

Abstract
On the mountains, along an elevation gradient, we generally observe an ample variation in temperature, with the associated difference in vegetation structure and composition and soil properties. With the aim of quantifying the relative importance of temperature, vegetation and edaphic properties on soil respiration (SR), we investigated changes in SR along an elevation gradient (404 to 2101 m a.s.l) in the southern slopes of the Alps in Northern Italy. We also analysed soil physicochemical properties, including soil organic carbon (SOC) and nitrogen (N) stocks, fine root C and N, litter C and N, soil bulk densities and soil pH at five forest sites, and also stand structural properties, including vegetation height, age and basal area. Our results indicated that SR rates increased with temperature in all sites, and 55–76% of SR variability was explained by temperature. Annual cumulative SR, ranging between 0.65–1.40 kg C m-2 yr-1, decreased along the elevation gradient, while temperature sensitivity (Q10) of SR increased with elevation. However, a high SR rate (1.27 kg C m-2 yr-1) and low Q10 were recorded in the mature conifer forest stand at 1731 m a.s.l., characterized by an uneven-aged structure and high dominant tree height, resulting in a nonlinear relationship between elevation and temperature. Reference SR at 10°C (SRref) was unrelated to elevation, but was related to tree height. A significant negative linear relationship was found between bulk density and elevation. Conversely, SOC, root C and N stock, pH, and litter mass were best fitted by nonlinear relationships with elevation. However, these parameters were not significantly correlated with SR when the effect of temperature was removed (SRref). These results demonstrate that the main factor affecting SR in forest ecosystems along this Alpine elevation gradient is temperature, but its regulating role can be strongly influenced by site biological characteristics, particularly vegetation type and structure, affecting litter quality and microclimate. This study also confirms that high elevation sites are rich in SOC and more sensitive to climate change, being prone to high C losses as CO2. Furthermore, our data indicate a positive relationship between Q10 and dominant tree height, suggesting that mature forest ecosystems characterized by an uneven-age structure, high SRref and moderate Q10, may be more resilient.
Introduction
Soil respiration (SR) is the largest biological carbon (C) flux after photosynthesis in terrestrial ecosystems, and is estimated to release 50–77 Pg C yr-1 globally [1–3]. This major natural flux largely determines the C balance between the terrestrial biosphere and the atmosphere [4–6] and plays a critical role in the carbon cycle. Soil is the largest C pool in the terrestrial biosphere and has been increasingly recognized to play a crucial role in mitigating global warming resulting from climate change [7–9]. Small changes in soil CO2 efflux or soil organic C stocks could severely impact the global C cycle [10]. The important role of forest productivity in the determination of SR was first demonstrated by Janssens et al. [11] and other studies have subsequently investigated and quantified the impact of productivity on SR modeling [12, 13]. In addition to productivity, SR is influenced by different abiotic and biotic factors such as soil temperature, soil moisture, and microbial community composition, introducing considerable uncertainty in SR estimates [14–16]. Among these factors affecting respiratory processes, the temperature has been the most well-studied [17, 18]. Many studies have addressed the prediction of SR response to increasing temperature (i.e. temperature sensitivity of SR), producing different equations relating soil CO2 efflux to temperature [19–21] or a combination of temperature and soil humidity [22]. However, the Q10 function [23], using the Q10 parameter to describe the temperature sensitivity of SR, is one of the most widely used models to quantify CO2 efflux from the soil in Earth system models.
The elevation is a key driver of climate properties, playing an essential role in soil organic matter content and mitigating the effects of climate change [16, 24–27]. Typically, temperature declines with elevation, thus elevation gradients have been widely used to assess soil respiration response to temperature. Studies suggest that CO2 exchange between soil and atmosphere varies along climatic gradients and that the temperature sensitivity (Q10) of SR increases with elevation [14, 28–30]. Furthermore, a positive relationship between soil organic carbon (SOC) and elevation indicates that global soil organic C stocks at high elevation are more sensitive to climate change and are predicted to decrease in a warming climate [16, 25, 31–34]. However, other studies have reported opposite trends, finding lower SOC content and higher SR at high elevations [32, 35, 36]. This variability may be partially due to confounding factors, other than temperature, affecting SR. Besides elevation, mountain landscapes are, in fact, characterized by substantial local changes in other parameters such as slope angle and orientation, which can affect microclimatic conditions and, therefore, soil C dynamics [37]. Furthermore, due to the heterogeneity of geological substrates, soils in mountain regions show high small-scale heterogeneity, which can generate marked differences in soil biogeochemical properties [38]. Studies have also found conflicting results in terms of the relationship between SR and SOC [28, 39].
Furthermore, the various plant communities can affect soil respiration rate through the diverse microclimate, soil and litter composition, and root distribution [20, 28, 40–42]. However, within the same plant biome, there is also high spatial heterogeneity in SR. Some authors have found a possible linkage between the topography, plant community structure (e.g., forest type and speed of regeneration), and SR within the same forest ecosystem [20, 40–42]. Furthermore, forest management can also play a crucial role in SR [43]. For example, tree removal can directly influence SR due to the removal itself (i.e., reduction of plant biomass) but also through indirectly changing the soil physicochemical properties and micrometeorological conditions [44].
Currently, the temperature dependency of SR and SOC decomposition is of major interest in the study of global climate change and the role of terrestrial ecosystems in regulating Earth´s climate [45, 46]. Therefore, there is a need to better understand the interactions between temperature and soil CO2 efflux. The general goal of this study was to disentangle the possible multi-effects of soil properties, temperature, SOC, and vegetation structure (in particular tree height and age) on SR along a plant biome elevation gradient. In particular, the existing differences in vegetation structure allowed us to investigate the extent to which these biological variables can alter the relation between elevation and SR.
Specifically, we tested the hypotheses that i) SR and SOC accumulation change linearly with elevation; and ii) the Q10 value increases linearly with elevation. Furthermore, we determined the main factors, other than temperature, affecting SR. To effectively isolate the effect of temperature on SR, the study was conducted along an altitudinal gradient in the Italian Alps, where confounding factors such as slope angle and orientation, and soil parent material were minimized. The differences in vegetation structure allowed us to investigate to which extent these biological variables, and the induced variation in microclimatology, can alter the relationship between elevation and SR.
Material and methods
Study sites
Five experimental sites were established between the top of the Rittner Horn and the city of Bozen, Italy, on the southern side of the Alps (Fig 1A). The overall elevation gradient between the highest (A) and the lowest site (E) is 1697 m and the elevation separation between each site is approximately 424 ± 60 m. All sites are characterized by the soil developed upon a glacial till laid on a porphyric bedrock and a SE slope orientation. Previous analyses performed at site B revealed that the soil was poor in ions forming carbonates (Ca++ = 106.2 meq/kg, Mg+ = 58.0 meq/kg), indicating that the spatial variability of soil C is largely determined by SOC. Soil CO2 flux measurements were taken in 2017 and 2018, during which time annual average precipitation ranged between 746 mm (Bozen, near the lowest site (site E)) and 1090 mm (measured near-site B at 1780 m) (data from the Hydrographic Office and the Agency for the Environment of the Autonomous Province of Bozen). Based on Wehren et al. [47], precipitation was expected to increase along the elevation gradient, with a minimum at site E and a maximum at site A. Details on the main characteristics of the research sites are reported in Table 1 and S1 File.
Fig 1.

(a) Map showing study sites (b) Scheme of vegetation along the elevation gradient.
Table 1. General characteristics of the study sites.
| Characteristics | Site A | Site B | Site C | Site D | Site E |
|---|---|---|---|---|---|
| Elevation (m a.s.l.) | 2101 | 1731 | 1354 | 865 | 404 |
| Mean annual temperature (°C) | 4 | 4 | 12 | 11 | 14 |
| Slope orientation | SE | SE | SE | SE | SE |
| Slope inclination (%) | 15 | 18 | 12 | 18 | 25 |
| Stand age (yr) | 61 | 202 | 153 | 54 | 88 |
| Land use | Shrubland | Forest | Forest | Forest | Forest |
| Dominant tree height (m) | 1.9 | 29 | 22.5 | 18.8 | 8.7 |
| Basal area (m2 ha-1) | 7.1 | 42.0 | 42.7 | 39.8 | 22.8 |
| Dominant overstory species | Dwarf Mountain pines (Pinus mugo) | Norway spruce (Picea abies), Swiss stone pine (Pinus cembra), Larch (Larix decidua) | Scotts pine (Pinus sylvestris) | Sessile oak (Quercus petrea), Scotts Pine (Pinus sylvestris), Chestnut (Castanea sativa) | Sessile oak (Quercus petrea), Flowering ash (Fraxinus ornus) |
| Main understory species | Intervening grasses (Festuca halleri) | Rusty leaved alprose (Rhododendron ferrugineum) | Heather (Erica carnea) | Understory almost absent | Smoke-bush (Cotinus coggygria), Succulent plants (Opuntia humifusa), Scorpion senna (Hippocrepis emerus). |
Site A was established in even-aged shrubland vegetation composed of Dwarf Mountain pine (Pinus mugo Turra) near the summit of the Rittner Horn. Pine trees at site A showed an even-aged distribution based on trunk diameter. The mean age was 61 yr.
