

Abstract
Background
Wolbachia pipientis are endosymbiotic bacteria present in a large proportion of terrestrial arthropods. The species is known to sometimes affect the ability of its host to transmit vector-borne pathogens. Central Sweden is endemic for Sindbis virus (SINV), where it is mainly transmitted by the vector species Culex pipiens and Culex torrentium, with the latter established as the main vector. In this study we investigated the Wolbachia prevalence in these two vector species in a region highly endemic for SINV.
Methods
Culex mosquitoes were collected using CDC light traps baited with carbon dioxide over 9 years at 50 collection sites across the River Dalälven floodplains in central Sweden. Mosquito genus was determined morphologically, while a molecular method was used for reliable species determination. The presence of Wolbachia was determined through PCR using general primers targeting the wsp gene and sequencing of selected samples.
Results
In total, 676 Cx. pipiens and 293 Cx. torrentium were tested for Wolbachia. The prevalence of Wolbachia in Cx. pipiens was 97% (95% CI 94.8–97.6%), while only 0.7% (95% CI 0.19–2.45%) in Cx. torrentium. The two Cx. torrentium mosquitoes that were infected with Wolbachia carried different types of the bacteria.
Conclusions
The main vector of SINV in the investigated endemic region, Cx. torrentium, was seldom infected with Wolbachia, while it was highly prevalent in the secondary vector, Cx. pipiens. The presence of Wolbachia could potentially have an impact on the vector competence of these two species. Furthermore, the detection of Wolbachia in Cx. torrentium could indicate horizontal transmission of the endosymbiont between arthropods of different species.
Graphical abstract
Keywords: Vector, Field, Mosquito, Endosymbiont, Alphavirus, Horizontal transmission, Scandinavia
Background
The transmission of arboviruses is influenced by a number of factors, including both abiotic (e.g., temperature) and biotic elements (e.g., vector immune status) [1–4]. One important biotic factor is the intracellular symbiont Wolbachia pipientis (Class: Alphaproteobacteria, Order: Rickettsiales), present in some nematode species and an estimated 40% of all terrestrial arthropods [5]. Wolbachia is a genetically diverse species, composed of 18 phylogenetically distinct supergroups described to date (A–R) [6]. Deeply involved in the reproduction of its host [7], Wolbachia is known for inducing cytoplasmic incompatibility and giving rise to crossing types, most studied in Culex pipiens and its Wolbachia strain wPip, which belongs to supergroup B [8–10]. Additionally, it is well established that Wolbachia infection in mosquitoes can influence their ability to become infected and transmit several arboviruses [11–14].
In a global context, Culex mosquitoes are important vectors for, e.g., West Nile virus (WNV) and Japanese encephalitis virus (JEV) [15–18]. In Central and Northern Europe, the morphologically identical vector species Cx. pipiens and Cx. torrentium are enzootic vectors of both WNV and Sindbis virus (SINV), transmitting these viruses among birds [19–22]. SINV is an arthritogenic alphavirus present throughout the Old World [23], although outbreaks of human disease are only reported from South Africa [24, 25] and Fennoscandia [26–29]. In Sweden, SINV is considered endemic to the central and northern parts of the country [28–30].
Culex torrentium is regarded as the most important enzootic vector in Sweden due to its high abundance in endemic areas, high infection rate, and superior vector competence to Cx. pipiens [31–34]. One difference between Cx. torrentium and Cx. pipiens is the prevalence of Wolbachia-infected individuals. Previous studies in Germany, Belgium, Russia, Belarus, Kazakhstan, and Kyrgyz Republic have found Wolbachia to be very common in Cx. pipiens but absent in Cx. torrentium [35–38]. It is therefore possible that these differences in Wolbachia infection status could account for part of the difference in vector competence seen between Cx. pipiens and Cx. torrentium. Previous studies have however only been performed in regions without intense SINV transmission. Therefore, this study aims at investigating the Wolbachia prevalence in Cx. pipiens and Cx. torrentium collected in a highly SINV-endemic region in central Sweden.
