

Abstract
Lung cancer represents the most common type of human malignancy and is the main cause of cancer-associated mortality worldwide. To improve the effectiveness of treatment strategies, a better understanding of the mechanisms of cancer progression and invasiveness is required. Recently, B-cell activating factor (BAFF) and a proliferation-inducing ligand (APRIL), two relatively newly described cytokines belonging to the tumor necrosis factor superfamily, have been shown to play a role in cancer progression. However, at present, the effects of both cytokines on lung cancer cells remain unclear. The present study aimed therefore to understand the direct effects of BAFF and APRIL on non-small cell lung cancer (NSCLC) progression. To do so, reverse transcription quantitative PCR and western blotting were used to evaluate whether A549 and H2030 NSCLC cells express receptors for both BAFF and APRIL. The results demonstrated that both investigated cell lines expressed BAFF-R (receptor specific to BAFF only) and transmembrane activator and CAML interactor (TACI; shared receptor for both cytokines). In addition, functional experiments were performed to determine the effects of BAFF and APRIL stimulation on cancer cell viability. The results demonstrated no direct effects of BAFF and APRIL on NSCLC cell proliferation and invasiveness. In summary, the present study demonstrated that NSCLC cells possess the ability to respond directly to both BAFF and APRIL. However, activation of BAFF-R and TACI signaling in cancer cells did not increase the proliferative capacity and invasiveness. Further investigation is thus required to better understand the role of BAFF and APRIL on the progression of NSCLC.
Keywords: lung cancer, B-cell activating factor, a proliferation-inducing ligand, B-cell maturation antigen, transmembrane activator and CAML interactor, B-cell activating factor receptor, tumor progression
Introduction
Lung cancer is the most common malignancy diagnosed worldwide and represents the leading cause of cancer associated-mortality. Among all types of lung cancer, non-small cell lung cancer (NSCLC) is the most frequent, accounting for ~85% of all lung cancer cases (1). Although lung neoplasm pathogenesis is known to rise constitutively, it is crucial to better understand the mechanisms of cancer progression, including mutual interactions between malignant and immune cells (2,3). Due to immune regulatory properties, cancer cells regulate innate and adaptive immune responses, abolishing antitumor activities and supporting mechanisms promoting tumor progression (4,5). Among tumor-infiltrating immune cells, the vast majority is represented by myeloid cells, such as monocytes/macrophages, which are referred to as tumor-associated macrophages (6,7). Due to pleiotropic biological activities, myeloid cells play a central role in the regulation of the tumor microenvironment by means of secreted soluble factors, including proteins belonging to the tumor necrosis factor (TNF) superfamily (8,9).
B-cell activating factor (BAFF) and a proliferation-inducing ligand (APRIL) are relatively newly discovered TNF superfamily members. Both ligands might act through direct interaction with shared receptors, such as B-cell maturation antigen (BCMA) and transmembrane activator and CML interactor (TACI), while BAFF is also explicitly recognized by BAFF receptor (BAFF-R). Both ligands serve a crucial role in B cell development, maturation and immunoglobulin class switching (10). Furthermore, BAFF was shown to stimulate anti-apoptotic signaling in tumor cells (8,11). Previous studies demonstrated that BAFF could play a role in neoplasm progression and aggressiveness (12–14). In addition, it was reported that both BAFF and APRIL signaling might increase tumor cell proliferation and enhance tumor cell viability in response to chemotherapeutic drugs in hematopoietic malignancies (8,9,15).
At present, to the best of our knowledge, the role of BAFF and APRIL in solid tumors development and progression remains unknown. However, the expression of their receptors was mainly reported in tumor-infiltrating cells (16,17). Interestingly, elevated blood levels of BAFF and APRIL are associated with a higher stage of the disease and cancer invasiveness, such as in breast cancer, chronic lymphocytic leukemia and pancreatic cancer (12,18). Furthermore, the elevated expression of BAFF and APRIL is associated with increased cell migration, epithelial-mesenchymal transistion and stemness, subsequently contributing to an aggressive phenotype of breast cancer (12,19,20). Similarly, APRIL overexpression was shown to be associated with tumor progression and was therefore proposed as a potential prognostic factor in rectal cancer, pancreatic adenocarcinoma and various B-cell malignances (13). In addition, in hepatocellular carcinoma cells, APRIL can play pleiotropic biological activities. Subsequently, APRIL may support tumor growth or reduce cell proliferation, depending on the particular pathway activation (21). Previous studies reported for the first time the importance of BAFF and APRIL signaling in NSCLC (22,23). However, to date, it remains unclear whether BAFF and APRIL may serve a direct role in regulating lung cancer cell proliferation and invasiveness (22,24). The present study aimed therefore to evaluate the effects of BAFF and APRIL on NSCLC cell proliferation and invasiveness.