Site B was established in a subalpine Norway spruce stand (Picea abies (L.) Karst.) at the Fluxnet research station of Renon-Mittelgrünwald (http://sites.fluxdata.org/IT-Ren/). The site is characterized by an uneven-aged distribution of individuals based on tree diameter, approaching the structure of old-growth forest stands [48, 49]. This type of structure was achieved by a traditional sylviculture treatment based on cutting small groups of trees. The oldest group of trees had a mean age of 202 yr.
Site C was located near Riggermoos, in a Scotts pine (Pinus sylvestris L:) stand, displaying a large basal area and an even-aged structure (mean age 153 yr.).
Site D was established in a mixed stand of Sessile oak (Quercus petrea (Matt.) Liebl.) and Chestnut (Castanea sativa L.), with some individuals of Scots pine and larch (Larix decidua L.) near the village of Signat. Forest vegetation at this site evolved from a former oak-chestnut coppice, with the presence of individuals of pines and larches grown as standards (mean suckers age 54 yr.).
Site E was located in a low stature stand dominated by Sessile oak and Flowering ash (Fraxinus ornus L.) on the hill slope of Sankt Magdalena, close to Bozen, with the entry in the plant community of tree species cultivated in the town gardens, for example, Lawson cypress (Chamaecyparis lawsoniana (A. Murray) Parl.
All sites except sites A and E are managed as high forests, mainly for wood harvesting. Site A is managed as natural vegetation with occasional harvesting at the forest margins to avoid expansion into the adjacent pasture areas. Site E has not been managed for decades. Tree age was assessed during sampling in 2018 and 2021 based on tree ring count. Forest tree diameter and height in circular plots of variable radius (5–25 m) were assessed during two sampling campaigns (2020 and 2021) with a TruePulse 360 B (Laser Tech, CO, USA).
The permission to carry out the experiment was obtained through the Forest Services of the Autonomous Province of Bozen (in case of public land, sites A and E) or by signed agreement with the landowners, in case of private land (sites B, C and D).
Soil respiration
To quantify SR, ten iron collars (10 cm height, 20 cm diameter) were inserted into the soil, three weeks before the first measurements at each site. Measurements were performed with an opaque survey chamber (Li-8100-104, LI-COR Biosciences, NE, USA) connected to an LI-8100 analyzer (LI-COR). By using this technique, an integrated value of SR is provided, which includes the heterotrophic CO2 emission from the different soil horizons and from the litter and the autotrophic CO2 emission from roots. On each collar, the measurement period was set to 120 s; the first 20 s of the measurement were considered dead-band, the last 20 s of purging, so the flux computation was limited to 80 s (see Montagnani et al. [10] for further details concerning the measurement settings). From July 21, 2017, SR measurements were performed periodically, about once per month, until July 20, 2018, for a total of 17 measurement days. The first four series of measurements were performed every three weeks at all the sites. During the winter, SR measurements were performed only in the lower elevation sites due to snow at higher elevations. The measurement calendar and flux data for the different sites are provided in S2 File. During measurements, the air temperature was measured inside the survey chamber (at 0.1 m above ground, RHT Plus, Skye Instruments, UK). A soil temperature profile was installed at control site B according to ICOS protocol [50]. Specifically, we used the -5 cm soil T data provided by a CS605 probe (Campbell Scientific, USA). We recorded soil temperature continuously at 30 min intervals throughout the experimental period.
Soil respiration data collected from each measurement point (collar) were related to chamber air temperature using a logistic model [28]:
| Eq 1 |
Where SR is soil respiration, a is the maximum value of SR, b determines the elongation of the SR curve along the x-axis, k is the logistic growth rate or steepness of the SR curve along the x-axis, and Ta is air temperature. Furthermore, SR data were also fitted with a Q10 model [51, 52]:
| Eq 2 |
Where SR is the soil respiration, SRref is the fitted SR at the reference temperature of 10°C (Tref), Q10 is the temperature sensitivity of SR, defined as the factor by which soil respiration increases with a 10°C temperature increase, and Ta is chamber air temperature.
Soil and plant analysis
After taking the last measurements (July 2018), the soil in each collar was sampled to a depth of 20 cm using a 4.8 split-corer (Eijkelkamp, NL). Litter present in each collar, which included leaf, cones and branch fragments was collected separately from the soil. Fine roots (<2 mm diameter) were separated from coarse roots using a caliper and were weighed after oven-drying at 105 ± 5°C. Larger roots (>2 mm) were analysed together with the soil. By using this fixed-depth approach of soil sampling, different soil horizons can be included in the sample. Being the soil organic (O horizon) depth different at the different locations, at site B, the O horizon only was included in the samples, while at the other sites the top of the mineral horizon was included as well, at least in a subset of samples.
In the laboratory, soil samples were weighed and sieved (2 mm mesh size) to separate roots, stones, and coarse organic matter fragments. The stones removed from each sample were weighed and their volume was determined on the base of their density. Soil bulk density was determined by dividing the weight of sieved soil by the sample volume, which was calculated substracting the volume of the stones from the core volume. Soil pH was measured using a pH-meter (Crison pH-Meter Basic 20+Electrod: Hach 50 10T CRISON, Barcelona, Spain) in a suspension with a soil:water ratio of 1:5. Fine roots, litter, and soil samples were analyzed for SOC and N content using a FlashEATM 1112 Elemental Analyzer (Thermo Fisher Scientific, Waltham, MA, USA).
Soil organic C stock was obtained as follows [53]:
| Eq 3 |
where C is the SOC content, BDsoil is the soil bulk density (kg dm−3) and 0.2 m is the sampling depth. Soil N stock was calculated with the same formula, using total N content in the place of SOC content. Since all sites were established on acidic soil and porphyric bedrock, carbonates contribution to total soil C were negligible and total soil C was almost equivalent to SOC.
Fine root C and N stocks were determined with the same computational approach, using fine root density (kg dm-3) instead of soil BD and fine root C and N content instead of SOC and N content. Litter C and N amounts were obtained by multiplying the litter dry weight by C or N content and dividing by the collar area. Data concerning soil and plant analysis is reported in S3 File.
In order to estimate the stand age at each site, three dominant trees of the most common species were cored with a 5 mm increment borer, then partially planed with a microtome blade to reveal growth rings. Lastly, age was measured through the LINTAB measuring device linked to the TSAP-Win software (Rinntech, Germany), following standard dendrochronological procedures [54].
Data analysis
Models were fitted to SR data using the nls package in R software [55]. Model fitness was evaluated based on Akaike’s Information Criterion (AIC), R-squared (R2), Mean Absolute Error (MAE), and Root Mean Squared Error (RMSE). The Q10 model was used to obtain the SRref and the Q10 value for every collar. Linear regression was used to compare the air temperature measured continuously in the reference plot (site B) and air temperature inside the chamber during SR measurements for each collar in each site. The obtained linear regression models were then used to predict chamber air temperature for the whole experimental period, for each collar, with a 30 min resolution (S4 File). Therefore, the predicted chamber air temperature was used to predict SR values simultaneously for the whole experimental period, based on the logistic models relating SR to chamber temperature. For some collars, it was not possible to obtain a good fit of the SR data using the logistic model; for these collars, the predicted soil respiration data from temperature was obtained using the Q10 model developed for the same collar. Finally, the total cumulative SR for the whole experimental period was determined for each collar at each site.
Soil respiration response to biological variables (SOC content, fine root C, litter C, fine root dry weight, soil N, fine root N, litter N, litter dry weight) was examined using Spearman’s Correlation Test and linear mixed-effects models (LMMs) fitted by restricted maximum likelihood (REML). Before applying LMMs, to avoid statistical errors, Variance Inflation Factor (VIF) was determined for biological variables, and variables with high VIF values were excluded from the model assessment. LMMs were built using the lme4 R package [56–58]. The models consisted of both fixed and random effects: biological variables were considered as fixed effects, and sampling plots (collars) nested in each site were used in the random-effects formula. R2 was used to summarize model goodness-of-fit together with AIC [59, 60]. Since computed R2 values by LMMs are a pseudo-R2 and technically incorrect, the r2glmm R package was used to compute R2 [60]. To exclude the confounding effect of temperature from LMMs and correlations tests, environmental variables were related to SRref instead of SR [22, 28, 61]. Furthermore, to assess the correlation between gross primary production (GPP) and SR, dominant tree height was used as a covariate in LMMs and Spearman’s correlation test. Statistical comparisons of average SOC and N content, fine root C and N, litter C and N, soil bulk density, soil pH, and soil respiration in the different sites were performed using the Kruskal-Wallis test (Dunn test, p < 0.05) for the non-normally distributed data, and a one-way ANOVA for the normally distributed data (Tukey test, p < 0.05). The normality of the data and homogeneity of variance were checked by the Shapiro–Wilk test and Levene’s test, respectively [62, 63]. To test linearity changes of SR, Q10 and SOC content with elevation, linear and nonlinear polynomial regressions were applied between elevation and environmental variables (SOC and soil N content, fine root C and N, litter C and N, soil bulk density, soil pH). The linearity changes of these variables with elevation were detected based on the lowest AIC and the highest R2. The association of Q10, SOC and soil N content with environmental variables was determined using Spearman’s Correlation Test. All statistical analyses were performed using R version 3.6.0 ([55], www.r-project.org).
