

Abstract
Objective
To determine the temporal expression of angiotensin-converting enzyme 2 (ACE2), a receptor for SARS-CoV-2, in dominant follicles throughout the periovulatory period in women and the regulatory mechanisms underlying ACE2 expression in human granulosa/lutein cells (hGLC).
Design
Experimental prospective clinical study and laboratory-based investigation.
Setting
University Medical Center and private in vitro fertilization center.
Patient(s)
Thirty premenopausal women undergoing surgery for tubal ligation and 16 premenopausal women undergoing in vitro fertilization.
Intervention(s)
Administration of human chorionic gonadotropin (hCG) and harvesting of preovulatory/ovulatory follicles by timed laparoscopy, and collection of granulosa/lutein cells and cumulus cells at the time of oocyte retrieval.
Main Outcome Measure(s)
Expression and localization of ACE2 in granulosa cells and dominant follicles collected throughout the periovulatory period of the menstrual cycle and in hGLC using quantitative polymerase chain reaction, immunoblotting, and immunohistochemistry.
Result(s)
ACE2 expression (mRNA and protein) is up-regulated in human ovulatory follicles after administration of hCG. ACE2 expression was higher in cumulus cells than in granulosa cells. hCG increased the expression of ACE2 in primary hGLC cultures; the increase was inhibited by RU486 (an antagonist for progesterone receptor and glucocorticoid receptor) and CORT125281 (a selective glucocorticoid receptor antagonist), but not by AG1478 (an EGF receptor tyrosine kinase inhibitor) or by dexamethasone.
Conclusion(s)
The hormone-regulated expression of ACE2 in granulosa cells suggests a potential role of ACE2 in the ovulatory process. These data also imply the possible impact of COVID-19 on a vital cyclic event of ovarian function and thus on women's overall reproductive health. However, SAR-CoV-2 infection in ovarian cells in vivo or in vitro has yet to be determined.
Key Words: ACE2, follicle, humans, ovary, ovulation
Abstract
Sobre-regulación ovulatoria de la enzima convertidora de angiotensina 2, un receptor para SARS-CoV-2, en folículos dominantes del ovario humano.
Objetivo
Determinar la expresión temporal de la enzima convertidora de angiotensina 2 (ACE2), un receptor para SARS-CoV-2, en los folículos dominantes de mujeres a través del periodo periovulatorio y los mecanismos regulatorios subyacentes de la expresión de ACE2 en células de la granulosa/luteínicas humanas (hGLC).
Diseño
Estudio clínico prospectivo experimental e investigación en laboratorio.
Lugar
Centro Médico Universitario y centro de fertilización in vitro privado.
Paciente(s)
30 mujeres premenopaúsicas sometidas a cirugía para ligadura tubárica y 16 mujeres premenopaúsicas sometidas a fecundación in vitro.
Intervencion(es)
Administración de gonadotropina coriónica humana (hCG) y aspiración de folículos preovulatorios y ovulatorios mediante programación con laparoscopia y recolección de células de granulosa/teca y células del cumulus al momento de la aspiración ovocitaria.
Resultado(s) principal(es) medible(s)
Expresión y localización de ACE2 en células de la granulosa y en folículos dominantes recolectados a través del periodo periovulatorio del ciclo menstrual y en hGLC usando reacción de la cadena de polimerasa cuantitativa, inmunoblotting e inmunohistoquímica.
Resultado(s)
La expresión de ACE2 (ARNm y proteína) está sobre-regulada en los folículos ovulatorios humanos después de la administración de hCG. La expresión de ACE2 fue mayor en células del cúmulus que en células de la granulosa. La hCG aumentó la expresión de ACE2 en los cultivos primarios de hGLC; el incremento fue inhibido por RU486 (antagonista para los receptores de progesterona y glucocorticoides) y por CORT125281 (antagonista selectivo del receptor de glucocorticoides), pero no por el AG1478 (inhibidor tirosina quinasa del receptor EGF) ni por la dexametasona.
Conclusion(es)
La regulación hormonal de la expresión de ACE2 en las células de la granulosa sugieren un rol potencial de ACE2 en el proceso ovulatorio. Estos datos también implican el posible impacto de la COVID-19 en un evento cíclico vital de la función ovárica y también en la salud reproductiva de las mujeres. Sin embargo, no se ha determinado aún la presencia de infección por SAR-CoV-2 en células ováricas in vivo o in vitro.
DIALOG: You can discuss this article with its authors and other readers at https://www.fertstertdialog.com/posts/33000
Since the beginning of the COVID-19 pandemic, heightened attention has been given to angiotensin-converting enzyme 2 (ACE2), since it acts as an entry receptor for SARS-CoV-2, a novel strain of the virus that causes COVID-19 (1). However, this role of ACE2 in viral cellular entry is in contrast to its original reported function as a component of the renin-angiotensin system (RAS).
Initially, ACE2 was described as a zinc metalloprotease that is capable of catalyzing the conversion of angiotensin I (Ang I) and angiotensin II (Ang II) to angiotensin-(1–9) and angiotensin-(1–7), respectively (2, 3). ACE2 displays high homology to human angiotensin-converting enzyme (ACE) (2, 4). ACE is well known for its ability to convert inactive angiotensin (Ang I) into the functionally active hormone (Ang II) (5). The best-elucidated actions of Ang II include vasoconstriction, stimulation of aldosterone and vasopressin secretion, vascular smooth muscle proliferation, and renal tubular sodium reuptake, thereby regulating body fluid volume, electrolyte balance, and arterial pressure (6, 7). These effects of Ang II are tightly controlled by ACE2, whose function is to limit the locally available Ang II concentration by rapidly hydrolyzing Ang II to Ang-(1–7) (8). Ang-(1–7) has also been shown to enhance vasodilation and exert antioxidative, antiinflammatory, and proresolution effects, likely through activation of the G protein-coupled receptor MAS1 (MAS1 proto-oncogene) (9). Therefore, ACE2 functions as a key counter-regulatory enzyme in the RAS. In addition to its role in the RAS, ACE2 can cleave other peptide substrates, such as neurotensin, kinetensin, des-Arg bradykinin, and apelin-13, -17, and -36 (2, 10). Besides these enzymatic functions, ACE2 was identified as a binding site for human coronavirus (HCoV-NL63) and the human severe acute respiratory syndrome coronaviruses, SARS-CoV and SARS-CoV-2 (11). The spike proteins on the surface of SARS coronaviruses bind to ACE2 on its target cells, initiating virus–cell membrane fusion and ultimately resulting in viral replication inside the host cells (11).
