

Abstract
Oxysterols, or cholesterol oxidation products, are naturally occurring lipids which regulate the physiology of cells, including those of the immune system. In contrast to effects that are mediated through nuclear receptors or by epigenetic mechanism, which take tens of minutes to occur, changes in the activities of cell-surface receptors caused by oxysterols can be extremely rapid, often taking place within subsecond timescales. Such cell-surface receptor effects of oxysterols allow for the regulation of fast cellular processes, such as motility, secretion and endocytosis. These cellular processes play critical roles in both the innate and adaptive immune systems. This review will survey the two broad classes of cell-surface receptors for oxysterols (G-protein coupled receptors (GPCRs) and ion channels), the mechanisms by which cholesterol oxidation products act on them, and their presence and functions in the different cell types of the immune system. Overall, this review will highlight the potential of oxysterols, synthetic derivatives and their receptors for physiological and therapeutic modulation of the immune system.
Keywords: oxysterols, ion channels, immune response, pharmacology, structure-function, inflammation, autoimmunity, infectious diseases
1. Introduction
Oxysterols are cholesterol oxidation products, which can be absorbed from the diet, or generated by auto-oxidation or by enzymatic mechanisms. Oxysterols result from oxidation of cholesterol on the sterol rings, the side chain, or both [1]. This generates a diverse range of oxysterol congeners that have distinct biophysical properties. In addition to being reaction intermediates in the synthesis of bile-acids and steroid hormones, many oxysterols are biologically active signalling molecules, regulating diverse cellular processes. Consequently, the dysregulation of oxysterol production and action is associated with a range of diseases, including cancers [2], atherosclerosis [3], age-related macular degeneration, neurodegeneration and osteoporosis [1]. The focus of the current review are the roles of oxysterols in immune system physiology and pathology [4,5,6].
Oxysterols exert their biological effects in a variety of ways, potentially including mechanisms that are yet to be elucidated. Most oxysterols have direct biophysical effects on lipid bilayers, including the modification of cholesterol organization and associated membrane subdomains [7]. Several side-chain oxysterols, including 25-hydroxycholesterol (25-HC, synthesized by cholesterol 25-hydroxylase) and 27-hydroxycholesterol (27-HC, generated by cytochrome p450 oxidase 27A1, CYP27A1), play roles in the innate immune system, blocking the infection and replication of viruses and bacteria. For example, 25-HC hinders the replication of SARS-CoV-2 by inhibiting membrane fusion [8]. It also alters the mechanical properties of lipid membranes, leading to local variations in stiffness, which can influence the entry and egress of pathogens [9]. Ring-modified oxysterols, such as 7-ketocholesterol (7-KC), also inhibit viral replication and infection, but exert robust cytotoxic effects on host cells [10]. Oxysterols can indirectly modify membrane properties: 25HC activates acyl-CoA:cholesterol acyltransferase (ACAT) in macrophages, leading to the rapid internalization of an accessible pool of cholesterol, thereby blocking infection by Shigella flexneri or Listeria monocytogenes [11].
Since oxysterols are hydrophobic, they readily diffuse into cells to interact with intracellular receptors. Oxysterol-binding proteins (OSBPs) and OSBP-related proteins (ORPs) are a family of cytosolic receptors for oxysterols. In humans, the OSBP/ORP family is encoded by 12 distinct genes, with all members possessing an OSBP-related ligand-binding domain and the majority having a pleckstrin-homology domain near the N-terminus, that allows for interaction with phosphatidtidylinositol phosphate lipids. It is thought that all OSBPs/ORPs participate in the transfer of sterols and other small molecules at sites of membrane contact between different organelles [12], but additional roles have also been identified, some of which are specific to particular members [13,14]. Several of these roles were first identified in cells derived from the immune system. Inositol 1,4,5-trisphosphate (IP3) is generated upon the activation of G protein-coupled receptors (GPCRs) or receptor tyrosine kinases, through the action of phospholipase C (PLC) isozymes on the lipid phosphatidylinositol-4,5-bisphosphate (PIP2). IP3 is a second messenger that releases Ca2+ from intracellular stores, such as the endoplasmic reticulum (ER), via IP3 receptor (IP3R) channels. Oxysterols exert congener- and cell-type-selective effects on cytoplasmic Ca2+ levels [15]. In T cell acute lymphoblastic leukemia cells, ORP4L assembles the receptor CD3ε, its transducers Gαq/11 and the effector PLC-β3 into a signalling complex, generating IP3, which triggers Ca2+ release via IP3Rs. This increases mitochondrial matrix Ca2+, thereby activating oxidative respiration [16]. In the Jurkat T-lymphoma cell line, ORP4L interacts directly with IP3Rs, enhancing IP3-induced Ca2+ release [17]. In mouse bone marrow-derived mast cells, ORP9S is phosphorylated by a type 2 phosphoinositide-dependent protein kinase (PKD-2, most likely a protein kinase C-β (PKC-β)) and inhibits phosphorylation of Akt/protein kinase B at S473 [18]. OSBPs/ORPs play roles in viral infection and replication. For example, the drug itraconazole inhibits the replication of a diverse range of viruses, including hepatitis C and poliovirus, via the antagonism of the lipid transfer activities of OSBP and of ORP4 [19].
A key class of intracellular receptors for oxysterols exert their cellular effects through changes in the transcription of target genes [20]. Archetypal nuclear receptors for oxysterols are the liver X receptors (LXRs), which exist as heterodimers with retinoid X receptors (RXRs). Of the two human LXRs, LXRα (nuclear receptor subfamily 1, group H, member 3 (NR1H3)) is abundant in macrophages, liver, intestine, adipose tissue, lung, kidney and adrenal gland, whereas LXRβ (NR1H2) is ubiquitous [21]. Binding of certain side-chain oxysterols (22(R)-HC, 24(S)-HC, 25-HC, or 27-HC) elicits conformational changes that promote dissociation of LXR-RXR complexes from transcriptional co-repressors, and stimulates LXR-RXR interaction with the sterol response elements (SREs) of target genes. These mechanisms increase the transcription of genes involved in cholesterol clearance (ATP-binding cassette A1 and G1, CYP7A1, apolipoprotein E) and those which regulate multiple components of the immune system [4].
There are three widely expressed members of the SRE-binding protein family (SREBP1a, SREBP1b and SREBP2), which are transcription factors tethered to the ER via a transmembrane-spanning domain. In the ER, SREBPs interact with SREBP cleavage-activating proteins (SCAPs). When the levels of cholesterol and oxysterols are low, SCAP transports SREBPs to the Golgi apparatus where it is cleaved by two resident proteases, releasing an N-terminal domain. This domain translocates to the nucleus, where it binds to SREs in the promoters of target genes [22]. These target genes include those involved in fatty acid metabolism, cholesterol transport and immune responses [6]. Cholesterol, or certain oxysterols (most notably 25-HC), bind to the ER-resident proteins insulin-induced gene-1 (INSIG-1) or INSIG-2, promoting interaction with SCAP [23], preventing its translocation to the Golgi and activation of SREBP. Consequently, cholesterol and some oxysterols are negative modulators of SREBP-dependent transcription.