Methods
Mosquitoes
Mosquitoes were collected at 50 different locations across the River Dalälven floodplains (Fig. 1) as part of a routine mosquito monitoring programme [39]. SINV is considered endemic to this region and some of the highest infection rates in mosquitoes have been detected here [22, 33]. Collections were performed every second week between May and September during the years 2010–2018 using CDC light traps baited with carbon dioxide. Mosquitoes were identified based on morphological characteristics [40], and Cx. pipiens/torrentium were sorted out and used for molecular identification to species. Briefly, individual mosquitoes were homogenized in 500 µl of phosphate-buffered saline (PBS) supplemented with 20% heat-inactivated fetal bovine serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 2.5 μg/ml amphotericin B (Thermo Fischer Scientific; Waltham, MA, USA) using two steel beads in the Qiagen TissueLyser II™ (Qiagen; Hilden, Germany). Five microliters (5 µl) of the homogenate was pretreated by incubating at 98 °C for 2 min in 20 µl of dilution buffer with 0.5 µl of DNA release additive, part of the Phire Tissue Direct PCR Master Mix kit (Thermo Scientific; Vilnius, Lithuania). The pretreated homogenate was stored at −20 °C before being used as a template in polymerase chain reaction (PCR). Conventional PCR of part of the cytochrome oxidase subunit I (COI) was performed in 20 µl reactions with 1 µl template using the forward primer C1-J-2183 (5′-CAACATTTATTTTGATTTTTTGG-3′) and the reverse primer TL2-N-3014 (5′-TCCAATGCACTAATCTGCCATATTA-3′) at a concentration of 0.5 µM each under the following thermocycler conditions: initial denaturation at 98 °C for 5 min, followed by 40 cycles of denaturation at 98 °C for 5 s, annealing at 54.5 °C for 5 s and extension at 72 °C for 20 s, and a final extension step at 72 °C for 1 min. A PCR-restriction fragment length polymorphism (PCR–RFLP) assay [41] was performed on the PCR product, using the restriction enzymes FspBi and SspI (Thermo Fischer Scientific; Vilnius, Lithuania).
Fig. 1.

Map showing the collection sites of Wolbachia-screened Culex mosquitoes. Collection sites are marked as black diamonds. Map data retrieved from
©OpenStreetMap contributors under the Open Database License
Culex pipiens molestus mosquitoes, originating from a field population sampled in Gothenburg, Sweden [42] and reared in our in-house mosquito rearing facility, were used as positive controls in PCR as they are naturally infected with a wPip strain of Wolbachia (data not shown). These were also used for PCR optimization.
Wolbachia detection
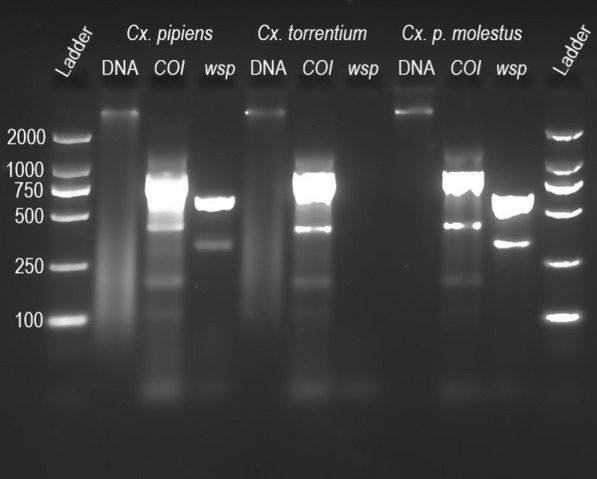
Wolbachia detection was performed through PCR on 5 µl of the mosquito homogenate, using the same Tissue Direct kit procedures as described above. Wolbachia primers 81F (5′-TGGTCCAATAAGTGATGAAGAAAC-3′) and 691R (5′-AAAAATTAAACGCTACTCCA-3′), designed for general detection of Wolbachia within supergroups A and B [43], were used at a final concentration of 0.5 µM each. The thermocycler conditions for Wolbachia detection were as described above but with the annealing temperature set to 58 °C. A subset of samples was also tested with a confirmatory PCR to determine whether the detected wsp gene Wolbachia belonged to that of the wPip strain using wPip-specific primers wPF (5′-CGACGTTAGTGGTGCAACATTTA-3′) and wPR (5′-AATAACGAGCACCAGCAAAGAGT-3′) [44] with the same PCR conditions as described previously but with the annealing temperature set to 56 °C. For primer optimization, DNA integrity was controlled by extraction of total DNA to make sure that a negative PCR result was not due to DNA degradation in the sample. DNA was extracted from 44 samples with the E.Z.N.A.® Tissue DNA Kit (Omega Bio-Tek, Inc., Norcross, GA, USA), and visual inspection of DNA integrity was done by gel electrophoresis. Extracted DNA and all PCR products were visualized on 1.8% agarose gel stained with GelRed® Nucleic Acid Gel Stain (Biotium, Fremont, CA, USA) (Fig. 2). A subset of PCR products was purified with ExoSAP-IT® (Thermo Fischer Scientific; Vilnius, Lithuania) and sequenced through Sanger sequencing (Macrogen; Amsterdam, The Netherlands) to validate the method and verify the results.
Fig. 2.
