

Abstract
Cancer as one of the leading causes of human deaths has always been one of the main health challenges in the world. Despite recent advances in therapeutic and diagnostic methods, there is still a high mortality rate among cancer patients. Late diagnosis is one of the main reasons for the high ratio of cancer related deaths. Therefore, it is required to introduce novel early detection methods. Various molecular mechanisms are associated with the tumor progression and metastasis. MicroRNAs (miRNAs) are a class of non-coding RNAs (ncRNAs) family that has important functions in regulation of the cellular processes such as cell proliferation, apoptosis, and tumor progression. Moreover, they have higher stability in body fluids compared with mRNAs which can be introduced as non-invasive diagnostic markers in cancer patients. MiR-132 has important functions as tumor suppressor or oncogene in different cancers. In the present review, we have summarized all of the studies which have been reported the role of miR-132 during tumor progressions. We categorized the miR-132 target genes based on their cell and molecular functions. Although, it has been reported that the miR-132 mainly functions as a tumor suppressor, it has also oncogenic functions especially in pancreatic tumors. MiR-132 mainly exerts its roles during tumor progressions by regulation of the transcription factors and signaling pathways. Present review clarifies the tumor specific molecular mechanisms of miR-132 to introduce that as an efficient non-invasive diagnostic marker in various cancers.
Keywords: MiR-132, Cancer, Diagnosis, Prognosis, Marker
Background
Cancer is one of the main causes of human deaths worldwide, with an estimated 10.0 million deaths in 2020 [1]. It is the second leading cause of mortality in the United States with about 606,520 deaths in 2020 [2, 3]. The financial burden of cancer poses different challenges for the patients and healthcare system [4]. As the morphologically similar tumors may exhibit different clinical symptoms due to their molecular differences, it is of high importance to introduce non-invasive methods to assess the molecular differences in tumors to select the most efficient therapeutic option. As the non-protein-coding DNA covers almost 97% of the human genome, non-coding RNAs (ncRNAs) have become the frontier of cancer biology [5, 6]. They are categorized into the various families such as microRNAs (miRNAs), long noncoding RNAs (lncRNAs), small interfering RNAs (siRNAs), and circular RNA (circRNA) [7–9]. MiRNAs are a class of the short ncRNAs involved in post-transcriptional regulation through binding to 3 untranslated region (3-UTR) of the target mRNA that results in mRNA degradation or translational inhibition [10]. Considering, the crucial functions of miRNAs in regulation of cellular mechanisms including cell proliferation, differentiation, growth, and apoptosis [11, 12], aberrant miRNA expression can be correlated with various cancers [13]. MiRNAs may serve as tumor suppressors, oncogenes, and regulators of the self-renewal process in cancer stem cells (CSC) [14]. Dysregulated miRNAs are promising diagnostic tumor markers and are also efficient as novel targets for the cancer therapy [15]. Since, they have higher stability in body fluids compared with mRNAs, the expression profiling of circulating miRNAs in body fluids can be utilized as a non-invasive method for cancer diagnosis and prognosis [16–20]. MiR-132 is a critical regulator of various cellular processes such as angiogenesis, cell proliferation, migration, and apoptosis [21–23]. Aberrant expression of miR-132 has been frequently reported in various cancers. It functions as a tumor suppressor or oncogene in different cancers [24–27]. Therefore, we have summarized all of the studies which have been reported the role of miR-132 during tumor progressions. We categorized the miR-132 target genes based on their cell and molecular functions (Table 1).
Table 1.
Molecular targets of miR-132 during tumor progressions
| Study | Year | Type | Gene | Target | Samples | MiR-132 function |
|---|---|---|---|---|---|---|
| Zhang et al. [23] | 2014 | Breast | miR-132 | HN1 |
10 NT* NMuMG, 4T1, MDA-MB-231, and MCF10A cell lines |
Tumor suppressor |
| Lian et al. [119] | 2016 | Laryngeal | miR-132 | FOXO1 |
10 NT Hep-G2 and AMC-HN-8 cell lines |
Oncogene |
| Han et al. [132] | 2020 | Retinoblastoma | ILF3-AS1 | miR-132 |
50 NT Y79, HXO-RB44, SO-RB50, and RB1 cell lines |
Tumor suppressor |
| Zhang et al. [129] | 2020 | Hepatocellular | LINC00160 | miR-132 |
68 NT HCCLM3, Huh7, Hep3B, and MHCC97 cell lines |
Tumor suppressor |
| Zhang et al. [123] | 2019 | Pancreatic | miR-132 | PTEN |
60 NT PAN-1, KLM-1, and PaCa-2 cell lines |
Oncogene |
| Renjie et al. [45] | 2015 | Pituitary | miR-132 | SOX5 |
16 T MMQ and GH3 cell lines |
Tumor suppressor |
| Xie et al. [124] | 2018 | Breast | miR-132 | PTEN |
53 NT MCF-7 cell line |
Oncogene |
| Chen et al. [94] | 2016 | Glioma | miR-132 | TTK |
46 T and 9 N U87 cell line |
Tumor suppressor |
| Song et al. [140] | 2017 | Colorectal | XIST | miR-132 |
50 NT SW480, SW620, LOVO,HT29, and HCT116 cell lines |
Tumor suppressor |
| Li et al. [68] | 2016 | Glioma | miR-132 | SIRT1 | U251 and U87 cell lines | Tumor suppressor |
| Zhao et al. [144] | 2019 | Pancreatic | miR-132 | SHH |
23 T and 25 N MiaPaCe-2a cell line |
Oncogene |
| Zhang et al. [79] | 2019 | Ovarian | miR-132 | BMI1 | SKOV3 | Tumor suppressor |
| Xue et al. [43] | 2020 | Nasopharyngeal | LINC01551 | miR-132 |
24 NT HNE1, SUNE2, HONE1, CNE2, and 6-10B cell lines |
Tumor suppressor |
| Liu et al. [89] | 2018 | Colorectal | MIAT | miR-132 |
30 NT Ht29, SW480, and LOVO cell lines |
Tumor suppressor |
| Zhou et al. [51] | 2018 | Glioma | NEAT1 | miR-132 |
14 T and 5 N U87, U251, SHG-44, and U-118MG cell lines |
Tumor suppressor |
| Liu et al. [42] | 2020 | Pancreatic | PTTG3P | miR-132 |
60 NT AsPc-1, BxPC-3, CaPAN-2, MiaPaCa-2, PANC-1, and SW1990 cell lines |
Oncogene |
| Chen et al. [36] | 2019 | Thyroid | miR-132 | FOXA1 |
30 NT TPC1 and GLAG-66 cell lines |
Tumor suppressor |
| Tian et al. [64] | 2016 | Ovarian | miR-132 | E2F5 |
32 NT SKOV3, OVCAR3, and A2780 cell lines |
Tumor suppressor |
| Guo et al. [84] | 2018 | Lung | miR-132 | USP9X | A549 and NCI-1299 cell lines | Tumor suppressor |
| Wang et al. [35] | 2018 | Breast | miR-132 | FOXA1 |
30 NT SK-BR3 and MDA-MB-468 cell lines |
Tumor suppressor |
| Li et al. [128] | 2019 | Breast | miR-132 | LAPTM4B |
131 T and 87 N MCF-7, MCF-10A, ZR-75–1, T470, and MDA-MB-231 cell lines |
Tumor suppressor |
| Lei et al. [147] | 2015 | Hepatocellular | miR-132 | YAP | Huh7 and HepG2 cell lines | Tumor suppressor |
| Liu et al. [49] | 2019 | Bladder | CIRC-DOCK1 | miR-132 |
23 T and 32 N BIU-87, EJ-m3, T24, and 5673 cell lines |
Tumor suppressor |
| Qu et al. [108] | 2016 | Prostate | miR-132 | GLUT1 | PC-3 and DU-145 cell lines | Tumor suppressor |
| Geng et al. [112] | 2016 | Astrocytoma | miR-132 | PEA15 | U251 and U87 cell lines | Tumor suppressor |
| Chen et al. [126] | 2018 | Thyroid | miR-132 | CSDE1 | BCPAP, TPC1, and 8505c cell lines | Tumor suppressor |
| Cheng et al. [90] | 2017 | Glioblastoma | miR-132 | TUSC3 | U87MG | Oncogene |
| Liu et al. [58] | 2015 | Osteosarcoma | miR-132 | SOX4 | MG63, HOS, 143B, U2OS, and SaOS-2 cell lines | Tumor suppressor |
| Abukiwan et al. [136] | 2019 | Pancreatic | miR-132 | TGFβ |
35 NT AsPC-1 and PANC-1 cell lines |
Oncogene |
| Zhao et al. [130] | 2015 | Cervical | miR-132 | SMAD2 |
20 NT HeLa and C33A cell lines |
Tumor suppressor |
| Li et al. [61] | 2015 | Lung | miR-132 | SOX4 | H460, A549, and YTMLC-9 cell lines | Tumor suppressor |
| Chen et al. [133] | 2020 | Oral | miR-132 | TGFβ |
37 NT SCC-9 and CAL-27 cell lines |
Tumor suppressor |
| Zhang et al. [70] | 2019 | Colorectal | SNHG5 | miR-132 |
25 NT RKO, SW480, and LOVO cell lines |
Tumor suppressor |
| Li et al. [30] | 2015 | Gastric | miR-132 | FOXO1 |
28 NT AGS and SNU-5 cell lines |
Oncogene |
| Lin et al. [74] | 2016 | Ovarian | miR-132 | CDH2, VIM | SKOV3 and OV2008 cell lines | Tumor suppressor |
| Liu et al. [139] | 2019 | Colorectal | miR-132 | ERK1 | NCM460, LOVO, and SW480 cell lines | Tumor suppressor |
| Liu et al. [102] | 2017 | Gastric | miR-132 | CD44, FN1 |
201 NT BGC823, AGS, and HGC27 cell lines |
Tumor suppressor |
| He et al. [107] | 2017 | Gastric | miR-132 | MUC13 |
40 NT MKN28 cell line |
Tumor suppressor |
| Huang et al. [62] | 2020 | Hepatocellular | miR-132 | SOX4 | HepG2, Huh7, and HccLM3 cell lines | Tumor suppressor |
| Liu et al. [78] | 2017 | Cervical | miR-132 | BMI1 |
104 NT HeLa, SiHa, and C33A cell lines |
Tumor suppressor |
| Zhang et al. [80] | 2018 | Lung | SOX2OT | miR-132 |
48 NT A549, H1299, NCI-H460, and HCC-827 cell lines |
Tumor suppressor |
| Wei et al. [131] | 2019 | Bladder | miR-132 | SMAD2 |
