

Abstract
Huntington’s disease (HD) is one of neurodegenerative diseases, and is defined as a monogenetic disease due to the mutation of Huntingtin gene. This disease affects several cellular functions in neurons, and further influences motor and cognitive ability, leading to the suffering of devastating symptoms in HD patients. MicroRNA (miRNA) is a non-coding RNA, and is responsible for gene regulation at post-transcriptional levels in cells. Since one miRNA targets to several downstream genes, it may regulate different pathways simultaneously. As a result, it raises a potential therapy for different diseases using miRNAs, especially for inherited diseases. In this review, we will not only introduce the update information of HD and miRNA, but also discuss the development of potential miRNA-based therapy in HD. With the understanding toward the progression of miRNA studies in HD, we anticipate it may provide an insight to treat this devastating disease, even applying to other genetic diseases.
Keywords: Huntington’s disease (HD), Pathogenesis, microRNA (miRNA), Gene regulation, miRNA-based therapy
Background
Huntington’s disease (HD) is one of inherited diseases, and leads to neurodegeneration in patients. Due to the progression of this disease, the patients suffer severe motor and cognitive deficits till death, and unfortunately there is no cure for this disease yet. As a result, numbers of studies have tried to demonstrate on molecular mechanisms of this disease, and anticipate to identify or develop novel strategies for therapy. Recently, microRNA (miRNA) is a raising candidate to apply in gene therapy due to critically regulatory roles of gene expression inside cells. Taking advantages from specific characteristics of miRNAs, researchers have been developing miRNA-based therapy for different inherited diseases, and hope to alleviate or cure these diseases, including neuronal diseases. In this article, we will review the molecular pathogenesis of HD, and show the recent development of therapy for this disease. In addition, we will also review the applications of miRNAs targeting on neuronal pathogenesis, and discuss the pons and cons of this potential miRNA-based strategy.
Huntington’s disease
General introduction
Genetic information
Huntington’s disease (HD) is an autosomal dominant disease, and the clinic description of this disease was first reported by the American physician, George Huntington, in 1872. This disease belongs to rare diseases, and the prevalence rate of HD in the US and Canada is approximately 6.52–13.7 patients per 100,000 population [1]. Additionally, HD is a monogenic disease, and the disease-causing gene, Huntingtin (HTT; also known as IT15), was first identified in 1993 [2]. This HTT gene is located in chromosome 4p16.3 region, and has 67 exons to be translated into an approximately 350 kDa HTT protein. Most importantly, there is a critical expansion of CAG trinucleotide repeats in the exon 1 region of HTT gene, and patients carrying mutant HTT (mHTT) with more than 36 CAG repeats will develop the symptoms of HD gradually. Due to that the abnormally expanded CAG trinucleotides could be translated into glutamines, HD is also considered as one of polyglutamine (ployQ) diseases [2, 3]. Commonly, the onset of HD is approximately at 40 years of age [4], and the cause of death usually is suicide due to unaffordable conditions of symptoms for patients [5]. Unfortunately, these is still no cure for this disease, and physicians only could treat this disease by alleviation of clinic symptoms.
Pathology
HTT is dominantly expressed in brains, and mHTT forms toxic gain-of function proteins to disrupt neuronal functions, leading to cell death and neurodegeneration. The most affected brain regions in HD patients include deeper layers of the cortex and striatum, and mHTT also additionally influences other brain regions, such as hippocampus, hypothalamus and cerebellum [3, 6]. Within different brain regions, medium spiny neurons in striatum, including caudate and putamen, are considered as the most susceptible cells affected by mHTT, resulting in neuronal death, striatum atrophy and enlarged lateral ventricles in HD patients [6, 7]. Due to deficits of striatum, HD also causes in the imbalance of glutamate and dopamine activity, further disrupting the circuitry in basal ganglia [8]. As to the cellular hallmarks of HD neuropathology, nuclear aggregates, intranuclear inclusions (also known as inclusion bodies) and neuropil aggregates are typical characteristics in postmortem brains of HD patients [6, 9], which are also often used as an indicator for the severity of HD.
Clinical symptoms
Due to disruption of neuronal cells in brains, HD patients display cognitive and motor dysfunctions. In HD patients, they suffer different illness, such as emotional problems, mental deterioration, fatigue, difficulty thinking, memory loss, impaired sleep or daytime sleepiness [3, 9, 10]. In addition, these patients also show chorea, dystonia, involuntary movements and difficulty walking, speaking and swallowing because of the impaired ability of motor control in the central nervous system (CNS) [3, 9, 11]. With the worse cognitive and motor functions, HD patients change behaviors and gradually lose independence, leading to huge burdens for their families and societies.
Molecular pathogenesis
Protein misfolding
Full length of HTT/mHTT contains several protease cleavage sites, where capsase-3, caspase-6, calpain or aspartic proteases could digest full length HTT to form different sizes of truncated N-terminal HTT/mHTT fragments [3, 12–14]. In HD, over-expanded polyQ accelerates N-terminal mHTT fragments to transform to β-sheet confirmations from α-helical coiled coils, and easily leads to protein misfolding. Especially, the first 17 residues of N-terminal HTT preferentially adopt the β-sheet conformations as the polyQ increases, further enhancing the accumulation of protein misfolding [15]. These misfolded mHTTs generate dimers, trimers or oligomers, and form pathological aggregates. As a result, the misfolded mHTTs disrupt gene regulation, vehicle trafficking, synaptic plasticity, protein clearance, neurogenesis, etc., and finally lead to cellular dysfunctions and death [3, 16–19] (Fig. 1). Since this protein misfolding is a critical cause for HD, several therapeutical strategies have addressed on this aspect.
Fig. 1.
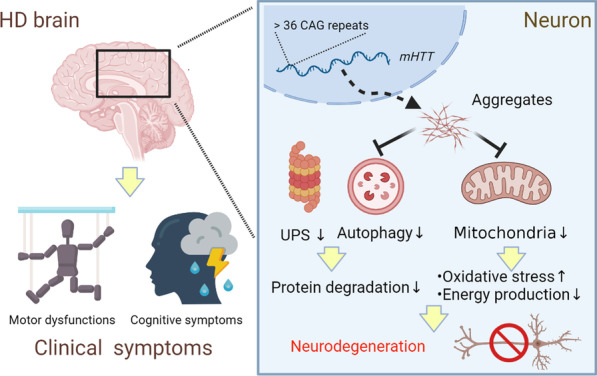
mHTT leads to cellular dysfunctions and clinical symptoms in HD patients. mHTT is transcribed from mHTT gene with more than 36 CAG repeats, and then forms aggregates to cause cellular dysfunctions, such as in protein degradation systems and mitochondria, finally leading to neurodegenerations. Due to neurodegenerations, HD patients suffer clinical symptoms, including motor and cognitive deficits
Mitochondrial dysfunctions
Mitochondrial dysfunction is a common characteristic in different neurodegenerative diseases, such as Alzheimer's disease (AD), Parkinson’s disease (PD) and Spinocerebellar Ataxia [20–22], and failure of mitochondrial functions results in increases of oxidative stress and deficits of energy production, further leading to neurodegenerations. In different HD models, several reports have shown the increase of oxidative stress and deficits of mitochondria in this disease [23–25]. These deficits include the impairments on mitochondrial biogenesis, dynamics, membrane potential, electron transport chains, adenosine triphosphate (ATP) productions, etc., and finally affect cellular functions, neuropathogenesis and clinic symptoms [24–27]. Due to the critical roles of mitochondrial functions in HD pathogenesis, several treatments targeting on this aspect have been demonstrated. For example, activation of mitochondria-related genes, such as Sirtuin 3 (SIRT3), Peroxisome proliferator-activated receptor gamma co-activator-1 alpha (PGC1alpha), Uncoupling protein (UCP), has shown to alleviate HD symptoms in different models [23, 24, 28], highly suggesting improvement of mitochondrial functions is an important therapeutical strategy for HD (Fig. 1).
Abnormal protein degradation
Protein degradation is an important step to control protein quality, and further maintains the cellular functions of proteins in different tissues. Ubiquitin-proteasome system (UPS) and autophagy, also known as macroautophagy, are two important systems responsible for protein degradations inside cells. The UPS clears up short-lived proteins by tagging a ubiquitin activating enzyme (E1), a conjugating enzyme (E2) and a ligase (E3), and further degrades these proteins in 26S proteasomes in the nucleus or cytoplasm. The autophagy system forms autophagosomes and autolysosomes, and then degrades long-lived proteins in lysosomes of cytoplasm. Based on previous studies, several different neurodegenerative diseases show poor treatments due to loss of functions in these two systems [29, 30]. In HD, mHTT has been reported to be degraded slowly, and impairment of two protein degradation systems has been found in different HD models [17, 31–34]. For example, a deficiency of a unique α-amine E2 enzyme, UBE2W, is highly correlated with the accumulation of mHTT [35], and polyQ of mHTT has been shown to block the activity of proteasomes [36], indicating the impairment of the UPS system in HD. In addition, mHTT disrupts autophagosome dynamics and vesicle trafficking, and also decreases the initiation of autophagy [37–39], indicating the impairment of the autophagy system. As a result, enhancement of these two protein degradation systems is also considered as a potential strategy of therapy in HD (Fig. 1).