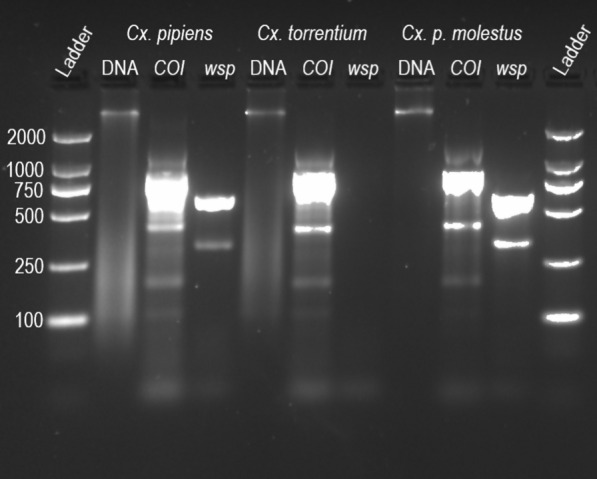
Representative agarose gel of extracted DNA and amplified PCR products. The extracted DNA (lanes 2, 5, and 7), amplified cytochrome oxidase subunit I (COI) gene (lanes 3, 6, and 9), and amplified wsp gene (lanes 4, 7, and 10) are shown for an individual of each of the species Cx. pipiens, Cx. torrentium, and Cx. pipiens molestus, respectively
Data analysis
All records were kept and analysed in Microsoft Excel 2016 (Microsoft; Redmond, CA, USA). Confidence intervals for Wolbachia prevalence were calculated assuming binomial distribution using the Wilson score interval through RStudio (RStudio team, Boston, USA). P-values to determine statistical significance for differences in Wolbachia prevalence between years were calculated using Fisher’s exact test with Bonferroni correction. Sequences of PCR fragments were analysed in the BioEdit sequence alignment editor version 7.2.5 [45].
Results
In total, 969 Culex mosquitoes (676 Cx. pipiens and 293 Cx. torrentium) were identified to species and tested for Wolbachia (Fig. 2). Wolbachia was present in 96.5% of the Cx. pipiens population (95% CI 94.8–97.6%) but could only be detected in two out of 293 Cx. torrentium individuals (0.68% prevalence, 95% CI 0.19–2.45%) (Table 1). Three of the Cx. pipiens that carried Wolbachia from each year were tested with primers specific to the wPip variant of wsp, of which all 27 were found to carry a wsp belonging to the wPip strain. In 2012, the prevalence of Wolbachia in Cx. pipiens was significantly lower than normal (Fisher’s exact test: P = 0.00455, OR: 0.389 CI [0.198–0.778], Bonferroni-corrected P = 0.041).
Table 1.
Results of the screening of Cx. pipiens and Cx. torrentium for Wolbachia
| Year | Species | Total tested | |||||
|---|---|---|---|---|---|---|---|
| Culex pipiens | Culex torrentium | ||||||
| Tested | Positive | % positive | Tested | Positive | % positive | ||
| 2010 | 93 | 92 | 98.9 | 49 | 0 | 0 | 142 |
| 2011 | 21 | 21 | 100 | 30 | 0 | 0 | 51 |
| 2012 | 208 | 190 | 91.3 | 71 | 0 | 0 | 279 |
| 2013 | 62 | 60 | 96.8 | 52 | 0 | 0 | 114 |
| 2014 | 6 | 5 | 83.3 | 2 | 0 | 0 | 8 |
| 2015 | 76 | 75 | 98.7 | 39 | 2 | 5.2 | 115 |
| 2016 | 30 | 30 | 100 | 30 | 0 | 0 | 60 |
| 2017 | 53 | 53 | 100 | 10 | 0 | 0 | 63 |
| 2018 | 127 | 126 | 99.2 | 10 | 0 | 0 | 137 |
| Total | 676 | 652 | 96.5 | 293 | 2 | 0.7 | 969 |
The mosquitoes were collected in central Sweden between 2010 and 2018. The prevalence of Wolbachia in Cx. pipiens differed significantly in year 2012 from the 9-year average (Fisher’s exact test: P = 0.00455, OR: 0.389 CI [0.198–0.778], Bonferroni-corrected P = 0.041). The differences for all other years are non-significant
Two Cx. torrentium were found to carry Wolbachia. Sequencing of the amplicons showed that the two partial wsp sequences were only 90% identical to each other. The wsp sequence from one of the Cx. torrentium individuals was very similar (> 99.8% identity) to the wsp of Wolbachia from Cx. pipiens (GenBank: KT964224.1), but also to isolates from the winter moth (Operophtera brumata: GenBank: KY587652.1), cabbage moth (Mamestra brassicae; GenBank: AB094375.1), and Toya propinqua (GenBank: KM386826.1). The other Cx. torrentium carried a Wolbachia whose wsp gene was highly similar (> 99.6% identity) to that of Wolbachia detected in several other insects, namely the spotted fritillary (Melitaea didyma; GenBank: MN322891.1), silverleaf whitefly (Bemisia tabaci; GenBank: AJ291379.1), azalea lace bug (Stephanitis pyrioides, GenBank: AB109622.1), Macrolophus pygmaeus (GenBank: FJ374283.1), and Amaurosoma flavipes (GenBank: JN601166.1), all of which carry Wolbachia from supergroup B. The sequencing results were confirmed by PCR using the wPip-specific wsp primers. This PCR amplified a correct fragment from only one of the two Wolbachia-positive Cx. torrentium (Fig. 3).
Fig. 3.
