Humans: - | ||||
| Spiroindoles | Antagonist at nicotinergic acetyl choline-receptor – paralysis | Nematodes | Small ruminants | + |
| Salicyanilides | Decoupling of oxidative phosphorylation – energy depletion | Haemonchus, Fasciola | Ruminants | Livestock: + |
| Dogs: - | ||||
| Cats: - | ||||
| Humans: - | ||||
| Pyrazinisoquinoles | Unresolved, tegument damage and inhibition of calcium permeability | Cestodes, selected trematodes | Human, pet and livestock animals | Livestock: |
| Dogs: (+) | ||||
| Cats: - | ||||
| Humans: - | ||||
| Benzenesulfonamides | Inhibtion of glycolysis – energy depletion | Fasciola | Livestock | – |
Anthelmintic resistance.
-no reports; (+) single report; + few isolated reports; ++ numerous reports; +++ widespread reports; ++++ widespread reports and multidrug resistance.
Declaration of competing interest
Manuela Schnyder, Peter Deplazes, Dwight Bowman and Georg von Samson-Himmelstjerna each declare that they repeatedly have acted as consultant for veterinary pharmaceutical and diagnostic companies and have previous and ongoing research collaborations with various veterinary pharmaceutical companies.
Jürgen Krücken declares that he has previous and ongoing research collaborations with various veterinary pharmaceutical companies.
Andrew Thompson declares that he has previous research collaborations with various veterinary pharmaceutical companies.
Warwick Grant declares no conflict of interest.
References
- Artho R., Schnyder M., Kohler L., Torgerson P., Hertzberg H. Avermectin-resistance in gastrointestinal nematodes of Boer goats and Dorper sheep in Switzerland. Vet. Parasitol. 2007;144:68–73. doi: 10.1016/j.vetpar.2006.09.032. [DOI] [PubMed] [Google Scholar]
- Bartram D.J., Leathwick D.M., Taylor M.A., Geurden T., Maeder S.J. The role of combination anthelmintic formulations in the sustainable control of sheep nematodes. Vet. Parasitol. 2012;186:151–158. doi: 10.1016/j.vetpar.2011.11.030. [DOI] [PubMed] [Google Scholar]
- Becker A.C., Rohen M., Epe C., Schnieder T. Prevalence of endoparasites in stray and fostered dogs and cats in Northern Germany. Parasitol. Res. 2012;111:849–857. doi: 10.1007/s00436-012-2909-7. [DOI] [PubMed] [Google Scholar]
- Beugnet F., Bourdeau P., Chalvet-Monfray K., Cozma V., Farkas R., Guillot J., Halos L., Joachim A., Losson B., Miró G., Otranto D., Renaud M., Rinaldi L. Parasites of domestic owned cats in Europe: co-infestations and risk factors. Parasites Vectors. 2014;7:291. doi: 10.1186/1756-3305-7-291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bindke J.D., Springer A., Janecek-Erfurth E., Böer M., Strube C. Helminth infections of wild European gray wolves (Canis lupus Linnaeus, 1758) in Lower Saxony, Germany, and comparison to captive wolves. Parasitol. Res. 2019;118:701–706. doi: 10.1007/s00436-018-6181-3. [DOI] [PubMed] [Google Scholar]
- Bowman D., Coles T., Eberhard M., Lighttowler M., Lynn R., Little S. tenth ed. 2014. Georgis ‘parasitology for Veterinarians; pp. 162–166. Sant Louis. [Google Scholar]
- Bridger K.E., Baggs E.M., Finney-Crawley J. Endoparasites of the coyote (Canis latrans), a recent migrant to insular newfoundland. J. Wildl. Dis. 2009;45:1221–1226. doi: 10.7589/0090-3558-45.4.1221. [DOI] [PubMed] [Google Scholar]
