

Abstract
The placenta of the Korean water deer was anatomically examined to accumulate basic information regarding its reproductive system. The convex placentomes with five to nine well-developed pedicles were observed in the whole uterine horns, and therefore, the placenta was classified as oligocotyledonary. The evidence indicating the migration of binucleate cells (BNCs) from trophectoderm to the uterine epithelium led to the histological classification of the placenta as synepitheliochorial. The number of fetuses was markedly higher than that in other ruminant species. However, the number of placentomes was found to be similar to the other Cervidae species. Therefore, these results suggest that the Korean water deer may possess special mechanisms or structures at the fetus attachment site to maintain this unusally high number of fetuses.
Keywords: fetus, Korean water deer, placentome
The placenta, which dominates the space between the maternal and fetal organs in eutherian mammals, functions as an immunomodulatory organ that protects the fetus from the maternal immune system and facilitates the exchange of gases and nutrients during the fetal development [5, 8]. Furthermore, this organ secretes several placenta-specific hormones (e.g., chorionic gonadotropin, interferon-τ, growth hormone, and prolactin) to support pregnancy [5, 8, 33].
The placenta can be classified into four main categories based on its shape and histological structure, including diffuse, cotyledonary or placentomal, zonary, and discoid. This classification is based on the distribution of the chorionic folds or villi in the endometrium of the uterus [32, 47]. The histological structure of the placenta is classified into three categories based on the number of tissue layers between the fetal and maternal blood vessels, including epitheliochorial, endotheliochorial, and hemochorial [9]. Especially in ruminants, the trophoblast binucleate cells (BNCs) are released form synepitheliochorial, a subtype of epitheliochorial placenta [29], via the cell fusion process, as mentioned later.
BNCs play an important role in the placenta of ruminants [44, 49]. For instance, in sheep and goats, BNCs are located in the fetal trophectoderm and transform the uterine epithelium to fetomaternal syncytium after migrating through the tight junctions [38, 39, 44, 48]. In fetomaternal syncytium, the placental lactogen and pregnancy-specific protein B from BNCs are mainly secreated close to the maternal blood vessels [44, 50]. Conversely, in cows and deer, the trinucleate cells produced by BNCs are reabsorbed by the trophectoderm after the secretion [45, 48, 52]. The fusion of BNCs and the uterine epithelium is essential for facilitating the transfer of fetal hormones and proteins to the maternal circulation [44,45,46, 49, 51, 53].
The suborder Ruminantia is one of the most successful mammals owing to its development and diversity in dentition, multichambered stomach, skull appendages, and locomotion [6, 42]. These marked characteristic differences observed among species have attracted the attention towards morphological studies of these species. There are still many species among the ruminants, including the water deer, for which the detailed structure of the placenta has not been examined yet. In this study, the detailed morphological structure of the uteroplacenta of the Korean water deer (Hydropotes inermis argyropus) was macroscopically and histologically examined. The genus Hydropotes, belonging to the subfamily Hydropotinae, and family Cervidae, is distributed across South Korea and the Northeastern China [7, 13], and it exhibits unique characteristics, including the presence of well-developed canines in the maxilla and absence of antlers [12]. Although the ecological [10, 11, 54] and genetic [21, 27] studies on this species have been conducted previously, there are limited studies on the anatomical examination of the Korean water deer, which include the male reproductive organ [36, 37], branching pattern of the aortic arch [1], rumen structure [19], and morphometric studies of the skull [22,23,24, 31].
Therefore, in this study, the uteroplacenta of the Korean water deer was corroborated through macroscopic and histological observations. This information may act as a prerequisite for the conservation and management of this species.
Ten uteri from the pregnant Korean water deer, who did not survive traffic accidents, were used in this study. The uteri were fixed using 10% formalin and stored until further observation. The number and size of the placentomes and fetuses were measured, and the crown-rump length (CRL) of the fetuses present in each uterus was measured. The volume of the placentome was evaluated, assuming that each placentome was an ellipsoid type, using the following formula:
where, a and b correspond to the minor and major axis of the placentome, respectively.