ACE2 mRNA is detected in most human tissues examined, yet the detection of ACE2 protein is limited to several tissues, including the lung, kidney, heart, and intestine (12). Specifically, abundant ACE2 expression is localized to lung alveolar epithelial cells, enterocytes of the small intestine, and vascular endothelial cells of the heart and kidney (13). The testis is also among the tissues that express high levels of ACE2 (mRNA and protein) (14). In contrast, little is known about the expression pattern of ACE2 in the human ovary. Two public databases for protein expression profiles (the Human Protein Atlas and GeneCards) contradict each other; the latter suggests a high expression of ACE2, whereas the former indicates little or no expression. A limited number of studies documented the expression of ACE2 in the ovary of animals (15, 16). Pereira et al. (16) reported that ACE2 expression was increased in the theca/interstitial layer of the immature rat ovary when eCG was administered to stimulate follicle development. In the bovine ovary, the follicular levels of ACE2 mRNA are transiently down-regulated during the ovulatory period (15). As a proxy indicator of ACE2 expression, Ang II and Ang-(1–7) were reported to be present in the ovaries of multiple species (15, 16, 17, 18). Previous studies showed that Ang II and Ang-(1–7) stimulated steroid hormone production and enhanced ovulation and oocyte maturation in rats, rabbits, and cattle (17, 19, 20, 21, 22). Similarly, Ang-(1–7) was detected in the follicular fluid of women undergoing in vitro fertilization (IVF); the level of this heptapeptide was correlated clinically with the oocyte maturation rate (23).
The ovary undergoes constant remodeling and cyclic ovulation that require precisely controlled angiogenesis and an acute inflammatory response (reviewed in Duffy et al. [24]). Given that ACE2 has been shown to play an important role in both angiogenesis and inflammatory responses, it is conceivable that this enzyme is expressed and involved in these processes in the human ovary. Moreover, accumulating evidence indicates that SARS-CoV-2 can infect multiple organs, including the ovary (25), but the long-term health impact of COVID-19 has yet to be determined. Therefore, it is critically important to assess the expression of ACE2 in the human ovary. In the present study, we sought to characterize the expression pattern of ACE2 (mRNA and protein) using dominant follicles collected before the luteinizing hormone (LH) surge and throughout the periovulatory period from naturally cycling women with ovulation induced by human chorionic gonadotropin (hCG), and to analyze the cellular regulatory mechanisms controlling ACE2 expression using primary human granulosa/lutein cells (hGLC).
Materials and methods
Materials
Unless otherwise noted, all reagents were purchased from MilliporeSigma (St. Louis, MO) or Thermo Fisher Scientific (Waltham, MA). CORT125281 was kindly provided by Corcept Therapeutics (Menlo Park, CA).
Human Tissue Collection
The protocol using human tissues was approved by the Human Ethics Committee of the Sahlgrenska Academy at the University of Gothenburg, and all patients gave informed written consent before participating. Whole follicles were collected from patients throughout the periovulatory period, as previously described (26). Women aged 30 to 38 years with regular menstrual cycles who had not used hormonal contraceptives for at least 3 months before enrollment in the study underwent laparoscopic sterilization. The women were monitored by transvaginal ultrasound for two or three menstrual cycles before surgery to ascertain cycle regularity and to monitor the growth of a dominant follicle during the follicular phase. These patients were divided into four groups: preovulatory, early ovulatory, late ovulatory, and postovulatory. In the preovulatory group, surgery was performed when the follicle reached >14 and ≤17.5 mm in diameter before the endogenous LH surge. These patients were not given hCG.
The remaining women were given recombinant hCG (Ovitrelle, 250 μg) and were divided into three groups: early ovulatory (surgery 12 to ≤18 hours post-hCG), ovulatory (surgery between >18 to ≤34 hours post-hCG), and post ovulatory phase (surgery >44 to ≤70 hours post-hCG). To confirm that these patients followed a normal hormonal pattern before the LH surge or after hCG administration, blood samples were taken at the time of surgery and serum progesterone and estradiol levels were measured (27). The whole intact follicle/early corpus luteum was removed with laparoscopic scissors and processed for either immunohistochemical or gene expression analysis. The follicle was bisected, and mural granulosa cells were gently scraped off from the interior of the follicle with small tissue forceps. For the gene expression study, the follicular fluid and cell suspension were combined and centrifuged at 500g to pellet and collect granulosa cells. For the immunohistochemistry study, the dominant follicles were fixed as described below.
Human Granulosa/Lutein Cell Cultures
Human granulosa/lutein cells (hGLC) were obtained from aspirates of patients undergoing IVF. The collection protocol was approved by the Institutional Review Board of the University of Kentucky Office of Research Integrity. Ovarian hyperstimulation was induced by the administration of recombinant human follicle-stimulating hormone in individualized doses to patients at the Bluegrass Fertility Center (Lexington, Kentucky). The IVF patients were then given hCG (10,000 U) on days 9 to 11 when one lead follicle reached 18 mm in diameter or two lead follicles reached 16 mm in diameter, and the dominant follicles were aspirated 36 hours later. The experiments with hGLC were performed as described previously (26, 28). Briefly, immediately after retrieval of the cumulus oocyte complexes, the remaining cells in the aspirates were subjected to Percoll gradient centrifugation to remove red blood cells. Cells from patients aged 24 to 40 years with nonovarian etiologies (e.g., male factor and egg donor) were used. The isolated cells were first examined under the microscope for their morphology, counted, and resuspended in OptiMEM medium supplemented with 10% fetal bovine serum and antibiotic-antimycotic and then seeded onto culture plates (2.5 × 105 cells/mL). The cells were acclimatized for 6 days, and the medium was changed every 24 hours. At the end of acclimation, the hGLC were treated with or without hCG (1 IU/mL) in the absence or presence of various reagents described in the Results section and cultured for the stated number of hours in OptiMEM medium supplemented with antibiotics–antimycotics.
