

Abstract
Recreational use of synthetic cannabinoids (SCs) is associated with desirable euphoric and relaxation effects as well as adverse effects including anxiety, agitation and psychosis. These SC-mediated actions represent a combination of potentiated cannabinoid receptor signaling and “off-target” receptor activity. The goal of this study was to compare the efficacy of various classes of SCs in stimulating CB1 receptors and activating “off-target” transient receptor potential (TRP) channels. Cannabinoid-type 1 (CB1) receptor activity was determined by measuring SC activation of G protein-gated inward rectifier K+ (GIRK) channels using a membrane potential-sensitive fluorescent dye assay. SC opening of vanilloid type-1 (TRPV1) channels was measured by recording intracellular Ca2+ transients. All of the SCs tested activated the GIRK channel with an efficacy of 4-fluoro MDMB-BUTINACA > 5-fluoro MDMB-PICA > MDMB-4en-PINACA ≈ WIN 55,212-2 > AB-FUBINACA > AM1220 ≈ JWH-122 N-(5-chloropentyl) > AM1248 > JWH-018 ≈ XLR-11 ≈ UR-144. The potency of the SCs at the CB1 receptor was 5-fluoro MDMB-PICA ≈ 4-fluoro MDMB-BUTINACA > AB-FUBINACA ≈ MDMB-4en-PINACA > JWH-018 > AM1220 > XLR-11 > JWH-122 N-(5-chloropentyl) > WIN 55,212-2 ≈ UR-144 > AM1248. In contrast, when tested at a SC concentration that produced a maximal effect on the Gi/GIRK channel, only XLR-11, UR-144 and AM1220 caused a significant activation of the TRPV1 channels. The TRPV1 channel/Ca2+ signal measured during application of 10 µM XLR-11 was similar to the signal induced by the endocannabinoid N-arachidonoylethanolamine (AEA). Thus, while various SCs share the ability to stimulate CB1 receptor/Gi signaling, they display limited efficacy in opening TRPV1 channels.
Keywords: synthetic cannabinoids, G protein-gated inward rectifier K+ channel, transient potential receptor channels, fluorescence measurements
1. Introduction
Synthetic cannabinoids (SCs) are found in recreational drug products that are sold under brand names such as “K2”, “Spice,” and “Black Mamba” (Gurney et al., 2014; Alves, et al., 2020). While these SCs products induce psychotropic effects similar to marijuana, they also produce serious side effects including seizures, respiratory depression, cardiac arrhythmias, stroke, kidney failure and even death (Thornton et al, 2013; Takematsu et al., 2014; Adams et al., 2016; Kelkar et al., 2018). Factors that mediate SC toxicity are postulated to be a combination of potentiated cannabinoid receptor signaling and “off-target” activity (Alipour et al., 2019; Yano et al., 2020). As the prevalence of both legal and illegal cannabinoid products continues to increase, it is critical to understand how receptor interactions and subsequent intracellular events bring about both desired and adverse effects of SCs.
The cannabinoid-type 1 (CB1) and cannabinoid-type 2 (CB2) receptors are the two major receptors in the endocannabinoid system (Kendall & Yudowski, 2016). The CB1 receptor is a G protein-coupled receptor (GPCR) expressed in the peripheral and central nervous systems (Howlett et al., 2002). Binding of ∆9-tetrahydrocannabinol (THC), the major psychoactive cannabinoid found in marijuana, to the neuronal CB1 receptor causes the dissociation of the βγ subunits of pertussis toxin-sensitive G proteins (Gi/Go) from the α subunit (Giα) (Glass & Northup, 1999; Mukhopadhyay & Howlett, 2005). Giα decreases intracellular levels of cAMP by inhibiting adenylyl cyclase (Howlett et al., 1986; Glass & Northup, 1999). In comparison, Giβγ activates G protein-gated inward rectifier K+ (GIRK) channels while inhibiting N- and Q-type Ca2+ channels ( Mackie et al., 1995; Guo & Ikeda, 2004). While the CB1 receptor predominately couples to Gi, SC signaling effects through Gs and Gq have also been reported under certain conditions (Glass & Northup, 1999; Lauckner et al., 2005).
Transient receptor potential (TRP) channels are a family of inotropic cation channels that serve as important “off-target” receptors for cannabinoids. Endocannabinoids such as anandamide (N-arachidonoylethanolamine [AEA]) and phytocannabinoids such as cannabidiol (CBD) bind to and activate TRP channels causing cellular Ca2+ influx and a resulting stimulation of Ca2+-dependent pathways (Zygmunt et al., 1999; Smart et al., 2000; De Petrocellis et al., 2008). CBD activates vanilloid type-1 (TRPV1) and type-2 (TRPV2) channels, along with ankyrin type-1 (TRPA1) channels (De Petrocellis et al., 2008; De Petrocellis et al., 2011). Cannabinoid activation of TRPV1 channels may partially underlie their effectiveness in treating inflammatory and neuropathic pain (Costa et al., 2007).
The naphthoylindole compound JWH-018 was one of the first SCs to be identified on the illicit drug market. As with other SCs, JWH-018 displays a higher CB1 receptor binding affinity and efficacy than THC (Wiley et al., 1998). Since the identification of JWH-018, other naphthoylindole (AM1220, AM2201), as well as tetramethylcyclopropyl indole (UR-144, XLR-11), indole carboxamide (AB-PICA, AB-FUBICA) and indazole carboxamide (AB-PINACA, MDMB-FUBINACA) SCs have appeared in K2/Spice products. While previous studies have characterized the CB1 receptor-mediated signaling profile of select SCs (Laprairie et al., 2014; Banister et al., 2015a; Finlay et al., 2019; Patel et al, 2020), the comparative effects of SCs on CB1 receptors and TRP channels have not been determined. The goal of this study was to compare the efficacy of various structural groups of SCs in activating GIRK channels (via CB1 receptors) and increasing intracellular levels of Ca2+ (via TRPV1 channels). All of the SCs tested in the study activated the GIRK channels with EC50s in the nanomolar to low micromolar range. In contrast, when tested at a SC concentration that caused maximal activation of the GIRK channels, only XLR-11, UR-144 and AM1220 stimulated TRPV1 channel opening.
2. Materials and methods
