

Abstract
Lactate, historically considered a waste product of anerobic metabolism, is a metabolite in whole-body metabolism needed for normal central nervous system (CNS) functions and a potent signaling molecule and hormone in the CNS. Neuronal activity signals normally induce its formation primarily in astrocytes and production is dependent on anerobic and aerobic metabolisms. Functions are dependent on normal dynamic, expansive, and evolving CNS functions. Levels can change under normal physiologic conditions and with CNS pathology. A readily combusted fuel that is sshuttled throughout the body, lactate is used as an energy source and is needed for CNS hemostasis, plasticity, memory, and excitability. Diffusion beyond the neuron active zone impacts activity of neurons and astrocytes in other areas of the brain. Barriergenesis, function of the blood-brain barrier, and buffering between oxidative metabolism and glycolysis and brain metabolism are affected by lactate. Important to neuroprotection, presence or absence is associated with L-lactate and heme oxygenase/carbon monoxide (a gasotransmitter) neuroprotective systems. Effects of carbon monoxide on L-lactate affect neuroprotection – interactions of the gasotransmitter with L-lactate are important to CNS stability, which will be reviewed in this article.
Keywords: astrocytes, biochemical interactions, CNS, gasotransmitters, microglia, neuropreservation, neuroprotection, oligodendrocytes
INTRODUCTION
Lactate (discovered in 1780, considered a waste product of metabolism) is a requisite molecule for whole body metabolism and normal central nervous system (CNS) development and functioning. It is the preferred substrate for energy purposes and a precursor of lipids in oligodendrocytes, astrocytes, and neurons during the perinatal period,1,2 but is preferentially associated with ischemia of the brain. Evidence now, however, indicates that lactate is found in the CNS under physiological conditions.3,4 Neuronal activity signals induce formation primarily in astrocytes. While it is used as an energy source, it is also needed for CNS hemostasis, plasticity, memory, and excitability and is now recognized as a necessary metabolite in the coordination of whole-body metabolism as a readily combusted fuel that is shuttled throughout the body, a potent signaling molecule, and a hormone.3,4,5,6,7 These functions are dependent on CNS requirements for normal dynamic, expansive, and evolving CNS physiological functions. Presence or absence is also important to CNS neuroprotection and is associated with L-lactate and heme oxygenase/carbon monoxide (CO) (a gasotransmitter) neuroprotective systems.8 Effects of CO on L-lactate affect neuroprotection - interactions of the gasotransmitter with L-lactate are important to CNS stability, which will be reviewed in this article. The following databases were systematically searched: MEDLINE, EMBASE, and Cochrane. Databases were searched from their inception to December 31, 2020.
SOURCES OF LACTATE IN THE CENTRAL NERVOUS SYSTEM
Considered a waste product of anerobic metabolism, lactate has known beneficial roles in the CNS and its production is not limited to anerobic metabolism. CNS concentration depends on serum lactate levels, oxygen availability, neuronal firing, degradation, and metabolic rate. Lactate serves as an important metabolic fuel and an intercellular messenger – its diffusion beyond the neuron active zone impacts activity of neurons and astrocytes in other areas of the brain.9,10,11 Contributing to neuronal metabolic support by transferring lactate through cytoplasmic myelinic channels and monocarboxylate transporters (MCTs), oligodendrocytes are active participants in this process – lactate is released by myelinating oligodendrocytes and then used by axons for generation of mitochondrial adenosine triphosphate.12,13 Oligodendrocytes are associated with the cell-to-cell lactate shuttle and the astrocyte-neuron lactate shuttle (lactate shuttling between cells producing lactate and cells consuming lactate is important to oligodendrocyte metabolism). These processes allow lactate to be used as a signaling molecule, energy source, and gluconeogenic precursor in the CNS.6,11 Lactate also affects barriergenesis and function of the blood-brain barrier (BBB),14 which is a buffer between oxidative metabolism and glycolysis,15 and may be the preferred fuel for brain metabolism.16 Lactate shuttles within and between cells and lactate uptake across the BBB suggests it is integral to CNS homeostasis although its transport across the BBB is rate-limited and lactate production within the brain reflects the needs of the CNS (metabolism and utilization is dynamic). Astrocyte-to-neuron lactate shuttling occurs during rest, but with excitation glycolysis of glucose to lactate (exceeding rate of mitochondrial fuel oxidation) in the neurons provides for increased energy needs.17,18 In the newborn mammalian brain, elevated blood lactate is utilized as a substrate and, in the adult resting brain with baseline blood lactate levels, about 10% of the brain energy needs are met by oxidation of lactate (can oxidize more as plasma lactate levels increase.)19,20 CNS lactate topography suggests region-specific biology/metabolism and is similar between individuals.21,22,23 Inherent to lactate homeostasis, elimination from the CNS is equally important – much of the lactate produced in the CNS during neuron activity is removed rather than being used as an energy source.
Uptake through the BBB
The highly vascularized BBB, a protective interface between the body and the CNS, selectively allows passage of substances, including lactate, into the brain. Initially identified by Paul Ehrlich24 (injected a dye into the bloodstream of mice and noticed that the dye did not enter the brain or spinal cord), the term BBB was introduced by Lewandowsky25 after studies showed that different substances injected into the ventricles of the brain resulted in neurologic symptoms that were not seen when injected intravenously. Electron microscopy showed that the brain endothelial cells construct a barrier between blood and CNS with a single layer of brain endothelial cells in contact with neurons, astrocytes, pericytes, and vascular smooth muscle cells. BBB function depends on signaling and crosstalk of this neurovascular unit.26,27 Tight junctions (barrier to passage of ions and molecules through the paracellular pathway and to the movement of proteins and lipids between the apical and the basolateral domains of the plasma membrane) and adherent junctions (allow epithelial cells to establish polarity with different proteins and lipids in the apical and basal plasma membranes) maintain the integrity of the BBB. Although it is best known for its barrier function of restricting transport of toxic and/or harmful substances from the blood to the brain, the BBB also has a carrier function that is responsible for the removal of metabolites and transport of nutrients to the brain. Its barrier function depends on the paracellular barrier, transcellular barrier, enzymatic barrier, and efflux transporters. Passive diffusion and specific transport proteins are necessary for its carrier function.28 Flow of blood affects BBB cell structure and function, increases BBB tightness, and affects endothelial cell differentiation.29 Clearance of substances from the CNS includes perivascular efflux, metabolism, and the BBB.30,31
L-lactate is considered a volume transmitter and can travel large distances from the site of production (i.e., muscle) to a site of consumption (i.e., CNS) with monocarboxylate transporters (MCTs) mediating diffusion of lactate between extracellular space and cells. Lactate passage across the BBB by nonsaturable diffusion or via MCTs in the plasma membrane – equilibration between lactate concentration in the blood and brain results (lactate then equilibrates with pyruvate via lactate dehydrogenase) – can be restricted.32 With normal plasma lactate levels, non-saturable diffusion can be as high as 50% of lactate transport from the blood to the CNS and, with high plasma lactate levels, near instantaneous equilibration occurs. MCTs carry L-lactate (most abundant monocarboxylate in the brain) across different cell membranes including the BBB – cotransport of monocarboxylate anion and proton occurs. Four carriers (MCTs 1–4) account for bidirectional, electroneural 1:1 co-transport of protons and monocarboxylic acids (i.e., lactate). MCT2 has the highest affinity for lactate – regionalization of MCTs 1–4 has been suggested to indicate involvement in different aspects of lactate metabolism and their production varies under different conditions. Accessory proteins (i.e., embigin, basigin, neuroplastin) are necessary for their activity.4,32,33,34,35,36 Efflux is by passive transport which is mediated by MCT1 which is present in both the abluminal and luminal membranes.30,37
Production of lactate in the brain
Important major metabolic pathways, catabolic and anabolic, necessary for CNS life/growth/division include gluconeogenesis/glycolysis, fatty acid β-oxidation, urea cycle, pentose phosphate pathway, oxidative phosphorylation, and the citric acid cycle. Glucose, the obligatory CNS energy substrate in mature humans, depends on glucose transport/metabolism, is mostly consumed by oligodendrocytes and astrocytes, and metabolites produced and released by astrocytes and oligodendrocytes are used by neurons as energy sources. The preferred energy substrate of the brain, lactate, is an important energy substrate for axons, is important in CNS health, and is an important source earlier in life and is now thought to be an important metabolite needed in the adult brain. Present in astrocytes, microglia, neurons, and oligodendrocytes, lactate production is via multiple pathways and its timely metabolism and elimination promotes CNS health and well-being – metabolic switching to lactate in the CNS has been linked to resistance to disease, stress, and injury. Metabolic processes in the CNS, dependent on the presence of multiple cell types and level of development/maturity, result in a pyruvate/lactate ratio that decreases with maturation and differs in regions of the brain. Astrocyte-neuron interactions, important to lactate metabolism, reflect dynamic needs and its production – glycolysis and glycogen metabolism are necessary for normal brain function, growth, and healing and can result in the production of lactate. CNS lactate production includes the following pathways: gluconeogenesis/glycolysis pathway, fatty acid β-oxidation, urea cycle, pentose phosphate pathway, oxidative phosphorylation, and citric acid cycle.4,7,38,39
Gluconeogenesis/glycolysis pathway
CNS generation of glucose from non-carbohydrate precursors (gluconeogenesis) is a multistep process – enzymes used in glycolysis may catalyze reversible reactions. There are, however, three irreversible reactions:
1) Conversion of pyruvate to phosphoenolpyruvate:
Pyruvate carboxylase: Pyruvate (cytosol) -> pyruvate (mitochondria) -> -> -> oxaloacetate (mitochondria) -> oxaloacetate (cytoplasm);
Phosphoenolpyruvate carboxykinase: Oxaloacetate (cytoplasm) ->->-> phosphoenolpyruvate;
2) Dephosphorylation of fructose-1,6 biphosphate:
Fructose 1,6-biphosphate (cytoplasm) ->->-> fructose-6 phosphate (cytoplasm);
3) Dephosphorylation of glucose 6-phosphate: Glucose-6 phosphate (cytoplasm) ->->-> glucose (cytoplasm).
Pyruvate kinase catalyzes the conversion of phosphoenolpyruvate to pyruvate. Pyruvate can then be converted to lactate via lactate dehydrogenase. Availability of enzymes varies between cells. For example, astrocytes exhibit significant 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase-3 activity, a key mechanism for regulating glycolysis and gluconeogenesis through synthesis or hydrolysis of fructose-2,6-bisphosphate. Preferential production of lactate via the gluconeogenesis/glycolysis pathway is dynamic and determined by the needs, metabolism, and health of the CNS.40,41,42,43,44,45
Fatty acid β-oxidation pathway
Lipids (classified into five major subcategories – fatty acids, triglycerides, sphingolipids, sterol lipids, and phospholipids) are important to normal CNS function and development and serve as bioactive molecules, energy susbstrates, acting as building blocks or a combination of these. Fatty acid imbalance affects CNS development and function. Fatty acids are the essential component of all of the lipids and consist of a carbon chain that ends in a carboxylic acid functional group (subclassified as short-chain fatty acids (2–4 carbons), medium-chain fatty acids (6–12 carbons), long-chain fatty acids (14–18 carbons), and very long-chain fatty acids with 18+ carbons.) If saturated, all of the carbons have hydrogen atoms and single bonds only exist between carbons whereas unsaturated fatty acids have carbon chains where a double bond has been introduced (mono- versus polyunsaturated defines whether there is more than one double bond.) Fatty acid β-oxidation results in the formation of acetyl-CoA as noted in the equation: β-oxidation of fatty acids->->->->->->->->Acetyl-CoA (Figure 1).
Figure 1.

Conversion of long-chain fatty acids to acetyl-CoA.
Note: Beta oxidation will proceed until only 2 carbon units remain. CoA: Coenzyme A; FAD: flavin adenine dinucleotide.
Mitochondrial β-oxidation of fatty acids results in energy – repeating sequence of four reactions catalyzed by acyl-CoA dehydrogenase, enoly-CoA thiolase, hydroxyacyl-CoA dehydrogenase, and ketoacyl-CoA thiolase. Acetyl-CoA results with a fatty acyl molecule that is two carbons shorter –reaction cycle repeats Figure 1. With an odd number of carbon atoms, propionyl-CoA is the final product. The acetyl-CoA enters the tricarboxylic acid cycle and pyruvate and, possibly, lactate may be generated. Propionyl-CoA is converted into succinyl-CoA (substrate for gluconeogenesis via oxaloacetate formation). Pyruvate and, possibly, lactate may be generated.46,47,48,49,50
Cytoplasmic fatty acyl CoA is converted to fatty acyl carnitine by carnitine acyl transferase I, an enzyme of the inner leaflet of the outer mitochondrial membrane. Fatty acyl carnitine is then transported by an antiport in exchange for free carnitine to the inner surface of the inner mitochondrial membrane. There carnitine acyl transferase II reverses the process, producing fatty acyl-CoA and carnitine. This shuttle mechanism is required only for longer chain fatty acids as follow:
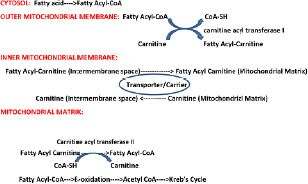
Medium- and short-chain fatty acids are carnitine-independent. They cross the mitochondrial membranes, and are activated in the mitochondrion. In the astrocyte and neuron mitochondria and cytosol, the four steps of β-oxidation include: dehydrogenation of the fatty acyl-CoA to make a trans double bond between α and β carbon (requires short, medium, and long chain acyl-CoA dehydrogenases, electron removed transferred to flavin adenine dinucleotide), hydration of the double bond, dehydrogenation of the β-hydroxyl group to a ketone (electron removed transferred to NAD+), acylation (addition of CoA and production of acetyl-CoA.

Urea cycle
While the urea cycle and the tricarboxylic acid cycle (TCA) are independent cycles, they are linked. Fumarate that is produced in the cycle is an intermediate in the TCA cycle, metabolized to malate, and then to pyruvate and lactate (Figure 2).
Figure 2.

Urea cycle and its relationship to Krebs cycle.
Urea cycle - Citrulline->->->argininosuccinate->->->arginine and fumarate
Pentose phosphate pathway
Taking place in the cytosol, the pentose phosphate pathway is anabolic more than catabolic, generates reducing equivalents, produces ribose 5-phosphate (needed for synthesis of nucleotides and nucleic acids) and erythrose 4-phosphate (needed for synthesis of aromatic amino acids), and is linked to glycolysis. During the oxidative phase (irreversible), glucose 6-phosphate is metabolized to ribulose 5-phosphate. The latter can be metabolized to fructose 6-phosphate during the nonoxidative phase (reversible.)