Results
Environmental factors variability along the altitudinal gradient
A significant difference in SOC stock was found only between site E (3891 ± 2756 g C m-2), at the lowest elevation where the C stock was smallest, and sites A, B, and C (Fig 2A). No significant differences were found in soil N stock between the different sites (Fig 2E). Fine root biomass and fine root C and N stocks in site A were significantly higher than in other sites (Fig 2B, 2D and 2F) and litter dry weight in site B was significantly higher than site E (Fig 2H). However, the accumulated C and N in the litter did not significantly differ along the altitudinal gradient (Fig 2C and 2G). Significant differences were found between pH values in the different sites (Fig 2I): the lowest soil pH was measured in site B (pH 3.8 ± 0.1) and the highest in site E (pH 5.9 ± 0.2, Fig 2I). The highest bulk density was found in site D (1.01 ± 0.27 g cm-3, Fig 2J) and the lowest was found in site B (0.26 ± 0.16 g cm-3; Fig 2J).
Fig 2.

Stocks of C and N in soil and biomass (a-g); litter dry weight (h); soil pH (i); and soil bulk density (j), in the examined sites (A-E). Different letters indicate significant differences between sites. Vertical bars represent the standard deviation of the mean.
Based on the AIC and R2, only soil bulk density was linearly related to elevation (Table 2). On the contrary, SOC and soil N stock, fine root dry weight, fine root C and N stock, pH and litter dry weight were better fitted with nonlinear relationships with elevation (Table 2; further detail about equation parameters can be found in S5 File). Furthermore, a significant correlation was found between SOC and soil N and soil pH, dominant tree height, stand age, basal area, and bulk density (Table 3).
Table 2. Linear (lm) and polynomial regressions (poly) between the different parameters tested and elevation.
| Parameters | Elevation | |||||
|---|---|---|---|---|---|---|
| Significance | Multiple R2 | AIC | ||||
| lm | Poly | lm | Poly-lm | lm | Poly-lm | |
| SOC stock (kg C m-2) | *** | *** | 0.28 | 0.40 | 764 | 758 |
| Soil N stock (g N m-2) | * | * | 0.15 | 0.20 | 510 | 509 |
| Soil respiration (kg C m-2 yr-1) | *** | *** | 0.33 | 0.36 | 34 | 34 |
| Temperature sensitivity of soil respiration (Q10) | *** | ** | 0.28 | 0.41 | 77 | 67 |
| Respiration at the 10°C reference temperature (SRref) (kg C m-2 yr-1) | n.s. | n.s. | <0.01 | 0.08 | 139 | 139 |
| Fine root dry weight (kg m-2) | * | ** | 0.10 | 0.24 | 717 | 712 |
| Fine root C stock (kg C m-2) | * | *** | 0.39 | 0.80 | 199 | 186 |
| Fine root N stock (g N m-2) | n.s. | *** | 0.28 | 0.80 | 102 | 88 |
| Litter dry weight stock (g m-2) | n.s. | * | 0.07 | 0.12 | 774 | 772 |
| Litter C stock (kg C m-2) | n.s. | n.s. | < 0.01 | 0.04 | 222 | 223 |
| Litter N stock (g N m-2) | n.s. | n.s. | < 0.01 | < 0.01 | 112 | 114 |
| pH | ** | *** | 0.63 | 0.88 | 22 | 8 |
| Soil bulk density (kg dm-3) | *** | *** | 0.32 | 0.29 | 9 | 11 |
*—p ≤ 0.05;
**- p ≤ 0.01;
***—p ≤ 0.001; n.s.–not significant. BD–soil bulk density; Q10 –temperature sensitivity of soil respiration; SR -–cumulative soil respiration; SRref–respiration at the 10°C reference temperature; AIC–Akaike information criterion.
Table 3. Spearman rank coefficients for the correlations between different variables.
| Variable | SOC stock (kg C m-2) | Soil N stock (g N m-2) | Temperature sensitivity of soil respiration (Q10) |
|---|---|---|---|
| SOC stock (kg C m-2) | 1.00 | 0.94*** | 0.15n.s. |
| Soil N stock (g N m-2) | 0.94*** | 1.00 | 0.13n.s. |
| Fine root dry weight (g m-2) | 0.14n.s. | 0.11n.s. | 0.06n.s. |
| Fine root C stock (kg m-2) | -0.11n.s. | 0.36n.s. | 0.37n.s. |
| Fine root N stock (g N m-2) | -0.07n.s. | 0.32n.s. | 0.46n.s. |
| Litter dry weight (kg m-2) | 0.02n.s. | 0.01n.s. | -0.08n.s. |
| Litter C stock (kg C m-2) | -0.17n.s. | -0.36n.s. | 0.01n.s. |
| Litter N stock (g N m-2) | -0.09n.s. | -0.23n.s. | -0.44n.s. |
| pH | -0.62* | -0.54* | -0.29n.s. |
| Soil bulk density (kg dm-3) | -0.52* | -0.39* | -0.23n.s. |
| Dominant tree height (m) | 0.33* | 0.21n.s. | -0.29* |
| Basal area (m2) | 0.31* | 0.16n.s. | -0.27* |
| Stand age (yr) | 0.32* | 0.26n.s. | -0.11n.s. |
Asterisks indicate significance levels: *–p ≤ 0.05,
**–p ≤ 0.01, and
***—p ≤ 0.001; n.s.–nonsignificant. SOC–soil organic carbon.
Soil respiration
Both logistic and Q10 models confirmed that soil respiration rates increased with temperature in all sites (Fig 3), and the seasonal pattern of SR was similar to that of air chamber temperature (S6 File).
Fig 3. Rates of soil respiration against chamber air temperature in the different sites.
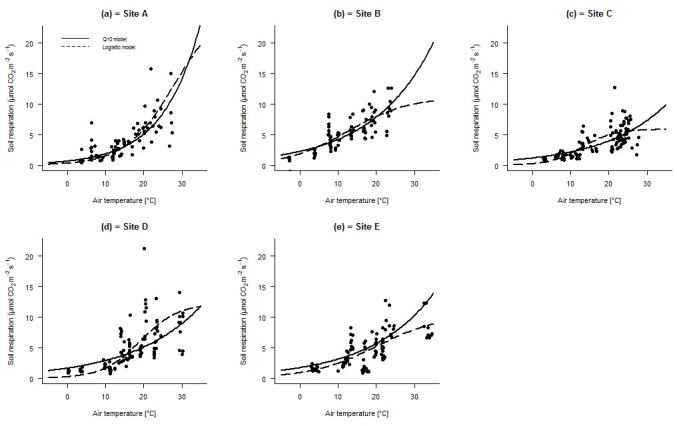
Regression lines were built using the mean values of model parameters obtained for different replicate collars of each site (n = 10).
A strong linear relationship was found between observed and predicted SR (R2 = 0.73; S7 File). Temperature explained between 55% and 76% of the variance in soil respiration at the experimental sites (Table 4). The Q10 and SRref values obtained for the different sites ranged between 1.75 and 2.96, and between 2.17 and 4.49 (μmol CO2 m–2 s–1), respectively (Table 4). The Q10 value recorded in site A (highest elevation) was significantly different from that of the other sites (Table 4). A significant linear relationship was identified between Q10 and elevation (Table 2). However, the trend between Q10 and temperature was best described by a nonlinear relationship (Table 2). Furthermore, a significant negative correlation was found between Q10 and mean dominant tree height and basal area (Table 3). No significant relationship between SRref and elevation was found (Table 2). However, significant differences were found between experimental sites; the highest SRref value was recorded in site B and the lowest values in sites A, C, and E (Table 4).
Table 4. Mean values of Q10 (temperature sensitivity) and SRref (soil respiration at the temperature of 10°C) for each site (A, B, C, D and E).
| A | B | C | D | E | |
|---|---|---|---|---|---|
| Q10 | 2.96 ± 0.72b | 1.83 ± 0.25a | 1.90 ± 0.24a | 1.75 ± 0.15a | 1.75 ± 0.32a |
| SRref (kg C m-2 yr-1) | 2.17 ± 0.60a | 4.49 ± 0.71c | 2.24 ± 0.58a | 3.17 ± 1.03b | 2.98 ± 0.41ab |
| AIC logistic model | 32.95 ± 10.45 | 34. 25 ± 6.45 | 34.19 ± 11.65 | 48.00 ± 14.55 | 49.76 ± 6.70 |
| AIC Q10 model | 30.42 ± 13.69 | 34.07 ± 7.31 | 35.39 ± 11.53 | 48.44 ± 14.13 | 49.11 ± 6.60 |
| R2 logistic model | 0.76 ± 021 | 0.75 ± 0.20 | 0.75 ± 0.16 | 0.67 ± 0.11 | 0.58 ± 0.10 |
| R2 Q10 model | 0.75 ± .23 | 0.71 ± 0.23 | 0.68 ± 0.21 | 0.61 ± 0.11 | 0.55 ± 0.11 |
| MAE logistic model | 1.06 ± 0.65 | 1.06 ± 0.45 | 0.68 ± 0.32 | 1.26 ± 0.62 | 1.29 ± 0.25 |
| MAE Q10 model | 1.07 ± 0.68 | 1.14 ± 0.52 | 0.79 ± 0.36 | 1.46 ± 0.76 | 1.39 ± 0.26 |
| RMSE logistic model | 1.36 ± 0.94 | 1.27 ± 0.50 | 0.86 ± 0.44 | 1.70 ± 0.89 | 1.71 ± 0.36 |
| RMSE Q10 model | 1.38 ± 1.0 | 1.36 ± 0.55 | 0.99 ± 0.49 | 1.87 ± 1.02 | 1.77 ± 0.38 |
MAE (Mean Absolute Error), RMSE (Root Mean Squared Error); R2 (Determination Coefficient) and AIC (Akaike information criterion). Different letters indicate significant differences between sites. Values are expressed as mean ± SD.