The RXR-related orphan receptor (ROR) family members α and γ can bind to certain oxysterols, particularly 7-position substituted congeners, with high affinity. This activates the transcription of target genes, including those involved in immunity. Other oxysterols, such as 24S-hydroxycholesterol (24S-HC) and 24S,25-epoxycholesterol, suppress the activities of these transcription factors [24].
Several oxysterols are selective estrogen receptor modulators. For example, 27-HC binds to both the α- and β-estrogen receptors, inducing transcription of their target genes [25]. In contrast, 27-HC antagonises the protective effects of estrogen against atherosclerosis in a diet-induced mouse model of hypercholesterolaemia [26].
Certain oxysterols influence the transcription of target genes via epigenetic mechanisms. During investigations of the effects of high extracellular glucose on the regulation of genes involved in non-alcoholic fatty liver disease, it was discovered that elevated 25-HC promoted methylation of CpG islands, acting as a direct, endogenous agonist of DNA methyltransferase 1 (DNMT1) [27]. Genes whose transcription is regulated in this manner include voltage-gated Ca2+ channel subunits, Ca2+-calmodulin kinases, those involved in lipid metabolism, inflammation and MAPK signalling. Within the immune system, cholesterol 25-hydroxylase, the enzyme that produces 25-HC, is particularly abundant in macrophages, and this oxysterol suppresses IgA synthesis [28]. In addition, 25-HC can be sulfated at its 3-position by sulfotransferase 2B1b to generate 25-HC3S, which can be further sulfated by sulfotransferase 2A1 [29]. In contrast to 25-HC, 25-HC3S is a potent antagonist of DMNT-1, -3a and -3b, and opposes its actions on target genes [27,30,31]. In a lipopolysaccharide-induced model of acute lung injury, 25-OHC concentrations are elevated, potentially promoting inflammation and the recruitment of leukocytes [32]. Since 25-HC3S has beneficial effects in models of non-alcoholic fatty liver disease, organ damage and inflammation, and has been evaluated favourably in clinical trials, further elucidation of the roles of this oxysterol metabolite in the regulation of immune cells will be of great value [31].
In addition to the direct effects of oxysterols on nuclear receptors, there is also considerable cross talk between signalling pathways modulated by cholesterol oxidation products and those regulated by other bioactive sterols. Some of these sterols are important modulators of the immune system. For example, active vitamin D (1,25-dihydroxycholesterol) inhibits the expression of CYP7A (cholesterol 7 hydroxlase α) by promoting the formation of a complex between the vitamin D receptor and LXRα [33]. The enzyme 11β-hydroxysteroid dehydrogenase type 1 (11β-HSD1) can convert 7-KC to 7β-hydroxycholesterol (7β-HC), and also interconverts active cortisol (a glucocorticoid hormone) into inactive cortisone. In adipocytes, co-incubation with 7-KC repressed, and 7β-HC enhanced, the biological effects of cortisol through a mechanism that involved changing the direction of substrate flux through the 11β-HSD1 and 11β-HSD2 enzyme pair [34].
Despite the potential for therapeutic exploitation of nuclear receptor-mediated oxysterol signalling in the regulation of immune cell biology, this does have two key limitations. Firstly, transcriptional mechanisms are slow, with a lag of tens of minutes to hours between the initial stimulus (change in the levels of a particular oxysterol) and the cellular response. This may be unsuitable for the control of cellular processes that can occur over much shorter timescales, such as motility, endocytosis and secretion. Secondly, in terms of the development of oxysterol-based immunomodulatory drugs, LXR agonists act on the liver to increase the expression of SREBP1c, which promotes the clinically unfavourable condition of hypertriglyceridemia [6,35,36]. Targeting the main focus of the current review, cell-surface receptors for oxysterols might provide effective solutions to these limitations. These targets can be divided into two classes: G protein-coupled receptors (GPRs) and ion channels. Evidence for the presence of these proteins in different immune cells was derived from two key sources: analyses of four independent transcriptomic databases from human immune cells (Supplemental Figure S1) [37,38,39] and a review of the scientific literature (Supplemental Figure S2).
2. G Protein-Coupled Receptors
G protein-coupled receptors (GPCRs) are predominantly located in the cell-surface membrane. They have a characteristic 7-transmembrane (7TM) topology and regulate the activities of second-messenger-producing enzymes. Figure 1 summarizes some of the properties of the three types of GPCR that are present in the mammalian immune system and which interact with oxysterols. Although there are four types of GPCR that are known to be modulated by oxysterols, one of these, GPR17, is undetectable in the immune cell types that we investigated, see Supplemental Figure S1. Figure 2 provides an overview of the distribution of these receptors among cells of the immune system.
Figure 1.
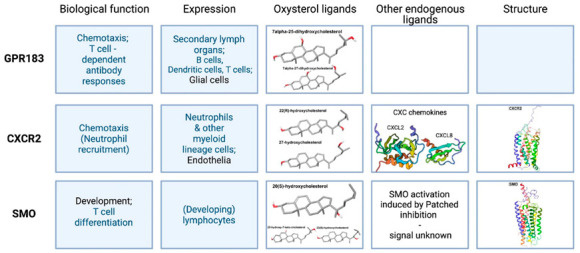
Overview of the biological functions and distribution (Expression)s of oxysterol activated GPCRs in cells of the immune system. The structures of their endogenous oxysterol ligands, details of other endogenous ligands and, where available, models of their three-dimensional structures of these receptors are also shown. This image was created using BioRender (BioRender.com).
Figure 2.
