

Abstract
The members of the Nedd4-like E3 family participate in various biological processes. However, their role in clear cell renal cell carcinoma (ccRCC) is not clear. This study systematically analyzed the Nedd4-like E3 family members in ccRCC data sets from multiple publicly available databases. NEDD4L was identified as the only NEDD4 family member differentially expressed in ccRCC compared with normal samples. Bioinformatics tools were used to characterize the function of NEDD4L in ccRCC. It indicated that NEDD4L might regulate cellular energy metabolism by co-expression analysis, and subsequent gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis. A prognostic model developed by the LASSO Cox regression method showed a relatively good predictive value in training and testing data sets. The result revealed that NEDD4L was associated with biosynthesis and metabolism of ccRCC. Since NEDD4L is downregulated and dysregulation of metabolism is involved in tumor progression, NEDD4L might be a potential therapeutic target in ccRCC.
Keywords: NEDD4L, Bioinformatics, Clear-cell renal cell cancer, Prognosis, Metabolism
Introduction
Renal cell carcinoma (RCC) is the third most common malignant tumor in the urinary system (Bray et al., 2018). Recently the incidence of RCC has been increasing, and approximately 403,262 new cases and 175,098 kidney cancer-related deaths occurred worldwide in 2018 (Siegel, Miller & Jemal, 2019). Clear-cell renal cell carcinoma (ccRCC) is the most common pathological type and accounts for about two-thirds of all RCCs (Kroeger et al., 2013). Nearly 30–40% of patients with ccRCC will develop metastases (Ghatalia et al., 2019; Pierorazio et al., 2016). The Von Hippel-Lindau/hypoxia-inducible factor (VHL/HIF) axis, the major carcinogenic pathway in ccRCC, promotes angiogenesis, cell growth and glycolysis by activating vascular endothelial growth factor A (VEGFA), transforming growth factor alpha and beta and platelet derived growth factor (Seles et al., 2020; Sanchez-Gastaldo et al., 2017; Maxwell et al., 1999). Compared to chemotherapy and radiotherapy, surgical resection of early-stage renal cancer is more effective (Chow et al., 2010). However, 20–40% of cases will relapse after surgery (Janzen et al., 2003). Accordingly, it is necessary to find biomarkers that can be effectively used to monitor RCC progression and evaluate its prognosis. Research shows that E3 ubiquitin ligases take part in various biological processes involved in the pathogenesis of carcinoma which could be considered as potential biomarker candidates (Wang et al., 2017).
The Nedd4-like E3 family mediates ubiquitination and proteasomal degradation of its substrate proteins as well as receptor-mediated endocytosis. In mammals, there are nine members: NEDD4, NEDD4L, ITCH, SMURF1, SMURF2, WWP1, WWP2, NEDL1 and NEDL2 (Wang et al., 2020). The Nedd4-like E3 family members have similar domains, including an N terminal C2 calcium-dependent phospholipid binding domain, 3–4 WW protein-protein interaction domains, and a C-terminal catalytic HECT ubiquitin ligase domain (Boase & Kumar, 2015). The Nedd4-like E3 family plays an important role in various biological processes. In recent years, the role of the NEDD4 family in urinary system tumors has attracted much attention. In bladder cancer, NEDD4 exerts its oncogenic function by regulating phosphatase and tensin homolog (PTEN) and notch receptor 1 (NOTCH1) expression (Wen et al., 2017). In prostate cancer, NEED4 exerts its oncogenic function by downregulating PTEN and androgen receptor (AR) (Li et al., 2015; Li et al., 2008a; Li et al., 2008b). NEDD4L is down-regulated in prostate cancer, and may exert an antitumor function by regulating TGFβ1 signaling (Li et al., 2015; Li et al., 2008a; Li et al., 2008b). However, research on the function of NEDD4 family members in ccRCC is lacking, and more attention needs to be paid to this topic.
The expression level of NEDD4L has been found significantly decreased in various tumors (Tanksley, Chen & Coffey, 2013; Yang et al., 2015; Hu et al., 2009; Gao et al., 2012; Zhao et al., 2015). It is reported that NEDD4L acts a pivotal part in the prognosis of various malignant carcinomas (Jiang et al., 2019). Also, NEDD4L has been shown to be involved in the regulation of certain major signaling pathways in carcinoma, including the WNT, and TGF-β signaling pathways (Zou, Levy-Cohen & Blank, 2015). In addition, NEDD4L has been shown to act as a cancer suppressor by inhibiting canonical WNT signaling in colorectal cancer (Tanksley, Chen & Coffey, 2013). Additionally, NEDD4L was found to suppress tumor metastasis by inhibiting the activity of TGF-β signaling pathway (Kuratomi et al., 2005). NEDD4L can bind to TGFBRI via SMAD7 and induce its degradation through ubiquitin proteasome pathway, thereby preventing the activation of R-SMADs. Moreover, NEDD4L can bind to SMAD2 and SMAD3 in a ligand-dependent manner and induce the degradation of SMAD2 (Kuratomi et al., 2005). However, NEDD4L was confirmed to have distinct functions in different carcinomas. Furthermore, NEDD4L was found to exert its pro-oncogenic function in gallbladder cancer, where it promotes invasion by regulating transcription of the matrix metallopeptidase 1 and 13 genes (Takeuchi et al., 2011). However, the function of NEDD4L in ccRCC remains unknown.
Fatty acids metabolism exerts new functions in cancer, such as protecting cells from lipotoxicity, facilitating cell migration and drug resistance by altering membrane fluidity, and meeting high energy demands of metastatic cells through fatty acid β-oxidation (FAO)Tumor progression depends on the reprogramming of cell metabolism (Pavlova & Thompson, 2016). In recent years, elevated lipid synthesis is considered to be an important metabolic abnormality for tumorigenesis (Swierczynski, Hebanowska & Sledzinski, 2014). The biosynthesis of fatty acids is increased, and the metabolism of fatty acids exerts new functions in cancer, such as protecting cells from lipotoxicity, facilitating cell migration and drug resistance by altering membrane fluidity, and meeting high energy demands of metastatic cells through fatty acid β-oxidation (FAO) (Chen & Huang, 2019). The rapid proliferation of tumor cells also increases their demand for amino acids. In addition to being used for protein synthesis, amino acids can also be used as metabolites and regulators to support the growth of tumor cells (Li & Zhang, 2016).
There is not a lot of literature on NEDD4L and cell metabolism and these topics need more attention from researchers. In this paper, we integrated data from multiple public databases like the Cancer Genome Atlas (TCGA), Oncomine and gene expression database (GEO) to methodically study the expression profile, prognostic significance and role of NEDD4L in ccRCC. The results indicate that NEDD4L is markedly downregulated in ccRCC, which is related to poor prognosis. Enrichment analysis indicates that NEDD4L may regulate biosynthesis of unsaturated fatty acids and fatty acids metabolism. These findings reveal for the first time the effect of NEDD4L in the prognosis of ccRCC and the potential involvement of this protein in the regulation of fatty acid synthesis and metabolism.
Materials and Methods
Data acquisition and processing
The kidney clear cell Carcinoma (KIRC) project of the TCGA (TCGA_KIRC) data set and GEO data sets (GSE40435 and GSE53757) were acquired and processed using the method described in our previous study (Zhang et al., 2020). Briefly, normalized RNA-seq data (607 cases, HTSeq-FPKM), phenotype information (985 cases), and survival data (979 cases) of the TCGA_KIRC were downloaded from the GDC hub of UCSC xena website (http://xena.ucsc.edu/public). Gene expression data of the GSE40435 and GSE53757 data sets (series matrix file) was downloaded from the Gene Expression Omnibus (GEO) database through the GEOquery package in the R software (version 3.6.2). After data processing, 526 tumor samples with survival data of the TCGA_KIRC data set, 202 samples of the GSE40435, and 144 samples of the GSE53757 were used for further analysis. All data for this study were acquired from public databases and did not require approval from the Ethics Committee.
Online database analysis
The transcriptional level of NEDD4L in renal cell carcinoma was verified in the Oncomine database (https://www.oncomine.org/resource/login.html). The expression data of 5 groups of renal cell carcinoma and normal tissues (transformed by log2) were retrieved and compared statistically. The default threshold was as follows: P <1E−4, multiple change >2, and the top 10% of genes. The staining intensity of NEDD4L in RCC samples and normal kidneys were downloaded from the Human Protein Atlas database (https://www.proteinatlas.org/).
Functional analysis and enrichment analysis
Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were carried out using the clusterProfiler package in R version 3.6.2 (Hsing et al., 2020). Gene set enrichment analysis (GSEA) was used to investigate pathways enriched in the high- and low-risk subgroups. C2.cp.kegg.v7.1.symbols.gmt was chosen as the gene set database. These pathways were considered to be significantly enriched when the following criteria were met: nominal p-value < 0.05, false discovery rate q-value < 0.25, and absolute normalized enrichment score >1.
Development of the prognostic model
The least absolute shrinkage and selection operator (LASSO) Cox regression analysis was performed using the glmnet package in R. The analysis generated key gene signatures, and their corresponding coefficients were obtained by multi-variate cox analysis. A new score was calculated by multiplying the gene expression value of each gene and their corresponding coeffecient, as follows:
Score = − 0.06919*SOWAHB − 0.18041*WDR72 − 0.22124*EPB41L4A-DT − 0.07547*C4orf19 − 0.22609*CDKL2 − 0.05303*IRF6 − 0.16642*DHRS7 − 0.31446*NUPR2.