1) Conversion of glucose 6-phosphate to ribulose 5 – phosphate
Oxidative phase
glucose 6-phosphate->->->->->->->->->->->ribulose 5- phosphate
2) Conversion of ribulose 5-phosphate to lactate
Non-oxidative phase
ribulose 5-phosphate←→xylulose 5-phosphate←→fructose 6-phosphate
or
glyceraldehyde 3-phosphate ->->->pyruvate->->->lactate.55,56,57
Oxidative phosphorylation
Cellular respiration depends on glycolysis (cytosol) which results in pyruvate, pyruvate transformation into acetyl-CoA (mitochondria), the citric acid cycle (acetyl-CoA is modified in the mitochondria to produce energy precursors), and oxidative phosphorylation (or electron transport-linked phosphorylation) – process where electron transport from the energy precursors from the citric acid cycle leads to the phosphorylation of adenosine diphosphate producing ATP (occurs in the mitochondria). Uncoupling between glycolysis and oxidative phosphorylation involves the partitioning between pyruvate (the primary substrate for glucose-driven oxidative phosphorylation) and lactate. In the mitochondria, succinate is reduced to fumarate by Complex II. The latter enters the Krebs cycle and pyruvate (and then lactate via lactate dehydrogenase (LDH)) can be produced.58,59,60 Nitric oxide production also inhibits the mitochondrial respiratory chain at cytochrome c oxidase in astrocytes and the majority of glucose consumed is released as lactate.61
Citric acid cycle
Oxidation of acetyl-CoA to CO2 by the citric acid cycle is the central process in energy production. The process also results in the formation of intermediates which can be converted primarily to fatty acids, glucose derivatives, γ-aminobutyric acid (GABA), glutamate, glutamine, and aspartate. Balance between the two biochemical processes related to the cycle (anaplerosis and cataplerosis) is essential to normal CNS physiology.62 Pyruvate carboxylation is important for the net synthesis of glutamate, GABA and aspartate (anaplerosis) and is associated with cerebral activity - flux through pyruvate carboxylase is higher in the awake state compared to deep phenobarbital anaesthesia.63 However, the brain cannot fully metabolize 4- and 5-carbon molecules – they must be removed by cataplerosis, a process that may be linked to biosynthetic processes resulting in synthesis of fatty acids and amino acids and gluconeogenesis.
Glutamate seems to be the molecule connecting anaplerosis and cataplerosis in the Krebs cycle. Cytosolic malic enzyme and phosphoenolpyruvate carboxykinase in astrocytes converts malate and oxaloacetate into lactate which leaves the brain (blood or periventricular system.) Efflux of lactate allows for ongoing pyruvate carboxylation.64,65,66
Movement of lactate in the brain
Lactate shuttles
The lactate shuttle hypothesis describes the movement of lactate within and between cells and is based on the observation that lactate is formed and utilized continuously in diverse cells under both anaerobic and aerobic conditions. Interconversion of lactate and pyruvate occurs via lactate dehydrogenase and is produced at sites with high rates of glycolysis and glycogenolysis. Lactate plays an important role as a shuttle and can be shuttled to adjacent or remote sites and can be used as a gluconeogenic precursor or substrate for oxidation. The hypothesis also addresses the role of lactate in lipolytic control, redox signaling, and gene expression and affirms the role of lactate in cell signaling, endocrine/autocrine/paracrine link between oxidative and glycolytic metabolism, and in the delivery of gluconeogenic and oxidative substrates – L-lactate is considered the link between aerobic and glycolytic pathways (substrate for mitochondrial respiration and a product of glycolysis.) Glycolytic lactate producer cells provide lactate that is taken up by the blood and mitochondrial reticulum (oxidative lactate consumer cells.)67,68 The anion is metabolically available under aerobic conditions and is important in a complicated feedback loop – lactate production is promoted with decreased ATP levels which leads to the cells ability to promote ATP homeostasis. Redox state, concentration gradient, and/or pH gradient are known to drive these shuttles.6 Cell-to-cell lactate interactions are important to CNS function. There is a normal physiological function and relationship between astrocytes and neurons – the astrocyte–neuron lactate shuttle. An increase in synaptic activity or low glucose conditions result in transfer of lactate from astrocytes to neurons which sustains neuronal oxidative metabolism. Astrocytes express MCT4 (a low affinity transporter for lactate suggesting that lactate produced by glycolysis is exported.) Neurons express MCT2 (a high affinity transporter for lactate.) The hypothesis is that astrocytes produce lactate which is then taken up by neurons and oxidized for energy.69,70,71 There are other CNS cell-to-cell lactate shuttles as well and these include microglia and their preferential utilization of lactate under pathologic conditions, the activity of lactate shuttles in traumatic brain injury and neurodegenerative diseases and the astrocyte–microglia lactate shuttle.6,15,72,73
Cell-to-cell, when coupled to intracellular, lactate shuttles allow for the simultaneous presence of glycolytic (i.e., glucose to lactate) and oxidative (i.e., lactate to pyruvate to acetyl-CoA) pathways. This intracellular lactate pathway – lactate exchange and conversion into pyruvate maintaining the redox balance in the cytosol and mitochondria – is thought to exist between cytosol and mitochondria and between cytosol and peroxisomes.15,74
Glymphatic system
About 60% to 68% of the brain’s total water content is within the intracellular space and 32–40% in the extracellular space (12% to 20% in the interstitial fluid (ISF), 10% in the cerebrospinal fluid (CSF), 10% in the blood.) The CSF, formed by the choroid plexuses (no BBB), depends on osmotic and hydrostatic movement of fluid, crystalloids, and colloids from plasma into stroma. The glymphatic system, integral to CNS fluid balance and removal of waste, has glial-dependent perivascular channels formed by astroglial cells, is dependent on aquaporin (AQP) 4 water channels on astrocyte endfeet, has peri-venous clearance and peri-arterial influx pathways, is connected to the lymphatic system associated with the dura, cranial nerves, and vessels which are located at skull exits, and allows continuous ISF and CSF removal of potentially harmful molecular products. Activity is increased during general anesthesia and during sleep and is decreased with stroke, diabetes mellitus, Alzheimer’s disease, and during aging.75,76,77,78,79,80,81,82 CNS lactate (higher while awake than during sleep and associated with reduction with cerebral glycolysis), a biomarker of the sleep-wake cycle, is exported as lactate by the glymphatic system – brain lactate concentration being inversely correlated with glymphatic-lymphatic clearance and resulting lactate/pyruvate ratio (and NADH/NAD redox potential) – and decreases rapidly with the onset of sleep. Elimination of lactate during sleep combined with the lack of elimination during wakefulness results in an increase of redox state with wakefulness (relative to sleep.)83,84,85
ELIMINATION OF LACTATE FROM THE CENTRAL NERVOUS SYSTEM
Maintaining a normal lactate level in the CNS depends on its utilization versus its accumulation. While the resting brain releases lactate (release from various brain structures differs under basal conditions), this increases incrementally with activation of the brain.86 Total lactate clearance is determined by its metabolism in the CNS, excretion via capillaries, or excretion through the CSF – clearance systems can be via CSF, ISF within the CNS, and blood.86,87,88 Three major models of lactate elimination via brain fluids have been hypothesized: 1) meningeal lymphatics,89 the glymphatic system,83,90 and the intramural periarterial system.91,92
The lymphatic system is a part of the vascular system. Peripherally, plasma is filtered through the vascular endothelium of capillaries in the extracellular space with over 80% being reabsorbed back into venous vessels and the remainder in the extracellular fluid or ISF. ISF is then filtered back into the lymphatics which return it into the venous system. In the CNS, the volume of brain fluid is fixed, there is BBB restriction of fluid filtration from plasma into the brain, and there is no common agreement about ISF formation in the brain. The three models of ISF formation that have been proposed include production by brain metabolism, cerebral capillary secretion of solutes, and ISF is a fraction of recycled CSF (flows from choroid plexus into subarachnoid space and then into perivascular space where CSF merges with ISF that has been generated by cerebral capillaries) – lymphatic communication with brain fluids affects the clearance of lactate as well as that of waste products, bacteria, viruses, toxins, etc.93,94,95 The extensive meningeal lymphatic vessel network that helps with macromolecular clearance and immune cell trafficking in the CNS also communicates with the glymphatic system, a system that allows CNS perfusion by ISF and CSF, and complements intramural, periarterial clearance.96,97,98
EXCESS OR TOO LITTLE LACTATE
Maintenance of lactate levels is necessary for normal health and well-being – an excess or relatively too little can result in changes in body metabolism. Fluctuations are not usually due to low oxygen levels as hyperlactatemia can occur with normal tissue perfusion/oxygenation and relative hypolactatemia with poor tissue perfusion/oxygenation. Although lactate (the major gluconeogenic precursor, a signaling molecule, and a major energy source) production occurs normally during rest, it is increased with sepsis, pancreatitis, trauma, heart failure, etc.68,99,100,101,102 The resulting lactic acidosis, overproduction or underutilization of lactic acid with the body not being able to adjust to these changes, is associated with a buildup of acid and an imbalance of the bodies pH level. Elevated levels are used as a biomarker for risk and therapy and high levels are associated with an increased risk of death independent of organ failure and shock.103,104,105 Cellular metabolic reprogramming, metabolic inflexibility, and a decrease in cellular proliferation occur.106,107,108,109,110,111 Effects on the cardiovascular system can result in hyporesponsiveness of the vascular system to vasopressors and reduced cardiac contractility. NAD+/NADH ratio, production of reactive oxygen species, regulation of genes, release of vascular endothelial growth factor/interlukin-1/transforming growth factor-β, and activation of Sirtuins are affected.101,109,110,111,112 Severe lactic acidosis has been suggested as a prerequisite of brain infarction from complete ischemia. This may be due to elevated brain glucose metabolism to lactate and its effect on astrocytes as excessive lactate is considered to be detrimental to the CNS and can affect recovery from brain trauma, anoxia, epilepsy, and ischemia.102 However, metabolism and cellular interactions determine if lactate is a waste product or a useful substrate. While acute and chronic pathologies alter brain metabolism resulting in changes in lactate concentration, lactate is a preferred fuel over glucose in brain preparations.32,112,113 When there is an increase in blood lactate in exercising humans (or when exogenous lactate is supplied), lactate is the major gluconeogenic precursor and substitutes for glucose as an energy substrate – cerebral lactate uptake increases. Improved cognitive function in brain-injured rats given intravenous lactate therapy and rapid increases of lactate MCT protein expression in rat brains following traumatic brain injury suggest that lactate metabolism in humans following traumatic brain injury is significant to recovery. The roles for lactate in normal cerebral metabolism and with CNS pathology are being redefined. Lactate supports hepatic and renal gluconeogenesis which is important to brain metabolism, production by the body indirectly supplies glucose to the injured brain, isotopically labeled lactate is directly consumed and used by the injured human brain, and exogenous lactate infusion augments cerebral substrate supply when glycolysis is affected.113,114,115,116 Lactate changes, reflective of overall metabolic health, are affected by heme-oxygenase (and CO) and affect the health of the CNS.
CARBON MONOXIDE EFFECTS ON LACTATE IN THE CENTRAL NERVOUS SYSTEM
CNS extracellular lactate concentration increases with neuropathology as well as CNS activation - both result in significant release of lactate.117 The latter, which can be associated with excitotoxicity, may result in neuronal damage and death as excitotoxicity is a major component of CNS pathology.118,119,120,121,122 L-lactate (protects neurons against excitotoxicity, a regulator and metabolite in the CNS, regulates the expression of synaptic plasticity and neuroprotection genes in cortical neurons, promotes angiogenesis/immune escape/cell migration, serves as a glucose-sparing substrate, regulates cerebral blood flow, and plays an important role in learning and memory)123,124,125,126,127 acts as a signaling molecule in pathological states,123,124,125 is neuroprotective, and effects are directed at neurotransmission. CO has been found to modulate L-lactate levels in astrocytes providing evidence that the heme oxygenase and L-lactate neuroprotective systems interact.8
The relationship between CNS L-lactate and the heme oxygenase/CO system is complex. The cytoprotective properties of the heme oxygenases are attributed to the production of CO. Endogenous production originates from heme metabolism (at least 86%) and heme-independent sources which include iron-ascorbate-catalyzed lipid peroxidation of microsomal lipids and phospholipids, photo-oxidation of organic compounds, auto- and enzymatic oxidation of phenols, and reduction of cybochrome b5. Heme oxygenase isozymes catalyze the first and rate-limiting step in the degradation of heme to CO, iron, and biliverdin.
Neuroprotective effects of CO are at metabolic (metabolic support, regulation of local blood flow, glycogen synthesis/storage, formation of neuro-glial-vascular unit and glial-vascular interface), molecular (regulation of pH, water transport/homeostasis, neurotransmitter homeostasis, ion homeostasis), organ (effects on lymphatic system and BBB), cellular/network (synaptic plasticity, synaptogenesis/maintenance/elimination, neurogenesis, neuronal development/guidance, defining architecture of CNS), and systemic levels (affects sleep, regulates energy balance, important to chemosensing) levels. Interactions between CNS L-lactate and the heme oxygenase/CO systems affecting neuroprotection can occur at multiple levels.
Metabolic
Metabolic support
Energy metabolism supports cerebral function, has moment-to-moment dynamic ranges, involves essential functional and metabolic interactions between neurons and astrocytes, and is compartmentalized. Neurons and astrocytes are dependent on each other for metabolic support of the CNS. Formation of lactate by astrocytes is a response to neuronal stimulation – lactate supplements CNS energy provision during euglycemia, may contribute to an increase in ATP reserves (brain prioritizes regulation of its own ATP concentration), and is a primary tricarboxylic acid cycle substrate. The rate of oxidative metabolism in astrocytes can increase as much as the rate of neuronal metabolism in response to sensory stimulation. In this milieu, a standing lactate gradient exists with higher lactate concentration in astrocytes than neurons and neuronal lactate may be extruded to nearby neurons with lower lactate concentrations or the extracellular space. Astrocytic K+ uptake, but not astrocytic Na+-coupled glutamate uptake, is important for establishment of a neuron-astrocyte metabolic partnership. A decrease in the oxygen glucose index – accompanied with a stimulation of pyruvate formation (and, therefore, lactate formation) and astrocyte respiration – is also seen with neuronal activation.3,60,65,128,129
Astrocytes are targets for treating neurological disorders as neurons are outnumbered by non-neuronal cells (i.e., astrocytes) and astrocytes contain glycogen which can be transformed to L-lactate (see above). Increased local levels of CO may increase astrocyte L-lactate formation and, therefore, affect the expression of neurologic diseases.8 Studies are needed to further evaluate the effects of CO on astrocyte lactate formation (and on metabolic support) and the interplay between CO, astrocyte lactate formation, metabolic support, and neurologic diseases.
