

Abstract
BACKGROUND
Gut microbial dysbiosis contributes to the development and progression of colorectal cancer (CRC). Natural killer (NK) cells are involved in early defense mechanisms to kill infective pathogens and tumor cells by releasing chemokines and cytokines. To better understand the relationship between the gut microbiome and CRC, it was hypothesized here that a high abundance of Fusobacterium nucleatum (F. nucleatum) in the gastrointestinal tract could cause reduced NK cell activity.
AIM
To identify associations between gastrointestinal tract F. nucleatum levels and NK cell activity.
METHODS
In vitro experiments were performed on NK cells treated with F. nucleatum, Peptostreptococcus anaerobius, and Parvimonas micra to identify the effects of gut microbiome species on NK cells. Following 24 and 48 h of treatment, NK cell counts were measured. In parallel studies, C57BL/6 mice were given broad-spectrum antibiotics in their drinking water to reduce resident gut flora. After 3 wk, the mice received the various bacterial species or phosphate-buffered saline (PBS) via oral gavage every 2 d for 6 wk. At the study end, blood samples were acquired to perform NK cell activity assessment and cytokine analysis. Intestinal tissues were collected and analyzed via immunohistochemistry (IHC).
RESULTS
The data show that after 3 wk of broad-spectrum antibiotic treatment, levels of total bacteria and F. nucleatum were markedly decreased in mice. Gavage of F. nucleatum significantly decreased NK cell activity relative to the activities of cells from mice treated with antibiotics only and PBS. The administration of F. nucleatum decreased the proportion of NK46+ cells based on IHC staining and increased the production of interleukin-1β and tumor necrosis factor-α.
CONCLUSION
High levels of F. nucleatum in the gastrointestinal tract reduced NK cell activity in mice, and the decrease in NK cell activity might be affected by increased pro-inflammatory cytokines after F. nucleatum treatment.
Keywords: Fusobacterium nucleatum, Natural killer cells, Colorectal cancer
Core Tip: Gut microbial dysbiosis contributes to the development and progression of colorectal cancer (CRC). Natural killer (NK) cells are involved in early defense mechanisms to kill infective pathogens and tumor cells by releasing chemokines and cytokines. To better understand the relationship between the gut microbiome and CRC, it was hypothesized here that a high abundance of Fusobacterium nucleatum (F. nucleatum) in the gastrointestinal tract could cause reduced NK cell activity. Accordingly, this association would impact the development of CRC in a host. To test this hypothesis, associations between the abundance of F. nucleatum in the murine gastrointestinal tract and NK cell activity were evaluated. In this study, we showed that a high abundance of F. nucleatum in the gastrointestinal tract reduced the activity of NK cells and that such outcomes might be associated with a concurrent increase in circulating levels of some pro-inflammatory cytokines, including interleukin-1β and tumor necrosis factor-α in these hosts. These findings suggest that increases in F. nucleatum in the gastrointestinal tract could be a factor in disrupting how the immune system prevents the development of CRC.
INTRODUCTION
Increasing evidence indicates that gut microbial dysbiosis contributes to the development and progression of colorectal cancer (CRC)[1-3]. Studies have suggested that the presence of several bacteria, including Fusobacterium nucleatum (F. nucleatum), Peptostreptococcus anaerobius (P. anaerobius), Parvimonas micra (P. micra), and Solobacterium moorei are significantly associated with CRC[4,5]. Other studies have described a strong correlation between the presence of F. nucleatum (a common oral anaerobic Gram-negative rod that is a typical cause of periodontal disease) with CRC development and pathogenicity[6-8]. The presence of F. nucleatum increases the proliferation of CRC cells and tumor development in mice by activating Toll-like receptor 4 signaling to nuclear factor-kappa B, and up-regulating the expression of miRNA-21. Other studies have shown that F. nucleatum can suppresses the tumor-immune microenvironment in a manner that could inhibit T-cell responses to antigens and arrest human T-cells in the G1 phase or induce apoptotic cell death[9-11]. In one report, the Fap2 protein of F. nucleatum was shown to play a crucial role in mediating fusobacterial CRC enrichment by Gal-GalNAc that is over-expressed in CRC[6]. In addition, this protein directly interacts with the inhibitory receptor TIGIT on natural killer (NK) cells, leading to an inhibition of NK cell cytotoxicity in humans[8].
NK cells are large granular lymphocytes (unique type of cytotoxic lymphocyte) that play a role in innate immunity[12]. NK cells are involved in early defense mechanisms to kill infective pathogens and tumor cells by releasing chemokines and cytokines, as well as by helping other immune cells in targeted cell elimination[13]. Imai et al[14] in a follow-up study of 154 patients with cancer for 11 years found that patients with high cytotoxic activity among their peripheral blood lymphocytes had decreased cancer risk and, conversely, those with low cytotoxic activity had increased cancer risk[14]. In a study that evaluated 140 patients with Stage III CRC who underwent surgery, it was seen that lower pre-operative NK cell activity resulted in significant effects on the development of metachronous distant metastases[15]. A cross-sectional study performed in Canada that evaluated NK cell activity in a high-risk population of subjects screened for CRC (via colonoscopy) showed that subjects with CRC had lower NK cell activity compared to those without CRC[16]. In addition, NK cells have been shown to be deficient when analyzing their localization and densities within CRC tissues[17].