The risk score was calculated by subtracting the minimum score of the cohort from this score, and dividing the difference by the absolute value of the maximum score of the cohort, namely riskscore = (Score-min(Score))/abs(max(Score)).
Development and evaluation of the nomogram
The independent risk factors determined by multivariate Cox regression analysis were chosen to develop a nomogram for prediction of total overall survival (OS) probability. The consistency index (C-index) was used to evaluate the consistency between the occurrence frequency of the actual results and the prediction probability of the model. The nomogram was generated using the rms package in R software (Li et al., 2019a; Li et al., 2019b).
Statistical analysis
The data collected were analyzed by default as described using web resources. The rest of the data was analyzed using R software (version 3.6.2). The patients were divided into two subgroups by the median value of gene expression or risk score. Briefly, the correlation between the expression of NEDD4L and the clinicopathological features of ccRCC patients was examined by chi-square test with SPSS software (IBM Corp., Armonk, NY, USA). The expression of NEDD4L and other genes was analyzed by Spearman correlation analysis with R software (version 3.6.2). Students’ t-test was used to compare the level of gene expression. Univariate/multivariate Cox regression were performed by using the ‘survminer’ package in R. Kaplan–Meier survival analysis was used to compare the OS, relapse-free survival (RFS), disease specific survival (DSS) and progression free interval (PFI) of high- and low-CIFI groups Time-dependent receiver operator characteristic (ROC) analyses and subsequent calculation of the area under the curve (AUC) (Le, 2019; Le et al., 2019) were performed using the ‘timeROC’ package in R. Packages in R used for data analysis and graph plotting included ggstatsplot, ggplot2, ggpubr, limma, survminer, survival, tidyverse, dplyr, and plyr. A value of p < 0.05 was considered to be statistically significant (*, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001).
Results
Patient characteristics
The data were collected from the TCGA_KIRC data set, including 72 normal subjects and 530 patients with primary tumors with clinical and gene expression data. The 526 patients with survival information were used for follow-up analysis (Table S1). There were 183 females (34.79%) and 343 males (65.21%). The onset age ranged from 26 to 88 years old, and the median age was 60 years old, 245 (46.58%) cases were younger than 60 years, and 281 (53.42%) cases were older than 60 years. The classification of the tumors according to the TNM (tumor, node, metastasis) staging system was as follows: 267 (50.76%) cases with T1 stage and 259 (49.24%) cases with T2-4 stage; 238 (45.25%) cases with N0 stage and 288 (54.75%) cases with N1/X stage; 418 (79.47%) cases with M0 stage, 106 (20.15%) cases with N1/X stage, and 2 (0.38%) cases of unknown stage; 239 (45.44%) cases with G1/2 stage, 279 (53.04%) cases with G3/4, and 8 (1.52%) cases of unknown stage. The classification of the tumors according to the Overall Stage Grouping (OSG) was as follows: 318 (60.46%) cases with stage I/II, 205 (38.97%) cases with stage III/IV, and 3 (0.57%) cases of unknown stage; 330 (60.46%) cases with stage I/II, 147 (38.97%) cases with stage III/IV, and 49 (0.57%) cases of unknown stage. The GSE40435 (101 normal subjects and 101 primary tumors), and GSE53757 (70 normal subjects and 70 primary tumors) data sets from the GEO database were used for verification.
NEDD4L is downregulated in ccRCC
We screened the transcription levels of NEDD4 family members in the TCGA_KIRC. The differentially expressed genes (DEGs) between normal and tumor tissues were analyzed using the following criteria: fold change (FC) >2 and FDR< 0.05. The results revealed that only NEDD4L was meaningfully down-regulated in tumors compared with that in normal tissues (Fig. 1A). Subsequently, two data sets from the GEO database were used for verification, and the results confirmed that NEDD4L was significantly down-regulated in ccRCC carcinoma tissues (Figs. 1B–1C). Data from the Oncomine database made up of five separate queues also confirmed that NEDD4L was meaningfully down-regulated in ccRCC (Fig. 1D). Immunohistochemical results showed that the expression level of NEDD4L in renal cell carcinoma was significantly decreased than that in normal kindey (Fig. S1).
Figure 1. Expression and prognostic analysis of NEDD4L in ccRCC.

(A) In TCGA_KIRC data set, DEGs were represented in a volcano plot. (B & C) Mining of the GEO database showed that NEDD4L was downregulated in tumor tissues. (D) Data from the Oncomine database confirmed that NEDD4L was significantly downregulated in tumor tissues. (E) Kaplan–Meier curves of the OS of the patients with ccRCC. (F) Kaplan–Meier curves of the PFI of the patients with ccRCC. (G) Kaplan–Meier curves of the DSS of the patients with ccRCC.