Regulation of local blood flow
Neurovascular coupling results in a local increase in blood flow to match energy demands and occurs physiologically and in disease at the capillary and arteriole levels (differ mechanistically.) At the capillary level, this is dependent on calcium signaling (calcium concentration changes in response to neuronal activity.) Arteriole dilation is dependent on N-methyl-D-aspartate (NMDA) receptor activation and nitric oxide synthesis.130 CO and lactate affect calcium signaling and NMDA receptors and, therefore, potentially local CNS blood flow and the latter may be dependent on CO concentration.
CO is known to be an endogenous vascular modulator functioning as both a vasoconstrictor and vasodilator and is highly protective to the vasculature. The vasoconstrictor action may be associated with the generation of reactive oxygen species.131 In the CNS, the gasotransmitter dilates cerebral arterioles. Vasodilatory stimuli (seizures, hypoxia, adenosine diphosphate, glutamatergic stimulation) result in an increase in CO. CO binds to smooth muscle cell Ca2+-activated K+ channel BKCa) channel-bound heme which leads to an increase in Ca2+ sparks-to-BKCa coupling (also binds directly to the BKCa channel at different locations) resulting in vasodilation. Inhibition of heme oxygenase affects dilation of the vasculature to these stimuli. Apoptosis and oxidant-generating pathways are also inhibited.132,133 Lactate may act as a dose-dependent regulator of cerebral microcirculation by affecting the hyperemic response, which may be related to a cytosolic redox impairment, and optimization of blood flow may interact with nitric oxide and arachidonic acid metabolites released by neuronal and glial cells during neural activity.109,134,135 As both CO and lactate affect calcium channels and NMDA, the relationship to each other, calcium channels/NMDA, and CNS local blood flow are important questions that have not been addressed. Further studies are needed.
Glycogen synthesis and storage
A polymer of glucose, glycogen is present in the CNS at low concentrations relative to the liver and skeletal muscle and, in developed brains in mammals under normal conditions, is located predominantly in astrocytes (has been found in embryonic neurons.) Synthesis is a multi-step process, is tightly regulated, and glucose transport across the BBB is needed. Glycogen synthase and the glycogen branching enzymes elongate glucose chains and introduce branch points. Astrocytes depleted of glycogen are not as successful at supporting neurons as are those not depleted of glycogen.136,137,138
Astrocyte glycogen metabolism is important for functioning of the CNS. The substance is too large to be released and travel between cells, but studies have shown that astrocytes release lactate into the media.1,3,7,8,19,22 Modulated by a number of factors, neuronal activity signals result in astrocyte release of lactate – formed predominantly in astrocytes from glucose or glycogen and transferred from astrocytes to neurons to match the neuronal energetic needs and to provide signals that modulate neuronal functions and important to CNS health and disease. CO modulates L-lactate levels (and, therefore, glycogen metabolism) in astrocytes and is being further studied.3,8,139,140
Formation of neuro-glial-vascular unit and glial-vascular interface
Composed of neurons, astrocytes, microglial cells, endothelial cells, pericytes, smooth muscle cells, and circulating blood cells, the neuro-glial-vascular unit interact to control synaptic communication, BBB function, local blood supply, neuronal development and surveillance/immune function. Models (Buxton-Want model of vascular dynamics, Hodgkin-Huxley formulation of neuronal membrane excitability, biophysical model of metabolic pathways) of the neuro-glial-vascular unit describe neural dynamics and propound that the energy inside of the neuron is dynamic and depends on neural activity – lactate in astrocytes can fuel neuronal activity – and local vasculature. The astrocyte-neuron lactate shuttle is predicted by these models – temporal dynamics of tissue lactate, tissue glucose and oxygen consumption, and of BOLD signal reported in human studies correctly predicted, transfer of lactate from astrocytes to neurons noted in response to activity, dynamics of extracellular lactate and oxygen as observed in vivo in rats correctly predicted, neuronal oxidative metabolism increased first upon activation with a subsequent delayed astrocytic glycolysis increase.141,142 This requires MCT. Nitric oxide has been shown to down-regulate MCT 1. The effect of CO on MCT’s and, therefore, lactate effects on the neuro-glial-vascular unit have not been studied.
Molecular
Regulation of pH
Normal functioning of the CNS is affected by pH as changes affect the tightly regulated micro-circuits. Several processes regulate H+ in the CNS – intracellular H+ sequestering, CO2 diffusion, H+ buffering, membrane transport of acid/base equivalents across cell membranes, carbonic anhydrase activity, monocarboxylic acid transporters, vacuolar-type proton ATPase, etc. Acid extrusion as well as Na+/H+ exchange, Na+-HCO3– cotransport, and Na+-dependent Cl–/HCO– exchange mediate pH levels.143,144,145,146 In the spinal cord, cells have two types of molecules in the cell membrane that detect pH levels with one reacting to acidic pH (acid sensing ion channel subunit 3) and the other to alkaline pH levels (PKD2L 1.) With neuronal activity, lactate is released and pH becomes more acidic. Lower pH and higher pH are inhibitory on motor activity via the secretion of somatostatin from nerve terminals of the pH-sensing central canal cells.147 Ion channel gating/conductances, metabolite exchange and neuronal excitability, synaptic transmission, and intercellular communication via gap junctions are dependent on pH and subsequent information processing. CNS lactate and CO can result in changes in pH. Normalization of pH can occur at multiple levels as noted above and both lactate and CO affect several of these processes. Studies to evaluate the interaction of lactate and CO on CNS pH have not been done.
Water transport and homeostasis
Water transport and homeostasis in the CNS is partly dependent on cotransporters – active and passive modes have been suggested. Numbers and types of cotransporters per cell and unit water permeability vary. Water channel proteins, AQPs, are from a large family of major intrinsic proteins that are integral to water transport and homeostasis in the human brain and mediate water flux between the four water compartments in the CNS. Eight AQPs are expressed in the brain – AQP1, AQP3, AQP4, AQP5, AQP7, AQP8, AQP9, and AQP11 with AQP1 and AQP4 expressed in the highest concentrations. Orthodox AQPs only conduct water while aquaglyceroporins also have the ability of conducting other small molecules. AQP9 is involved in water flux through plasma membranes and is also permeable to monocarboxylates (i.e., lactate). AQP4, the most abundant AQP in the brain, is concentrated in the astrocytic foot processes, is important to BBB defense, neuroplasticity, removal of waste, and homeostasis of water (transports water into the brain but also moves water out of the brain in times of edema).148,149,150
AQPs also conduct gas molecules. AQP1 has been associated with nitric oxide permeation when reconstituted in lipid vesicle and in vivo. Nitric oxide is produced by both neuronal nitric oxide synthase and endothelial nitric oxide synthase – AQP4 is found in the proximity of both neuronal nitric oxide synthase and endothelial nitric oxide synthase (exists in plasma membranes of end feet of astrocytes which surround endothelial cells of capillaries). AQP4 is more adapted for gas conduction when compared to AQP1.151,152,153,154 Whether AQPs can conduct gas molecules (and which gases) and whether AQP-mediated gas conduction is clinically important are not clear. Initial studies suggest that lactate and CO are probably affected by AQP’s. Further studies are needed to determine affects by AQPs on lactate and CO and interactions between lactate, CO and AQPs.
Neurotransmitter homeostasis
There are more than 40 neurotransmitters in the human CNS – excitatory (glutamate, acetylcholine, histamine, dopamine, norepinephrine, epinephrine), inhibitory (GABA, serotonin, dopamine), neuromodulators (dopamine, serotonin, acetylcholine, histamine, norepinephrine), and neurohormones (releasing hormones, oxytocin, vasopressin). Their homeostasis, ability of a cell/system of cells in the brain to identify a change and to respond with a compensatory response that restores baseline function, results as an interaction of several mechanisms including regulation of release of neurotransmitters by autoreceptors, non-synaptic production of the neurotransmitters, neurotransmitter vesicular release from the presynapse, uptake by transporters, and diffusion of the neurotransmitters. Neurotransmitter gradients exist and their maintenance is crucial for neurological processes – glia surrounding the synapse are a major effector of outcomes. Computational models have shown that specific glial configurations may be necessary, non-synaptic sources are needed to maintain extracellular concentrations of neurotransmitters, transporter and non-synaptic source densities can co-vary to provide tone on presynaptic autoreceptors, and synaptic glial configuration can be suggested for a given neurotransmitter and a given extracellular neurotransmitter concentration.155,156,157 CO (and nitric oxide) is a neurotransmitter as well. Not stored in synaptic vesicles, release of CO with depolarization results in activation of biosynthetic enzymes – heme-oxygenase 2 is activated by phosphorylation by casein kinase 2.158,159
The effects of lactate and CO on homeostasis of the CNS neurotransmitters continue to be studied. Lactate serves as a signal to neurons to release more noradrenaline and CO-mediated facilitation of CNS cell interactions affects neural circuits.160,161,162 The interaction between lactate and CO in affecting these responses is unknown and need to be studied.
Ion homeostasis
Ca2+
CO affects Ca2+ signaling (necessary for synaptic transmission, gene transcription, CNS excitation, and memory storage) and may control the switch between glucose and lactate utilization during CNS activation, synaptic activity, and with neuropathology. Maintenance of normal CNS function requires regulation of calcium homeostasis – CO regulation of calcium channels (calcium activated K+ – distributed in both excitable and non-excitable cells, highly sensitive to intracellular calcium concentrations and voltage, and able to decrease voltage-dependent Ca2+ entry through membrane hyperpolarization and serve as negative feedback regulators – and Ca2+ channel families) is necessary to normal CNS function. The mechanisms by which CO regulates the calcium channels are unclear, remain controversial, and require further study. Telezhkin et al.163 found that cysteine residue 911 in the C terminal tail of human BKCaα subunit is important for activation by CO. Bak et al.164 proposed a compartmentalized Ca2+-induced limitation of the malate-aspartate shuttle that defines pre- and post-synaptic compartments metabolizing glucose and glucose plus lactate in which the latter displays a positive correlation between oxidative metabolism of glucose and Ca2+ signaling. Lactate is essential to calcium homeostasis as well. Takata and colleagues165 determined that support of synaptic function by lactate utilization is associated with activation of NMDA and L-type calcium channels. Studies to determine the relationship between lactate and CO on calcium homeostasis are needed.
Cl–
Chloride (Cl–) homeostasis involves multiple complex interactions and is critical for synaptic inhibition, volume regulation, and determines effects of glycine-mediated and GABA neurotransmission.166 Absence/presence and function of chloride transporters and channels (differ in brain cell types) affect direction of Cl– fluxes and cell physiology.
GABA, an excitatory neurotransmitter during CNS development is inhibitory in the adult. The switching of GABA from excitatory to inhibitory has been correlated with the intracellular concentration of Cl– – in early development intracellular Cl– is higher than extracellular concentrations, but in adults intracellular Cl– is lower than extracellular Cl–. When GABA binds to GABAA receptors, the Cl– ion channel opens, Cl– influxes, and postsynaptic membrane hyperpolarizes with inhibition of the Ca2+ influx of the postsynaptic neuron.167 The role switch is related to the expression of cation/chloride cotransporters NKCC1 and KCC2 and parallels the maturation of the CNS. Inhibition by GABA depends on Cl– influx and its continuation on Cl– extrusion from the cell (electrochemical Cl– gradient does not get depleted) – in the adult CNS, KCC2 is crucial for the inhibitory effects of GABA (and glycine) in synaptic circuits.168
Cation/chloride cotransporters have fundamental roles in differentiation, disease, neuronal proliferation, damage, recovery, and functions associated with plasmalemmal receptor- and channel-mediated effects (i.e., synaptic plasticity). These ion transporters depend on the electrochemical force of Na+ and K+ ions which interact with the Na+-K+ ATPase and its modulators – regulation of NKCC1 and KCC2 is by insulin-like growth factor, brain-derived neurotrophic factor, and cystic fibrosis transmembrane conductance regulator, and by phosphorylation of enzymes.169 Structure and mechanism of these two cation/chloride cotransporters determine electroneutral transport of sodium, chloride, and/or potassium across membranes and, thereby, impact ion absorption and secretion, maintenance of ion homeostasis, and regulation of cell volume – ion-translocation pathways, key residues for transport activity, and ion-binding sites have been defined.170,171 These ion transporters determine GABA actions – lactate has been associated with GABA release and CO (as an excitatory molecule) effects in the hypothalamic supraoptic nucleus are dependent on GABA.172,173 These relationships require further studies defining interactions between Cl–, GABA, CO, and lactate on health and disease of the CNS.
K+
Four main ions fluctuate with rhythmic firing in the CNS. These include Ca2+, Na+, Cl–, and K+. Synergy between these ions results in an oscillatory activity. Astrocytes regulate neuronal firing by affecting extracellular concentrations. Extracellular K+ levels in the CNS are low relative to intracellular levels and the gradient is important for synaptic communication and electrical signaling (with neuronal activity, a transient rise in extracellular levels is seen). If homeostasis is not maintained, wide-spread depolarization of CNS cells can occur and compromise normal synapses, neuronal firing, and uptake of neurotransmitters.174,175,176,177
Neuronal discharge results in the activation of the voltage-gated potassium channels with K+ moving to the extracellular space. The accumulating K+ is taken up by neighboring astrocytes and redistributed through the syncytium via the activation of gap junctions.178,179 In astrocytes, glycogenolysis is needed for the uptake of increased extracellular K+.180,181 Kir channels (seven subfamilies with each having multiple members) and the Na+-K+ pump are required for regulation of the extracellular K+.174 Kir channels establish and regulate the resting membrane potential of excitable cells and, in astrocytes, affect potassium buffering (and, therefore, neuronal excitability.)182 In the CNS, Kir4.1 is a modulator of brain-derived neurotrophic factor. Dysfunction of astrocytic Kir4.1 channels facilitates its expression in astrocytes (activating the Ras/Raf/mitogen-activated protein kinase kinase/extracellular signal-regulated kinase pathway) and may be linked to psychiatric disorders.183,184
The relationship of CO and lactate to Kir channels and possibly brain-derived neurotrophic factor in the CNS has not been defined. CO, however, inhibits Kir channels in cardiomyocytes as does H2S.185,186 The effects of gasotransmitters on Kir channels in the CNS have not been studied. Studies are needed to evaluate CO effects on Kir channels and brain-derived neurotrophic factor in the brain and the role of lactate in these metabolic processes (glycogenolysis is important).
Organ
Effects on the lymphatic system
CNS immune privilege and drainage are affected by the relationship between the CNS glymphatic system (system that allows CNS perfusion by the CSF and ISF) and meningeal lymphatics (major role in drainage of ISF, CSF, CNS-derived molecules, and immune cells from CNS and meninges to peripheral lymph nodes). Meningeal lymphatic vessels, also important in exchange of soluble contents between the CSF and ISF, are keys to removing cellular waste, solutes, and immune traffic from the CNS.97,187,188 Connection between the two systems allows continuous flow of CSF that perfuses the brain to be drained to peripheral nodes.96 Lactate is partially cleared from the CNS by these two systems. While heme oxygenase is involved in subarachnoid hemorrhage and its clearance through meningeal lymphatics,189 interactions between lactate clearance, CO, and the CNS lymphatic systems have not been defined and requires study.