Immunohistochemistry
The follicles were fixed in 4% formaldehyde, embedded in paraffin, and sectioned at 7 μm. Immunostaining was performed in the Markey Biospecimen Procurement and Translational Pathology Shared Resource Facility at the University of Kentucky, as previously described (28). Briefly, heat-induced epitope retrieval was performed in a Biocare Medical Decloaking chamber using Dako's low pH Target Retrieval Solution. Primary antibody incubation was performed at 4oC overnight for ACE2 (MilliporeSigma, HPA000288, 1:200 dilution) and for 2 hours at room temperature for platelet and endothelial cell adhesion molecule 1 (PECAM1) (Roche Diagnostics, Nederlands JC70 monoclonal antibody, predilute), respectively. Rabbit IgG was used in place of primary antibodies as a negative control. Antibody staining was detected with an appropriate Immpress alkaline phosphatase kit and Vector Red AP chromogen (Vector Laboratories, Burlingame, CA).
Gene Expression Analysis
The levels of mRNA for ACE2 were measured by a previously described method (28). Briefly, total RNA was isolated from granulosa cells using an RNeasy mini kit (Qiagen, Germantown, MD). The synthesis of the first-strand cDNA was performed by reverse transcription of 500 ng of total RNA using superscript III. The levels of mRNA for the genes examined were measured by quantitative polymerase chain reaction (qPCR) using Brilliant 3 Ultra-Fast SYBR green (Stratagene, Santa Clara, CA). Oligonucleotide primers corresponding to ACE2 were designed using Primer3 software (5'-GGTGGGAGATGAAGCGAGAG-3', 5'-ACATGGAACAGAGATGCGGG-3'). The relative abundance of the target transcript was normalized to internal reference genes (Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) for in vivo sample and RNA, 18S ribosomal 5 (RNA18S5) for in vitro sample) as previously described (26, 28) and calculated according to the 2-ΔΔCT method (29).
Western Blot Analysis
Nuclear extracts were isolated from cultured cells, denatured, run on a 10% polyacrylamide gel, and then transferred onto a nitrocellulose membrane as described previously (28). The membrane was incubated overnight at 4°C in 5% skim milk in Tris-buffered saline including 0.1% Tween-20 solution containing primary antibodies against ACE2 (MilliporeSigma, HPA000288, 1:100 dilution) and ACTB (Santa Cruz Biotechnology, Dallas, TX, sc-47778, 1:1,000 dilution). The blots were incubated with the respective secondary HRP-conjugated antibody for 1 hour at room temperature. Peroxidase activity was visualized using the SuperSignal West Pico Chemiluminescent Substrate.
Statistical Analyses
All data are presented as means ± SEM. Data were tested for homogeneity of variance by Levene's test, and log transformations were performed as appropriate. The paired sample t-test or analysis of variance was used to test differences between cell types in levels of mRNA for ACE2, or throughout the time of tissue collection or time of culture, or among treatments in vitro, as appropriate. If the test revealed significant effects, the means were compared by Duncan's test, with P<.05 considered to indicate statistical significance.
Results
Expression of ACE2 in Dominant Follicles During the Ovulatory Period
To determine whether ACE2 is expressed and regulated in human preovulatory follicles during the ovulatory period, the levels of mRNA for ACE2 were assessed in granulosa cells isolated from dominant follicles collected during the preovulatory, early ovulatory, late ovulatory, and postovulatory phases. The levels of mRNA for ACE2 were markedly increased in granulosa cells during the early and late ovulatory phases (>40-fold) compared with those obtained before hCG administration (Fig. 1 ). After ovulation, the levels of ACE2 mRNA in granulosa cells of postovulatory follicles were similar to those of dominant follicles collected during the early and late ovulatory phases, but higher than those of dominant follicles collected before hCG administration (Fig. 1A). Next, to determine whether ACE2 is also expressed in cumulus cells of ovulatory follicles, we collected granulosa cells and cumulus cells from women undergoing IVF procedures at the time of oocyte retrieval. As shown in Figure 1B, we found abundant expression of ACE2 in cumulus cells of ovulatory follicles. The levels of mRNA and protein for ACE2 were higher in cumulus cells than in granulosa cells from all patients tested.
Figure 1.

Levels of mRNA for angiotensin-converting enzyme 2 (ACE2) in granulosa cells (GCs) and cumulus cells of periovulatory follicles. (A) Dominant follicles were retrieved from the ovaries of women undergoing laparoscopic tubal sterilization before the luteinizing hormone surge or at various times after recombinant human chorionic gonadotropin administration and divided into four phases: preovulatory (Pre, n = 6), early ovulatory (EO, n = 5), late ovulatory (LO, n = 6), and postovulatory (PO, n = 2). The levels of mRNA for ACE2 were measured by quantitative polymerase chain reaction (qPCR) in granulosa cells isolated from a dominant follicle collected at Pre, EO, and LO and whole follicles retrieved at PO and normalized to the levels of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA in each sample. The levels are presented as fold change to Pre values. Bars with no common superscripts are significantly different (P<.05). (B) Cumulus cells and granulosa cells were collected at the time of oocyte retrieval from women undergoing a standardized in vitro fertilization procedure. a) The levels of mRNA for ACE2 were measured by qPCR and normalized to the levels of RNA, 18S Ribosomal 5 (RNA18S5) in each sample (n = 5 independent samples). ∗P<.05. b) A representative Western blot image detecting ACE2 protein. The samples loaded were from indicated independent patients. ACTB detection in each lane was used as a protein loading control.
To determine whether the increase in ACE2 mRNA levels translated to ACE2 protein and what types of cells expressed ACE2, the dominant follicles collected throughout the periovulatory phases were analyzed by immunohistochemistry. Staining for ACE2 was negligible in the granulosa cells and theca cell layer of dominant follicles obtained before the endogenous LH surge (Fig. 2 A). In dominant follicles obtained during the early ovulatory phase, positive staining for ACE2 became evident in the granulosa cells and theca cell layer (Fig. 2B). During the late ovulatory period, intensive staining of ACE2 was localized to both the granulosa cells and theca layer of dominant follicles (Fig. 2C and D). Interestingly, some cells in the granulosa and theca layer showed more robust staining (arrows) compared with the rest of the cells in late ovulatory follicles. This concentrated immunopositive staining of ACE2 was more prominent in the late ovulatory follicles that showed the morphologic changes typical of follicles immediately before ovulation (e.g., dispersed granulosa cells, dissolution of the basement membrane, and thinning of the theca interna layer) (Fig. 2D). In addition to the granulosa and theca cell layers, intense staining of ACE2 was detected throughout the stroma layer of late ovulatory follicles (Fig. 2D, wavy arrow). After ovulation, staining of ACE2 was persistent in both the granulosa and theca lutein cells of the postovulatory follicle as well as in the stroma layer, showing intense staining in some, but not all, follicular cells (arrows and wavy arrows in Fig. 2E and F).