The cumulative SR in site A was significantly lower than in the other sites (Fig 4). A nonlinear relationship between cumulative SR and elevation was also found (Table 2).
Fig 4. Total cumulative soil respiration (kg C m-2 yr-1) for the different sites.
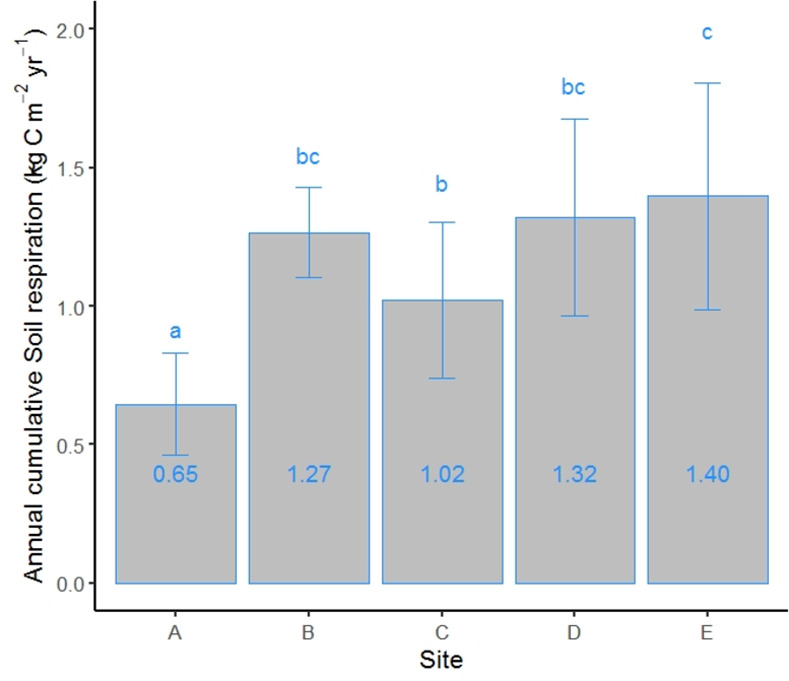
Error bars indicate standard deviation. Different letters indicate significant differences between sites.
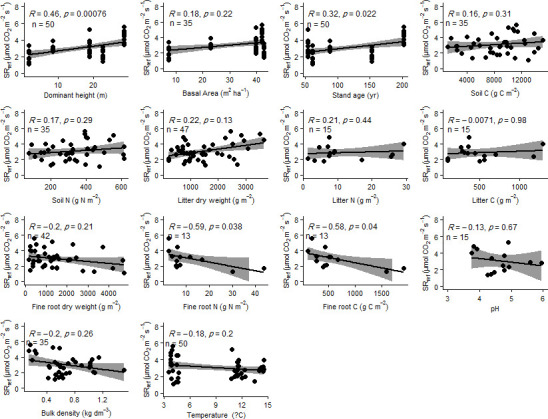
Soil organic C stock, mean dominant tree height, and litter dry weight best explained SR (SRref, μmol CO2 m–2 s–1 at 10°C) in LMMs (VIF<10; Table 5). According to the model, about 16% of SR was explained by tree height (R2 = 0.16, Table 5). A positive association between SR and mean dominant tree height and stand age, and a negative association between SR and fine root C and N, were found by Spearman’s Correlation Test (Fig 5).
Table 5. Results of linear mixed-effects models testing the impact of biological variables on the SRref at 10°C.
| Model variables | Value | VIF | p-value | R2 |
|---|---|---|---|---|
| Intercept | 1.51 | 0.004 | 0.29 | |
| SOC stock (kg C m-2) | < 0.01 | 1.20 | 0.67 | < 0.01 |
| Dominant tree height (m) | 0.05 | 1.21 | 0.01 | 0.16 |
| Litter dry weight (kg m-2) | < 0.01 | 1.05 | 0.09 | 0.07 |
Parameters in bold indicate significant correlations. VIF–Variance Inflation Factor.
Fig 5. Relationships between reference soil respiration at 10°C (SRref) and different site properties.
Discussion
Organic C and N content along the altitudinal gradient
The recorded ranges of SOC and soil N stocks were 3891–10270 g C m-2 and 219–397 g N m-2 respectively (Fig 2). These values were within the ranges reported by other studies along elevation gradients [28, 64]. The lowest value was recorded at the lowest elevation, and our data corroborated an increase of the soil organic C stock with increasing elevation shown in other studies [9, 26, 27, 29, 64–67]. Being the main driver for the loss of soil organic C, low temperature can limit the decomposition of organic matter at high elevations. For this reason, elevation could induce a significant increase in SOC [25, 65, 67–69]. However, an increment in SOC stock was not linearly linked with elevation gradient and, therefore, our first hypothesis was not confirmed. The non-linear relationship of SOC stock with elevation and the high value of SOC stock in site C (with a mean temperature of 12°C) suggests that factors other than temperature have influenced SOC accumulation. Different microclimatic or micromorphological conditions caused by differences in slope angle and orientation can influence C storage in soils [35, 37, 70]. However, in the present study, all the sites were characterized by a similar slope and were all south or south-east facing; therefore, we exclude an influence of these factors on SOC accumulation. According to recent studies, SOC is not consistently related to variation in climatic conditions along elevation gradients, but is also strongly affected by productivity or by vegetation type/composition [32, 35, 71, 72]. Our data confirm a significant positive correlation of SOC stock with mean dominant tree height, basal area and stand age (Table 3). Generally, tall trees in a mature forest stand can increase litterfall production. Therefore, the higher amount of SOC found in the present study at intermediate elevation in the older stands (B and C) could be explained by higher site productivity, which is indicated by the mean dominant tree height and basal area [73, 74]. Standing leaf area and litterfall increase with tree size, although this relation is strongly species-dependent, with leaf area increasing much more in spruce than in Scotts pine forests.
Soil pH and bulk density are considered to be two of the main variables influencing soil properties other than SOC, soil microbial activity and soil respiration [75, 76]. Generally, at high elevation, higher precipitation and lower evapotranspiration rates reduce soil pH by increasing the leaching of basic cations [70, 77–80]. This was confirmed by the significant relationship between elevation and soil pH found in the present study (Table 2 and Fig 2). In addition, soil bulk density significantly diminished with increasing elevation (Fig 2 and Table 2). One of the main factors affecting soil bulk density is SOC content [81]. Therefore, the lowest values of soil bulk density at high elevation could be explained by the high stock of SOC, as confirmed by the negative association found between SOC stock and soil bulk density (Table 3), previously reported in other studies [9, 64, 67, 82].
Factors affecting soil respiration
The total SR observed in our study sites is within the range reported for similar forests [28, 66]. The decrease in SR observed along the elevation increase could be explained by the reduction in temperature with elevation. The temperature was the main factor controlling and explaining most of the variability of SR, in agreement with other studies performed along altitudinal gradients reporting that temperature can explain between 55% and 76% of SR variability [8, 11, 16, 29, 68, 83]. Yet, the annual cumulative SR in site B was double that of site A, despite both sites experiencing the same mean annual temperature. This finding, together with nonlinear SR correlation with elevation and temperature, suggests that other environmental factors can have a role in regulating SR [36, 66]. For example, Grand et al. [38] reported that soil and vegetation heterogeneity strongly affect soil carbon efflux in complex geomorphic terrain. The five study sites were established on the same bedrock; therefore, the high SR rate in site B could not have resulted from a confounding effect of the soil parent material. Site B is an uneven-aged dense mature forest stand, with a mean dominant tree height of approximately 29 m (Table 1). Since tree height can be used as a proxy of GPP, the high SR rate in this site could be attributed to high GPP [11, 31, 71, 84–86], which can provide substrates for root and microbial respiration through photosynthesis [71, 87], supported by a significant positive relation between SRref and dominant tree height (Fig 5 and Table 5). Above- and below-ground tree size increases with age and, therefore, the growth of root biomass leads to higher SR as forest stands develop [39, 88–90]. In this context, our findings suggest that after removing the effect of temperature, productivity and tree size are principal factors affecting SR.
At a global scale, SR has been related to SOC stock, litter production and pH, and negatively correlated with soil bulk density; therefore, a high accumulation of SOC and litter could lead to an increase in soil respiration [22, 71, 91–93]. In the present study, the highest amount of SOC stock and dry litter weight were also observed in site B. However, we did not find a significant correlation between SR and SOC stock or litter dry weight (Fig 5 and Table 5).