- Bružinskaitè-Schmidhalter R., Šarkūnas M., Malakausas A., Mathis A., Torgerson P.R., Deplazes P. Helminths of red foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) in Lithuania. Parasitology. 2012;139:120–127. doi: 10.1017/S0031182011001715. [DOI] [PubMed] [Google Scholar]
- Bryan H.M., Darimont C.T., Hill J.E., Paquet P.C., Thompson R.C., Wagner B., Smits J.E. Seasonal and biogeographical patterns of gastrointestinal parasites in large carnivores: wolves in a coastal archipelago. Parasitology. 2012;139:781–790. doi: 10.1017/S0031182011002319. [DOI] [PubMed] [Google Scholar]
- Chaudhry U., Redman E.M., Ashraf K., Shabbir M.Z., Rashid M.I., Ashraf S., Gilleard J.S. Microsatellite marker analysis of Haemonchus contortus populations from Pakistan suggests that frequent benzimidazole drug treatment does not result in a reduction of overall genetic diversity. Parasites Vectors. 2016;9:349. doi: 10.1186/s13071-016-1624-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaudhry U., Redman E.M., Raman M., Gilleard J.S. Genetic evidence for the spread of a benzimidazole resistance mutation across southern India from a single origin in the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2015;45:721–728. doi: 10.1016/j.ijpara.2015.04.007. [DOI] [PubMed] [Google Scholar]
- Chelladurai J.J., Kifleyohannes T., Scott J., Brewer M.T. Praziquantel resistance in the zoonotic cestode Dipylidium caninum. Am. J. Trop. Med. Hyg. 2018;99:1201–1205. doi: 10.4269/ajtmh.18-0533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornelius M., Jacobson C., Dobson R., Besier R. Computer modelling of anthelmintic resistance and worm control outcomes for refugia-based nematode control strategies in Merino ewes in Western Australia. Vet. Parasitol. 2016;220:59–66. doi: 10.1016/j.vetpar.2016.02.030. [DOI] [PubMed] [Google Scholar]
- Cotter J.L., Van Burgel A., Besier R.B. Anthelmintic resistance in nematodes of beef cattle in south-west Western Australia. Vet. Parasitol. 2015;207:276–284. doi: 10.1016/j.vetpar.2014.11.019. [DOI] [PubMed] [Google Scholar]
- Craig P., Hegglin D., Lightowlers M., Torgerson P.R., Wang Q. Echinococcosis: control and prevention. Adv. Parasitol. 2017;96:55–158. doi: 10.1016/bs.apar.2016.09.002. [DOI] [PubMed] [Google Scholar]
- Craig P., Larrieu E. Control of cystic echinococcosis/hydatidosis: 1863–2002. Adv. Parasitol. 2006;61:443–508. doi: 10.1016/S0065-308X(05)61011-1. [DOI] [PubMed] [Google Scholar]
- Dantas-Torres F., Ketzis J., Mihalca A.D., Baneth G., Otranto D., Tort G.P., Watanabe M., Linh B.K., Inpankaew T., Jimenez Castro P.D., Borrás P., Arumugam S., Penzhorn B.L., Ybañez A.P., Irwin P., Traub R.J. TroCCAP recommendations for the diagnosis, prevention and treatment of parasitic infections in dogs and cats in the tropics. Vet. Parasitol. 2020;283:109167. doi: 10.1016/j.vetpar.2020.109167. [DOI] [PubMed] [Google Scholar]
- Demeler J., Van Zeveren A.M., Kleinschmidt N., Vercruysse J., Höglund J., Koopmann R., Cabaret J., Claerebout E., Areskog M., von Samson-Himmelstjerna G. Monitoring the efficacy of ivermectin and albendazole against gastro intestinal nematodes of cattle in Northern Europe. Vet. Parasitol. 2009;160:109–115. doi: 10.1016/j.vetpar.2008.10.030. [DOI] [PubMed] [Google Scholar]