Low expression of NEDD4L correlated to dismal prognosis in ccRCC
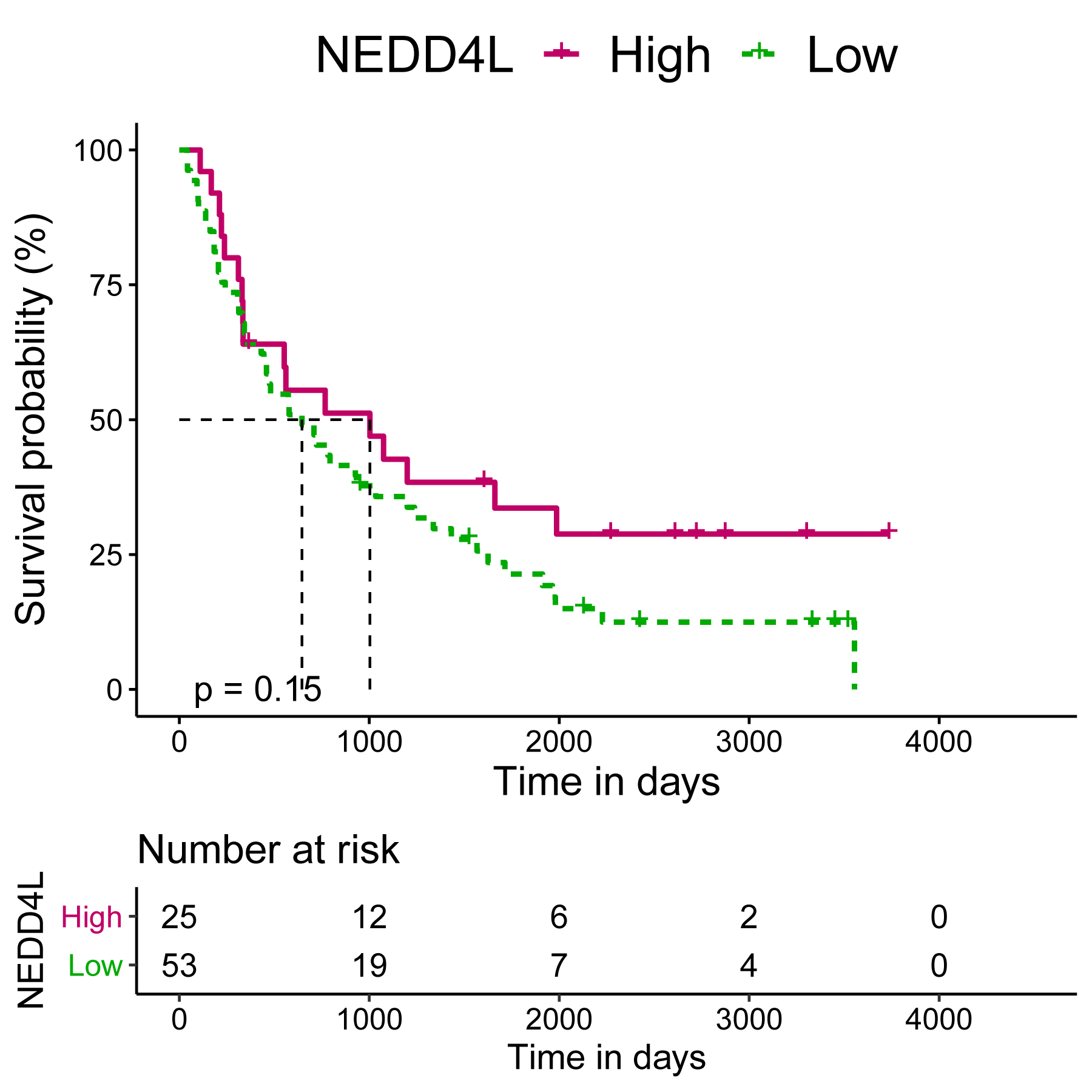
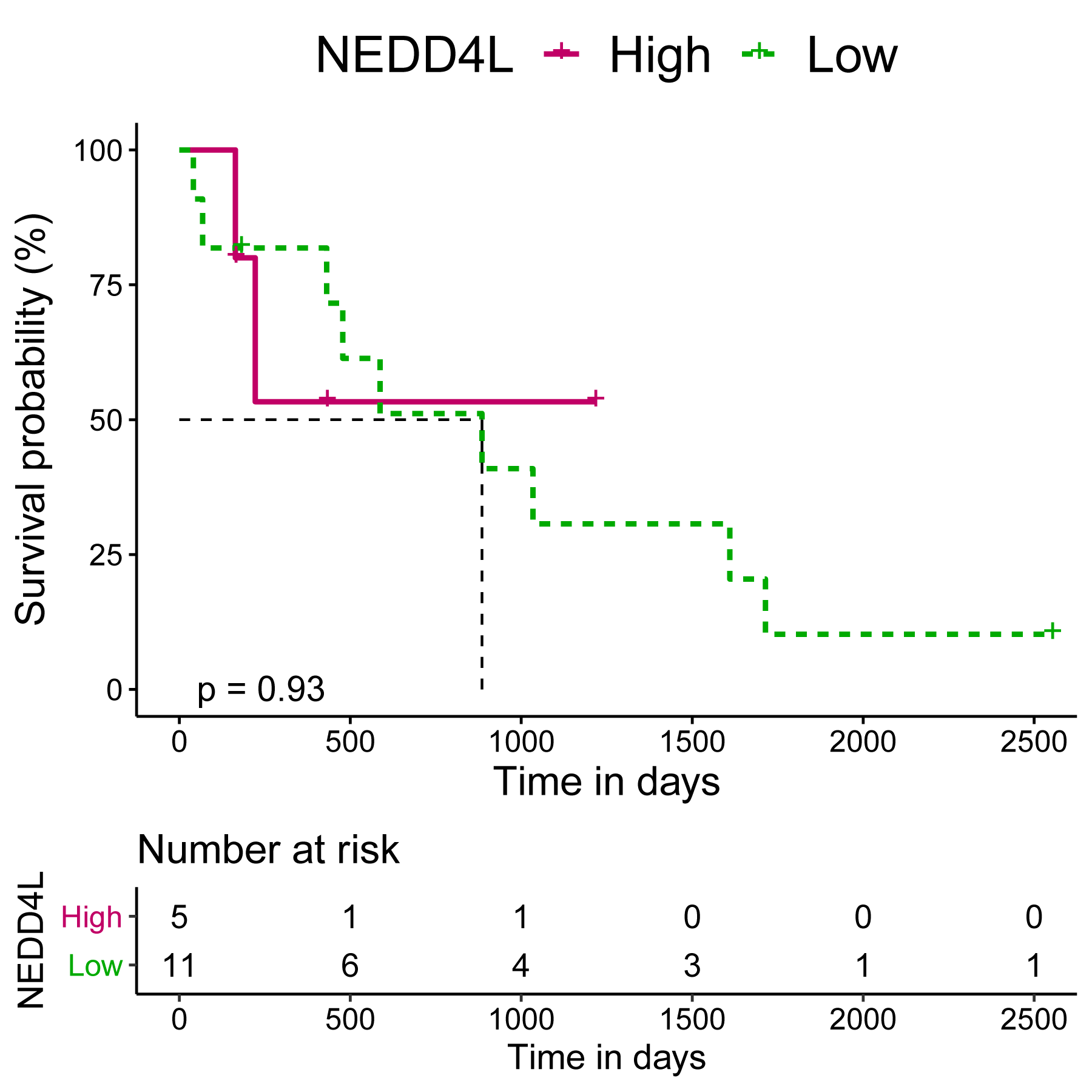
The TCGA_KIRC data set was used to investigate the prognostic value of NEDD4L in ccRCC. After data filtering, a total of 526 samples with gene expression and survival data were included. The median of the gene expression was set as the cut-off value. Kaplan–Meier survival analysis showed that patients with low NEDD4L expression had shorter OS (P < 0.0001), PFS (P < 0.0001) and DSS (P < 0.0001) (Figs. 1E–1G). Patients were divided into different subgroups according to sex, age, laterality, cancer status, histological grade, TNM stage, and OSG tumor stage. Analysis results of all but the N1 and M1 subgroups showed that patients with high NEDD4L expression had longer OS (Figs. 2 and 3, Figs. S2 & S3). Univariate and multivariate analysis were used to validate the prognostic value of NEDD4L in ccRCC. The univariate analysis results demonstated that low NEDD4L expression was associated with shorter OS (hazard ratio [HR]: 2.323; 95% CI [1.683−3.208]; P = 0.001) (Table 1). Multivariate COX regression analysis revealed that low expression of NEDD4L was still an independent factor associated with the deterioration of OS (HR: 1.905; CI [1.33–2.73], P < 0.001) (Table 1). These results suggested that NEDD4L may be an independent prognostic factor for ccRCC.
Figure 2. Prognostic analysis of NEDD4L in ccRCC patients grouped by sex, age, laterality, and cancer status.

(A–D) Kaplan–Meier plot of the OS in high- and low-NEDD4L expression subgroups of ccRCC patients who were grouped according to sex (A), age (B), laterality (C), and cancer status (D).
Figure 3. Prognostic analysis of NEDD4L in ccRCC patients grouped by histological grade, TNM stage, and tumor stage.

(A–D) Kaplan–Meier plot of the OS in high- and low-NEDD4L expression subgroups of ccRCC patients who were grouped according to histological grade (A), TNM stage (B & C), and tumor stage (D).
Table 1. Univariate and multivariate analysis of overall survival in TCGA patients with ccRCC.
| Variables | Univariate analyses | Multivariate analyses | ||
|---|---|---|---|---|
| Hazard ratio (95% CI) | p -value | Hazard ratio (95% CI) | p -value | |
| Age (<60 vs. ≥60) | 0.557 (0.406–0.746) | <0.001 | 0.681 (0.487–0.953) | 0.025 |
| Sex (female vs. male) | 1.056 (0.773–1.442) | 0.734 | ||
| Laterality (left vs. right) | 1.409 (1.042–1.904) | 0.026 | 1.345 (0.972–1.860) | 0.073 |
| NEDD4L (low vs. high) | 2.323 (1.683–3.208) | <0.001 | 1.905 (1.330–2.730) | <0.001 |
| T (T1 vs. T2/3/4) | 0.336 (0.240–0.470) | <0.001 | 1.328 (0.707–2.495) | 0.378 |
| N (N0 vs.N1/X) | 1.106 (0.819–1.493) | 0.511 | ||
| M (M0 vs. M1/X) | 0.265 (0.195–0.362) | <0.001 | 0.828 (0.558–1.231) | 0.351 |
| Cancer status (tumor free vs. with tumor) | 0.195 (0.140–0.273) | <0.001 | 0.334 (0.225–0.495) | <0.001 |
| Stage (stage I vs. stage II/III/IV) | 0.253 (0.183–0.349) | <0.001 | 0.447 (0.240–0.834) | 0.011 |
| Grade (G1/2 vs. G3/4) | 0.374 (0.265–0.529) | <0.001 | 0.759 (0.508–1.134) | 0.178 |
NEDD4L downregulation correlated with worse clinicopathological features of ccRCC
To investigate the relationship between the expression of NEDD4L and clinicopathological features in renal cell carcinoma, patients were divided into low and high expression subgroup on the base of the median expression of NEDD4L. Chi-square test showed that lower levels of NEDD4L were significantly associated with adverse prognostic features, including ccRCC with higher T stages (OR = 0.456 (0.322−0.646) for T1 vs. T2/3/4, P < 0.001), M stage (OR = 0.530 (0.342−0.821) for M0 vs. M1/X, P = 0.004), OSG tumor stage (OR = 0.429 (0.299−0.615) for stage I/II vs. stage III/IV, P < 0.001), histological grade (OR = 0.456 (0.321−0.647) for G1/2 vs. G3/4, P < 0.001) (Table 2).
Table 2. NEDD4L expression associated with clinical characteristics.
| Clinical characteristics | Total (N) | OR (95% CI) | P |
|---|---|---|---|
| T (T1 vs. T2/3/4) | 526 | 0.456 (0.322–0.646) | <0.001 |
| M (M0 vs. M1/X) | 524 | 0.530 (0.342–0.821) | 0.004 |
| N (N0 vs.N1/X) | 526 | 1.063 (0.754–1.499) | 0.726 |
| Stage (stage I/II vs. stage III/IV) | 523 | 0.429 (0.299–0.615) | <0.001 |
| Grade (G1/2 vs. G3/4) | 523 | 0.456 (0.321–0.647) | <0.001 |
| Sex (female vs. male) | 526 | 0.702 (0.490–1.008) | 0.055 |
| Age (<60 vs. ≥60) | 526 | 0.771 (0.547–1.087) | 0.137 |
| Laterality (left vs. right) | 525 | 1.411 (1.000–1.991) | 0.049 |
| Cancer status (tumor free vs. with tumor) | 477 | 0.443 (0.296–0.662) | <0.001 |
Notes.
- OR
- odds ratio
- CI
- confidence interval
P values were calculated from two-sided chi-square test.