To better understand any relationship between the gut microbiome and CRC, it was hypothesized here that a high abundance of F. nucleatum in the gastrointestinal tract could cause reduced NK cell activity. Accordingly, this association would impact upon the development of CRC in a host. To test this hypothesis, associations between the abundance of F. nucleatum in the murine gastrointestinal tract and NK cell activity were evaluated.
MATERIALS AND METHODS
Bacterial culture
F. nucleatum (ATCC 25586), P. anaerobius (ATCC 27337), and P. micra (ATCC 33270) were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). All bacteria were cultured at 37°C in Anaerobe basal broth (CM0957, OXOID, Thermo Fisher Scientific, West Palm Beach, FL, USA) in an anaerobic jar (Hardy Diagnostics, Santa Maria, CA, USA). Anaerobic conditions were maintained using AnaeroGen packets (Thermo Fisher Scientific).
Culture of NK cells
For in vitro studies, NK92 cells (CRL-2407, human NK line) were obtained from ATCC. The cells were cultured in a minimum essential medium (MEM, Sigma, St. Louis, MO, USA) containing 12.5% fetal bovine serum (HyClone, Logan, UT, USA), 12.5% horse serum (HyClone), 200 U recombinant IL-2/mL (Invitrogen, Carlsbad, CA, USA), 2 mmol/L L-glutamine, 0.2 mmol/L myoinositol, 0.1 mmol/L 2-mercaptoethanol, and 0.02 mmol/L folic acid (all Sigma, St. Louis, MO, USA) at 37°C in a 5% CO2 incubator. Fresh medium was replaced every 2 d; cells reached confluence at 5 d of culture.
Cell proliferation assay
Cell viability was determined by trypsin blue exclusion. Here, NK92 cells were plated at 105 cells/well in 6-well plates. The cells were infected with bacteria at 100:1, 250:1, and 500:1 multiplicity of infection (MOI); control cells received phosphate-buffered saline (PBS, pH 7.4). After 24 and 48 h of culture, cell viabilities were assessed.
Animal experiments
C57BL/6 mice (male, 6-wk-of-age) were purchased from Orient Bio (Gyeonggi, Korea) and housed in pathogen-free facilities maintained at 20°C with 50% relative humidity and a 12-h light:dark cycle. All mice had ad libitum access to standard rodent chow and filtered water. The mice were acclimated for 1 wk and randomly divided into four groups (5 mice per group): Control, anti-control (antibiotics only), PBS (PBS after antibiotics), and FN (F. nucleatum after antibiotics). All mice (except control) were treated with a cocktail of broad-spectrum antibiotics (0.2 g/L ampicillin, 0.1 g/L vancomycin, 0.2 g/L neomycin, and 0.2 g/L metronidazole) in their drinking water for 3 wk to establish flora-deficient status. After 3 wk, the mice were administered 109 CFU F. nucleatum or the same dose of PBS by oral gavage every 2 d for 6 wk. At the end of the experiments, mice were anaesthetized with chloroform and euthanized by CO2 gas to permit collection of biomaterials. All experimental procedures were performed with the approval of the Animal Experimentation Ethics Committee of Kosin University.
F. nucleatum quantification
Stool samples were collected from all mice once a week to assess the flora-deficient status and to compare the levels of total bacteria and F. nucleatum. For this, total DNA was isolated using the GeneAll Exgene Stool DNA mini kit (GeneAll Biotechnology Co. Seoul, Korea) according to the manufacturer’s instructions. After quantification and confirmation of purity using the Exgene Stool DNA mini kit (GeneAll, Seoul, Korea), an aliquot of total DNA (1 mg) was used as a template to be mixed with TB Green Premix Ex Taq II (Takara Bio, Shiga, Japan). The primers used for amplification were: F. nucleatum 5’-CGAGGAACCTTACCAGCGTT-3’ (F) and 5’-CCCAACATCTCACGACACGA-3’ (R); total bacteria 5’-GTGSTGCAYGGYTGTCGTCA-3’ (F) and 5’-ACGTCRTCCMCACCTTCCTC-3’ (R); and GAPDH 5’-TGGCCTTCCGTGTTCCTAC-3’ (F) and 5’-GAG-TTGCTGTTGAAGTCGCA-3’ (R). All reactions were performed in triplicate in a 7300 Real-time polymerase chain reaction (PCR) system (Applied Biosystems, Waltham, MA, USA).
Assay of ex vivo NK cell activity
At necropsy, whole blood was collected from the heart and placed in sodium heparin-coated tubes for subsequent isolation of plasma. NK cell activity was then assessed (in triplicate) using a Murine NK activity kit (ATGen Co. Ltd., Seoul, Korea) according to the manufacturer’s instructions. The absorbance generated for each sample reflected the relative amount of NK cell activity in the sample.