Effects on the BBB
BBB integrity is a prerequisite of normal CNS health. Its disruption, as with stroke, is associated with cerebral edema and neurologic deficiency.190,191 This disruption has been shown to be alleviated by CO-releasing molecule-3, which reduces Evans blue leakage and brain edema, increases expression of platelet derived growth factor receptor β, Zonula occludens-1, and laminin, decreases the expression of matrix metallopeptidase-9, thus protecting pericyptes, tight junction proteins, and matrix proteins.192 The heme oxygenase-1 pathways are also important to the migration and differentiation of endothelial progenitor cells important to BBB integrity.193,194 Lactate also affects barriergenesis and functioning of BBB.14 Interactions between CO and lactate on the BBB have not been defined and need to be studied.
Cellular and network
Synaptic plasticity
Synaptic plasticity is important for maintaining homeostasis in the body, for sharing essential information among neurons, and for regulating synaptic transmission or electrical signal transduction to neuronal networks. Upregulation or downregulation of CO affects this plasticity and, therefore, neuropatholgies.195,196 Lactate is also a signaling molecule in synaptic plasticity – L-lactate stimulates the expression of synaptic plasticity-related genes such as Arc, c-Fos, and Zif268 through a mechanism involving NMDA receptor activity and its downstream signaling cascade extracellular signal-regulated kinase 1/2.129,197,198 Inhibition of the latter increases synaptic vesicle exocytosis by increasing calcium influx through L-type calcium channels.199 Downregulation of mitogen-activated protein kinase kinase/extracellular signal-regulated kinase 1/2 signaling pathways by CO has been suggested as a possible mechanism.200,201 The interactions between CO and lactate and their impact on synaptic plasticity require further study.
Synaptogenesis/maintenance/elimination
Changes in brain structure and organization as we experience, learn, and adapt is neuroplasticity (continues throughout life). Connections within the brain are constantly becoming stronger or weaker. This intricately regulated and multi-step process is defined by neuronal synapse formation/maturation and depends on key regulators, occurs simultaneously in different brain regions and between different types of neurons (a single neuron receives thousands of synaptic inputs), and are affected by extrinsic input, neuronal activity, intrinsic signaling pathways, and sensory experience.
Changes in synaptic or neuroplasticity are associated with multiple neuropsychiatric conditions. CO is important to restoring neuroplasticity in the CNS and in structuring synaptic plasticity and is being evaluated as a neurotherapeutic.194,195 The biochemical changes that occur with CO is not well defined and being further evaluated. Lactate is a regulator of synaptic genes and affects synaptogenesis, maintenance, and elimination.70,194,202 The relationship between CO and lactate on synaptogenesis/maintenance/elimination has not been defined.
Neurogenesis
The process by which new neurons are formed in the brain, neurogenesis, is crucial when an embryo is developing and continues in certain brain regions after birth and throughout our lifespan. Diversity of neurons results from regulated neurogenesis during embryonic development – neural stem cells (complex identity/functions ranging between quiescence/activation/intermediary subtypes) differentiate and become a specialized cell type at specific times and regions in the CNS and are affected by external factors. Mitochondrial dynamics are now known to be important during this process and neural stem cell fate decisions – mitochondria-regulated energetics is linked to neuronal development.
Integral to this process, mitochondria are central to determining replication and differentiation of these precursor cells in the CNS. Transition to a more differentiated cell type is accompanied by downregulation of glycolysis and fatty acid oxidation pathways and mitochondrial biogenesis.203,204,205,206 Lactate levels affect neurogenesis as does the presence of CO.116,207,208,209,210,211 Studies are needed to further evaluate the interactions of lactate and CO on neurogenesis (and effects on neuropathologies).
Neuronal development and guidance
The human brain contains more than 1 × 1011 neurons and each neuron connects to other neurons – axons and dendrites grow out, interact with other cells, and selectively protrude to create a functional, integrated, coordinated, working network. Initiated from the late embryonic stage and defining neural progenitor differentiation, neuronal development is determined by axon targeting/growth, dendrite maturation, synapse formation requiring integration of intrinsic genetic programs and the extrinsic factors, and chemoaffinity. Synaptic connections, continuously formed and eliminated, determine neural circuit plasticity.212,213,214,215
Pruning of dendrites and axons to mature neural circuitry is necessary for normal CNS health. Within the first 2 years of life (in humans), more than half of the connections formed in utero are eliminated. Pruned neurites are important for pre-remodeling networks, may serve as pioneering neurons that can guide other neurons, eliminate initially formed extensive connections made in the earlier wiring process, eliminate excessive branches, and simplify developmental programs.216,217,218,219 Microglia (constitute up to 10–15% of all cells in mammalian CNS) are associated with synapse elimination and influence synaptic strength – neuronal activity can directly activate microglia (detect functional state of neuron.) As the CNS matures, microglia localize in different regions throughout the brain. This is thought to be due to the microenvironment and functional differences – classical activation and alternative activation related to anti-inflammatory reactions and tissue remodeling.
Microglia monitor CNS parenchyma and affect clearance of cells in the healthy brain, neuronal excitability, neurogenesis, and synaptic activity. Phenotype can change from surveillant to pro- or anti-inflammatory in response to conditions. Upregulation of specific pathways based on stimuli can alter mitochondrial metabolism – changes in metabolism are seen with activation of microglia. CO affects microglia respiration and depends on the concentration.220,221 A CO-releasing molecule 3 affects microglia activation.222 Microglial metabolism (including lactate metabolism) is affected with neuroinflammation.222,223 Interaction between these two systems with pruning and neurodegenerative diseases has not been defined.
Defining the architecture of the CNS
Brain architecture continues to develop over time and is an ongoing process that begins before birth and continues into adulthood. It is comprised of billions of connections between individual neurons across different areas of the brain – simpler neural connections and skills form first followed by more complex circuits and skills – and is dependent on the interaction of genes and experiences. Whole brain volume changes throughout life – growth occurs during childhood and into adolescence, a gradual volume decrease is then seen and possible more growth or no tissue loss between around 18 and 35 years of age, followed by volume loss which increases with age. Brain reference atlases to identify the complex anatomical architecture of the brain help to link structure to function.224,225
After birth, connections continue to be modulated based on environment and experiences with genes affecting the building of the different cellular membranes and defining the molecules and ions that are used for the functioning CNS. Synapses and microcircuits as well as neurons, local circuits, and systems/pathways are affected. Experiences are crucial to the development of brain architecture and affect genes – genes can be turned on or off.226,227,228
Ability to change the architecture of the brain in response to experiences decreases over time. Anatomical architecture and structural/functional networks affect functional interactions in the brain – changes of structural connectivity are paralleled by changes in functional connectivity and characterize brain changes in aging.228 Rise in brain lactate is seen with aging of the brain - the mechanism varies with different brain regions (LDH status is important.)229,230 The change architecture of the aging brain is under oxidative stress – heme oxygenase genes are induced. CO (products of reaction catalyzed by heme oxygenase) has profound effects on the CNS.231,232 The roles and interactions of lactate and CO on changing CNS architecture needs further study.
Systemic level
Effects on sleep
Lactate is a biomarker of sleeping – lactate concentrations remain higher during waking relative to sleep and with forced sleep deprivation, increase with waking, and decrease within minutes of sleep.84,233,234 During sleep, there is decreased ISF concentration, decreased production, and increased perivascular clearance of lactate.37 Activation of the glymphatic system with change-of-state is associated with lowering CNS lactate during sleep.82 This changes the lactate/pyruvate ratio (and, therefore, the NADH/NAD redox potential).83 In those patients with obstructive sleep apnea, exhaled CO correlates with hypoxia during sleep,235 postsleep circulating CO level is increased initially,236 and venous lactate is increased (improved with continuous positive airway pressure).237,238 The proinflammatory transcription factor nuclear factor kappaB (affected by lactate and CO) is also increased.239 Interactions between CO, lactate, and nuclear factor kappaB during sleep and in obstructive sleep apnea need further studies.
Regulation of energy balance
The brain is integral to maintaining energy balance – the hypothalamic nuclei and the brainstem, main brain areas for energy balance regulation, are affected by nonhomeostatic and homeostatic circuits (interrelated and integrated). Neural circuits in the CNS process information regarding food consumption, food, food-related cues, and feeding and energy balance are associated with higher brain functions and degenerative processes.240,241,242 5′ adenosine monophosphate-activated protein kinase (AMPK), metabolic regulator of the entire organism, senses AMP:ATP and/or adenosine diphosphate/ATP ratios and maintains energy balance by promoting ATP production by increasing the activity/expression of proteins involved in catabolism and conserving ATP by switching off biosynthetic pathways. Selective activation of hypothalamic AMPK negates hypothalamic glucose/lactate-sensing mechanisms.243 Activated AMPK also stimulates heme oxygenase-1 gene expression and elevates the nuclear factor erythroid 2-related factor 2/heme oxygenase-1 signaling axis.244,245 Further studies are needed to define the relationships between AMPK, lactate, and CO associated with regulation of energy balance.
Chemosensing
Chemosensing (generation of a response to the presence of a chemical stimulus) is a fundamental sensory function and depends on a complex combination of receptor-mediated transmembrane signals, lipid modifications, protein translocations, and differential activation/deactivation of membrane-bound and cytosolic components. Some are mediated by somatosensory nerve endings of the trigeminal, glossopharyngeal, and vagus nerves – chemicals activate chemosensory receptors via mechanisms distinct from other sensory systems and the nerve endings can trigger a range of sensations and reflexes. Overlapping sensitivity of individual ion channels to chemical agonist can result in combinatorial activation of different classes of nerve endings by a single chemical. Complex interplay between chemical agonists and chemosensory receptors allow for chemosensory mimicry by chemically distinct agents. Multiple areas of the CNS are involved in eliciting a response to a particular chemical stimulus.
Chemosensory mechanisms may be an important causative factor in some neurologic diseases, but studies to evaluate the roles of lactate and/or CO in chemosensing are lacking. However, lactate infusion is a provocative challenge for panic disorder and is thought to be related to chemosensory mechanisms.246,247,248 CO effects on this are unknown. Studies to evaluate lactate and CO interactions on chemosensing are needed.
SUMMARY
Lactate is no longer considered “just a waste product of metabolism.” Overall and CNS health are affected by its presence or absence and its biochemical properties. CO is now known to be critical to the production of lactate and its roles (as a fuel, hormone, and/or signaling molecule) in the CNS. The neuroprotective ability of L-lactate partially depends on the integrity of the heme-oxygenase system and, in particular, CO. Further evaluation is needed of the L-lactate/heme-oxygenase/CO interactions to better understand the effects on its application in CNS health and neuropathologies.
Footnotes
Conflicts of interest
The author has no conflicts of interest.
Financial support
None.
Copyright license agreement
The Copyright License Agreement has been signed by the author before publication.
Plagiarism check
Checked twice by iThenticate.
Peer review
Externally peer reviewed.