For histological examination, multiple placentomes were randomly harvested from each uterus and re-fixed using Bouin solution for 1–2 days before paraffin embedding. A paraffin block was cut into four µm thickness sections, following which, hematoxylin and eosin (H&E), periodic acid–Schiff (PAS), and Azan stainings were performed. After PAS staining, the cytoplasm of BNCs was slightly reddish. Using this as an index, the distribution and cell density of BNCs were evaluated visually using a microscope (Biozero, Keyence, Osaka, Japan). The tissue areas on the vertical sections of the placentomes were divided into the deep, intermediate, and surface layer starting from the center of the maternal organ (Fig. 2B), and three examination fields were randomly selected from each layer. The villi were evenly divided into one-third region from the base to the tip of each villus, which was based on the evidence that BNCs tend to localize in the deep layer of placentomes in Sika deer (unpublished data).
Fig. 2.

The vertical section of the placentome from specimen 007 stained using Azan staining (A), and its schematic diagram (B). A pedicle (arrow) extending from the maternal organ of the placentome to the uterus, and the villi (arrowheads) were observed. Scale bar=5 mm.
The uterus of the Korean water deer was bicornuate, which existed with partial fusion in the common uterine body (Fig. 1A). About one to four fetuses were observed in the uteroplacenta (Table 1), while five–nine placentomes were found in the whole uterine horns (Table 1), with volumes ranging from 8.5 to 56.2 cm3. The characteristics of all the fetuses and placentomes are presented in Table 1. The placenta of the suborder Ruminantia, except infraorder Tragulina, can be classified as cotyledonary as it possesses a placentome formed of fetal villi (cotyledon) and maternal crypts (caruncle) [25]. The placentome assists in establishing the maternofetal vascular systems, which is an essential process for the maternal system to support the growth of the fetus throughout the gestation period [34, 35]. The number of placentomes among ruminants varies between three and 150, and the placenta are classified in two different types, including polycotyledonary (cow: 70–142, sheep: 88–96, goat and giraffe: 160–180, impala: 40–50, and Japanese serow: 100), and oligocotyledonary (red deer, roe deer, and fallow deer: eight, and Sika deer, and reindeer: six) [16, 30]. In this study, the placenta of the Korean water deer was classified as oligocotyledonary, similar to other Cervidae families with six–eight placentomes [16]. The shape of the placentomes of this species was convex, which was revealed using both macroscopic and microscopic observations (Fig. 2). Additionally, the villi were well-developed and exhibited clear pedicles in each placentome. The placentome of ruminants can be classified based on the combination of the shape and branching conditions of the villi. The flat-shaped placentome (pronghorn [4, 30], musk deer [4], kob [3, 4, 20], and greater kudu [4, 20]) possesses unbranched or branched villi. The convex-shaped placentome can be classified based on unbranched or lightly-branched villi (fallow deer [17, 49], red deer [16, 50], giraffe [41]), and well-branched villi (reindeer [3, 16], cattle [3, 26, 28, 45]). Moreover, the concave-shaped placentome (goats [3, 45], sheep [3, 4, 28, 45], and impala [40]) have been shown to possess well-branched villi.
Fig. 1.

The fixed uteroplacenta of the Korean water deer (specimen 003). A: The external view of the partially fused (arrow) bicornuate uterus and palpable swollen pregnant uterine horns (asterisks) that are distinguishable. B: The internal view of the placentomes is indicated through asterisks. Here, three fetuses were confirmed.
Table 1. Size and number of fetuses and placentomes in Korean water deer.
| Specimen ID | CRL (mm) Average (min–max) |
Volume of placentome (cm3) Average (min–max) |
No. of placentomes (Left/Right uterine horn) |
No. of fetuses (Left/Right uterine horn) |
|---|---|---|---|---|
| 1 | 13.6 (13.6–13.6) | NA | NA | 3 (3/0) |
| 2 | 92.2 (91.5–92.8) | 8.5 (5.2–16.7) | 5 (2/3) | 2 (1/1) |
| 3 | 94.2 (92.1–97.0) | 17.1 (11.9–22.9) | 5 (2/3) | 3 (1/2) |
| 4 | 104.1 (98.4–112.2) | 21.6 (11.9–28.2) | 5 (3/2) | 3 (1/2) |
| 5 | 111.4 (104.0–116.4) | 28.9 (3.6–47.5) | 7 (4/3) | 4 (2/2) |
| 6 | 114.5 (113.6–115.4) | 12.5 (8.5–18.3) | 6 (3/3) | 2 (1/1) |
| 7 | 123.6 (112.0–129.6) | 23.4 (2.7–57.0) | 9 (4/5) | 3 (2/1) |
| 8 | 142.4 (142.4–142.4) | 25.4 (13.5–36.0) | 5 (2/3) | 1 (1/0) |
| 9 | 174.9 (162.8–194.6) | 56.2 (7.4–82.0) | 7 (4/3) | 3 (1/2) |
| 10 | 183.7 (183.7–183.7) | 28.2 (22.2–32.7) | 6 (3/3) | 1 (0/1) |
CRL, crown rump length; NA, not available.