Materials and methods
Cell lines
The human NSCLC cell lines A549 and H2030 were purchased from the American Type Culture Collection. Cells were cultured in Dulbecco's modified Eagles medium (PAN Biotech UK, Ltd.) supplemented with 10% of heat-inactivated and filtered FBS (PAN Biotech UK, Ltd.) and gentamycin (50 µg/ml; Gibco; Thermo Fisher Scientific, Inc.) and placed at 37°C in a humidified incubator containing 5% CO2. Cells were passaged when at 70–80% confluence. Cells from the second to fifth passage were used for all experiments.
Reverse transcription quantitative PCR
To assess the expression level of BAFF, APRIL, BAFF-R, BCMA and TACI, cells were harvested, washed with PBS and lysed in RLT buffer (Qiagen) supplemented with 1% β-mercaptoethanol (Sigma-Aldrich; Merck KGaA) at room temperature. Total RNA was extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturers' protocol and quantified using NanoDrop (NanoDrop 2000c/2000; Thermo Fisher Scientific, Inc.). Reverse transcription was performed using the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Inc.) according to manufacturers' instructions. Commercially available TaqMan assays (Thermo Fisher Scientific, Inc.; Table I) and TaqMan Universal PCR Master Mix (Thermo Fisher Scientific, Inc.) were used according to manufacturers' instructions. The thermocyclig conditions were as follows: Polymerase activation at 95°C for 10 min followed by 40 cycles of denaturation at 95°C for 15 sec and annealing/extension at 60°C for 1 min. Samples were analyzed in the StepOne Plus system (Thermo Fisher Scientific, Inc.). Data were analyzed using StepOne Software v2.3 (Thermo Fisher Scientific, Inc.). The relative expression levels were normalized to endogenous control and were expressed as 2−ΔΔCq (25).
Table I.
TaqMan assays used for reverse transcription quantitative PCR.
| Assay | Target species | Catalogue number | Manufacturer |
|---|---|---|---|
| TNFSF13 (BAFF) | Human | Hs00198106_m1 | Thermo Fisher Scientific, Inc. |
| TNFSF13 (APRIL) | Human | Hs00601664_g1 | Thermo Fisher Scientific, Inc. |
| TNFRSF1 (BAFF-R) | Human | Hs00606874_g1 | Thermo Fisher Scientific, Inc. |
| TNFRSF1 (TACI) | Human | Hs00963364_m1 | Thermo Fisher Scientific, Inc. |
| TNFRSF1 (BCMA) | Human | Hs03045080_m1 | Thermo Fisher Scientific, Inc. |
| GADPH | Human | Hs02786624_g1 | Thermo Fisher Scientific, Inc. |
APRIL, a proliferation-inducing ligand; BAFF, B-cell activating factor; BAFF-R, BAFF receptor; TACI, transmembrane activator and CAML interactor; BCMA, B-cell maturation antigen.
Western blotting
To evaluate the protein expression of BAFF, APRIL, BAFF-R, BCMA and TACI, cells were harvested, washed in PBS and lysed in RIPA buffer (Thermo Fisher Scientific, Inc.) supplemented with complete protease inhibitor cocktail (Roche Diagnostics) for 15 min on ice. The cell debris were removed by centrifugation at 13,000 × g for 5 min at 4°C. The concentration of total protein was assessed using Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Inc.) according to manufacturers' instructions. Proteins (20 µg) were mixed with Lammli sample buffer (1:4; Bio-Rad Laboratories, Inc.) and heated at 95°C for 5 min. Proteins were separated on 4–10% TGX gel (Bio-Rad Laboratories, Inc.) and transferred onto nitrocellulose membranes (Bio-Rad Laboratories, Inc.) using the TransBlot turbo system (Bio-Rad Laboratories, Inc.). The membranes were blocked using 5% skimmed milk dissolved in PBS and supplemented with 0.1% Tween-20 (Sigma-Aldrich; Merck KGaA; T-PBS) for 1 h at room temperature. Membranes were incubated with primary antibodies overnight at 4°C (Table II). Subsequently, membranes were washed using 10X PBS (Corining) supplemented with 0,1% Tween-20 and incubated with specific HRP-conjugated secondary antibodies (Table II) for 1 h in room temperature. Bands were detected using the SuperSignal West Femto chemiluminescence substrate kit (Thermo Fisher Scientific, Inc.) and visualized with ChemiDoc Touch System (Bio-Rad Laboratories, Inc.). Relative expression levels were normalized to endogenous control (β-actin) using ImageJ software v1.53 (National Institutes of Health).