Figure 2.

Expression pattern of angiotensin-converting enzyme 2 (ACE2) protein in periovulatory follicles. Dominant follicles were retrieved from the ovaries of women undergoing laparoscopic tubal sterilization before the luteinizing hormone surge or at various times after recombinant human chorionic gonadotropin administration. The paraffin-embedded sections of dominant follicles obtained at various time points (A, preovulatory phase, n = 2; B, early ovulatory phase, n = 4; C and D, late ovulatory phase, n = 4; E, late ovulatory phase, n = 1) were subjected to immunohistochemical analyses to detect ACE2 protein. The sections in C and D are from different patients obtained from the late ovulatory period. F is an antrum region of the postovulatory follicle shown in panel E. Pink/red staining indicates positive signals for ACE2. All sections were lightly stained with hematoxylin (blue staining) for nuclear counterstaining. Arrows point to granulosa and theca cells where intense staining for ACE2 was detected. Wavy arrows point to ACE2 staining in the stroma layer. GC = granulosa cells; GLC = granulosa lutein cells; RBC = red blood cells; TC = theca cells; TLC = theca lutein cells; Scale bar in A = 100 μm.
In the ovary, endothelial cells and leukocytes infiltrate into the granulosa and theca cell layer of ovulating follicles during transformation of the follicle into the corpus luteum (30). With the current finding that a subpopulation of cells in the granulosa and theca cell layer exhibited intense staining of ACE2, we questioned whether ACE2 was expressed in infiltrating endothelial cells and leukocytes. Therefore, serial sections of late ovulatory and postovulatory follicles used to detect ACE2 protein were evaluated for PECAM1, which was known to be expressed on the surface of endothelial cells, platelets, monocytes, neutrophils, and some types of T cells (31). As expected, immunopositive staining for PECAM1 was localized to endothelial cells lining various types of blood vessels and leukocytes inside blood vessels surrounding the ovulatory and postovulatory follicles (Fig. 3A and E). PECAM1 staining was also detected in endothelial cells located between the granulosa and theca interna layer throughout the late ovulatory follicle (Fig. 3B, arrows) and the theca lutein layer of the postovulatory follicle (Fig. 3F, arrows). However, the endothelial cell and leukocyte expression of PECAM1 in the ovulatory and postovulatory follicles was not colocalized to the distribution of ACE2 (the arrows in Fig. 3F and H point to the same location).
Figure 3.

Comparison of the expression pattern between platelet and endothelial cell adhesion molecule 1 (PECAM1) and angiotensin-converting enzyme 2 (ACE2) in late ovulatory and postovulatory follicles. Dominant follicles were retrieved from the ovaries of women undergoing laparoscopic tubal sterilization. The serial sections of dominant follicles obtained during the late ovulatory (A and B) and postovulatory (C and D) phases were subjected to immunohistochemical analyses to detect PECAM1 and ACE2. The square bars in A, C, E, and G are amplified in B, D, F, and H, respectively. PECAM1 was used to detect endothelial cells; it also stained a subpopulation of leukocytes. Asterisks in F and H were used to locate the same structure in the serial sections of the same postovulatory follicle. Pink/red staining indicates positive signals for PECAM1 and ACE2. All sections were lightly stained with hematoxylin (blue staining) for nuclear counterstaining. Arrows (B, D, F, H) and arrowheads (E) point to endothelial cells and leukocytes stained with PECAM1, respectively. GC = granulosa cells; GLC = granulosa lutein cells; TC = theca cells; TLC = theca lutein cells. Scale bar in A = 100 μm.
Regulation of ACE2 Expression in Human Granulosa Cells
To investigate the cellular mechanisms underlying the marked upregulation of ACE2 expression in periovulatory follicles after hCG administration in vivo, we used primary hGLC that were acclimated in cultures for 6 days without any hormone treatments to regain hCG responsiveness. As shown in Figure 4 A, we found that hCG treatment increased the expression of ACE2 at the levels of both mRNA and protein compared with that in control samples, similar to that observed during the periovulatory period of the menstrual cycle.
Figure 4.

Regulation of angiotensin-converting enzyme 2 (ACE2) expression in human granulosa cells. Human granulosa/lutein cells (hGLC) obtained from women undergoing a standardized in vitro fertilization procedure were acclimated for 6 days. (A) hGLC were treated without (Control) or with human chorionic gonadotropin (hCG) (1 IU/mL) for 0, 12, 24, and 36 hours. Right panel: The levels of mRNA for ACE2 were measured by quantitative polymerase chain reaction (qPCR) and normalized to the levels of RNA, 18S ribosomal 5 (RNA18S5) in each sample (n ≥7 independent samples). Bars with no common superscripts are significantly different (P<.05). Left panel: Representative Western blot image detecting ACE2 protein. ACTB detection in each lane was used as a protein loading control. The experiments were repeated three times, each with independent samples. (B) The cells were treated without (Control) or with hCG in the presence or absence of AG1478 (10 μM), RU486 (20 μM), CORT125281 (50 μM), or dexamethasone (20 μM) for 24 and 36 hours. Right panel: The levels of mRNA for ACE2 were measured by qPCR and normalized to the levels of RNA18S5 in each sample (n = 7 independent patient samples). Bars with no common superscripts are significantly different (P<.05). Left panel: A representative Western blot image detecting ACE2 protein. ACTB detection in each lane was used as a protein loading control. The experiments were repeated three times, each with independent patient samples.