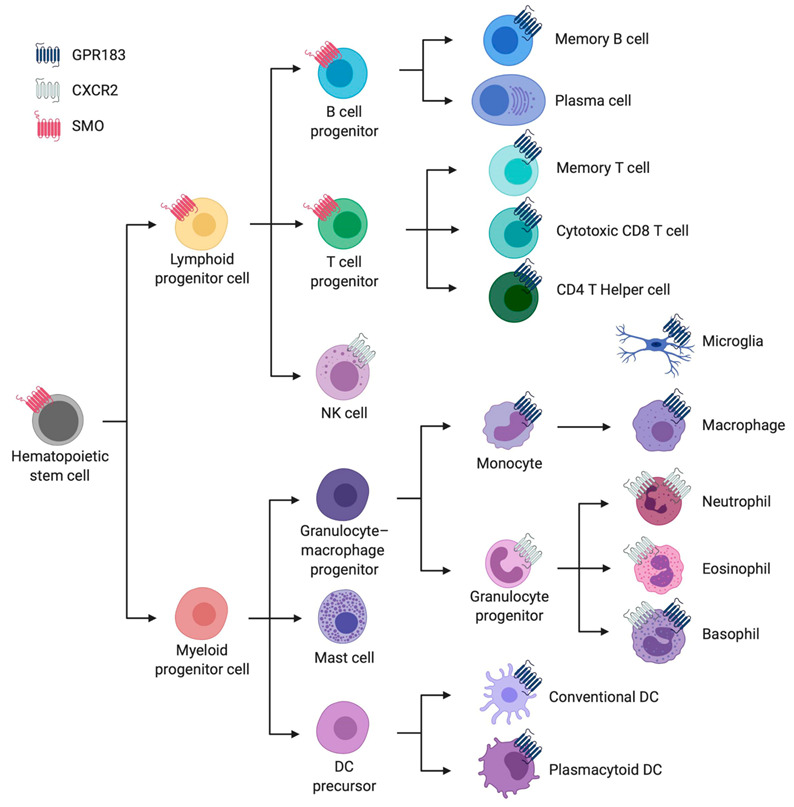
Overview of the main immune cell types from the hematopoietic lineage with an abstraction of their cellular expression of the three GPCRs discussed in this article. These GPCRs are represented by three distinct colours: purple for GPR183, light grey for CXCR2 and red for SMO. This overview was created in BioRender (BioRender.com) and is based on a consensus from four independent transcriptomic data sets from human immune cells [37,38,39], as summarized in Supplemental Figure S1.
2.1. Epstein–Barr Virus-Induced Receptor 2, or GPR183
2.1.1. Cellular Expression Pattern and Overall Biological Function
Epstein–Barr virus-induced receptor 2 (EBI2), or GPR183, is mainly expressed by immune cells and related tissues. Secondary lymph organs have an especially abundant expression, but the lungs and the gastrointestinal tract also show comparably high expression levels [40].
The best-established role for the receptor lies in the positioning of cells in secondary lymph organs, and especially the facilitation of T cell-dependent antibody responses. Through the interplay and differential expression of enzymes in the secondary lymphoid organs, a gradient of 7α,25-OHC is created, with the highest concentration at the B/T-cell border and the outer follicular regions [41]. B cells undergo the sequential up and down regulation of GPR183, in parallel with the expression of several other chemotactic GPCRs. Activation of B cells by B-cell receptor (BCR) binding leads to an upregulation of GPR183. This, in turn, results in the lymphocyte alignment on the T-cell border, a high ligand concentration region, by integrated signalling via both GPR183 and C-C chemokine receptor 7 (CCR7) [42,43]. Here, they can receive secondary differentiation signals from T-helper (Th) cells. If they are T cell-stimulated, GPR183 expression is downregulated, which allows the B cells to move to the germinal centres in the B-cell zone for antibody refinement or escape to the circulation. Expression in the germinal centre is generally lowered, but the differential expression of GPR183 still correlates with localization in the light or dark zone [43,44]. Memory B cells, on the other hand, stay on the fringes of the follicular area, due to high GPR183, normal CCR7 levels and downregulated CCR5 expression. By a similar mechanism, corresponding specialized TH cells are localized to the same region [45].
T cells also receive cues from oxysterols via GPR183. In their development process, GPR183 influences central tolerance, especially in CD4 single positive (SP) cells. It regulates CD4+ effector and regulator cell generation by increasing encounters with antigen-presenting cells, such as dendritic cells (DCs), and thus increases the number of encounters with different, rare self-antigens [46]. GPR183 remains highly expressed on CD4+ T cells compared to CD8+ T cells and also enhances CD4–DC encounters in secondary lymph organs. Thus, it enables the early differentiation of CD4+ helper cells [47], which in turn facilitates more efficient CD8+ T- and B-cell responses. In exemplum, GPR183 stimulates T follicular helper cell (TfH) differentiation and then induces secondary migration to the follicles [48].
Bringing it back full circle, DCs also express GPR183, and react to oxysterol cues by migration. This has mainly been studied in context of correct positioning in the spleen, in the bridging channels unto activation [49]. In activated DCs GPR183 is also responsible for localization at the outer T zone, where they determine TH cell differentiation to TfH cells [50]. GPR183 expression has also been found relevant in plasmacytoid (pDC) and myeloid (mDC) subsets, where it downregulates interferon responses upon Toll-like receptor (TLR) binding, as well as migration towards the spleen [51]. These regulatory effects are also supported by an observed interplay between tuberculosis infection and type two diabetes, where reduced GPR183 expression correlated with increased interferon levels and infection severity [21].
The expression of GPR183 in eosinophils, together with increased oxysterol production in inflamed airways, could be responsible for recruitment to the tissue [52]. Another specialized immune compartment is the gastrointestinal tract, where lymphoid tissue inducer (LTi)-like group 3 innate lymphoid cells (ILC3) have been found to be organized in a GPR183 dependent manner and this affects inflammatory processes in the bowel [53,54].
GPR183 is also expressed on astrocytes and microglia in the brain, where it may thus effect neuroinflammatory diseases [55,56,57,58]. In the context of neuropathic pain, GPR183-expressing astrocytes and microglia accumulate in the spinal cord and allodynic effects can be attenuated by the inhibition of this receptor in vivo [59].
Related to this, several studies have looked at the effects of the GPR183–oxysterol axis in multiple sclerosis. GPR183 dependent T-lymphocyte trafficking seems to be of significant relevance here. In particular, TH17 and memory lymphocytes were attracted by high 7α,25-OHC concentrations, which can be established by activated microglia in inflammatory lesions [60,61,62].
Unrelated to the immune system, GPR183 is also expressed on monocyte/osteoclasts precursors, and thus heightened 7α,25-OHC secretion by osteoblasts, and in an autocrine manner by osteoclast precursors themselves, provides signals for bone resorption [63]. This might also be of relevance in the control of bone mineral density during systemic inflammatory or high cholesterol settings.
2.1.2. The Chemistry and Production of Endogenous Oxysterols That Bind GPR183
7α,25-OHC has been determined to be the endogenous ligand with the highest affinity and strongest activity for GPR183, though other oxysterols like 7α,27-OHC or 7β,25 OHC also activate this receptor, albeit with lower potency and efficacy [64]. 7α,25-OHC is synthesized from cholesterol in two steps. The first additional hydroxyl group is added in position 25 facilitated by cholesterol-25 hydroxylase (CH25H); the second is catalysed by CYP7B1 [65]. Alternatively, 7α,25-OHC can be produced by a combination of CYP7A1 and CYP3A4 [66,67]. However, this is insufficient to launch normal T cell-dependent antibody responses [64].