Table II.
Description of antibodies used for western blotting.
| Antibody | Target species | Host | Isotype | Clone | Dilution | Catalogue number | Manufacturer |
|---|---|---|---|---|---|---|---|
| BAFF-R | Human, mouse and rat | Mouse | IgG2b | H-1 | 1:150 | sc-365410 | Santa Cruz Biotechnology, Inc. |
| TACI | Human, mouse and rat | Mouse | IgG2b | C-9 | 1:200 | sc-365253 | Santa Cruz Biotechnology, Inc. |
| BCMA | Human | Rat | IgG2a | 335004 | 1:100 | MAB193 | R&D Systems, Inc. |
| β-actin (Direct-Blot HRP) | Human, mouse and rat | Mouse | IgG2b | 2F1-1 | 1:1000 | 643807 | BioLegend, Inc. |
| Anti-mouse IgG HRP-conjugated | Mouse | Goat | IgG | Polyclonal | 1:1000 | HAF007 | R&D Systems, Inc. |
| Anti-rat IgG HRP-conjugated | Rat | Goat | IgG | Polyclonal | 1:1000 | HAF005 | R&D Systems, Inc. |
BAFF-R, B-cell activating factor receptor; TACI, transmembrane activator and CAML interactor; BCMA, B-cell maturation antigen. HRP, horseradish peroxidase; Ig, immunoglobulin.
Ligand receptor interaction blocking
To block the interactions of BAFF, APRIL and their receptors in A549 and H2030 cell lines, blocking antibodies for TACI (monoclonal mouse IgG1 clone #165609; R&D Systems, Inc.; cat. no. MAB174) and BAFF-R (monoclonal mouse IgG2a, kappa clone #8A7; Thermo Fisher Scientific, Inc.) were used separately or in combination at 10 µg/ml for 24 h in standard cell culture conditions (37°C, 5% CO2).
Cell proliferation assay
To assess the effects of BAFF and APRIL on lung cancer cell proliferation, the cells were harvested and labeled with carboxyfluorescein succinimidyl ester (CFSE; Sigma-Aldrich; Merck KGaA). The cells were cultured for 72 h in 24 well plates (37°C, 5% CO2) in the presence of recombinant human BAFF or APRIL proteins at the concentrations of 50, 100 or 150 ng/ml or without any stimulation (vehicle). The cells were acquired by using TrypLE Select (Thermo Fisher Scientific, Inc.) at 24, 48 and 72 h. Subsequently, cells were washed in PBS and stained with 7-AAD as suggested by the manufacturer (0,25 µg per 1×106 cells; Becton-Dickinson and Company), and incubated for 15 min at 4°C. Cells were analyzed on FACSCalibur flow cytometry system (Becton-Dickinson and Company). Data were analyzed using FlowJo software version 10.6.1 (Tree Star, Inc.). To set the gates for the analysis of proliferation (negative control), colcemid (50 ng/ml) was used to arrest the mitosis in metastatasis stage. Cell viability was assessed according to 7-AAD staining and viable cells were considered as 7-AAD negative.
Invasion assay
To evaluate the influence of BAFF and APRIL on non-small cell lung cancer invasiveness, a commercially available invasion assay (Cell Biolabs, Inc.) was used according to the manufacturers' instruction. Briefly, the membrane was rehydrated, and cell culture media containing 10% FBS (vehicle) or 10% FBS with BAFF (150 ng/ml) or 10% FBS with APRIL (150 ng/ml) was added to the lower well of the invasion plate. Subsequently, cells in single-cell suspension were placed in the insert. After 48 h incubation at 37°C, the invasive cells were stained with cell stain solution, lysed with extraction solution and the absorbance was measured at 560 nm using a plate reader (Ledetect; Labexim Products). The data were analyzed using MicroWin2000 software (OEM version; Labexim Products).
Statistical analysis
Statistical analysis was performed using GraphPad Prism 6 software (GraphPad Software, Inc.). Data were compared using Mann-Whitney U test or Kruskal-Wallis followed by Dunn's post hoc test. P<0.05 was considered to indicate a statistically significant difference. The results from at least five independent experiments are presented as the median ± interquartile range.