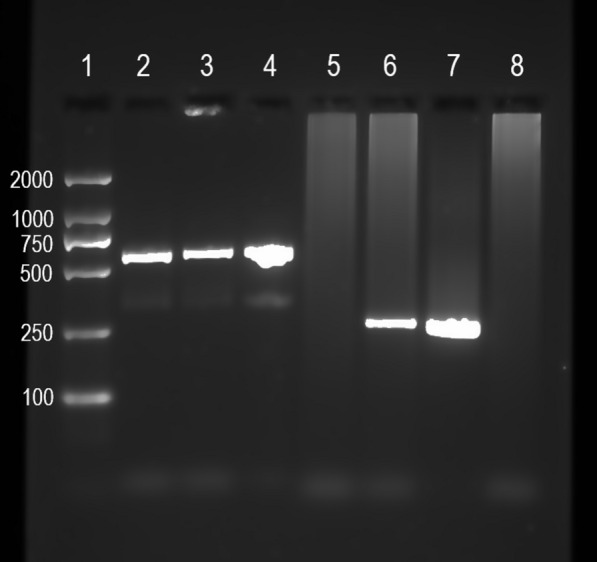
Gel showing amplification with general wsp primers (Cx. torrentium in lanes 2–3, Cx. pipiens molestus in lane 4), and with specific wPip primers (Cx. torrentium in lanes 5–6, Cx. pipiens molestus in lane 7). Lane 1 shows a size ladder and lane 8 shows a combined non-template control for general and specific wsp amplification
Discussion
We found that Wolbachia was highly prevalent in Cx. pipiens collected around the River Dalälven floodplains, while it was nearly absent from Cx. torrentium. This is in line with previous European studies investigating large samples of Cx. pipiens, with reported Wolbachia prevalence of 91% in western Russia [35], 95% in central Russia, 81% in Belarus [36], and 93% in Germany [37]. Raharimalala et al. [38] detected Wolbachia in nine out of nine tested adult Cx. pipiens and 26 out of 48 larvae collected in Belgium, which also supports the generally high prevalence of Wolbachia in this species. Interestingly, the different populations studied by Khrabrova et al. [36] had varying levels of Wolbachia prevalence, some with as few as 34.5% of individuals carrying the endosymbiont. Wolbachia is reported to approach fixation in most Cx. pipiens populations worldwide [46, 47], but this does not seem to hold true for all European populations.
Only Ricci et al. [48] have, to our knowledge, previously found Wolbachia in Cx. torrentium, after testing only two individuals collected in Italy. Raharimalala et al. [38], Leggewie et al. [37], Vinogradova et al. [35], and Khrabrova et al. [36] detected no Wolbachia in Cx. torrentium despite having tested 42 Belgian, 188 German, 321 Russian, and 853 Eastern European individuals, respectively. Our study, as well as the study by Ricci et al. [47], tested adult mosquitoes, while the four that failed to detect Wolbachia in Cx. torrentium tested field-collected larvae and pupae. Wolbachia is usually inherited and should thus be present in all life stages of the mosquito; however, life stage is still potentially an important consideration when screening for Wolbachia, both to avoid analysing siblings and to detect potential horizontal transmission.
Due to the low prevalence of Wolbachia in Cx. torrentium, we hypothesize that the two positive individuals or their recent ancestors acquired the infection horizontally. Transmission could potentially have occurred by feeding on the same plants as other arthropods [49, 50] or through arthropod parasites, such as through mites sometimes feeding on mosquitoes [51, 52]. Despite wsp being a poor marker of Wolbachia strain due to its tendency to recombine [53], the lineage of the Cx. torrentium whose wsp gene matched that of the wPip strain could have acquired its infection from a Cx. pipiens through their shared habitat and ecological niche. Alternative sources are also possible, since a highly similar wsp sequence has also been found in other Palearctic insects. Further studies on the mechanisms for horizontal Wolbachia transmission involving mosquitoes are needed to fully explain the occasional spread of Wolbachia to Cx. torrentium.
The restriction of SINV outbreaks to Northern Europe has been suggested to be connected to the relatively higher abundance of the competent vector species Cx. torrentium in SINV-endemic regions [31, 32]. Under laboratory conditions, Cx. torrentium is significantly more susceptible to SINV infection than Cx. pipiens [34, 54]. The presence of Wolbachia in Cx. pipiens may contribute to its lower susceptibility to SINV. Such reduction in vector competence is often seen when transferring a novel Wolbachia strain into a mosquito species that is naturally Wolbachia-free or naturally carries a different strain [11, 55–58], but the impact of a naturally occurring Wolbachia infection (i.e., native infection) is not as clear, with reports of both reduced vector competence [13, 14, 59] and no observed effect [60–63]. No vector competence studies have been done on the role of Wolbachia in alphavirus transmission in Culex mosquitoes. With relatively few data to extrapolate from, empirical investigation is needed to evaluate the impact of Wolbachia on the SINV transmission cycle.