- Denver D.R., Dolan P.C., Wilhelm L.J., Sung W., Lucas-Lledó J.I., Howe D.K., Lewis S.C., Okamoto K., Thomas W.K., Lynch M. A genome-wide view of Caenorhabditis elegans base-substitution mutation processes. Proc. Natl. Acad. Sci. U.S.A. 2009;106:16310–16314. doi: 10.1073/pnas.0904895106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deplazes P., Eckert J., Mathis A., Samson-Himmelstjerna G.von, Zahner H. vol. 333. Wageningen Academic Press Publisheers; 2016. Phylum nematoda. (Parasitology in Veterinary Medicine). ISBN:978-0-90-8686-274. [Google Scholar]
- Deplazes P., van Knapen F., Schweiger A., Overgaauw P.A. Role of pet dogs and cats in the transmission of helminthic zoonoses in Europe, with a focus on echinococcosis and toxocarosis. Vet. Parasitol. 2011;182:41–53. doi: 10.1016/j.vetpar.2011.07.014. [DOI] [PubMed] [Google Scholar]
- Dobson R.J., Barnes E.H., Tyrrell K.L., Hosking B.C., Larsen J.W., Besier R.B., Love S., Rolfe P.F., Bailey J.N. A multi-species model to assess the effect of refugia on worm control and anthelmintic resistance in sheep grazing systems. Aust. Vet. J. 2011;89:200–208. doi: 10.1111/j.1751-0813.2011.00719.x. [DOI] [PubMed] [Google Scholar]
- Doyle S.R., Bourguinat C., Nana-Djeunga H.C., Kengne-Ouafo J.A., Pion S.D.S., Bopda J., Kamgno J., Wanji S., Che H., Kuesel A.C., Walker M., Basáñez M.G., Boakye D.A., Osei-Atweneboana M.Y., Boussinesq M., Prichard R.K., Grant W.N. Genome-wide analysis of ivermectin response by Onchocerca volvulus reveals that genetic drift and soft selective sweeps contribute to loss of drug sensitivity. PLoS Neglected Trop. Dis. 2017;11 doi: 10.1371/journal.pntd.0005816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drake J., Carey T. Seasonality and changing prevalence of common canine gastrointestinal nematodes in the USA. Parasites Vectors. 2019;12:430. doi: 10.1186/s13071-019-3701-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drogemuller M., Failing K., Schnieder T., von Samson-Himmelstjerna G. Effect of repeated benzimidazole treatments with increasing dosages on the phenotype of resistance and the beta-tubulin codon 200 genotype distribution in a benzimidazole-resistant cyathostomin population. Vet. Parasitol. 2004;123:201–213. doi: 10.1016/j.vetpar.2004.06.019. [DOI] [PubMed] [Google Scholar]
- Dyachenko V., Pantchev N., Gawlowska S., Vrhovec M.G., Bauer C. Echinococcus multilocularis infections in domestic dogs and cats from Germany and other European countries. Vet. Parasitol. 2008;157:244–253. doi: 10.1016/j.vetpar.2008.07.030. [DOI] [PubMed] [Google Scholar]
- Elsemore D.A., Geng J., Cote J., Hanna R., Lucio-Forster A., Bowman D.D. Enzyme-linked immunosorbent assays for coproantigen detection of Ancylostoma caninum and Toxocara canis in dogs and Toxocara cati in cats. J. Vet. Diagn. Invest. 2017;29:645–653. doi: 10.1177/1040638717706098. [DOI] [PubMed] [Google Scholar]
- Fairweather I., Brennan G.P., Hanna R.E.B., Robinson M.W., Skuce P.J. Drug resistance in liver flukes. Int J Parasitol Drugs Drug Resist. 2020;12:39–59. doi: 10.1016/j.ijpddr.2019.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falzon L.C., O'Neill T.J., Menzies P.I., Peregrine A.S., Jones-Bitton A., vanLeeuwen J., Mederos A. A systematic review and meta-analysis of factors associated with anthelmintic resistance in sheep. Prev. Vet. Med. 2014;117:388–402. doi: 10.1016/j.prevetmed.2014.07.003. [DOI] [PubMed] [Google Scholar]