2.1. AtT20/SEPCB1 and HEK293T cell culture and plating.
The AtT20 pituitary cell line was obtained from ATCC (AtT-20/D16y-F2, CRL-1795) and grown in Dulbecco’s Modified Eagle Medium (DMEM) with 10 % fetal bovine serum and Pen-Strep. AtT20 cells were stably transfected with lentivirus vectors containing either the human cannabinoid type-1 (CB1) receptor tagged with a super-ecliptic pHluorin (SEP CB1) (from Dr. Andrew Irving, University College Dublin) (McDonald et al., 2007) or GFP. The tagged-CB1 receptor displays a CB1 receptor signaling response similar to the unmodified receptor (Andersen et al., 2018). CB1 receptor expression was verified using a fluorescent microscope (Leica DM IL). Cells were plated in poly-l-lysine-coated wells of black 96-well plates (Greiner Bio-One) (30,000 cells per well). HEK293T cells were cultured as above with the addition of 2 mM glutamine to the media and transiently transfected with the human TRPV1 channel (Origene cDNA #SC128105) or GFP using Lipofectamine 2000 (Invitrogen). AtT20 and HEK293T cells were stored in an incubator at 37° C (5 % O2 / 95 % CO2) and used on days 1–3 after plating.
2.2. GIRK and TRPV1 channel fluorescent assays.
GIRK channel activation was monitored in the 96-well plates by fluorescently recording the cell membrane potential (MP) as previously described (Walsh, 2011; Andersen et al., 2018). For the MP measurements, the AtT20-SEPCB1 cells were incubated for 30 min in buffer solution consisting of; 132 mM NaCl, 5 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 5 mM dextrose, 5 mM HEPES, pH 7.4 (with NaOH), with a MP-sensitive fluorescent dye (MPSD) (FLIPR Membrane Potential kit BLUE; Molecular Devices). Prior to the fluorescent measurements, the cells were loaded with MPSD in buffer solution (as above), but containing 1 mM KCl and incubated for an additional 5 min. Fluorescent signals were recorded using a Synergy2 microplate reader (Biotek) (Walsh, 2011; Andersen et al., 2018). Cannabinoids were dissolved in DMSO at stock concentrations of 20 mM to 50 mM (final DMSO = 0.05 to 0.1 %). All cannabinoids were diluted to working concentrations in 1 mM KCl buffer solution containing the MPSD. Due to solubility issues, concentrations of the SCs above 20 µM were not tested. The cannabinoids or control solution (10 or 20 µl) were injected into each well (total volume = 110 or 220 µl) at time zero. Data points were collected at 5 s intervals over a 250 s sampling period at excitation and emission wavelengths of 520 and 560 nm, respectively. SCs that stimulate the CB1 receptor cause a decrease in the fluorescent signal (see supplementary Fig. 1A and Fig. 1B). In contrast, no changes in the signal are observed with CB2 receptor agonists or control solution (see supplementary Fig. 1A).
For the Ca2+ fluorescent measurements, HEK293T-TRPV1 cells were incubated for 30 min in buffer solution containing 10 µM Fluo-4 acetoxymethyl ester (Fluo-4/AM) and 0.01 % Pluronic (Invitrogen). The cells were then be washed several times without the dye and incubated for 10–15 min to facilitate the de-esterification of the dye. Fluorescent measurements were made using the Synergy plate reader equipped with a 480 nm excitation/ 520 nm emission filter set. In order to enhance the TRPV1 channel/Ca2+ signal, fluorescent recordings were made in buffer solution (as above), but containing 5 mM Ca2+. For each experimental plate the SC-induced signal was normalized to the peak signal obtained with 1 µM capsaicin.
2.3. Data analysis
Concentration versus response curves obtained from the MP and Ca2+ fluorescent experiments were fit using four-parameter, non-linear regression analysis: y = ymin + (ymax – ymin / 1 + (Drug/EC50)-Hillslope), where EC50 is the concentration producing a 50 % increase in the maximal response ymax (Emax), ymin is defined as a minimum fluorescent channel response and Hill slope is the slope factor. Relative Emax values for the GIRK channel signal were determined by normalizing the GIRK channel fluorescent response of each SC to the maximal GIRK channel response measured in each plate to 10 µM WIN 55,212-2. The SC Ca2+ fluorescent signals were normalized to the response obtained with 1 µM capsaicin.
2.4. Statistical analysis
Statistical significance of the comparison of cannabinoid GIRK channel Emax data was determined using a one-way, multiple measures ANOVA with significance set at P < 0.05. Significant differences were identified using a Holms-Ŝídák posthoc analysis. Curve fitting and statistical analyses were performed using Sigmaplot 14.0.
2.5. Drugs and Chemicals
The following compounds were purchased from Cayman Chemical: 4-fluoro MDMB-BUTINACA, 5-fluoro MDMB-PICA, AB-FUBINACA, AM1220, AM1248, JWH-018, JWH-122 N-(5-chloropentyl), MDMB-4en-PINACA, WIN 55,212-2, UR-144, XLR-11, anandamide (AEA) and JWH-133. Capsaicin was obtained from Sigma-Aldrich (St. Louis, MO). Controlled substances were purchased using the Walsh Laboratory DEA license.
3. Results
3.1. SC activation of GIRK channels
The AtT20-SEPCB1 cells used in these experiments endogenously express GIRK (Kir3.1/3.2) channels and were stably transfected with a tagged human CB1 receptor using lentivirus. Binding of SCs to the CB1 receptor stimulates the dissociation of the Giβγ subunits of Gi from the Gα subunit. Giβγ then activates the GIRK channels causing a cellular efflux of K+ and a concomitant decrease in the cell resting MP. Therefore, CB1 receptor/Gi activity was determined by using a fluorescent MPSD assay (Walsh, 2011; Andersen et al., 2018). GIRK channel fluorescent responses were measured for ten SCs commonly found in illicit substances and compared with the prototypic aminoalkylindole SC WIN 55,212-2 (see Table 1). SC concentration versus response curves were then obtained and fit to determine the potency (EC50) and efficacy (Emax) of the compounds.
Table 1:
Synthetic Cannabinoid Structures
| Cannabinoid | Cannabinoid | ||
|---|---|---|---|
| WIN 55, 212-2 |