The branching condition of the fetal villi in the convex-shaped placentome is known to differ among the Cervidae family [2, 16]. Although the fetal villi of the subfamily Cervidae are unbranched or branched, those of the reindeer are extensively branched [16]. The roe deer placentome exhibits intermediate villi branching [16]. In this study, the Korean water deer was found to exhibit a convex-shaped placentome and well-branched villi, similar to that in the reindeer.
The number of fetuses in the uterus of the Korean water deer ranged between one and four as shown in this study (Table 1). According to Gilbert et al. [14], Hassanin et al. [18], and Gustafson [15], the family Cervidae can be divided into the subfamily Cervinae, consisting of two tribes (Muntiacini and Cervini), and the subfamily Capreolinae, consisting of three tribes (Capreolini, Alceini, and Odocoileini). The number of fetuses in the subfamilies Cervinae and Caprepolinae is usually one, with an exception in the tribe Capreolini, which includes the water deer and roe deer [3, 16]. Although the number of fetuses is similar between the roe deer and the Korean water deer, the branching of the villi in the placentome of the roe deer was not as developed as in the case of the Korean water deer [16]. The number of fetuses among the members of the Cervidae family is similar based on the classifications by Gilbert et al. [14] and Hassanin et al. [18]; however, the placentome shape and branching of the villi did not demonstrate a strong correlation among ruminant species, including the Cervidae family.
The shape of the placentome and pedicle, which is located inside the placentome through the histological examination using Azan staining (Fig. 2A). BNCs were observed using PAS staining on both sides of the uterine and trophectodermal epithelium (Fig. 3), which indicated that the histological structure of the placenta was synepitheliochorial. The number of BNCs was higher on the fetal side than on the maternal side (Fig. 4). The localization of BNCs on the maternal side confirmed the migration of BNCs from the trophectoderm to the uterine epithelium. The reindeer is similar to the Korean water deer as it exhibits a well-developed placentome and pedicle, well-branched villi, and the existence of BNCs in the trophectoderm [16, 43]. Conversely, the number of fetuses in the roe deer is similar to that of the Korean water deer; however, the branching of the villi is not as developed as in the water deer [16].
Fig. 3.
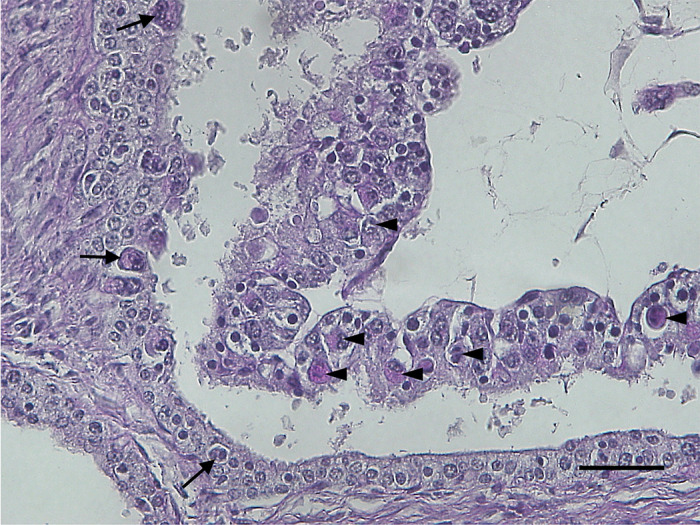
The placentome from specimen 007 stained using periodic acid–Schiff (PAS). Binucleated cells (BNCs) in the trophectoderm (arrowheads) and uterine epithelium (arrows) were observed. Scale bar=50 µm.