The gradient in the lymph nodes stems mainly from stromal cells. Here, the cell-specific knockout of these key enzymes disrupted B-cell positioning. Several hematopoietic cell types, for example, follicular DCs, have also been shown to express Ch25H and Cyp7b1, but ligand expression in the lymph tissue was not affected by the lack of these enzymes in bone marrow-derived cells [41]. This becomes relevant in normal tissue immune responses, as the key enzymes can be differentially regulated. Innate immune cells, such as DCs and macrophages, increase Ch25H expression via multiple immune pathways [68]. Lipopolysaccharide (LPS) challenge of mouse macrophages, mimicking bacterial infection, resulted in heightened Ch25H expression and, correspondingly, increased the plasma concentration of 25-HC in human test subjects [69]. Human primary monocytes express both receptor and ligand producing enzymes, with a marked upregulation upon LPS stimulation [70]. The lungs are suggested to be a tissue where this holds central significance, as CH25H is especially highly expressed and LPS challenge in KO mice results in prolonged neutrophilia [71]. Epithelial cells in the airways also upregulate these oxysterol-producing enzymes in chronic obstructive pulmonary disease conditions. Here, the higher concentration was shown to attract B cells via GPR183 (and stimulate local lymphoid tissue formation) [72]. An older study also found a correlation between increased 25-HC secretion and decreased IgA class switching, with the same effect in CYP7B1 KO mice [28]. Under normal physiological conditions, 25-HC and 27-HC concentrations are increased in Cyp7b1 KO mice but, otherwise, the phenotype remains mostly normal. Enzyme function in bile acid synthesis seems to be substitutable [73].
CYP7B1 expression is regulated by nuclear factor kappa B (NF-κB) and the Janus kinase signalling pathway, tying it to inflammatory and metabolic stress contexts. In rheumatoid arthritis, for example, upregulation of tumour necrosis factor alpha (TNFα), interleukin 1 (IL-1) and IL-17 induces heightened CYP7B1 expression by fibroblastic cells [74]. During acute lung inflammation Cyp7b1 is also upregulated [75]. In insulin resistance states, on the other hand, CYP7B1 expression is low, while GPR183 and CH25H are upregulated, leading to accumulation of intermediate oxysterols in non-alcoholic fatty liver disease [76,77]. However, a direct correlation between disease progression and GPR183 signalling was not found. Breast cancer cells downregulate CYP7B1 expression by methylation, and promote CYP27A1 expression by specifically differentiating and recruiting M2 macrophages, leading to a feedback loop increasing 27-HC concentrations. This promotes tumour growth and metastasis [78].
Hydroxysterol dehydrogenase-3 B7 (HSD3B7) is an enzyme relevant in the degradation of 7α,25-OHC and a number of other oxysterols to form bile acids. HSD3B7 is expressed in secondary lymph organ stromal cells and fDC and is necessary to create a balanced gradient [41]. It has also been found in the intestines, especially in stromal cells surrounding the lymphoid structures [53]. Interestingly, the cue for fDC localization itself is given by 7α,27-OHC, which is produced by another enzyme, Cyp27a1, also expressed in stromal cells, but also degraded by HSD3B7 [50]. This might be another mechanism to achieve differential signalling.
Overall, these possible relation certainly shows there is room to explore unravelling the signalling pathways governing the immune system and, in particular, the antibody production capabilities.
2.1.3. GPR183 Receptor Structure
To date, there is no resolved receptor structure. However, several attempts at homology modelling have been made, accompanied by extensive mutagenesis studies. Developments in this structure guided approach, with the goal of advancement towards targeted drug discovery, were mainly made by two groups: those of Changlu Liu and of Mette Rosenkilde. Confirmation of a specific binding mode of either the endogenous ligands or existing small molecules remains elusive.
Mutagenesis efforts concentrated on residues in the orthosteric, major and minor binding pocket area, where several amino acids showed effects on signal transduction to Gαi or β-arrestin. Since GPR183 had been described as an orphan receptor until around 2010, several mutagenesis studies were performed, where either the inhibition of supposed constitutive activity occurred, or where binding of the first synthetic compound were analyzed [79,80]. By these means, an initial set of functionally important residues was identified. The strongest effects of mutants were seen in R87, increasing activity, and F257 of the CFxP-motif, decreasing it. Several partially conserved chemokine receptor (CXCR) motifs were also detected, in conformity with GPR183 now generally believed to be closely related to that receptor family. Both groups proposed binding poses on homology models based on the phylogenically closest known resolved structure of CXCR4 (PDB:3ODU) or a CXCR4 chimera, but those differ [81]. In the model from Zhang et al., the key interacting residues were proposed to be R87, N114 and E183 [81], of which N114 was later proven to belong to an allosteric metal ion site. However, the groups did agree on the relevance of four residues: R87, Y112, Y116, Y260, which were determined as the key binding residues in the model from Benned-Jensen et al. [82]. A third homology model based on CXCR4 exists, made to compare the orthosteric sites of the three class A GPCRs characterized to be oxysterol binding [83]. The one residue not appearing in their binding site prognosis is Y116, since it is rotated in a different direction in the template structure. The modelling of GPR183 on more recent crystal structures includes the PAR1 receptor, where possibilities for biased agonism were in focus. In this process, the receptor has been shown to contain the conserved allosteric sodium binding site found in several class A receptors, with special importance of the main residues D77, N114 and D304 [84].
Further analysis to confirm the specific binding residues is in the making though, to improve identification opportunities for pharmacologically relevant new drug candidates. These days, there are several endogenous oxysterols and small molecule modulators, but a definite site for neither is known. Recent discoveries include a number of allosteric sites in class A GPCRs formerly unconsidered [85,86]. Of special interest are binding sites outside of the helices, in the lipid bilayer, since oxysterols, with their highly lipophilic character, would have easy access here. This is also supported by cholesterol molecules, which favour specific sites in resolved structures or even enter receptors from here [87].
2.2. CXC Chemokine Receptor 2 (CXCR2)
2.2.1. Cellular Expression Pattern and Overall Biological Function
CXCR2 is a chemokine receptor. It is phylogenetically related to CXCR1, with which it shares high sequence similarity and has some overlapping ligands and mechanisms. CXCR2 is highly expressed by innate immune cells and endothelia. One of the main functions is neutrophil recruitment and activation, leading to the creation of a pro-inflammatory environment to promote pathogen clearance. In cancer, neutrophils recruited by this mechanism can also further promote tumour growth and metastasis by the induction of neoangiogenesis and inhibition of cytotoxic T cells [88,89]. Opposing effects have also been demonstrated. Angiogenic effects might also be attributed to CXCR2 expression on endothelial cells, which also leads to chemotaxis in these cells [90].
Several other diseases have been associated with CXCR2 signalling, mostly related to neutrophil activity. Among these are neurodegenerative diseases, such as multiple sclerosis, where increased ROS production leads to sustained neuronal damage [91] and neutrophil activation facilitates blood–brain barrier breakdown [92]. Several other inflammatory diseases are influenced by CXCR2 dysregulated neutrophils. Pancreatitis can be attenuated by the inhibition of CXCR2 signalling on neutrophils, where the main effects are attributed to decreased neutrophil chemotaxis-reducing activation of monocytes and, thus, cell death in the pancreatic tissue [93]. The same holds true in ulcerative colitis, where attenuated chemotaxis leads to a decrease in inflammatory cytokine expression [94,95].