- Fisher M.A., Jacobs D.E., Hutchinson M.J., Dick I.G. Studies on the control of Toxocara canis in breeding kennels. Vet. Parasitol. 1994;55:87–92. doi: 10.1016/0304-4017(94)90058-2. [DOI] [PubMed] [Google Scholar]
- Fritzen B., Rohn K., Schnieder T., von Samson-Himmelstjerna G. Endoparasite control management on horse farms--lessons from worm prevalence and questionnaire data. Equine Vet. J. 2010;42:79–83. doi: 10.2746/042516409X471485. [DOI] [PubMed] [Google Scholar]
- Gasbarre L.C., Smith L.L., Hoberg E., Pilitt P.A. Further characterization of a cattle nematode population with demonstrated resistance to current anthelmintics. Vet. Parasitol. 2009;166:275–280. doi: 10.1016/j.vetpar.2009.08.019. [DOI] [PubMed] [Google Scholar]
- Geary T.G., Hosking B.C., Skuce P.J., von Samson-Himmelstjerna G., Maeder S., Holdsworth P., Pomroy W., Vercruysse J. World Association for the Advancement of Veterinary Parasitology (WAAVP) Guideline: anthelmintic combination products targeting nematode infections of ruminants and horses. Vet. Parasitol. 2012;190:306–316. [Google Scholar]
- Gemmell M., Johnstone P., Oudemans G. The effect of route of administration on the efficacy of praziquantel against Echinococcus granulosus infections in dogs. Res. Vet. Sci. 1980;29:131–132. [PubMed] [Google Scholar]
- Gilleard J., Redman E. Genetic diversity and population structure of Haemonchus contortus. Adv. Parasitol. 2016;93:31–68. doi: 10.1016/bs.apar.2016.02.009. [DOI] [PubMed] [Google Scholar]
- Gilleard J.S. Haemonchus contortus as a paradigm and model to study anthelmintic drug resistance. Parasitology. 2013;140:1506–1522. doi: 10.1017/S0031182013001145. [DOI] [PubMed] [Google Scholar]
- Hanna R.E., McMahon C., Ellison S., Edgar H.W., Kajugu P.E., Gordon A., Irwin D., Barley J.P., Malone F.E., Brennan G.P., Fairweather I. Fasciola hepatica: a comparative survey of adult fluke resistance to triclabendazole, nitroxynil and closantel on selected upland and lowland sheep farms in Northern Ireland using faecal egg counting, coproantigen ELISA testing and fluke histology. Vet. Parasitol. 2015;207:34–43. doi: 10.1016/j.vetpar.2014.11.016. [DOI] [PubMed] [Google Scholar]
- Hegglin D., Deplazes P. Control of Echinococcus multilocularis: strategies, feasibility and cost–benefit analyses. Int. J. Parasitol. 2013;43:327–337. doi: 10.1016/j.ijpara.2012.11.013. [DOI] [PubMed] [Google Scholar]
- Jackson R., Lance D., Townsend K., Stewart K. Isolation of anthelmintic resistant Ancylostoma caninum. N. Z. Vet. J. 1987;35:215–216. doi: 10.1080/00480169./1987.35456. [DOI] [PubMed] [Google Scholar]
- Jenkins D.J. Does the presence of Spirometra erinacei reduce the efficacy of praziquantel against Echinococcus granulosus in dogs? Int. J. Parasitol. 1998;28:1943. doi: 10.1016/s0020-7519(98)00158-1. [DOI] [PubMed] [Google Scholar]
- Jimenez Castro P.D., Kaplan R. Persistent or suspected-resistant hookworm infections. Clin. Brief. 2020:61–68. https://www.cliniciansbrief.com/article/persistent-or-suspected-resistant-hookworm-infections August. [Google Scholar]