Current and potential treatments
Current treatment/drugs
Till now, there is no treatment to cure HD, and the clinic treatments only could alleviate symptoms to improve the quality of life (Table 1). Chorea is one of motor symptoms in HD, and decreased neurotransmission of dopamine has shown the reduction of chorea [40], indicating decrease of dopamine signaling is a therapeutic target for HD patients. Tetrabenazine (TBZ) and Deutetrabenazine, both approved by the Food and Drug Administration (FDA), are inhibitors of the vesicular monoamine transporter type 2 (VMAT2), and functions through depletion of dopamine in the presynaptic terminals to improve chorea [40, 41]. In addition, dopamine receptor blockers, such as tiapride, haloperidol, fluphenazine, olanzapine and risperidone, are also used to treat HD to suppress chorea. Since dopamine receptor blockers belong to one type of antipsychotics, these drugs also alleviate comorbid psychiatric symptoms in HD [40, 42]. Besides to the decrease of dopamine signaling, suppression of glutamate transmission is also a strategy to reduce chorea. These drugs used in clinic include riluzole and amantadine [8, 43]. Moreover, alleviations of psychiatric and cognitive symptoms are also important to improve quality of life. For example, selective serotonin reuptake inhibitors (SSRIs) or serotonin-norepinephrine reuptake inhibitors (SNRIs) has been used to deal with depression and anxiety, and above dopamine receptor blockers have used as antipsychotics in HD [40, 43]. Since these drugs target on alleviation of symptoms only, it is important to develop treatments to cure HD.
Table 1.
Current and potential treatments for HD
| Current and potential treatments for HD | ||
|---|---|---|
| Current drugs | Working mechanisms | References |
| Tetrabenazine; Deutetrabenazine | VMAT2 inhibitors | [41, 5] |
| Tiapride; Haloperidol; Fluphenazine; Olanzapine; Risperidone | Dopamine receptor blockers | [42, 40] |
| Riluzole; Amantadine | Suppression of glutamate transmission | [8, 43] |
| Cell therapy | Cell replacement; Secretion of beneficial factors | [44, 44–46] |
| Gene therapy | Reduction of mutant HTT; Enhancement of cellular functions | [47, 47–55] |
Cell therapy
Cell-based therapy, which may provide structural and functional reconstruction in brain regions, is a potential strategy for neurodegenerative diseases (Table 1) [20, 44, 45]. In HD, these related studies have been examined in different animal models. For example, intranasal administration of mesenchymal stem cells or intrastriatal administration of dental pulp stem cells has ameliorated neuropathological and behavioral phenotypes in different HD rodent models [46], and these results highly suggest potential applications in clinic. Indeed, several worldwide clinical trials of cell transplantations in HD have been conducted, and showed the amelioration of brain structures, neural circuitry and motor functions [45]. Although some HD patients showed long-lasting improvements of their symptoms, graft failure was also observed in several patients due to different individual characteristics, immune responses, graft protocols or cell sources in different trials [45]. As a result, the guide line for cell-based therapy is still being developed.
Gene therapy
Since the HD is a monogenetic disease, mHTT-lowering strategy is considered as a potential direction for the therapy. Taking advantages of different molecular techniques, such as antisense oligonucleotides, gene deletion and RNA interference, these alterations suppress the expression of mHTT and rescue HD phenotypes in different models as well (Table 1) [47–50]. Based on the information via ClinicalTrials.gov provided by the U.S. National Library of Medicine, antisense oligonucleotides designed for mHTT are already in phase II and III of clinical trials (NCT02519036, NCT03225833, NCT03225846, etc), highly suggesting foreseeable potential of these therapies. In addition to targeting on mHTT, increases of neuroprotective effects through other functional genes, such as neurotrophic factors, growth factors, anti-oxidative stress genes and microRNAs, have also alleviated HD symptoms [25, 51–55]. These results suggest gene therapies may not only offer temporary relief, but also cure the root cause in HD.
MicroRNA
Cellular biofunctions
microRNA biogenesis
microRNA (miRNA) is a small non-coding RNA, and plays an important role in gene regulation. miRNA transcripts are generated from non-coding regions of primary miRNAs (Pri-miRNAs), and the Pri-miRNAs are processed by a nuclear ribonuclease III enzyme, Drosha, to form hairpin precursor miRNAs (Pre-miRNAs) inside nucleus. Pre-miRNAs are transported to cytoplasm by exportin proteins, such as exportin 5, and then are processed again by the Dicer to form mature miRNAs. Mature miRNAs further interact with a RNA-induced silencing complex (RISC), and bind to target mRNAs to mostly cleave mRNA or suppress translation at the post-transcriptional level [56]. Although it is well known miRNAs function through downregulating gene expression, miRNAs also have been reported to offer opposite effects to upregulate translation during different stages of cell cycle [57]. Notably, one miRNA usually has targeted on multiple genes (Winter, Jung et al. 2009), suggesting the importance to control the expressions of miRNAs under physiological conditions.
microRNAs related to mitochondrial functions
As described above, mitochondrial functions could be determined through mitochondrial biogenesis, dynamics, membrane potential and ATP productions. These mitochondrial functions are closely regulated by different miRNAs during neuronal development and disease progression. For example, miR-137 upregulates nuclear factor erythroid 2-related factor 2 (NRF2) to enhance mitochondrial biogenesis, and also further modulates mitochondrial fusion and fission to affect mitochondrial dynamics in neural stem cells [58]. In addition, miR-338 targets to cytochrome-c oxidase subunit 4I1 (COX4I1) to regulate mitochondrial ATP production in brains [59]. Furthermore, miR-204 is upregulated to target transient receptor potential mucolipin-1 (TRPML1) to damage mitochondrial membrane potential and ATP production in AD, and suppression of miR-204 rescues those mitochondrial damages and reactive oxygen species (ROS) productions through upregulation of TRPML1 [60]. These results indicate brain-enriched miRNAs, such as miR-137, miR-338 and miR-204, play critical roles to maintain mitochondrial biogenesis and biofunctions during brain development, and unbalance of these miRNA regulations leads to the progression of neuronal diseases (Fig. 2).
Fig. 2.
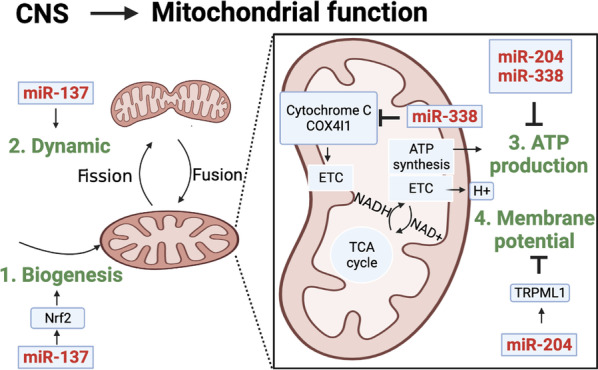
microRNAs regulate mitochondrial functions in CNS. Different miRNAs involve in mitochondrial functions, such as mitochondrial dynamic, biogenesis, ATP production and membrane potential
microRNAs related to protein degradations
Two protein degradation systems, UPS and autophagy, are regulated by several miRNAs as well. Based on previous reports, several miRNAs have shown to target different E1, E2 or E3 enzymes of UPS to affect different neuronal functions [61–63]. Similar to UPS, the autophagy system is also closely regulated by miRNAs in the neuronal system, and autophagy-related proteins, such as Sequestosome 1 (Sqstm1), Optineurin (Optn), BACE1 and ATG5, are directly controlled by miRNAs to affect autophagic activity [64, 65]. Due to dysfunctions of these protein-degradation-related miRNAs, the cells, including neurons, glia cells and microglia cells, in CNS fail to process genesis, migration, maturation or advanced interactive functions. For example, higher expression level of miR-30a-5p has shown to worsen UPS to downregulate glutamate transporter 1 (GLT-1), further leading to glutamate excitotoxicity [62]. In addition, higher expression levels of miR-331-3p and miR-9-5p have shown to target autophagy regulators, Sqstm1 and Optn, further impairing autophagic degradation activity [64]. These results all support the critical roles of miRNAs in the cellular functions of protein degradations (Fig. 3), implying a potential application to adjust these cellular functions through miRNAs to treat different neuronal diseases.
Fig. 3.