Results
NSCLC cells express BAFF, APRIL and their receptors
First, we aimed to investigate whether NSCLC cells could express BAFF, APRIL and their receptors BCMA, TACI and BAFF-R using RT-qPCR and western blotting (Fig. 1). The results demonstrated that A549 and H2030 cell lines expressed BAFF, APRIL and BAFF-R (Fig. 1A-C). Conversely, the expression of TACI and BCMA was detected in H2030 cells only, while in the A549 cell line, the expression of both receptors was under the detection limit (data not shown). Furthermore, no difference was found in the expression levels of BAFF, APRIL and BAFF-R among the analyzed cell lines (Fig. 1A-C). In addition, results from western blotting demonstrated that both cell lines expressed BAFF-R and TACI at the protein level, while no expression of BCMA was detected (Fig. 1D). These findings suggested that NSCLC cells could respond directly to BAFF and APRIL stimulation via BAFF-R and TACI signaling.
Figure 1.
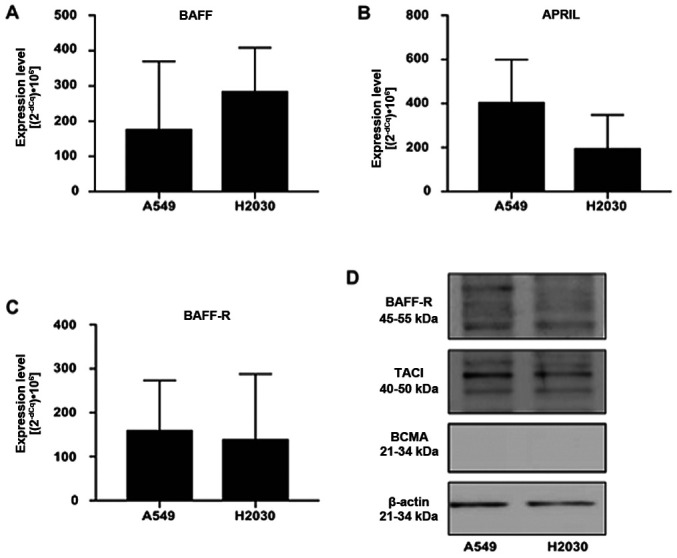
BAFF-R and TACI are expressed in non-small cell lung cancer cells. (A) BAFF, (B) APRIL and (C) BAFF-R gene expression levels in A549 and H2030 cell lines. (D) Representative western blottings for BAFF-R, TACI and BCMA in A549 and H2030 cell lines. No significant differences were observed. APRIL, a proliferation-inducing ligand; BAFF-R, B-cell activating factor receptor; TACI, transmembrane activator and CAML interactor; BCMA, B-cell maturation antigen.
Effect of different BAFF and APRIL doses on NSCLC cells viability
Having found that both cell lines possess the potential to directly respond to BAFF and APRIL stimulation by the interaction with TACI and BAFF-R, we subsequently aimed to investigate the direct effects of both ligands on cancer cell viability. As expected, the results demonstrated that stimulation of H2030 and A549 cells with 50, 100 and 150 ng/ml BAFF (Fig. 2A and B) and APRIL (Fig. 2C and D) did not affect NSCLC cell viability. These findings indicated that BAFF and APRIL were not toxic for NSCLC cells and could be used in further functional experiments.
Figure 2.
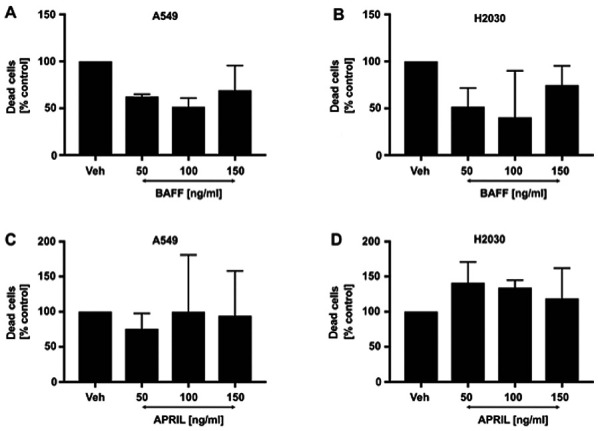
Effects of BAFF and APRIL signaling on non-small cell lung cancer cell viability. Effect of BAFF signaling on (A) A549 and (B) H2030 cell viability. Effects of APRIL signaling on (C) A549 and (D) H2030 cell viability. Cells were cultured in the presence of different concentrations of BAFF and APRIL or untreated (veh alone). No significant differences were observed. APRIL, a proliferation-inducing ligand; BAFF, B-cell activating factor; veh, vehicle.
NSCLC-derived BAFF and APRIL do not influence cell proliferation
Having found that both cell lines can produce the analyzed ligands, we aimed to investigate whether A549 and H2030 cell-derived BAFF and APRIL may directly influence their proliferation (Fig. 3). Functional experiments were performed with receptor blocking using functional-grade monoclonal antibodies. The results demonstrated that blocking TACI and BAFF-R separately or in combination did not influence the proliferation of A549 (Fig. 3C) and H2030 (Fig. 3D) cells. These results indicated that endogenous BAFF and APRIL may not increase tumor growth directly by increasing cancer cell proliferation.