32 NT T24 cell line |
Tumor suppressor |
| He et al. [86] | 2020 | Colorectal | SNHG16 | miR-132 |
50 NT SW480 and SW620 cell lines |
Tumor suppressor |
| Fu et al. [60] | 2016 | Prostate | miR-132 | SOX4 |
57 NT LnCap and VCap cell lines |
Tumor suppressor |
| Zheng et al. [81] | 2014 | Colorectal | miR-132 | ZEB2 |
62 NT HT29, LOVO, HTC116, SW480, and SW620 cell lines |
Tumor suppressor |
| Wang et al. [113] | 2014 | Osteosarcoma | miR-132 | CCNE1 |
10 NT HOS, MG63, 143B, and Saos-2 cell lines |
Tumor suppressor |
| Li et al. [32] | 2016 | Nasopharyngeal | miR-132 | FOXA1 | CNE2 | Tumor suppressor |
* Tumor (T) tissues and Normal (N) margins
Transcription factors
Forkhead box proteins (Fox) transcription factors
There are increasing numbers of the feedback loop interactions between transcription factors and miRNAs in which the transcription factors up or down regulate the miRNAs, while the miRNAs inhibit the transcription factors in a negative feedback [28]. FOXO1 belongs to the Forkhead box proteins (Fox) transcription factors that functions as a negative regulator of cell cycle progression [29]. It has been shown that miR-132 significantly promoted gastric tumor cell growth by FOXO1 targeting. There was also significant miR-132 up regulation in gastric cancer (GC) tissues in comparison with normal margins [30]. Forkhead box protein A1 (FOXA1) is a pivotal transcription factor involved in cell proliferation, apoptosis, and differentiation, organogenesis, and tumor progression [31, 32]. It is required for the chromatin recruitment of estrogen receptor that regulates chromatin remodeling, estrogen receptor-related gene expressions, and tumor cell proliferation [33, 34]. It has been shown that there was an inverse correlation between the levels of miR-132 and FOXA1 expressions. MiR-132 reduced the breast tumor cells proliferation via FOXA1 targeting [35]. There was also miR-132 down regulation in thyroid tumor tissues and cell lines. It reduced thyroid tumor cell proliferation and invasion by FOXA1 inhibition [36]. Cisplatin (CDPP) is one of the main therapeutic drugs in nasopharyngeal carcinoma (NPC), however there is a noticeable ratio of resistance among the patients [37]. It has been reported that there was miR-132 down regulation in NPC patients. It also induced CDDP sensitivity in NPC cells through FOXA1 suppression [32]. Long non-coding RNAs (lncRNAs) are a family of the ncRNAs that regulate cell growth and tumorigenesis by post-transcriptional regulation and miRNAs sponging [38]. They are involved in tumorigenesis, tissue development, embryogenesis, and inflammation [39–41]. Pseudogene belongs to the lncRNAs family that regulates the gene expression during tumor progressions. PTTG3P is a pseudogene that is up regulated in pancreatic ductal adenocarcinoma (PDAC) tissues. It has been observed that there were correlations between the PTTG3P up regulation, larger tumor size, poor prognosis, and poor differentiation in PDAC tissues. PTTG3P induced tumor growth and invasion through miR-132-3p sponging that resulted in FOXM1 targeting [42]. It has been observed that there were LINC01551 up regulation in NPC tissues and cells. LINC01551 induced malignant transformation of NPC by miR-132-5p sponging [43].
Developmental transcription factors
SOX5 belongs to the Sox family of developmental transcription factors involved in regulation of embryogenesis, cell differentiation, proliferation, and migration [44]. There were miR-132 down regulations in invasive pituitary tumor tissues and cell lines. It reduced cell proliferation and invasion through SOX5 suppression [45]. Circular RNAs (CircRNAs) are endogenous RNAs characterized by closed continuous loops without polyadenylated tail [46]. They are involved in different cellular mechanisms such as chromatin remodeling, cell proliferation, apoptosis, invasion, and differentiation [47, 48]. It has been observed that there was circDOCK1 up regulation in bladder cancer (BCa) cells. CircDOCK1 induced cell proliferation and migration by miR‐132‐3p sponging that resulted in SOX5 up regulation [49]. SOX2 is a developmental transcription factor that participates in self-renewal process and tumor progression [50]. NEAT1 sponged miR-132 to up regulate SOX2 in glioma cells [51]. Epithelial-mesenchymal transition (EMT) is a pivotal process during tumor progression in which the tumor cells lose their epithelial feature and cell–cell adhesion to gain mesenchymal feature with high migratory and invasive properties [52–54]. EMT is orchestrated by various structural factors such as CDH1 and VIM that are regulated by EMT-related transcription factors including SNAI1, SNAI2, and TWIST [55–57]. SOX4 is a developmental transcription factor with critical functions during embryogenesis and tumorigenesis. It has been shown that miR-132 reduced osteosarcoma (OS) cell proliferation and EMT via SOX4 targeting. There was a miR-132 down regulation in OS cell lines in comparison with normal cells. It also regulated apoptosis by BCL-2 targeting. Moreover, miR-132 significantly inhibited OS invasion by CDH1 up regulation, while down regulation of the mesenchymal factors such as CDH2 and VIM [58]. SOX4 has a critical role in promotion of EMT process during the prostate cancer (PCa) progression [59]. There was a significant association between miR-132 down regulation, high Gleason score, and distant metastasis. MiR-132 inhibited prostate tumor cell migration, colony formation, and TGF-b-induced EMT by SOX4 targeting [60]. Other studies have been reported that miR-132-3p inhibited the lung and liver tumor cells invasions by SOX4 targeting [61, 62]. E2F5 belongs to the E2F family of transcription factors that regulate cell cycle progression [63]. It has been observed that there were significant miR-132 down regulation in ovarian tumor tissues and cell lines. It suppressed ovarian tumor cell proliferation and invasion via E2F5 targeting [64]. SIRT1 is an NAD dependent deacetylase that regulates cell death in oxidative and genotoxic stresses [65, 66]. SREBP is a leucine zipper transcription factor involved in cholesterogenesis and lipogenesis [67]. It has been observed that miR-132 reduced glioma cell proliferation by down regulations of SIRT1 and SREBP-1c [68]. CAMP-responsive element binding protein 5 (CREB5) is a zinc-finger DNA-binding protein with pivotal functions in cell proliferation and differentiation [69]. There was a significant SNHG5 up regulation in colorectal cancer (CRC) cells. It induced CRC invasion, while inhibited apoptosis through CREB5 up regulation following the miR-132-3p sponging [70].
EMT-related transcription factors
Enhancer of zeste homolog 2 (EZH2) is one of the components of Polycomb repressor complex 2 (PRC2) that is involved in DNA methylation using DNA methyl transferases (DNMTs) recruitment [71]. It has a pivotal role in epigenetic silencing by catalyzing the H3K27me3 in promoter sequences [72]. EZH2 is also up regulated by various transcription factors like SOX4 [73]. It has been reported that the SOX4/EZH2 complex induced H3K27me3 in miR-132 promoter sequence. MiR-132 reduced EMT process in ovarian tumor cells by CDH1 up regulation, while CDH2 and VIM down regulations. Therefore, SOX4 was suggested as the effector of miR-132 during EMT regulation in ovarian cancer (OC) [74]. BMI-1 is a ring finger component of PRC1 complex involved in epigenetic suppression [75]. It is an epigenetic modification protein involved in CSC self-renewal, tumor progression, and metastasis [21]. There were correlations between the BMI-1 up regulation, poor prognosis, increased invasion, and radio resistance [76, 77]. It was observed that there was miR-132 down regulation in cervical cancer. There was also a direct association between the levels of miR-132 expressions and radiation intensity. MiR-132 increased radio sensitivity through BMI-1 targeting [78]. It has been shown that there was significant miR-132 down regulation in SKOV3/CDDP cells compared with maternal SKOV3 cells. Reduced levels of miR-132 induced the CDDP resistance in ovarian tumor cells via BMI-1 targeting and subsequent apoptosis inhibition [79]. ZEB2 is a zinc finger transcription factor that functions as a transcriptional co-repressor via R-SMADs binding. There were significant SOX2OT up regulations in Non-small-cell lung carcinoma (NSCLC) tissues and cell lines. SOX2OT silencing significantly reduced cell proliferation, invasion, and EMT process by miR-132 sponging that resulted in ZEB2 up regulation [80]. There was also a significant miR-132 down-regulation in metastatic CRC tissues in comparison with non-metastatic tumor tissues. It reduced the CRC invasion and EMT process via ZEB2 targeting. The levels of miR-132 expressions were inversely correlated with stage, tumor size, survival, and distant metastasis in CRC patients [81].