|
UR-144 |

|
| JWH-018 |

|
XLR-11 |

|
| JWH-122 N-(5-chloropentyl) |

|
5F-MDMB-PICA |

|
| AM1220 |
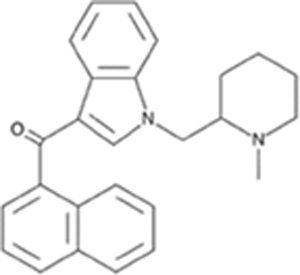
|
AB-FUBINACA |

|
| AM1248 |

|
4F-MDMB-BUTINACA |

|
| JWH-133 |

|
MDMB-4en-PINACA |

|
Fig. 1 displays MPSD fluorescent responses and concentration vs. response curves for representative naphthoylindole (JWH-018 & AM1220) and tetramethylcyclopropyl indole (XLR-11) SCs, while Fig. 2 shows the results obtained with representative indazole carboxylate compounds (AB-FUBINACA, 4-fluoro MDMB-BUTINACA & MDMB-4en-PINACA). All of the SCs induced a GIRK channel fluorescent response with EC50s in the nanomolar to low micromolar concentration range (Table 2). Only AM1248 was less potent than WIN 55,212-2 (EC50 = 523 nM) with an EC50 of 2530 nM (Table 2). Overall, the rank order of potencies was 5-fluoro MDMB-PICA ≈ 4-fluoro MDMB-BUTINACA > AB-FUBINACA ≈ MDMB-4en-PINACA > JWH-018 > AM1220 > XLR-11 > JWH-122 N-(5-chloropentyl) > WIN 55,212-2 ≈ UR-144 > AM1248. In order to quantify SC efficacies, concentration vs. response curves were normalized to the Emax of WIN 55,212-2 (Figs. 1 & 2). When compared with WIN 55,212-2, most of the SCs were significantly less effective in activating the GIRK channels. The indole and indazole carboxamides, 5-fluoro MDMB-PICA and 4-fluoro MDMB-BUTINACA, were the only SCs to stimulate a CB1 receptor/GIRK channel response greater than WIN 55,212-2 (Fig. 2, Table 2). The order of efficacies was 4-fluoro MDMB-BUTINACA > 5-fluoro MDMB-PICA > MDMB-4en-PINACA ≈ WIN 55,212-2 > AB-FUBINACA > AM1220 ≈ JWH-122 N-(5-chloropentyl) > AM1248 > JWH-018 ≈ XLR-11 ≈ UR-144.
Fig. 1.

Activation of GIRK channel fluorescent signal by SCs. A: Changes in the MPSD signal following injection of 10 µM of the indicated SCs into wells containing the AtT20-SEPCB1 cells. Each point represents the mean ± S.E.M. obtained in 4–6 wells. The SCs were added at time zero (↓). B: Concentration versus response curve for the GIRK channel fluorescent signal. The maximal response of each SC (as in panel A) was normalized to the GIRK channel signal measured with 10 µM WIN 55,212-2 and the resulting curve fit using the 4 parameter model described in the Methods.
Fig. 2.