Immunohistochemical analyses of intestinal CD3 and NKp46 cells
Murine colon tissues were collected and placed in 10% buffered formalin overnight for fixing. Sections 4-μm thick were then prepared for immunohistochemical (IHC) staining using an automated immunostainer (Bond-MAX, Leica, Buffalo Grove, IL, USA). Antibodies against mouse CD3 (rabbit polyclonal, Dako, Agilent Technologies, city, state) and NKp46 (rabbit polyclonal, R&D Systems, Minneapolis, MN, USA) were used. Stained samples were then evaluated by light microscopy and immuno-activity was assessed. The CD3+ or NKp46+ cells were counted in five different fields (total = 500 counted cells) under 400 × magnification; cell counts were then converted to percentages for data presentation.
Biochemical assays
Plasma levels of pro-inflammatory interleukin (IL)-6, -1β, -18, and -12p70, and tumor necrosis factor (TNF)-α were measured (in duplicate) using commercial ELISA kits (Mouse IL-6 Uncoated ELISA, Mouse IL-18 Platinum ELISA, Mouse IL-1β Uncoated ELISA, Mouse IL-12p70 Platinum ELISA and Mouse TNF-α Uncoated ELISA; all Invitrogen). All experimental steps were performed at room temperature and in duplicate. Samples were quantified by spectrophotometry at 450 nm. The sensitivity levels of the kits were 4-500 pg/mL for IL-6, 8-1000 pg/mL for IL-1β, 31.2-2000 pg/mL for IL-18, 15.6-1000 pg/mL for IL-12p70, and 8-1000 pg/mL for TNF-α.
Statistical analysis
Each experiment was repeated at least three times. Data are expressed as means ± SD from each independent experiment. A Student’s t-test was performed for continuous variables; outcomes with P ≤ 0.05 were considered statistically significant. Statistical analyses were performed using SPSS Statistics software (v. 23.0, IBM, Armonk, NY, USA).
RESULTS
Cytotoxic effect of F. nucleatum, P. anaerobius, and P. micra on NK cells in vitro
The effects of various gut microbiota species on NK cell survival were first assessed. Here, F. nucleatum, P. anaerobius, and P. micra -all known to be associated with CRC- were cultured with NK cells for 24 or 48 h. To identify optimal amounts of gut microbiome species affecting NK cells, experiments with each were carried out at 100:1, 250:1, or 500:1 MOI. As shown in Figure 1, NK cell counts were decreased in co-cultures with F. nucleatum, P. anaerobius, and P. micra. The results were remarkable in co-cultures at both 24 and 48 h; however, F. nucleatum most markedly decreased NK cell counts after 48 h. Therefore, F. nucleatum was selected for further study here as the representative microbiome species for use in subsequent in vivo studies.
Figure 1.

Effects of Fusobacteriumnucleatum, Peptostreptococcusanaerobius, and Parvimonasmicra on natural killer cells in vitro.Natural killer 92 cells were cultured for 24 or 48 h with each organism at a multiplicity of infection of 100:1, 250:1 or 500:1. Post-incubation, viability in the cultures was assessed using trypan blue staining. A: Fusobacterium nucleatum; B: Peptostreptococcus anaerobius; C: Parvimonas micra. F. nucleatum: Fusobacterium nucleatum; P. anaerobius: Peptostreptococcus anaerobius; P. micra: Parvimonas micra.
NK cell activity decreased in mice treated with F. nucleatum
To test whether F. nucleatum could reduce the activity of NK cells in mice, the animals were divided into four groups (5 mice per group): Control, anti-control, PBS and FN (Figure 2A). To establish flora-deficient status, mice were treated with a cocktail of broad-spectrum antibiotics for 3 wk. Confirmation of flora-deficiency was then assessed by quantitative PCR analyses of stool samples; the data confirmed that the levels of total bacteria and F. nucleatum were decreased (Figure 2B). At that point, F. nucleatum was re-introduced by oral gavage to the flora-deficient mice every 2 d for 6 wk. Analyses of their stools showed that levels of total bacteria and F. nucleatum gradually increased and then attained a plateau (Figure 2C).
Figure 2.

Experimental design. A: Mice were divided into four goups: Control, anti-control (antibiotics only), phosphate-buffered saline (PBS) (PBS after antibiotics), and Fusobacterium nucleatum (F. nucleatum) (F. nucleatum after antibiotics). The mice were treated with broad-spectrum antibiotics for 3 wk (except control), F. nucleatum was introduced to mice every 2 d for 6 wk by oral gavage; B: After treatments, the levels of total gut bacteria (left) and F. nucleatum (right) were evaluated; C: After administration of F. nucleatum, levels of total bacteria (left) and F. nucleatum (right) in mice were again evaluated. Data are expressed as means ± SD of three independent experiments. PBS: Phosphate-buffered saline.