Co-expression and enrichment analysis
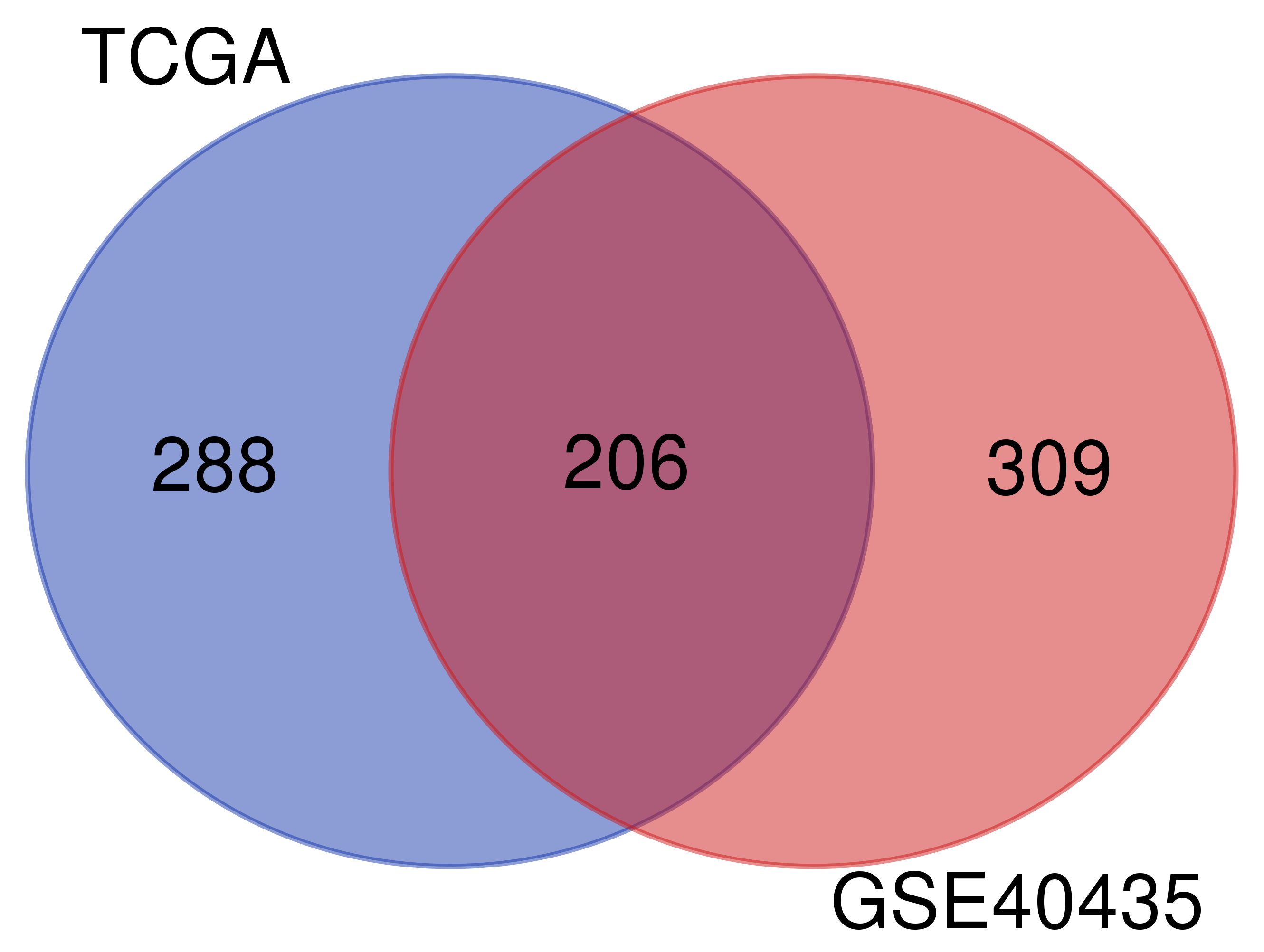
Since NEDD4L was downregulated in ccRCC and correlated with worse prognosis of patients with ccRCC, we undertook to elucidate its potential role in this disease. In TCGA_KIRC and GSE40435 data sets, the correlation between the expression of NEDD4L and other genes in tumor samples was analyzed. A total of 494 genes in the TCGA_KIRC and 515 genes in the GSE40435 were identified as NEDD4L-related genes, using the following criteria: coefficient >0.5 and p value < 0.05. Among them, 206 NEDD4L-related genes were shared between the two datasets (Fig. S4, Table S2 ) and were subjected to GO and KEGG analysis. GO analysis revealed that the proteins encoded by these genes were mainly located in mitochondrial membrane and matrix, involved in the catabolism of many metabolites, such as lipid oxidation, fatty acid oxidation, fatty acid catabolism, monocarboxylic acid catabolism, and performed various molecular functions, such as DNA binding and ATP enzyme activity (Fig. 4A, Table S3). Accordingly, KEGG analysis showed that these genes play a major role in amino acid metabolism, oxidative phosphorylation and citrate cycle (Fig. 4B). These results suggest that NEDD4L is closely related to ccRCC cell metabolism.
Figure 4. GO and KEGG enrichment analysis of NEDD4L.

(A) GO enrichment analysis of NEDD4L. (B) KEGG enrichment analysis of NEDD4L.
Development of a NEDD4L-related prognostic model
To develop a NEDD4L-related prognostic model for ccRCC, we first performed univariate Cox regression analysis on the aforementioned 206 NEDD4L-related genes, which identified 183 genes with significant prognostic relevance (p < 0.05, Table S2). Patients from TCGA_KIRC (n = 526) were randomly and equally divided into two groups, one is to establish a LASSO Cox regression model, and the other is to evaluate the effectiveness of the model. The 183 NEDD4L-related genes with significant prognostic relevance were input into a LASSO Cox regression model, which generated eight key genes (Figs. 5A–5B), namely DHRS7, NUPR2, C4orf19, CDKL2, SOWAHB, WDR72, EPB41L4A-DT, and IRF6 (Fig. 5C). The risk score was calculated by inputting the selected signature genes into the above formula. The median risk score was set to a cutoff value, dividing patients into low-risk subgroups and high-risk subgroups. Prognostic analysis with Kaplan–Meier method showed that patients with ccRCC in the low risk subgroup had significantly longer DSS (Figs. 6A–6C). The analysis of the time-related receiver operating characteristic (ROC) in the TCGA training data set shows that the risk score had a favorable predictive value (Fig. 6D, area under the curve (AUC) at 1 year = 0.72, AUC at 3 years = 0.75, AUC at 5 years = 0.8). The predictions of the established risk scores were further confirmed by the TCGA test data set and the entire TCGA data set. The ROC analysis of the TCGA testing dataset showed that the AUC was 0.76 at 1 year, 0.69 at 3 years, and 0.68 at 5 years (Fig. 6E), while that of the TCGA entire dataset showed that the AUC was 0.74 at 1 year, 0.71 at 3 years, and 0.74 at 5 years (Fig. 6F). As reflected by the AUC values at 1 year, 3 year and 5 years, the signatures developed in our work had relatively better performance in predicting outcome of ccRCC patients (Fig. 6F, Fig. S5) (Zhang, Qiu & Yang, 2021; Fei, Chen & Xu, 2021; Chu et al., 2021; Xing et al., 2021).
Figure 5. Construction of LASSO Cox regression model.

(A–C) A LASSO Cox regression model was developed from NEDD4L and 183 related genes, calculating the tuning parameter (λ) based on the partial likelihood deviance with tenfold cross-validation. Each dot represents a patient, and the heatmap represents the genes expression levels. The optimal log λ value is indicated by the vertical black line in the plot.
Figure 6. Reliability verification of NEDD4L-related prognostic model.

(A–C) Kaplan–Meier plot of the DDS in the high- and low-risk groups of the TCGA_KIRC cohort. (D–F) Time-dependent ROC analysis of the risk score for the DDS and survival status in the TCGA_KIRC cohort.
Identification of NEDD4L-related signaling pathways by GSEA
Various biological processes are recognized to be involved in the occurrence and development of ccRCC. Accordingly, we hypothesized that the worse survival related to low expression of NEDD4L may be associated with some activated signal pathways in ccRCC. To evaluate this hypothesis, we performed a GSEA of the differences in the NEDD4L-related low risk group in TCGA_KIRC to verify the key signaling pathways related to NEDD4L expression. There were significant differences in the richness of MSigDB sets (c2.cp.v6.2 symbols) of multiple pathways (normalized P < 0.05, FDR < 0.25). According to the normalized enrichment score (NES), the signaling pathways most significantly associated with NEDD4L expression are shown in Fig. 7 and Table 3. In particular, NEDD4L expression was associated with biosynthesis of unsaturated fatty acids, insulin signaling pathway, and fatty acid metabolism, supporting a possible role of NEDD4L in the regulation of various metabolic processes
Figure 7. GSEA between ccRCC patients with high and low NEDD4L expression in the TCGA_SKCM and GSE40435 cohorts (A–H).

Table 3. GSEA of the NEDD4L-related low risk group in TCGA_KIRC.
| MSigDB collection | Gene set name | NES | NOM p-value |
|---|---|---|---|
| c2.cp.v6.2.symbols.gmt | ERBB signaling pathway | 1.812 | 0.002 |
| Vasopressin regulated water reabsorption | 1.713 | 0.012 | |
| Propanoate metabolism | 1.708 | 0.041 | |
| Neurotrophin signaling pathway | 1.660 | 0.008 | |
| Ubiquitin mediated proteolysis | 1.653 | 0.018 | |
| RNA degradation | 1.650 | 0.008 |
Notes.
- NES
- normalized enrichment score
- NOM
- nominal
- FDR
- false discovery rate
Gene sets with NOM p-value < 0.05 and FDR q-value < 0.25 are considered as significant.