CXCR2 is also expressed in hematopoietic stem cells, where signalling plays a role in homeostasis [96]. It also acts as a mobilization signal for neutrophil release from the bone marrow [97]. Furthermore, the fate of differentiating cells of the myeloid line is dependent on the receptor and a bias towards suppressor cells exists in ligand rich tumour settings [98].
2.2.2. The Chemistry and Production of Endogenous CXCR2 Ligands
CXCR2 has several endogenous ligands. The receptor is sensitive to all ELR+ CXC chemokines to varying degrees [99]. Different sets of these are expressed in different settings, but CXCR2 is recognized as their main receptor. CXCL1 and 2, for example, provide the cue for mobilization of neutrophils from the bone marrow [97] and are contributing factors in CNS autoimmune diseases due to increased immune cell infiltration upon TH17 induced chemokine upregulation [92]. CXCL8, on the other hand, plays a significant role in recruitment to sites of infection and vascular escape [100]. Here, it has been shown that concentration, steepness of the gradient, as well as ligand conformation and complexation, play differential roles in signalling outcomes.
Interestingly, in non-inflammatory, non-disease settings a key regulator of endothelial ELR+ CXCL expression seems to be low shear stress as seen in capillaries Shaik et al., [101]. This is regulated via p38, NF-κB signalling pathway, which relates well to increased expression in inflammatory settings.
Oxysterols are another wide group of endogenous ligands for CXCR2. A broad range of single hydroxylated cholesterol derivates proved to be able to induce migration in neutrophils [102,103]. The focus in this study was on 22-HC, due to its higher concentration in tumours. The group reported higher neutrophil recruitment of CXCR2-expressing cells, accompanied by the described pro-tumour effects. They did not, however, see synergistic effects between oxysterols and endogenous CXCLs. The effects seem to be independent from each other, with oxysterols sufficient to induce chemotaxis. A later study also elucidated the effects of 27-HC [89], the second highly abundant oxysterol in cancer. In comparison, they observed increased metastasis with 27-HC treatment. This effect was facilitated by γ/δ T cells, next to the intrinsically important neutrophils, through a vicious cycle between these populations.
2.2.3. CXCR2 Structure
Since antagonism of CXCR2 has been suggested as a therapeutic tool in a wide spectrum of issues for several decades, a lot of effort was put into finding, refining and proving the concepts behind a wide panel of compound classes. Methods range from early chimeric proteins and site-directed mutagenesis to partial structure resolution. Finally, in 2019, a full receptor structure, including an endogenous ligand and G protein and a version with a small molecule, were resolved (PDB:6LFL/M/O) [104].
Mutagenesis studies revealed several distinct areas that are responsible for the binding of different ligands. Endogenous CXCLs are thought to bind on the extracellular side, with an emphasis on N-terminal involvement for CXCL1 and probably other ELR+ CXC chemokines not inducing signalling on CXCR1, and increased importance of ECL2 for binding of CXCL8 [105].
Known CXCR2 antagonists can be divided into several classes, which seem to, at least in part, bind distinct sites on the receptor. Two main sites have been described, one intracellular, possibly interfering with G protein binding, and one inside the transmembrane helical bundle, probably interfering with activation-related conformational changes [106,107,108]. This intrahelical pocket is located inside of TM 3, 5 and 6, with an entry point from the TM5/6 interface. Interestingly, the environment of the entry channel seems conductive for lipid binding, as seen in the CXCR4 crystal structure, which one might speculate to be a possible site for interaction with oxysterols too.
The intracellular C-terminus of CXCR2 carries a PDZ binding motif, which is relevant for signal transduction via Na+/H+ exchanger regulatory factor (NHERF1), which is especially relevant in neutrophil chemotaxis. Interference with this interaction has also been proven to modulate disease mechanisms [109,110].
The full length cryo-EM receptor structures resolved by Liu et al., 2020 [102] further elucidated the binding of CXCL8, Gi and an antagonist at the intracellular site, overlapping with the G-protein site. CXCL8 turned out to have a fairly shallow binding compared to other chemokine receptor/ligand complexes. In comparison with CXCR1, two major interaction sites are likely key to permissive ELR+ CXCL recognition, the PP-PC motif region on the N terminus and CSR2 that directly interacts with the ELR motif. Another interesting finding was a resolved cholesterol located on the TM 2, 3, 4 interface at the height of the inner membrane leaflet in the active state structure. This might indicate a different oxysterol binding site, stabilizing the receptor in the active conformation. There have been studies looking at possible effects of cholesterol in general chemokine receptor function [111], and some of the mechanisms might be correlated or overlapping.
2.3. Smoothened (SMO)
2.3.1. Cellular Expression Pattern and Overall Biological Function
Smoothened (SMO) is a class F GPCR best known for its actions in development and cancer. It is an oncoprotein and part of the hedgehog (Hh) signalling pathway, where it is responsible for transmembrane signal transduction. This happens when Hh binding to its receptor Patched relieves inhibition of SMO. Several direct ligands and modulators for SMO have been known for a while [112,113]. These can activate or inhibit different steps in the activation cascade, before relocalisation to the basal cilia, or at the stage of final activation [114]. Signalling results in Gli family transcription factor cleavage, where pathway activation leads to activator fragments, while downregulation increases the repressor fragment availability. There are three different mammalian hedgehog proteins, and three Gli factors.
Hh signalling also acts as a regulator in the immune system, Figure 3, and has direct regulatory influences on the adaptive side. Lymphocyte development and fate can be dependent on the Hh pathway and thus on SMO, Figure 3. Differential Hh signalling has been shown to be important in various steps of T cell differentiation, as well as effector function. In the development sonic hedgehog (SHh)-dependent Gli activation is important for transition of DN1 to DN2 and survival of DN4 cells. Intermediate concentrations accelerate transition to DP cells, while very high or very low concentrations lead to arrest [115,116]. In contrast, DN3 survival and later proliferation of DP to SP effector cells increased upon downregulation of Hh pathway activation [117,118].
Figure 3.
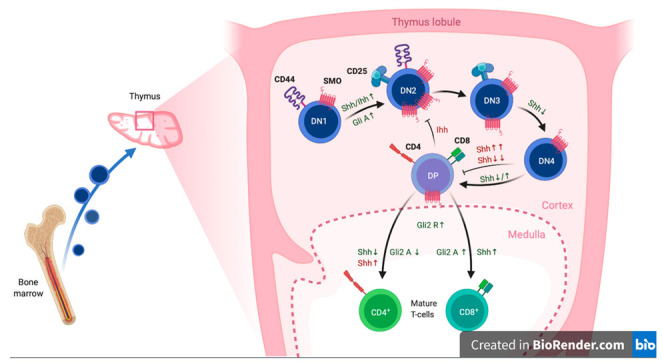
Summary of the differential expression and signalling effects of SMO, including its upstream signals (Hh) and its downstream transcription factors (Gli) on T-cell development in the thymus. Inhibitory signals and effects are highlighted in red, differentiation stimulating effects are marked green. This figure was adapted from “T-Cell Development in Thymus 2”, by BioRender.com (2021), retrieved from https://app.biorender.com/biorender-templates.