Figure 3.

Endogenous BAFF and APRIL do not change non-small cell lung cancer cell proliferation. Representative flow cytometry density plots for CFSE based proliferation of (A) A549 and (B) H2030 cell lines with blocked TACI (anti-TACI) and/or BAFF-R (anti-BAFF-R). Effects of blocking TACI and/or BAFF-R in (C) A549 and (D) H2030 cell lines. Colcemid was used as a negative control of cancer cell proliferation (for gate set up). No significant differences were observed. BAFF-R, B-cell activating factor receptor; TACI, transmembrane activator and CAML interactor; vh, vehicle
Exogenous BAFF and APRIL did not influence NSCLC cell proliferation
Next, we wished to analyze whether exogenous BAFF and APRIL could directly affect NSCLC cell proliferation. Cells were stimulated with recombinant human BAFF and APRIL at 50, 100 and 150 ng/ml. The results demonstrated no effect of exogenous stimulation with both ligands on A549 (Fig. 4A) and H2030 (Fig. 4B) cell proliferation, since no differences in the frequency of proliferating cells was observed (Fig. 4C-F). Furthermore, no effect on cancer cell invasiveness in response to BAFF and APRIL stimulation was observed compared with vehicle alone (Fig. 5).
Figure 4.

Direct effects of exogenous BAFF and APRIL signaling on non-small cell lung cancer cell proliferation. Representative flow cytometry density plots for CFSE based proliferation in BAFF and APRIL-treated (A) A549 and (B) H2030 cells. Effects of different concentrations BAFF and APRIL on A549 (C and E, respectively) and H2030 (D and F, respectively) cells. Colcemid was used as a negative control of cancer cell proliferation (for gate set up). No significant differences were observed. APRIL, a proliferation-inducing ligand; BAFF, B-cell activating factor; veh, vehicle.
Figure 5.

The influence of BAFF and APRIL signaling on non-small cell lung cancer cell aggressiveness. Effect of 150 ng/ml BAFF and APRIL treatments on A549 and H2030 cancer cell aggressiveness. 10% FBS (vehicle) was used as a control. No significant differences were observed. APRIL, a proliferation-inducing ligand; BAFF, B-cell activating factor; veh, vehicle.
Discussion
The present study demonstrated that NSCLC cells possess the ability to respond directly to BAFF and APRIL stimulation via interaction with TACI and BAFF-R, while BCMA was undetectable at the protein level. Previous studies reported that BAFF and APRIL can enhance the proliferation of normal and malignant B cells (11,20). In addition, previous studies from our group demonstrated that the expression of BCMA might serve as a prognostic factor for treatment response in patients with acute myeloid leukemia (8,9). In solid tumors, BAFF and APRIL signaling is associated with cancer cell proliferation, such as in breast cancer (12), esophageal cancer (26), clear cell renal cell carcinoma (27), adult male germ cell tumor (28) and invasive bladder carcinoma (29). Furthermore, elevated local and systemic levels of BAFF and APRIL were shown to be associated with a higher stage and disease progression, such as in chronic graft versus host disease, systemic lupus erythormatosus and breast cancer tumor size (12,13). Surprisingly, the present study demonstrated that BAFF and APRIL direct signaling did not serve an essential role in NSCLC aggressiveness, as no effect of BAFF and APRIL on NSCLC cell proliferation and invasiveness was reported. Conversely, previous studies reported that APRIL signaling in NSCLC cells promote tumor proliferation, migration and metastasis (23,30). Consequently, both ligands may act as tumor supporters and pro-metastatic factors in an indirect manner. In fact, high BAFF and APRIL expression were shown in NSCLC tissues (22). It is therefore essential to determine whether BAFF and APRIL signaling in tumor-infiltrating or stromal cells, including tumor-infiltrating macrophages or tumor-associated fibroblasts, may play a role in tumor progression and spread.
Macrophages (alternatively activated) and fibroblasts are crucial components of tumor stroma. The progressive effects of macrophages and fibroblasts on tumor growth are associated with the release of growth factors and anti-inflammatory mediators, their promotion of cancer invasiveness by releasing matrix degradation enzymes, and their involvement in the recruitment of suppressive cells, including T regulatory cells to the tumor side (31,32). Both aforementioned cell subsets have been shown to produce high levels of BAFF and APRIL; however, the effects of both ligands on the activation and biological function of tumor-infiltrating macrophages and tumor-associated fibroblasts remain elusive (18,24,33–35). BAFF via direct interaction with TACI may therefore induce monocyte maturation towards macrophages and, thus, contribute to tumor progression.