An increase in SR has also been observed as a consequence of increasing soil pH between 4 and 7, because of the positive effect of pH on soil microbial activity within this range [1, 75, 94–96]. Conversely, SR and bulk density are generally negatively correlated, as a low SR indicates increasing rates of SOC accumulation and therefore a decrease in bulk density [92]. Furthermore, SR declines with increasing bulk density due to the lower soil porosity and oxygen availability for microbial activity in compacted soils [20, 65, 97]. However, our analysis did not confirm a significant correlation of SR with pH or bulk density (Table 5 and Fig 5). Prediction of SR is difficult because of a range of factors such as slope angle and orientation, and soil type [37, 38, 66, 70]. In the present study, by minimizing the confounding role of these parameters, we conclude that the most important factors controlling SR along an Alpine altitude gradient were temperature and dominant tree height. Therefore, a tall adult or mature forest stand with high productivity or GPP, and even more so a more complex and uneven-aged structured forest able to maintain high biomass levels over time such as site B, can significantly affect SR, suggesting that tree size and stand biomass are more relevant than stand age itself.
Temperature sensitivity of soil respiration (Q10)
The temperature sensitivity of SR is an important ecological model parameter, and according to previous studies it is mainly controlled by temperature [16, 19, 98, 99]. The Q10 and SRref values found at site B (2.0 and 4.09, respectively) confirm those found at the same site by Acosta et al. [100]. Although at a smaller spatial scale, Acosta et al. [100] also found an increasing SRref to be a function of stand age (and consequently height). The significant trend of Q10 with elevation in our study confirms previous results, highlighting the higher sensitivity of high elevation ecosystems to global warming [16, 101–104]. However, the only significant difference was found between the Q10 value at site A (higher elevation) and the other sites (Table 4). The Q10 value in site A was also significantly higher than site B, which is characterized by a similar mean annual temperature. Previous studies have found that Q10 is negatively related to pH and positively related to SOC stock [61, 105]. However, in our study SOC stock and pH could not be the cause of the lower Q10 value found in site B which, compared with the other sites, is characterized by a lower pH value and a similar SOC stock content. The temperature sensitivity of SR can also be affected by forest structure [106]. Dense forest stands with a closed canopy can create a specific understory microclimate by providing a cool shelter during heat waves, which can decrease daily maximum air temperature by up to 5.1°C [107, 108]. Therefore, we can hypothesize that, since temperature range is positively linked with ecosystem respiration [109], the dense forest stand in site B, which has the highest dominant tree height, is less sensitive to global warming. This confirms what found by Niu et al. [110] in the same site and may explain the significant negative correlation between Q10 and mean dominant tree height (Table 3). Our study suggests that a close-to natural, continuous cover forestry system with a significant permanent presence of bigger individuals, particularly on sites with good forest productivity, can not only maintain current C stock in the biomass but also lead to a reduced sensitivity to temperature of SOC stock.
Role of stand structural properties
While the correlation between forest productivity and respiration is well known [11], this research opens a new perspective regarding the possible link between stand structural parameters and temperature sensitivity of respiration in forest soils. In a mature forest several characteristics, differing from those of a younger forest, may influence soil respiration. While stand age is the most obvious, it does not appear to be tightly linked to soil respiration [39], and our research corroborates this. Forest stature can also affect forest soil microclimate; intact forests show a reduced temperature above the canopy [111, 112] and current studies are evaluating the impact of vegetation structural properties on soil temperature [113]. In addition, there is evidence of a positive trend between forest development and efficiency in the enhancement of soil C storage, most likely related to the presence of microbial communities and plant roots more efficient in SOC storage [114].
Among the other variables that may affect SR that should be evaluated there is the overstory LAI. A significant positive correlation between LAI and SR was reported by Migliavacca et al. [39]. This positive relationship can be explained by the relation between LAI and productivity. However, a high LAI can also intercept direct radiation, reducing thermal fluctuations at the soil level and hence SR sensitivity to temperature [102, 115]. The mature stand in site B has not only a high overstory LAI, but also a relevant presence of understory [116] able to intercept direct radiation passing through the uneven-aged canopy. The role of litter quality, and particularly of N content, is still debated. Long-standing leaves present in the mature stand (site B), which are depleted in N, may have (or not) a longer degradation time [117, 118]. However, only a more detailed experimental design would allow us to determine which of these different features of mature vs. young forest affect SR sensitivity to temperature.
Conclusions
In this study, a significant nonlinear relationship between SR, SOC, Q10, and elevation was detected along the examined Alpine altitudinal gradient, rejecting our initial hypothesis of linearity. Our data confirmed a negative trend between SR and elevation, but a positive trend of SOC and Q10 with elevation. We, therefore, conclude that temperature is the major factor controlling annual SR, Q10, and SOC, but its regulating role may be strongly affected by site biological characteristics, particularly GPP or vegetation type/composition. The high Q10 value detected at high elevations confirmed a greater potential vulnerability of high mountain ecosystems to climate change, where small temperature changes can induce a stronger increase in CO2 emissions. However, the site with the tallest dominant height and more complex structure showed a high SRref and moderate Q10, indicating that maintaining a forest close to an old growth system, with a heterogenous and uneven-age structure, can reduce, to some extent, the effects of climate change on ecosystems and decrease the positive feedback due to soil CO2 emissions to the atmosphere.
Supporting information
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(DOCX)
(TIF)
(TIF)
Data Availability
All relevant data are within the manuscript and its Supporting Information files. Additional data can be made available upon journal request.
Funding Statement
This project was funded by the internal resources of the Faculty of Science and Technology, Free University of Bolzano, Project CliMicrobe, RTD call 2017 (grant number CUP I52F17000560005). The Open Access Publishing Fund of the Free University of Bozen-Bolzano paid the article publication fee.
References
- 1.Luo Y, Zhou X. Soil respiration and the environment. Sandiego: Academic Press: 2006. [Google Scholar]
- 2.Schlesinger WH. Carbon balance in terrestrial detritus. Ann Rev Ecol Systematics 1977; 8: 51–81. [Google Scholar]
- 3.Raich JW, Potter CS. Global patterns of carbon dioxide emissions from soils. Global Biogeochem Cycles. 1995; 9: 23–26. [Google Scholar]
- 4.Valentini R, Matteucci G, Dolman AJ, Schulze ED, Rebmann C, Moors EJ, et al. Respiration as the main determinant of carbon balance in European forests. Nature. 2000; 404: 861–865. doi: 10.1038/35009084 [DOI] [PubMed] [Google Scholar]
- 5.Scharlemann JP, Tanner EV, Hiederer R, Kapos V. Global soil carbon: understanding and managing the largest terrestrial carbon pool. Carbon Manag. 2014; 5: 81–91. [Google Scholar]
- 6.Makita N, Kosugi Y, Sakabe A, Kanazawa A, Ohkubo S, Tani M. Seasonal and diurnal patterns of soil respiration in an evergreen coniferous forest: Evidence from six years of observation with automatic chambers. PLoS ONE. 2018; 13(2): e0192622. 10.1371/journal.pone.0192622 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cong WF, Van Ruijven J, Mommer L, De Deyn GB, Berendse F, Hoffland E. Plant species richness promotes soil carbon and nitrogen stocks in grasslands without legumes. Journal of Ecology. 2014; 102(5): 1163–1170. doi: 10.1111/1365-2745.12280 [DOI] [Google Scholar]