Next, to determine whether the hCG-induced increases in ACE2 expression were mediated by key ovulatory mediators, such as the EGF receptor signaling, progesterone/progesterone receptor (PGR) or glucocorticoids/glucocorticoid receptor (NR3C1), the hGLC were treated with or without hCG in the absence or presence of AG1478 (an EGF receptor inhibitor), RU486 (a dual antagonist of progesterone receptor and glucocorticoid receptor), CORT125281 (a selective NR3C1 antagonist), or dexamethasone (a synthetic glucocorticoid). As expected, hCG increased ACE2 expression, but the hCG-induced increases were completely abolished by RU486 at both 24 and 36 hours (Fig. 4B). CORT125281 treatment also resulted in a partial reduction of hCG-induced ACE2 expression at both time points. In contrast, neither AG1478 nor dexamethasone affected basal or hCG-induced ACE2 expression.
Discussion
In women throughout their reproductive years, the ovary undergoes a series of cyclic changes during the menstrual cycle, culminating in ovulation. The preovulatory LH surge initiates these cyclic changes by inducing the expression of specific genes. The encoded products of these ovulatory genes exert their biological actions to bring about necessary cellular and extracellular changes required for ovulation and corpus luteum formation. In the present study, we showed that the expression of ACE2, an enzyme with carboxypeptidase activity and a primary receptor for SARS-CoV-2, is rapidly and dramatically induced after hCG administration in dominant follicles obtained from naturally cycling women throughout the periovulatory period. Using the primary hGLC model that can recapitulate key ovulatory changes in gene expression, this study provided further experimental evidence that the ovulatory induction of ACE2 expression was mediated by hCG and the hCG-induced steroid hormones progesterone and glucocorticoids in ovulatory follicles. Another noteworthy finding is the higher expression of ACE2 in cumulus cells than in granulosa cells collected immediately before ovulation. Together, this novel information not only suggests the potential involvement of ACE2 as a critical enzyme for the LH surge-induced cyclic events of ovulation, cumulus expansion, oocyte maturation, and luteal formation, but also implies a possible impact of COVID-19 on vital cyclic ovarian functions and thus on women’s overall reproductive health.
To the best of our knowledge, our comprehensive in vivo study is novel in that it is the first report documenting the ovulatory induction of ACE2 expression in naturally cycling women and the only report showing the dramatic and rapid upregulation of ACE2 expression after ovulatory induction in any species studied so far. In cattle, ACE2 mRNA levels were initially down-regulated in granulosa cells after induction of the LH surge but increased back to the preovulatory level 24 hours after gonadotropin-releasing hormone injection, whereas no changes were observed in thecal levels of ACE2 mRNA throughout the periovulatory period (15). No significant changes in the ACE2 expression pattern were observed in preovulatory follicles obtained before and throughout the ovulatory period in the nonhuman primate (Supplemental Fig. 1, available online) (32). In the mouse ovary, the ACE2 mRNA profile showed low abundance without a clear pattern throughout follicular development and the ovulatory period (http://okdb.appliedbioinfo.net and Supplemental Fig. 2). Therefore, our data indicate that the dramatic upregulation of ACE2 expression is specific to human ovulatory follicles and further point to the differences in specific mechanisms underlying the ovulatory process among different species. Of note, in the present study, hCG (rhCG for in vivo and urinary hCG for in vitro studies) was given to patients to mimic the endogenous LH surge. This was done to control exactly when the ovulatory trigger was initiated in individual patients. Therefore, it remains to be determined whether the endogenous LH surge and hCG administration would exert the same effect on ACE2 expression.
Another intriguing finding is the unique localization pattern of ACE2 protein observed in human ovulatory follicles. The initial induction of ACE2 staining was localized evenly throughout the granulosa and theca cell layer during the early ovulatory phase, and then the staining became more intense and progressively more sporadic among cells when the dominant follicle progressed toward ovulation and transformed into the corpus luteum. The uneven, localized staining of ACE2 did not appear in nonfollicular cells such as endothelial cells or leukocytes when compared with the PECAM1 staining pattern in serial sections of the same follicle. Consistent with this finding, qPCR analysis of ACE2 mRNA levels showed little to undetectable levels of ACE2 mRNA in leukocytes compared with granulosa cells (Supplemental Fig. 3). One possible explanation for this unique localization pattern is that ACE2 exists in two different forms, the membrane-bound form and the secreted form (33). During the early ovulatory phase when the level of ACE2 mRNA was rapidly up-regulated, ACE2 would have been processed and expressed as a single-pass type I membrane protein. Then, with the progression toward the late and postovulatory phases, the enzymatically active extracellular domain of ACE2 could be cleaved, released, and aggregated onto specific cells or the area where the substrates for ACE2 were present. This possibility needs to be explored in future studies.
To explore the cellular mechanism underlying ovulatory ACE2 expression, we used granulosa cells pooled from multiple follicles of each patient obtained during IVF oocyte retrieval. Despite the potential limitations of pooling granulosa cells that may have come from different development stages of the follicle, these cells (hGLC) increased the expression of genes involved in the ovulatory process, including the genes for PGR and EGF-like peptides, and the production of progesterone and prostaglandins in response to hCG (28). Similarly, consistent with the in vivo expression pattern, hCG stimulated ACE2 expression in hGLC in vitro. This induction was completely inhibited by RU486, a dual antagonist of PGR and NR3C1, and partially inhibited by CORT125281, a selective antagonist of NR3C1. Our pilot study showed that hCG increased NR3C1 expression and cortisol production in hGLC (Supplemental Fig. 4). These data, taken together, indicate that the up-regulated expression of ACE2 was mediated by the actions of progesterone and glucocorticoids in granulosa cells of the ovulatory follicle. Similar to our findings from granulosa cells, a recent study by Chadchan et al. (34) reported that progesterone promoted ACE2 expression in the endometrial stroma of both humans and mice. The existing data on the regulation of ACE2 expression by glucocorticoids or its receptor appear to be dependent on the cell types and models tested (35, 36). Therefore, our findings of the regulatory mechanisms involved in ACE2 expression provide new insights into the ovulatory process and valuable data that may be useful in understanding SARS-CoV-2 infection in reproductive-aged women.