Establishment of a nomogram based on NEDD4L
Univariate and multivariate Cox regression analysis were achieved on the TCGA_KIRC ccRCC data set to investigate whether the risk score was an independent predictor of prognosis in patients with ccRCC. The adjustment of conventional clinical models, including age, sex, laterality, TNM stage, cancer status, histological grade, and tumor stage, indicated that the risk score was also an independent prognostic factor, which confirmed it had a strong ability to predict OS in patients with ccRCC (OR = 1.9329 (1.3429–2.782), P < 0.001, Figs. 8A–8B)
Figure 8. Integration of risk score and clinical characteristics.

(A & B) Univariate and multivariate regression analysis of the relation between the risk score and clinicopathological characteristics regarding OS in the TCGA_KIRC. (C) Nomogram developed to predict the 1-, 3-, and 5-year OS in the TCGA_KIRC data set.
To establish a quantitative method for ccRCC prognosis, we used independent clinical risk factors to develop a nomogram (Fig. 8C). The point scale of nomogram is used to assign points to each variable based on multivariate Cox analysis. We drew a horizontal line to determine the points of each variable, then added the points of all variables together to calculate the total score of each patient, and then normalize it to a distribution of 0 to 45. The estimated 1 -, 3-and 5-year survival rates of ccRCC patients were calculated by drawing a vertical line between the total point axis and each prognostic axis (Li et al., 2019a; Li et al., 2019b). The C-index of the nomogram 0.7627102 (0.7255857–0.7998348) suggesting a good prediction effect. Thus, our results prove that nomogram was the best model for predicting the prognosis of renal cell carcinoma compared with a single risk factor.
Discussion
The Nedd4-like E3 family includes nine members, which regulate key signaling pathways in tumorigenesis, such as TGFβ, EGF, IGF, VEGF, CXCL12, and TNF (Chen & Matesic, 2007). In cancers, NEDD4, WWP1, WWP2, SMURF1, and SMURF2 have been shown to function as oncogenes, while NEDD4L acts as an antioncogene (Chen & Matesic, 2007; Tanksley, Chen & Coffey, 2013; Zhao et al., 2015). This study systematically investigated the role of the NEDD4 family in ccRCC through the comprehensive analysis of TCGA_KIRC and GEO data sets and Oncomine database. The analysis revealed that only NEDD4L was significantly downregulated in ccRCC, and low expression of NEDD4L was associated with shorter survival (Fig. 1).
NEDD4 is shown to be overexpressed in various types of human cancers including gastric cancer (Kim et al., 2008; Sun et al., 2014; Reichert-Penetrat et al., 1998), colorectal cancer (Kim et al., 2008), breast cancer (Kim et al., 2008), non-small-cell lung carcinoma (Amodio et al., 2010; Shao et al., 2018), and hepatocellular carcinoma (Hang et al., 2016; Huang et al., 2017). NEDD4 acts as a double-edged sword in tumors. On the one hand, NEDD4 promotes tumor survival by inducing cell proliferation, inhibiting apoptosis, disrupting the cell cycle, promoting cell migration and invasion, and enhancing drug resistance (Wang et al., 2020). On the other hand, NEDD4 inhibits tumor proliferation by binding to MYC, inhibiting HER3 expression, and targeting PIP5K1A. SMURFs also plays a dual role in tumors, both as tumor promoters and suppressors (David, Nair & Pillai, 2013). By controlling the stability of several key proteins, these proteins play a central role in cell cycle progression, proliferation, differentiation, metastasis, genome stability and senescence. WWP1 is frequently upregulated in multiple human malignancies, and has been found to play an important role in cell proliferation, apoptosis and invasion (Nourashrafeddin et al., 2015; Zhi & Chen, 2012; Chen et al., 2007a; Chen et al., 2007b; Chen & Zhang, 2018). WWP2 functions as an oncogene in liver cancer, breast cancer, endometrial cancer, gastric cancer, lung cancer, oral cancer, and ovarian cancer (Clements et al., 2015; Fang et al., 2020; Fukumoto et al., 2014; Jung et al., 2014; Wang et al., 2020; Butt et al., 2019; Sakashita et al., 2013; Wan et al., 2019). The expression of NEDL1 in benign neuroblastoma is higher than that in non-malignant neuroblastoma (Li et al., 2008a; Li et al., 2008b). So far as we know, there are no reportes on ITCH and NEDL2 in tumors, thus their function in ccRCC is unclear. Our study showed that their expression levels were no significantly changed in ccRCC, which is inconsistent with reports in other tumors, which may be due to tumor specificity.
NEDD4L has been considered as a tumor suppressor. Decreased NEDD4L expression has been found in colorectal cancer (Tanksley, Chen & Coffey, 2013), gastric cancer (Jiang et al., 2019), ovarian cancer (Yang et al., 2015), prostate cancer (Hu et al., 2009), and non-small cell lung cancer (Hu et al., 2009). Consistent with previous reports, this study suggested that NEDD4L acts as a tumor suppressor in ccRCC. Subsequent analysis showed that NEDD4L is an independent prognostic factor for ccRCC and might be associated with energy metabolism. Lee et al. reported that NEDD4L inhibited autophagy and mitochondrial metabolism by downregulating the expression of ULK1 or ASCT2, thereby inhibiting the growth and survival of pancreatic cancer cells. Kim et al. (2018) reported that NEDD4L acts a pivotal part in the feedback regulation of cAMP signal by limiting CREB-regulated transcription coactivator 3 protein levels. The findings of the above studies are consistent with the findings of our study, indicating that NEDD4L plays an important role in tumor metabolism.
We also established a prognostic model based on NEDD4L using the LASSO Cox regression, a broadly selected machine learning algorithm used to minimize the risk of overfitting. The model had good predictive value for training and test queues (Fig. 6). A total of 526 cases from TCGA_KIRC were randomly divided into low-risk and high-risk subgroups, and then analyzed by GSEA, GO, and KEGG enrichment analysis, which are widely used bioinformatics tools in the functional characterization of specific genes.
All of these enrichment analyses suggested that the main difference between the two groups is in metabolism (Figs. 4 and 7, Table 3). The decreased expression of NEDD4L may lead to metabolic disorder and promote the development of ccRCC.
Lee et al. (2020) study found that NEDD4L could inhibit the growth and survival of pancreatic cancer cells by inhibiting autophagy and mitochondrial metabolism. Zhang et al. reported that NEDD4L also played vital roles in bone metabolism. These studies provided strong evidence for the involvement of NEDD4L in cell metabolism (PMID: 22957059). The prediction model developed in this study showed that DHRS7, NUPR2, C4orf19, CDKL2, SOWAHB, WDR72, EPB41L4A_DT, and IRF6 were significantly associated with NEDD4L function.
DHRS7 can recognize steroids and retinoids as potential substrates, and might act a pivotal part in the metabolism of these signal molecules (Stambergova et al., 2016). Lopez et al. (2015) reported that NUPR2 could induce G1 phase cell cycle arrest, decrease cell survival rate and proliferation ability. C4orf19 was found to be significantly downregulated in multicentric breast cancer, but its function is unclear (Lang et al., 2017). Two studies showed that the decreased expression of CDKL2 was associated with the progression and poor prognosis of gliomas (Yi et al., 2018; Yi et al., 2020), and another study showed that CDKL2 promoted EMT and progression of breast cancer (Li et al., 2014). WDR72 was found to be crucial in ameloblasts and the kidneys, and may act a pivotal part in enamel maturation (Rungroj et al., 2018). EPB41L4A_DT was reported to inhibit the progression of ovarian cancer, hepatocellular carcinoma, non-small cell lung cancer, and breast cancer (Sun, Yang & Gao, 2020; Wang et al., 2019; Shu et al., 2018; Xu et al., 2016). IRF6 has been proved to be a tumor suppressor in breast and gastric cancer (Li et al., 2019a; Li et al., 2019b; Xu et al., 2019). All these genes play important roles in a variety of solid tumors, indirectly confirming the importance of NEDD4L in ccRCC.
A NEDD4L-related prognostic model of ccRCC was established by LASSO Cox regression. After adjusting the routine clinical features, the risk score was confirmed as an independent prognostic factor. It seemed that the risk score may have a stronger predictive power than the traditional prognostic factors. We also conducted a comprehensive assessment in combination with risk scores and other important clinical models. According to the calibration graph, there was a significant consistency between the actual and predicted values of the 1 -, 3-and 5-year OS (Fig. 8). In order to show the reliability of our risk model, we compared it with other ccRCC risk models, and the results showed that our risk model has better prediction ability (Fig. S5) (Zhang, Qiu & Yang, 2021; Fei, Chen & Xu, 2021; Chu et al., 2021; Xing et al., 2021). Based on the complementary perspective of the corresponding tumor, the model provides a personalized score for individual patients and a valuable new prognostic evaluation method for clinicians.
Although this study provides new clues for the relationship between NEDD4L expression and ccRCC progression, it still has some limitations. First, it is necessary to further verify the function of NEDD4L in ccRCC and cellular metabolic processes, and completely elucidate the specific mechanism involved. Second, our risk score requires a large sample of prospective studies to be demonstrated, which is just a retrospective analysis.