Structural factors
USP9X belongs to the ubiquitin-specific peptidase (USP) family involved in various cellular processes via deubiquitinaton and stabilization of target proteins. USP9X up regulation is associated with tumor cell proliferation, drug resistance, and invasion [82]. It also deubiquitinates the MCL1 as an anti-apoptotic factor to suppress cell death in NSCLC [83]. It has been reported that miR-132 reduced NSCLC invasion via USP9X targeting [84]. USP22 belongs to the deubiquitinating enzyme (DUB) family of proteins involved in tumor relapse and progression [85]. USP22 silencing inhibits the tumor cell proliferation [76]. It has been reported that SNHG16 induced colorectal tumor cell proliferation and invasion through miR-132-3p sponging and subsequent USP22 up regulation [86]. HN1 promotes the ubiquitin-related degradation of b-catenin that results in loss of CDH1 interaction, actin organization, and cell migration [87]. It has been reported that there was miR-132 down regulation in breast cancer (BC) tissues in comparison with normal margins. MiR-132 significantly inhibited BC cell proliferation and metastasis through HN1 targeting. There was also a direct association between the levels of HN1 expression and poor survival in BC patients [23].
Derlin1 belongs to the derlin protein family that participates in endoplasmic reticulum (ER)-related degradation of misfolded proteins. It mediates retro translocation of misfolded proteins from ER to cytoplasm for the proteasomal degradation. Myocardial infarction associated transcript (MIAT) is an lncRNA associated with various human disorders such as diabetes and cancer [88]. There were significant MIAT up regulations in CRC tissues and cells. Silencing of MIAT promoted apoptosis, while suppressed CRC invasion. MIAT induced CRC cell proliferation and invasion through miR-132 sponging that resulted in Derlin-1 up regulation [89]. Tumor suppressor candidate 3 (TUSC3) is a component of the oligosaccharyl transferase complex involved in regulation of the N-linked protein glycosylation. It is a tumor suppressor frequently down regulated in different cancers. It has been reported that miR-132 promoted temozolomide resistance and glioblastoma initiating cells (GICs) phenotype formation by TUSC3 targeting in glioblastoma (GBM). TUSC3 also significantly down regulated the STAT3 and MDM2, while up regulate p53 [90].
TTK is a pivotal dual specificity kinase during mitotic checkpoint, centrosome duplication, and chromosome stability [91]. It induces cell proliferation and migration via AKT activation [92]. HLF is a transcription factor involved in resistance toward oxidative stress-induced apoptosis [93]. It has been reported that there were miR-132 down regulations in glioma tissues and cell lines that were associated with advanced tumor grades. HLF-mediated miR-132 inhibited glioma cell invasion and radio resistance via TTK inhibition [94]. P21-activated kinase 1 (Pak1) is a serine/threonine kinase that has key functions in cell migration, apoptosis, and neoplastic transformation [95, 96]. It regulates various cellular processes such as tumor cell invasion, drug resistance, angiogenesis, and EMT [97]. It exerts its oncogenic function by preventing apoptosis using different cascades including FOXO1, CLL/BCL-2, or DLC1 [98, 99]. ATF2 belongs to the b-ZIP family of transcription factors that regulates cellular differentiation and survival [100]. FN1 is an extracellular matrix glycoprotein involved in angiogenesis and tumor cell invasion [101]. It has been observed that miR-132 affected the hematogenous metastasis in GC. PAK1 down regulated the miR-132 via phosphorylation of ATF2 that prevents ATF2 to enter to the nucleus where it functions as an inducer of miR-132 expression. MiR-132 also reduced the levels of CD44 and FN1 expressions to promote lymphocytic mediated apoptosis of tumor cells. There were significant miR-132 down regulations in GC tissues that were associated with hematogenous metastasis. ATF2 up regulated the miR-132 that subsequently regulated the CD44/FN1/SIRT1/BDNF axis to recruit lymphocytes to suppress hematogenous metastasis in GC [102]. Receptor tyrosine kinases (RTKs) are the cell surface receptors for many extracellular signals such as hormones and growth factors. Aberrant RTK activation is implicated in progression of different tumors [103, 104]. MUC13 is a trans-membrane mucin associated with abnormal cell proliferation and tumor growth [105]. It activates the HER2, ERK, and AKT, while suppresses p53 expression [106]. It has been reported that there was a significant MUC13 up regulation in GC tissues in comparison with normal margins. MiR-132-3p suppressed GC progression by MUC13 targeting that resulted in activation of HER2 signaling [107].
Glucose transporter 1 (GLUT1) is a glucose uniporter across the erythrocytes plasma membranes. It has been shown that there was significant miR-132 down regulation in prostate tumor cells. MiR-132 silencing promoted the cell proliferation by induced glycolysis following the GLUT1 up regulation [108]. PEA-15 is an anti-apoptotic factor involved in TRAIL resistance of tumor cells. PEA15 over expression has been reported in GBM, leukemia, and NSCLC patients who were resistant against TRAIL [109–111]. MiR-132 reduced tumor cell proliferation and invasion, while increased apoptosis by targeting PEA-15 in astrocytoma. It was also observed that the miR-132 was regulated by CREB and KLF transcription factors [112]. Cyclin E1 (CCNE1) belongs to the cyclin family of proteins that regulates cyclin-dependent kinase 2 (CDK2) during cell cycle G1/S transition. It has been observed that there were miR-132 down regulations in OS tissues compared with normal bone tissues. MiR-132 reduced OS cell proliferation, colony formation, and in vivo growth via CCNE1 targeting [113].
PI3K/AKT pathway
The PI3K/AKT is an important signaling pathway that transfers the extracellular signals such as growth factors and hormones into the cells to regulate cell proliferation, metabolism, and apoptosis. PI3K activation by the RTKs and G-protein coupled receptors (GPCRs) subsequently phosphorylates and activates the AKT (Fig. 1). AKT is a serine/threonine kinase that has various effectors including CREB, FOXO, and mTOR [114, 115]. FOXO1 phosphorylation by AKT results in nuclear export and proteasome-dependent degradation [116]. It is a transcriptional regulator of apoptosis and CDK inhibitors such as BIM, FASL, p27, and p21 that inhibit G1/S transition and promote apoptosis [117, 118]. It has been reported that there was a significant miR-132 up regulation in laryngeal squamous cell carcinoma (LSCC) cells. MiR-132 promoted LSCC cell proliferation and tumor growth by PI3K/AKT activation and FOXO1 targeting [119]. PTEN as a tyrosine phosphatase inhibits the PI3K/AKT signaling by PIP3 dephosphorylation that results in AKT inhibition [120]. Therefore, PTEN down regulation activates the AKT/ERK pathway to regulate tumor cell proliferation and invasion. PTEN up regulation also promotes tumor cells apoptosis [121]. Moreover, it is a potent regulator of EMT progression [122]. It has been reported that there was miR-132 up regulation in pancreatic carcinoma that was associated with poor prognosis. MiR-132 reduced cell invasion and proliferation of pancreatic tumor cells through PTEN targeting [123]. MiR-132 increased doxorubicin resistance of BC cells through PTEN targeting [124]. Cold shock domain containing E1 (CSDE1) is an RNA binding protein (RBP) that is involved in tumor progression [125]. It has been observed that CSDE1 reduced thyroid tumor cell proliferation. CSDE1 down regulated the PTEN that resulted in AKT activation. MiR-132 also targeted the CSDE1 in thyroid tumor cells [126]. LAPTM4B is an inducer of tumor cell proliferation, invasion, and drug resistant by activation of PI3K/AKT pathway [127]. There were correlations between miR‐132‐3p down regulation, TNM staging, and tumor relapse in BC patients in which the patients with stage II/III had lower levels of miR-132-3p expressions compared with patients with stage I, and patients with recurrence had significantly lower levels of miR-132-3p expression. MiR‐132‐3p suppressed the breast tumor cell proliferation and invasion through LAPTM4B inhibition that resulted in inhibition of the PI3K/AKT/mTOR axis [128]. PIK3R3 is the regulatory subunit of the PI3K that phosphorylates phosphatidylinositol as a second messenger in intracellular signal transductions. It binds to the activated tyrosine kinases by SH2 domains to exert its functions. It has been reported that the LINC00160 knock down reduced the levels of PIK3R3 through miR-132 up regulation that resulted in reduced hepatocellular carcinoma (HCC) tumor cell drug resistance. There were also LINC00160 and PIK3R3 up regulations in HCC tissues. LINC00160 sponged the miR-132 to up regulate PIK3R3. LINC00160 silencing inhibited the HCC cell autophagy and proliferation, while induced apoptosis through PIK3R3 and ATG5 down regulations via promotion of miR-132 [129].
Fig. 1.