BAFF and APRIL signaling stimulate the expression of anti-apoptotic proteins, such as Bcl-2, Bcl-XL and Bcl-2-related protein A1 (36,37). Both ligands may therefore serve as anti-apoptotic mediators and increase the survival of cancer cells subjected to chemotherapy (11). Interestingly, APRIL gene silencing was shown to increase the apoptotic susceptibility of gastric cancer cells (38). However, the present study demonstrated that BAFF and APRIL stimulation had no effects on the viability of NSCLC cells in normal conditions. Further investigation is therefore required to elucidate the impact of BAFF and APRIL on cancer cell susceptibility to cytotoxic therapy.
In summary, the results from the present study demonstrated that, despite the presence of TACI and BAFF-R in NSCLC cells, both BAFF and APRIL did not exert direct effects on cancer cell proliferation and invasiveness. Further studies are needed to elucidate the mechanisms of previously reported ex vivo associations between BAFF and APRIL and cancer progression (12,13).
Acknowledgements
Not applicable.
Funding Statement
This study was supported by the statutory founds of Medical University of Bialystok, Poland (grant no. N/ST/ZB/16/001/1113).
Funding
This study was supported by the statutory founds of Medical University of Bialystok, Poland (grant no. N/ST/ZB/16/001/1113).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
AE, MM and LB developed the research concept. AE and LB designed the experiments. MW, MT, DL and KG performed the experiments. MW, MT and JD performed statistical analysis and designed figures. MW and MT drafted the mancuscript. AE, LB, JD and MM revised the draft. AE, MM and BL confirmed the authenticity of all the raw data. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable
Patient consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
References
- 1.Molina JR, Yang P, Cassivi SD, Schild SE, Adjei AA. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin Proc. 2008;83:584–594. doi: 10.4065/83.5.584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol. 2013;14:1014–1022. doi: 10.1038/ni.2703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hirsch FR, Scagliotti GV, Mulshine JL, Kwon R, Curran WJ, Jr, Wu YL, Paz-Ares L. Lung cancer: Current therapies and new targeted treatments. Lancet. 2017;389:299–311. doi: 10.1016/S0140-6736(16)30958-8. [DOI] [PubMed] [Google Scholar]
- 4.Stankovic B, Bjørhovde HAK, Skarshaug R, Aamodt H, Frafjord A, Müller E, Hammarström C, Beraki K, Bækkevold ES, Woldbæk PR, et al. Immune Cell Composition in Human Non-small Cell Lung Cancer. Front Immunol. 2019;9:3101. doi: 10.3389/fimmu.2018.03101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Liu X, Wu S, Yang Y, Zhao M, Zhu G, Hou Z. The prognostic landscape of tumor-infiltrating immune cell and immunomodulators in lung cancer. Biomed Pharmacother. 2017;95:55–61. doi: 10.1016/j.biopha.2017.08.003. [DOI] [PubMed] [Google Scholar]
- 6.Poh AR, Ernst M. Targeting macrophages in cancer: From bench to bedside. Front Oncol. 2018;8:49. doi: 10.3389/fonc.2018.00049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Loyher PL, Hamon P, Laviron M, Meghraoui-Kheddar A, Goncalves E, Deng Z, Torstensson S, Bercovici N, Baudesson de Chanville C, Combadière B, et al. Macrophages of distinct origins contribute to tumor development in the lung. J Exp Med. 2018;215:2536–2553. doi: 10.1084/jem.20180534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bolkun L, Grubczak K, Schneider G, Zembko P, Radzikowska U, Singh P, Kloczko J, Ratajczak MZ, Moniuszko M, Eljaszewicz A. Involvement of BAFF and APRIL in Resistance to Apoptosis of Acute Myeloid Leukemia. J Cancer. 2016;7:1979–1983. doi: 10.7150/jca.15966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bolkun L, Lemancewicz D, Jablonska E, Szumowska A, Bolkun-Skornicka U, Ratajczak-Wrona W, Dzieciol J, Kloczko J. The impact of TNF superfamily molecules on overall survival in acute myeloid leukaemia: Correlation with biological and clinical features. Ann Hematol. 2015;94:35–43. doi: 10.1007/s00277-014-2178-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Meinl E, Thaler FS, Lichtenthaler SF. Shedding of BAFF/APRIL receptors controls B cells. Trends Immunol. 2018;39:673–676. doi: 10.1016/j.it.2018.07.002. [DOI] [PubMed] [Google Scholar]