Supplemental Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Acknowledgments
We thank Editorbar for providing language assistance in the preparation of this article.
Funding Statement
This work was supported by the National Natural Science Foundation of China (81903031), China Postdoctoral Science Foundation (2020M682334), Henan Postdoctoral Foundation (202003002), the Open Project of State Key Laboratory of Cancer Biology of China (CBSKL2019KF12, CBSKL2019KF13). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Contributor Information
Baile Zuo, Email: zuobaile@xxmu.edu.cn.
Guoyin Li, Email: 20181004@zknu.edu.cn, guoyin0604@163.com.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Hui Zhao, Junjun Zhang and Xiaoliang Fu performed the experiments, prepared figures and/or tables, authored or reviewed drafts of the paper, and approved the final draft.
Dongdong Mao, Xuesen Qi, Gang Meng, Zewen Song, Ru Yang, Zhenni Guo, Binghua Tong and Meiqing Sun analyzed the data, prepared figures and/or tables, and approved the final draft.
Baile Zuo and Guoyin Li conceived and designed the experiments, authored or reviewed drafts of the paper, and approved the final draft.
Data Availability
References
- Amodio et al. (2010).Amodio N, Scrima M, Palaia L, Salman AN, Quintiero A, Franco R, Botti G, Pirozzi P, Rocco G, De Rosa N, Viglietto G. Oncogenic role of the E3 ubiquitin ligase NEDD4-1, a PTEN negative regulator, in non-small-cell lung carcinomas. American Journal of Pathology. 2010;177:2622–2634. doi: 10.2353/ajpath.2010.091075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boase & Kumar (2015).Boase NA, Kumar S. NEDD4: the founding member of a family of ubiquitin-protein ligases. Gene. 2015;557:113–122. doi: 10.1016/j.gene.2014.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bray et al. (2018).Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians. 2018;68:394–424. doi: 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- Butt et al. (2019).Butt G, Yaylim I, Attar R, Aras A, Romero MA, Qureshi MZ, Purenovic J, Farooqi AA. NEDD4 family of E3 ubiquitin ligases in breast cancer: spotlight on SMURFs, WWPs and NEDD4. Advances in Experimental Medicine and Biology. 2019;1152:365–375. doi: 10.1007/978-3-030-20301-6_19. [DOI] [PubMed] [Google Scholar]
- Chen & Matesic (2007).Chen C, Matesic LE. The Nedd4-like family of E3 ubiquitin ligases and cancer. Cancer and Metastasis Reviews. 2007;26:587–604. doi: 10.1007/s10555-007-9091-x. [DOI] [PubMed] [Google Scholar]
- Chen et al. (2007a).Chen C, Sun X, Guo P, Dong XY, Sethi P, Zhou W, Zhou Z, Petros J, Frierson HJ, Vessella RL, Atfi A, Dong JT. Ubiquitin E3 ligase WWP1 as an oncogenic factor in human prostate cancer. Oncogene. 2007a;26:2386–2394. doi: 10.1038/sj.onc.1210021. [DOI] [PubMed] [Google Scholar]
- Chen et al. (2007b).Chen C, Zhou Z, Ross JS, Zhou W, Dong JT. The amplified WWP1 gene is a potential molecular target in breast cancer. International Journal of Cancer. 2007b;121:80–87. doi: 10.1002/ijc.22653. [DOI] [PubMed] [Google Scholar]
- Chen & Zhang (2018).Chen JJ, Zhang W. High expression of WWP1 predicts poor prognosis and associates with tumor progression in human colorectal cancer. American Journal of Cancer Research. 2018;8:256–265. [PMC free article] [PubMed] [Google Scholar]
- Chen & Huang (2019).Chen M, Huang J. The expanded role of fatty acid metabolism in cancer: new aspects and targets. Precision Clinical Medicine. 2019;2:183–191. doi: 10.1093/pcmedi/pbz017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chow et al. (2010).Chow TF, Youssef YM, Lianidou E, Romaschin AD, Honey RJ, Stewart R, Pace KT, Yousef GM. Differential expression profiling of microRNAs and their potential involvement in renal cell carcinoma pathogenesis. Clinical Biochemistry. 2010;43:150–158. doi: 10.1016/j.clinbiochem.2009.07.020. [DOI] [PubMed] [Google Scholar]
- Chu et al. (2021).Chu G, Xu T, Zhu G, Liu S, Niu H, Zhang M. Identification of a novel protein-based signature to improve prognosis prediction in renal clear cell carcinoma. Frontiers in Molecular Biosciences. 2021;8:623120. doi: 10.3389/fmolb.2021.623120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clements et al. (2015).Clements AE, Bravo V, Koivisto C, Cohn DE, Leone G. WWP2 and its association with PTEN in endometrial cancer. Gynecologic Oncology Reports. 2015;13:26–29. doi: 10.1016/j.gore.2015.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- David, Nair & Pillai (2013).David D, Nair SA, Pillai MR. Smurf E3 ubiquitin ligases at the cross roads of oncogenesis and tumor suppression. Biochimica Et Biophysica Acta/General Subjects. 2013;1835:119–128. doi: 10.1016/j.bbcan.2012.11.003. [DOI] [PubMed] [Google Scholar]
- Fang et al. (2020).Fang S, Zhang D, Weng W, Lv X, Zheng L, Chen M, Fan X, Mao J, Mao C, Ye Y, Xu M, Ji J. CPSF7 regulates liver cancer growth and metastasis by facilitating WWP2-FL and targeting the WWP2/PTEN/AKT signaling pathway. Biochimica et Biophysica Acta - Molecular Cell Research. 2020;1867:118624. doi: 10.1016/j.bbamcr.2019.118624. [DOI] [PubMed] [Google Scholar]
- Fei, Chen & Xu (2021).Fei H, Chen S, Xu C. Construction autophagy-related prognostic risk signature combined with clinicopathological validation analysis for survival prediction of kidney renal papillary cell carcinoma patients. BMC Cancer. 2021;21:411. doi: 10.1186/s12885-021-08139-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukumoto et al. (2014).Fukumoto C, Nakashima D, Kasamatsu A, Unozawa M, Shida-Sakazume T, Higo M, Ogawara K, Yokoe H, Shiiba M, Tanzawa H, Uzawa K. WWP2 is overexpressed in human oral cancer, determining tumor size and poor prognosis in patients: downregulation of WWP2 inhibits the AKT signaling and tumor growth in mice. Oncoscience. 2014;1:807–820. doi: 10.18632/oncoscience.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao et al. (2012).Gao C, Pang L, Ren C, Ma T. Decreased expression of Nedd4L correlates with poor prognosis in gastric cancer patient. Medical Oncology. 2012;29:1733–1738. doi: 10.1007/s12032-011-0061-3. [DOI] [PubMed] [Google Scholar]
- Ghatalia et al. (2019).Ghatalia P, Gordetsky J, Kuo F, Dulaimi E, Cai KQ, Devarajan K, Bae S, Naik G, Chan TA, Uzzo R, Hakimi AA, Sonpavde G, Plimack E. Prognostic impact of immune gene expression signature and tumor infiltrating immune cells in localized clear cell renal cell carcinoma. Journal for Immuno Therapy of Cancer. 2019;7:139. doi: 10.1186/s40425-019-0621-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hang et al. (2016).Hang X, Zhu S, Di H, Wu Z, Chu K, Wang J, Xin H, Yu G, Peng H, Miao X, Xu W. NEDD4 depletion inhibits hepatocellular carcinoma growth via targeting PTEN. Cellular Physiology and Biochemistry. 2016;39:768–779. doi: 10.1159/000445667. [DOI] [PubMed] [Google Scholar]
- Hsing et al. (2020).Hsing M, Wang Y, Rennie PS, Cox ME, Cherkasov A. ETS transcription factors as emerging drug targets in cancer. Medicinal Research Reviews. 2020;40:413–430. doi: 10.1002/med.21575. [DOI] [PubMed] [Google Scholar]
- Hu et al. (2009).Hu XY, Xu YM, Fu Q, Yu JJ, Huang J. Nedd4L expression is downregulated in prostate cancer compared to benign prostatic hyperplasia. European Journal of Surgical Oncology (EJSO) 2009;35:527–531. doi: 10.1016/j.ejso.2008.09.015. [DOI] [PubMed] [Google Scholar]
- Huang et al. (2017).Huang ZJ, Zhu JJ, Yang XY, Biskup E. NEDD4 promotes cell growth and migration via PTEN/PI3K/AKT signaling in hepatocellular carcinoma. Oncology Letters. 2017;14:2649–2656. doi: 10.3892/ol.2017.6532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janzen et al. (2003).Janzen NK, Kim HL, Figlin RA, Belldegrun AS. Surveillance after radical or partial nephrectomy for localized renal cell carcinoma and management of recurrent disease. Urologic Clinics of North America. 2003;30:843–852. doi: 10.1016/s0094-0143(03)00056-9. [DOI] [PubMed] [Google Scholar]