REFERENCES
- 1.Bouzier-Sore AK, Voisin P, Bouchaud V, Bezancon E, Franconi JM, Pellerin L. Competition between glucose and lactate as oxidative energy substrates in both neurons and astrocytes: a comparative NMR study. Eur J Neurosci. 2006;24:1687–1694. doi: 10.1111/j.1460-9568.2006.05056.x. [DOI] [PubMed] [Google Scholar]
- 2.Medina JM, Tabernero A. Lactate utilization by brain cells and its role in CNS development. J Neurosci Res. 2005;79:2–10. doi: 10.1002/jnr.20336. [DOI] [PubMed] [Google Scholar]
- 3.Magistretti PJ, Allaman I. Lactate in the brain: from metabolic endproduct to signalling molecule. Nat Rev Neurosci. 2018;19:235–249. doi: 10.1038/nrn.2018.19. [DOI] [PubMed] [Google Scholar]
- 4.Proia P, Di Liegro CM, Schiera G, Fricano A, Di Liegro I. Lactate as a metabolite and a regulator in the central nervous system. Int J Mol Sci. 2016;17:1450. doi: 10.3390/ijms17091450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Adeva-Andany M, López-Ojén M, Funcasta-Calderón R, et al. Comprehensive review on lactate metabolism in human health. Mitochondrion. 2014;17:76–100. doi: 10.1016/j.mito.2014.05.007. [DOI] [PubMed] [Google Scholar]
- 6.Brooks GA. The science and translation of lactate shuttle theory. Cell Metab. 2018;27:757–785. doi: 10.1016/j.cmet.2018.03.008. [DOI] [PubMed] [Google Scholar]
- 7.Dienel GA. The metabolic trinity, glucose-glycogen-lactate, links astrocytes and neurons in brain energetics, signaling, memory, and gene expression. Neurosci Lett. 2017;637:18–25. doi: 10.1016/j.neulet.2015.02.052. [DOI] [PubMed] [Google Scholar]
- 8.Horvat A, Vardjan N, Zorec R. Targeting astrocytes for treating neurological disorders: carbon monoxide and noradrenaline-induced increase in lactate. Curr Pharm Des. 2017;23:4969–4978. doi: 10.2174/1381612823666170622112734. [DOI] [PubMed] [Google Scholar]
- 9.Ames A., 3rd CNS energy metabolism as related to function. Brain Res Brain Res Rev. 2000;34:42–68. doi: 10.1016/s0165-0173(00)00038-2. [DOI] [PubMed] [Google Scholar]
- 10.Baltan S. Can lactate serve as an energy substrate for axons in good times and in bad, in sickness and in health? Metab Brain Dis. 2015;30:25–30. doi: 10.1007/s11011-014-9595-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Barros LF. Metabolic signaling by lactate in the brain. Trends Neurosci. 2013;36:396–404. doi: 10.1016/j.tins.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 12.Philips T, Rothstein JD. Oligodendroglia: metabolic supporters of neurons. J Clin Invest. 2017;127:3271–3280. doi: 10.1172/JCI90610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Saab AS, Tzvetanova ID, Nave KA. The role of myelin and oligodendrocytes in axonal energy metabolism. Curr Opin Neurobiol. 2013;23:1065–1072. doi: 10.1016/j.conb.2013.09.008. [DOI] [PubMed] [Google Scholar]
- 14.Salmina AB, Kuvacheva NV, Morgun AV, et al. Glycolysis-mediated control of blood-brain barrier development and function. Int J Biochem Cell Biol. 2015;64:174–184. doi: 10.1016/j.biocel.2015.04.005. [DOI] [PubMed] [Google Scholar]
- 15.Brooks GA. Cell-cell and intracellular lactate shuttles. J Physiol. 2009;587:5591–5600. doi: 10.1113/jphysiol.2009.178350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Smith D, Pernet A, Hallett WA, Bingham E, Marsden PK, Amiel SA. Lactate: a preferred fuel for human brain metabolism in vivo. J Cereb Blood Flow Metab. 2003;23:658–664. doi: 10.1097/01.WCB.0000063991.19746.11. [DOI] [PubMed] [Google Scholar]
- 17.Ashrafi G, Wu Z, Farrell RJ, Ryan TA. GLUT4 mobilization supports energetic demands of active synapses. Neuron. 2017;93:606–615. doi: 10.1016/j.neuron.2016.12.020. e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Díaz-García CM, Mongeon R, Lahmann C, Koveal D, Zucker H, Yellen G. Neuronal stimulation triggers neuronal glycolysis and not lactate uptake. Cell Metab. 2017;26:361–374.e4. doi: 10.1016/j.cmet.2017.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Boumezbeur F, Petersen KF, Cline GW, et al. The contribution of blood lactate to brain energy metabolism in humans measured by dynamic 13C nuclear magnetic resonance spectroscopy. J Neurosci. 2010;30:13983–13991. doi: 10.1523/JNEUROSCI.2040-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pellerin L. Food for thought: the importance of glucose and other energy substrates for sustaining brain function under varying levels of activity. Diabetes Metab. 2010;36(Suppl 3):S59–63. doi: 10.1016/S1262-3636(10)70469-9. [DOI] [PubMed] [Google Scholar]
- 21.Grist JT, McLean MA, Riemer F, et al. Quantifying normal human brain metabolism using hyperpolarized [1-(13)C] pyruvate and magnetic resonance imaging. Neuroimage. 2019;189:171–179. doi: 10.1016/j.neuroimage.2019.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lee CY, Soliman H, Geraghty BJ, et al. Lactate topography of the human brain using hyperpolarized (13)C-MRI. Neuroimage. 2020;204:116202. doi: 10.1016/j.neuroimage.2019.116202. [DOI] [PubMed] [Google Scholar]
- 23.Miloushev VZ, Granlund KL, Boltyanskiy R, et al. Metabolic imaging of the human brain with hyperpolarized (13)C pyruvate demonstrates (13)C lactate production in brain tumor patients. Cancer Res. 2018;78:3755–3760. doi: 10.1158/0008-5472.CAN-18-0221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ehrlich P. Das Sauerstaff-Bedürfriss des Organismus: eine Farbenanalytische Studie. Berlin: Hirschwald; 1885. [Google Scholar]
- 25.Lewandowsky M. Zur Lehre von der Cerebrospinalflüssgkeit. Z Clin Med. 1990;40:480–494. [Google Scholar]
- 26.Banks WA. From blood-brain barrier to blood-brain interface: new opportunities for CNS drug delivery. Nat Rev Drug Discov. 2016;15:275–292. doi: 10.1038/nrd.2015.21. [DOI] [PubMed] [Google Scholar]
- 27.Chow BW, Gu C. The molecular constituents of the blood-brain barrier. Trends Neurosci. 2015;38:598–608. doi: 10.1016/j.tins.2015.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wilhelm I, Fazakas C, Krizbai IA. In vitro models of the blood-brain barrier. Acta Neurobiol Exp (Wars) 2011;71:113–128. doi: 10.55782/ane-2011-1828. [DOI] [PubMed] [Google Scholar]
- 29.Santaguida S, Janigro D, Hossain M, Oby E, Rapp E, Cucullo L. Side by side comparison between dynamic versus static models of blood-brain barrier in vitro: a permeability study. Brain Res. 2006;1109:1–13. doi: 10.1016/j.brainres.2006.06.027. [DOI] [PubMed] [Google Scholar]
- 30.Hladky SB, Barrand MA. Metabolite clearance during wakefulness and sleep. Handb Exp Pharmacol. 2019;253:385–423. doi: 10.1007/164_2017_37. [DOI] [PubMed] [Google Scholar]
- 31.Knudsen GM, Pettigrew KD, Patlak CS, Paulson OB. Blood-brain barrier permeability measurements by double-indicator method using intravenous injection. Am J Physiol. 1994;266:H987–999. doi: 10.1152/ajpheart.1994.266.3.H987. [DOI] [PubMed] [Google Scholar]
- 32.Bergersen LH. Lactate transport and signaling in the brain: potential therapeutic targets and roles in body-brain interaction. J Cereb Blood Flow Metab. 2015;35:176–185. doi: 10.1038/jcbfm.2014.206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Morland C, Lauritzen KH, Puchades M, et al. The lactate receptor, G-protein-coupled receptor 81/hydroxycarboxylic acid receptor 1: Expression and action in brain. J Neurosci Res. 2015;93:1045–1055. doi: 10.1002/jnr.23593. [DOI] [PubMed] [Google Scholar]
- 34.Ovens MJ, Manoharan C, Wilson MC, Murray CM, Halestrap AP. The inhibition of monocarboxylate transporter 2 (MCT2) by AR-C155858 is modulated by the associated ancillary protein. Biochem J. 2010;431:217–225. doi: 10.1042/BJ20100890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Takado Y, Cheng T, Bastiaansen JAM, et al. Hyperpolarized (13)C magnetic resonance spectroscopy reveals the rate-limiting role of the blood-brain barrier in the cerebral uptake and metabolism of l-lactate in vivo. ACS Chem Neurosci. 2018;9:2554–2562. doi: 10.1021/acschemneuro.8b00066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wilson MC, Kraus M, Marzban H, et al. The neuroplastin adhesion molecules are accessory proteins that chaperone the monocarboxylate transporter MCT2 to the neuronal cell surface. PLoS One. 2013;8:e78654. doi: 10.1371/journal.pone.0078654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hladky SB, Barrand MA. Elimination of substances from the brain parenchyma: efflux via perivascular pathways and via the blood-brain barrier. Fluids Barriers CNS. 2018;15:30. doi: 10.1186/s12987-018-0113-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sobral-Monteiro-Junior R, Maillot P, Gatica-Rojas V, et al. Is the “lactormone” a key-factor for exercise-related neuroplasticity? A hypothesis based on an alternative lactate neurobiological pathway. Med Hypotheses. 2019;123:63–66. doi: 10.1016/j.mehy.2018.12.013. [DOI] [PubMed] [Google Scholar]
- 39.Zhang W, Guo C, Jiang K, Ying M, Hu X. Quantification of lactate from various metabolic pathways and quantification issues of lactate isotopologues and isotopmers. Sci Rep. 2017;7:8489. doi: 10.1038/s41598-017-08277-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Adeva M, González-Lucán M, Seco M, Donapetry C. Enzymes involved in l-lactate metabolism in humans. Mitochondrion. 2013;13:615–629. doi: 10.1016/j.mito.2013.08.011. [DOI] [PubMed] [Google Scholar]
- 41.Cerdán S. Twenty-seven years of cerebral pyruvate recycling. Neurochem Res. 2017;42:1621–1628. doi: 10.1007/s11064-017-2173-4. [DOI] [PubMed] [Google Scholar]
- 42.Daikhin Y, Yudkoff M. Compartmentation of brain glutamate metabolism in neurons and glia. J Nutr. 2000;130:1026S–1031S. doi: 10.1093/jn/130.4.1026S. [DOI] [PubMed] [Google Scholar]
- 43.Hertz L, Dienel GA. Energy metabolism in the brain. Int Rev Neurobiol. 2002;51:1–102. doi: 10.1016/s0074-7742(02)51003-5. [DOI] [PubMed] [Google Scholar]
- 44.Melkonian EA, Asuka E, Schury MP. Physiology, gluconeogenesis. StatPearls. Treasure Island (FL): StatPearls Publishing; 2021. [PubMed] [Google Scholar]
- 45.Yip J, Geng X, Shen J, Ding Y. Cerebral gluconeogenesis and diseases. Front Pharmacol. 2016;7:521. doi: 10.3389/fphar.2016.00521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.de Carvalho C, Caramujo MJ. The various roles of fatty acids. Molecules. 2018;23:2583. doi: 10.3390/molecules23102583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hofmann K, Rodriguez-Rodriguez R, Gaebler A, Casals N, Scheller A, Kuerschner L. Astrocytes and oligodendrocytes in grey and white matter regions of the brain metabolize fatty acids. Sci Rep. 2017;7:10779. doi: 10.1038/s41598-017-11103-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lei E, Vacy K, Boon WC. Fatty acids and their therapeutic potential in neurological disorders. Neurochem Int. 2016;95:75–84. doi: 10.1016/j.neuint.2016.02.014. [DOI] [PubMed] [Google Scholar]
- 49.Panov A, Orynbayeva Z, Vavilin V, Lyakhovich V. Fatty acids in energy metabolism of the central nervous system. Biomed Res Int 2014. 2014 doi: 10.1155/2014/472459. 472459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Schönfeld P, Wojtczak L. Short- and medium-chain fatty acids in energy metabolism: the cellular perspective. J Lipid Res. 2016;57:943–954. doi: 10.1194/jlr.R067629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ipata PL, Pesi R. Understanding the interrelationship between the synthesis of urea and gluconeogenesis by formulating an overall balanced equation. Adv Physiol Educ. 2017;41:286–290. doi: 10.1152/advan.00180.2016. [DOI] [PubMed] [Google Scholar]
- 52.Pesi R, Balestri F, Ipata PL. Metabolic interaction between urea cycle and citric acid cycle shunt: A guided approach. Biochem Mol Biol Educ. 2018;46:182–185. doi: 10.1002/bmb.21099. [DOI] [PubMed] [Google Scholar]
- 53.Shambaugh GE., 3rd Urea biosynthesis I. The urea cycle and relationships to the citric acid cycle. Am J Clin Nutr. 1977;30:2083–2087. doi: 10.1093/ajcn/30.12.2083. [DOI] [PubMed] [Google Scholar]
- 54.Yudkoff M. Urea cycle. In: Siegel GJ, Agranoff BW, Albers RW, Fisher SK, Uhler MD, editors. Basic neurochemistry: molecular, cellular and medical aspects. 6th ed. Philadelphia: Lippincott-Raven; 1999. [Google Scholar]
- 55.Horecker BL. The pentose phosphate pathway. J Biol Chem. 2002;277:47965–47971. doi: 10.1074/jbc.X200007200. [DOI] [PubMed] [Google Scholar]
- 56.Lee MH, Malloy CR, Corbin IR, Li J, Jin ES. Assessing the pentose phosphate pathway using [2, 3-(13) C(2)] glucose. NMR Biomed. 2019;32:e4096. doi: 10.1002/nbm.4096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Stincone A, Prigione A, Cramer T, et al. The return of metabolism: biochemistry and physiology of the pentose phosphate pathway. Biol Rev Camb Philos Soc. 2015;90:927–963. doi: 10.1111/brv.12140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hall CN, Klein-Flügge MC, Howarth C, Attwell D. Oxidative phosphorylation, not glycolysis, powers presynaptic and postsynaptic mechanisms underlying brain information processing. J Neurosci. 2012;32:8940–8951. doi: 10.1523/JNEUROSCI.0026-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Nortley R, Attwell D. Control of brain energy supply by astrocytes. Curr Opin Neurobiol. 2017;47:80–85. doi: 10.1016/j.conb.2017.09.012. [DOI] [PubMed] [Google Scholar]
- 60.Yellen G. Fueling thought: Management of glycolysis and oxidative phosphorylation in neuronal metabolism. J Cell Biol. 2018;217:2235–2246. doi: 10.1083/jcb.201803152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bolaños JP. Bioenergetics and redox adaptations of astrocytes to neuronal activity. J Neurochem. 2016;139(Suppl 2):115–125. doi: 10.1111/jnc.13486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sonnewald U. Glutamate synthesis has to be matched by its degradation - where do all the carbons go? J Neurochem. 2014;131:399–406. doi: 10.1111/jnc.12812. [DOI] [PubMed] [Google Scholar]
- 63.Serres S, Raffard G, Franconi JM, Merle M. Close coupling between astrocytic and neuronal metabolisms to fulfill anaplerotic and energy needs in the rat brain. J Cereb Blood Flow Metab. 2008;28:712–724. doi: 10.1038/sj.jcbfm.9600568. [DOI] [PubMed] [Google Scholar]
- 64.Jády AG, Nagy Á M, Kőhidi T, Ferenczi S, Tretter L, Madarász E. Differentiation-dependent energy production and metabolite utilization: a comparative study on neural stem cells, neurons, and astrocytes. Stem Cells Dev. 2016;25:995–1005. doi: 10.1089/scd.2015.0388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Juaristi I, Contreras L, González-Sánchez P, et al. The response to stimulation in neurons and astrocytes. Neurochem Res. 2019;44:2385–2391. doi: 10.1007/s11064-019-02803-7. [DOI] [PubMed] [Google Scholar]
- 66.Juaristi I, Llorente-Folch I, Satrústegui J, Del Arco A. Extracellular ATP and glutamate drive pyruvate production and energy demand to regulate mitochondrial respiration in astrocytes. Glia. 2019;67:759–774. doi: 10.1002/glia.23574. [DOI] [PubMed] [Google Scholar]
- 67.Bendahan D, Chatel B, Jue T. Comparative NMR and NIRS analysis of oxygen-dependent metabolism in exercising finger flexor muscles. Am J Physiol Regul Integr Comp Physiol. 2017;313:R740–R753. doi: 10.1152/ajpregu.00203.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Rogatzki MJ, Ferguson BS, Goodwin ML, Gladden LB. Lactate is always the end product of glycolysis. Front Neurosci. 2015;9:22. doi: 10.3389/fnins.2015.00022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bastian C, Quinn J, Doherty C, et al. Role of brain glycogen during ischemia, aging and cell-to-cell interactions. Adv Neurobiol. 2019;23:347–361. doi: 10.1007/978-3-030-27480-1_12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Calì C, Tauffenberger A, Magistretti P. The strategic location of glycogen and lactate: from body energy reserve to brain plasticity. Front Cell Neurosci. 2019;13:82. doi: 10.3389/fncel.2019.00082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Mächler P, Wyss MT, Elsayed M, et al. In vivo evidence for a lactate gradient from astrocytes to neurons. Cell Metab. 2016;23:94–102. doi: 10.1016/j.cmet.2015.10.010. [DOI] [PubMed] [Google Scholar]
- 72.Mason S. Lactate shuttles in neuroenergetics-homeostasis, allostasis and beyond. Front Neurosci. 2017;11:43. doi: 10.3389/fnins.2017.00043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Mason S, van Furth AM, Mienie LJ, et al. A hypothetical astrocyte-microglia lactate shuttle derived from a (1)H NMR metabolomics analysis of cerebrospinal fluid from a cohort of South African children with tuberculous meningitis. Metabolomics. 2015;11:822–837. doi: 10.1007/s11306-014-0741-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Cruz RS, de Aguiar RA, Turnes T, Penteado Dos Santos R, de Oliveira MF, Caputo F. Intracellular shuttle: the lactate aerobic metabolism. Scientific World Journal. 2012;2012:420984. doi: 10.1100/2012/420984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Benveniste H, Heerdt PM, Fontes M, Rothman DL, Volkow ND. Glymphatic system function in relation to anesthesia and sleep states. Anesth Analg. 2019;128:747–758. doi: 10.1213/ANE.0000000000004069. [DOI] [PubMed] [Google Scholar]