Fig. 4.

The density of binucleate cells (BNCs) in the placentome. The density of fetal BNCs was higher than that of maternal BNCs in all specimens.
To the best of our knowledge, this is the first study presenting the detailed structure of the placentome of the Korean water deer. The Korean water deer possesses an average of six convex-shaped placentomes with well-developed pedicles and villi. The placenta of the Korean water deer can be classified as oligocotyledonary and synepitheliochorial, which is similar to other Cervidae subfamilies. The number of fetuses was markedly higher than that in other species of Cervidae family. Although the number of placentomes is similar to other species of Cervidae family, the Korean water deer might possess special mechanisms or structures in the attachment site of the fetus to maintain the unusally high number of fetuses. Therefore, the unknown underlying mechanisms need to be elucidated in the future studies.
CONFLICT OF INTEREST
The authors declare that there are no conflicts of interest.
REFERENCES
- 1.Ahn D. C., Kim H. C., Tae H. J., Kang H. S., Kim N. S., Park S. Y., Kim I. S.2008. Branching pattern of aortic arch in the Korean water deer. J. Vet. Med. Sci. 70: 1051–1055. doi: 10.1292/jvms.70.1051 [DOI] [PubMed] [Google Scholar]
- 2.Amoroso E. C., Parkes A. S.1952. Marshall’s Physiology of Reproduction. pp. 127–311. (Park, A. C. ed.), Longmans Green, London. [Google Scholar]
- 3.Andresen A.1927. Die plazentome der wiederkäuer. Morphol. Jahrb. 57: 410–485. [Google Scholar]
- 4.Benirschke K.2006. Comparative placentation, http://placentation.ucs d.edu/ [accessed on May 1, 2020].
- 5.Carter A. M.2012. Evolution of placental function in mammals: the molecular basis of gas and nutrient transfer, hormone secretion, and immune responses. Physiol. Rev. 92: 1543–1576. doi: 10.1152/physrev.00040.2011 [DOI] [PubMed] [Google Scholar]
- 6.Chen L., Qiu Q., Jiang Y., Wang K., Lin Z., Li Z., Bibi F., Yang Y., Wang J., Nie W., Su W., Liu G., Li Q., Fu W., Pan X., Liu C., Yang J., Zhang C., Yin Y., Wang Y., Zhao Y., Zhang C., Wang Z., Qin Y., Liu W., Wang B., Ren Y., Zhang R., Zeng Y., da Fonseca R. R., Wei B., Li R., Wan W., Zhao R., Zhu W., Wang Y., Duan S., Gao Y., Zhang Y. E., Chen C., Hvilsom C., Epps C. W., Chemnick L. G., Dong Y., Mirarab S., Siegismund H. R., Ryder O. A., Gilbert M. T. P., Lewin H. A., Zhang G., Heller R., Wang W.2019. Large-scale ruminant genome sequencing provides insights into their evolution and distinct traits. Science 364: eaav6202. doi: 10.1126/science.aav6202 [DOI] [PubMed] [Google Scholar]
- 7.Cooke A. S., Farrell L.1998. Chinese Water Deer, Mammal Society and The British Deer Society, London and Fordingbridge, Hampshire. [Google Scholar]
- 8.Cross J. C., Baczyk D., Dobric N., Hemberger M., Hughes M., Simmons D. G., Yamamoto H., Kingdom J. C. P.2003. Genes, development and evolution of the placenta. Placenta 24: 123–130. doi: 10.1053/plac.2002.0887 [DOI] [PubMed] [Google Scholar]
- 9.Dantzer V., Leiser R.2006. Placentation. pp. 279–297. In: Dellmann’s Textbook of Veterinary Histology, 6th ed. (Eurell, J. A. and Frappier, B. L. eds.), Blackwell Publishing, Oxford. [Google Scholar]
- 10.Dubost G., Charron F., Courcoul A., Rodier A.2008. Population characteristics of a semi-free-ranging polytocous cervid, Hydropotes inermis. Mammalia 72: 333–343. doi: 10.1515/MAMM.2008.050 [DOI] [Google Scholar]