- Jiang et al. (2019).Jiang X, Zhang S, Yin Z, Sheng Y, Yan Q, Sun R, Lu M, Zhang Z, Li Y. The correlation between NEDD4L and HIF-1alpha levels as a gastric cancer prognostic marker. International Journal of Medical Sciences. 2019;16:1517–1524. doi: 10.7150/ijms.34646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung et al. (2014).Jung JG, Stoeck A, Guan B, Wu RC, Zhu H, Blackshaw S, Shih I, Wang TL. Notch3 interactome analysis identified WWP2 as a negative regulator of Notch3 signaling in ovarian cancer. PLOS Genetics. 2014;10:e1004751. doi: 10.1371/journal.pgen.1004751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim et al. (2008).Kim SS, Yoo NJ, Jeong EG, Kim MS, Lee SH. Expression of NEDD4-1, a PTEN regulator, in gastric and colorectal carcinomas. APMIS. 2008;116:779–784. doi: 10.1111/j.1600-0463.2008.00999.x. [DOI] [PubMed] [Google Scholar]
- Kim et al. (2018).Kim YH, Yoo H, Hong AR, Kwon M, Kang SW, Kim K, Song Y. NEDD4L limits cAMP signaling through ubiquitination of CREB-regulated transcription coactivator 3. FASEB Journal. 2018;32:4053–4062. doi: 10.1096/fj.201701406R. [DOI] [PubMed] [Google Scholar]
- Kroeger et al. (2013).Kroeger N, Xie W, Lee JL, Bjarnason GA, Knox JJ, Mackenzie MJ, Wood L, Srinivas S, Vaishamayan UN, Rha SY, Pal SK, Yuasa T, Donskov F, Agarwal N, Kollmannsberger CK, Tan MH, North SA, Rini BI, Choueiri TK, Heng DY. Metastatic non-clear cell renal cell carcinoma treated with targeted therapy agents: characterization of survival outcome and application of the International mRCC Database Consortium criteria. Cancer. 2013;119:2999–3006. doi: 10.1002/cncr.28151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuratomi et al. (2005).Kuratomi G, Komuro A, Goto K, Shinozaki M, Miyazawa K, Miyazono K, Imamura T. NEDD4-2 (neural precursor cell expressed, developmentally down-regulated 4-2) negatively regulates TGF-beta (transforming growth factor-beta) signalling by inducing ubiquitin-mediated degradation of Smad2 and TGF-beta type I receptor. Biochemical Journal. 2005;386:461–470. doi: 10.1042/BJ20040738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang et al. (2017).Lang ZQ, Wu YQ, Pan XB, Qu GM, Zhang TG. The identification of multifocal breast cancer-associated long non-coding RNAs. European Review for Medical and Pharmacological Sciences. 2017;21:5648–5654. doi: 10.26355/eurrev_201712_14008. [DOI] [PubMed] [Google Scholar]
- Le (2019).Le NQK. Fertility-GRU: identifying Fertility-Related Proteins by Incorporating Deep-Gated Recurrent Units and Original Position-Specific Scoring Matrix Profiles. Journal of Proteome Research. 2019;18:3503–3511. doi: 10.1021/acs.jproteome.9b00411. [DOI] [PubMed] [Google Scholar]
- Le et al. (2019).Le NQK, Yapp E, Nagasundaram N, Yeh HY. Classifying promoters by interpreting the hidden information of DNA sequences via deep learning and combination of continuous fasttext N-Grams. Frontiers in Bioengineering and Biotechnology. 2019;7:305. doi: 10.3389/fbioe.2019.00305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee et al. (2020).Lee DE, Yoo JE, Kim J, Kim S, Kim S, Lee H, Cheong H. NEDD4L downregulates autophagy and cell growth by modulating ULK1 and a glutamine transporter. Cell Death & Disease. 2020;11:38. doi: 10.1038/s41419-020-2242-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li et al. (2019a).Li D, Cheng P, Wang J, Qiu X, Zhang X, Xu L, Liu Y, Qin S. IRF6 is directly regulated by ZEB1 and ELF3, and predicts a favorable prognosis in gastric cancer. Frontiers in Oncology. 2019a;9:220. doi: 10.3389/fonc.2019.00220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li et al. (2014).Li L, Liu C, Amato RJ, Chang JT, G Du, Li W. CDKL2 promotes epithelial-mesenchymal transition and breast cancer progression. Oncotarget. 2014;5:10840–10853. doi: 10.18632/oncotarget.2535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li et al. (2015).Li H, Mohamed AA, Sharad S, Umeda E, Song Y, Young D, Petrovics G, McLeod DG, Sesterhenn IA, Sreenath T, Dobi A, Srivastava S. Silencing of PMEPA1 accelerates the growth of prostate cancer cells through AR, NEDD4 and PTEN. Oncotarget. 2015;6:15137–15149. doi: 10.18632/oncotarget.3526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li et al. (2008b).Li Y, Ozaki T, Kikuchi H, Yamamoto H, Ohira M, Nakagawara A. A novel HECT-type E3 ubiquitin protein ligase NEDL1 enhances the p53-mediated apoptotic cell death in its catalytic activity-independent manner. Oncogene. 2008b;27:3700–3709. doi: 10.1038/sj.onc.1211032. [DOI] [PubMed] [Google Scholar]
- Li et al. (2008a).Li H, Xu LL, Masuda K, Raymundo E, McLeod DG, Dobi A, Srivastava S. A feedback loop between the androgen receptor and a NEDD4-binding protein, PMEPA1, in prostate cancer cells. Journal of Biological Chemistry. 2008a;283:28988–28995. doi: 10.1074/jbc.M710528200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li et al. (2019b).Li G, Xu W, Zhang L, Liu T, Jin G, Song J, Wu J, Wang Y, Chen W, Zhang C, Chen X, Ding Z, Zhu P, Zhang B. Development and validation of a CIMP-associated prognostic model for hepatocellular carcinoma. EBioMedicine. 2019b;47:128–141. doi: 10.1016/j.ebiom.2019.08.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li & Zhang (2016).Li Z, Zhang H. Reprogramming of glucose, fatty acid and amino acid metabolism for cancer progression. Cellular and Molecular Life Sciences. 2016;73:377–392. doi: 10.1007/s00018-015-2070-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez et al. (2015).Lopez MB, Garcia MN, Grasso D, Bintz J, Molejon MI, Velez G, Lomberk G, Neira JL, Urrutia R, Iovanna J. Functional characterization of nupr1l, A novel p53-regulated isoform of the high-mobility group (HMG)-related protumoral protein nupr1. Journal of Cellular Physiology. 2015;230:2936–2950. doi: 10.1002/jcp.25022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maxwell et al. (1999).Maxwell PH, Wiesener MS, Chang GW, Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER, Ratcliffe PJ. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999;399:271–275. doi: 10.1038/20459. [DOI] [PubMed] [Google Scholar]
- Nourashrafeddin et al. (2015).Nourashrafeddin S, Aarabi M, Modarressi MH, Rahmati M, Nouri M. The evaluation of WBP2NL-related genes expression in breast cancer. Pathology & Oncology Research. 2015;21:293–300. doi: 10.1007/s12253-014-9820-8. [DOI] [PubMed] [Google Scholar]
- Pavlova & Thompson (2016).Pavlova NN, Thompson CB. The emerging hallmarks of cancer metabolism. Cell Metabolism. 2016;23:27–47. doi: 10.1016/j.cmet.2015.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierorazio et al. (2016).Pierorazio PM, Johnson MH, Patel HD, Sozio SM, Sharma R, Iyoha E, Bass EB, Allaf ME. Management of renal masses and localized renal cancer: systematic review and meta-analysis. Journal D Urologie. 2016;196:989–999. doi: 10.1016/j.juro.2016.04.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reichert-Penetrat et al. (1998).Reichert-Penetrat S, Barbaud A, Martin S, Omhover L, Weber M, Schmutz JL. Pemphigus vulgaris on an old surgical scar: Koebner’s phenomenon? European Journal of Dermatology. 1998;8:60–62. [PubMed] [Google Scholar]
- Rungroj et al. (2018).Rungroj N, Nettuwakul C, Sawasdee N, Sangnual S, Deejai N, Misgar RA, Pasena A, Khositseth S, Kirdpon S, Sritippayawan S, Vasuvattakul S, Yenchitsomanus PT. Distal renal tubular acidosis caused by tryptophan-aspartate repeat domain 72 (WDR72) mutations. Clinical Genetics. 2018;94:409–418. doi: 10.1111/cge.13418. [DOI] [PubMed] [Google Scholar]
- Sakashita et al. (2013).Sakashita H, Inoue H, Akamine S, Ishida T, Inase N, Shirao K, Mori M, Mimori K. Identification of the NEDD4L gene as a prognostic marker by integrated microarray analysis of copy number and gene expression profiling in non-small cell lung cancer. Annals of Surgical Oncology. 2013;20(Suppl 3):S590–S598. doi: 10.1245/s10434-013-3059-6. [DOI] [PubMed] [Google Scholar]
- Sanchez-Gastaldo et al. (2017).Sanchez-Gastaldo A, Kempf E, Gonzalez DAA, Duran I. Systemic treatment of renal cell cancer: a comprehensive review. Cancer Treatment Reviews. 2017;60:77–89. doi: 10.1016/j.ctrv.2017.08.010. [DOI] [PubMed] [Google Scholar]