Activation of GIRK channel fluorescent signal by SCs. A: Changes in the MPSD signal following injection of either 1 µM or 10 µM of the indicted SCs into wells containing the AtT20-SEPCB1 cells. Each point represents the mean ± S.E.M. obtained in 4–6 wells. The SCs were added at time zero (↓). B: Concentration versus response curve for the GIRK channel fluorescent signal. The maximal response of each SC (as in panel A) was normalized to the GIRK channel signal measured with 10 µM WIN 55,212-2 and the resulting curve fit using the 4-parameter model described in the Methods.
Table 2:
Sythetic Cannabinoid Potency and Efficacy
| Class | Cannabinoid | EC50 (nM) ± S.E.M. | GIRK Response (%) ± S.E.M. |
|---|---|---|---|
| Indole-Based | |||
| Aminoalkylindole Prototype | WIN 55, 212-2 | 523 ± 68.0 | Reference |
| Napthoylindoles | JWH-018 | 46.3 ± 4.8 | 55.8 ± 6.1 |
| JWH-122 N-(5chloropentyl) | 231 ± 35.4 | 74.0 ± 3.4a | |
| AM1220 | 172 ± 6.2 | 78.3 ± 2.7a | |
| Adamantoylindole | AM1248 | 2530 ± 805 | 63.5 ± 3.9a |
| Tetracyclopropylindoles | UR-144 | 532 ± 33.9 | 51.3 ± 3.3a |
| XLR-11 | 214 ± 14.9 | 53.0 ± 2.6a | |
| Indole Carboxamide | 5-fluoro-MDMB-PICA | 3.33 ± 0 .3 | 108 ± 4.3 |
| Indazole-Based | |||
| Indazole Carboxamide | AB-FUBINACA | 18.1 ± 7.0 | 89 ± 2.9 |
| 4-fluoro-MDMBBUTINACA | 4.97 ± 2.6 | 115 ± 2.9b | |
| MDMB-4en-PINACA | 19.7 ± 2.9 | 101 ± 5.2 |
P < .001 compared to WIN 55, 212-2
P < .05 compared to WIN 55, 212-2
3.2. SC activation of TRPV1 channels
HEK293T cells were transiently transfected with TRPV1 channels and intracellular Ca2+ measured following the injection of the TRPV1 channel agonist capsaicin, the endocannabinoid AEA or the SCs (Fig. 3). Application of 1 µM capsaicin resulted in a large Ca2+ fluorescent signal that rapidly decayed over the 250 second recording time. Compared with capsaicin, AEA (10 µM) produced a statistically smaller Ca2+ transient that showed little decay. High concentrations of SCs such as HU-201, CP 55,940 and others are associated with non-receptor-mediated increases in Ca2+ in CHO cells (Felder et al., 1992). For this reason, SCs were tested at a concentration causing a maximum CB1 receptor/GIRK channel response (as determined above). As shown in Fig. 3A, 10 µM XLR-11 produced a Ca2+ transient of the same amplitude and with similar kinetics to AEA. The maximal effect of each of the SCs on TRPV1 channel activation was normalized by the TRPV1 signal measured with capsaicin. Of the SCs tested in the assay, only XLR-11, UR-144 and AM1220 caused a significant activation of the TRPV1 channels over the control solution (containing DMSO) (Fig. 3B). Concentration versus response curves for these SCs were generated and compared to that of AEA in Fig. 3C. The rank order of potencies was AEA > XLR-11 > UR-144 ≈ AM1220. The order of efficacies was AEA ≈ XLR-11 > UR-144 ≈ AM1220. Ca2+ transients were not observed in nontransfected HEK293T cells treated with a 10 µM concentration of XLR-11, UR-144 or AM1220 (not shown). We also confirmed that the CB2 receptor agonist JWH-133, previously reported to activate TRPV1 channels (Soethoudt et al., 2017), also produced a Ca2+ signal in the TRPV1-expressing cells (Fig. 3B). As expected, no CB1 receptor-mediated activation of the GIRK channels was measured in the presence of JWH-133 (supplementary Fig. 1B).
Fig. 3.