Shh levels also influence cell fate. Homeostasis and maturation of γ/δ T cells in adult mice are reduced upon inhibition of Hh signalling [119,120]. Low SHh increases the rate of differentiation to single positive cells, while skewing balance between CD4+ and CD8+ towards CD4+ T cells [120]. CD4+ cells are themselves skewed towards Th2 differentiation upon higher signalling via Gli2A [121]. Indian hedgehog (IHh), another Hh ligand, is needed for efficient CD8+ cytotoxicity [122]. Natural killer T (NKT) cells, another T-cell subpopulation, also possess an increased cytotoxic profile upon SHh stimulation, which they can produce themselves, thus opening the possibility of entering an autocrine loop [123].
Other cells of the adaptive immune system are also subject to Hh signalling. Early B-cell development is dependent on SHh, such that lower levels stimulate differentiation, while higher levels reduce B cell commitment and maturation. This is regulated via Gli3 repression of SHh expression in the stromal cells [124,125]. A similar mechanism, as has been shown for thymic epithelial cells [120] and used in skin inflammation experiments to check on the influence of Hh signalling on peripheral immune cells [126].
2.3.2. Chemistry and Production of Endogenous SMO Ligands
There is a certain degree of speculation about the endogenous ligand of SMO. Neither the mechanism of how Patched inhibits SMO activity, nor which lipid or small molecule might be endogenously responsible for signal transduction, has been elucidated so far. Sterols have been proven to modulate SMO activation though, and their widespread natural occurrence qualifies them as possible endogenous ligands or synergistic allosteric modulators.
In vitro studies found that oxysterol 20(S)-HC is the most potent agonist, though 22(S)-HC, 7-keto-25-HC and 7-keto-27-HC and others activate the receptor [127]. All of these have the normal cholesterol hydroxyl group in position 3 in common. Several of these are able to synergize with each other, or with some of the other known small molecule ligands. Unmodified cholesterol also binds SMO, and can activate the receptor, as well as synergizing with Hh signals or other ligands. An important relation between SMO activation and cholesterol biosynthesis was established long ago [128], but the question of whether cholesterol itself is the endogenous ligand has been raised multiple times over the years and remains unanswered. Fairly recently, a study showed that covalent cholesterol modification takes place on residue D95 in human SMO (in the extracellular CRD binding site) [129]. It might be that either cholesterol esterification, binding or oxysterol availability are regulated by Patched, or that they provide a second signal, maybe ensuring capability of the cell to support the induced changes in transcription. One way in which oxysterols could be enriched for SMO activation has been described by Raleigh et al. [130]. They found that a number of oxysterols, which can synergistically activate SMO via two sites, are enriched in the cilia membrane, where final SMO activation usually takes place.
2.3.3. SMO Receptor Structure
There are several receptor structures for SMO, published dating back to 2013, using methods ranging from NMR to conventional X-ray diffraction, to cryo-electron microscopy. A number of ligands have been resolved, bound to fragments, the full-length receptor, or receptor G protein complexes. Especially relevant for this review are the structures containing cholesterols and derivates.
A cysteine-rich domain (CRD) crystal structure from [131] (PDB:5KZV) shows a binding site for 20(S)-OHC. The authors highlighted the binding mode and allosteric transition induced by binding and discovered key residues that are important for both oxysterol and cholesterol binding, which both induce conformational changes and are able to activate the receptor for/with SHh stimulation. The group hypothesized that cholesterol is the endogenous ligand, due to its physiologically relevant concentration. Further studies found that cholesterol seems to be covalently bound in that site at some point. Residue D95, which had been mutated by the aforementioned group and abolished binding in their assays, showed up with a correspondingly increased molecular weight in MS/MS. However, the mechanism of this modification and its explicit regulatory function remain unclear [129].
Comparing different structures resolved in recent years, some major differences can be seen in the tilt of the CRD region. This might be due to different activation states [132,133]. Deshpande et al. [134] solved a full receptor structure, stabilized in the active state by nanobody binding, with cholesterol bound in a deep pocket inside the 7TM region (PDB:6O3C). This binding produces the activating changes for GPCRs, independent of Hh, especially the marked shift in TM6. The binding pocket opens up with the receptor in a more active-like state; for example, with agonists bound higher up in the receptor, or by stimulation from the CRD site. The known antagonists bind at a site that overlaps partly with this deep cholesterol site, thus inhibiting activation this way as well as stabilizing the inactive state.
A year later, Qi et al. [135] solved more structures of WT and SMO-Gi complexes with several oxysterols and an agonist, SAG, bound (PDB:6XBM). In this way they discovered more, interconnected binding sites, which seem to allow for transfer through the receptor, from the deep 7TM pocket outwards to the CRD.
2.4. Gamma-Amino Butyric Acid (GABA) Type B Receptors
GABA is the main inhibitory neurotransmitter in the vertebrate central nervous system. It exerts biological effects by interacting with ionotropic GABAARs (see Section 3.1.3), which contain an intrinsic chloride channel, and with metabotropic GABABRs, which are GPCRs. There is limited evidence supporting a modulatory role for oxysterols in GABABR signalling. In brain slices from the rat lateral septum, the superfusion of 25-HC selectively reduced GABABR- but not GABAAR-dependent inhibitory post-synaptic potentials [136]. Microglia are known to possess both GABABR subtypes. The activation of these receptors inhibits microglial IL-6 and IL-12p40 secretion in response to LPS [137]. The GABABR-selective antagonist, baclofen, is reported to inhibit DC cell activation and their priming of Th17+ T cells [138].
3. Ion Channels
Gating of ion channels permits rapid changes in intracellular ion concentrations in response to diverse stimuli, leading to alterations in membrane potential, the second messenger Ca2+, cell volume, cell-death, gene expression, secretion, endocytosis, or motility. The gating of a limited range of ion channels is known to be modulated by oxysterols, often in a congener- and channel subtype-selective channel fashion. In addition to cell-surface channel proteins, IP3R Ca2+ release channels are located predominantly on the ER and are also modulated by oxysterols. This includes the stimulation of the proteolytic degradation of IP3Rs [139], and the assembly of IP3R signalling complexes by [16], and direct interactions with, ORP4L [17]. The cell-surface ion channels that are modulated by oxysterols can be divided into three functional categories: those gated by ligands; those gated by changes in membrane potential (voltage-gated); and those gated by multiple stimuli (multi-modal gating). The presence and roles of these channels in the immune system are largely unexplored. Their levels of transcription in immune cells are summarized in Figure 4. These channels have considerable potential as targets for the development of new therapies to combat immune disorders, including autoimmune diseases [4,5].