The body weights of the mice were checked every week and compared. The weights of PBS and FN mice were lower compared to those of the control and anti-control groups after oral gavage; however, the differences were not significant (Figure 3A). At sacrifice (i.e., after 6 wk of gavage treatments), the mice were euthanized and the entire colon of each was isolated, longitudinally-opened, and examined. No significant differences in colonic length or colonic mucosal surface were found between the groups (Figure 3B).
Figure 3.

Body weight, colon length, and mucosal surface among the experimental groups. A: Weights of mice; B: Colon length and mucosal surface. PBS: Phosphate-buffered saline; FN: F. nucleatum.
To evaluate the activity of NK cells in situ, levels of interferon-gamma (IFN-γ) in the blood of each mouse was evaluated. The data indicated that the activity of NK cells in the FN group was significantly lower than in the control, anti-control, and PBS mice (Figure 4).
Figure 4.
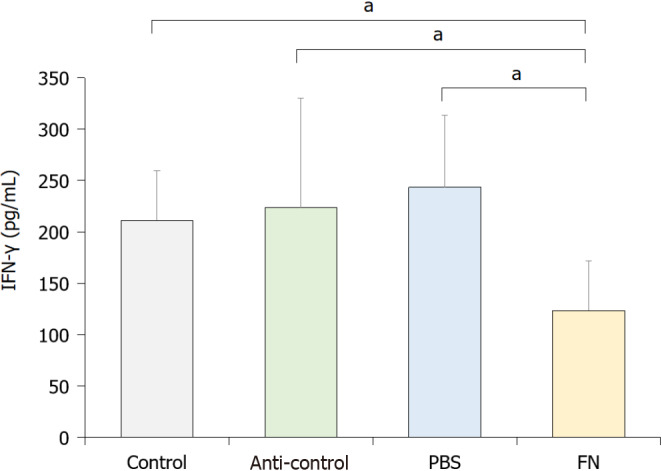
Blood levels of interferon-gamma. To indirectly evaluate the activity of natural killer cells, whole blood from each mouse was collected at necropsy and levels of interferon-gamma evaluated by ELISA. Data are expressed as means ± SD of three independent experiments. Value significantly different at aP < 0.05. IFN-γ: Interferon-gamma; PBS: Phosphate-buffered saline; FN: F. nucleatum.
Local NK cell numbers in the gut decreased in mice treated with F. nucleatum
Using IHC staining, the intestinal tract of the mice was evaluated for the presence of NK cells. The data indicate that the levels of CD3+ cells were significantly lower in the anti-control, PBS, and FN mice than in the control mice (Figure 5). The levels of NKp46+ cells were lower in the FN group than in the control group, although the difference was not significant.
Figure 5.

Intestinal levels of CD3 and NKp46 cells. A: CD3+ cells; B: NKp46+ cells. Data are expressed as means ± SD of three independent experiments. Value significantly different at aP < 0.05. NK: Natural killer; PBS: Phosphate-buffered saline; FN: F. nucleatum.
F. nucleatum treatments resulted in increased plasma pro-inflammatory cytokine levels
As shown in Figure 6, plasma levels of select pro-inflammatory cytokines were also evaluated at the end of the treatment regimens. The data indicated that circulating levels of IL-1β and TNF-α in the FN mice were significantly higher than in the control mice; values in the anti-control and PBS groups were not significantly higher than in the controls. Plasma levels of IL-6, IL-18, and IL-12p70 in the FN mice were not significantly higher than in the control. In addition, the values in the anti-control and PBS groups were not significantly changed compared to the control group.
Figure 6.

Plasma pro-inflammatory cytokine levels. A: Interleukin (IL)-1β; B: Tumor necrosis factor-α; C: IL-6; D: IL-18; E: IL-12p70. Data are expressed as means ± SD of three independent experiments. Value significantly different at aP < 0.05. PBS: Phosphate-buffered saline; FN: F. nucleatum; IL: Interleukin; TNF-α: Tumor necrosis factor-α.
DISCUSSION
Emerging evidence has shown a role for the gut microbiome, especially F. nucleatum, in cancer biology. Although F. nucleatum, a common bacterial resident in the oral cavity, has been shown to be involved in the proliferation of CRC cells and tumor development, data on how the Fusobacteria engage in the development of CRC are lacking. This study showed that a high abundance of F. nucleatum in the gastrointestinal tract reduced the activity of NK cells and that such outcomes might be associated with a concurrent increase in circulating levels of some pro-inflammatory cytokines, including IL-1β and TNF-α in these hosts. These findings suggest that increases in F. nucleatum in the gastrointestinal tract could be a factor in disrupting how the immune system prevents development of CRC.
The importance of the gut microbiome in immunity/inflammation has been more clearly elucidated in the past two decades. In a healthy state, the host immune system responds to intestinal microbiome strictly confined to the mucosal surface[18]. The mucus layer, epithelial cells, immunoglobulin A, dendritic cells, and T-cells comprise the mucosal barrier; these prevent the exposure and passage of gut microbiome to the gut-associated lymphoid tissue[19]. When the maintenance of tissue homeostasis is destroyed, translocating bacteria into the mucosal tissue causes a local (and systemic) effect on inflammatory cells, i.e., tissue inflammatory monocytes can respond to microbial-derived ligands by producing prostaglandin E2 which limits neutrophil activation and tissue damage[20].