- Seles et al. (2020).Seles M, Hutterer GC, Fosselteder J, Svoboda M, Resel M, Barth DA, Pichler R, Bauernhofer T, Zigeuner RE, Pummer K, Slaby O, Klec C, Pichler M. Long non-coding RNA PANTR1 is associated with poor prognosis and influences angiogenesis and apoptosis in clear-cell renal cell cancer. Cancers (Basel) 2020;12 doi: 10.3390/cancers12051200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shao et al. (2018).Shao G, Wang R, Sun A, Wei J, Peng K, Dai Q, Yang W, Lin Q. The E3 ubiquitin ligase NEDD4 mediates cell migration signaling of EGFR in lung cancer cells. Molecular Cancer. 2018;17:24. doi: 10.1186/s12943-018-0784-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shu et al. (2018).Shu J, Li S, Chen YB, Zhu QF, Yu XH. Long non-coding RNA EPB41L4A-AS2 inhibited non-small cell lung cancer proliferation, invasion and promoted cell apoptosis. Neoplasma. 2018;65:664–672. doi: 10.4149/neo_2018_170713N480. [DOI] [PubMed] [Google Scholar]
- Siegel, Miller & Jemal (2019).Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA: A Cancer Journal for Clinicians. 2019;69:7–34. doi: 10.3322/caac.21551. [DOI] [PubMed] [Google Scholar]
- Stambergova et al. (2016).Stambergova H, Zemanova L, Lundova T, Malcekova B, Skarka A, Safr M, Wsol V. Human DHRS7, promising enzyme in metabolism of steroids and retinoids? Journal of Steroid Biochemistry and Molecular Biology. 2016;155:112–119. doi: 10.1016/j.jsbmb.2015.09.041. [DOI] [PubMed] [Google Scholar]
- Sun et al. (2014).Sun A, Yu G, Dou X, Yan X, Yang W, Lin Q. Nedd4-1 is an exceptional prognostic biomarker for gastric cardia adenocarcinoma and functionally associated with metastasis. Molecular Cancer. 2014;13:248. doi: 10.1186/1476-4598-13-248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun, Yang & Gao (2020).Sun T, Yang P, Gao Y. Long non-coding RNA EPB41L4A-AS2 suppresses progression of ovarian cancer by sequestering microRNA-103a to upregulate transcription factor RUNX1T1. ExperimentaL Physiology. 2020;105:75–87. doi: 10.1113/EP087847. [DOI] [PubMed] [Google Scholar]
- Swierczynski, Hebanowska & Sledzinski (2014).Swierczynski J, Hebanowska A, Sledzinski T. Role of abnormal lipid metabolism in development, progression, diagnosis and therapy of pancreatic cancer. World Journal of Gastroenterology. 2014;20:2279–2303. doi: 10.3748/wjg.v20.i9.2279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeuchi et al. (2011).Takeuchi T, Adachi Y, Nagayama T, Furihata M. Nedd4L modulates the transcription of metalloproteinase-1 and -13 genes to increase the invasive activity of gallbladder cancer. International Journal of Experimental Pathology. 2011;92:79–86. doi: 10.1111/j.1365-2613.2010.00740.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanksley, Chen & Coffey (2013).Tanksley JP, Chen X, Coffey RJ. NEDD4L is downregulated in colorectal cancer and inhibits canonical WNT signaling. PLOS ONE. 2013;8:e81514. doi: 10.1371/journal.pone.0081514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan et al. (2019).Wan L, Liu T, Hong Z, Pan Y, Sizemore ST, Zhang J, Ma Z. NEDD4 expression is associated with breast cancer progression and is predictive of a poor prognosis. Breast Cancer Research. 2019;21:148. doi: 10.1186/s13058-019-1236-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang et al. (2020).Wang Z, Hu X, Ye M, Lin M, Chu M, Shen X. NEDD4 E3 ligase: functions and mechanism in human cancer. Seminars in Cancer Biology. 2020;67:92–101. doi: 10.1016/j.semcancer.2020.03.006. [DOI] [PubMed] [Google Scholar]
- Wang et al. (2020).Wang K, Liu J, Zhao X, Li H, Luo G, Yu Y, Guo Y, Zhang L, Zhu J, Wang S, Hua W, Yang A, Zhang R, Li J. WWP2 regulates proliferation of gastric cancer cells in a PTEN-dependent manner. Biochemical and Biophysical Research Communications. 2020;521:652–659. doi: 10.1016/j.bbrc.2019.10.179. [DOI] [PubMed] [Google Scholar]
- Wang et al. (2017).Wang D, Ma L, Wang B, Liu J, Wei W. E3 ubiquitin ligases in cancer and implications for therapies. Cancer and Metastasis Reviews. 2017;36:683–702. doi: 10.1007/s10555-017-9703-z. [DOI] [PubMed] [Google Scholar]
- Wang et al. (2019).Wang YG, Wang T, Shi M, Zhai B. Long noncoding RNA EPB41L4A-AS2 inhibits hepatocellular carcinoma development by sponging miR-301a-5p and targeting FOXL1. Journal of Experimental & Clinical Cancer Research. 2019;38:153. doi: 10.1186/s13046-019-1128-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen et al. (2017).Wen W, Li J, Wang L, Xing Y, Li X, Ruan H, Xi X, Xiong J, Kuang R. Inhibition of NEDD4 inhibits cell growth and invasion and induces cell apoptosis in bladder cancer cells. Cell Cycle. 2017;16:1509–1514. doi: 10.1080/15384101.2017.1338220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xing et al. (2021).Xing Q, Zeng T, Liu S, Cheng H, Ma L, Wang Y. A novel 10 glycolysis-related genes signature could predict overall survival for clear cell renal cell carcinoma. BMC Cancer. 2021a;21:381. doi: 10.1186/s12885-021-08111-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu et al. (2019).Xu HF, Huang TJ, Yang Q, Xu L, Lin F, Lang YH, Hu H, Peng LX, Meng DF, Xie YJ, Tan L, Qian CN, Huang BJ. Candidate tumor suppressor gene IRF6 is involved in human breast cancer pathogenesis via modulating PI3K-regulatory subunit PIK3R2 expression. Cancer Management and Research. 2019;11:5557–5572. doi: 10.2147/CMAR.S203060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu et al. (2016).Xu S, Wang P, You Z, Meng H, Mu G, Bai X, Zhang G, Zhang J, Pang D. The long non-coding RNA EPB41L4A-AS2 inhibits tumor proliferation and is associated with favorable prognoses in breast cancer and other solid tumors. Oncotarget. 2016;7:20704–20717. doi: 10.18632/oncotarget.8007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang et al. (2015).Yang Q, Zhao J, Cui M, Gi S, Wang W, Han X. Nedd4L expression is decreased in ovarian epithelial cancer tissues compared to ovarian non-cancer tissue. Journal of Obstetrics and Gynaecology Research. 2015a;41:1959–1964. doi: 10.1111/jog.12808. [DOI] [PubMed] [Google Scholar]
- Yi et al. (2020).Yi R, Yang S, Liao Y, Hu Z, Long H, Zeng Y, Wang X, Qiu C, Xu A, Lin J, Wu Z. Decreased CDKL2 expression is correlated with the progression and poor prognosis of glioma. Pathology Research and Practice. 2020;216:152920. doi: 10.1016/j.prp.2020.152920. [DOI] [PubMed] [Google Scholar]
- Yi et al. (2018).Yi R, Yang S, Wen E, Hu Z, Long H, Zeng Y, Wang X, Huang X, Liao Y, Luo M, Wang J, Zhou M, Wang W, Xu A, Lin J, Wu Z, Song Y. Negative nuclear expression of CDKL2 correlates with disease progression and poor prognosis of glioma. International Journal of Clinical and Experimental Pathology. 2018;11:712–719. [PMC free article] [PubMed] [Google Scholar]
- Zhang, Qiu & Yang (2021).Zhang H, Qiu X, Yang G. The CSRNP gene family serves as a prognostic biomarker in clear cell renal cell carcinoma. Frontiers in Oncology. 2021;11:620126. doi: 10.3389/fonc.2021.620126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang et al. (2020).Zhang J, Zhang X, Li J, Song Z. Systematic analysis of the ABC transporter family in hepatocellular carcinoma reveals the importance of ABCB6 in regulating ferroptosis. Life Sciences. 2020;257:118131. doi: 10.1016/j.lfs.2020.118131. [DOI] [PubMed] [Google Scholar]
- Zhao et al. (2015).Zhao R, Cui T, Han C, Zhang X, He J, Srivastava AK, Yu J, Wani AA, Wang QE. DDB2 modulates TGF-beta signal transduction in human ovarian cancer cells by downregulating NEDD4L. Nucleic Acids Research. 2015;43:7838–7849. doi: 10.1093/nar/gkv667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhi & Chen (2012).Zhi X, Chen C. WWP1: a versatile ubiquitin E3 ligase in signaling and diseases. Cellular and Molecular Life Sciences. 2012;69:1425–1434. doi: 10.1007/s00018-011-0871-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou, Levy-Cohen & Blank (2015).Zou X, Levy-Cohen G, Blank M. Molecular functions of NEDD4 E3 ubiquitin ligases in cancer. Biochimica Et Biophysica Acta/General Subjects. 2015;1856:91–106. doi: 10.1016/j.bbcan.2015.06.005. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The following information was supplied regarding data availability:
The raw data are available from TCGA (project ID TCGA_KIRC) and from NCBI GSE40435 and GSE53757.