Figure 4.
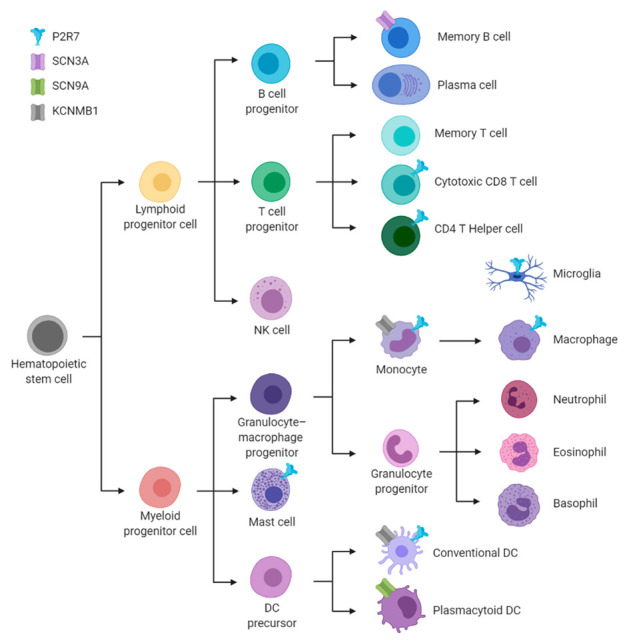
Overview of the key immune cell types from the hematopoietic lineage with an abstraction of their cellular expression of the ion channels discussed in this article. These channels are represented by icons of different colours: blue Figure 2. X7R, purple for SCN3A, green for SCN9A and grey for KCNMB1. This overview is derived from consensus of four independent transcriptomic data sets from human immune cells [31,32,33], as summarized in Supplemental Figure S1. Figure 4 was created in BioRender (BioRender.com).
3.1. Ligand-Gated Ion Channels
These are gated by the binding of a specific ligand to a site within the ion channel complex. Only a limited number of this type of channel have been reported to interact with oxysterols: the P2X7 purinoreceptor (P2X7R), the N-methyl-d-aspartate receptor (NMDAR) subtype 2B (encoded by GRIN2B) and the GABAARs.
3.1.1. The P2X7 Purinoreceptor (P2X7R)
P2X7R is a member of the P2X family of purinoreceptors that bind ATP, opening a non-selective cation channel that is highly permeable to Ca2+. P2X7R monomers contain two transmembrane helices and a cytoplasmic C-terminus that is extended relative to other members of the P2X family. P2X7R is distinct from other members of its family in that it is formed only by homotrimeric complexes, as opposed to heterotrimeric ones, and in that it binds ATP with low affinity. When stimulated by low concentrations of ATP, the P2XR channel attains a conformational state that is permeable to inorganic cations. When stimulated with higher concentrations of ATP for sustained durations, the channel pore dilates, or recruits additional components, such as pannexin-1, so that it forms a pathway that is permeable to much larger organic cations [140]. This dilated pore conformation is thought to underlie the subtype-specific cytotoxicity resulting from P2X7 activation. The molecular structures of rat P2X7R in both its apo, closed state (PDB:6U9V) and ATP-bound, open state (PDB:6U9W) have been determined using cryo-electron microscopy, but these provide no evidence supporting a dilated pore conformation. However, these structures indicate a role for cysteine palmitoylation in maintaining the channel in a non-desensitizing state [141].
P2X7R channel gating is modulated by several types of sterol. The depletion of cholesterol from the plasma membranes of cells transfected with P2X7R increases permeation of organic cations, whereas cholesterol loading inhibits this mechanism. An N-terminal domain and multiple cholesterol recognition amino acid consensus motifs near the C-terminus are key determinants of this cholesterol sensitivity [142]. Vitamin D (1,25-dihydroxycholesterol) rapidly increases in Ca2+ in peripheral blood mononuclear cells via a mechanism dependent on P2X7R [143], but it is not known if this effect is mediated by binding to the same sites as cholesterol. There is also evidence that oxysterols modify the gating of P2X7R channels. In a model of human skin, 25-HC promotes both caspase-dependent apoptosis and pyroptosis via a P2X7R-dependent mechanism [144]. In human retinal pigment epithelial cells, both 7KC and 25-HC activate P2X7R, with the latter oxysterol requiring pannexin-1 for this effect [145].
P2X7R is present in many cells of the immune system, with the highest levels in monocytes, followed by NK cells and CD4+ memory Treg cells; Figure 4 and Supplemental Figure S1. The roles of oxysterols in modifying P2X7R responses in immune cells are yet to be elucidated. This receptor plays a key role, as in the recognition of extracellular ATP, an archetypal damage associated molecular pattern (DAMP) molecule that is released from injured cells [146]. In monocytes, the acute stimulation of P2X7R enhances the secretion of IL-1β [147], whereas sustained activation promotes apoptotic cell death. Extracellular ATP promotes the activation of the NLRP3-inflammosome in monocytes and this process is impaired during sepsis [148].
Substantial experimental evidence supports the roles of P2X7R in T-lymphocyte biology [149]. P2X7R antagonists inhibit the TCR-dependent activation of mouse T cells, as assessed by their effect on IL-2 production [150]. Through P2X7R, ATP inhibits both the differentiation of naïve CD4+ cells into regulatory CD4+ CD25+ T cells (Treg) and the immunosuppressive capabilities of this subtype [151]. The activation of P2X7R drives the decision for differentiation to an αβ rather than a γδ TCR phenotype [152]. Human CD4+CD45RO+ memory T cells indirectly inhibit NLRP3 inflammasome activation in ATP-stimulated monocytes, via downregulation of P2X7R [153]. In response to Plasmodium chabaudi malaria infection, P2X7R stimulates Th-cell differentiation and decreases the Tfh subset [154]. Extracellular ATP promotes metabolic fitness in long-lived CD8+ memory T cells, through a mechanism involving P2X7R and AMP-activated protein kinase [155]. P2X7R stimulates non-classical secretion in numerous cell types, including T lymphocytes [156]. It also impairs the MHC-I-dependent presentation of oligopeptides by antigen presenting cells, thereby suppressing the activation of CD8+ T cells [157]. ATP drives apoptotic death in thymocytes via the activation of P2X7R. This receptor also underlies inflammation in a model of colitis, by promoting apoptosis in Treg cells [158].