- 8.Luo W, Li MH, Sardans J, Lü XT, Wang C, Peñuelas J, et al. Carbon and nitrogen allocation shifts in plants and soils along aridity and fertility gradients in grasslands of China. Ecol Evol. 2017; 7(17): 6927–6934. doi: 10.1002/ece3.3245 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Devi SB, Sherpa SSSS. Soil carbon and nitrogen stocks along the altitudinal gradient of the Darjeeling Himalayas, India. Environ Monit Assess. 2019; 191 (361). 10.1007/s10661-019-7470-8 [DOI] [PubMed] [Google Scholar]
- 10.Montagnani L, Badraghi A, Speak AF, Wellstein C, Borruso L, Zerbe S, et al. Evidence for a non-linear carbon accumulation pattern along an Alpine glacier retreat chronosequence in Northern Italy. PeerJ. 2019; 7:e7703, doi: 10.7717/peerj.7703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Janssens I.A, Lankreijer H, Matteucci G, Kowalski AS, Buchmann N, Epron D. Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Glob Chang Biol. 2001; 7: 269–278. [Google Scholar]
- 12.Hopkins F, Gonzalez-Meler MA, Flower CE, Lynch DJ, Czimczik C, Tang J, et al. Ecosystem-level controls on root-rhizosphere respiration, New Phytol. 2013; 199: 339e351. 10.1111/nph.12271 [DOI] [PubMed] [Google Scholar]
- 13.Scandellari F, Zanotelli D, Ceccon C, Bolognesi M, Montagnani L, Cassol P, et al. Enhancing prediction accuracy of soil respiration in an apple orchard by integrating photosynthetic activity into a temperature-related model, Eur J Soil Biol. 2015; 70: 77–87. doi: 10.1016/j.ejsobi.2015.07.006 [DOI] [Google Scholar]
- 14.Zhang ZS, Dong XJ, Xu BX, Chen YL, Zhao Y, Gao YH, et al. Soil respiration sensitivities to water and temperature in a revegetated desert. J. Geophys. Res. Biogeosci. 2015; 120: 773–787. doi: 10.1002/2014JG002805 [DOI] [Google Scholar]
- 15.Chen D, Yu M, González G, Zou X, Gao Q. Climate Impacts on Soil Carbon Processes along an Elevation Gradient in the Tropical Luquillo Experimental Forest. Forests. 2017; 90: doi: 10.3390/f8030090 [DOI] [Google Scholar]
- 16.Ma M, Zang Z, Xie Z, Chen Q, Xu W, Zhao C, et al. Soil respiration of four forests along elevation gradient in northern subtropical China. Ecol Evol. 2019; 9: 12846–12857. doi: 10.1002/ece3.5762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Reichstein M, Beer C. Soil respiration across scales: the importance of a model-data integration framework for data interpretation. J Plant Nutr Soil Sci. 2008; 171: 344e354. [Google Scholar]
- 18.Subke JA, Bahn M. On the ‘temperature sensitivity’ of soil respiration: Can we use the immeasurable to predict the unknown? Soil Biol Biochem. 2008; 42 (9): 1653–1656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lloyd J, Taylor JA. On the temperature dependence of soil respiration, Funct Ecol. 1994; 8(3): 315–323. doi: 10.2307/2389824 [DOI] [Google Scholar]
- 20.Xu M, Qi Y. Soil surface CO2 efflux and its spatial and temporal variation in a young ponderosa pine plantation in California. Global Change Biol. 2001; 7: 667–677. 10.1046/j.1354-1013.2001.00435.x [DOI] [Google Scholar]
- 21.Janssens IA, Dore S, Epron D, Lankreijer H, Buchmann N, Longdoz B, et al. Climatic influences on seasonal and spatial differences in soil CO2 efflux. In: Valentini R (ed) Fluxes of carbon, water and energy of European forests. Berlin: Springer; 2003. p. 235–255. [Google Scholar]
- 22.Reichstein M, Rey A, Freibauer A, Tenhunen J, Valentini R, Banza R. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Global Biogeochem Cycles. 2003; 17: doi: 10.1029/2003GB002035 [DOI] [Google Scholar]
- 23.Van’t Hoff JH. Lectures on theoretical and physical chemistry. In Chemical Dynamics London: Edward Arnold; 1898. [Google Scholar]
- 24.Lomolino MV. Elevation gradients of species-density: historical and prospective views. Glob Ecol Biogeogr. 2001; 10: 3–13. [Google Scholar]
- 25.Prietzel J, Zimmermann L, Schubert A, Christophel D. Organic matter losses in German Alps forest soils since the 1970s most likely caused by warming. Nat Geosci. 2016; 1–8, doi: 10.1038/NGEO2732 [DOI] [Google Scholar]
- 26.Shedayi AA, Xu M, Naseer L, Khan B. Altitudinal gradients of soil and vegetation carbon and nitrogen in a high altitude nature reserve of Karakoram ranges. SpringerPlus. 2016; 5: 320. doi: 10.1186/s40064-016-1935-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jiang L, He Z, Liu J, Xing C, Gu X, Wei C, et al. Elevation Gradient Altered Soil C, N, and P Stoichiometry of Pinus taiwanensis Forest on Daiyun Mountain. Forests. 2019; 10: 1089. doi: 10.3390/f10121089 [DOI] [Google Scholar]
- 28.Rodeghiero M, Cescatti A. Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps. Glob Chang Biol. 2005; 11: 1024–1041. doi: 10.1111/j.1365-2486.2005.00963.x [DOI] [Google Scholar]
- 29.Shi Z, Wang JS, He R, Fang YH, Xu ZK, Quan W, et al. Soil respiration and its regulating factor along an elevation gradient in Wuyi Mountain of Southeast China. Chinese J Ecol. 2008; 27 (4): 563–568. [Google Scholar]
- 30.Luo S, Liu G, Li Z, Hu C, Gong L, Wang M, et al. Soil respiration along an altitudinal gradient in a subalpine secondary forest in China. iForest. 2014; 8: 526–532. [Google Scholar]
- 31.Kane ES, Valentine DW, Schuur EAG, Dutta K. Soil carbon stabilization along climate and stand productivity gradients in black spruce forests of interior Alaska. Can J For Res. 2005; 35: 2118–2129. [Google Scholar]
- 32.Djukic L, Zehetner F, Tatzber M, Gerzabek MH. Soil organic-matter stocks and characteristics along an Alpine elevation gradient. J Plant Nutr Soil Sci. 2010; 173: 30–38. doi: 10.1002/jpln.200900027 [DOI] [Google Scholar]
- 33.Kunkel ML, Flores AN, Smith TJ, McNamara JP, Benner SG. A simplified approach for estimating soil carbon and nitrogen stocks in semi-arid complex terrain. Geoderma. 2011; 165 (1): 1–11. [Google Scholar]
- 34.Tian Q, He H, Cheng W, Bai Z, Wang Y, Zhan X. Factors controlling soil organic carbon stability along a temperate forest altitudinal gradient. Sci Rep. 2016; 6: 18783. doi: 10.1038/srep18783 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Garcia-Pausas J, Casals P, Camarero L, Huguet C, Sebastia MT, Thompson R, et al. Soil organic carbon storage in mountain grasslands of the Pyre-nees, effects of climate and topography. Biogeochemistry. 2007; 82: 279–289. 10.1007/s10533-007-9071-9. [DOI] [Google Scholar]
- 36.Zimmermann M, Meir P, Bird MI, Malhi Y, Ccahuana AJQ. Temporal variation and climate dependence of soil respiration and its components along a 3000 m altitudinal tropical forest gradient. Global Biogeochem Cy. 2010; 24: GB4012. doi: 10.1029/2010GB003787 [DOI] [Google Scholar]
- 37.Kobler J, Zehetgruber B, Jandl R, Dirnböck T, Schindlbacher A. Effects of slope aspect and site elevation on seasonal soil carbon dynamics in a forest catchment in the Austrian Limestone Alps. 19th EGU General Assembly (EGU2017); 2017 April 23–28; Vienna, Austria. 2017.
- 38.Grand S, Rubin A, Verrecchia EP, Vittoz P. Variation in Soil Respiration across Soil and Vegetation Types in an Alpine Valley. PLoS ONE. 2016; 11 (9): e0163968. 10.1371/journal.pone.0163968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Migliavacca M, Reichstein M, Richardson AD, Colombo R, Sutton MA, Lasslop G. Semiempirical modeling of abiotic and biotic factors controlling ecosystem respiration across eddy covariance sites. Global Change Biology. 2011; 17(1): 390–409. doi: 10.1111/j.1365-2486.2010.02243.x [DOI] [Google Scholar]
- 40.Bréchet L, Ponton S, Alméras T, Bonal D, Epron D. Does spatial distribution of tree size account for spatial variation in soil respiration in a tropical forest?. Plant Soil. 2011; 347 (293): 10.1007/s11104-011-0848-1 [DOI] [Google Scholar]
- 41.Xu X, Shi Z, Li D, Zhou X, Sherry RA, Luo Y. Plant community structure regulates responses of prairie soil respiration to decadal experimental warming. Glob Change Biol. 2015; 21: 3846–3853. 10.1111/gcb.12940 [DOI] [PubMed] [Google Scholar]
- 42.Tian Q, Wang D, Tang Y, Li Y, Wang M, Liao C, et al. Topographic controls on the variability of soil respiration in a humid subtropical forest. Biogeochemistry. 2019; 145: 177–192. 10.1007/s10533-019-00598-x [DOI] [Google Scholar]
- 43.Tonon G, Dezi S, Ventura M, Scandellari F. The Effect of Forest Management on Soil Organic Carbon. In: Sauer TJ, Eiler JM, Sivakumar MVK (eds) Sustaining Soil Productivity in Response to Global Climate Change: Science, Policy, and Ethics. John Wiley & Sons, Inc.; 2011. p. 225–238.