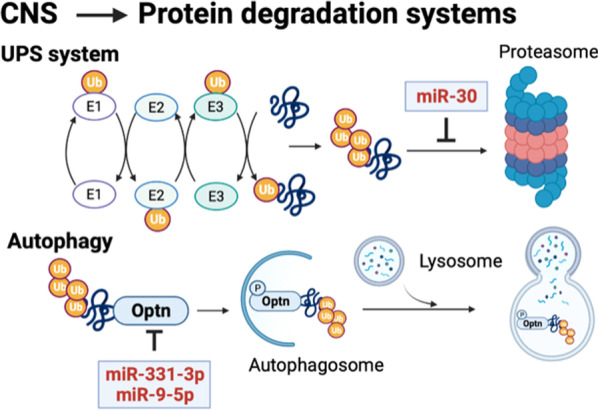
microRNAs regulate two protein degradation systems in CNS. Different miRNAs involve at different stages of different protein degradation systems, such as the UPS and autophagy systems
microRNA toward neuronal diseases
microRNAs in neurodegenerative diseases
Abnormal expression of global miRNAs or specific miRNAs has been reported to cause neurodegenerative diseases. Taking advantages of gene-modified animals, deletion of Dicer, which causes abnormal expression of global miRNAs, has led to neuronal degenerations, impairments and symptoms [66–68]. Deletion of specific miRNAs, such as miR-124-1 or miR-8, has also shown to develop neuronal degenerations and apoptosis in different models [69, 70]. These indicate loss of miRNA functions could be a potential disease-causing factor for neurodegenerative diseases; however, these is still limited information regarding to mutations of specific miRNAs lead to neurodegenerative diseases in clinic. This provides an insight that searching mutations in non-coding miRNAs could be one of potential directions for identification of disease-causing factors in the sporadic type of neurodegenerative diseases. Moreover, although miRNAs might not be main causes for neurodegenerative diseases, these diseases, including AD, PD and HD, do lead to alterations of miRNA expression profiling, which further regulates downstream cellular functions and finally leads to disease-specific pathology and symptoms [61, 62, 64, 71, 72]. Since the expression profiling of certain specific miRNAs is associated with the disease progression, neuropathology or symptoms, these miRNAs could be used as biomarkers for disease diagnoses and also could be applied for therapy potentially.
microRNAs in Huntington’s disease
The expression profiling of miRNAs in HD patients has been reported in several studies, and altered miRNAs are highly associated with the regulation of molecular or pathological phenotypes [73–75]. In addition, global miRNA alterations have also been reported in HD because of the accumulation of Argonaute-2 (AGO2) protein, which is a critical component for RISC [76]. These results show HD could lead to abnormal expression of global miRNAs or specific miRNAs as well. As a result, miRNAs are recently addressed as biomarkers to indicate the progression of HD in clinic [73, 77].
Due to limited manipulations of miRNA alterations in HD patients, miRNA studies have been widely demonstrated in different animal models. In transgenic monkey studies, HD transgenic monkeys showed downregulation of miR-128a and upregulation of miR-196a, which is similar in HD patients [61, 78, 79]. In transgenic mouse studies, miR-9/9*, miR-124 and miR-132 have been shown to be suppressed in both human HD patients and mouse models [75, 79, 80]. Since HD patients and animals display certain similar miRNA profiling, these animal models have been used to investigate that miRNAs as potential therapeutical targets in HD. Indeed, the treatments of miR-132 in R6/2 HD transgenic mice show improvements of behavioral symptoms and delay of disease progression [81]. In addition, miR-196a also improves molecular, neuropathological and behavioral phenotypes in HD transgenic mice through enhancements of neuronal cytoskeletons [52, 61]. Most interestingly, several studies have addressed on artificial miRNAs targeting on human HTT [50, 82–84]. Although these miRNAs do not selectively target on mutant HTT and do reduce the expression of normal HTT, these miRNAs offer functional improvement, such as decrease of aggregates and alleviation of HD symptoms, and also do not lead to severe side-effects. Indeed, this strategy, as known as AMT-130, of non-selectively lowering HTT using artificial miRNAs has been undergone in a clinical phase 1/2 trial (NCT04120493) in 2021[85], and aims to examine the safety and efficacy of this miRNA therapy in early stage of HD patients. Due to the similar approach examined in different animal models already, physicians are looking forward to seeing positive results. Overall, since these in vivo data show strong evidence to alleviate HD progression in animal models, it highly raises this potential therapy using miRNA-based strategies in HD.
Potential treatments using miRNAs
Different types of miRNA mimics or inhibitors
miRNAs have been considered as a pharmacologic strategy against diseases, and different types of miRNA mimics or inhibitors altering the expression of miRNAs have been developed. Regarding to miRNA mimics, mature form of miRNAs is commonly synthesized to deliver into cells or tissues. However, the half-life of these raw mimics is usually less than 30 min due to degradation by RNases, resulting in limited effects of functional miRNAs [86]. As a result, modified miRNAs have been developed to prevent RNase attacks. For example, phosphorothioate (PS) RNA is generated through using sulfur in the phosphodiester bonds of the RNAs in order to decrease the attacks of RNases, and modifications in 2’-OH group of RNAs to form 2'-O-methyl (2'-O-Me) RNAs are also used to increase the miRNA stability, further enhancing the effects of miRNA mimics [86–88]. Regarding to miRNA inhibitors, antisense RNAs are broadly used to block miRNAs; however, quick degradation of antisense RNAs is observed as well, which is similar to that of miRNA mimics. Therefore, PS and 2’-OMe RNA techniques are also broadly used as miRNA antisense inhibitors to block the functions of miRNAs [86, 88]. Even more, new modified mesyl phosphoramidate RNA targeting to miRNAs has shown more efficient to block miRNA functions [89]. These results suggest the increase of miRNA stability to prolong half-life is a critical step to apply miRNA therapy, and synthetic biology would also play an important role for this application.
Delivery systems
Brains with blood–brain barrier (BBB) result in obstacles for drug delivery into cells; as a result, a suitable delivery system is necessary to transport miRNAs into target brain cells. Nanoparticle is a powerful platform able to penetrate BBB to deliver RNA-based treatments into brain regions, and has been developed for more than 10 years. Several types of nanoparticles carrying miRNAs have been examined in different models of neuronal diseases in vivo, including quercetin-conjugated superparamagnetic iron oxide nanoparticles in AD [90], traceable polymeric nanoparticles in PD [91], poly(lactic-co-glycolic acid) nanoparticles in stroke [92], etc. These materials could not only show high capacity and low cytotoxicity to deliver miRNAs into brain regions, but also harbor other biological purposes, such as antioxidant effects and distribution monitoring, showing that nanoparticles with dual or more functions are a trend to develop this type of delivery systems.
In addition to nanoparticles, delivering miRNAs by viral systems has also been applied in several neuronal diseases. Especially, adeno-associated viruses (AAVs) are most often used because higher titer of AAVs is relatively easier generated, higher levels of miRNAs could be produced in AAVs, AAVs do not easily integrate into genomes and AAVs are relatively safer than other viral systems. Therefore, AAVs, such as Luxturna, has been approved by FDA to apply in clinical trials [93]. In animal studies, AAVs expressing miRNAs have been broadly demonstrated to target neuronal diseases. For example, AAVs carrying miR-23a have improved neuropathology and survival rate in spinal muscular atrophy (SMA) mice in vivo [94]. AAV-miRNAs targeting on spinocerebellar ataxia type 3, another polyglutamine disease, have shown to reduce the expression of disease-causing gene in different models [95]. In HD, AAV-miRNAs against mutant HTT have also been examined in different in vivo models, and shown different beneficial effects on molecular or neuropathological phenotypes [96–98]. Due to approval of clinical trials previously, these animal studies highly bridge the potential applications of AAVs carrying miRNAs to target neuronal diseases in clinic.
Exosomes, which are small extracellular vesicles with a diameter of less than 100 nm, are an emerging carrier to deliver miRNAs into brains recently because of lipid bilayer structures. Exosomes carry proteins, mRNAs, miRNAs, etc., and deliver these materials to specific target cells to further control cellular signals or gene regulations [86, 99]. Since exosomes are endogenous components inside cells, higher stability, lower immunogenicity and lower cytotoxicity are advantages to apply to deliver miRNAs in vivo. As a result, the exosome-based delivery systems carrying miRNAs have been used to protect brains from injury in several neuronal diseases, such as brain glioblastoma, ischemia or hemorrhage [100–103]. In HD mouse models, this exosome system has been applied to deliver miR-124 to suppress the target gene, RE1-silencing transcription factor (REST) which represses neuronal genes, in R6/2 HD transgenic mice; however, this treatment does not improve HD behaviors [104]. Due to rapid progression of exosome-related studies, optimization of this strategy is anticipated to apply to deliver miRNAs efficiently and safely in vivo.