F. nucleatum, whose presence has been proposed to correlate with CRC, can induce inflammation and suppress host immunity. There is increased evidence that F. nucleatum modulates immune cells in the CRC microenvironment and promotes intestinal tumorigenesis[21]. F. nucleatum can also stimulate autophagic pathways and alter chemotherapeutic responses of patients[22]. The current study focused on the impact of F. nucleatum upon gastrointestinal tract immune cells, especially NK cells. This study found that F. nucleatum could reduce local NK cell counts and decrease the activity of NK cells in the gut. Normally, NK cell activity is controlled by a balance of various signals that are delivered to the cells via inhibitory and activating NK cell receptors[23]. One study revealed that the ability of NK cells to kill tumors was inhibited by hemagglutinating F. nucleatum strains and that F. nucleatum inhibited immune cell activity by specifically targeting the inhibitory receptor TIGIT via the Fap2 protein[8]. Although the present study did not investigate the detailed mechanism for the observed inhibitory effects of F. nucleatum on the activity of NK cells in mice, the findings suggest that the gut microbiome could play an inhibitory role.
NK cells usually present as CD56+CD3- cells and can be categorized into CD3-CD56dimCD16+ and CD3-CD56brightCD16- cell classes. CD56dim cells appear to have more cytotoxic activity, while CD56bright cells produce high levels of cytokines including IFN-γ and TNF-α[24]. The activation and inhibition of NK cells are induced by an interplay among the cells’ activating and inhibitory receptors[25]. NKp46 is expressed on NK cells in the tumor bed and is a major determinant of NK cell function[26]. In the current study, levels of CD3+ cells significantly decreased after antibiotic treatment and NKp46+ cells decreased after administration of F. nucleatum, although the difference was not significant. Based on these results, it is thought that the microenvironment change after antibiotic treatment could affect these alterations, and the administration of F. nucleatum might decrease NK cell number.
Several inflammatory cytokines (including TNF-α, IL-6, IL-18, and IL-1β) are known to be involved in the initiation and progression of cancers[27], and could modulate the activation of NK cells. TNF-α can induce DNA damage through reactive oxygen and nitrogen species, IL-18 can induce the production of angiogenic and tumor growth stimulating factors, IL-6 can induce oxidative stress and promote early tumorigenesis, and IL-1β can promote neo-angiogenesis and evoke antiapoptotic signaling in tumor cells[28-30]. It is plausible that any changes in the expression in one or more of these could modulate tumor growth/survival in situ by also modulating the activation of NK cells[31]. For example, IL-12 can enhance the activity of NK cells and CD8+ T-lymphocytes, and so is expected to impart anti-tumor activity[32,33]. In this study, it was seen that circulating levels of IL-1β and TNF-α (but not IL-6, IL-18, or IL-12p70) were significantly increased after administration of F. nucleatum to mice. Further studies are needed to determine if these changes directly/indirectly impact the activity of NK cells themselves in hosts.
This study has some limitations. First, whether decreased NK cell activity induces the development of CRC was not assessed. Further studies including the association between NK cell activity and CRC could strengthen the results. Second, the pro-inflammatory cytokine levels were not evaluated in NK92 cells and in colonic tissues treated with F. nucleatum. Therefore, changes in plasma pro-inflammatory cytokine levels in NK92 cells and in colonic tissues treated with F. nucleatum were not confirmed.
CONCLUSION
In summary, the current study identified that high levels of F. nucleatum in the gastrointestinal tract reduced NK cell activity in mice and suggests insights on how NK cell activity was affected, i.e., F. nucleatum induced systemic pro-inflammatory responses in the hosts. Based on these findings, we suggest a possible mechanism by which high levels of F. nucleatum in the gastrointestinal tract induce an increase in plasma pro-inflammatory cytokines, which reduces NK cell activity (Figure 7). Our findings illustrate another potential means by which gut microbiome can influence NK cells which, in turn could impact the development of CRC.
Figure 7.
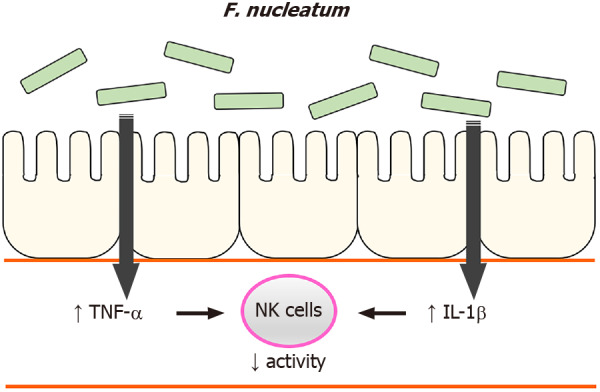
A possible mechanism. High levels of F. nucleatum in the gastrointestinal tract induce an increase in plasma pro-inflammatory cytokines, which reduces natural killer cell activity. IL: Interleukin; TNF-α: Tumor necrosis factor-α; NK: Natural killer.