- 44.Greiser C, Meineri E, Luoto M, Ehrlén J, Hylander K. Monthly microclimate models in a managed boreal forest landscape. Agri For Meteorol. 2018; 250–251: 147–158. 10.1016/j.agrformet.2017.12.252. [DOI] [Google Scholar]
- 45.Carey JC, Tang J, Templer PH, Kroeger KD, Crowther T, Burton AJ, et al. Temperature response of soil respiration largely unaltered with experimental warming. PNAS. 2016; 113 (48): 13797–13802. doi: 10.1073/pnas.1605365113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tang J, Cheng H, Fang C. The temperature sensitivity of soil organic carbon decomposition is not related to labile and recalcitrant carbon. PLoS ONE. 2017; 12 (11): e0186675. 10.1371/journal.pone.0186675 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wehren B, Weingartner R, Schädler B, Viviroli D. General Characteristics of Alpine Waters. In: Bundi U. (eds) Alpine Waters. The Handbook of Environmental Chemistry. Heidelberg, Berlin: Springer; 2010. 10.1007/978-3-540-88275-6_2 [DOI]
- 48.Montagnani L, Manca G, Canepa E, Georgieva E, Acosta M, Feigenwinter C, et al. A new mass conservation approach to the study of CO2 advection in an alpine forest. J. Geophys. Res. Atmos. 2009; 114: D07306, doi: 10.1029/2008JD010650 [DOI] [Google Scholar]
- 49.Xu X, Yi C, Montagnani L, Kutter E. Numerical Study of the Interplay between Thermo-topographic Slope Flow and Synoptic Flow on Canopy Transport Processes, Agric For Meteorol. 2018; 255: 3–16. 10.1016/j.agrformet.2017.03.004. [DOI] [Google Scholar]
- 50.Gielen B, Acosta M, Altimir N, Buchmann N, Cescatti A, Ceschia E, et al. Soil-meteorological measurements at ICOS monitoring stations in terrestrial ecosystems. Int Agrophys. 2019; 32: 645–664. doi: 10.1515/intag-2017-0048 [DOI] [Google Scholar]
- 51.Richards FJ. A flexible growth function for empirical use. J Exp Bot. 1959; 10: 290–300. [Google Scholar]
- 52.Janssens IA, Pilegaard K. Large seasonal changes in Q10 of soil respiration in a beech forest. Glob Chang Biol. 2003; 9: 911–918. [Google Scholar]
- 53.Ventura M, Panzacchi P, Muzzi E, Magnani F, Tonon G. Carbon balance and soil carbon input in a poplar short rotation coppice plantation as affected by nitrogen and wood ash application. New Forests. 2019. 10.1007/s11056-019-09709-w [DOI] [Google Scholar]
- 54.Cook ER, Kairiukstis LA. Methods of dendrochronology: applications in the environmental sciences. Dordrecht: Kluwer Academic Publishers; 1990. [Google Scholar]
- 55.R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2019. Available online at https://www.R-project.org/.
- 56.Zuur A, Ieno EN, Walker N, Saveliev AA, Smith GM. Mixed Effects Models and Extensions in Ecology with R. New York: Springer; 2009. [Google Scholar]
- 57.Bates D, Maechler M, Bolker B, Walker S. lme4: linear mixed-effects models using Eigen and S4. R package version 1.1–7; 2014.
- 58.Crawley M.J. The R Book. Chichester: John Wiley & Sons, Ltd.; 2007.
- 59.Nakagawa S, Holger S. A general and simple method for obtaining R2 from generalized linear mixed effects models. Methods Ecol Evol. 2013; 4: 133–142. [Google Scholar]
- 60.Jaeger BC, Edwards LJ, Das K, Sen PK. An R2 Squared Statistic for Fixed Effects in the Generalized Linear Mixed Model. J Appl Stat. 2016; 44 (6): 1086–1105 [Google Scholar]
- 61.Meyer N, Welp G, Amelung W. The temperature sensitivity (Q10) of soil respiration: Controlling factors and spatial prediction at regional scale based on environmental soil classes. Global Biogeochem Cy. 2018; 32: 306–323. 10.1002/2017GB005644 [DOI] [Google Scholar]
- 62.Shapiro SS, Wilk MB. Analysis of variance test for normality. Biometrika. 1965; 52: 591–611. [Google Scholar]
- 63.Levene H. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling, I. Olkin et al. eds., Standford: Stanford University Press; 1960. p. 278–292.
- 64.Tsozué D, Nghonda JP, Tematio P, Basga SD. Changes in soil properties and soil organic carbon stocks along an elevation gradient at Mount Bambouto, Central Africa. Catena. 2019; 175: 251–262. 10.1016/j.catena.2018.12.028 [DOI] [Google Scholar]
- 65.Wang G, Zhou Y, Xu X, Ruan H, Wang J. Temperature Sensitivity of Soil Organic Carbon Mineralization along an Elevation Gradient in the Wuyi Mountains, China. PLoS ONE. 2013; 8(1): e53914. doi: 10.1371/journal.pone.0053914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Chatterjee A, Jenerette GD. Variation in soil organic matter accumulation and metabolic activity along an elevation gradient in the Santa Rosa Mountains of Southern California, USA. J Arid Land. 2015; 7(6): 814–819. doi: 10.1007/s40333-015-0085-1 [DOI] [Google Scholar]
- 67.de la Cruz-Amo L, Bañares-de-Dios G, Cala V, Granzow-de la Cerda I, Espinosa CI, Ledo A, et al. Trade-offs among aboveground, belowground, and soil organic carbon stocks along altitudinal gradients in Andean Tropical Montane Forests. Front. Plant Sci. 2020; 11:106. doi: 10.3389/fpls.2020.00106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.He X, Hou E, Liu Y, Wen D. Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometryin subtropical China. Sci Rep. 2016; 6: 24261. doi: 10.1038/srep24261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kirschbaum MUF. The temperature dependence of organic-matter decomposition-still a topic of debate. Soil Biol Biochem. 2006; 38: 2510–2518 [Google Scholar]
- 70.Griffiths PR, Madritch MD, Swanson AK. The effects of topography on forest soil characteristics in the Oregon Cascade Mountains (USA): Implications for the effects of climate change on soil properties. For Ecol Manag. 2009; 257(1): 1–7. [Google Scholar]
- 71.Bahn M, Rodeghiero M, Anderson-Dunn M, Dore S, Gimeno C, Drösler M, et al. Soil respiration in European grasslands in relation to climate and assimilate supply. Ecosystems. 2008; 11: 1352–1367. doi: 10.1007/s10021-008-9198-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Shi Y, Baumann F, Ma Y, Song C, Uhn PK, Scholten T, He JS. Organic and inorganic carbon in the topsoil of the Mongolian and Tibetan grasslands: pattern, control and implications. Biogeosciences. 2012; 9: 2287–2299. doi: 10.5194/bg-9-2287-2012 [DOI] [Google Scholar]
- 73.Cao J, Li MT, Deo RC, Adamowskiet JF, Cerdà D, Feng Q, et al. Comparison of socialecological resilience between two grassland management patterns driven by grassland land contract policy in the Maqu, Qinghai-Tibetan Plateau. Land Use Policy. 2018a; 74: 88–96. 10.1016/j.landusepol.2017.07.027. [DOI] [Google Scholar]
- 74.Cao J, Zhang X, Deo R, Gong Y, Feng Q. Influence of stand type and stand age on soil carbon storage in China’s arid and semi-arid regions. Land Use Policy. 2018b; 78: 258–265, 10.1016/j.landusepol.2018.07.002 [DOI] [Google Scholar]
- 75.Vanhala P. Seasonal variation in the soil respiration rate in coniferous forest soils. Soil Biol Biochem. 2002; 34: 1375–1379. [Google Scholar]
- 76.Zhang YY, Wu W, Liu H. Factors affecting variations of soil pH in different horizons in hilly regions. PLoS One. 2019; 14(6): e0218563. doi: 10.1371/journal.pone.0218563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Martí C, Badía D. Characterization and classification of soils along two altitudinal transects in the Eastern Pyrenees, Spain. Arid Soil Res Rehabil. 1995; 9: 367–383. doi: 10.1080/15324989509385905 [DOI] [Google Scholar]
- 78.Smith JL, Halvorson JJ, Jr HB. Soil properties and microbial activity across a 500 m elevation gradient in a semi-arid environment. Soil Biol Biochem. 2002; 34: 1749–1757. [Google Scholar]
- 79.Seibert J, Stendahl J, Sørensen R. Topographical influences on soil properties in boreal forests. Geoderma. 2007; 141(1–2): 139–148. [Google Scholar]
- 80.Badía D, Ruiz A, Girona A, Martí C, Casanova J, Ibarra P, Zufiaurre R. The influence of elevation on soil properties and forest litter in the Siliceous Moncayo Massif, SW Europe. J Mt Sci. 2016; 13: 2155–2169. 10.1007/s11629-015-3773-6 [DOI] [Google Scholar]
- 81.Klopfenstein ST, Hirmas DR, Johnson W. Relationships between soil organic carbon and precipitation along a climosequence in loess-derived soils of the Central Great Plains, USA. Catena. 2015; 133: 25–34. 10.1016/j.catena.2015.04.015 [DOI] [Google Scholar]
- 82.Schrumpf M, Schulze ED, Kaiser K, Schumacher, J. How accurately can soil organic carbon stocks and stock changes be quantified by soil inventories?. Biogeosciences. 2011; 8: 1193–1212. 10.5194/bg-8-1193-2011. [DOI] [Google Scholar]