Activation of TRPV1 channel Ca2+ fluorescent signal by cannabinoids. A: Changes in the Ca2+ signal following injection of capsaicin, AEA or XLR-11 into wells containing the HEK239T-TRPV1 cells. Control data were obtained by adding capsaicin to nontransfected HEK239T cells. Each point represents the mean ± S.E.M. obtained in 6–8 wells. Compounds were added at time zero (↓). Ca2+ transients were not observed in nontransfected cells during treatment with capsaicin, AEA or XLR-11. B: Summary of cannabinoid activation of TRPV1 channels in HEK239T-TRPV1 cells. The maximal response of each cannabinoid was normalized to the Ca2+ fluorescent signal measured with 1 µM capsaicin measured in the same plate. Control data were obtained by injection of the solution containing solvent (DMSO) into the wells. Each bar displays the mean ± S.E.M. normalized signal obtained in 3–4 experiments as in panel A. *P < 0.05 compared with control. C: Concentration versus response curve for the TRPV1 channel fluorescent signal. The maximal response of each compound (as in panel B) was normalized to the signal measured with 1 µM capsaicin and the resulting curve fit using the 4-parameter model described in the Methods. Fitted parameters for the EC50 and Emax were: AEA = 2.1 µM & 0.35, XLR-11 = 5.3 µM & 0.39, UR-144 = 7.9 µM & 0.23 and AM1220 = 6.8 µM & 0.26.
4. Discussion
The availability of a growing list of novel SCs has resulted in cases of acute toxicity in which medical personnel struggle to develop an effective treatment plan. While SCs act as CB1 and CB2 receptor agonists and antagonists, recent observations indicate that some of their effects are mediated through binding to “off target” receptors (De Petrocellis et al., 2011; Yano et al., 2020). Suggested “off target” sites for SCs include 5-HT1a receptors, the G-protein-coupled receptor 55 (GPR55), glycine and glutamate receptors, voltage-gated Na+ channels and TRP channels (De Petrocellis et al., 2011; Soethoudt et al., 2017; Senn et al., 2020; Yano et al., 2020). In this study, we compared the activity of a structurally diverse group of SCs on the CB1 receptor and TRPV1 channel. While all the SCs tested in this study activated the GIRK channels with EC50s in the nanomolar to low micromolar concentration range, only a small number of compounds were effective in opening the TRPV1 channels.
As summarized in Table 2, most of the SCs examined in our study had a greater potency than WIN 55,212-2 at the CB1 receptor. In contrast, only 4-fluoro MDMB-BUTINACA caused a significantly greater GIRK channel fluorescent signal when compared to WIN 55,212-2. Overall, the indole and indazole carboxylates were the most potent and efficacious of the SCs. Members of this family of SCs, such as AB-FUBINACA, have been recently linked to severe toxicity and death (Adams et al., 2016). Using the GIRK channel/membrane potential hyperpolarization assay to measure CB1 receptor activity, Banister et al. (2015a) and Longworth et al. (2017) have reported that indole and indazole carboxamide SCs have the greatest CB1 receptor potency. For example, AB-FUBINACA and 5-fluoro ABD-PINACA display ten- and one hundred-fold greater potencies respectively, when compared with the naphthoylindole compounds JWH-018 and AM2201. In general, this is consistent with the greater potency and activity of indole and indazole carboxamide-type SCs in inhibiting adenylyl cyclase, stimulating mitogen-activated protein kinases such as pERK1/2 and recruiting β-arrestin proteins to the CB1 receptor (Finlay et al., 2019; Patel et al., 2020). In addition, EC50s reported in our study were comparable to previous EC50s measured using the GIRK channel assay. EC50s reported for AB-FUBINACA, JWH-018, XLR-11, WIN 55,212-2 and UR-144 were 18 nM, 46 nM, 214 nM, 523 nM and 532 nM, respectively (Table 2), compared with 1.8 nM, 18 & 102 nM, 98 nM, 282 nM and 417 nM (Banister et al., 2015a; Banister et al., 2015b).
Terminal fluorination of pentylindole SCs is a popular chemical modification found in K2/Spice products. We found that XLR-11, a fluorinated tetramethylcyclopropyl compound, was more potent in activating the GIRK channels than the nonfluorinated parent compound UR-144 (Table 2). In addition, the fluorinated carboxamide indole and indazole SCs, 5-fluoro MDMB-PICA and 4-fluoro MDMB-BUTINACA, were the most potent activators of the GIRK channels tested in our study. However, the activities of the nonfluorinated parent compounds were not determined. Previous results have shown that addition of a fluoride atom in AM2201 (terminal fluorination of JWH-018) results in greater CB1 receptor binding affinity (Makriyannis & Deng, 2001). The fluorinated analogs of the SCs UR-144, PB-22 and APICA (XLR-11, 5F-PB-22 and STS-135, respectively) also show increased CB1 receptor potency (Banister et al., 2015b). In addition to fluoride, halogenated SC compounds including Cl, Br and I analogs of JWH-122 display activity at the CB1 receptor (Noble et al., 2019). In agreement with this finding, JWH-122 N-(5-chloropentyl) produced a strong GIRK channel response in our assay. However, it was less potent than JWH-018 indicating that addition of a methyl group and terminal chlorination (in JWH-122 N-(5-chloropentyl)) decreases CB1 receptor activity.