- 11.Dubost G., Charron F., Courcoul A., Rodier A.2011. Social organization in the Chinese water deer, Hydropotes inermis. Acta Theriol. (Warsz.) 56: 189–198. doi: 10.1007/s13364-010-0008-7 [DOI] [Google Scholar]
- 12.Dubost G., Charron F., Courcoul A., Rodier A.2011. The Chinese water deer, Hydropotes inermis-A fast-growing and productive ruminant. Mamm. Biol. 76: 190–195. doi: 10.1016/j.mambio.2010.04.001 [DOI] [Google Scholar]
- 13.Geist V.1998. Deer of the World: Their Evolution, Behaviour, and Ecology, Stackpole Books, Mechanicsburg. [Google Scholar]
- 14.Gilbert C., Ropiquet A., Hassanin A.2006. Mitochondrial and nuclear phylogenies of Cervidae (Mammalia, Ruminantia): Systematics, morphology, and biogeography. Mol. Phylogenet. Evol. 40: 101–117. doi: 10.1016/j.ympev.2006.02.017 [DOI] [PubMed] [Google Scholar]
- 15.Gustafson E. P.2015. An early pliocene north american deer: bretzia pseudalces, its osteology, biology, and place in cervid history. Bull. Museum Nat. Hist. Univ. Oregon. 25: 1–75. [Google Scholar]
- 16.Hamilton W. J., Harrison R. J., Young B. A.1960. Aspects of placentation in certain cervidae. J. Anat. 94: 1–33. [PMC free article] [PubMed] [Google Scholar]
- 17.Harrison R. J., Hyett A. R.1954. The development and growth of the placentomes in the fallow deer (Dama dama L.). J. Anat. 88: 338–355. [PMC free article] [PubMed] [Google Scholar]
- 18.Hassanin A., Delsuc F., Ropiquet A., Hammer C., Jansen van Vuuren B., Matthee C., Ruiz-Garcia M., Catzeflis F., Areskoug V., Nguyen T. T., Couloux A.2012. Pattern and timing of diversification of Cetartiodactyla (Mammalia, Laurasiatheria), as revealed by a comprehensive analysis of mitochondrial genomes. C. R. Biol. 335: 32–50. doi: 10.1016/j.crvi.2011.11.002 [DOI] [PubMed] [Google Scholar]
- 19.Hofmann R. R., Kock R. A., Ludwig J., Axmacher H.1988. Seasonal changes in rumen papillary development and body condition in free ranging Chinese water deer (Hydropotes inermis). J. Zool. (Lond.) 216: 103–117. doi: 10.1111/j.1469-7998.1988.tb02418.x [DOI] [Google Scholar]
- 20.Hradecký P., Mossman H. W., Stott G. G.1988. Comparative development of ruminant placentomes. Theriogenology 29: 715–729. doi: 10.1016/S0093-691X(88)80016-5 [DOI] [PubMed] [Google Scholar]
- 21.Hu J., Fang S. G., Wan Q. H.2006. Genetic diversity of Chinese water deer (Hydropotes inermis inermis): implications for conservation. Biochem. Genet. 44: 161–172. doi: 10.1007/s10528-006-9020-7 [DOI] [PubMed] [Google Scholar]
- 22.Kim Y. K., Koyabu D., Lee H., Kimura J.2013. Sexual dimorphism of craniomandibular size in the Korean water deer, Hydropotes inermis argyropus. J. Vet. Med. Sci. 75: 1153–1159. doi: 10.1292/jvms.13-0125 [DOI] [PubMed] [Google Scholar]
- 23.Kim Y. K., Koyabu D., Lee H., Kimura J.2015. Cranial morphological homogeneity in two subspecies of water deer in China and Korea. J. Vet. Med. Sci. 77: 1427–1435. doi: 10.1292/jvms.15-0037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kim Y., Suzuki S., Na K. J., Lee H., Kimura J.2013. Skull growth of the Korean water deer, Hydropotes inermis argyropus. J. Vet. Med. Sci. 75: 867–878. doi: 10.1292/jvms.13-0019 [DOI] [PubMed] [Google Scholar]
- 25.Kimura J., Sasaki M., Endo H., Fukuta K.2004. Anatomical and histological characterization of the female reproductive organs of mouse deer (Tragulidae). Placenta 25: 705–711. doi: 10.1016/j.placenta.2004.02.009 [DOI] [PubMed] [Google Scholar]