- 83.Keenan TF, Migliavacca M, Papale D, Baldocchi D, Reichstein M, Torn M, et al. Widespread inhibition of daytime ecosystem respiration. Nat Ecol Evol. 2019; 3(3): 407–415. doi: 10.1038/s41559-019-0809-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Reichstein M, Ciais P, Papale D, Valentini R, Running S, Viovy N. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: a joint flux tower, remote sensing and modelling analysis. Glob Change Biol. 2007; 13(3): 634–651. doi: 10.1111/j.1365-2486.2006.01224.x [DOI] [Google Scholar]
- 85.Yu GR, Zhu XJ, Fu YL, He HL, Wang QF, Wen XF. Spatial patterns and climate drivers of carbon fluxes in terrestrial ecosystems of China Global Change Bio. 2013; l19: 798–810. 10.1111/gcb.12079 [DOI] [PubMed] [Google Scholar]
- 86.Chen S, Zou J, Hu Z, Lu Y. Climate and vegetation drivers of terrestrial carbon fluxes: a global data synthesis. Adv Atmos Sci. 2019; 36: 679–696. [Google Scholar]
- 87.Ma J, Liu R, Li C, Fan L, Xu G, Li Y. Herbaceous layer determines the relationship between soil respiration and photosynthesis in a shrub-dominated desert plant community. Plant Soil. 2020, 449: 193–207. 10.1007/s11104-020-04484-6 [DOI] [Google Scholar]
- 88.Powers M, Kolka R, Bradford J, Palik B, Jurgensen M. Forest floor and mineral soil respiration rates in a northern Minnesota red pine chronosequence. Forests. 2018; 9 (1): 16. 10.3390/f9010016 [DOI] [Google Scholar]
- 89.Xiao W, Ge X, Zeng L, Huang Z, Lei J, Zhou B, et al. Rates of litter decomposition and soil respiration in relation to soil temperature and water in differentaged Pinus massoniana forests in the three gorges reservoir area, China. PLoS One. 2014; 9 (7): e101890. doi: 10.1371/journal.pone.0101890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yu K, Yaoa X, Denga Y, Laia Z, Lina L, Liu J. Effects of stand age on soil respiration in Pinus massoniana plantations in the hilly red soil region of Southern China. Catena. 2019; 178: 313–321. 10.1016/j.catena.2019.03.038 [DOI] [Google Scholar]
- 91.Raich JW, Tufekciogul A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry. 2000; 48: 71–90. 10.1023/A:1006112000616 [DOI] [Google Scholar]
- 92.Chen Q, Wang Q, Han X, Wan S, Li L. Temporal and spatial variability and controls of soil respiration in a temperate steppe in northern China. Global Biogeochem Cycles. 2010; 24: GB2010. doi: 10.1029/2009GB003538 [DOI] [Google Scholar]
- 93.Oertel C, Matschullat J, Zurba K, Zimmermann F, Erasmi S. Greenhouse Gas Emissions From Soil—A review, Chemie der Erde. 2016; 76: 327–352, doi: 10.1016/j.chemer.2016.04.002 [DOI] [Google Scholar]
- 94.Andersson S, Nilsson SI. Influence of pH and temperature on microbial activity, substrate availability of soil-solution bacteria and leaching of dissolved organic carbon in a mor humus. Soil Biol Biochem. 2001; 33 (9): 1181–1191. 10.1016/S0038-0717(01)00022-0 [DOI] [Google Scholar]
- 95.Reth S, Reichstein M, Falge E. The effect of soil water content, soil Temperature, soil pH value and the root mass on soil CO2 efflux- a modified model. Plant Soil. 2005; 268: 21–33. [Google Scholar]
- 96.Chappell C, Johnson A. Influence of pH and bulk density on carbon dioxide efflux in tree urban wetland types. Professional Agricultural Workers Journal. 2015; 3 (1, 5). htp://tuspubs.tuskegee.edu/pawj/vol3/iss1/5 [Google Scholar]
- 97.Mordhorst A, Peth S, Horn R. Influence of mechanical loading on static and dynamic CO2 efflux on differently textured and managed Luvisols. Geoderma. 2014; 219–220: 1–13. [Google Scholar]
- 98.Chen B, Liu S, Ge J, Chu J. Annual and seasonal variations of Q10 soil respiration in the sub-alpine forests of the Eastern Qinghai-Tibet Plateau, China. Soil Biol Biochem. 2010; 42: 1735–1742 [Google Scholar]
- 99.Feng J, Wang J, Song Y, Zhu B. Patterns of soil respiration and its temperature sensitivity in grassland ecosystems across China, Biogeosciences. 2018; 15: 5329–5341, 10.5194/bg-15-5329-2018 [DOI] [Google Scholar]
- 100.Acosta M, Pavelka M, Montagnani L, Kutsch W, Lindroth A, Juszczak R, et al. Soil surface CO2 efflux measurements in Norway spruce forests. Comparison between four different sites acrossEurope—from boreal to alpine forest. Geoderma. 2013; 192: 295–303. doi: 10.1016/j.geoderma.2012.08.027 [DOI] [Google Scholar]
- 101.Zhou T, Shi P, Hui D, Luo Y. Global pattern of temperature sensitivity of soil heterotrophic respiration (Q10) and its implications for carbon-climate feedback, J Geophys Res-Biogeo. 2009; 114: 271–274, 10.1029/2008JG000850, 2009. [DOI] [Google Scholar]
- 102.Song X, Peng C, Zhao Z, Zhang Z, Guo B, Wang W, et al. Quantification of soil respiration in forest ecosystems across China. Atmos Environ. 2014; 94: 546–551, 10.1016/j.atmosenv.2014.05.071 [DOI] [Google Scholar]
- 103.Zhao J, Li R, Li X, Tian L. Environmental controls on soil respiration in alpine meadow along a large altitudinal gradient on the central Tibetan Plateau. Catena. 2017; 159: 84–92. 10.1016/j.catena.2017.08.007 [DOI] [Google Scholar]
- 104.Zhou W, Hui D, Shen W. Effects of Soil Moisture on the Temperature Sensitivity of Soil Heterotrophic Respiration: A Laboratory Incubation Study. PLoS ONE. 2014; 9(3): e92531. doi: 10.1371/journal.pone.0092531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Zhou Z, Guo C, Meng H. Temperature sensitivity and basal rate of soil respiration and their determinants in temperate forests of North China. PLoS One. 2013; 8(12): e81793. 10.1371/journal.pone.0081793 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Quan Q, Wang C, He N, Zhang Z, Wen X, Su H, et al. Forest type affects the coupled relationships of soil C and N mineralization in the temperate forests of northern China. Sci Rep. 2014; 4: 6584, doi: 10.1038/srep06584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Rambo TR, North MP. Canopy microclimate response to pattern and density of thinning in a Sierra Nevada forest. For Ecol Manag. 2009; 257 (2): 435–442, 10.1016/j.foreco.2008.09.029 [DOI] [Google Scholar]
- 108.Arx GV, Dobbertin M, Rebetez M. Spatio-temporal effects of forest canopy on understory microclimate in along-term experiment in Switzerland. Agr Forest Meteorol. 2012; 166–167: 144–155, 10.1016/j.agrformet.2012.07.018 [DOI] [Google Scholar]
- 109.Song B; Niu S; Luo R; Luo Y; Chen J, et al. Divergent apparent temperature sensitivity of terrestrial ecosystem respiration. Journal of Plant Ecology 7(5):419–428, 10.1093/jpe/rtu014. [DOI] [Google Scholar]
- 110.Niu S, Luo Y, Fei S, Yuan W, Schimel D, Law BE, et al. Thermal Optimality of Net Ecosystem Exchange of Carbon Dioxide and Underlying Mechanisms. New Phytol. 2012; 194 (3): 775–783, doi: 10.1111/j.1469-8137.2012.04095.x [DOI] [PubMed] [Google Scholar]
- 111.Alkama R, Cescatti A. Biophysical climate impacts of recent changes in global forest cover, Science. 2016Feb5; 351(6273): 600–4. doi: 10.1126/science.aac8083 [DOI] [PubMed] [Google Scholar]
- 112.Lin H, Chen Y, Song Q, Fua P, Cleverly J, Magliulo V, et al. Quantifying deforestation and forest degradation with thermal response. Science of the Total Environment. 2017; 607–608: 1286–1292. 10.1016/j.scitotenv.2017.07.062 [DOI] [PubMed] [Google Scholar]
- 113.Lembrecht JJ, Hoogen JVD, Aalto J, Ashcroft MB, Frenne PD, Kemppinen J, et al. Mismatches between soil and air temperature. 2021; doi: 10.32942/osf.io/pksqw [DOI] [Google Scholar]
- 114.Huang W, Han T, Liu J, Wang G, and Zhou G. Changes in soil respiration components and their specific respiration along three successional forests in the subtropics. Funct Ecol. 2016; 30: 1466–1474. 10.1111/1365-2435.12624 [DOI] [Google Scholar]
- 115.Tanaka K, Hashimoto S, Plant canopy effects on soil thermal and hydrological properties and soil respiration, Ecological Modelling, 2006; 196(2006)32–44, doi: 10.1016/j.ecolmodel.2006.01.004 [DOI] [Google Scholar]
- 116.Pisek J, Erb A, Korhonen L, Biermann T, Carrara A, Cremonese E, et al. Retrieval and validation of forest background reflectivity from daily MODIS bidirectional reflectance distribution function (BRDF) data across European forests. Biogeosciences. 2021; 18 (2): 621–635. [doi: 10.5194/bg-18-621-2021] [DOI] [Google Scholar]
- 117.Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol Lett. 2008; 11: 1065–1071. doi: 10.1111/j.1461-0248.2008.01219.x [DOI] [PubMed] [Google Scholar]
- 118.Zhou G, Zhang J, Qiu X, Wei F, Xu X. Decomposing litter and associated microbial activity responses to nitrogen deposition in two subtropical forests containing nitrogen-fixing or non-nitrogen-fixing tree species. Sci Rep. 2018; 8: 12934. 10.1038/s41598-018-30666-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(DOCX)
(TIF)
(TIF)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files. Additional data can be made available upon journal request.