SCs are non-polar, lipid soluble compounds that readily cross the blood brain barrier and exert effects on the CNS. The toxicological and mechanistic actions of the SCs have previously been investigated in neuronal cell cultures. SCs such as CP-55,490, JWH-018 and MAM-2201 cause cytotoxic effects in mouse forebrain, human neuroblastoma SH-SY5Y and D384 astrocyte cultures, as well as neuronal cells derived from human umbilical cord mesenchymal stem cells (Tomiyama & Funada, 2014; Wojcieszak et al., 2016; Coccini et al., 2021). Atwood et al., (2010) reported that JWH-018 inhibited excitatory postsynaptic currents (EPSCs) in cultured autaptic hippocampal neurons. JWH018 inhibited EPSCs at low concentrations (IC50 = 15 nM) and this inhibition was prevented by the CB1 receptor antagonist rimonabant. Similar neuronal inhibitory effects, including a suppression of Ca2+ spikes and an inhibition of electrical activity in hippocampal neurons cultured on multi-electrode arrays, were observed with JWH-018, MAM-2201 and other SCs (Tauskela et al., 2016). In addition, MAM-2201 suppresses glutamatergic and GABAergic neurotransmission via CB1 receptors when measured in rodent brain slice preparations (Irie et al., 2015). Thus, these neuronal toxicological and inhibitory actions may account in part for the various neurological manifestations noted during recreational use of SCs (Gurney et al., 2014; Adams et al., 2016).
Several TRP channels, including the TRPV and TRPA subtypes, represent novel molecular targets for cannabinoids. TRPV1 channels are expressed in both peripheral and CNS nerve terminals where they co-localize with CB1 receptors (Cristino et al., 2006). Endocannabinoids, such as anandamide (AEA) and phytocannabinoids including CBD, cannabidivarin (CBDV), cannabigerol (CBG) and cannabigevarin (CBGV) activate TRPV1 channels (De Petrocellis et al., 2011; Muller et al., 2019). In addition, several SC CB2 receptor ligands activate TRPV1 channels with EC50s in the micromolar concentration range (Soethoudt et al., 2017). Of these CB2 ligands JWH-133 (EC50 = 8.2 µM) was reported to be the most potent (Soethoudt et al., 2017). In the present study, the effects of the SCs were examined on TRPV1 channel activation using a SC concentration that produced a maximal activation of the CB1 receptor/Gi signaling pathway. At this concentration only XLR-11, UR-144 and AM1220 elicited a significant TRPV1 channel Ca2+ signal (Fig. 3). The Ca2+ signal produced by 10 µM XLR-11 was similar in amplitude and kinetics to the signal measured in the presence of 10 µM AEA. We also confirmed that JWH-133 activates the TRPV1 channel at high micromolar concentrations.
5. Conclusion
Data from the present study demonstrate that while various structural classes of SCs share the ability to stimulate CB1 receptor/Gi signaling, they display limited efficacy in opening TRPV1 channels. The tetramethylcyclopropyl indole compound XLR-11 was the most efficacious of the SCs examined in activating the TRPV1 channel and produced a Ca2+ signal comparable to AEA. Thus, analogs of XLR-11 may provide a promising source for identifying new cannabinoid TRP channel agonists.
Supplementary Material
Funding
This work was supported by US Public Health Service award NS-071530 and National Science Foundation award CBET-1606882 (KBW).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Declaration of Interest: None
References
- Adams AJ, Banister SD, Irizarry L, Trecki J, Schwartz M, & Gerona R (2016). “Zombie” outbreak caused by the synthetic cannabinoid AMB-FUBINACA in new york. New Eng. J. Med 376(3), 235–242. doi: 10.1056/NEJMoa1610300 [DOI] [PubMed] [Google Scholar]
- Alipour A, Patel PB, Shabbir Z, & Gabrielson S (2019). Review of the many faces of synthetic cannabinoid toxicities. Mental Health Clin 9(2), 93–99. doi: 10.9740/mhc.2019.03.093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alves VL, Gonçalves JL, Aguiar J, Teixeira HM, & Câmara JS (2020). The synthetic cannabinoids phenomenon: from structure to toxicological properties. A review. Critical Rev.Toxicol 50(5) 359–382. doi: 10.1080/10408444.2020.1762539 [DOI] [PubMed] [Google Scholar]
- Andersen HK, Piroli GG, & Walsh KB (2018). A real time screening assay for cannabinoid CB1 receptor-mediated signaling. J. Pharmacol. Toxicol. Methods 94, 44–49. doi: 10.1016/j.vascn.2018.05.001 [DOI] [PubMed] [Google Scholar]
- Atwood BK, Huffman J, Straiker A, & Mackie K (2010). JWH-018, a common constituent of ‘Spice’ herbal blends, is a potent and efficacious cannabinoid CB1 receptor agonist. Br. J. Pharmacol 160, 585–593. doi: 10.1111/j.1476-5381.2009.00582x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banister SD, Moir M, Stuart J, Kevin RC, Wood KE, Longworth M, et al. , (2015a). Pharmacology of indole and indazole synthetic cannabinoid designer drugs AB-FUBINACA, ADB-FUBINACA, AB-PINACA, ADB-PINACA, 5F-AB-PINACA, 5F-ADB-PINACA, ADBICA, and 5F-ADBICA. ACS Chemical Neurosci 6(9), 1546–1559. doi: 10.1021/acschemneuro.5b00112 [DOI] [PubMed] [Google Scholar]