Limitations and challenges
Although miRNAs show several advantages for potential applications of therapy in neuronal diseases, there are still limitations and challenges (Fig. 4). The major limitation of miRNA applications is one miRNA usually has multiple targets. As a result, treatments of miRNAs may encounter unexpected side-effects due to multiple bindings in different direct targets. Therefore, safety or toxicity studies should be evaluated before clinical trials. Indeed, certain miRNA treatments for HD have been examined. For example, the side-effects of miRNAs targeting on HTT have been demonstrated in mice, rats, minipigs and non-human primates, and results show the tolerability in these in vivo models [50, 83, 84]. However, this characteristic may oppositely be considered as an advantage because miRNAs might also target multiple pathways to regulate downstream phenotypes. For example, miR-196a has bound to different target genes to decrease aggregates of mutant HTT, improve mitochondrial dynamic, enhance neurite outgrowth, etc., to alleviate the progression of HD in different models [52, 61, 105, 106]. This indicates miRNAs may function through different regulatory pathways to further provide synergistic effects.
Fig. 4.

miRNA-based therapy may encounter potential challanges in HD. miRNA-based therapy needs to overcome the miRNA instability, suitable delivery systems and functional effects inside brains. Nucleotide modifications, such as 2'-O-methyl- and phosphorothioate- structure, may increase the stability of miRNAs. Delivery systems, such as viruses, exosomes and nanoparticles, may faciliate the efficiency and safety to transport miRNAs into brains. Most importantly, several potential challenges which may be encountered inside brains are listed
In addition, the miRNAs are relatively unstable, and the half-life of miRNAs is shorter as described in “Different types of miRNA mimics or inhibitors”. Although modified miRNAs have efficiently extended the half-life, long-term expression of these miRNAs is desired if miRNAs are used as a therapeutical strategy to cure genetic diseases. The viral system, such as AAV- or lentivirus-based system, is one of options to achieve long-term expression. The AAV delivery system is able to continuously express exogenous transgenes for more than one year without integrating into host genomes [107], and the lentivirus-based system even constitutively expresses transgenes due to integrating into genomes [108]. However, the random integration of lentiviruses into genomes may disrupt the endogenous gene expressions [109], which raises another drawback of the viral system.
Moreover, cell-, tissue- or region-specific expression of miRNAs is also a challenge for treatments of miRNAs in CNS. Although miRNAs could be stereotaxically injected into regions of interest, the invasive penetrations are not always allowed in CNS. To overcome this obstacle, cell- or tissue-specific promoters could be manipulated to achieve this purpose if the viral delivery systems are used to transport miRNAs. Besides, targeted exosomes, which could transport siRNAs to specific tissues via targeting peptides, are an option to reach this goal as well [110]. Additionally, specific coating of antibodies, ligands, etc., on nanoparticles to carry miRNAs is also a potential strategy to deliver to specific cells/regions [111, 112]. However, these platforms are still needed to be optimized to increase specificity, efficacy and safety as applied in clinic.
Conclusion
HD is a devastating neurodegenerative disease, and leads to unimaginable burdens to patients, families and societies. Since HD displays several pathological phenotypes, such as protein misfolding, mitochondrial dysfunctions and abnormal protein degradation, it raises a potential therapy for HD if a treatment could target multiple pathological phenotypes simultaneously. miRNAs have multiple target genes, and may function through several pathways to influence different phenotypes simultaneously. With the development of modified miRNAs and delivery systems, miRNAs raise a potential direction for therapy to treat HD; however, this idea is still at the proof-of-concept stage. In future, the optimization of miRNA structures and delivery systems would be a critical step to apply to the clinic. In addition, overcoming drawbacks and limitations would also be an important step to apply miRNA therapy.
Acknowledgements
All figures are created with BioRender.com.
Abbreviations
- HD
Huntington’s disease
- miRNA
MicroRNA
- HTT
Huntingtin
- mHTT
Mutant HTT
- ploy Q
Polyglutamine
- CNS
Central nervous system
- AD
Alzheimer's disease
- PD
Parkinson’s disease
- ATP
Adenosine triphosphate
- SIRT3
Sirtuin 3
- PGC1alpha
Peroxisome proliferator-activated receptor gamma co-activator-1 alpha
- UCP
Uncoupling protein
- UPS
Ubiquitin–proteasome system
- TBZ
Tetrabenazine
- FDA
Food and Drug Administration
- VMAT2
Vesicular monoamine transporter type 2
- SSRIs
Selective serotonin reuptake inhibitors
- SNRIs
Serotonin-norepinephrine reuptake inhibitors
- Pri-miRNAs
Primary miRNAs
- Pre-miRNAs
Precursor miRNAs
- RISC
RNA-induced silencing complex
- NRF2
Nuclear factor erythroid 2-related factor 2
- COX4I1
Cytochrome-c oxidase subunit 4I1
- TRPML1
Transient receptor potential mucolipin-1
- ROS
Reactive oxygen species
- Sqstm1
Sequestosome 1
- Optn
Optineurin
- GLT-1
Glutamate transporter 1
- AGO2
Argonaute-2
- PS
Phosphorothioate
- 2'-OMe
2'-O-methyl
- BBB
Blood–brain barrier
- AAV
Adeno-associated virus
- SMA
Spinal muscular atrophy
- REST
RE1-silencing transcription factor
Authors’ contributions
CWT, PYH, SCC, PHC and SHY collected and organized references. SHY drafted the paper. All authors read and approved the final manuscript.
Funding
This work was supported by the Ministry of Science and Technology (108-2314-B-006 -079 -MY3).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
None declared.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Chih-Wei Tung, Pin-Yu Huang and SiewChin Chan contributed equally to this paper
References
- 1.Yohrling G, Raimundo K, Crowell V, Lovecky D, Vetter L, Seeberger L. Prevalence of Huntington’s Disease in the US (954) Neurology. 2020;94(15 Supplement):954. [Google Scholar]
- 2.Group THsDCR. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. The Huntington's Disease Collaborative Research Group. Cell. 1993;72(6):971–983. doi: 10.1016/0092-8674(93)90585-E. [DOI] [PubMed] [Google Scholar]
- 3.Yang SH, Chan AW. Transgenic animal models of Huntington's disease. Curr Top Behav Neurosci. 2011;7:61–85. doi: 10.1007/7854_2010_105. [DOI] [PubMed] [Google Scholar]
- 4.Oosterloo M, Bijlsma EK, van Kuijk SM, Minkels F, de Die-Smulders CE. Clinical and genetic characteristics of late-onset Huntington's disease. Parkinsonism Relat Disord. 2019;61:101–105. doi: 10.1016/j.parkreldis.2018.11.009. [DOI] [PubMed] [Google Scholar]
- 5.Solberg OK, Filkuková P, Frich JC, Feragen KJB. Age at death and causes of death in patients with Huntington disease in Norway in 1986–2015. J Huntingtons Dis. 2018;7(1):77–86. doi: 10.3233/JHD-170270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gutekunst CA, Li SH, Yi H, Mulroy JS, Kuemmerle S, Jones R, Rye D, Ferrante RJ, Hersch SM, Li XJ. Nuclear and neuropil aggregates in Huntington's disease: relationship to neuropathology. J Neurosci. 1999;19(7):2522–2534. doi: 10.1523/JNEUROSCI.19-07-02522.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Portera-Cailliau C, Hedreen JC, Price DL, Koliatsos VE. Evidence for apoptotic cell death in Huntington disease and excitotoxic animal models. J Neurosci. 1995;15(5 Pt 2):3775–3787. doi: 10.1523/JNEUROSCI.15-05-03775.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.André VM, Cepeda C, Levine MS. Dopamine and glutamate in Huntington's disease: a balancing act. CNS Neurosci Ther. 2010;16(3):163–178. doi: 10.1111/j.1755-5949.2010.00134.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Li SH, Li XJ. Huntingtin and its role in neuronal degeneration. Neuroscientist. 2004;10(5):467–475. doi: 10.1177/1073858404266777. [DOI] [PubMed] [Google Scholar]