ARTICLE HIGHLIGHTS
Research background
Gut microbial dysbiosis contributes to the development and progression of colorectal cancer (CRC). Natural killer (NK) cells are involved in early defense mechanisms to kill infective pathogens and tumor cells by releasing chemokines and cytokines.
Research motivation
This study was designed to better understand the relationship between the gut microbiome and CRC.
Research objectives
The objective of this study was to identify associations between gastrointestinal tract Fusobacterium nucleatum (F. nucleatum) levels and NK cell activity.
Research methods
In vitro experiments were performed on NK cells treated with F. nucleatum, Peptostreptococcus anaerobius, and Parvimonas micra to identify the effects of gut microbiome species on NK cells. After 24 and 48 h of treatment, NK cell counts were measured. In parallel studies, C57BL/6 mice were given broad-spectrum antibiotics in their drinking water to reduce resident gut flora. After 3 wk, the mice received the various bacterial species or phosphate-buffered saline via oral gavage every 2 d for 6 wk. At the study end, blood samples were acquired to perform NK cell activity assessment and cytokine analysis. Intestinal tissues were collected and analyzed via immunohistochemistry (IHC).
Research results
After 3 wk of broad-spectrum antibiotic treatment, levels of total bacteria and F. nucleatum were markedly decreased in mice. F. nucleatum by gavage significantly decreased NK cell activity relative to the activities of cells from mice treated with antibiotics only and phosphate-buffered saline. The administration of F. nucleatum decreased the proportion of NK46+ cells based on IHC staining and increased the production of interleukin-1β and tumor necrosis factor-α.
Research conclusions
High levels of F. nucleatum in the gastrointestinal tract reduced NK cell activity in mice, and the decrease in NK cell activity might be affected by increased pro-inflammatory cytokines after F. nucleatum treatment.
Research perspectives
Based on these findings, we suggest that increases in F. nucleatum in the gastrointestinal tract could be a factor in disrupting how the immune system prevents development of CRC.
Footnotes
Institutional review board statement: No human experiments were conducted and no human samples were used in this study.
Institutional animal care and use committee statement: All experimental procedures were performed with the approval of the Animal Experimentation Ethics Committee of Kosin University College of Medicine, No. KMAP-18-06.
Conflict-of-interest statement: The authors declare no conflicts of interest. The authors alone are responsible for the content of this manuscript.
ARRIVE guidelines statement: The authors have read the ARRIVE guidelines, and the manuscript was prepared and revised according to the ARRIVE guidelines.
Manuscript source: Unsolicited manuscript
Peer-review started: February 19, 2021
First decision: May 13, 2021
Article in press: July 5, 2021
Specialty type: Gastroenterology and hepatology
Country/Territory of origin: South Korea
Peer-review report’s scientific quality classification
Grade A (Excellent): 0
Grade B (Very good): B, B
Grade C (Good): C
Grade D (Fair): D
Grade E (Poor): 0
P-Reviewer: Gao F, Perisetti A, Wu HT, Yau TO S-Editor: Fan JR L-Editor: Webster JR P-Editor: Yuan YY
Contributor Information
Yeon Ji Kim, Institute of Gastroenterology, Kosin University College of Medicine, Busan 49267, Spain.
Bu Kyung Kim, Department of Internal Medicine, Kosin University College of Medicine, Busan 49267, South Korea.
Seun Ja Park, Department of Internal Medicine, Kosin University College of Medicine, Busan 49267, South Korea.
Jae Hyun Kim, Department of Internal Medicine, Kosin University College of Medicine, Busan 49267, South Korea. kjh8517@daum.net.
Data sharing statement
Technical appendix and dataset available from the corresponding author at kjh8517@daum.net.