- Banister SD, Stuart J, Kevin RC, Edington A, Longworth M, Wilkinson SM, et al. (2015b). Effects of bioisosteric fluorine in synthetic cannabinoid designer drugs JWH-018, AM-2201, UR-144, XLR-11, PB-22, 5F-PB-22, APICA, and STS-135. ACS Chem. Neurosci, 6(8), 1445–1458. doi: 10.1021/acschemneuro.5b00107 [DOI] [PubMed] [Google Scholar]
- Coccini T, De Simone U, Lonati D, Scaravaggi G, Marti M & Locatelli CA (2021). MAM-2201, one of the most potent-naphthoyl indole derivative-synthetic cannabinoids, exerts toxic effects on human cell-based models of neurons and astrocytes. Neurotoxicity Res May 4th online. doi: 10.1007/s12640-021-00369-3 [DOI] [PubMed] [Google Scholar]
- Costa B, Trovato AE, Comelli F, Giagnoni G, & Colleoni M (2007). The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and neuropathic pain. Eur. J. Pharmacol 556(1), 75–83. doi: 10.1016/j.ejphar.2006.11.006 [DOI] [PubMed] [Google Scholar]
- Cristino L, de Petrocellis L, Pryce G, Baker D, Guglielmotti V, & Di Marzo V (2006). Immunohistochemical localization of cannabinoid type 1 and vanilloid transient receptor potential vanilloid type 1 receptors in the mouse brain. Neurosci, 139(4), 1405–1415. doi: 10.1016/j.neuroscience.2006.02.074 [DOI] [PubMed] [Google Scholar]
- De Petrocellis L, Ligresti A, Moriello AS, Allarà M, Bisogno T, Petrosino S, & Di Marzo V (2011). Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Brit. J. Pharmacol, 163(7), 1479–1494. doi: 10.1111/j.1476-5381.2010.01166.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Petrocellis L, Vellani V, Schiano-Moriello A, Marini P, Magherini PC, Orlando P, & Di Marzo V (2008). Plant-derived cannabinoids modulate the activity of transient receptor potential channels of ankyrin type-1 and melastatin type-8. J. Pharmacol. Exp.Ther 325, 1007–1015. [DOI] [PubMed] [Google Scholar]
- Felder CC, Veluz JSH, Williams HL, Briley EM & Matsuda LA (1992). Cannabinoid agonists stimulate both receptor and non-receptor-mediated signal transduction pathways in cell transfected with and expressing cannabinoid receptor clones. Mol. Pharm 42(5), 838–845. [PubMed] [Google Scholar]
- Finlay DB, Manning JJ, Ibsen MS, Macdonald CE, Patel M, Javitch JA, et al. , (2019). Do toxic synthetic cannabinoid receptor agonists have signature in vitro activity profiles? A case study of AMB-FUBINACA. ACS Chem. Neurosci 10(10), 4350–4360. doi: 10.1021/acschemneuro.9b00429 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glass M, & Northup JK (1999). Agonist selective regulation of G proteins by cannabinoid CB1 and CB2 receptors. Mol. Pharmacol 56, 1362–1369. doi: 10.1124/mol.56.6.1362 [DOI] [PubMed] [Google Scholar]
- Guo J, & Ikeda SR (2004). Endocannabinoids modulate N-type calcium channels and G-protein-coupled inwardly rectifying potassium channels via CB1 cannabinoid receptors heterologously expressed in mammalian neurons. Mol. Pharmacol 65, 665–674. doi: 10.1124/mol.65.3.665 [DOI] [PubMed] [Google Scholar]
- Gurney SMR, Scott KS, Kacinko SL, Presley BC, & Logan BK (2014). Pharmacology, toxicology, and adverse effects of synthetic cannabinoid drugs. Forensic Sci. Rev 26(1), 53–78. [PubMed] [Google Scholar]
- Howlett AC, Barth F, Bonner TI, Cabral G, Casellas P, Devane WA, et al. (2002). International Union of Pharmacology. XXVII. Classification of Cannabinoid Receptors. Pharmacol. Rev, 54(2), 161. doi: 10.1124/pr.54.2.161 [DOI] [PubMed] [Google Scholar]
- Howlett AC, Qualy JM, & Khachatrian LL (1986). Involvement of Gi in the inhibition of adenylate cyclase by cannabimimetic drugs. Mol. Pharmacol 29(3), 307. [PubMed] [Google Scholar]
- Irie T, Kikura-Hanajiri R, Usami M, Uchiyama N, Goda Y & Sekino Y (2015). MAM-2201, a synthetic cannabinoid drug of abuse, suppresses the synaptic input to cerebellar Purkinje cells via activation of presynaptic CB1 receptors. Neuropharmacol 95, 479–491. [DOI] [PubMed] [Google Scholar]
- Kelkar AH, Smith NA, Martial A, Moole H, Tarantino MD, & Roberts JC (2018). An outbreak of synthetic cannabinoid–associated coagulopathy in Illinois. New Eng. J. Med 379(13), 1216–1223. doi: 10.1056/NEJMoa1807652 [DOI] [PubMed] [Google Scholar]
- Kendall DA, & Yudowski GA (2016). Cannabinoid receptors in the central nervous system: Their signaling and roles in disease. Front. Cell. Neurosci 10. doi: 10.3389/fncel.2016.00294 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laprairie RB, Bagher AM, Kelly MEM, Dupré DJ, & Denovan-Wright EM (2014). Type 1 cannabinoid receptor ligands display functional selectivity in a cell culture model of striatal medium spiny projection neurons. J. Biol. Chem 289(36), 24845–24862. doi: 10.1074/jbc.M114.557025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauckner JE, Hille B, & Mackie K (2005). The cannabinoid agonist WIN 55,212-2 increases intracellular calcium via CB1 receptor coupling to Gq/11 G proteins. Proc. Natl. Acad. Sci. USA 102, 19144–191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Longworth M, Connor M, Banister SD, & Kassiou M (2017). Synthesis and pharmacological profiling of the metabolites of synthetic cannabinoid drugs APICA, STS-135, ADB-PINACA, and 5F-ADB-PINACA. ACS Chem. Neurosci 8(8), 1673–1680 [DOI] [PubMed] [Google Scholar]