- 26.Klisch K., Pfarrer C., Schuler G., Hoffmann B., Leiser R.1999. Tripolar acytokinetic mitosis and formation of feto-maternal syncytia in the bovine placentome: different modes of the generation of multinuclear cells. Anat. Embryol. (Berl.) 200: 229–237. doi: 10.1007/s004290050275 [DOI] [PubMed] [Google Scholar]
- 27.Koh H. S., Lee B. K., Wang J., Heo S. W., Jang K. H.2009. Two sympatric phylogroups of the Chinese water deer (Hydropotes inermis) identified by mitochondrial DNA control region and cytochrome b gene analyses. Biochem. Genet. 47: 860–867. doi: 10.1007/s10528-009-9285-8 [DOI] [PubMed] [Google Scholar]
- 28.Leiser R., Krebs C., Klisch K., Ebert B., Dantzer V., Schuler G., Hoffmann B.1997. Fetal villosity and microvasculature of the bovine placentome in the second half of gestation. J. Anat. 191: 517–527. doi: 10.1046/j.1469-7580.1997.19140517.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.MacGeady T. A., Quinn P. J., Fitzpatrick E. S., Ryan M. T., Kilroy D., Lonergan P.2017. Forms of implantation and placentation. pp. 92–111. In: Veterinary Embryology, 2nd ed., John Wiley & Sons, West Sussex. [Google Scholar]
- 30.Mossman H. W.1987. Vertebrate Fetal Membranes, Rutgers University Press, New Brunswick. [Google Scholar]
- 31.Oh J., Kim Y. K., Yasuda M., Koyabu D., Kimura J.2017. Cranial suture closure pattern in water deer and implications of suture evolution in cervids. Mamm. Biol. 86: 17–20. doi: 10.1016/j.mambio.2017.03.004 [DOI] [Google Scholar]
- 32.Ramsey E. M.1982. The Placenta: Human and Animal, Praeger, New York. [Google Scholar]
- 33.Rawn S. M., Cross J. C.2008. The evolution, regulation, and function of placenta-specific genes. Annu. Rev. Cell Dev. Biol. 24: 159–181. doi: 10.1146/annurev.cellbio.24.110707.175418 [DOI] [PubMed] [Google Scholar]
- 34.Reynolds L. P., Redmer D. A.1995. Utero-placental vascular development and placental function. J. Anim. Sci. 73: 1839–1851. doi: 10.2527/1995.7361839x [DOI] [PubMed] [Google Scholar]
- 35.Reynolds L. P., Redmer D. A.2001. Angiogenesis in the placenta. Biol. Reprod. 64: 1033–1040. doi: 10.1095/biolreprod64.4.1033 [DOI] [PubMed] [Google Scholar]
- 36.Sohn J. H., Kimura J.2012. Observation of male reproductive organ in Korean water deer (Hydropotes inermis argyropus). Asian J. Anim. Vet. Adv. 7: 30–37. doi: 10.3923/ajava.2012.30.37 [DOI] [Google Scholar]
- 37.Sohn J., Sasaki M., Yasuda M., Kim Y., Shin N. S., Kimura J.2013. Immunolocalization of cytoskeletal proteins in the testes of two Asian cervids: water deer (Hydropotes inermis) and Reeves’ muntjac (Muntiacus reevesi). J. Vet. Med. Sci. 75: 1071–1075. doi: 10.1292/jvms.13-0079 [DOI] [PubMed] [Google Scholar]
- 38.Wango E. O., Wooding F. B., Heap R. B.1990. The role of trophoblastic binucleate cells in implantation in the goat: a morphological study. J. Anat. 171: 241–257. [PMC free article] [PubMed] [Google Scholar]
- 39.Wango E. O., Wooding F. B. P., Heap R. B.1990. The role of trophoblast binucleate cells in implantation in the goat: a quantitative study. Placenta 11: 381–394. doi: 10.1016/S0143-4004(05)80214-0 [DOI] [PubMed] [Google Scholar]
- 40.Wilsher S., Greenwood R. E. S., Mahon G. D., Allen W. R.2020. Placentation and hormonal maintenance of pregnancy in the impala (Aepyceros melampus). Placenta 95: 91–105. doi: 10.1016/j.placenta.2020.04.009 [DOI] [PubMed] [Google Scholar]