- Jimenez Castro P.D., Howell S.B., Schaefer J.J., Avramenko R.W., Gilleard J.S., Kaplan R.M. Multiple drug resistance in the canine hookworm Ancylostoma caninum: an emerging threat? Parasites Vectors. 2019;12:576. doi: 10.1186/s13071-019-3828-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jimenez Castro P.D., Mansour A., Charles S., Hostetler J., Settje T., Kulke D., Kaplan R.M. Efficacy evaluation of anthelmintic products against an infection with the canine hookworm (Ancylostoma caninum) isolate Worthy 4.1F3P in dogs. Int. J. Parasitol. Drugs Drug Resist. 2020;13:22–27. doi: 10.1016/j.ijpddr.2020.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan R.M., Vidyashankar A.N. An inconvenient truth: global worming and anthelmintic resistance. Vet. Parasitol. 2012;186:70–78. doi: 10.1016/j.vetpar.2011.11.048. [DOI] [PubMed] [Google Scholar]
- Kenyon F., Jackson F. Targeted flock/herd and individual ruminant treatment approaches. Vet. Parasitol. 2012;186:10–17. doi: 10.1016/j.vetpar.2011.11.041. [DOI] [PubMed] [Google Scholar]
- Kitchen S., Ratnappan R., Han S., Leasure C., Grill E., Iqbal Z., Granger O., O'Halloran D.M., Hawdon J.M. Isolation and characterization of a naturally occurring multidrug-resistant strain of the canine hookworm, Ancylostoma caninum. Int. J. Parasitol. 2019;49:397–406. doi: 10.1016/j.ijpara.2018.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitchen S., Ratnappan R., Han S., Leasure C., Grill E., Iqbal Z., Granger O., O'Halloran D.M., Hawdon J.M. Isolation and characterization of a naturally occurring multidrug-resistant strain of the canine hookworm, Ancylostoma caninum. Int. J. Parasitol. 2019;49:397–406. doi: 10.1016/j.ijpara.2018.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knapp-Lawitzke F., Krücken J., Ramünke S., von Samson-Himmelstjerna G., Demeler J. Rapid selection for β-tubulin alleles in codon 200 conferring benzimidazole resistance in an Ostertagia ostertagi isolate on pasture. Vet. Parasitol. 2015;209:84–92. doi: 10.1016/j.vetpar.2015.02.005. [DOI] [PubMed] [Google Scholar]
- Kopp S.R., Coleman G.T., McCarthy J.S., Kotze A.C. Application of in vitro anthelmintic sensitivity assays to canine parasitology: detecting resistance to pyrantel in Ancylostoma caninum. Vet. Parasitol. 2008;152:284–293. doi: 10.1016/j.vetpar.2007.12.020. [DOI] [PubMed] [Google Scholar]
- Kopp S.R., Kotze A.C., McCarthy J.S., Coleman G.T. High-level pyrantel resistance in the hookworm Ancylostoma caninum. Vet. Parasitol. 2007;143:299–304. doi: 10.1016/j.vetpar.2006.08.036. [DOI] [PubMed] [Google Scholar]
- Learmount J., Taylor M.A., Bartram D.J. A computer simulation study to evaluate resistance development with a derquantel-abamectin combination on UK sheep farms. Vet. Parasitol. 2012;187:244–253. doi: 10.1016/j.vetpar.2011.12.033. [DOI] [PubMed] [Google Scholar]
- Leathwick D.M., Sauermann C.W., Nielsen M.K. Managing anthelmintic resistance in cyathostomin parasites: investigating the benefits of refugia-based strategies. Int. J. Parasitol. Drugs Drug. Resist. 2019;10:118–124. doi: 10.1016/j.ijpddr.2019.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leathwick D.M., Sauermann C.W., Geurden T., Nielsen M.K. Managing anthelmintic resistance in Parascaris spp.: a modelling exercise. Vet. Parasitol. 2017;240:75–81. doi: 10.1016/j.vetpar.2017.03.026. [DOI] [PubMed] [Google Scholar]
- Little S.E., Johnson E.M., Lewis D., Jaklitsch R.P., Payton M.E., Blagburn B.L., Bowman D.D., Moroff S., Tams T., Rich L., Aucoin D. Prevalence of intestinal parasites in pet dogs in the United States. Vet. Parasitol. 2009;166:144–152. doi: 10.1016/j.vetpar.2009.07.044. [DOI] [PubMed] [Google Scholar]