In normal physiology, ACE2 is best known for its ability to hydrolyze Ang II to Ang-(1–7) (8, 9). Therefore, the marked increases in ACE2 expression could mean a shift in the balance between Ang II and Ang-(1–7), reducing Ang II levels while increasing Ang-(1–7) levels. Both Ang II and Ang-(1–7) are present in the follicular fluid collected at the time of IVF in women (18). Studies in animal models have shown that Ang II and Ang-(1–7) stimulate steroid production, regulate blood flow, and promote oocyte maturation and ovulation (20, 37, 38, 39). Therefore, the current findings showing rapid induction of ACE2 expression in human ovulatory follicles and high expression of ACE2 in cumulus cells support the notion that Ang II/ACE2/Ang-(1–7) is likely involved in cumulus expansion, oocyte maturation, and the ovulatory process in the human ovary.
With the current global challenge of COVID-19, our findings take on another level of significance, since ACE2 serves as a primary entry receptor for SARS-CO-2. A recent study reported ACE2 expression in oocytes, cumulus cells, and granulosa cells collected from patients at the time of IVF procedure (40). In this study, the expression of TMPRSS2, CD147, and CTSL was also documented in these ovarian cells as factors involved in SARS-CoV-2 infection (40). Therefore, our current findings showing the rapid and dramatic induction of ACE2 expression, together with the findings from the previous study (40), offer a compelling possibility that the ovary could be a target of SARS-CoV-2, being particularly vulnerable during the periovulatory period. In the scenario of SARS-CoV-2 infection in the ovary, this virus could have a negative impact on female fertility, not only by blocking the physiological action of ACE2 necessary for oocyte maturation, ovulation, and corpus luteum formation, but also by destroying mature oocytes and ovulatory follicles. In support of these possibilities, Orvieto et al. (41) reported that the proportion of top-quality embryos was significantly reduced in couples undergoing consecutive IVF cycles after recovering from COVID-19 infection compared with the proportion before the infection. Therefore, it will be vital to give adequate attention to the reproductive health of women who have had COVID-19 infection and to provide appropriate precautions and screening to women seeking IVF treatment. Once more data and information become available, it will be of great interest to assess the impact of COVID-19 infection on female fertility.
Acknowledgments
The authors thank Drs. Patrick Hannon, Ph.D. and Ketan Shrestha, Ph.D. for critical reading of the manuscript.
DIALOG: You can discuss this article with its authors and other readers at https://www.fertstertdialog.com/posts/33000
Footnotes
Y.C. has nothing to disclose. H.J. has nothing to disclose. M.B. has nothing to disclose. J.W.A. has nothing to disclose. T.E.C. has nothing to disclose. M.J. has nothing to disclose.
Supported by grants P01HD71875 (to M.J. and T.E.C.), R03HD095098 (to M.J.), and R01HD096077 (to M.J.) and the Biospecimen and Tissue Procurement Shared Resource Facility of the University of Kentucky Markey Cancer Center (P30CA177558).
Supplementary data
Supplemental Figure 1.

ACE2 mRNA profile extracted from GEODatasets (Accession: GDS3863). Microarray analysis of ovulatory, luteinizing follicles from a primate model of controlled ovulation at 0, 12, 24, 36 hours after exposure to an ovulatory (exogenous hCG) stimulus during the menstrual cycle (PMID: 21036944). A bar graph above was generated by imputing the term “ACE2” (https://www.ncbi.nlm.nih.gov/ geoprofiles?term = GDS3863[ACCN]+ACE2). A red bar represents the expression measurement from the value of each sample.
Supplemental Figure 2.

The profile of Ace2 mRNA during the follicular development and ovulatory period in the mouse ovary. The profile of mRNA for Ace2 depicted in the Ovarian Kaleidoscope Database (http://okdb.appliedbioinfo.net) was redrawn. This data set was based on microarray data using immature mice that were injected with HMG (FSH+LH activity) and 48 hours later with hCG to induce ovulation. Ovaries were collected before and every 2 hours after HMG and every 1 hour after hCG administration.
Supplemental Figure 3.
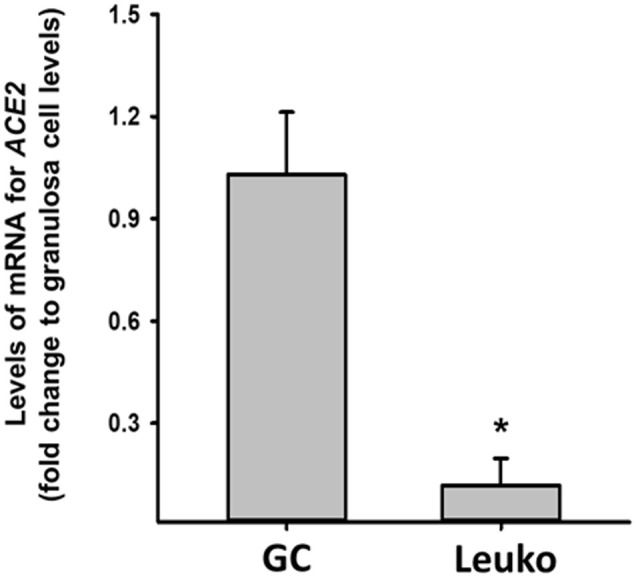
Comparison of levels of mRNA for ACE2 in granulosa cells (GC) vs. leukocytes (Leuko). Follicular aspirates collected at the time of oocyte retrieval from IVF patients were incubated in red blood cell lysis buffer and filtered through a 40-μm cell strainer. The flow-through cells were mixed and incubated with a human CD45 microbead (Miltenyi Biotec) to magnetically label leukocytes and then subjected to a Magnetic-activated cell sorting (MACS) column placed in an AutoMAC Pro Separator (Miltenyi Biotec). Negative (GCs) and positive (Leuko) fractions were separately collected and used to isolate to total RNA. Experiments were performed from 3 individual patient samples. ∗, p < 0.05
Supplemental Figure 4.

Effects of hCG on NR3C1 expression and cortisol production in hGLCs. Granulosa/lutein cells obtained from IVF patients were cultured for 6 d and then treated with or without hCG (1 IU/) for 36 hours. (A) NR3C1 was detected by Western blots using whole-cell extracts. The membrane was reprobed with a monoclonal antibody against ACTB to assess protein loading in each lane. (B) Cortisol levels in culture media were measured by a cortisol ELISA kit (R&D systems) after performing extraction procedures described by Yong et. al (PMID: 11134135) (n = 4 individual patient samples; ∗, p ≤ 0.05).