- McDonald NA, Henstridge CM, Connolly CN, & Irving AJ (2007). Generation and functional characterization of fluorescent, N-terminally tagged CB1 receptor chimeras for live-cell imaging. Mol. Cell. Neurosci 35, 237–248. [DOI] [PubMed] [Google Scholar]
- Mackie K, Lai Y, Westenbroek R, & Mitchell R (1995). Cannabinoids activate an inwardly rectifying potassium conductance and inhibit Q-type calcium currents in AtT20 cells transfected with rat brain cannabinoid receptor. J. Neurosci 15(10), 6552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makriyannis A, & Deng H (2001). Preparation of cannabimimetic indole deriviatives with cannabinoid CB1 or CB2 receptor binding affinity. World patent WO 2001/028557.
- Mukhopadhyay S, & Howlett AC (2005). Chemically distinct ligands promote differential CB1 cannabinoid receptor-Gi protein interactions. Mol. Pharmacol 67(6), 2016–2024. doi: 10.1124/mol.104.003558 [DOI] [PubMed] [Google Scholar]
- Muller C, Morales P, & Reggio PH (2019). Cannabinoid ligands targeting TRP channels. Front. Mol. Neurosci 11, 487–487. doi: 10.3389/fnmol.2018.00487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noble C, Cannaert A, Linnet K, & Stove CP (2019). Application of an activity-based receptor bioassay to investigate the in vitro activity of selected indole- and indazole-3-carboxamide-based synthetic cannabinoids at CB1 and CB2 receptors. Drug Test. Anal 11(3), 501–511. doi: 10.1002/dta.2517 [DOI] [PubMed] [Google Scholar]
- Patel M, Manning JJ, Finlay DB, Javitch JA, Banister SD, Grimsey NL, & Glass M (2020). Signalling profiles of a structurally diverse panel of synthetic cannabinoid receptor agonists. Biochem. Pharmacol 175, 113871. doi: 10.1016/j.bcp.2020.113871 [DOI] [PubMed] [Google Scholar]
- Senn L, Cannazza G, & Biagini G (2020). Receptors and channels possibly mediating the effects of phytocannabinoids on seizures and epilepsy. Pharmaceuticals (Basel, Switzerland), 13(8), 174. doi: 10.3390/ph13080174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smart D, Gunthorpe MJ, Jerman JC, Nasir S, Gray J, Muir AI, et al. (2000). The endogenous lipid anandamide is a full agonist at the human vanilloid receptor (hVR1). Brit. J. Pharmacol 129(2), 227–230. doi: 10.1038/sj.bjp.0703050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soethoudt M, Grether U, Fingerle J, Grim TW, Fezza F, De Petrocellis, et al. (2017). Cannabinoid CB2 receptor ligand profiling reveals biased signalling and off-target activity. Nature Comm 8(1), 13958. doi: 10.1038/ncomms13958 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takematsu M, Hoffman RS, Nelson LS, Schechter JM, Moran JH, & Wiener SW (2014). A case of acute cerebral ischemia following inhalation of a synthetic cannabinoid. Clin.Toxicol 52(9), 973–975. doi: 10.3109/15563650.2014.958614 [DOI] [PubMed] [Google Scholar]
- Tauskela JS, Comas T, Hewitt M, Aylsworth A, Zhao X, et al. , (2016). Effect of synthetic cannabinoids on spontaneous neuronal activity: Evaluation using Ca2+ spiking and multi-electrode arrays. Eur. J. Pharmacol 786, 148–160. [DOI] [PubMed] [Google Scholar]
- Thornton SL, Wood C, Friesen MW, & Gerona RR (2013). Synthetic cannabinoid use associated with acute kidney injury. Clin.Toxicol 51(3), 189–190. doi: 10.3109/15563650.2013.770870 [DOI] [PubMed] [Google Scholar]
- Tomiyama KI, & Funada M (2014). Cytotoxicity of synthetic cannabinoids on primary neuronal cells of the forebrain: the involvement of cannabinoid CB1 receptors and apoptotic cell death. Toxicol. App. Pharmacol 274, 17–23. [DOI] [PubMed] [Google Scholar]
- Walsh KB (2011). Targeting GIRK channels for the development of new therapeutic agents. Front. Pharmacol 2, 64. doi: 10.3389/fphar.2011.00064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiley JL, Compton DR, Dai D, Lainton JAH, Phillips M, Huffman JW, & Martin BR (1998). Structure-activity relationships of indole- and pyrrole-derived cannabinoids. J. Pharmacol. Exp.Ther, 285(3), 995–1004. [PubMed] [Google Scholar]
- Wojcieszak J, Krzemién W, & Zawilska JB (2016). JWH-133, a selective cannabinoid CB2 receptor agonist, exerts toxic effects on neuroblastoma SH-SY5Y cells. J Mol. Neurosci 58, 441–445 [DOI] [PubMed] [Google Scholar]
- Yano H, Adhikari P, Naing S, Hoffman AF, Baumann MH, Lupica CR, & Shi L (2020). Positive allosteric modulation of the 5-HT1A receptor by indole-based synthetic cannabinoids abused by humans. ACS Chem. Neurosci, 11(10), 1400–1405. doi: 10.1021/acschemneuro.0c00034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zygmunt PM, Petersson J, Andersson DA, Chuang H-H, Sørgård M, Di Marzo V, et al. (1999). Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 400(6743), 452–457. doi: 10.1038/22761 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.