- 10.Glidden AM, Luebbe EA, Elson MJ, Goldenthal SB, Snyder CW, Zizzi CE, Dorsey ER, Heatwole CR. Patient-reported impact of symptoms in Huntington disease: PRISM-HD. Neurology. 2020;94(19):e2045–e2053. doi: 10.1212/WNL.0000000000008906. [DOI] [PubMed] [Google Scholar]
- 11.Lang C, Gries C, Lindenberg KS, Lewerenz J, Uhl S, Olsson C, Samzelius J, Landwehrmeyer GB. Monitoring the motor phenotype in Huntington's disease by analysis of keyboard typing during real life computer use. J Huntingtons Dis. 2021;10(2):259–268. doi: 10.3233/JHD-200451. [DOI] [PubMed] [Google Scholar]
- 12.DeGuire SM, Ruggeri FS, Fares MB, Chiki A, Cendrowska U, Dietler G, Lashuel HA. N-terminal Huntingtin (Htt) phosphorylation is a molecular switch regulating Htt aggregation, helical conformation, internalization, and nuclear targeting. J Biol Chem. 2018;293(48):18540–18558. doi: 10.1074/jbc.RA118.004621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Skotte NH, Pouladi MA, Ehrnhoefer DE, Huynh K, Qiu X, Nielsen SMB, Nielsen TT, Norremolle A, Hayden MR. Compromised IGF signaling causes caspase-6 activation in Huntington disease. Exp Neurol. 2020;332:113396. doi: 10.1016/j.expneurol.2020.113396. [DOI] [PubMed] [Google Scholar]
- 14.Yang H, Yang S, Jing L, Huang L, Chen L, Zhao X, Yang W, Pan Y, Yin P, Qin ZS, Tang B, Li S, Li XJ. Truncation of mutant huntingtin in knock-in mice demonstrates exon1 huntingtin is a key pathogenic form. Nat Commun. 2020;11(1):2582. doi: 10.1038/s41467-020-16318-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Priya SB, Gromiha MM. Structural insights into the aggregation mechanism of huntingtin exon 1 protein fragment with different polyQ-lengths. J Cell Biochem. 2019;120(6):10519–10529. doi: 10.1002/jcb.28338. [DOI] [PubMed] [Google Scholar]
- 16.Cheng PH, Li CL, Her LS, Chang YF, Chan AW, Chen CM, Yang SH. Significantly differential diffusion of neuropathological aggregates in the brain of transgenic mice carrying N-terminal mutant huntingtin fused with green fluorescent protein. Brain Struct Funct. 2013;218(1):283–294. doi: 10.1007/s00429-012-0401-x. [DOI] [PubMed] [Google Scholar]
- 17.Her LS, Lin JY, Fu MH, Chang YF, Li CL, Tang TY, Jhang YL, Chang CY, Shih MC, Cheng PH, Yang SH. The differential profiling of ubiquitin-proteasome and autophagy systems in different tissues before the onset of Huntington's disease models. Brain Pathol. 2015;25(4):481–490. doi: 10.1111/bpa.12191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kocerha J, Liu Y, Willoughby D, Chidamparam K, Benito J, Nelson K, Xu Y, Chi T, Engelhardt H, Moran S, Yang SH, Li SH, Li XJ, Larkin K, Neumann A, Banta H, Yang JJ, Chan AW. Longitudinal transcriptomic dysregulation in the peripheral blood of transgenic Huntington's disease monkeys. BMC Neurosci. 2013;14:88. doi: 10.1186/1471-2202-14-88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li XJ, Li H, Li S. Clearance of mutant huntingtin. Autophagy. 2010;6(5):663–664. doi: 10.4161/auto.6.5.12336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fu MH, Li CL, Lin HL, Chen PC, Calkins MJ, Chang YF, Cheng PH, Yang SH. Stem cell transplantation therapy in Parkinson's disease. Springerplus. 2015;4:597. doi: 10.1186/s40064-015-1400-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hsu JY, Jhang YL, Cheng PH, Chang YF, Mao SH, Yang HI, Lin CW, Chen CM, Yang SH. The truncated C-terminal fragment of mutant ATXN3 disrupts mitochondria dynamics in spinocerebellar ataxia type 3 models. Front Mol Neurosci. 2017;10:196. doi: 10.3389/fnmol.2017.00196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Medala VK, Gollapelli B, Dewanjee S, Ogunmokun G, Kandimalla R, Vallamkondu J. Mitochondrial dysfunction, mitophagy, and role of dynamin-related protein 1 in Alzheimer's disease. J Neurosci Res. 2021;99(4):1120–1135. doi: 10.1002/jnr.24781. [DOI] [PubMed] [Google Scholar]
- 23.Jin YN, Johnson GV. The interrelationship between mitochondrial dysfunction and transcriptional dysregulation in Huntington disease. J Bioenerg Biomembr. 2010;42(3):199–205. doi: 10.1007/s10863-010-9286-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Naia L, Carmo C, Campesan S, Fão L, Cotton VE, Valero J, Lopes C, Rosenstock TR, Giorgini F, Rego AC. Mitochondrial SIRT3 confers neuroprotection in Huntington's disease by regulation of oxidative challenges and mitochondrial dynamics. Free Radic Biol Med. 2021;163:163–179. doi: 10.1016/j.freeradbiomed.2020.11.031. [DOI] [PubMed] [Google Scholar]
- 25.Yusuf IO, Chen HM, Cheng PH, Chang CY, Tsai SJ, Chuang JI, Wu CC, Huang BM, Sun HS, Yang SH. Fibroblast growth factor 9 activates anti-oxidative functions of Nrf2 through ERK signalling in striatal cell models of Huntington's disease. Free Radic Biol Med. 2019;130:256–266. doi: 10.1016/j.freeradbiomed.2018.10.455. [DOI] [PubMed] [Google Scholar]
- 26.Cherubini M, Lopez-Molina L, Gines S. Mitochondrial fission in Huntington's disease mouse striatum disrupts ER-mitochondria contacts leading to disturbances in Ca(2+) efflux and Reactive Oxygen Species (ROS) homeostasis. Neurobiol Dis. 2020;136:104741. doi: 10.1016/j.nbd.2020.104741. [DOI] [PubMed] [Google Scholar]
- 27.Dubinsky JM. Towards an understanding of energy impairment in Huntington's disease brain. J Huntingtons Dis. 2017;6(4):267–302. doi: 10.3233/JHD-170264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jamwal S, Blackburn JK, Elsworth JD. PPARgamma/PGC1alpha signaling as a potential therapeutic target for mitochondrial biogenesis in neurodegenerative disorders. Pharmacol Therapy. 2020;219:107705. doi: 10.1016/j.pharmthera.2020.107705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Boland B, Yu WH, Corti O, Mollereau B, Henriques A, Bezard E, Pastores GM, Rubinsztein DC, Nixon RA, Duchen MR, Mallucci GR, Kroemer G, Levine B, Eskelinen EL, Mochel F, Spedding M, Louis C, Martin OR, Millan MJ. Promoting the clearance of neurotoxic proteins in neurodegenerative disorders of ageing. Nat Rev Drug Discov. 2018;17(9):660–688. doi: 10.1038/nrd.2018.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Harding RJ, Tong YF. Proteostasis in Huntington's disease: disease mechanisms and therapeutic opportunities. Acta Pharmacol Sin. 2018;39(5):754–769. doi: 10.1038/aps.2018.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bennett EJ, Shaler TA, Woodman B, Ryu KY, Zaitseva TS, Becker CH, Bates GP, Schulman H, Kopito RR. Global changes to the ubiquitin system in Huntington's disease. Nature. 2007;448(7154):704–708. doi: 10.1038/nature06022. [DOI] [PubMed] [Google Scholar]
- 32.Huang ZN, Chen JM, Huang LC, Fang YH, Her LS. Inhibition of p38 mitogen-activated protein kinase ameliorates HAP40 depletion-induced toxicity and proteasomal defect in Huntington's disease model. Mol Neurobiol. 2021;58(6):2704–2723. doi: 10.1007/s12035-020-02280-y. [DOI] [PubMed] [Google Scholar]
- 33.Huang ZN, Her LS. The ubiquitin receptor ADRM1 modulates HAP40-induced proteasome activity. Mol Neurobiol. 2017;54(9):7382–7400. doi: 10.1007/s12035-016-0247-y. [DOI] [PubMed] [Google Scholar]