References
- 1.Yan R., Zhang Y., Li Y., Xia L., Guo Y., Zhou Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science. 2020;367:1444–1448. doi: 10.1126/science.abb2762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Donoghue M., Hsieh F., Baronas E., Godbout K., Gosselin M., Stagliano N., et al. A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9. Circ Res. 2000;87:E1–E9. doi: 10.1161/01.res.87.5.e1. [DOI] [PubMed] [Google Scholar]
- 3.Vickers C., Hales P., Kaushik V., Dick L., Gavin J., Tang J., et al. Hydrolysis of biological peptides by human angiotensin-converting enzyme-related carboxypeptidase. J Biol Chem. 2002;277:14838–14843. doi: 10.1074/jbc.M200581200. [DOI] [PubMed] [Google Scholar]
- 4.Tipnis S.R., Hooper N.M., Hyde R., Karran E., Christie G., Turner A.J. A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J Biol Chem. 2000;275:33238–33243. doi: 10.1074/jbc.M002615200. [DOI] [PubMed] [Google Scholar]
- 5.Soubrier F., Hubert C., Testut P., Nadaud S., Alhenc-Gelas F., Corvol P. Molecular biology of the angiotensin I converting enzyme: I. Biochemistry and structure of the gene. J Hypertens. 1993;11:471–476. doi: 10.1097/00004872-199305000-00001. [DOI] [PubMed] [Google Scholar]
- 6.Brewster U.C., Perazella M.A. The renin-angiotensin-aldosterone system and the kidney: effects on kidney disease. Am J Med. 2004;116:263–272. doi: 10.1016/j.amjmed.2003.09.034. [DOI] [PubMed] [Google Scholar]
- 7.Benigni A., Cassis P., Remuzzi G. Angiotensin II revisited: new roles in inflammation, immunology and aging. EMBO Mol Med. 2010;2:247–257. doi: 10.1002/emmm.201000080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Santos R.A., Ferreira A.J., Verano-Braga T., Bader M. Angiotensin-converting enzyme 2, angiotensin-(1–7) and Mas: new players of the renin-angiotensin system. J Endocrinol. 2013;216:R1–R17. doi: 10.1530/JOE-12-0341. [DOI] [PubMed] [Google Scholar]
- 9.Santos R.A.S., Sampaio W.O., Alzamora A.C., Motta-Santos D., Alenina N., Bader M., et al. The ACE2/angiotensin-(1–7)/MAS axis of the renin-angiotensin system: focus on angiotensin-(1–7) Physiol Rev. 2018;98:505–553. doi: 10.1152/physrev.00023.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang W., McKinnie S.M., Farhan M., Paul M., McDonald T., McLean B., et al. Angiotensin-converting enzyme 2 metabolizes and partially inactivates Pyr-Apelin-13 and Apelin-17: physiological effects in the cardiovascular system. Hypertension. 2016;68:365–377. doi: 10.1161/HYPERTENSIONAHA.115.06892. [DOI] [PubMed] [Google Scholar]
- 11.Li W., Zhang C., Sui J., Kuhn J.H., Moore M.J., Luo S., et al. Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. EMBO J. 2005;24:1634–1643. doi: 10.1038/sj.emboj.7600640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang Y., Wang Y., Luo W., Huang L., Xiao J., Li F., et al. A comprehensive investigation of the mRNA and protein level of ACE2, the putative receptor of SARS-CoV-2, in human tissues and blood cells. Int J Med Sci. 2020;17:1522–1531. doi: 10.7150/ijms.46695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hamming I., Timens W., Bulthuis M.L., Lely A.T., Navis G., van Goor H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J Pathol. 2004;203:631–637. doi: 10.1002/path.1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Douglas G.C., O'Bryan M.K., Hedger M.P., Lee D.K., Yarski M.A., Smith A.I., et al. The novel angiotensin-converting enzyme (ACE) homolog, ACE2, is selectively expressed by adult Leydig cells of the testis. Endocrinology. 2004;145:4703–4711. doi: 10.1210/en.2004-0443. [DOI] [PubMed] [Google Scholar]
- 15.Tonellotto dos Santos J., Ferreira R., Gasperin B.G., Siqueira L.C., de Oliveira J.F., Santos R.A., et al. Molecular characterization and regulation of the angiotensin-converting enzyme type 2/angiotensin-(1–7)/MAS receptor axis during the ovulation process in cattle. J Renin Angiotensin Aldosterone Syst. 2012;13:91–98. doi: 10.1177/1470320311417273. [DOI] [PubMed] [Google Scholar]
- 16.Pereira V.M., Reis F.M., Santos R.A., Cassali G.D., Santos S.H., Honorato-Sampaio K., et al. Gonadotropin stimulation increases the expression of angiotensin-(1–7) and MAS receptor in the rat ovary. Reprod Sci. 2009;16:1165–1174. doi: 10.1177/1933719109343309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Viana G.E., Pereira V.M., Honorato-Sampaio K., Oliveira C.A., Santos R.A., Reis A.M. Angiotensin-(1–7) induces ovulation and steroidogenesis in perfused rabbit ovaries. Exp Physiol. 2011;96:957–965. doi: 10.1113/expphysiol.2011.058453. [DOI] [PubMed] [Google Scholar]
- 18.Reis F.M., Bouissou D.R., Pereira V.M., Camargos A.F., dos Reis A.M., Santos R.A. Angiotensin-(1–7), its receptor Mas, and the angiotensin-converting enzyme type 2 are expressed in the human ovary. Fertil Steril. 2011;95:176–181. doi: 10.1016/j.fertnstert.2010.06.060. [DOI] [PubMed] [Google Scholar]
- 19.Costa A.P., Fagundes-Moura C.R., Pereira V.M., Silva L.F., Vieira M.A., Santos R.A., et al. Angiotensin-(1–7): a novel peptide in the ovary. Endocrinology. 2003;144:1942–1948. doi: 10.1210/en.2002-220787. [DOI] [PubMed] [Google Scholar]
- 20.Honorato-Sampaio K., Pereira V.M., Santos R.A., Reis A.M. Evidence that angiotensin-(1–7) is an intermediate of gonadotrophin-induced oocyte maturation in the rat preovulatory follicle. Exp Physiol. 2012;97:642–650. doi: 10.1113/expphysiol.2011.061960. [DOI] [PubMed] [Google Scholar]