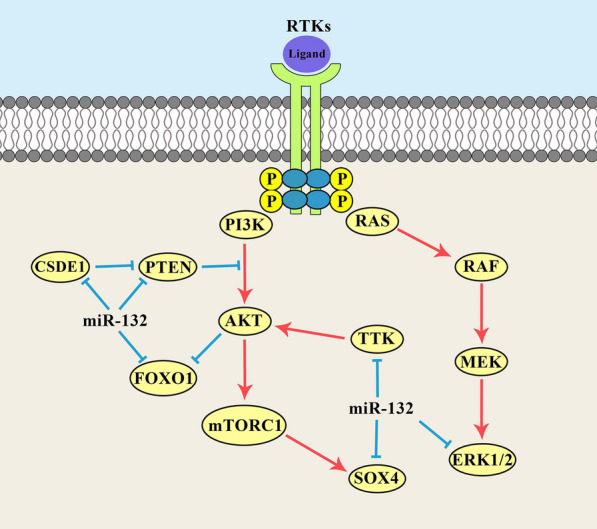
Molecular mechanisms of miR-132 in regulation of PI3K/AKT and MAPK and signaling pathways during tumor progression
TGF-β pathway
Transforming growth factor b (TGF-β) is a secreted multi-faceted cytokine involved in regulation of embryogenesis, apoptosis, inflammation, and tissue homeostasis using SMAD family of transducer proteins. It triggers and maintains the EMT process by promotion of signaling pathways and transcription factors. Both SMAD-dependent and independent cascades are recruited by TGFβ to induce EMT during tumor progression. It has been reported that there was miR-132 down regulation in cervical cancer samples. MiR-132 reduced cervical tumor cell growth and invasion by SMAD2 targeting that resulted in EMT and cell cycle regulations. MiR-132 silencing promoted EMT via CDH1 down regulation, while VIM, FN1, SNAI1, SNAI2, and TWIST2 up regulations [130]. A significant miR-132 down regulation was also observed in BC tissues with metastatic lymph nodes. MiR-132 silencing promoted the breast tumor cell invasion and increased the levels of EMT-related markers and TGFβ1/SMAD2 expressions. There was an inverse association between SMAD2 and miR-132 expression levels in BC tissues. MiR-132 inhibited the EMT by down regulations of CDH2, ZEB1, SNAI1, and VIM in BC cells. It regulated the EMT process through TGFβ1/SMAD2 signaling pathway [131]. It has been observed that there was ILF3-AS1 up regulation in retinoblastoma (RB) tissues compared with normal controls. Levels of ILF3-AS1 expressions were directly correlated with advanced stage and optic nerve metastasis. ILF3-AS1 silencing significantly decreased malignant behaviors and in vivo tumor growth. ILF3-AS1 promoted RB progression through miR-132–3p sponging that up regulated the SMAD2 [132]. MiR-132 was reported to increase cisplatin sensitivity in Oral squamous cell carcinoma (OSCC) cells. There was also significant TGFβ1 up regulation in OSCC tissues that was conversely associated with miR-132 expression. MiR-132 also reduced OSCC cell proliferation and invasion by targeting the TGFβ1/SMAD2-3 axis [133]. Glucocorticoids are a class of corticosteroids with therapeutic values in lymphoid cancer, however some of the patients are insensitive to this treatment option [134]. Dexamethasone (DEX) is a glucocorticoid medication of tumor progression that promotes EMT and self-renewal via activation of the JNK and TGFβ pathways [135]. It has been observed that the DEX was involved in regulation of miR-132 promoter methylation. MiR-132 increased pancreatic tumor cell clonogenicity and EMT through TGFβ regulation [136].
Other signaling pathways
Mitogen-activated protein kinase (MAPK) signaling pathway is categorized to the ERK, JNK, and p38 cascades in mammalian cells which are involved in regulation of stress responses, cell proliferation, and differentiation. This signaling pathway transmits the extracellular signals via a sequential activation of MAP4K, MAP3K, and MAPKAPK. JNK and p38 are mainly activated in stress response, while the ERK1/2 are associated with cell proliferation and differentiation [137]. ERK1 is involved in tumor relapse, invasion, and drug resistance [138]. It can be regulated by the miR-132 during tumor progressions (Fig. 1). MiR-132 suppressed CRC cell proliferation and Adriamycin (ADM) resistance, while promoted apoptosis through ERK1 targeting [139]. There were XIST up regulations in CRC tissues and cells that were directly associated with TNM stage and tumor size. XIST induced colorectal tumor cell proliferation via the miR-132-3p/ERK2 axis [140]. Hedgehog (Hh) is a developmental signaling pathway involved in cell differentiation and embryogenesis. It is activated by Hh ligands binding with PTCH receptor that results in activation of GLI transcription factors [141]. Aberrant Shh activation induces the cell proliferation by Myc, PTCH, and CCND1 up regulations [142, 143]. It has been reported that miR-132 increased pancreatic tumor cell proliferation via Hh pathway [144]. Hippo signaling is involved in regulation of organ volume by the maintenance of cell proliferation/apoptosis balance [145, 146]. Yes-associated protein (YAP) is one of the key effectors of Hippo signaling pathway which has a pivotal function in induction of cell proliferation and invasion, while apoptosis suppression. It has been reported that miR-132 induced hepatoma cell apoptosis, while suppressed their proliferation and invasion through YAP targeting [147].
Conclusions
In present review we summarized all of the studies that have been evaluated the role of miR-132 in different cancers. This review clarifies the cell and molecular mechanisms that are regulated by miR-132 during tumor progressions. It has been reported that the miR-132 mainly functions as a tumor suppressor; it has also oncogenic functions especially in pancreatic tumors. It mainly exerts its roles during tumor progressions by regulation of the transcription factors and signaling pathways. Present review clarifies the tumor specific molecular mechanisms of miR-132 to introduce that as an efficient non-invasive diagnostic marker in various cancers.
Acknowledgements
Not applicable.
Abbreviations
- miRNAs
MicroRNAs
- ncRNAs
Non-coding RNAs
- lncRNAs
Long noncoding RNAs
- siRNAs
Small interfering RNAs
- circRNA
Circular RNA
- CSC
Cancer stem cells
- Fox
Forkhead box proteins
- FOXA1
Forkhead box protein A1
- NPC
Nasopharyngeal carcinoma
- BCa
Bladder cancer
- EMT
Epithelial-mesenchymal transition
- OS
Osteosarcoma
- PCa
Prostate cancer
- CREB5
CAMP-responsive element binding protein 5
- CRC
Colorectal cancer
- EZH2
Enhancer of zeste homolog 2
- PRC2
Polycomb repressor complex 2
- DNMTs
DNA methyl transferases
- OC
Ovarian cancer
- NSCLC
Non-small-cell lung carcinoma
- USP
Ubiquitin-specific peptidase
- MIAT
Myocardial infarction associated transcript
- TUSC3
Tumor suppressor candidate 3
- GICs
Glioblastoma initiating cells
- GBM
Glioblastoma
- Pak1
P21-activated kinase 1
- GLUT1
Glucose transporter 1
- CCNE1
Cyclin E1
- CDK2
Cyclin-dependent kinase 2
- GPCRs
G-protein coupled receptors
- LSCC
Laryngeal squamous cell carcinoma
- CSDE1
Cold shock domain containing E1
- RBP
RNA binding protein
- HCC
Hepatocellular carcinoma
- TGF-β
Transforming growth factor b
- RB
Retinoblastoma
- OSCC
Oral squamous cell carcinoma
- DEX
Dexamethasone
- MAPK
Mitogen-activated protein kinase
- Hh
Hedgehog
- YAP
Yes-associated protein
- CDPP
Cisplatin
- ADM
Adriamycin
Authors' contributions
ASZ, ZNN, and NT were involved in search strategy and drafting. MM supervised the project and revised and edited the manuscript. All authors read and approved the final manuscript.
Funding
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Meysam Moghbeli, Email: moghbelim@mums.ac.ir, Email: Meysam_moghbeli@yahoo.com.
Amir Sadra Zangouei, Email: zangoueias961@mums.ac.ir.
Zahra Nasrpour Navaii, Email: Zhr.nasrpour@gmail.com.
Negin Taghehchian, Email: n_taghehchian@yahoo.com.
References
- 1.Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;17(3):209–49. doi: 10.3322/caac.21660. [DOI] [PubMed] [Google Scholar]
- 2.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020;70(1):7–30. doi: 10.3322/caac.21590. [DOI] [PubMed] [Google Scholar]
- 3.Park J, Look KA. Health care expenditure burden of cancer care in the United States. INQUIRY J Health Care Org Provis Financing. 2019;56:0046958019880696. doi: 10.1177/0046958019880696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Altice CK, Banegas MP, Tucker-Seeley RD, Yabroff KR. Financial hardships experienced by cancer survivors: a systematic review. J Natl Cancer Inst. 2017;109(2):djw205. doi: 10.1093/jnci/djw205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Grillone K, Riillo C, Scionti F, Rocca R, Tradigo G, Guzzi PH, et al. Non-coding RNAs in cancer: platforms and strategies for investigating the genomic “dark matter”. 2020;39(1):1–19. [DOI] [PMC free article] [PubMed]
- 6.Zangouei AS, Rahimi HR, Mojarrad M, Moghbeli M. Non coding RNAs as the critical factors in chemo resistance of bladder tumor cells. Diagn Pathol. 2020;15(1):136. doi: 10.1186/s13000-020-01054-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Guttman M, Rinn JLJN. Modular regulatory principles of large non-coding RNAs. Nature. 2012;482(7385):339–346. doi: 10.1038/nature10887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Esteller MJ. Non-coding RNAs in human disease. Nat Genet Res. 2011;12(12):861–74. doi: 10.1038/nrg3074. [DOI] [PubMed] [Google Scholar]