- 34.Zhao T, Hong Y, Li S, Li XJ. Compartment-dependent degradation of mutant huntingtin accounts for its preferential accumulation in neuronal processes. J Neurosci. 2016;36(32):8317–8328. doi: 10.1523/JNEUROSCI.0806-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang B, Zeng L, Merillat SA, Fischer S, Ochaba J, Thompson LM, Barmada SJ, Scaglione KM, Paulson HL. The ubiquitin conjugating enzyme Ube2W regulates solubility of the Huntington's disease protein, huntingtin. Neurobiol Dis. 2018;109(Pt A):127–136. doi: 10.1016/j.nbd.2017.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Holmberg CI, Staniszewski KE, Mensah KN, Matouschek A, Morimoto RI. Inefficient degradation of truncated polyglutamine proteins by the proteasome. Embo J. 2004;23(21):4307–4318. doi: 10.1038/sj.emboj.7600426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Brattås PL, Hersbach BA, Madsen S, Petri R, Jakobsson J, Pircs K. Impact of differential and time-dependent autophagy activation on therapeutic efficacy in a model of Huntington disease. Autophagy. 2021;17(6):1316–1329. doi: 10.1080/15548627.2020.1760014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.del Toro D, Alberch J, Lázaro-Diéguez F, Martín-Ibáñez R, Xifró X, Egea G, Canals JM. Mutant huntingtin impairs post-Golgi trafficking to lysosomes by delocalizing optineurin/Rab8 complex from the Golgi apparatus. Mol Biol Cell. 2009;20(5):1478–1492. doi: 10.1091/mbc.e08-07-0726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wong YC, Holzbaur EL. The regulation of autophagosome dynamics by huntingtin and HAP1 is disrupted by expression of mutant huntingtin, leading to defective cargo degradation. J Neurosci. 2014;34(4):1293–1305. doi: 10.1523/JNEUROSCI.1870-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Stahl CM, Feigin A. Medical, surgical, and genetic treatment of Huntington disease. Neurol Clin. 2020;38(2):367–378. doi: 10.1016/j.ncl.2020.01.010. [DOI] [PubMed] [Google Scholar]
- 41.Claassen DO, Philbin M, Carroll B. Deutetrabenazine for tardive dyskinesia and chorea associated with Huntington's disease: a review of clinical trial data. Expert Opin Pharmacother. 2019;20(18):2209–2221. doi: 10.1080/14656566.2019.1674281. [DOI] [PubMed] [Google Scholar]
- 42.McColgan P, Tabrizi SJ. Huntington's disease: a clinical review. Eur J Neurol. 2018;25(1):24–34. doi: 10.1111/ene.13413. [DOI] [PubMed] [Google Scholar]
- 43.Coppen EM, Roos RA. Current pharmacological approaches to reduce chorea in Huntington's disease. Drugs. 2017;77(1):29–46. doi: 10.1007/s40265-016-0670-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ahmadian-Moghadam H, Sadat-Shirazi MS, Zarrindast MR. Therapeutic potential of stem cells for treatment of neurodegenerative diseases. Biotechnol Lett. 2020;42(7):1073–1101. doi: 10.1007/s10529-020-02886-1. [DOI] [PubMed] [Google Scholar]
- 45.Bachoud-Levi AC, Massart R, Rosser A. Cell therapy in Huntington's disease: taking stock of past studies to move the field forward. Stem Cells. 2021;39(2):144–155. doi: 10.1002/stem.3300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yu-Taeger L, Stricker-Shaver J, Arnold K, Bambynek-Dziuk P, Novati A, Singer E, Lourhmati A, Fabian C, Magg J, Riess O, Schwab M, Stolzing A, Danielyan L, Nguyen HHP. Intranasal administration of mesenchymal stem cells ameliorates the abnormal dopamine transmission system and inflammatory reaction in the R6/2 mouse model of huntington disease. Cells. 2019;8(6):595. doi: 10.3390/cells8060595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Cheong RY, Baldo B, Sajjad MU, Kirik D, Petersén Å. Effects of mutant huntingtin inactivation on Huntington disease-related behaviours in the BACHD mouse model. Neuropathol Appl Neurobiol. 2020;47(4):564–578. doi: 10.1111/nan.12682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dabrowska M, Olejniczak M. Gene therapy for Huntington's disease using targeted endonucleases. Methods Mol Biol. 2020;2056:269–284. doi: 10.1007/978-1-4939-9784-8_17. [DOI] [PubMed] [Google Scholar]
- 49.Leavitt BR, Tabrizi SJ. Antisense oligonucleotides for neurodegeneration. Science. 2020;367(6485):1428–1429. doi: 10.1126/science.aba4624. [DOI] [PubMed] [Google Scholar]
- 50.Spronck EA, Vallès A, Lampen MH, Montenegro-Miranda PS, Keskin S, Heijink L, Evers MM, Petry H, Deventer SJV, Konstantinova P, Haan M. Intrastriatal administration of AAV5-miHTT in non-human primates and rats is well tolerated and results in miHTT transgene expression in key areas of Huntington disease pathology. Brain Sci. 2021;11(2):129. doi: 10.3390/brainsci11020129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Connor B, Sun Y, von Hieber D, Tang SK, Jones KS, Maucksch C. AAV1/2-mediated BDNF gene therapy in a transgenic rat model of Huntington's disease. Gene Ther. 2016;23(3):283–295. doi: 10.1038/gt.2015.113. [DOI] [PubMed] [Google Scholar]
- 52.Her LS, Mao SH, Chang CY, Cheng PH, Chang YF, Yang HI, Chen CM, Yang SH. miR-196a enhances neuronal morphology through suppressing RANBP10 to Provide neuroprotection in Huntington's disease. Theranostics. 2017;7(9):2452–2462. doi: 10.7150/thno.18813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yusuf IO, Chen HM, Cheng PH, Chang CY, Tsai SJ, Chuang JI, Wu CC, Huang BM, Sun HS, Chen CM, Yang SH. FGF9 induces neurite outgrowth upon ERK signaling in knock-in striatal Huntington's disease cells. Life Sci. 2021;267:118952. doi: 10.1016/j.lfs.2020.118952. [DOI] [PubMed] [Google Scholar]
- 54.Yusuf IO, Chen HM, Cheng PH, Chang CY, Tsai SJ, Chuang JI, Wu CC, Huang BM, Sun HS, Chen CM, Yang SH. Fibroblast growth factor 9 stimulates neuronal length through NF-kB signaling in striatal cell Huntington's disease models. Mol Neurobiol. 2021;58(5):2396–2406. doi: 10.1007/s12035-020-02220-w. [DOI] [PubMed] [Google Scholar]
- 55.Yusuf IO, Cheng PH, Chen HM, Chang YF, Chang CY, Yang HI, Lin CW, Tsai SJ, Chuang JI, Wu CC, Huang BM, Sun HS, Yang SH. Fibroblast growth factor 9 suppresses striatal cell death dominantly through ERK signaling in Huntington's disease. Cell Physiol Biochem. 2018;48(2):605–617. doi: 10.1159/000491889. [DOI] [PubMed] [Google Scholar]
- 56.Winter J, Jung S, Keller S, Gregory RI, Diederichs S. Many roads to maturity: microRNA biogenesis pathways and their regulation. Nat Cell Biol. 2009;11(3):228–234. doi: 10.1038/ncb0309-228. [DOI] [PubMed] [Google Scholar]
- 57.Vasudevan S, Tong Y, Steitz JA. Switching from repression to activation: microRNAs can up-regulate translation. Science. 2007;318(5858):1931–1934. doi: 10.1126/science.1149460. [DOI] [PubMed] [Google Scholar]
- 58.Channakkar AS, Singh T, Pattnaik B, Gupta K, Seth P, Adlakha YK. MiRNA-137-mediated modulation of mitochondrial dynamics regulates human neural stem cell fate. Stem Cells. 2020;38(5):683–697. doi: 10.1002/stem.3155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Li L, Voloboueva L, Griffiths BB, Xu L, Giffard RG, Stary CM. MicroRNA-338 inhibition protects against focal cerebral ischemia and preserves mitochondrial function in vitro in astrocytes and neurons via COX4I1. Mitochondrion. 2021;59:105–112. doi: 10.1016/j.mito.2021.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhang L, Fang Y, Zhao X, Zheng Y, Ma Y, Li S, Huang Z, Li L. miR-204 silencing reduces mitochondrial autophagy and ROS production in a murine AD model via the TRPML1-activated STAT3 pathway. Mol Therapy Nucleic Acids. 2021;24:822–831. doi: 10.1016/j.omtn.2021.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Cheng PH, Li CL, Chang YF, Tsai SJ, Lai YY, Chan AW, Chen CM, Yang SH. miR-196a ameliorates phenotypes of Huntington disease in cell, transgenic mouse, and induced pluripotent stem cell models. Am J Hum Genet. 2013;93(2):306–312. doi: 10.1016/j.ajhg.2013.05.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Meng X, Zhong J, Zeng C, Yung KKL, Zhang X, Wu X, Qu S. MiR-30a-5p regulates GLT-1 function via a PKCα-mediated ubiquitin degradation pathway in a mouse model of Parkinson's disease. ACS Chem Neurosci. 2021;12(9):1578–1592. doi: 10.1021/acschemneuro.1c00076. [DOI] [PubMed] [Google Scholar]