References
- 1.Mima K, Nishihara R, Qian ZR, Cao Y, Sukawa Y, Nowak JA, Yang J, Dou R, Masugi Y, Song M, Kostic AD, Giannakis M, Bullman S, Milner DA, Baba H, Giovannucci EL, Garraway LA, Freeman GJ, Dranoff G, Garrett WS, Huttenhower C, Meyerson M, Meyerhardt JA, Chan AT, Fuchs CS, Ogino S. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut. 2016;65:1973–1980. doi: 10.1136/gutjnl-2015-310101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Louis P, Hold GL, Flint HJ. The gut microbiota, bacterial metabolites and colorectal cancer. Nat Rev Microbiol. 2014;12:661–672. doi: 10.1038/nrmicro3344. [DOI] [PubMed] [Google Scholar]
- 3.Irrazábal T, Belcheva A, Girardin SE, Martin A, Philpott DJ. The multifaceted role of the intestinal microbiota in colon cancer. Mol Cell. 2014;54:309–320. doi: 10.1016/j.molcel.2014.03.039. [DOI] [PubMed] [Google Scholar]
- 4.Nakatsu G, Li X, Zhou H, Sheng J, Wong SH, Wu WK, Ng SC, Tsoi H, Dong Y, Zhang N, He Y, Kang Q, Cao L, Wang K, Zhang J, Liang Q, Yu J, Sung JJ. Gut mucosal microbiome across stages of colorectal carcinogenesis. Nat Commun. 2015;6:8727. doi: 10.1038/ncomms9727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yu J, Feng Q, Wong SH, Zhang D, Liang QY, Qin Y, Tang L, Zhao H, Stenvang J, Li Y, Wang X, Xu X, Chen N, Wu WK, Al-Aama J, Nielsen HJ, Kiilerich P, Jensen BA, Yau TO, Lan Z, Jia H, Li J, Xiao L, Lam TY, Ng SC, Cheng AS, Wong VW, Chan FK, Yang H, Madsen L, Datz C, Tilg H, Wang J, Brünner N, Kristiansen K, Arumugam M, Sung JJ. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut. 2017;66:70–78. doi: 10.1136/gutjnl-2015-309800. [DOI] [PubMed] [Google Scholar]
- 6.Abed J, Emgård JE, Zamir G, Faroja M, Almogy G, Grenov A, Sol A, Naor R, Pikarsky E, Atlan KA, Mellul A, Chaushu S, Manson AL, Earl AM, Ou N, Brennan CA, Garrett WS, Bachrach G. Fap2 Mediates Fusobacterium nucleatum Colorectal Adenocarcinoma Enrichment by Binding to Tumor-Expressed Gal-GalNAc. Cell Host Microbe. 2016;20:215–225. doi: 10.1016/j.chom.2016.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yang Y, Weng W, Peng J, Hong L, Yang L, Toiyama Y, Gao R, Liu M, Yin M, Pan C, Li H, Guo B, Zhu Q, Wei Q, Moyer MP, Wang P, Cai S, Goel A, Qin H, Ma Y. Fusobacterium nucleatum Increases Proliferation of Colorectal Cancer Cells and Tumor Development in Mice by Activating Toll-Like Receptor 4 Signaling to Nuclear Factor-κB, and Up-regulating Expression of MicroRNA-21. Gastroenterology. 2017;152:851–866.e24. doi: 10.1053/j.gastro.2016.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gur C, Ibrahim Y, Isaacson B, Yamin R, Abed J, Gamliel M, Enk J, Bar-On Y, Stanietsky-Kaynan N, Coppenhagen-Glazer S, Shussman N, Almogy G, Cuapio A, Hofer E, Mevorach D, Tabib A, Ortenberg R, Markel G, Miklić K, Jonjic S, Brennan CA, Garrett WS, Bachrach G, Mandelboim O. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity. 2015;42:344–355. doi: 10.1016/j.immuni.2015.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nosho K, Sukawa Y, Adachi Y, Ito M, Mitsuhashi K, Kurihara H, Kanno S, Yamamoto I, Ishigami K, Igarashi H, Maruyama R, Imai K, Yamamoto H, Shinomura Y. Association of Fusobacterium nucleatum with immunity and molecular alterations in colorectal cancer. World J Gastroenterol. 2016;22:557–566. doi: 10.3748/wjg.v22.i2.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Huynh T, Kapur RV, Kaplan CW, Cacalano N, Kinder Haake S, Shi W, Sieling P, Jewett A. The role of aggregation in Fusobacterium nucleatum- induced immune cell death. J Endod. 2011;37:1531–1535. doi: 10.1016/j.joen.2011.06.034. [DOI] [PubMed] [Google Scholar]
- 11.Shenker BJ, Datar S. Fusobacterium nucleatum inhibits human T-cell activation by arresting cells in the mid-G1 phase of the cell cycle. Infect Immun. 1995;63:4830–4836. doi: 10.1128/iai.63.12.4830-4836.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kiessling R, Klein E, Pross H, Wigzell H. "Natural" killer cells in the mouse. II. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Characteristics of the killer cell. Eur J Immunol. 1975;5:117–121. doi: 10.1002/eji.1830050209. [DOI] [PubMed] [Google Scholar]
- 13.Bryceson YT, Chiang SC, Darmanin S, Fauriat C, Schlums H, Theorell J, Wood SM. Molecular mechanisms of natural killer cell activation. J Innate Immun. 2011;3:216–226. doi: 10.1159/000325265. [DOI] [PubMed] [Google Scholar]
- 14.Imai K, Matsuyama S, Miyake S, Suga K, Nakachi K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general population. Lancet. 2000;356:1795–1799. doi: 10.1016/S0140-6736(00)03231-1. [DOI] [PubMed] [Google Scholar]