- 9.Vo JN, Zhang Y, Shukla S, Xiao L, Robinson D, Wu Y-M, et al. The landscape of circular RNA in cancer. Cell. 2018;176(4):869–8. doi: 10.1016/j.cell.2018.12.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Moreno-Moya JM, Vilella F, Simon C. MicroRNA: key gene expression regulators. Fertil Steril. 2014;101(6):1516–1523. doi: 10.1016/j.fertnstert.2013.10.042. [DOI] [PubMed] [Google Scholar]
- 11.Peláez N, Carthew RW. Biological robustness and the role of microRNAs: a network perspective. Curr Top Dev Biol. 2012;99:237–255. doi: 10.1016/B978-0-12-387038-4.00009-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pasquinelli AE. MicroRNAs and their targets: recognition, regulation and an emerging reciprocal relationship. Nat Rev Genet. 2012;13(4):271–82. doi: 10.1038/nrg3162. [DOI] [PubMed] [Google Scholar]
- 13.Shenouda SK, Alahari SK. MicroRNA function in cancer: oncogene or a tumor suppressor? Cancer Metastasis Rev. 2009;28(3):369–78. doi: 10.1007/s10555-009-9188-5. [DOI] [PubMed] [Google Scholar]
- 14.Cho WC. MicroRNAs: potential biomarkers for cancer diagnosis, prognosis and targets for therapy. Int J Biochem Cell Biol. 2010;42(8):1273–81. doi: 10.1016/j.biocel.2009.12.014. [DOI] [PubMed] [Google Scholar]
- 15.Hayes J, Peruzzi PP, Lawler S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Mol Med. 2014;20(8):460–469. doi: 10.1016/j.molmed.2014.06.005. [DOI] [PubMed] [Google Scholar]
- 16.Kosaka N, Iguchi H, Ochiya T. Circulating microRNA in body fluid: a new potential biomarker for cancer diagnosis and prognosis. Cancer Sci. 2010;101(10):2087–92. doi: 10.1111/j.1349-7006.2010.01650.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Weber JA, Baxter DH, Zhang S, Huang DY, How Huang K, Jen Lee M, et al. The microRNA spectrum in 12 body fluids. Clin Chem. 2010;56(11):1733–41. doi: 10.1373/clinchem.2010.147405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gablo NA, Prochazka V, Kala Z, Slaby O, Kiss I. Cell-free microRNAs as non-invasive diagnostic and prognostic biomarkers in pancreatic cancer. Curr Genomics. 2019;20(8):569–80. doi: 10.2174/1389202921666191217095017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Imaoka H, Toiyama Y, Fujikawa H, Hiro J, Saigusa S, Tanaka K, et al. Circulating microRNA-1290 as a novel diagnostic and prognostic biomarker in human colorectal cancer. Ann Oncol. 2016;27(10):1879–86. doi: 10.1093/annonc/mdw279. [DOI] [PubMed] [Google Scholar]
- 20.Jin Y, Wong YS, Goh BK, Chan CY, Cheow PC, Chow PK, et al. Circulating microRNAs as potential diagnostic and prognostic biomarkers in hepatocellular carcinoma. Sci Rep. 2019;9(1):1–12. doi: 10.1038/s41598-019-46872-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li H, Song F, Chen X, Li Y, Fan J, Wu X. Bmi-1 regulates epithelial-to-mesenchymal transition to promote migration and invasion of breast cancer cells. Int J Clin Exp Pathol. 2014;7(6):3057–3064. [PMC free article] [PubMed] [Google Scholar]
- 22.Nudelman AS, DiRocco DP, Lambert TJ, Garelick MG, Le J, Nathanson NM, et al. Neuronal activity rapidly induces transcription of the CREB-regulated microRNA-132, in vivo. Hippocampus. 2010;20(4):492–498. doi: 10.1002/hipo.20646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang ZG, Chen WX, Wu YH, Liang HF, Zhang BX. MiR-132 prohibits proliferation, invasion, migration, and metastasis in breast cancer by targeting HN1. Biochem Biophys Res Commun. 2014;454(1):109–114. doi: 10.1016/j.bbrc.2014.10.049. [DOI] [PubMed] [Google Scholar]
- 24.Wang Y, Han J, Fan S, Yang W, Zhang Y, Xu T, et al. miR-132 weakens proliferation and invasion of glioma cells via the inhibition of Gli1. Eur Rev Med Pharmacol Sci. 2018;22(7):1971–8. doi: 10.26355/eurrev_201804_14724. [DOI] [PubMed] [Google Scholar]
- 25.Li Y, Zu L, Wang Y, Wang M, Chen P, Zhou Q. miR-132 inhibits lung cancer cell migration and invasion by targeting SOX4. J Thorac Dis. 2015;7(9):1563. doi: 10.3978/j.issn.2072-1439.2015.09.06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhao D, Hou Y, Sun F, Han B, Li SJ. Effects of miR-132 on proliferation and apoptosis of pancreatic cancer cells via Hedgehog signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(5):1978–85. doi: 10.26355/eurrev_201903_17236. [DOI] [PubMed] [Google Scholar]
- 27.Zhang H, Liu A, Feng X, Tian L, Bo W, Wang H, et al. MiR-132 promotes the proliferation, invasion and migration of human pancreatic carcinoma by inhibition of the tumor suppressor gene PTEN. Prog Biophys Mol Biol. 2019;148:65–72. doi: 10.1016/j.pbiomolbio.2017.09.019. [DOI] [PubMed] [Google Scholar]
- 28.Aguda BD. Modeling microRNA-transcription factor networks in cancer. Adv Exp Med Biol. 2013;774:149–167. doi: 10.1007/978-94-007-5590-1_9. [DOI] [PubMed] [Google Scholar]
- 29.Gross DN, van den Heuvel AP, Birnbaum MJ. The role of FoxO in the regulation of metabolism. Oncogene. 2008;27(16):2320–2336. doi: 10.1038/onc.2008.25. [DOI] [PubMed] [Google Scholar]
- 30.Li W, Zhang J, Chen T, Yin P, Yang J, Cao Y. miR-132 upregulation promotes gastric cancer cell growth through suppression of FoxO1 translation. Tumour Biol. 2015 doi: 10.1007/s13277-015-3924-y. [DOI] [PubMed] [Google Scholar]
- 31.He K, Zeng H, Xu X, Li A, Cai Q, Long X. Clinicopathological significance of forkhead box protein A1 in breast cancer: a meta-analysis. Exp Ther Med. 2016;11(6):2525–2530. doi: 10.3892/etm.2016.3229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li YL, Zhao YG, Chen B, Li XF. MicroRNA-132 sensitizes nasopharyngeal carcinoma cells to cisplatin through regulation of forkhead box A1 protein. Pharmazie. 2016;71(12):715–718. doi: 10.1691/ph.2016.6764. [DOI] [PubMed] [Google Scholar]
- 33.Droog M, Nevedomskaya E, Kim Y, Severson T, Flach KD, Opdam M, et al. Comparative cistromics reveals genomic cross-talk between FOXA1 and ERalpha in tamoxifen-associated endometrial carcinomas. Can Res. 2016;76(13):3773–3784. doi: 10.1158/0008-5472.CAN-14-1813. [DOI] [PubMed] [Google Scholar]
- 34.Hurtado A, Holmes KA, Ross-Innes CS, Schmidt D, Carroll JS. FOXA1 is a key determinant of estrogen receptor function and endocrine response. Nat Genet. 2011;43(1):27–33. doi: 10.1038/ng.730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang D, Ren J, Ren H, Fu JL, Yu D. MicroRNA-132 suppresses cell proliferation in human breast cancer by directly targeting FOXA1. Acta Pharmacol Sin. 2018;39(1):124–131. doi: 10.1038/aps.2017.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen X, Li M, Zhou H, Zhang L. miR-132 targets FOXA1 and exerts tumor-suppressing functions in thyroid cancer. Oncol Res. 2019;27(4):431–437. doi: 10.3727/096504018X15201058168730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cao LH, Li HT, Lin WQ, Tan HY, Xie L, Zhong ZJ, et al. Morphine, a potential antagonist of cisplatin cytotoxicity, inhibits cisplatin-induced apoptosis and suppression of tumor growth in nasopharyngeal carcinoma xenografts. Sci Rep. 2016;6:18706. doi: 10.1038/srep18706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Liang R, Han B, Li Q, Yuan Y, Li J, Sun D. Using RNA sequencing to identify putative competing endogenous RNAs (ceRNAs) potentially regulating fat metabolism in bovine liver. Sci Rep. 2017;7(1):6396. doi: 10.1038/s41598-017-06634-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li Z, Han K, Zhang D, Chen J, Xu Z, Hou L. The role of long noncoding RNA in traumatic brain injury. Neuropsychiatr Dis Treat. 2019;15:1671–1677. doi: 10.2147/NDT.S206624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pirogov SA, Gvozdev VA, Klenov MS. Long noncoding RNAs and stress response in the nucleolus. Cells. 2019;8(7):688. doi: 10.3390/cells8070668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rahmani Z, Mojarrad M, Moghbeli M. Long non-coding RNAs as the critical factors during tumor progressions among Iranian population: an overview. Cell Biosci. 2020;10:6. doi: 10.1186/s13578-020-0373-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Liu W, Tang J, Zhang H, Kong F, Zhu H, Li P, et al. A novel lncRNA PTTG3P/miR-132/212-3p/FoxM1 feedback loop facilitates tumorigenesis and metastasis of pancreatic cancer. Cell Death Discov. 2020;6(1):136. doi: 10.1038/s41420-020-00360-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Xue MY, Cao HX. LINC01551 promotes metastasis of nasopharyngeal carcinoma through targeting microRNA-132-5p. Eur Rev Med Pharmacol Sci. 2020;24(7):3724–3733. doi: 10.26355/eurrev_202004_20836. [DOI] [PubMed] [Google Scholar]
- 44.Hersh CP, Silverman EK, Gascon J, Bhattacharya S, Klanderman BJ, Litonjua AA, et al. SOX5 is a candidate gene for chronic obstructive pulmonary disease susceptibility and is necessary for lung development. Am J Respir Crit Care Med. 2011;183(11):1482–1489. doi: 10.1164/rccm.201010-1751OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Renjie W, Haiqian L. MiR-132, miR-15a and miR-16 synergistically inhibit pituitary tumor cell proliferation, invasion and migration by targeting Sox5. Cancer Lett. 2015;356(2 Pt B):568–78. doi: 10.1016/j.canlet.2014.10.003. [DOI] [PubMed] [Google Scholar]