Conclusions
Our study, performed in a SINV-endemic region of Sweden, confirmed previously reported general patterns of Wolbachia infection in Culex mosquitoes, with most Cx. pipiens and very few Cx. torrentium carrying the endosymbiont, which potentially has implications for their differences in vector competence. Our findings, paired with the specific conditions under which SINV is transmitted in Sweden, prompt more research into Wolbachia’s role in the SINV transmission cycle as well as the horizontal routes of Wolbachia transmission among mosquitoes.
Acknowledgements
The authors are grateful to Jan O. Lundström for providing mosquito material, and for the financial support received from the funders described above.
Abbreviations
- CDC
Centers for Disease Control and Prevention
- CHIKV
Chikungunya virus
- COI
Cytochrome oxidase subunit I
- Cx.
Culex
- JEV
Japanese encephalitis virus
- PBS
Phosphate buffered saline
- PCR–RFLP
Polymerase chain reaction–restriction fragment length polymorphism
- SINV
Sindbis virus
- WNV
West Nile virus
Authors’ contributions
JCH conceptualized, supervised, and administered the project, acquired mosquito material and funding, and reviewed and contributed to the manuscript. AB conducted DNA extraction, Wolbachia screening and species verification, and wrote the manuscript. Both authors have read and approved the final manuscript.
Funding
Open access funding provided by Uppsala University. The research was funded by the Swedish Society for Medical Research, the Carl Trygger Foundation, and the European Union’s Horizon 2020 research and innovation programme under Grant No. 874735 (VEO). None of the funding bodies had any role in the design of the study or collection, analysis, and interpretation of data, or in drafting of the manuscript.
Availability of data and materials
For amplicon sequences of the Wolbachia detected in Cx. torrentium and Cx. pipiens, the data sets generated and/or analysed during the current study are available in the GenBank repository, accession numbers MW622245-MW622247. All other data sets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Alexander Bergman, Email: alexander.brg@outlook.com.
Jenny C. Hesson, Email: jenny.hesson@imbim.uu.se
References
- 1.Ramirez JL, Souza-Neto J, Cosme RT, Rovira J, Ortiz A, Pascale JM, et al. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl Trop Dis. 2012;6:e1561. doi: 10.1371/journal.pntd.0001561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sim S, Jupatanakul N, Dimopoulos G. Mosquito immunity against arboviruses. Viruses. 2014;6:4479–4504. doi: 10.3390/v6114479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zouache K, Fontaine A, Vega-Rua A, Mousson L, Thiberge J-M, Lourenco-De-Oliveira R, et al. Three-way interactions between mosquito population, viral strain and temperature underlying chikungunya virus transmission potential. Proc R Soc B. 2014;281:20141078. doi: 10.1098/rspb.2014.1078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ciota AT, Keyel AC. The role of temperature in transmission of zoonotic arboviruses. Viruses. 2019;11:1013. doi: 10.3390/v11111013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zug R, Hammerstein P. Still a host of hosts for Wolbachia: analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE. 2012;7:e38544. doi: 10.1371/journal.pone.0038544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Landmann F. The Wolbachia endosymbionts. Microbiol Spectr. 2019 doi: 10.1128/microbiolspec.BAI-0018-2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol. 2008;6:741–751. doi: 10.1038/nrmicro1969. [DOI] [PubMed] [Google Scholar]
- 8.Hoffmann AA. Incompatible mosquitoes. Nature. 2005;436:189. doi: 10.1038/436189a. [DOI] [PubMed] [Google Scholar]
- 9.Sinkins SP, Walker T, Lynd AR, Steven AR, Makepeace BL, Godfray JHC, et al. Wolbachia variability and host effects on crossing type in Culex mosquitoes. Nature. 2005;436:257–60. doi: 10.1038/nature03629. [DOI] [PubMed] [Google Scholar]
- 10.Bonneau M, Atyame C, Beji M, Justy F, Cohen-Gonsaud M, Sicard M, et al. Culex pipiens crossing type diversity is governed by an amplified and polymorphic operon of Wolbachia. Nat Commun. 2018;9:319. doi: 10.1038/s41467-017-02749-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Walker T, Johnson PH, Moreira LA, Iturbe-Ormaetxe I, Frentiu FD, McMeniman CJ, et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature. 2011;476:450–453. doi: 10.1038/nature10355. [DOI] [PubMed] [Google Scholar]