- 76.Benveniste H, Lee H, Volkow ND. The glymphatic pathway: waste removal from the CNS via cerebrospinal fluid transport. Neuroscientist. 2017;23:454–465. doi: 10.1177/1073858417691030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Benveniste H, Liu X, Koundal S, Sanggaard S, Lee H, Wardlaw J. The glymphatic system and waste clearance with brain aging: a review. Gerontology. 2019;65:106–119. doi: 10.1159/000490349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Harrison IF, Siow B, Akilo AB, et al. Non-invasive imaging of CSF-mediated brain clearance pathways via assessment of perivascular fluid movement with diffusion tensor MRI. Elife. 2018;7:e34028. doi: 10.7554/eLife.34028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Jessen NA, Munk AS, Lundgaard I, Nedergaard M. The Glymphatic system: a beginner’s guide. Neurochem Res. 2015;40:2583–2599. doi: 10.1007/s11064-015-1581-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Plog BA, Nedergaard M. The glymphatic system in central nervous system health and disease: past, present, and future. Annu Rev Pathol. 2018;13:379–394. doi: 10.1146/annurev-pathol-051217-111018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Ratner V, Gao Y, Lee H, et al. Cerebrospinal and interstitial fluid transport via the glymphatic pathway modeled by optimal mass transport. Neuroimage. 2017;152:530–537. doi: 10.1016/j.neuroimage.2017.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wang LH, Wang ZL, Chen WY, Chen MJ, Xu GY. The glymphatic system: concept, function and research progresses. Sheng Li Xue Bao. 2018;70:52–60. [PubMed] [Google Scholar]
- 83.Lundgaard I, Lu ML, Yang E, et al. Glymphatic clearance controls state-dependent changes in brain lactate concentration. J Cereb Blood Flow Metab. 2017;37:2112–2124. doi: 10.1177/0271678X16661202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mintun MA, Vlassenko AG, Rundle MM, Raichle ME. Increased lactate/pyruvate ratio augments blood flow in physiologically activated human brain. Proc Natl Acad Sci U S A. 2004;101:659–664. doi: 10.1073/pnas.0307457100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Naylor E, Aillon DV, Barrett BS, et al. Lactate as a biomarker for sleep. Sleep. 2012;35:1209–1222. doi: 10.5665/sleep.2072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Deng W, Liu C, Parra C, et al. Quantitative imaging of the clearance systems in the eye and the brain. Quant Imaging Med Surg. 2020;10:1–14. doi: 10.21037/qims.2019.11.18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Verheggen ICM, Van Boxtel MPJ, Verhey FRJ, Jansen JFA, Backes WH. Interaction between blood-brain barrier and glymphatic system in solute clearance. Neurosci Biobehav Rev. 2018;90:26–33. doi: 10.1016/j.neubiorev.2018.03.028. [DOI] [PubMed] [Google Scholar]
- 88.Xie L, Kang H, Xu Q, et al. Sleep drives metabolite clearance from the adult brain. Science. 2013;342:373–377. doi: 10.1126/science.1241224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Louveau A, Smirnov I, Keyes TJ, et al. Structural and functional features of central nervous system lymphatic vessels. Nature. 2015;523:337–341. doi: 10.1038/nature14432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ringstad G, Valnes LM, Dale AM, et al. Brain-wide glymphatic enhancement and clearance in humans assessed with MRI. JCI Insight. 2018;3:e121537. doi: 10.1172/jci.insight.121537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Albargothy NJ, Johnston DA, MacGregor-Sharp M, et al. Convective influx/glymphatic system: tracers injected into the CSF enter and leave the brain along separate periarterial basement membrane pathways. Acta Neuropathol. 2018;136:139–152. doi: 10.1007/s00401-018-1862-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Aldea R, Weller RO, Wilcock DM, Carare RO, Richardson G. Cerebrovascular smooth muscle cells as the drivers of intramural periarterial drainage of the brain. Front Aging Neurosci. 2019;11:1. doi: 10.3389/fnagi.2019.00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Abbott NJ. Evidence for bulk flow of brain interstitial fluid: significance for physiology and pathology. Neurochem Int. 2004;45:545–552. doi: 10.1016/j.neuint.2003.11.006. [DOI] [PubMed] [Google Scholar]
- 94.Rapoport SI. Blood-brain barrier in physiology and medicine. New York, NY, USA: Raven Press; 1976. [Google Scholar]
- 95.Semyachkina-Glushkovskaya O, Postnov D, Kurths J. Blood, brain barrier, lymphatic clearance, and recovery: Ariadne’s thread in labyrinths of hypotheses. Int J Mol Sci. 2018;19:3818. doi: 10.3390/ijms19123818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Aspelund A, Antila S, Proulx ST, et al. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J Exp Med. 2015;212:991–999. doi: 10.1084/jem.20142290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Louveau A, Plog BA, Antila S, Alitalo K, Nedergaard M, Kipnis J. Understanding the functions and relationships of the glymphatic system and meningeal lymphatics. J Clin Invest. 2017;127:3210–3219. doi: 10.1172/JCI90603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Tarasoff-Conway JM, Carare RO, Osorio RS, et al. Clearance systems in the brain-implications for Alzheimer disease. Nat Rev Neurol. 2015;11:457–470. doi: 10.1038/nrneurol.2015.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Brooks GA. Lactate as a fulcrum of metabolism. Redox Biol. 2020;35:101454. doi: 10.1016/j.redox.2020.101454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ferguson BS, Rogatzki MJ, Goodwin ML, Kane DA, Rightmire Z, Gladden LB. Lactate metabolism: historical context, prior misinterpretations, and current understanding. Eur J Appl Physiol. 2018;118:691–728. doi: 10.1007/s00421-017-3795-6. [DOI] [PubMed] [Google Scholar]
- 101.Schurr A, Payne RS. Lactate, not pyruvate, is neuronal aerobic glycolysis end product: an in vitro electrophysiological study. Neuroscience. 2007;147:613–619. doi: 10.1016/j.neuroscience.2007.05.002. [DOI] [PubMed] [Google Scholar]
- 102.Schurr A. Lactate: a major and crucial player in normal function of both muscle and brain. J Physiol. 2008;586:2665–2666. doi: 10.1113/jphysiol.2008.155416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Khacho M, Tarabay M, Patten D, et al. Acidosis overrides oxygen deprivation to maintain mitochondrial function and cell survival. Nat Commun. 2014;5:3550. doi: 10.1038/ncomms4550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Lamonte G, Tang X, Chen JL, et al. Acidosis induces reprogramming of cellular metabolism to mitigate oxidative stress. Cancer Metab. 2013;1:23. doi: 10.1186/2049-3002-1-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Wilson DF. Regulation of cellular metabolism: programming and maintaining metabolic homeostasis. J Appl Physiol (1985) 2013;115:1583–1588. doi: 10.1152/japplphysiol.00894.2013. [DOI] [PubMed] [Google Scholar]
- 106.Baufeld A, Koczan D, Vanselow J. L-lactate induces specific genome wide alterations of gene expression in cultured bovine granulosa cells. BMC Genomics. 2019;20:273. doi: 10.1186/s12864-019-5657-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Gabriel-Costa D, da Cunha TF, Bechara LR, et al. Lactate up-regulates the expression of lactate oxidation complex-related genes in left ventricular cardiac tissue of rats. PLoS One. 2015;10:e0127843. doi: 10.1371/journal.pone.0127843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Haas R, Smith J, Rocher-Ros V, et al. Lactate regulates metabolic and pro-inflammatory circuits in control of T cell migration and effector functions. PLoS Biol. 2015;13:e1002202. doi: 10.1371/journal.pbio.1002202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Hollyer TR, Bordoni L, Kousholt BS, van Luijk J, Ritskes-Hoitinga M, Østergaard L. The evidence for the physiological effects of lactate on the cerebral microcirculation: a systematic review. J Neurochem. 2019;148:712–730. doi: 10.1111/jnc.14633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Tauffenberger A, Fiumelli H, Almustafa S, Magistretti PJ. Lactate and pyruvate promote oxidative stress resistance through hormetic ROS signaling. Cell Death Dis. 2019;10:653. doi: 10.1038/s41419-019-1877-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Veech RL, Todd King M, Pawlosky R, Kashiwaya Y, Bradshaw PC, Curtis W. The “great” controlling nucleotide coenzymes. IUBMB Life. 2019;71:565–579. doi: 10.1002/iub.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Schurr A. Lactate: the ultimate cerebral oxidative energy substrate? J Cereb Blood Flow Metab. 2006;26:142–152. doi: 10.1038/sj.jcbfm.9600174. [DOI] [PubMed] [Google Scholar]
- 113.Glenn TC, Martin NA, Horning MA, et al. Lactate: brain fuel in human traumatic brain injury: a comparison with normal healthy control subjects. J Neurotrauma. 2015;32:820–832. doi: 10.1089/neu.2014.3483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hashimoto T, Hussien R, Cho HS, Kaufer D, Brooks GA. Evidence for the mitochondrial lactate oxidation complex in rat neurons: demonstration of an essential component of brain lactate shuttles. PLoS One. 2008;3:e2915. doi: 10.1371/journal.pone.0002915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Jalloh I, Helmy A, Howe DJ, et al. A Comparison of Oxidative Lactate Metabolism in Traumatically Injured Brain and Control Brain. J Neurotrauma. 2018;35:2025–2035. doi: 10.1089/neu.2017.5459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Zhou J, Liu T, Guo H, et al. Lactate potentiates angiogenesis and neurogenesis in experimental intracerebral hemorrhage. Exp Mol Med. 2018;50:1–12. doi: 10.1038/s12276-018-0113-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Dienel GA. Brain lactate metabolism: the discoveries and the controversies. J Cereb Blood Flow Metab. 2012;32:1107–1138. doi: 10.1038/jcbfm.2011.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ernst AS, Böhler LI, Hagenston AM, et al. EphB2-dependent signaling promotes neuronal excitotoxicity and inflammation in the acute phase of ischemic stroke. Acta Neuropathol Commun. 2019;7:15. doi: 10.1186/s40478-019-0669-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Fern R, Matute C. Glutamate receptors and white matter stroke. Neurosci Lett. 2019;694:86–92. doi: 10.1016/j.neulet.2018.11.031. [DOI] [PubMed] [Google Scholar]
- 120.Kaur R, Mehan S, Singh S. Understanding multifactorial architecture of Parkinson’s disease: pathophysiology to management. Neurol Sci. 2019;40:13–23. doi: 10.1007/s10072-018-3585-x. [DOI] [PubMed] [Google Scholar]
- 121.Valko K, Ciesla L. Amyotrophic lateral sclerosis. Prog Med Chem. 2019;58:63–117. doi: 10.1016/bs.pmch.2018.12.001. [DOI] [PubMed] [Google Scholar]
- 122.Woitzik J, Pinczolits A, Hecht N, et al. Excitotoxicity and metabolic changes in association with infarct progression. Stroke. 2014;45:1183–1185. doi: 10.1161/STROKEAHA.113.004475. [DOI] [PubMed] [Google Scholar]
- 123.Bouzat P, Oddo M. Lactate and the injured brain: friend or foe? Curr Opin Crit Care. 2014;20:133–140. doi: 10.1097/MCC.0000000000000072. [DOI] [PubMed] [Google Scholar]
- 124.Castillo X, Rosafio K, Wyss MT, et al. A probable dual mode of action for both L- and D-lactate neuroprotection in cerebral ischemia. J Cereb Blood Flow Metab. 2015;35:1561–1569. doi: 10.1038/jcbfm.2015.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Jourdain P, Allaman I, Rothenfusser K, Fiumelli H, Marquet P, Magistretti PJ. L-lactate protects neurons against excitotoxicity: implication of an ATP-mediated signaling cascade. Sci Rep. 2016;6:21250. doi: 10.1038/srep21250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Margineanu MB, Mahmood H, Fiumelli H, Magistretti PJ. L-lactate regulates the expression of synaptic plasticity and neuroprotection genes in cortical neurons: a transcriptome analysis. Front Mol Neurosci. 2018;11:375. doi: 10.3389/fnmol.2018.00375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Sobieski C, Warikoo N, Shu HJ, Mennerick S. Ambient but not local lactate underlies neuronal tolerance to prolonged glucose deprivation. PLoS One. 2018;13:e0195520. doi: 10.1371/journal.pone.0195520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.DiNuzzo M, Giove F, Maraviglia B, Mangia S. Computational flux balance analysis predicts that stimulation of energy metabolism in astrocytes and their metabolic interactions with neurons depend on uptake of K(+) rather than glutamate. Neurochem Res. 2017;42:202–216. doi: 10.1007/s11064-016-2048-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Sonnay S, Gruetter R, Duarte JMN. How energy metabolism supports cerebral function: insights from (13)C magnetic resonance studies in vivo. Front Neurosci. 2017;11:288. doi: 10.3389/fnins.2017.00288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Mishra A, Reynolds JP, Chen Y, Gourine AV, Rusakov DA, Attwell D. Astrocytes mediate neurovascular signaling to capillary pericytes but not to arterioles. Nat Neurosci. 2016;19:1619–1627. doi: 10.1038/nn.4428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Lamon BD, Zhang FF, Puri N, Brodsky SV, Goligorsky MS, Nasjletti A. Dual pathways of carbon monoxide-mediated vasoregulation: modulation by redox mechanisms. Circ Res. 2009;105:775–783. doi: 10.1161/CIRCRESAHA.109.197434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Leffler CW, Parfenova H, Jaggar JH. Carbon monoxide as an endogenous vascular modulator. Am J Physiol Heart Circ Physiol. 2011;301:H1–H11. doi: 10.1152/ajpheart.00230.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Xi Q, Umstot E, Zhao G, Narayanan D, Leffler CW, Jaggar JH. Glutamate regulates Ca2+ signals in smooth muscle cells of newborn piglet brain slice arterioles through astrocyte- and heme oxygenase-dependent mechanisms. Am J Physiol Heart Circ Physiol. 2010;298:H562–569. doi: 10.1152/ajpheart.00823.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Pournaras CJ, Rungger-Brändle E, Riva CE, Hardarson SH, Stefansson E. Regulation of retinal blood flow in health and disease. Prog Retin Eye Res. 2008;27:284–330. doi: 10.1016/j.preteyeres.2008.02.002. [DOI] [PubMed] [Google Scholar]
- 135.Vafaee MS, Vang K, Bergersen LH, Gjedde A. Oxygen consumption and blood flow coupling in human motor cortex during intense finger tapping: implication for a role of lactate. J Cereb Blood Flow Metab. 2012;32:1859–1868. doi: 10.1038/jcbfm.2012.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Obel LF, Müller MS, Walls AB, et al. Brain glycogen-new perspectives on its metabolic function and regulation at the subcellular level. Front Neuroenergetics. 2012;4:3. doi: 10.3389/fnene.2012.00003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Pederson BA. Structure and regulation of glycogen synthase in the brain. Adv Neurobiol. 2019;23:83–123. doi: 10.1007/978-3-030-27480-1_3. [DOI] [PubMed] [Google Scholar]
- 138.Waitt AE, Reed L, Ransom BR, Brown AM. Emerging roles for glycogen in the CNS. Front Mol Neurosci. 2017;10:73. doi: 10.3389/fnmol.2017.00073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Falkowska A, Gutowska I, Goschorska M, Nowacki P, Chlubek D, Baranowska-Bosiacka I. Energy metabolism of the brain, including the cooperation between astrocytes and neurons, especially in the context of glycogen metabolism. Int J Mol Sci. 2015;16:25959–25981. doi: 10.3390/ijms161125939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Rich LR, Harris W, Brown AM. The role of brain glycogen in supporting physiological function. Front Neurosci. 2019;13:1176. doi: 10.3389/fnins.2019.01176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Chander BS, Chakravarthy VS. A computational model of neuro-glio-vascular loop interactions. PLoS One. 2012;7:e48802. doi: 10.1371/journal.pone.0048802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Jolivet R, Coggan JS, Allaman I, Magistretti PJ. Multi-timescale modeling of activity-dependent metabolic coupling in the neuron-glia-vasculature ensemble. PLoS Comput Biol. 2015;11:e1004036. doi: 10.1371/journal.pcbi.1004036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Chu XP, Zhu XM, Wei WL, et al. Acidosis decreases low Ca(2+)-induced neuronal excitation by inhibiting the activity of calcium-sensing cation channels in cultured mouse hippocampal neurons. J Physiol. 2003;550:385–399. doi: 10.1113/jphysiol.2003.043091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Deitmer JW, Rose CR. pH regulation and proton signalling by glial cells. Prog Neurobiol. 1996;48:73–103. doi: 10.1016/0301-0082(95)00039-9. [DOI] [PubMed] [Google Scholar]
- 145.Obara M, Szeliga M, Albrecht J. Regulation of pH in the mammalian central nervous system under normal and pathological conditions: facts and hypotheses. Neurochem Int. 2008;52:905–919. doi: 10.1016/j.neuint.2007.10.015. [DOI] [PubMed] [Google Scholar]
- 146.Schlue WR, Dörner R. The regulation of pH in the central nervous system. Can J Physiol Pharmacol. 1992;70 Suppl:S278–285. doi: 10.1139/y92-273. [DOI] [PubMed] [Google Scholar]
- 147.Karolinska Institutet. New mechanism for sensing and regulating pH in the nervous system. [Accessed by February 2, 2021]. https://medicalxpress.com/news/2016-04-mechanism-ph-nervous.html .