- 11.Kern C, Cornuel JF, Billard C, Tang R, Rouillard D, Stenou V, Defrance T, Ajchenbaum-Cymbalista F, Simonin PY, Feldblum S, et al. Involvement of BAFF and APRIL in the resistance to apoptosis of B-CLL through an autocrine pathway. Blood. 2004;103:679–688. doi: 10.1182/blood-2003-02-0540. [DOI] [PubMed] [Google Scholar]
- 12.Pelekanou V, Notas G, Athanasouli P, Alexakis K, Kiagiadaki F, Peroulis N, Kalyvianaki K, Kampouri E, Polioudaki H, Theodoropoulos P, et al. BCMA (TNFRSF17) induces APRIL and BAFF mediated breast cancer cell stemness. Front Oncol. 2018;8:301. doi: 10.3389/fonc.2018.00301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Moreaux J, Veyrune JL, De Vos J, Klein B. APRIL is overexpressed in cancer: Link with tumor progression. BMC Cancer. 2009;9:83. doi: 10.1186/1471-2407-9-83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Quinn J, Glassford J, Percy L, Munson P, Marafioti T, Rodriguez-Justo M, Yong K. APRIL promotes cell-cycle progression in primary multiple myeloma cells: Influence of D-type cyclin group and translocation status. Blood. 2011;117:890–901. doi: 10.1182/blood-2010-01-264424. [DOI] [PubMed] [Google Scholar]
- 15.Bolkun L, Lemancewicz D, Jablonska E, Kulczynska A, Bolkun-Skornicka U, Kloczko J, Dzieciol J. BAFF and APRIL as TNF superfamily molecules and angiogenesis parallel progression of human multiple myeloma. Ann Hematol. 2014;93:635–644. doi: 10.1007/s00277-013-1924-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yang S, Li JY, Xu W. Role of BAFF/BAFF-R axis in B-cell non-Hodgkin lymphoma. Crit Rev Oncol Hematol. 2014;91:113–122. doi: 10.1016/j.critrevonc.2014.02.004. [DOI] [PubMed] [Google Scholar]
- 17.Tai YT, Acharya C, An G, Moschetta M, Zhong MY, Feng X, Cea M, Cagnetta A, Wen K, van Eenennaam H, et al. APRIL and BCMA promote human multiple myeloma growth and immunosuppression in the bone marrow microenvironment. Blood. 2016;127:3225–3236. doi: 10.1182/blood-2016-01-691162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Koizumi M, Hiasa Y, Kumagi T, Yamanishi H, Azemoto N, Kobata T, Matsuura B, Abe M, Onji M. Increased B cell-activating factor promotes tumor invasion and metastasis in human pancreatic cancer. PLoS One. 2013;8:e71367. doi: 10.1371/journal.pone.0071367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kampa M, Notas G, Stathopoulos EN, Tsapis A, Castanas E. The TNFSF members APRIL and BAFF and their receptors TACI, BCMA, and BAFFR in oncology, with a special focus in breast cancer. Front Oncol. 2020;10:827. doi: 10.3389/fonc.2020.00827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Novak AJ, Grote DM, Stenson M, Ziesmer SC, Witzig TE, Habermann TM, Harder B, Ristow KM, Bram RJ, Jelinek DF, et al. Expression of BLyS and its receptors in B-cell non-Hodgkin lymphoma: Correlation with disease activity and patient outcome. Blood. 2004;104:2247–2253. doi: 10.1182/blood-2004-02-0762. [DOI] [PubMed] [Google Scholar]
- 21.Notas G, Alexaki VI, Kampa M, Pelekanou V, Charalampopoulos I, Sabour-Alaoui S, Pediaditakis I, Dessirier V, Gravanis A, Stathopoulos EN, et al. APRIL binding to BCMA activates a JNK2-FOXO3-GADD45 pathway and induces a G2/M cell growth arrest in liver cells. J Immunol. 2012;189:4748–4758. doi: 10.4049/jimmunol.1102891. [DOI] [PubMed] [Google Scholar]