- 12.Hoffmann AA, Montgomery BL, Popovici J, Iturbe-Ormaetxe I, Johnson PH, Muzzi F, et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature. 2011;476:454–457. doi: 10.1038/nature10356. [DOI] [PubMed] [Google Scholar]
- 13.Tsai C-H, Chen T-H, Lin C, Shu P-Y, Su C-L, Teng H-J. The impact of temperature and Wolbachia infection on vector competence of potential dengue vectors Aedes aegypti and Aedes albopictus in the transmission of dengue virus serotype 1 in southern Taiwan. Parasites Vectors. 2017;10(1):551. doi: 10.1186/s13071-017-2493-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Glaser RL, Meola MA. The native Wolbachia endosymbionts of Drosophila melanogaster and Culex quinquefasciatus increase host resistance to West Nile virus infection. PLoS ONE. 2010;5:e11977. doi: 10.1371/journal.pone.0011977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.van den Hurk AF, Ritchie SA, Mackenzie JS. Ecology and geographical expansion of Japanese encephalitis virus. Annu Rev Entomol. 2009;54:17–35. doi: 10.1146/annurev.ento.54.110807.090510. [DOI] [PubMed] [Google Scholar]
- 16.Reuben R, Tewari SC, Hiriyan J, Akiyama J. Illustrated keys to species of Culex (Culex) associated with Japanese encephalitis in Southeast Asia (Diptera: Culicidae) Mosq Syst. 1994;26:75–96. [Google Scholar]
- 17.Fros JJ, Miesen P, Vogels CB, Gaibani P, Sambri V, Martina BE, et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health. 2015;1:31–36. doi: 10.1016/j.onehlt.2015.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fortuna C, Remoli ME, Di Luca M, Severini F, Toma L, Benedetti E, et al. Experimental studies on comparison of the vector competence of four Italian Culex pipiens populations for West Nile virus. Parasites Vectors. 2015;8:463. doi: 10.1186/s13071-015-1067-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Francy DB, Jaenson TG, Lundström JO, Schildt EB, Espmark A, Henriksson B, et al. Ecologic studies of mosquitoes and birds as hosts of Ockelbo virus in Sweden and isolation of Inkoo and Batai viruses from mosquitoes. Am J Trop Med Hyg. 1989;41:355–363. doi: 10.4269/ajtmh.1989.41.355. [DOI] [PubMed] [Google Scholar]
- 20.Jöst H, Bialonski A, Storch V, Günther S, Becker N, Schmidt-Chanasit J. Isolation and phylogenetic analysis of Sindbis viruses from mosquitoes in Germany. J Clin Microbiol. 2010;48:1900–1903. doi: 10.1128/JCM.00037-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Leggewie M, Badusche M, Rudolf M, Jansen S, Börstler J, Krumkamp R, et al. Culex pipiens and Culex torrentium populations from Central Europe are susceptible to West Nile virus infection. One Health. 2016;2:88–94. doi: 10.1016/j.onehlt.2016.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lundström JO, Hesson JC, Schäfer ML, Östman Ö, Semmler T, Bekaert M, et al. Sindbis virus polyarthritis outbreak signalled by virus prevalence in the mosquito vectors. PLoS Negl Trop Dis. 2019;13:e0007702. doi: 10.1371/journal.pntd.0007702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Adouchief S, Smura T, Sane J, Vapalahti O, Kurkela S. Sindbis virus as a human pathogen—epidemiology, clinical picture and pathogenesis. Rev Med Vir. 2016;26:221–241. doi: 10.1002/rmv.1876. [DOI] [PubMed] [Google Scholar]
- 24.Storm N, Weyer J, Markotter W, Kemp A, Leman PA, Dermaux-Msimang V, et al. Human cases of Sindbis fever in South Africa, 2006–2010. Epidemiol Infect. 2013;142:234–238. doi: 10.1017/S0950268813000964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jupp PG, Blackburn NK, Thompson DL, Meenehan GM. Sindbis and West Nile virus infections in the Witwatersrand-Pretoria region. S Afr Med J. 1986;70:218–220. [PubMed] [Google Scholar]
- 26.Brummer-Korvenkontio M, Vapalahti O, Kuusisto P, Saikku P, Manni T, Koskela P, et al. Epidemiology of Sindbis virus infections in Finland 1981–96: possible factors explaining a peculiar disease pattern. Epidemiol Infect. 2002;129:335–345. doi: 10.1017/S0950268802007409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sane J, Guedes S, Kurkela S, Lyytikäinen O, Vapalahti O. Epidemiological analysis of mosquito-borne Pogosta disease in Finland, 2009. Euro Surveill. 2010;15:19462. doi: 10.2807/ese.15.02.19462-en. [DOI] [PubMed] [Google Scholar]
- 28.Lundström JO, Vene S, Espmark Å, Engvall M, Niklasson B. Geographical and temporal distribution of Ockelbo disease in Sweden. Epidemiol Infect. 1991;106:567–574. doi: 10.1017/S0950268800067637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bergqvist J, Forsman O, Larsson P, Näslund J, Lilja T, Engdahl C, et al. Detection and isolation of Sindbis virus from mosquitoes captured during an outbreak in Sweden, 2013. Vector Borne Zoonotic Dis. 2015;15:133–140. doi: 10.1089/vbz.2014.1717. [DOI] [PubMed] [Google Scholar]