- 148.Badaut J, Fukuda AM, Jullienne A, Petry KG. Aquaporin and brain diseases. Biochim Biophys Acta. 2014;1840:1554–1565. doi: 10.1016/j.bbagen.2013.10.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Filippidis AS, Carozza RB, Rekate HL. Aquaporins in brain edema and neuropathological conditions. Int J Mol Sci. 2016;18:55. doi: 10.3390/ijms18010055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Lambert J, Mejia S, Vojdani A. Plant and human aquaporins: pathogenesis from gut to brain. Immunol Res. 2019;67:12–20. doi: 10.1007/s12026-018-9046-z. [DOI] [PubMed] [Google Scholar]
- 151.Boron WF. Sharpey-Schafer lecture: gas channels. Exp Physiol. 2010;95:1107–1130. doi: 10.1113/expphysiol.2010.055244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Herrera M, Garvin JL. Aquaporins as gas channels. Pflugers Arch. 2011;462:623–630. doi: 10.1007/s00424-011-1002-x. [DOI] [PubMed] [Google Scholar]
- 153.Mohammadi MT, Dehghani GA. Nitric oxide as a regulatory factor for aquaporin-1 and 4 gene expression following brain ischemia/reperfusion injury in rat. Pathol Res Pract. 2015;211:43–49. doi: 10.1016/j.prp.2014.07.014. [DOI] [PubMed] [Google Scholar]
- 154.Rodriguez-Grande B, Konsman JP. Gas diffusion in the CNS. J Neurosci Res. 2018;96:207–218. doi: 10.1002/jnr.24077. [DOI] [PubMed] [Google Scholar]
- 155.Mohan A, Pendyam S, Kalivas PW, Nair SS. Molecular diffusion model of neurotransmitter homeostasis around synapses supporting gradients. Neural Comput. 2011;23:984–1014. doi: 10.1162/NECO_a_00101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Murphy-Royal C, Dupuis J, Groc L, Oliet SHR. Oliet SHR. Astroglial glutamate transporters in the brain: Regulating neurotransmitter homeostasis and synaptic transmission. J Neurosci Res. 2017;95:2140–2151. doi: 10.1002/jnr.24029. [DOI] [PubMed] [Google Scholar]
- 157.Pendyam S, Mohan A, Kalivas PW, Nair SS. Role of perisynaptic parameters in neurotransmitter homeostasis--computational study of a general synapse. Synapse. 2012;66:608–621. doi: 10.1002/syn.21547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Boehning D, Moon C, Sharma S, et al. Carbon monoxide neurotransmission activated by CK2 phosphorylation of heme oxygenase-2. Neuron. 2003;40:129–137. doi: 10.1016/s0896-6273(03)00596-8. [DOI] [PubMed] [Google Scholar]
- 159.Xue L, Farrugia G, Miller SM, Ferris CD, Snyder SH, Szurszewski JH. Carbon monoxide and nitric oxide as coneurotransmitters in the enteric nervous system: evidence from genomic deletion of biosynthetic enzymes. Proc Natl Acad Sci U S A. 2000;97:1851–1855. doi: 10.1073/pnas.97.4.1851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.University of Bristol. New research sheds light on how the body regulates fundamental neuro-hormone. [Accessed by February 2, 2021]. https://www.eurekalert.org/pub_releases/2014-02/uob-nrs021014.php .
- 161.Jung E, Koh SH, Yoo M, Choi YK. Regenerative potential of carbon monoxide in adult neural circuits of the central nervous system. Int J Mol Sci. 2020;21:2273. doi: 10.3390/ijms21072273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Mahar M, Cavalli V. Intrinsic mechanisms of neuronal axon regeneration. Nat Rev Neurosci. 2018;19:323–337. doi: 10.1038/s41583-018-0001-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Telezhkin V, Brazier SP, Mears R, Müller CT, Riccardi D, Kemp PJ. Cysteine residue 911 in C-terminal tail of human BK(Ca)α channel subunit is crucial for its activation by carbon monoxide. Pflugers Arch. 2011;461:665–675. doi: 10.1007/s00424-011-0924-7. [DOI] [PubMed] [Google Scholar]
- 164.Bak LK, Obel LF, Walls AB, et al. Novel model of neuronal bioenergetics: postsynaptic utilization of glucose but not lactate correlates positively with Ca2+ signalling in cultured mouse glutamatergic neurons. ASN Neuro. 2012;4:e00083. doi: 10.1042/AN20120004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Takata T, Sakurai T, Yang B, Yokono K, Okada Y. Effect of lactate on the synaptic potential, energy metabolism, calcium homeostasis and extracellular glutamate concentration in the dentate gyrus of the hippocampus from guinea-pig. Neuroscience. 2001;104:371–378. doi: 10.1016/s0306-4522(01)00086-0. [DOI] [PubMed] [Google Scholar]
- 166.Benarroch EE. Cation-chloride cotransporters in the nervous system: general features and clinical correlations. Neurology. 2013;80:756–763. doi: 10.1212/WNL.0b013e318283bb1c. [DOI] [PubMed] [Google Scholar]
- 167.Li K, Xu E. The role and the mechanism of gamma-aminobutyric acid during central nervous system development. Neurosci Bull. 2008;24:195–200. doi: 10.1007/s12264-008-0109-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Doyon N, Vinay L, Prescott SA, De Koninck Y. Chloride regulation: a dynamic equilibrium crucial for synaptic inhibition. Neuron. 2016;89:1157–1172. doi: 10.1016/j.neuron.2016.02.030. [DOI] [PubMed] [Google Scholar]
- 169.Watanabe M, Fukuda A. Development and regulation of chloride homeostasis in the central nervous system. Front Cell Neurosci. 2015;9:371. doi: 10.3389/fncel.2015.00371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Blaesse P, Schmidt T. K-Cl cotransporter KCC2--a moonlighting protein in excitatory and inhibitory synapse development and function. Pflugers Arch. 2015;467:615–624. doi: 10.1007/s00424-014-1547-6. [DOI] [PubMed] [Google Scholar]
- 171.Chew TA, Orlando BJ, Zhang J, et al. Structure and mechanism of the cation-chloride cotransporter NKCC1. Nature. 2019;572:488–492. doi: 10.1038/s41586-019-1438-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Coxon JP, Cash RFH, Hendrikse JJ, et al. GABA concentration in sensorimotor cortex following high-intensity exercise and relationship to lactate levels. J Physiol. 2018;596:691–702. doi: 10.1113/JP274660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Reis WL, Biancardi VC, Son S, Antunes-Rodrigues J, Stern JE. Carbon monoxide and nitric oxide interactions in magnocellular neurosecretory neurones during water deprivation. J Neuroendocrinol. 2015;27:111–122. doi: 10.1111/jne.12245. [DOI] [PubMed] [Google Scholar]
- 174.Kadala A, Verdier D, Morquette P, Kolta A. Ion homeostasis in rhythmogenesis: the interplay between neurons and astroglia. Physiology (Bethesda) 2015;30:371–388. doi: 10.1152/physiol.00023.2014. [DOI] [PubMed] [Google Scholar]
- 175.Larsen BR, Holm R, Vilsen B, MacAulay N. Glutamate transporter activity promotes enhanced Na(+) /K(+) -ATPase-mediated extracellular K(+) management during neuronal activity. J Physiol. 2016;594:6627–6641. doi: 10.1113/JP272531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Larsen BR, Stoica A, MacAulay N. Managing brain extracellular K(+) during neuronal activity: the physiological role of the Na(+)/K(+)-ATPase subunit isoforms. Front Physiol. 2016;7:141. doi: 10.3389/fphys.2016.00141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Morquette P, Verdier D, Kadala A, et al. An astrocyte-dependent mechanism for neuronal rhythmogenesis. Nat Neurosci. 2015;18:844–854. doi: 10.1038/nn.4013. [DOI] [PubMed] [Google Scholar]
- 178.Scemes E, Spray DC. Extracellular K+ and astrocyte signaling via connexin and pannexin channels. Neurochem Res. 2012;37:2310–2316. doi: 10.1007/s11064-012-0759-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Xu L, Zeng LH, Wong M. Impaired astrocytic gap junction coupling and potassium buffering in a mouse model of tuberous sclerosis complex. Neurobiol Dis. 2009;34:291–299. doi: 10.1016/j.nbd.2009.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Hertz L, Gerkau NJ, Xu J, et al. Roles of astrocytic Na(+), K(+)-ATPase and glycogenolysis for K(+) homeostasis in mammalian brain. J Neurosci Res. 2015;93:1019–1030. doi: 10.1002/jnr.23499. [DOI] [PubMed] [Google Scholar]
- 181.Xu J, Song D, Xue Z, Gu L, Hertz L, Peng L. Requirement of glycogenolysis for uptake of increased extracellular K+ in astrocytes: potential implications for K+ homeostasis and glycogen usage in brain. Neurochem Res. 2013;38:472–485. doi: 10.1007/s11064-012-0938-3. [DOI] [PubMed] [Google Scholar]
- 182.Sepúlveda FV, Pablo Cid L, Teulon J, Niemeyer MI. Molecular aspects of structure, gating, and physiology of pH-sensitive background K2P and Kir K+-transport channels. Physiol Rev. 2015;95:179–217. doi: 10.1152/physrev.00016.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Kinboshi M, Mukai T, Nagao Y, et al. inhibition of inwardly rectifying potassium (Kir) 4.1 channels facilitates brain-derived neurotrophic factor (BDNF) expression in astrocytes. Front Mol Neurosci. 2017;10:408. doi: 10.3389/fnmol.2017.00408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Ohno Y, Kinboshi M, Shimizu S. Inwardly rectifying potassium channel Kir4.1 as a novel modulator of BDNF expression in astrocytes. Int J Mol Sci. 2018;19:3313. doi: 10.3390/ijms19113313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Ha J, Xu Y, Kawano T, et al. Hydrogen sulfide inhibits Kir2 and Kir3 channels by decreasing sensitivity to the phospholipid phosphatidylinositol 4,5-bisphosphate (PIP(2)) J Biol Chem. 2018;293:3546–3561. doi: 10.1074/jbc.RA117.001679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Liang S, Wang Q, Zhang W, et al. Carbon monoxide inhibits inward rectifier potassium channels in cardiomyocytes. Nat Commun. 2014;5:4676. doi: 10.1038/ncomms5676. [DOI] [PubMed] [Google Scholar]
- 187.Dupont G, Schmidt C, Yilmaz E, et al. Our current understanding of the lymphatics of the brain and spinal cord. Clin Anat. 2019;32:117–121. doi: 10.1002/ca.23308. [DOI] [PubMed] [Google Scholar]
- 188.Raper D, Louveau A, Kipnis J. How Do Meningeal Lymphatic Vessels Drain the CNS? Trends Neurosci. 2016;39:581–586. doi: 10.1016/j.tins.2016.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Thomas JL, Jacob L, Boisserand L. Lymphatic system in central nervous system. Med Sci (Paris) 2019;35:55–61. doi: 10.1051/medsci/2018309. [DOI] [PubMed] [Google Scholar]
- 190.Chen J, Wang L, Xu H, et al. Meningeal lymphatics clear erythrocytes that arise from subarachnoid hemorrhage. Nat Commun. 2020;11:3159. doi: 10.1038/s41467-020-16851-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Jiang X, Andjelkovic AV, Zhu L, et al. Blood-brain barrier dysfunction and recovery after ischemic stroke. Prog Neurobiol. 2018;163-164:144–171. doi: 10.1016/j.pneurobio.2017.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Sifat AE, Vaidya B, Abbruscato TJ. Blood-brain barrier protection as a therapeutic strategy for acute ischemic stroke. AAPS J. 2017;19:957–972. doi: 10.1208/s12248-017-0091-7. [DOI] [PubMed] [Google Scholar]
- 193.Wang J, Zhang D, Fu X, et al. Carbon monoxide-releasing molecule-3 protects against ischemic stroke by suppressing neuroinflammation and alleviating blood-brain barrier disruption. J Neuroinflammation. 2018;15:188. doi: 10.1186/s12974-018-1226-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Yang Y, Dong B, Lu J, Wang G, Yu Y. Hemopexin reduces blood-brain barrier injury and protects synaptic plasticity in cerebral ischemic rats by promoting EPCs through the HO-1 pathway. Brain Res. 2018;1699:177–185. doi: 10.1016/j.brainres.2018.08.008. [DOI] [PubMed] [Google Scholar]
- 195.Choi YK. Role of carbon monoxide in neurovascular repair processing. Biomol Ther (Seoul) 2018;26:93–100. doi: 10.4062/biomolther.2017.144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Shefa U, Kim D, Kim MS, Jeong NY, Jung J. Roles of gasotransmitters in synaptic plasticity and neuropsychiatric conditions. Neural Plast 2018. 2018 doi: 10.1155/2018/1824713. 1824713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Shefa U, Yeo SG, Kim MS, et al. Role of gasotransmitters in oxidative stresses, neuroinflammation, and neuronal repair. Biomed Res Int 2017. 2017 doi: 10.1155/2017/1689341. 1689341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Wang Q, Hu Y, Wan J, Dong B, Sun J. Lactate: A novel signaling molecule in synaptic plasticity and drug addiction. Bioessays. 2019;41:e1900008. doi: 10.1002/bies.201900008. [DOI] [PubMed] [Google Scholar]