- 63.Smrt RD, Szulwach KE, Pfeiffer RL, Li X, Guo W, Pathania M, Teng ZQ, Luo Y, Peng J, Bordey A, Jin P, Zhao X. MicroRNA miR-137 regulates neuronal maturation by targeting ubiquitin ligase mind bomb-1. Stem Cells. 2010;28(6):1060–1070. doi: 10.1002/stem.431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Chen ML, Hong CG, Yue T, Li HM, Duan R, Hu WB, Cao J, Wang ZX, Chen CY, Hu XK, Wu B, Liu HM, Tan YJ, Liu JH, Luo ZW, Zhang Y, Rao SS, Luo MJ, Yin H, Wang YY, Xia K, Tang SY, Xie H, Liu ZZ. Inhibition of miR-331-3p and miR-9-5p ameliorates Alzheimer's disease by enhancing autophagy. Theranostics. 2021;11(5):2395–2409. doi: 10.7150/thno.47408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhou Y, Ge Y, Liu Q, Li YX, Chao X, Guan JJ, Diwu YC, Zhang Q. LncRNA BACE1-AS promotes autophagy-mediated neuronal damage through the miR-214-3p/ATG5 signalling axis in Alzheimer's disease. Neuroscience. 2021;455:52–64. doi: 10.1016/j.neuroscience.2020.10.028. [DOI] [PubMed] [Google Scholar]
- 66.Hébert SS, Papadopoulou AS, Smith P, Galas MC, Planel E, Silahtaroglu AN, Sergeant N, Buée L, De Strooper B. Genetic ablation of Dicer in adult forebrain neurons results in abnormal tau hyperphosphorylation and neurodegeneration. Hum Mol Genet. 2010;19(20):3959–3969. doi: 10.1093/hmg/ddq311. [DOI] [PubMed] [Google Scholar]
- 67.Li T, Wang J, Wang H, Yang Y, Wang S, Huang N, Wang F, Gao X, Niu J, Li Z, Mei F, Xiao L. The deletion of dicer in mature myelinating glial cells causes progressive axonal degeneration but not overt demyelination in adult mice. Glia. 2018;66(9):1960–1971. doi: 10.1002/glia.23450. [DOI] [PubMed] [Google Scholar]
- 68.Tao J, Wu H, Lin Q, Wei W, Lu XH, Cantle JP, Ao Y, Olsen RW, Yang XW, Mody I, Sofroniew MV, Sun YE. Deletion of astroglial Dicer causes non-cell-autonomous neuronal dysfunction and degeneration. J Neurosci. 2011;31(22):8306–8319. doi: 10.1523/JNEUROSCI.0567-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Karres JS, Hilgers V, Carrera I, Treisman J, Cohen SM. The conserved microRNA miR-8 tunes atrophin levels to prevent neurodegeneration in Drosophila. Cell. 2007;131(1):136–145. doi: 10.1016/j.cell.2007.09.020. [DOI] [PubMed] [Google Scholar]
- 70.Sanuki R, Onishi A, Koike C, Muramatsu R, Watanabe S, Muranishi Y, Irie S, Uneo S, Koyasu T, Matsui R, Chérasse Y, Urade Y, Watanabe D, Kondo M, Yamashita T, Furukawa T. miR-124a is required for hippocampal axogenesis and retinal cone survival through Lhx2 suppression. Nat Neurosci. 2011;14(9):1125–1134. doi: 10.1038/nn.2897. [DOI] [PubMed] [Google Scholar]
- 71.Wu X, Meng X, Tan F, Jiao Z, Zhang X, Tong H, He X, Luo X, Xu P, Qu S. Regulatory mechanism of miR-543-3p on GLT-1 in a mouse model of Parkinson's disease. ACS Chem Neurosci. 2019;10(3):1791–1800. doi: 10.1021/acschemneuro.8b00683. [DOI] [PubMed] [Google Scholar]
- 72.Wu YY, Kuo HC. Functional roles and networks of non-coding RNAs in the pathogenesis of neurodegenerative diseases. J Biomed Sci. 2020;27(1):49. doi: 10.1186/s12929-020-00636-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Chang KH, Wu YR, Chen CM. Down-regulation of miR-9* in the peripheral leukocytes of Huntington's disease patients. Orphanet J Rare Dis. 2017;12(1):185. doi: 10.1186/s13023-017-0742-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hoss AG, Lagomarsino VN, Frank S, Hadzi TC, Myers RH, Latourelle JC. Study of plasma-derived miRNAs mimic differences in Huntington's disease brain. Mov Disord. 2015;30(14):1961–1964. doi: 10.1002/mds.26457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Packer AN, Xing Y, Harper SQ, Jones L, Davidson BL. The bifunctional microRNA miR-9/miR-9* regulates REST and CoREST and is downregulated in Huntington's disease. J Neurosci. 2008;28(53):14341–14346. doi: 10.1523/JNEUROSCI.2390-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Pircs K, Petri R, Madsen S, Brattås PL, Vuono R, Ottosson DR, St-Amour I, Hersbach BA, Matusiak-Brückner M, Lundh SH, Petersén Å, Déglon N, Hébert SS, Parmar M, Barker RA, Jakobsson J. Huntingtin aggregation impairs autophagy, leading to argonaute-2 accumulation and global microRNA dysregulation. Cell Rep. 2018;24(6):1397–1406. doi: 10.1016/j.celrep.2018.07.017. [DOI] [PubMed] [Google Scholar]
- 77.Reed ER, Latourelle JC, Bockholt JH, Bregu J, Smock J, Paulsen JS, Myers RH. MicroRNAs in CSF as prodromal biomarkers for Huntington disease in the PREDICT-HD study. Neurology. 2018;90(4):e264–e272. doi: 10.1212/WNL.0000000000004844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kocerha J, Xu Y, Prucha MS, Zhao D, Chan AW. microRNA-128a dysregulation in transgenic Huntington's disease monkeys. Mol Brain. 2014;7:46. doi: 10.1186/1756-6606-7-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Martinez B, Peplow PV. Altered microRNA expression in animal models of Huntington's disease and potential therapeutic strategies. Neural Regen Res. 2021;16(11):2159–2169. doi: 10.4103/1673-5374.310673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Johnson R, Buckley NJ. Gene dysregulation in Huntington's disease: REST, microRNAs and beyond. Neuromol Med. 2009;11(3):183–199. doi: 10.1007/s12017-009-8063-4. [DOI] [PubMed] [Google Scholar]
- 81.Fukuoka M, Takahashi M, Fujita H, Chiyo T, Popiel HA, Watanabe S, Furuya H, Murata M, Wada K, Okada T, Nagai Y, Hohjoh H. Supplemental treatment for Huntington's disease with miR-132 that is deficient in Huntington's disease brain. Mol Ther Nucleic Acids. 2018;11:79–90. doi: 10.1016/j.omtn.2018.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Caron NS, Southwell AL, Brouwers CC, Cengio LD, Xie Y, Black HF, Anderson LM, Ko S, Zhu X, van Deventer SJ, Evers MM, Konstantinova P, Hayden MR. Potent and sustained huntingtin lowering via AAV5 encoding miRNA preserves striatal volume and cognitive function in a humanized mouse model of Huntington disease. Nucleic Acids Res. 2020;48(1):36–54. doi: 10.1093/nar/gkz976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Spronck EA, Brouwers CC, Vallès A, de Haan M, Petry H, van Deventer SJ, Konstantinova P, Evers MM. AAV5-miHTT gene therapy demonstrates sustained huntingtin lowering and functional improvement in Huntington disease mouse models. Mol Therapy Methods Clin Dev. 2019;13:334–343. doi: 10.1016/j.omtm.2019.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Vallès A, Evers MM, Stam A, Sogorb-Gonzalez M, Brouwers C, Vendrell-Tornero C, Acar-Broekmans S, Paerels L, Klima J, Bohuslavova B, Pintauro R, Fodale V, Bresciani A, Liscak R, Urgosik D, Starek Z, Crha M, Blits B, Petry H, Ellederova Z, Motlik J, van Deventer S, Konstantinova P. Widespread and sustained target engagement in Huntington's disease minipigs upon intrastriatal microRNA-based gene therapy. Sci Transl Med. 2021;13(588):eabb8920. doi: 10.1126/scitranslmed.abb8920. [DOI] [PubMed] [Google Scholar]
- 85.Rodrigues FB, Wild EJ. Huntington's disease clinical trials corner: April 2020. J Huntingtons Dis. 2020;9(2):185–197. doi: 10.3233/JHD-200002. [DOI] [PubMed] [Google Scholar]
- 86.Lee TJ, Yuan X, Kerr K, Yoo JY, Kim DH, Kaur B, Eltzschig HK. Strategies to modulate microRNA functions for the treatment of cancer or organ injury. Pharmacol Rev. 2020;72(3):639–667. doi: 10.1124/pr.119.019026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Durso M, Gaglione M, Piras L, Mercurio ME, Terreri S, Olivieri M, Marinelli L, Novellino E, Incoronato M, Grieco P, Orsini G, Tonon G, Messere A, Cimmino A. Chemical modifications in the seed region of miRNAs 221/222 increase the silencing performances in gastrointestinal stromal tumor cells. Eur J Med Chem. 2016;111:15–25. doi: 10.1016/j.ejmech.2016.01.047. [DOI] [PubMed] [Google Scholar]