- Lyons E.T., Tolliver S.C., Collins S.S. Field studies on endoparasites of thoroughbred foals on seven farms in central Kentucky in 2004. Parasitol. Res. 2006;98:496–500. doi: 10.1007/s00436-005-0091-x. [DOI] [PubMed] [Google Scholar]
- Martin P.J., Anderson N., Jarrett R.G. Detecting benzimidazole resistance with faecal egg count reduction tests and in vitro assays. Aust. Vet. J. 1989;66:236–240. doi: 10.1111/j.1751-0813.1989.tb13578.x. [DOI] [PubMed] [Google Scholar]
- Martin P.J., Le Jambre L.F., Claxton J.H. The impact of refugia on the development of thiabendazole resistance in Haemonchus contortus. Int. J. Parasitol. 1981;11:35–41. doi: 10.1016/0020-7519(81)90023-0. [DOI] [PubMed] [Google Scholar]
- McCall J.W., Genchi C., Kramer L.H., Guerrero J., Venco L. Heartworm disease in animals and humans. Adv. Parasitol. 2008;66:193–285. doi: 10.1016/S0065-308X(08)00204-2. [DOI] [PubMed] [Google Scholar]
- Mehlhorn H., Hanser E., Harder A., Hansen O., Mencke N., Schaper R. Synergistic effects of pyrantel and the febantel metabolite fenbendazole on adult Toxocara canis. Parasitol. Res. 2003;90:S151–S153. doi: 10.1007/s00436-003-0923-5. [DOI] [PubMed] [Google Scholar]
- Miró G., Gálvez R., Montoya A., Delgado B., Drake J. Survey of Spanish pet owners about endoparasite infection risk and deworming frequencies. Parasites Vectors. 2020;13:101. doi: 10.1186/s13071-020-3976-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miró G., Mateo M., Montoya A., Vela E., Calonge R. Survey of intestinal parasites in stray dogs in the Madrid area and comparison of the efficacy of three anthelmintics in naturally infected dogs. Parasitol. Res. 2007;100:317–320. doi: 10.1007/s00436-006-0258-0. [DOI] [PubMed] [Google Scholar]
- Nielsen M.K. Evidence-based considerations for control of Parascaris spp. infections in horses. Equine Vet. Educ. 2016;28:224–231. [Google Scholar]
- Nijsse R., Mughini-Gras L., Wagenaar J.A., Franssen F., Ploeger H.W. Environmental contamination with Toxocara eggs: a quantitative approach to estimate the relative contributions of dogs, cats and foxes, and to assess the efficacy of advised interventions in dogs. Parasites Vectors. 2015;8:397. doi: 10.1186/s13071-015-1009-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noack S., Harrington J., Carithers D.S., Kaminsky R., Selzer P.M. Heartworm disease - overview, intervention, and industry perspective. Int. J. Parasitol. Drugs Drug Resist. 2021;16:65–89. doi: 10.1016/j.ijpddr.2021.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer C.S., Robertson I.D., Traub R.J., Rees R., Thompson R.C. Intestinal parasites of dogs and cats in Australia: the veterinarian's perspective and pet owner awareness. Vet. J. 2010;183:358–361. doi: 10.1016/j.tvjl.2008.12.007. [DOI] [PubMed] [Google Scholar]
- Redman E., Whitelaw F., Tait A., Burgess C., Bartley Y., Skuce P.J., Jackson F., Gilleard J.S. The emergence of resistance to the benzimidazole anthlemintics in parasitic nematodes of livestock is characterised by multiple independent hard and soft selective sweeps. PLoS Neglected Trop. Dis. 2015;9 doi: 10.1371/journal.pntd.0003494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redman W.K., Bryant J.E., Ahmad G. Gastrointestinal helminths of coyotes (Canis latrans) from Southeast Nebraska and Shenandoah area of Iowa. Vet. World. 2016;9:970–975. doi: 10.14202/vetworld.2016.970-975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reperant L.A., Hegglin D., Fischer C., Kohler L., Weber J.M., Deplazes P. Influence of urbanization on the epidemiology of intestinal helminths of the red fox (Vulpes vulpes) in Geneva, Switzerland. Parasitol. Res. 2007;101:605–611. doi: 10.1007/s00436-007-0520-0. [DOI] [PubMed] [Google Scholar]