- 22.Dou H, Yan Z, Zhang M, Xu X. APRIL, BCMA and TACI proteins are abnormally expressed in non-small cell lung cancer. Oncol Lett. 2016;12:3351–3355. doi: 10.3892/ol.2016.5095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dou H, Yan Z, Zhang M, Xu X. APRIL promotes non-small cell lung cancer growth and metastasis by targeting ERK1/2 signaling. Oncotarget. 2017;8:109289–109300. doi: 10.18632/oncotarget.22672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Qian Z, Qingshan C, Chun J, Huijun Z, Feng L, Qiang W, Qiang X, Min Z. High expression of TNFSF13 in tumor cells and fibroblasts is associated with poor prognosis in non-small cell lung cancer. Am J Clin Pathol. 2014;141:226–233. doi: 10.1309/AJCP4JP8BZOMHEAW. [DOI] [PubMed] [Google Scholar]
- 25.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 26.Hao Y, Triadafilopoulos G, Sahbaie P, Young HS, Omary MB, Lowe AW. Gene expression profiling reveals stromal genes expressed in common between Barrett's esophagus and adenocarcinoma. Gastroenterology. 2006;131:925–933. doi: 10.1053/j.gastro.2006.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lee C, Park JW, Suh JH, Moon KC. High expression of APRIL correlates with poor prognosis in clear cell renal cell carcinoma. Pathol Res Pract. 2015;211:824–828. doi: 10.1016/j.prp.2015.07.011. [DOI] [PubMed] [Google Scholar]
- 28.Korkola JE, Houldsworth J, Chadalavada RS, Olshen AB, Dobrzynski D, Reuter VE, Bosl GJ, Chaganti RS. Down-regulation of stem cell genes, including those in a 200-kb gene cluster at 12p13.31, is associated with in vivo differentiation of human male germ cell tumors. Cancer Res. 2006;66:820–827. doi: 10.1158/0008-5472.CAN-05-2445. [DOI] [PubMed] [Google Scholar]
- 29.Sanchez-Carbayo M, Socci ND, Lozano J, Saint F, Cordon-Cardo C. Defining molecular profiles of poor outcome in patients with invasive bladder cancer using oligonucleotide microarrays. J Clin Oncol. 2006;24:778–789. doi: 10.1200/JCO.2005.03.2375. [DOI] [PubMed] [Google Scholar]
- 30.Sun B, Wang H, Wang X, Huang H, Ding W, Jing R, Shi G, Zhu L. A proliferation-inducing ligand: A new biomarker for non-small cell lung cancer. Exp Lung Res. 2009;35:486–500. doi: 10.1080/01902140902759274. [DOI] [PubMed] [Google Scholar]
- 31.Sahai E, Astsaturov I, Cukierman E, DeNardo DG, Egeblad M, Evans RM, Fearon D, Greten FR, Hingorani SR, Hunter T, et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat Rev Cancer. 2020;20:174–186. doi: 10.1038/s41568-019-0238-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chen X, Song E. Turning foes to friends: Targeting cancer-associated fibroblasts. Nat Rev Drug Discov. 2019;18:99–115. doi: 10.1038/s41573-018-0004-1. [DOI] [PubMed] [Google Scholar]
- 33.Chen J, He D, Chen Q, Guo X, Yang L, Lin X, Li Y, Wu W, Yang Y, He J, et al. BAFF is involved in macrophage-induced bortezomib resistance in myeloma. Cell Death Dis. 2017;8:e3161. doi: 10.1038/cddis.2017.533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Manfroi B, McKee T, Mayol JF, Tabruyn S, Moret S, Villiers C, Righini C, Dyer M, Callanan M, Schneider P, et al. CXCL-8/IL8 produced by diffuse large B-cell lymphomas recruits neutrophils expressing a proliferation-inducing ligand APRIL. Cancer Res. 2017;77:1097–1107. doi: 10.1158/0008-5472.CAN-16-0786. [DOI] [PubMed] [Google Scholar]
- 35.Lowin T, Anssar TM, Bäuml M, Classen T, Schneider M, Pongratz G. Positive and negative cooperativity of TNF and Interferon-γ in regulating synovial fibroblast function and B cell survival in fibroblast/B cell co-cultures. Sci Rep. 2020;10:780. doi: 10.1038/s41598-020-57772-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mackay F, Tangye SG. The role of the BAFF/APRIL system in B cell homeostasis and lymphoid cancers. Curr Opin Pharmacol. 2004;4:347–354. doi: 10.1016/j.coph.2004.02.009. [DOI] [PubMed] [Google Scholar]
- 37.He B, Chadburn A, Jou E, Schattner EJ, Knowles DM, Cerutti A. Lymphoma B cells evade apoptosis through the TNF family members BAFF/BLyS and APRIL. J Immunol. 2004;172:3268–3279. doi: 10.4049/jimmunol.172.8.5128-c. [DOI] [PubMed] [Google Scholar]
- 38.Ni SZ, Cao HY, Chen Z, Zhu Y, Xu ZK. siRNA interference with a proliferation-inducing ligand gene in the Sgr-7901 gastric carcinoma cell line. Asian Pac J Cancer Prev. 2012;13:1511–1514. doi: 10.7314/APJCP.2012.13.4.1511. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.