- 88.Kumar V, Kumar V, Luo J, Mahato RI. Therapeutic potential of OMe-PS-miR-29b1 for treating liver fibrosis. Mol Therapy. 2018;26(12):2798–2811. doi: 10.1016/j.ymthe.2018.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Patutina OA, Gaponova Miroshnichenko SK, Sen’kova AV, Savin IA, Gladkikh DV, Burakova EA, Fokina AA, Maslov MA, Shmendel EV, Wood MJA, Vlassov VV, Altman S, Stetsenko DA, Zenkova MA. Mesyl phosphoramidate backbone modified antisense oligonucleotides targeting miR-21 with enhanced in vivo therapeutic potency. Proc Natl Acad Sci U S A. 2020;117(51):32370–32379. doi: 10.1073/pnas.2016158117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Amanzadeh JE, Esmaeili A, Rahgozar S, Noorbakhshnia M. Quercetin-conjugated superparamagnetic iron oxide nanoparticles protect AlCl(3)-induced neurotoxicity in a rat model of Alzheimer's disease via antioxidant genes, APP gene, and miRNA-101. Front Neurosci. 2020;14:598617. doi: 10.3389/fnins.2020.598617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Saraiva C, Paiva J, Santos T, Ferreira L, Bernardino L. MicroRNA-124 loaded nanoparticles enhance brain repair in Parkinson's disease. J Control Release. 2016;235:291–305. doi: 10.1016/j.jconrel.2016.06.005. [DOI] [PubMed] [Google Scholar]
- 92.Saraiva C, Talhada D, Rai A, Ferreira R, Ferreira L, Bernardino L, Ruscher K. MicroRNA-124-loaded nanoparticles increase survival and neuronal differentiation of neural stem cells in vitro but do not contribute to stroke outcome in vivo. PLoS ONE. 2018;13(3):e0193609. doi: 10.1371/journal.pone.0193609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Mendell JR, Al-Zaidy SA, Rodino-Klapac LR, Goodspeed K, Gray SJ, Kay CN, Boye SL, Boye SE, George LA, Salabarria S, Corti M, Byrne BJ, Tremblay JP. Current clinical applications of in vivo gene therapy with AAVs. Mol Therapy. 2021;29(2):464–488. doi: 10.1016/j.ymthe.2020.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kaifer KA, Villalón E, O'Brien BS, Sison SL, Smith CE, Simon ME, Marquez J, O'Day S, Hopkins AE, Neff R, Rindt H, Ebert AD, Lorson CL. AAV9-mediated delivery of miR-23a reduces disease severity in Smn2B/-SMA model mice. Hum Mol Genet. 2019;28(19):3199–3210. doi: 10.1093/hmg/ddz142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Martier R, Sogorb-Gonzalez M, Stricker-Shaver J, Hübener-Schmid J, Keskin S, Klima J, Toonen LJ, Juhas S, Juhasova J, Ellederova Z, Motlik J, Haas E, van Deventer S, Konstantinova P, Nguyen HP, Evers MM. Development of an AAV-based microRNA gene therapy to treat Machado-Joseph disease. Mol Therapy Methods Clin Dev. 2019;15:343–358. doi: 10.1016/j.omtm.2019.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Evers MM, Miniarikova J, Juhas S, Vallès A, Bohuslavova B, Juhasova J, Skalnikova HK, Vodicka P, Valekova I, Brouwers C, Blits B, Lubelski J, Kovarova H, Ellederova Z, van Deventer SJ, Petry H, Motlik J, Konstantinova P. AAV5-miHTT gene therapy demonstrates broad distribution and strong human mutant Huntingtin lowering in a Huntington's disease minipig model. Mol Ther. 2018;26(9):2163–2177. doi: 10.1016/j.ymthe.2018.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Miniarikova J, Zimmer V, Martier R, Brouwers CC, Pythoud C, Richetin K, Rey M, Lubelski J, Evers MM, van Deventer SJ, Petry H, Déglon N, Konstantinova P. AAV5-miHTT gene therapy demonstrates suppression of mutant huntingtin aggregation and neuronal dysfunction in a rat model of Huntington's disease. Gene Ther. 2017;24(10):630–639. doi: 10.1038/gt.2017.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Pfister EL, DiNardo N, Mondo E, Borel F, Conroy F, Fraser C, Gernoux G, Han X, Hu D, Johnson E, Kennington L, Liu P, Reid SJ, Sapp E, Vodicka P, Kuchel T, Morton AJ, Howland D, Moser R, Sena-Esteves M, Gao G, Mueller C, DiFiglia M, Aronin N. Artificial miRNAs reduce human mutant huntingtin throughout the striatum in a transgenic sheep model of Huntington's disease. Hum Gene Therapy. 2018;29(6):663–673. doi: 10.1089/hum.2017.199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–659. doi: 10.1038/ncb1596. [DOI] [PubMed] [Google Scholar]
- 100.Chen X, Jiang M, Li H, Wang Y, Shen H, Li X, Zhang Y, Wu J, Yu Z, Chen G. CX3CL1/CX3CR1 axis attenuates early brain injury via promoting the delivery of exosomal microRNA-124 from neuron to microglia after subarachnoid hemorrhage. J Neuroinflammation. 2020;17(1):209. doi: 10.1186/s12974-020-01882-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Kim G, Kim M, Lee Y, Byun JW, Hwang DW, Lee M. Systemic delivery of microRNA-21 antisense oligonucleotides to the brain using T7-peptide decorated exosomes. J Control Release. 2020;317:273–281. doi: 10.1016/j.jconrel.2019.11.009. [DOI] [PubMed] [Google Scholar]
- 102.Lai N, Wu D, Liang T, Pan P, Yuan G, Li X, Li H, Shen H, Wang Z, Chen G. Systemic exosomal miR-193b-3p delivery attenuates neuroinflammation in early brain injury after subarachnoid hemorrhage in mice. J Neuroinflammation. 2020;17(1):74. doi: 10.1186/s12974-020-01745-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Yang J, Zhang X, Chen X, Wang L, Yang G. Exosome mediated delivery of miR-124 promotes neurogenesis after ischemia. Mol Therapy Nucleic Acids. 2017;7:278–287. doi: 10.1016/j.omtn.2017.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Lee ST, Im W, Ban JJ, Lee M, Jung KH, Lee SK, Chu K, Kim M. Exosome-based delivery of miR-124 in a Huntington's disease model. J Mov Disord. 2017;10(1):45–52. doi: 10.14802/jmd.16054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Fu MH, Li CL, Lin HL, Tsai SJ, Lai YY, Chang YF, Cheng PH, Chen CM, Yang SH. The potential regulatory mechanisms of miR-196a in Huntington's disease through bioinformatic analyses. PLoS ONE. 2015;10(9):e0137637. doi: 10.1371/journal.pone.0137637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kunkanjanawan T, Carter RL, Prucha MS, Yang J, Parnpai R, Chan AW. miR-196a ameliorates cytotoxicity and cellular phenotype in transgenic Huntington's disease monkey neural cells. PLoS ONE. 2016;11(9):e0162788. doi: 10.1371/journal.pone.0162788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Eykens C, Rossaert E, Duqué S, Rué L, Bento-Abreu A, Hersmus N, Lenaerts A, Kerstens A, Corthout N, Munck S, Van Damme P, Holt MG, von Jonquires G, Klugmann M, Van Den Bosch L, Robberecht W. AAV9-mediated gene delivery of MCT1 to oligodendrocytes does not provide a therapeutic benefit in a mouse model of ALS. Mol Ther Methods Clin Dev. 2021;20:508–519. doi: 10.1016/j.omtm.2021.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Cheng PH, Chang YF, Mao SH, Lin HL, Chen CM, Yang SH. Lentiviral transgenesis in mice via a simple method of viral concentration. Theriogenology. 2016;86(6):1427–1435. doi: 10.1016/j.theriogenology.2016.04.088. [DOI] [PubMed] [Google Scholar]
- 109.Yang SH, Cheng PH, Sullivan RT, Thomas JW, Chan AW. Lentiviral integration preferences in transgenic mice. Genesis. 2008;46(12):711–718. doi: 10.1002/dvg.20435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–345. doi: 10.1038/nbt.1807. [DOI] [PubMed] [Google Scholar]
- 111.Jang E, Kim E, Son HY, Lim EK, Lee H, Choi Y, Park K, Han S, Suh JS, Huh YM, Haam S. Nanovesicle-mediated systemic delivery of microRNA-34a for CD44 overexpressing gastric cancer stem cell therapy. Biomaterials. 2016;105:12–24. doi: 10.1016/j.biomaterials.2016.07.036. [DOI] [PubMed] [Google Scholar]
- 112.Tivnan A, Heilinger T, Ramsey JM, O'Connor G, Pokorny JL, Sarkaria JN, Stringer BW, Day BW, Boyd AW, Kim EL, Lode HN, Cryan SA, Prehn JH. Anti-GD2-ch14.18/CHO coated nanoparticles mediate glioblastoma (GBM)-specific delivery of the aromatase inhibitor, Letrozole, reducing proliferation, migration and chemoresistance in patient-derived GBM tumor cells. Oncotarget. 2017;8(10):16605–16620. doi: 10.18632/oncotarget.15073. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.