- Reynoldson J.A., Behnke J.M., Pallant L.J., Macnish M.G., Gilbert F., Giles S., Spargo R., Thompson R.A. Failure of pyrantel in treatment of human hookworm infections (Ancylostoma duodenale) in the Kimberley region of north west Australia. Acta Trop. 1997;68:301–312. doi: 10.1016/s0001-706x(97)00106-x. [DOI] [PubMed] [Google Scholar]
- Ridley R.K., Dryden M.W., Gabbert N.H., Schoning P. Epidemiology and control of helminth parasites in Greyhound breeding farms. Compend. Contin. Educ. Vet. 1994;6:585–599. [Google Scholar]
- Romig T., Deplazes P., Jenkins D., Giraudoux P., Massolo A., Craig P.S., Wassermann M., Takahashi K., De La Rue M. Ecology and life cycle patterns of Echinococcus species. Adv. Parasitol. 2017;95:213–314. doi: 10.1016/bs.apar.2016.11.002. [DOI] [PubMed] [Google Scholar]
- Rose H., Rinaldi L., Bosco A., Mavrot F., de Waal T., Skuce P., Charlier J., Torgerson P.R., Hertzberg H., Hendrickx G., Vercruysse J., Morgan E.R. Widespread anthelmintic resistance in European farmed ruminants: a systematic review. Vet. Rec. 2015;176:546. doi: 10.1136/vr.102982. [DOI] [PubMed] [Google Scholar]
- Rose H., Rinaldi L., Bosco A., Mavrot F., de Waal T., Skuce P., Charlier J., Torgerson P.R., Hertzberg H., Hendrickx G., Vercruysse J., Morgan E.R. Widespread anthelmintic resistance in European farmed ruminants: a systematic review. Vet. Rec. 2015;176:546. doi: 10.1136/vr.102982. [DOI] [PubMed] [Google Scholar]
- Rose Vineer H., Morgan E.R., Hertzberg H., Bartley D.J., Bosco A., Charlier J., Chartier C., Claerebout E., de Waal T., Hendrickx G., Hinney B., Höglund J., Ježek J., Kašný M., Keane O.M., Martínez-Valladares M., Mateus T.L., McIntyre J., Mickiewicz M., Munoz A.M., Phythian C.J., Ploeger H.W., Rataj A.V., Skuce P.J., Simin S., Sotiraki S., Spinu M., Stuen S., Thamsborg S.M., Vadlejch J., Varady M., von Samson-Himmelstjerna G., Rinaldi L. Increasing importance of anthelmintic resistance in European livestock: creation and meta-analysis of an open database. Parasite. 2020;27:69. doi: 10.1051/parasite/2020062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saeed I., Maddox-Hyttel C., Monrad J., Kapel C.M. Helminths of red foxes (Vulpes vulpes) in Denmark. Vet. Parasitol. 2006;139:168–179. doi: 10.1016/j.vetpar.2006.02.015. [DOI] [PubMed] [Google Scholar]
- Sanchez J., Dharmarajan G., George M.M., Pulaski C., Wolstenholme A.J., Gilleard J.S., Kaplan R.M. Using population genetics to examine relationships of Dirofilaria immitis based on both macrocyclic lactone-resistance status and geography. Vet. Parasitol. 2020;283:109–125. doi: 10.1016/j.vetpar.2020.109125. [DOI] [PubMed] [Google Scholar]
- Sauermann C.W., Nielsen M.K., Luo D., Leathwick D.M. Modelling the development of anthelmintic resistance in cyathostomin parasites: the importance of genetic and fitness parameters. Vet. Parasitol. 2019;269:28–33. doi: 10.1016/j.vetpar.2019.04.007. [DOI] [PubMed] [Google Scholar]