- 41.Wilsher S., Stansfield F., Greenwood R. E. S., Trethowan P. D., Anderson R. A., Wooding F. B. W., Allen W. R.2013. Ovarian and placental morphology and endocrine functions in the pregnant giraffe (Giraffa camelopardalis). Reproduction 145: 541–554. doi: 10.1530/REP-13-0060 [DOI] [PubMed] [Google Scholar]
- 42.Wilson D. E., Reeder D. M.2005. Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed., John Hopkins University Press, Baltimore. [Google Scholar]
- 43.Wimsatt W. A.1951. Observations on the morphogenesis, cytochemistry, and significance of the binocleate giant cells of the placenta of ruminants. Am. J. Anat. 89: 233–281. doi: 10.1002/aja.1000890204 [DOI] [PubMed] [Google Scholar]
- 44.Wooding F. B. P.1984. Role of binucleate cells in fetomaternal cell fusion at implantation in the sheep. Am. J. Anat. 170: 233–250. doi: 10.1002/aja.1001700208 [DOI] [PubMed] [Google Scholar]
- 45.Wooding F. B. P.1992. Current topic: the synepitheliochorial placenta of ruminants: binucleate cell fusions and hormone production. Placenta 13: 101–113. doi: 10.1016/0143-4004(92)90025-O [DOI] [PubMed] [Google Scholar]
- 46.Wooding F. B. P., Beckers J. F.1987. Trinucleate cells and the ultrastructural localisation of bovine placental lactogen. Cell Tissue Res. 247: 667–673. doi: 10.1007/BF00215761 [DOI] [PubMed] [Google Scholar]
- 47.Wooding F. B. P., Burton G.2008. Comparative Placentation: Structures, Functions and Evolution, Springer-Verlag, Berlin. [Google Scholar]
- 48.Wooding F. B. P., Wathes D. C.1980. Binucleate cell migration in the bovine placentome. J. Reprod. Fertil. 59: 425–430. doi: 10.1530/jrf.0.0590425 [DOI] [PubMed] [Google Scholar]
- 49.Wooding F. B. P., Hobbs T., Morgan G., Heap R. B., Flint A. P. F.1993. Cellular dynamics of growth in sheep and goat synepitheliochorial placentomes: an autoradiographic study. J. Reprod. Fertil. 98: 275–283. doi: 10.1530/jrf.0.0980275 [DOI] [PubMed] [Google Scholar]
- 50.Wooding F. B. P., Morgan G., Adam C. L.1997. Structure and function in the ruminant synepitheliochorial placenta: central role of the trophoblast binucleate cell in deer. Microsc. Res. Tech. 38: 88–99. doi: [DOI] [PubMed] [Google Scholar]
- 51.Wooding F. B. P., Morgan G., Forsyth I. A., Butcher G., Hutchings A., Billingsley S. A., Gluckman P. D.1992. Light and electron microscopic studies of cellular localization of oPL with monoclonal and polyclonal antibodies. J. Histochem. Cytochem. 40: 1001–1009. doi: 10.1177/40.7.1607634 [DOI] [PubMed] [Google Scholar]
- 52.Wooding F. B. P., Osborn D., Killian G. J.2018. Trinucleate uterine epithelial cells as evidence for White-tail Deer trophoblast binucleate cell migration and as markers of placental binucleate cell dynamics in a variety of wild ruminants. Placenta 62: 34–42. doi: 10.1016/j.placenta.2017.12.012 [DOI] [PubMed] [Google Scholar]
- 53.Wooding F. B. P., Roberts R. M., Green J. A.2005. Light and electron microscope immunocytochemical studies of the distribution of pregnancy associated glycoproteins (PAGs) throughout pregnancy in the cow: possible functional implications. Placenta 26: 807–827. doi: 10.1016/j.placenta.2004.10.014 [DOI] [PubMed] [Google Scholar]
- 54.Zhu H., Qin P., Wang H.2004. Functional group classification and target species selection for Yancheng Nature Reserve, China. Biodivers. Conserv. 13: 1335–1353. doi: 10.1023/B:BIOC.0000019401.52391.a9 [DOI] [Google Scholar]