- 46.Wilusz JE, Sharp PA. Molecular biology. A circuitous route to noncoding RNA. Science. 2013;340(6131):440–1. doi: 10.1126/science.1238522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chen I, Chen CY, Chuang TJ. Biogenesis, identification, and function of exonic circular RNAs. Wiley Interdiscipl Rev RNA. 2015;6(5):563–579. doi: 10.1002/wrna.1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Salzman J, Circular RNA. Expression: its potential regulation and function. Trends Genet. 2016;32(5):309–316. doi: 10.1016/j.tig.2016.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Liu P, Li X, Guo X, Chen J, Li C, Chen M, et al. Circular RNA DOCK1 promotes bladder carcinoma progression via modulating circDOCK1/hsa-miR-132–3p/Sox5 signalling pathway. Cell Prolif. 2019;52(4):e12614. doi: 10.1111/cpr.12614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gopal K, Gupta N, Zhang H, Alshareef A, Alqahtani H, Bigras G, et al. Oxidative stress induces the acquisition of cancer stem-like phenotype in breast cancer detectable by using a Sox2 regulatory region-2 (SRR2) reporter. Oncotarget. 2016;7(3):3111–3127. doi: 10.18632/oncotarget.6630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhou K, Zhang C, Yao H, Zhang X, Zhou Y, Che Y, et al. Knockdown of long non-coding RNA NEAT1 inhibits glioma cell migration and invasion via modulation of SOX2 targeted by miR-132. Mol Cancer. 2018;17(1):105. doi: 10.1186/s12943-018-0849-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Abbaszadegan MR, Moghbeli M. Role of MAML1 and MEIS1 in esophageal squamous cell carcinoma depth of invasion. Pathol Oncol Res. 2018;24(2):245–250. doi: 10.1007/s12253-017-0243-1. [DOI] [PubMed] [Google Scholar]
- 53.Moghbeli M, Mosannen Mozaffari H, Memar B, Forghanifard MM, Gholamin M, Abbaszadegan MR. Role of MAML1 in targeted therapy against the esophageal cancer stem cells. J Transl Med. 2019;17(1):126. doi: 10.1186/s12967-019-1876-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Moghbeli M, Rad A, Farshchian M, Taghehchian N, Gholamin M, Abbaszadegan MR. Correlation between Meis1 and Msi1 in esophageal squamous cell carcinoma. J Gastrointest Cancer. 2016;47(3):273–277. doi: 10.1007/s12029-016-9824-6. [DOI] [PubMed] [Google Scholar]
- 55.Bolos V, Peinado H, Perez-Moreno MA, Fraga MF, Esteller M, Cano A. The transcription factor Slug represses E-cadherin expression and induces epithelial to mesenchymal transitions: a comparison with Snail and E47 repressors. J Cell Sci. 2003;116(Pt 3):499–511. doi: 10.1242/jcs.00224. [DOI] [PubMed] [Google Scholar]
- 56.Cano A, Perez-Moreno MA, Rodrigo I, Locascio A, Blanco MJ, del Barrio MG, et al. The transcription factor snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat Cell Biol. 2000;2(2):76–83. doi: 10.1038/35000025. [DOI] [PubMed] [Google Scholar]
- 57.Yang J, Mani SA, Donaher JL, Ramaswamy S, Itzykson RA, Come C, et al. Twist, a master regulator of morphogenesis, plays an essential role in tumor metastasis. Cell. 2004;117(7):927–939. doi: 10.1016/j.cell.2004.06.006. [DOI] [PubMed] [Google Scholar]
- 58.Liu Y, Li Y, Liu J, Wu Y, Zhu Q. MicroRNA-132 inhibits cell growth and metastasis in osteosarcoma cell lines possibly by targeting Sox4. Int J Oncol. 2015;47(5):1672–1684. doi: 10.3892/ijo.2015.3147. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 59.Wang L, Zhang J, Yang X, Chang YW, Qi M, Zhou Z, et al. SOX4 is associated with poor prognosis in prostate cancer and promotes epithelial-mesenchymal transition in vitro. Prostate Cancer Prostatic Dis. 2013;16(4):301–307. doi: 10.1038/pcan.2013.25. [DOI] [PubMed] [Google Scholar]
- 60.Fu W, Tao T, Qi M, Wang L, Hu J, Li X, et al. MicroRNA-132/212 upregulation inhibits TGF-beta-mediated epithelial-mesenchymal transition of prostate cancer cells by targeting SOX4. Prostate. 2016;76(16):1560–1570. doi: 10.1002/pros.23241. [DOI] [PubMed] [Google Scholar]
- 61.Li Y, Zu L, Wang Y, Wang M, Chen P, Zhou Q. miR-132 inhibits lung cancer cell migration and invasion by targeting SOX4. J Thorac Dis. 2015;7(9):1563–1569. doi: 10.3978/j.issn.2072-1439.2015.09.06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Huang J, Lu D, Xiang T, Wu X, Ge S, Wang Y, et al. MicroRNA-132-3p regulates cell proliferation, apoptosis, migration and invasion of liver cancer by targeting Sox4. Oncol Lett. 2020;19(4):3173–3180. doi: 10.3892/ol.2020.11431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Chen HZ, Tsai SY, Leone G. Emerging roles of E2Fs in cancer: an exit from cell cycle control. Nat Rev Cancer. 2009;9(11):785–797. doi: 10.1038/nrc2696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tian H, Hou L, Xiong YM, Huang JX, Zhang WH, Pan YY, et al. miR-132 targeting E2F5 suppresses cell proliferation, invasion, migration in ovarian cancer cells. Am J Transl Res. 2016;8(3):1492–1501. [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 65.Haigis MC, Guarente LP. Mammalian sirtuins–emerging roles in physiology, aging, and calorie restriction. Genes Dev. 2006;20(21):2913–2921. doi: 10.1101/gad.1467506. [DOI] [PubMed] [Google Scholar]
- 66.Longo VD, Kennedy BK. Sirtuins in aging and age-related disease. Cell. 2006;126(2):257–268. doi: 10.1016/j.cell.2006.07.002. [DOI] [PubMed] [Google Scholar]
- 67.Eberle D, Hegarty B, Bossard P, Ferre P, Foufelle F. SREBP transcription factors: master regulators of lipid homeostasis. Biochimie. 2004;86(11):839–848. doi: 10.1016/j.biochi.2004.09.018. [DOI] [PubMed] [Google Scholar]
- 68.Li Y, Zhang J, He J, Zhou W, Xiang G, Xu R. MicroRNA-132 cause apoptosis of glioma cells through blockade of the SREBP-1c metabolic pathway related to SIRT1. Biomed Pharmacother. 2016;78:177–84. doi: 10.1016/j.biopha.2016.01.022. [DOI] [PubMed] [Google Scholar]
- 69.He S, Deng Y, Liao Y, Li X, Liu J, Yao S. CREB5 promotes tumor cell invasion and correlates with poor prognosis in epithelial ovarian cancer. Oncol Lett. 2017;14(6):8156–8161. doi: 10.3892/ol.2017.7234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zhang M, Li Y, Wang H, Yu W, Lin S, Guo J. LncRNA SNHG5 affects cell proliferation, metastasis and migration of colorectal cancer through regulating miR-132-3p/CREB5. Cancer Biol Ther. 2019;20(4):524–536. doi: 10.1080/15384047.2018.1537579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Vire E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C, et al. The Polycomb group protein EZH2 directly controls DNA methylation. Nature. 2006;439(7078):871–874. doi: 10.1038/nature04431. [DOI] [PubMed] [Google Scholar]
- 72.Cao R, Zhang Y. The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr Opin Genet Dev. 2004;14(2):155–164. doi: 10.1016/j.gde.2004.02.001. [DOI] [PubMed] [Google Scholar]
- 73.Tiwari N, Tiwari VK, Waldmeier L, Balwierz PJ, Arnold P, Pachkov M, et al. Sox4 is a master regulator of epithelial-mesenchymal transition by controlling Ezh2 expression and epigenetic reprogramming. Cancer Cell. 2013;23(6):768–783. doi: 10.1016/j.ccr.2013.04.020. [DOI] [PubMed] [Google Scholar]
- 74.Lin L, Wang Z, Jin H, Shi H, Lu Z, Qi Z. MiR-212/132 is epigenetically downregulated by SOX4/EZH2-H3K27me3 feedback loop in ovarian cancer cells. Tumour Biol. 2016 doi: 10.1007/s13277-016-5339-9. [DOI] [PubMed] [Google Scholar]
- 75.Weng MY, Li L, Feng SY, Hong SJ. Expression of Bmi-1, P16, and CD44v6 in uterine cervical carcinoma and its clinical significance. Cancer Biol Med. 2012;9(1):48–53. doi: 10.3969/j.issn.2095-3941.2012.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Liu YL, Jiang SX, Yang YM, Xu H, Liu JL, Wang XS. USP22 acts as an oncogene by the activation of BMI-1-mediated INK4a/ARF pathway and Akt pathway. Cell Biochem Biophys. 2012;62(1):229–235. doi: 10.1007/s12013-011-9287-0. [DOI] [PubMed] [Google Scholar]
- 77.Tong YQ, Liu B, Zheng HY, He YJ, Gu J, Li F, et al. Overexpression of BMI-1 is associated with poor prognosis in cervical cancer. Asia Pac J Clin Oncol. 2012;8(4):e55–62. doi: 10.1111/j.1743-7563.2012.01564.x. [DOI] [PubMed] [Google Scholar]
- 78.Liu GF, Zhang SH, Li XF, Cao LY, Fu ZZ, Yu SN. Overexpression of microRNA-132 enhances the radiosensitivity of cervical cancer cells by down-regulating Bmi-1. Oncotarget. 2017;8(46):80757–80769. doi: 10.18632/oncotarget.20358. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 79.Zhang XL, Sun BL, Tian SX, Li L, Zhao YC, Shi PP. MicroRNA-132 reverses cisplatin resistance and metastasis in ovarian cancer by the targeted regulation on Bmi-1. Eur Rev Med Pharmacol Sci. 2019;23(9):3635–44. doi: 10.26355/eurrev_201905_17787. [DOI] [PubMed] [Google Scholar]