- 30.Ahlm C, Eliasson M, Vapalahti O, Evander M. Seroprevalence of Sindbis virus and associated risk factors in northern Sweden. Epidemiol Infect. 2013;142:1559–1565. doi: 10.1017/S0950268813002239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hesson JC, Östman Ö, Schäfer M, Lundström JO. Geographic distribution and relative abundance of the sibling vector species Culex torrentium and Culex pipiens in Sweden. Vector Borne Zoonotic Dis. 2011;11:1383–1389. doi: 10.1089/vbz.2011.0630. [DOI] [PubMed] [Google Scholar]
- 32.Hesson JC, Rettich F, Merdić E, Vignjević G, Östman Ö, Schäfer M, et al. The arbovirus vector Culex torrentium is more prevalent than Culex pipiens in northern and central Europe. Med Vet Entomol. 2013;28:179–186. doi: 10.1111/mve.12024. [DOI] [PubMed] [Google Scholar]
- 33.Hesson JC, Verner-Carlsson J, Larsson A, Ahmed R, Lundkvist Å, Lundström JO. Culex torrentium mosquito role as major enzootic vector defined by rate of Sindbis virus infection, Sweden, 2009. Emerg Infect Dis. 2015;21:875–878. doi: 10.3201/eid2105.141577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lundström JO, Niklasson B, Francy DB. Swedish Culex torrentium and Cx.pipiens (Diptera: Culicidae) as experimental vectors of Ockelbo virus. J Med Entomol. 1990;27:561–3. doi: 10.1093/jmedent/27.4.561. [DOI] [PubMed] [Google Scholar]
- 35.Vinogradova EB, Shaikevich EV, Ivanitsky AV. A study of the distribution of Culex pipiens complex (Insecta: Diptera: Culicidae) mosquitoes in the European part of Russia by molecular methods of identification. Comp Cytogenet. 2007;1:129–138. [Google Scholar]
- 36.Khrabrova NV, Bukhanskaya ED, Sibataev AK, Volkova TV. The distribution of strains of endosymbiotic bacteria Wolbachia pipientis in natural populations of Culex pipiens mosquitoes (Diptera: Culicidae) Eur Mosq Bull. 2009;27:18–22. [Google Scholar]
- 37.Leggewie M, Krumkamp R, Badusche M, Heitmann A, Jansen S, Schmidt-Chanasit J, et al. Culex torrentium mosquitoes from Germany are negative for Wolbachia. Med Vet Entomol. 2017;32:115–120. doi: 10.1111/mve.12270. [DOI] [PubMed] [Google Scholar]
- 38.Raharimalala FN, Boukraa S, Bawin T, Boyer S, Francis F. Molecular detection of six (endo-) symbiotic bacteria in Belgian mosquitoes: first step towards the selection of appropriate paratransgenesis candidates. Parasitol Res. 2015;115:1391–1399. doi: 10.1007/s00436-015-4873-5. [DOI] [PubMed] [Google Scholar]
- 39.Schäfer M, Wahlqvist P, Lundström JO. Stickmyggmängder vid nedre Dalälven [Amounts of mosquitoes at lower Dalälven]. Biologisk Myggkontroll inom NEDAB. 2018. http://www.utslappshandel.se/upload/stod-i-miljoarbetet/remisser-och-yttranden/remisser-2018/ansokan-mygg-nedre-dalalven/bilaga5-stickmyggmangder-vid-nedre-dalalven.pdf. Accessed 22 Oct 2020.
- 40.Becker N, Petric D, Zgomba M, Boase C, Dahl C, Lane J, et al. Mosquitoes and their control. 2. Heidelberg: Springer; 2010. [Google Scholar]
- 41.Hesson JC, Lundström JO, Halvarsson P, Erixon P, Collado A. A sensitive and reliable restriction enzyme assay to distinguish between the mosquitoes Culex torrentium and Culex pipiens. Med Vet Entomol. 2010;24:142–149. doi: 10.1111/j.1365-2915.2010.00871.x. [DOI] [PubMed] [Google Scholar]
- 42.Hesson JC, Schäfer M, Lundström JO. First report on human-biting Culex pipiens in Sweden. Parasit Vectors. 2016;9:632. doi: 10.1186/s13071-016-1925-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhou W, Rousset F, O’Neill S. Phylogeny and PCR–based classification of Wolbachia strains using wsp gene sequences. Proc R Soc Lond B. 1998;265:509–515. doi: 10.1098/rspb.1998.0324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Calvitti M, Moretti R, Skidmore AR, Dobson SL. Wolbachia strain wPip yields a pattern of cytoplasmic incompatibility enhancing a Wolbachia-based suppression strategy against the disease vector Aedes albopictus. Parasites Vectors. 2012;5:254. doi: 10.1186/1756-3305-5-254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95–98. [Google Scholar]
- 46.Duron O, Lagnel J, Raymond M, Bourtzis K, Fort P, Weill M. Transposable element polymorphism of Wolbachia in the mosquito Culex pipiens: evidence of genetic diversity, superinfection and recombination. Mol Ecol. 2005;14:1561–1573. doi: 10.1111/j.1365-294X.2005.02495.x. [DOI] [PubMed] [Google Scholar]