- 21.Ferreira R., Oliveira J.F., Fernandes R., Moraes J.F., Gonçalves P.B. The role of angiotensin II in the early stages of bovine ovulation. Reproduction. 2007;134:713–719. doi: 10.1530/REP-07-0239. [DOI] [PubMed] [Google Scholar]
- 22.Mikuni M., Brännström M., Hellberg P., Peterson C.A., Pall M., Edwin S.S., et al. Saralasin-induced inhibition of ovulation in the in vitro perfused rat ovary is not replicated by the angiotensin II type-2 receptor antagonist PD123319. Am J Obstet Gynecol. 1998;179:35–40. doi: 10.1016/s0002-9378(98)70248-0. [DOI] [PubMed] [Google Scholar]
- 23.Cavallo I.K., Dela Cruz C., Oliveira M.L., Del Puerto H.L., Dias J.A., Lobach V.N., et al. Angiotensin-(1–7) in human follicular fluid correlates with oocyte maturation. Hum Reprod. 2017;32:1318–1324. doi: 10.1093/humrep/dex072. [DOI] [PubMed] [Google Scholar]
- 24.Duffy D.M., Ko C., Jo M., Brannstrom M., Curry T.E. Ovulation: parallels with inflammatory processes. Endocr Rev. 2019;40:369–416. doi: 10.1210/er.2018-00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bian X.W., Team T.C.-P. Autopsy of COVID-19 patients in China. Natl Sci Rev. 2020;7:1414–1418. doi: 10.1093/nsr/nwaa123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Al-Alem L., Puttabyatappa M., Rosewell K., Brännström M., Akin J., Boldt J., et al. Chemokine ligand 20: a signal for leukocyte recruitment during human ovulation? Endocrinology. 2015;156:3358–3369. doi: 10.1210/en.2014-1874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lind A.K., Dahm-Kähler P., Weijdegård B., Sundfeldt K., Brännström M. Gelatinases and their inhibitors during human ovulation: increased expression of tissue inhibitor of matrix mtalloproteinase-1. Mol Hum Reprod. 2006;12:725–736. doi: 10.1093/molehr/gal086. [DOI] [PubMed] [Google Scholar]
- 28.Choi Y., Wilson K., Hannon P.R., Rosewell K.L., Brännström M., Akin J.W., et al. Coordinated regulation among progesterone, prostaglandins, and EGF-like factors in human ovulatory follicles. J Clin Endocrinol Metab. 2017;102:1971–1982. doi: 10.1210/jc.2016-3153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Livak K.J., Schmittgen T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(–delta delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 30.Trau H.A., Davis J.S., Duffy D.M. Angiogenesis in the primate ovulatory follicle is stimulated by luteinizing hormone via prostaglandin E2. Biol Reprod. 2015;92:15. doi: 10.1095/biolreprod.114.123711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bergom C., Paddock C., Gao C., Holyst T., Newman D.K., Newman P.J. An alternatively spliced isoform of PECAM-1 is expressed at high levels in human and murine tissues, and suggests a novel role for the C-terminus of PECAM-1 in cytoprotective signaling. J Cell Sci. 2008;121:1235–1242. doi: 10.1242/jcs.025163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Xu F., Stouffer R.L., Müller J., Hennebold J.D., Wright J.W., Bahar A., et al. Dynamics of the transcriptome in the primate ovulatory follicle. Mol Hum Reprod. 2011;17:152–165. doi: 10.1093/molehr/gaq089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Grobe N., Di Fulvio M., Kashkari N., Chodavarapu H., Somineni H.K., Singh R., et al. Functional and molecular evidence for expression of the renin angiotensin system and ADAM17-mediated ACE2 shedding in COS7 cells. Am J Physiol Cell Physiol. 2015;308:C767–C777. doi: 10.1152/ajpcell.00247.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chadchan S.B., Popli P., Maurya V.K., Kommagani R. The SARS-CoV-2 receptor, angiotensin-converting enzyme 2, is required for human endometrial stromal cell decidualization. Biol Reprod. 2021;104:336–343. doi: 10.1093/biolre/ioaa211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Xiang Z., Liu J., Shi D., Chen W., Li J., Yan R., et al. Glucocorticoids improve severe or critical COVID-19 by activating ACE2 and reducing IL-6 levels. Int J Biol Sci. 2020;16:2382–2391. doi: 10.7150/ijbs.47652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Young M.J., Clyne C.D., Chapman K.E. Endocrine aspects of ACE2 regulation: RAAS, steroid hormones and SARS-CoV-2. J Endocrinol. 2020;247:R45–R62. doi: 10.1530/JOE-20-0260. [DOI] [PubMed] [Google Scholar]
- 37.Domińska K. Involvement of ACE2/Ang-(1–7)/MAS1 axis in the regulation of ovarian function in mammals. Int J Mol Sci. 2020;21:4572. doi: 10.3390/ijms21134572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gonçalves P.B., Ferreira R., Gasperin B., Oliveira J.F. Role of angiotensin in ovarian follicular development and ovulation in mammals: a review of recent advances. Reproduction. 2012;143:11–20. doi: 10.1530/REP-11-0192. [DOI] [PubMed] [Google Scholar]
- 39.Mitsube K., Mikuni M., Matousek M., Zackrisson U., Brännström M. Role of the angiotensin II system in regulation of ovulation and blood flow in the rat ovary. Reproduction. 2003;125:425–435. doi: 10.1530/rep.0.1250425. [DOI] [PubMed] [Google Scholar]
- 40.Rajput S.K., Logsdon D.M., Kile B., Engelhorn H.J., Goheen B., Khan S., et al. Human eggs, zygotes, and embryos express the receptor angiotensin 1-converting enzyme 2 and transmembrane serine protease 2 protein necessary for severe acute respiratory syndrome coronavirus 2 infection. F S Sci. 2021;2:33–42. doi: 10.1016/j.xfss.2020.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Orvieto R., Segev-Zahav A., Aizer A. Does COVID-19 infection influence patients' performance during IVF-ET cycle?: an observational study. Gynecol Endocrinol. 2021;11:1–3. doi: 10.1080/09513590.2021.1918080. [DOI] [PubMed] [Google Scholar]