- 80.Zhang K, Li Y, Qu L, Ma X, Zhao H, Tang Y. Long noncoding RNA Sox2 overlapping transcript (SOX2OT) promotes non-small-cell lung cancer migration and invasion via sponging microRNA 132 (miR-132) Onco Targets Ther. 2018;11:5269–5278. doi: 10.2147/OTT.S168654. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 81.Zheng YB, Luo HP, Shi Q, Hao ZN, Ding Y, Wang QS, et al. miR-132 inhibits colorectal cancer invasion and metastasis via directly targeting ZEB2. World J Gastroenterol. 2014;20(21):6515–6522. doi: 10.3748/wjg.v20.i21.6515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Murtaza M, Jolly LA, Gecz J, Wood SA. La FAM fatale: USP9X in development and disease. Cell Mol Life Sci. 2015;72(11):2075–2089. doi: 10.1007/s00018-015-1851-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kushwaha D, O'Leary C, Cron KR, Deraska P, Zhu K, D'Andrea AD, et al. USP9X inhibition promotes radiation-induced apoptosis in non-small cell lung cancer cells expressing mid-to-high MCL1. Cancer Biol Ther. 2015;16(3):392–401. doi: 10.1080/15384047.2014.1002358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Guo H, Zhang X, Chen Q, Bao Y, Dong C, Wang X. miR-132 suppresses the migration and invasion of lung cancer cells by blocking USP9X-induced epithelial-mesenchymal transition. Am J Transl Res. 2018;10(1):224–234. [PMC free article] [PubMed] [Google Scholar]
- 85.Liu T, Liu J, Chen Q, Jin S, Mi S, Shao W, et al. Expression of USP22 and the chromosomal passenger complex is an indicator of malignant progression in oral squamous cell carcinoma. Oncol Lett. 2019;17(2):2040–2046. doi: 10.3892/ol.2018.9837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.He X, Ma J, Zhang M, Cui J, Yang H. Long Non-Coding RNA SNHG16 Activates USP22 Expression to Promote Colorectal Cancer Progression by Sponging miR-132-3p. Onco Targets Ther. 2020;13:4283–4294. doi: 10.2147/OTT.S244778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Varisli L, Ozturk BE, Akyuz GK, Korkmaz KS. HN1 negatively influences the beta-catenin/E-cadherin interaction, and contributes to migration in prostate cells. J Cell Biochem. 2015;116(1):170–178. doi: 10.1002/jcb.24956. [DOI] [PubMed] [Google Scholar]
- 88.Liao J, He Q, Li M, Chen Y, Liu Y, Wang J. LncRNA MIAT: myocardial infarction associated and more. Gene. 2016;578(2):158–161. doi: 10.1016/j.gene.2015.12.032. [DOI] [PubMed] [Google Scholar]
- 89.Liu Z, Wang H, Cai H, Hong Y, Li Y, Su D, et al. Long non-coding RNA MIAT promotes growth and metastasis of colorectal cancer cells through regulation of miR-132/Derlin-1 pathway. Cancer Cell Int. 2018;18:59. doi: 10.1186/s12935-017-0477-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Cheng ZX, Yin WB, Wang ZY. MicroRNA-132 induces temozolomide resistance and promotes the formation of cancer stem cell phenotypes by targeting tumor suppressor candidate 3 in glioblastoma. Int J Mol Med. 2017;40(5):1307–1314. doi: 10.3892/ijmm.2017.3124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Liu X, Winey M. The MPS1 family of protein kinases. Annu Rev Biochem. 2012;81:561–585. doi: 10.1146/annurev-biochem-061611-090435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Liu X, Liao W, Yuan Q, Ou Y, Huang J. TTK activates Akt and promotes proliferation and migration of hepatocellular carcinoma cells. Oncotarget. 2015;6(33):34309–34320. doi: 10.18632/oncotarget.5295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ritchie A, Gutierrez O, Fernandez-Luna JL. PAR bZIP-bik is a novel transcriptional pathway that mediates oxidative stress-induced apoptosis in fibroblasts. Cell Death Differ. 2009;16(6):838–846. doi: 10.1038/cdd.2009.13. [DOI] [PubMed] [Google Scholar]
- 94.Chen S, Wang Y, Ni C, Meng G, Sheng X. HLF/miR-132/TTK axis regulates cell proliferation, metastasis and radiosensitivity of glioma cells. Biomed Pharmacother. 2016;83:898–904. doi: 10.1016/j.biopha.2016.08.004. [DOI] [PubMed] [Google Scholar]
- 95.Kelly ML, Chernoff J. Getting smart about p21-activated kinases. Mol Cell Biol. 2011;31(3):386–387. doi: 10.1128/MCB.01267-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Liu F, Li X, Wang C, Cai X, Du Z, Xu H, et al. Downregulation of p21-activated kinase-1 inhibits the growth of gastric cancer cells involving cyclin B1. Int J Cancer. 2009;125(11):2511–2519. doi: 10.1002/ijc.24588. [DOI] [PubMed] [Google Scholar]
- 97.Kumar R, Sanawar R, Li X, Li F. Structure, biochemistry, and biology of PAK kinases. Gene. 2017;605:20–31. doi: 10.1016/j.gene.2016.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Mazumdar A, Kumar R. Estrogen regulation of Pak1 and FKHR pathways in breast cancer cells. FEBS Lett. 2003;535(1–3):6–10. doi: 10.1016/S0014-5793(02)03846-2. [DOI] [PubMed] [Google Scholar]
- 99.Xu J, Liu H, Chen L, Wang S, Zhou L, Yun X, et al. Hepatitis B virus X protein confers resistance of hepatoma cells to anoikis by up-regulating and activating p21-activated kinase 1. Gastroenterology. 2012;143(1):199–212. doi: 10.1053/j.gastro.2012.03.053. [DOI] [PubMed] [Google Scholar]
- 100.Claps G, Cheli Y, Zhang T, Scortegagna M, Lau E, Kim H, et al. A transcriptionally inactive ATF2 variant drives melanomagenesis. Cell Rep. 2016;15(9):1884–1892. doi: 10.1016/j.celrep.2016.04.072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Van Obberghen-Schilling E, Tucker RP, Saupe F, Gasser I, Cseh B, Orend G. Fibronectin and tenascin-C: accomplices in vascular morphogenesis during development and tumor growth. Int J Dev Biol. 2011;55(4–5):511–525. doi: 10.1387/ijdb.103243eo. [DOI] [PubMed] [Google Scholar]
- 102.Liu F, Cheng Z, Li X, Li Y, Zhang H, Li J, et al. A novel Pak1/ATF2/miR-132 signaling axis is involved in the hematogenous metastasis of gastric cancer cells. Mol Ther Nucleic Acids. 2017;8:370–382. doi: 10.1016/j.omtn.2017.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Chung C. Tyrosine kinase inhibitors for epidermal growth factor receptor gene mutation-positive non-small cell lung cancers: an update for recent advances in therapeutics. J Oncol Pharm Pract. 2016;22(3):461–476. doi: 10.1177/1078155215577810. [DOI] [PubMed] [Google Scholar]
- 104.Shabani M, Naseri J, Shokri F. Receptor tyrosine kinase-like orphan receptor 1: a novel target for cancer immunotherapy. Expert Opin Ther Targets. 2015;19(7):941–955. doi: 10.1517/14728222.2015.1025753. [DOI] [PubMed] [Google Scholar]
- 105.Chauhan SC, Ebeling MC, Maher DM, Koch MD, Watanabe A, Aburatani H, et al. MUC13 mucin augments pancreatic tumorigenesis. Mol Cancer Ther. 2012;11(1):24–33. doi: 10.1158/1535-7163.MCT-11-0598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Khan S, Ebeling MC, Zaman MS, Sikander M, Yallapu MM, Chauhan N, et al. MicroRNA-145 targets MUC13 and suppresses growth and invasion of pancreatic cancer. Oncotarget. 2014;5(17):7599–7609. doi: 10.18632/oncotarget.2281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.He L, Qu L, Wei L, Chen Y, Suo J. Reduction of miR1323p contributes to gastric cancer proliferation by targeting MUC13. Mol Med Rep. 2017;15(5):3055–3061. doi: 10.3892/mmr.2017.6347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Qu W, Ding SM, Cao G, Wang SJ, Zheng XH, Li GH. miR-132 mediates a metabolic shift in prostate cancer cells by targeting Glut1. FEBS Open Bio. 2016;6(7):735–741. doi: 10.1002/2211-5463.12086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Eramo A, Pallini R, Lotti F, Sette G, Patti M, Bartucci M, et al. Inhibition of DNA methylation sensitizes glioblastoma for tumor necrosis factor-related apoptosis-inducing ligand-mediated destruction. Can Res. 2005;65(24):11469–11477. doi: 10.1158/0008-5472.CAN-05-1724. [DOI] [PubMed] [Google Scholar]
- 110.Garofalo M, Romano G, Quintavalle C, Romano MF, Chiurazzi F, Zanca C, et al. Selective inhibition of PED protein expression sensitizes B-cell chronic lymphocytic leukaemia cells to TRAIL-induced apoptosis. Int J Cancer. 2007;120(6):1215–1222. doi: 10.1002/ijc.22495. [DOI] [PubMed] [Google Scholar]
- 111.Zanca C, Garofalo M, Quintavalle C, Romano G, Acunzo M, Ragno P, et al. PED is overexpressed and mediates TRAIL resistance in human non-small cell lung cancer. J Cell Mol Med. 2008;12(6A):2416–2426. doi: 10.1111/j.1582-4934.2008.00283.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Geng F, Wu JL, Lu GF, Liang ZP, Duan ZL, Gu X. MicroRNA-132 targets PEA-15 and suppresses the progression of astrocytoma in vitro. J Neurooncol. 2016;129(2):211–220. doi: 10.1007/s11060-016-2173-2. [DOI] [PubMed] [Google Scholar]
- 113.Wang J, Xu G, Shen F, Kang Y. miR-132 targeting cyclin E1 suppresses cell proliferation in osteosarcoma cells. Tumour Biol. 2014;35(5):4859–4865. doi: 10.1007/s13277-014-1637-2. [DOI] [PubMed] [Google Scholar]