- 15.Kondo E, Koda K, Takiguchi N, Oda K, Seike K, Ishizuka M, Miyazaki M. Preoperative natural killer cell activity as a prognostic factor for distant metastasis following surgery for colon cancer. Dig Surg. 2003;20:445–451. doi: 10.1159/000072714. [DOI] [PubMed] [Google Scholar]
- 16.Jobin G, Rodriguez-Suarez R, Betito K. Association Between Natural Killer Cell Activity and Colorectal Cancer in High-Risk Subjects Undergoing Colonoscopy. Gastroenterology. 2017;153:980–987. doi: 10.1053/j.gastro.2017.06.009. [DOI] [PubMed] [Google Scholar]
- 17.Halama N, Braun M, Kahlert C, Spille A, Quack C, Rahbari N, Koch M, Weitz J, Kloor M, Zoernig I, Schirmacher P, Brand K, Grabe N, Falk CS. Natural killer cells are scarce in colorectal carcinoma tissue despite high levels of chemokines and cytokines. Clin Cancer Res. 2011;17:678–689. doi: 10.1158/1078-0432.CCR-10-2173. [DOI] [PubMed] [Google Scholar]
- 18.Konrad A, Cong Y, Duck W, Borlaza R, Elson CO. Tight mucosal compartmentation of the murine immune response to antigens of the enteric microbiota. Gastroenterology. 2006;130:2050–2059. doi: 10.1053/j.gastro.2006.02.055. [DOI] [PubMed] [Google Scholar]
- 19.Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell. 2014;157:121–141. doi: 10.1016/j.cell.2014.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schreiber S, Rosenstiel P, Albrecht M, Hampe J, Krawczak M. Genetics of Crohn disease, an archetypal inflammatory barrier disease. Nat Rev Genet. 2005;6:376–388. doi: 10.1038/nrg1607. [DOI] [PubMed] [Google Scholar]
- 21.Brennan CA, Garrett WS. Gut Microbiota, Inflammation, and Colorectal Cancer. Annu Rev Microbiol. 2016;70:395–411. doi: 10.1146/annurev-micro-102215-095513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yu T, Guo F, Yu Y, Sun T, Ma D, Han J, Qian Y, Kryczek I, Sun D, Nagarsheth N, Chen Y, Chen H, Hong J, Zou W, Fang JY. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell. 2017;170:548–563.e16. doi: 10.1016/j.cell.2017.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol. 2013;34:182–191. doi: 10.1016/j.it.2013.01.003. [DOI] [PubMed] [Google Scholar]
- 24.Robertson MJ, Ritz J. Biology and clinical relevance of human natural killer cells. Blood. 1990;76:2421–2438. [PubMed] [Google Scholar]
- 25.Cooper MA, Fehniger TA, Caligiuri MA. The biology of human natural killer-cell subsets. Trends Immunol. 2001;22:633–640. doi: 10.1016/s1471-4906(01)02060-9. [DOI] [PubMed] [Google Scholar]
- 26.Chretien AS, Devillier R, Fauriat C, Orlanducci F, Harbi S, Le Roy A, Rey J, Bouvier Borg G, Gautherot E, Hamel JF, Ifrah N, Lacombe C, Cornillet-Lefebvre P, Delaunay J, Toubert A, Arnoulet C, Vey N, Blaise D, Olive D. NKp46 expression on NK cells as a prognostic and predictive biomarker for response to allo-SCT in patients with AML. Oncoimmunology. 2017;6:e1307491. doi: 10.1080/2162402X.2017.1307491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Landskron G, De la Fuente M, Thuwajit P, Thuwajit C, Hermoso MA. Chronic inflammation and cytokines in the tumor microenvironment. J Immunol Res. 2014;2014:149185. doi: 10.1155/2014/149185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Palma G, Barbieri A, Bimonte S, Palla M, Zappavigna S, Caraglia M, Ascierto PA, Ciliberto G, Arra C. Interleukin 18: friend or foe in cancer. Biochim Biophys Acta. 2013;1836:296–303. doi: 10.1016/j.bbcan.2013.09.001. [DOI] [PubMed] [Google Scholar]
- 29.Murata M, Thanan R, Ma N, Kawanishi S. Role of nitrative and oxidative DNA damage in inflammation-related carcinogenesis. J Biomed Biotechnol. 2012;2012:623019. doi: 10.1155/2012/623019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Meng F, Wehbe-Janek H, Henson R, Smith H, Patel T. Epigenetic regulation of microRNA-370 by interleukin-6 in malignant human cholangiocytes. Oncogene. 2008;27:378–386. doi: 10.1038/sj.onc.1210648. [DOI] [PubMed] [Google Scholar]
- 31.Zwirner NW, Domaica CI. Cytokine regulation of natural killer cell effector functions. Biofactors. 2010;36:274–288. doi: 10.1002/biof.107. [DOI] [PubMed] [Google Scholar]
- 32.Tugues S, Burkhard SH, Ohs I, Vrohlings M, Nussbaum K, Vom Berg J, Kulig P, Becher B. New insights into IL-12-mediated tumor suppression. Cell Death Differ. 2015;22:237–246. doi: 10.1038/cdd.2014.134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lu X. Impact of IL-12 in Cancer. Curr Cancer Drug Targets. 2017;17:682–697. doi: 10.2174/1568009617666170427102729. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Technical appendix and dataset available from the corresponding author at kjh8517@daum.net.