- 199.Yang J, Ruchti E, Petit JM, et al. Lactate promotes plasticity gene expression by potentiating NMDA signaling in neurons. Proc Natl Acad Sci U S A. 2014;111:12228–12233. doi: 10.1073/pnas.1322912111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Subramanian J, Morozov A. Erk1/2 inhibit synaptic vesicle exocytosis through L-type calcium channels. J Neurosci. 2011;31:4755–4764. doi: 10.1523/JNEUROSCI.6594-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Kaizu T, Ikeda A, Nakao A, et al. Protection of transplant-induced hepatic ischemia/reperfusion injury with carbon monoxide via MEK/ERK1/2 pathway downregulation. Am J Physiol Gastrointest Liver Physiol. 2008;294:G236–244. doi: 10.1152/ajpgi.00144.2007. [DOI] [PubMed] [Google Scholar]
- 202.Park SJ, Lee SK, Lim CR, et al. Heme oxygenase-1/carbon monoxide axis suppresses transforming growth factor-β1-induced growth inhibition by increasing ERK1/2-mediated phosphorylation of Smad3 at Thr-179 in human hepatocellular carcinoma cell lines. Biochem Biophys Res Commun. 2018;498:609–615. doi: 10.1016/j.bbrc.2018.03.030. [DOI] [PubMed] [Google Scholar]
- 203.Agostini M, Romeo F, Inoue S, et al. Metabolic reprogramming during neuronal differentiation. Cell Death Differ. 2016;23:1502–1514. doi: 10.1038/cdd.2016.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Beckervordersandforth R, Ebert B, Schäffner I, et al. Role of mitochondrial metabolism in the control of early lineage progression and aging phenotypes in adult hippocampal neurogenesis. Neuron. 2017;93:560–573.e6. doi: 10.1016/j.neuron.2016.12.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Maffezzini C, Calvo-Garrido J, Wredenberg A, Freyer C. Metabolic regulation of neurodifferentiation in the adult brain. Cell Mol Life Sci. 2020;77:2483–2496. doi: 10.1007/s00018-019-03430-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Zheng X, Boyer L, Jin M, et al. Metabolic reprogramming during neuronal differentiation from aerobic glycolysis to neuronal oxidative phosphorylation. Elife. 2016;5:e13374. doi: 10.7554/eLife.13374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Almeida AS, Soares NL, Sequeira CO, Pereira SA, Sonnewald U, Vieira HLA. Improvement of neuronal differentiation by carbon monoxide: Role of pentose phosphate pathway. Redox Biol. 2018;17:338–347. doi: 10.1016/j.redox.2018.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Almeida AS, Soares NL, Vieira M, Gramsbergen JB, Vieira HL. Carbon monoxide releasing molecule-A1 (CORM-A1) improves neurogenesis: increase of neuronal differentiation yield by preventing cell death. PLoS One. 2016;11:e0154781. doi: 10.1371/journal.pone.0154781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Jha MK, Lee IK, Suk K. Metabolic reprogramming by the pyruvate dehydrogenase kinase-lactic acid axis: Linking metabolism and diverse neuropathophysiologies. Neurosci Biobehav Rev. 2016;68:1–19. doi: 10.1016/j.neubiorev.2016.05.006. [DOI] [PubMed] [Google Scholar]
- 210.Lev-Vachnish Y, Cadury S, Rotter-Maskowitz A, et al. L-lactate promotes adult hippocampal neurogenesis. Front Neurosci. 2019;13:403. doi: 10.3389/fnins.2019.00403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Wang J, Cui Y, Yu Z, et al. Brain endothelial cells maintain lactate homeostasis and control adult hippocampal neurogenesis. Cell Stem Cell. 2019;25:754–767.e9. doi: 10.1016/j.stem.2019.09.009. [DOI] [PubMed] [Google Scholar]
- 212.Choi YK, Maki T, Mandeville ET, et al. Dual effects of carbon monoxide on pericytes and neurogenesis in traumatic brain injury. Nat Med. 2016;22:1335–1341. doi: 10.1038/nm.4188. [DOI] [PubMed] [Google Scholar]
- 213.Chechik G, Meilijson I, Ruppin E. Neuronal regulation: A mechanism for synaptic pruning during brain maturation. Neural Comput. 1999;11:2061–2080. doi: 10.1162/089976699300016089. [DOI] [PubMed] [Google Scholar]
- 214.Lemke G. Glial control of neuronal development. Annu Rev Neurosci. 2001;24:87–105. doi: 10.1146/annurev.neuro.24.1.87. [DOI] [PubMed] [Google Scholar]
- 215.Lieberman OJ, McGuirt AF, Tang G, Sulzer D. Roles for neuronal and glial autophagy in synaptic pruning during development. Neurobiol Dis. 2019;122:49–63. doi: 10.1016/j.nbd.2018.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Fort PE, Xu NJ, Zhang W. Editorial: Neuronal development and degeneration. Front Genet. 2019;10:1213. doi: 10.3389/fgene.2019.01213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Kondo Y, Yada Y, Haga T, et al. Temporal relation between neural activity and neurite pruning on a numerical model and a microchannel device with micro electrode array. Biochem Biophys Res Commun. 2017;486:539–544. doi: 10.1016/j.bbrc.2017.03.082. [DOI] [PubMed] [Google Scholar]
- 218.Riccomagno MM, Kolodkin AL. Sculpting neural circuits by axon and dendrite pruning. Annu Rev Cell Dev Biol. 2015;31:779–805. doi: 10.1146/annurev-cellbio-100913-013038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Schuldiner O, Yaron A. Mechanisms of developmental neurite pruning. Cell Mol Life Sci. 2015;72:101–119. doi: 10.1007/s00018-014-1729-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Ulbrich F, Goebel U, Böhringer D, Charalambous P, Lagrèze WA, Biermann J. Carbon monoxide treatment reduces microglial activation in the ischemic rat retina. Graefes Arch Clin Exp Ophthalmol. 2016;254:1967–1976. doi: 10.1007/s00417-016-3435-6. [DOI] [PubMed] [Google Scholar]
- 221.Wilson JL, Bouillaud F, Almeida AS, et al. Carbon monoxide reverses the metabolic adaptation of microglia cells to an inflammatory stimulus. Free Radic Biol Med. 2017;104:311–323. doi: 10.1016/j.freeradbiomed.2017.01.022. [DOI] [PubMed] [Google Scholar]
- 222.Bani-Hani MG, Greenstein D, Mann BE, Green CJ, Motterlini R. A carbon monoxide-releasing molecule (CORM-3) attenuates lipopolysaccharide- and interferon-gamma-induced inflammation in microglia. Pharmacol Rep. 2006;58 Suppl:132–144. [PubMed] [Google Scholar]
- 223.Lauro C, Limatola C. Metabolic reprograming of microglia in the regulation of the innate inflammatory response. Front Immunol. 2020;11:493. doi: 10.3389/fimmu.2020.00493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Ding SL, Royall JJ, Sunkin SM, et al. Comprehensive cellular-resolution atlas of the adult human brain. J Comp Neurol. 2016;524:3127–3481. doi: 10.1002/cne.24080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Hedman AM, van Haren NE, Schnack HG, Kahn RS, Hulshoff Pol HE. Human brain changes across the life span: a review of 56 longitudinal magnetic resonance imaging studies. Hum Brain Mapp. 2012;33:1987–2002. doi: 10.1002/hbm.21334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Cheng L, Zhu Y, Sun J, et al. Principal states of dynamic functional connectivity reveal the link between resting-state and task-state brain: an fMRI study. Int J Neural Syst. 2018;28:1850002. doi: 10.1142/S0129065718500028. [DOI] [PubMed] [Google Scholar]
- 227.Garcés P, Pereda E, Hernández-Tamames JA, Del-Pozo F, Maestú F, Pineda-Pardo J. Multimodal description of whole brain connectivity: A comparison of resting state MEG, fMRI, and DWI. Hum Brain Mapp. 2016;37:20–34. doi: 10.1002/hbm.22995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Zimmermann J, Ritter P, Shen K, Rothmeier S, Schirner M, McIntosh AR. Structural architecture supports functional organization in the human aging brain at a regionwise and network level. Hum Brain Mapp. 2016;37:2645–2661. doi: 10.1002/hbm.23200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Datta S, Chakrabarti N. Age related rise in lactate and its correlation with lactate dehydrogenase (LDH) status in post-mitochondrial fractions isolated from different regions of brain in mice. Neurochem Int. 2018;118:23–33. doi: 10.1016/j.neuint.2018.04.007. [DOI] [PubMed] [Google Scholar]
- 230.Ross JM, Öberg J, Brené S, et al. High brain lactate is a hallmark of aging and caused by a shift in the lactate dehydrogenase A/B ratio. Proc Natl Acad Sci U S A. 2010;107:20087–20092. doi: 10.1073/pnas.1008189107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Poon HF, Calabrese V, Scapagnini G, Butterfield DA. Free radicals: key to brain aging and heme oxygenase as a cellular response to oxidative stress. J Gerontol A Biol Sci Med Sci. 2004;59:478–493. doi: 10.1093/gerona/59.5.m478. [DOI] [PubMed] [Google Scholar]
- 232.Schipper HM, Song W, Tavitian A, Cressatti M. The sinister face of heme oxygenase-1 in brain aging and disease. Prog Neurobiol. 2019;172:40–70. doi: 10.1016/j.pneurobio.2018.06.008. [DOI] [PubMed] [Google Scholar]
- 233.Cocks JA. Change in the concentration of lactic acid in the rat and hamster brain during natural sleep. Nature. 1967;215:1399–1400. doi: 10.1038/2151399a0. [DOI] [PubMed] [Google Scholar]
- 234.Reich P, Geyer SJ, Karnovsky ML. Metabolism of brain during sleep and wakefulness. J Neurochem. 1972;19:487–497. doi: 10.1111/j.1471-4159.1972.tb01358.x. [DOI] [PubMed] [Google Scholar]
- 235.Azuma M, Murase K, Tachikawa R, et al. Relationship between obstructive sleep apnea and endogenous carbon monoxide. J Appl Physiol (1985) 2017;122:104–111. doi: 10.1152/japplphysiol.00658.2016. [DOI] [PubMed] [Google Scholar]
- 236.Kobayashi M, Miyazawa N, Takeno M, et al. Circulating carbon monoxide level is elevated after sleep in patients with obstructive sleep apnea. Chest. 2008;134:904–910. doi: 10.1378/chest.07-2904. [DOI] [PubMed] [Google Scholar]
- 237.Jensen MLF, Vestergaard MB, Tønnesen P, Larsson HBW, Jennum PJ. Cerebral blood flow, oxygen metabolism, and lactate during hypoxia in patients with obstructive sleep apnea. Sleep. 2018;41:zsy001. doi: 10.1093/sleep/zsy001. [DOI] [PubMed] [Google Scholar]
- 238.Lin T, Huang JF, Lin QC, et al. The effect of CPAP treatment on venous lactate and arterial blood gas among obstructive sleep apnea syndrome patients. Sleep Breath. 2017;21:303–309. doi: 10.1007/s11325-016-1409-x. [DOI] [PubMed] [Google Scholar]
- 239.Htoo AK, Greenberg H, Tongia S, et al. Activation of nuclear factor kappaB in obstructive sleep apnea: a pathway leading to systemic inflammation. Sleep Breath. 2006;10:43–50. doi: 10.1007/s11325-005-0046-6. [DOI] [PubMed] [Google Scholar]
- 240.Matafome P, Seiça R. The role of brain in energy balance. Adv Neurobiol. 2017;19:33–48. doi: 10.1007/978-3-319-63260-5_2. [DOI] [PubMed] [Google Scholar]
- 241.Rui L. Brain regulation of energy balance and body weight. Rev Endocr Metab Disord. 2013;14:387–407. doi: 10.1007/s11154-013-9261-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Waterson MJ, Horvath TL. Neuronal regulation of energy homeostasis: beyond the hypothalamus and feeding. Cell Metab. 2015;22:962–970. doi: 10.1016/j.cmet.2015.09.026. [DOI] [PubMed] [Google Scholar]
- 243.Yang CS, Lam CK, Chari M, et al. Hypothalamic AMP-activated protein kinase regulates glucose production. Diabetes. 2010;59:2435–2443. doi: 10.2337/db10-0221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Liu XM, Peyton KJ, Shebib AR, Wang H, Korthuis RJ, Durante W. Activation of AMPK stimulates heme oxygenase-1 gene expression and human endothelial cell survival. Am J Physiol Heart Circ Physiol. 2011;300:H84–93. doi: 10.1152/ajpheart.00749.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Zimmermann K, Baldinger J, Mayerhofer B, Atanasov AG, Dirsch VM, Heiss EH. Activated AMPK boosts the Nrf2/HO-1 signaling axis--A role for the unfolded protein response. Free Radic Biol Med. 2015;88:417–426. doi: 10.1016/j.freeradbiomed.2015.03.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Maddock RJ. The lactic acid response to alkalosis in panic disorder : an integrative review. J Neuropsychiatry Clin Neurosci. 2001;13:22–34. doi: 10.1176/jnp.13.1.22. [DOI] [PubMed] [Google Scholar]
- 247.Vollmer LL, Strawn JR, Sah R. Acid-base dysregulation and chemosensory mechanisms in panic disorder: a translational update. Transl Psychiatry. 2015;5:e572. doi: 10.1038/tp.2015.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Wemmie JA. Neurobiology of panic and pH chemosensation in the brain. Dialogues Clin Neurosci. 2011;13:475–483. doi: 10.31887/DCNS.2011.13.4/jwemmie. [DOI] [PMC free article] [PubMed] [Google Scholar]