- 114.Peltier J, O'Neill A, Schaffer DV. PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation. Dev Neurobiol. 2007;67(10):1348–1361. doi: 10.1002/dneu.20506. [DOI] [PubMed] [Google Scholar]
- 115.Rafalski VA, Brunet A. Energy metabolism in adult neural stem cell fate. Prog Neurobiol. 2011;93(2):182–203. doi: 10.1016/j.pneurobio.2010.10.007. [DOI] [PubMed] [Google Scholar]
- 116.Zhao X, Gan L, Pan H, Kan D, Majeski M, Adam SA, et al. Multiple elements regulate nuclear/cytoplasmic shuttling of FOXO1: characterization of phosphorylation- and 14-3-3-dependent and -independent mechanisms. Biochem J. 2004;378(Pt 3):839–849. doi: 10.1042/bj20031450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Ho KK, Myatt SS, Lam EW. Many forks in the path: cycling with FoxO. Oncogene. 2008;27(16):2300–2311. doi: 10.1038/onc.2008.23. [DOI] [PubMed] [Google Scholar]
- 118.Lam EW, Brosens JJ, Gomes AR, Koo CY. Forkhead box proteins: tuning forks for transcriptional harmony. Nat Rev Cancer. 2013;13(7):482–495. doi: 10.1038/nrc3539. [DOI] [PubMed] [Google Scholar]
- 119.Lian R, Lu B, Jiao L, Li S, Wang H, Miao W, et al. MiR-132 plays an oncogenic role in laryngeal squamous cell carcinoma by targeting FOXO1 and activating the PI3K/AKT pathway. Eur J Pharmacol. 2016;792:1–6. doi: 10.1016/j.ejphar.2016.10.015. [DOI] [PubMed] [Google Scholar]
- 120.Cai XM, Tao BB, Wang LY, Liang YL, Jin JW, Yang Y, et al. Protein phosphatase activity of PTEN inhibited the invasion of glioma cells with epidermal growth factor receptor mutation type III expression. Int J Cancer. 2005;117(6):905–912. doi: 10.1002/ijc.21251. [DOI] [PubMed] [Google Scholar]
- 121.Wu D, Li M, Tian W, Wang S, Cui L, Li H, et al. Hydrogen sulfide acts as a double-edged sword in human hepatocellular carcinoma cells through EGFR/ERK/MMP-2 and PTEN/AKT signaling pathways. Sci Rep. 2017;7(1):5134. doi: 10.1038/s41598-017-05457-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Liu H, Pan Y, Han X, Liu J, Li R. MicroRNA-216a promotes the metastasis and epithelial-mesenchymal transition of ovarian cancer by suppressing the PTEN/AKT pathway. Onco Targets Ther. 2017;10:2701–2709. doi: 10.2147/OTT.S114318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Zhang H, Liu A, Feng X, Tian L, Bo W, Wang H, et al. MiR-132 promotes the proliferation, invasion and migration of human pancreatic carcinoma by inhibition of the tumor suppressor gene PTEN. Prog Biophys Mol Biol. 2019;148:65–72. doi: 10.1016/j.pbiomolbio.2017.09.019. [DOI] [PubMed] [Google Scholar]
- 124.Xie M, Fu Z, Cao J, Liu Y, Wu J, Li Q, et al. MicroRNA-132 and microRNA-212 mediate doxorubicin resistance by down-regulating the PTEN-AKT/NF-kappaB signaling pathway in breast cancer. Biomed Pharmacother. 2018;102:286–94. doi: 10.1016/j.biopha.2018.03.088. [DOI] [PubMed] [Google Scholar]
- 125.Agami R. microRNAs, RNA binding proteins and cancer. Eur J Clin Invest. 2010;40(4):370–374. doi: 10.1111/j.1365-2362.2010.02279.x. [DOI] [PubMed] [Google Scholar]
- 126.Chen T, Lu M, Zhou X, Pan X, Han Y, Zhang Y, et al. miR-132 and miR-212 cluster function as a tumor suppressor in thyroid cancer cells by CSDE1 mediated post-transcriptional program. Int J Clin Exp Pathol. 2018;11(2):963–971. [PMC free article] [PubMed] [Google Scholar]
- 127.Li L, Wei XH, Pan YP, Li HC, Yang H, He QH, et al. LAPTM4B: a novel cancer-associated gene motivates multidrug resistance through efflux and activating PI3K/AKT signaling. Oncogene. 2010;29(43):5785–5795. doi: 10.1038/onc.2010.303. [DOI] [PubMed] [Google Scholar]
- 128.Li S, Xu JJ, Zhang QY. MicroRNA-132-3p inhibits tumor malignant progression by regulating lysosomal-associated protein transmembrane 4 beta in breast cancer. Cancer Sci. 2019;110(10):3098–3109. doi: 10.1111/cas.14164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Zhang W, Liu Y, Fu Y, Han W, Xu H, Wen L, et al. Long non-coding RNA LINC00160 functions as a decoy of microRNA-132 to mediate autophagy and drug resistance in hepatocellular carcinoma via inhibition of PIK3R3. Cancer Lett. 2020;478:22–33. doi: 10.1016/j.canlet.2020.02.014. [DOI] [PubMed] [Google Scholar]
- 130.Zhao JL, Zhang L, Guo X, Wang JH, Zhou W, Liu M, et al. miR-212/132 downregulates SMAD2 expression to suppress the G1/S phase transition of the cell cycle and the epithelial to mesenchymal transition in cervical cancer cells. IUBMB Life. 2015;67(5):380–394. doi: 10.1002/iub.1381. [DOI] [PubMed] [Google Scholar]
- 131.Wei XC, Lv ZH. MicroRNA-132 inhibits migration, invasion and epithelial-mesenchymal transition via TGFbeta1/Smad2 signaling pathway in human bladder cancer. Onco Targets Ther. 2019;12:5937–5945. doi: 10.2147/OTT.S201731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Han S, Song L, Chen Y, Hou M, Wei X, Fan D. The long non-coding RNA ILF3-AS1 increases the proliferation and invasion of retinoblastoma through the miR-132–3p/SMAD2 axis. Exp Cell Res. 2020;393(2):112087. doi: 10.1016/j.yexcr.2020.112087. [DOI] [PubMed] [Google Scholar]
- 133.Chen L, Zhu Q, Lu L, Liu Y. MiR-132 inhibits migration and invasion and increases chemosensitivity of cisplatin-resistant oral squamous cell carcinoma cells via targeting TGF-beta1. Bioengineered. 2020;11(1):91–102. doi: 10.1080/21655979.2019.1710925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Pufall MA. Glucocorticoids and cancer. Adv Exp Med Biol. 2015;872:315–333. doi: 10.1007/978-1-4939-2895-8_14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Liu L, Aleksandrowicz E, Schonsiegel F, Groner D, Bauer N, Nwaeburu CC, et al. Dexamethasone mediates pancreatic cancer progression by glucocorticoid receptor, TGFbeta and JNK/AP-1. Cell Death Dis. 2017;8(10):e3064. doi: 10.1038/cddis.2017.455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Abukiwan A, Nwaeburu CC, Bauer N, Zhao Z, Liu L, Gladkich J, et al. Dexamethasone-induced inhibition of miR-132 via methylation promotes TGF-beta-driven progression of pancreatic cancer. Int J Oncol. 2019;54(1):53–64. doi: 10.3892/ijo.2018.4616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y, Hu LL. ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med. 2020;19(3):1997–2007. doi: 10.3892/etm.2020.8454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Song XF, Chang H, Liang Q, Guo ZF, Wu JW. ZEB1 promotes prostate cancer proliferation and invasion through ERK1/2 signaling pathway. Eur Rev Med Pharmacol Sci. 2017;21(18):4032–4038. [PubMed] [Google Scholar]
- 139.Liu Y, Zhang M. miR-132 regulates adriamycin resistance in colorectal cancer cells through targeting extracellular signal-regulated kinase 1. Cancer Biother Radiopharm. 2019;34(6):398–404. doi: 10.1089/cbr.2018.2749. [DOI] [PubMed] [Google Scholar]
- 140.Song H, He P, Shao T, Li Y, Li J, Zhang Y. Long non-coding RNA XIST functions as an oncogene in human colorectal cancer by targeting miR-132-3p. J BUON. 2017;22(3):696–703. [PubMed] [Google Scholar]
- 141.Bangs F, Anderson KV. Primary cilia and mammalian hedgehog signaling. Cold Spring Harb Perspect Biol. 2017;9(5):a028175. doi: 10.1101/cshperspect.a028175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Jung Y, McCall SJ, Li YX, Diehl AM. Bile ductules and stromal cells express hedgehog ligands and/or hedgehog target genes in primary biliary cirrhosis. Hepatology. 2007;45(5):1091–1096. doi: 10.1002/hep.21660. [DOI] [PubMed] [Google Scholar]
- 143.Zhao L, Yu Y, Deng C. Protein and mRNA expression of Shh, Smo and Gli1 and inhibition by cyclopamine in hepatocytes of rats with chronic fluorosis. Toxicol Lett. 2014;225(2):318–324. doi: 10.1016/j.toxlet.2013.12.022. [DOI] [PubMed] [Google Scholar]
- 144.Zhao DW, Hou YS, Sun FB, Han B, Li SJ. Effects of miR-132 on proliferation and apoptosis of pancreatic cancer cells via Hedgehog signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(5):1978–1985. doi: 10.26355/eurrev_201903_17236. [DOI] [PubMed] [Google Scholar]
- 145.Camargo FD, Gokhale S, Johnnidis JB, Fu D, Bell GW, Jaenisch R, et al. YAP1 increases organ size and expands undifferentiated progenitor cells. Curr Biol. 2007;17(23):2054–2060. doi: 10.1016/j.cub.2007.10.039. [DOI] [PubMed] [Google Scholar]
- 146.Zhao B, Li L, Lei Q, Guan KL. The Hippo-YAP pathway in organ size control and tumorigenesis: an updated version. Genes Dev. 2010;24(9):862–874. doi: 10.1101/gad.1909210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Lei CJ, Li L, Gao X, Zhang J, Pan QY, Long HC, et al. Hsa-miR-132 inhibits proliferation of hepatic carcinoma cells by targeting YAP. Cell Biochem Funct. 2015;33(5):326–333. doi: 10.1002/cbf.3119. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.