- Schnieder T., Laabs E.M., Welz C. Larval development of Toxocara canis in dogs. Vet. Parasitol. 2011;175:193–206. doi: 10.1016/j.vetpar.2010.10.027. [DOI] [PubMed] [Google Scholar]
- Silvestre A., Cabaret J., Humbert J.F. Effect of benzimidazole under-dosing on the resistant allele frequency in Teladorsagia circumcincta (Nematoda) Parasitology. 2001;123:103–111. doi: 10.1017/s0031182001008009. [DOI] [PubMed] [Google Scholar]
- Siyadatpanah A., Gholami S., Daryani A., Sarvi S., Sharif M., Seguel M., Boundenga L., Amouei A., Pagheh A.S., Rahimi M.T., Hosseini S.A., Anvari D. The prevalence of intestinal helminths in free-ranging canids of Mazandaran, northern Iran. Iran. J. Parasitol. 2019;14:563–571. [PMC free article] [PubMed] [Google Scholar]
- Sowemimo O., Asaolu S. The daily egg production of Ancylostoma caninum and the distribution of the worm along the digestive tract of the dog. J. Parasitol. 2008;3:92–97. [Google Scholar]
- Stafford K., Kollasch T.M., Duncan K.T., Horr S., Goddu T., Heinz-Loomer C., Rumschlag A.J., Ryan W.G., Sweet S., Little S.E. Detection of gastrointestinal parasitism at recreational canine sites in the USA: the DOGPARCS study. Parasites Vectors. 2020;13:275. doi: 10.1186/s13071-020-04147-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strube C., Neubert A., Springer A., von Samson-Himmelstjerna G. Survey of German pet owners quantifying endoparasitic infection risk and implications for deworming recommendations. Parasites Vectors. 2019;12:203. doi: 10.1186/s13071-019-3410-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szafrańska E., Wasielewski O., Bereszyński A. A faecal analysis of helminth infections in wild and captive wolves, Canis lupus L., in Poland. J. Helminthol. 2010;84:415–419. doi: 10.1017/S0022149X10000106. [DOI] [PubMed] [Google Scholar]
- Thompson R.C., Roberts M.G. Does pet helminth prophylaxis increase the rate of selection for drug resistance? Trends Parasitol. 2001;17:576–578. doi: 10.1016/s1471-4922(01)02084-0. [DOI] [PubMed] [Google Scholar]
- Van Wyk J.A. Refugia-overlooked as perhaps the most potent factor concerning the development of anthelmintic resistance. Onderstepoort J. Vet. Res. 2001;68:55–67. [PubMed] [Google Scholar]
- Vienažindienė Ž., Joekel D.E., Schaper R., Deplazes P., Šarkūnas M. Longitudinal study for anthelmintic efficacy against intestinal helminths in naturally exposed Lithuanian village dogs: critical analysis of feasibility and limitations. Parasitol. Res. 2018;117:1581–1590. doi: 10.1007/s00436-018-5843-5. [DOI] [PubMed] [Google Scholar]
- von Samson-Himmelstjerna G. Anthelmintic resistance in equine parasites–detection, potential clinical relevance and implications for control. Vet. Parasitol. 2012;185:2–8. doi: 10.1016/j.vetpar.2011.10.010. [DOI] [PubMed] [Google Scholar]
- Wapenaar W., Barkema H.W., O'Handley R. Fecal shedding of Toxocara canis and other parasites in foxes and coyotes on Prince Edward Island, Canada. J. Wildl. Dis. 2013;49:394–397. doi: 10.7589/2012-04-113. [DOI] [PubMed] [Google Scholar]
- Wolstenholme A.J., Fairweather I., Prichard R., von Samson-Himmelstjerna G., Sangster N.C. Drug resistance in veterinary helminths. Trends Parasitol. 2004;20:469–476. doi: 10.1016/j.pt.2004.07.010. [DOI] [PubMed] [Google Scholar]