- 47.Dumas E, Atyame CM, Milesi P, Fonseca DM, Shaikevich EV, Unal S, et al. Population structure of Wolbachia and cytoplasmic introgression in a complex of mosquito species. BMC Evol Biol. 2013;13:181. doi: 10.1186/1471-2148-13-181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ricci I, Cancrini G, Gabrielli S, D’amelio S, Favia G. Searching for Wolbachia (Rickettsiales: Rickettsiaceae) in mosquitoes (Diptera: Culicidae): large polymerase chain reaction survey and new identifications. J Med Entomol. 2002;39:562–7. doi: 10.1603/0022-2585-39.4.562. [DOI] [PubMed] [Google Scholar]
- 49.Li S-J, Ahmed MZ, Lv N, Shi P-Q, Wang X-M, Huang J-L, et al. Plantmediated horizontal transmission of Wolbachia between whiteflies. ISME J. 2016;11:1019–1028. doi: 10.1038/ismej.2016.164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sintupachee S, Milne JR, Poonchaisri S, Baimai V, Kittayapong P. Closely related Wolbachia strains within the pumpkin arthropod community and the potential for horizontal transmission via the plant. Microb Ecol. 2006;51:294–301. doi: 10.1007/s00248-006-9036-x. [DOI] [PubMed] [Google Scholar]
- 51.Cook JM, Butcher RDJ. The transmission and effects of Wolbachia bacteria in parasitoids. Res Popul Ecol. 1999;41:15–28. doi: 10.1007/PL00011978. [DOI] [Google Scholar]
- 52.Brown AN, Lloyd VK. Evidence for horizontal transfer of Wolbachia by a Drosophila mite. Exp Appl Acarol. 2015;66:301–311. doi: 10.1007/s10493-015-9918-z. [DOI] [PubMed] [Google Scholar]
- 53.Baldo L, Lo N, Werren JH. Mosaic nature of the Wolbachia surface protein. JB. 2005;187:5406–5418. doi: 10.1128/JB.187.15.5406-5418.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lwande OW, Näslund J, Lundmark E, Ahlm K, Ahlm C, Bucht G, et al. Experimental infection and transmission competence of Sindbis virus in Culex torrentium and Culex pipiens mosquitoes from northern Sweden. Vector Borne Zoonotic Dis. 2019;19:128–133. doi: 10.1089/vbz.2018.2311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, Hedges LM, et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, chikungunya, and Plasmodium. Cell. 2009;139:1268–1278. doi: 10.1016/j.cell.2009.11.042. [DOI] [PubMed] [Google Scholar]
- 56.Blagrove MSC, Arias-Goeta C, Di Genua C, Failloux A-B, Sinkins SP. A WolbachiawMel transinfection in Aedesalbopictus is not detrimental to host fitness and inhibits chikungunya virus. PLoS Negl Trop Dis. 2013;7:e2152. doi: 10.1371/journal.pntd.0002152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ant TH, Herd CS, Geoghegan V, Hoffmann AA, Sinkins SP. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathog. 2018;14:e1006815. doi: 10.1371/journal.ppat.1006815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Caragata EP, Rocha MN, Pereira TN, Mansur SB, Dutra HLC, Moreira LA. Pathogen blocking in Wolbachia-infected Aedes aegypti is not affected by Zika and dengue virus co-infection. PLoS Negl Trop Dis. 2019;13:e0007443. doi: 10.1371/journal.pntd.0007443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Mousson L, Zouache K, Arias-Goeta C, Raquin V, Mavingui P, Failloux A-B. The native Wolbachia symbionts limit transmission of dengue virus in Aedes albopictus. PLoS Negl Trop Dis. 2012;6:e1989. doi: 10.1371/journal.pntd.0001989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mousson L, Martin E, Zouache K, Madec Y, Mavingui P, Failloux AB. Wolbachia modulates chikungunya replication in Aedes albopictus. Mol Ecol. 2010;19:1953–1964. doi: 10.1111/j.1365-294X.2010.04606.x. [DOI] [PubMed] [Google Scholar]
- 61.Micieli MV, Glaser RL. Somatic Wolbachia (Rickettsiales: Rickettsiaceae) levels in Culex quinquefasciatus and Culex pipiens (Diptera: Culicidae) and resistance to West Nile virus infection. J Med Entom. 2014;51:189–199. doi: 10.1603/ME13152. [DOI] [PubMed] [Google Scholar]
- 62.Ahmad NA, Vythilingam I, Lim YAL, Zabari NZAM, Lee HL. Detection of Wolbachia in Aedes albopictus and their effects on chikungunya virus. Am J Trop Med Hyg. 2016;96:148–156. doi: 10.4269/ajtmh.16-0516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Joanne S, Vythilingam I, Teoh B-T, Leong C-S, Tan K-K, Wong M-L, et al. Vector competence of Malaysian Aedes albopictus with and without Wolbachia to four dengue virus serotypes. Trop Med Int Health. 2017;22:1154–1165. doi: 10.1111/tmi.12918. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
For amplicon sequences of the Wolbachia detected in Cx. torrentium and Cx. pipiens, the data sets generated and/or analysed during the current study are available in the GenBank repository, accession numbers MW622245-MW622247. All other data sets used and/or analysed during the current study are available from the corresponding author on reasonable request.