3.1.2. Ionotropic Glutamate Receptors
Glutamate is the most abundant excitatory neurotransmitter in the mammalian central nervous system. Receptors for glutamate are divided between those that are ionotropic, containing an intrinsic ion channel, and metabotropic GPCRs. Ionotropic glutamate receptors are named after their definitive pharmacological ligands: N-methyl-d-aspartate (NMDA), γ-amino butyric acid (GABA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA). Endogenous agonists of NMDAR channel complexes are glutamate, d-serine and/or glycine, depending on their subunit composition. They are heterotetrameric channel complexes, usually consisting of two obligatory GluN1 subunits (encoded by GRIN1) and two GluN2A, GluN2B, GluN2C, or GluN2D subunits (GRIN2A, GRIN2B, GRIN2C, or GRIN2D). Two alternative subunits, GluN3A and GluN3B (GRIN3A, GRIN3B), can combine with GluN1 to form a complex that has reduced current, which is gated by glycine and not by glutamate [159,160,161]. The opening of NMDAR channels usually results in the influx of Ca2+ and Na+, and an efflux of K+. In vivo, the permeation pathway can be blocked by Mg2+, with this inhibition being relieved by depolarization. NMDARs are predominantly present in the central nervous system, where they act as post-synaptic glutamate receptors. They have also been identified in multiple peripheral, non-neuronal cell-types [162,163]. The presence of NMDARs in cell types that circulate in the blood is potentially problematic, since the concentrations of amino acid agonists in the extracellular fluid far exceed those required for the maximal opening of these channels. This situation would result in the excessive influx of Ca2+ and consequent cytotoxicity. This issue may be resolved by peripheral cells producing NMDARs that are less sensitive to such stimuli, either through alternate splicing of the mRNA(s) encoding their subunits, distinctive assembles of subunits, or by the presence of additional inhibitory modulators that do not operate in the nervous system [162].
NMDARS are modulated by oxysterols. The most abundant cholesterol metabolite in the brain is 24(S)-HC, which is sometimes referred to as cerebrosterol. This oxysterol potentiates NMDAR-dependent excitatory post-synaptic currents (EPSCs) in rat hippocampal neurons at submicrolar concentrations; an effect that was not caused by unmodified cholesterol, or by other oxysterols, at concentrations up to 10 µM. 24(S)-HC did not modify the gating of other ionotropic glutamate receptors, the GABA receptors and AMPA receptors [164]. SGE-201, a synthetic analogue of 24(S)-HC, influenced the electrophysiological properties of NMDARs present either in single neurons, heterologously expressed in Xenopus oocytes, or within hippocampal slices [165]. 24(S)-HC is produced in the central nervous system by the action of CYP46A1 on cholesterol. In hippocampal slices from CYP46A1−/− knockout mice, both the concentration of 24(S)-HC and the basal activity of NMDARs are greatly reduced relative to those from wildtype animals [166]. Use of chimeric receptors to map the binding of positive allosteric modulators to NMDARs indicates that 24(S)-HC interacts with the transmembrane domains of these proteins, at a binding site distinct from those of pregnenolone sulfate, or of docosahexaenoic acid [167]. In mouse hippocampal slices, the GluN2B-selective antagonist Ro25-6981 blocked the 24(S)-HC enhancement of NMDAR-dependent currents, whereas the GluN2A antagonist PEAQX did not [168]. Although 25-HC weakly potentiated NMDAR currents when administered alone, it non-competitively inhibited the potentiation elicited by 24(S)-HC. This suggests the presence of two distinct binding sites for oxysterol modulators within NMDAR complexes [169]. In an oxygen-glucose deprivation model of ischemia, 24(S)-HC exacerbates neuronal death, whereas 25-HC opposes it and can reverse the effects of 24(S)-HC [169]. Another oxysterol congener, 27-HC, reduces NMDAR protein levels in mouse brain, possibly through the downregulation of the activity-regulated cytoskeleton-associated protein (Arc), required for trafficking of these channels to the cell-surface [170].
Negative modulation of NMDARs by some oxysterol congeners might provide an explanation for an absence of cytotoxicity in immune cells, in the face of high concentrations of excitatory amino acids encountered in the blood. NMDARs are present in some immune cells, as reviewed by Boldyrev in 2005 [171]. Using fluorescence-activated cytometry, GluN1 was detected in human peripheral lymphocytes, with the activation of NMDARs in these cells being associated with Ca2+ influx, ROS production and cell death. In NK cells derived from this population, NMDA had no effect on IFN-γ production, but suppressed that stimulated by IL-2 [172]. In mouse splenic B-lymphocytes, non-competitive NMDAR antagonists reduced proliferation, chemotaxis and immunoglobulin secretion, but enhanced IL-10 production. However, these antagonists block K+ currents through voltage-gated Kv1.3 and KCa3.1 present in these and other immune cells (see Section 3.2.1.1 and Section 3.3.1 ) [173,174]. These observations highlight the need for caution when interpreting the effects of small molecule drugs on immune cells.
3.1.3. Ionotropic GABAARs
As described in Section 2.4, GABA is the predominant central inhibitory neurotransmitter, which exerts its effects via both metabotropic (GABABR) and ionotropic (GABAAR) receptors. GABAAR complexes are pentameric in structure, with 19 distinct genes encoding subunits, and possess an intrinsic chloride channel [175]. Although no evidence for the effects of oxysterols on the gating of GABAARs has been reported to date, these channels are modulated by neurosteroids, such as progesterone, allopregnanolone, and their synthetic analogues [176]. GABAAR regulates multiple aspects of immune function, including the migration of DC and the activation of T lymphocytes [177].
3.2. Voltage-Gated Ion Channels
Voltage-gated ion channels respond to changes in membrane potential in the order of a few tens of millivolts. This allows for the flow of ions, altering membrane potential, in addition to the transfer of electrolytes and, in some cases, the influx of the second messenger Ca2+ [15]. This class of channel are often extremely selective in terms of the ions that they conduct, relative to other channel families. Voltage-gated ion channels regulate diverse processes, including secretion, cell motility and the generation of action potentials. Two types of voltage-gated ion channels are known to be modulated by oxysterols: voltage-gated potassium channels and voltage-gated sodium channels.
3.2.1. Voltage-Gated Potassium Channels
There is considerable evidence to support roles of voltage-gated K+ channels in immune cell function. These channels are present in most cell types and regulate multiple aspects of physiology, including membrane potential, secretion, cell volume and death. They are formed from tetrameric complexes that interact with a range of accessory proteins. Voltage-gated K+ channels are diverse in terms of their structures, electrophysiology and pharmacology. To date, of the voltage-gated K+ channel family, only Kv3.1 (belonging to the Shaw, or C, subfamily and encoded by KCNC1) and KCa1.1 (Slo-1, or large conductance calcium-activated potassium channel, subfamily M, alpha member 1, encoded by KCNMA1) have been reported to be modulated by oxysterols. Since it is gated by a combination of membrane depolarization and increased cytoplasmic Ca2+, KCa1.1 will be discussed in greater detail in Section 3.3.1, on channels opened by multiple stimuli.
3.2.1.1. Kv3.1
The Kv3 family of voltage-gated K+ channels was originally identified as the Shaw family in the fruit-fly Drosophila melanogaster and consists of three distinct genes encoding Kv3.1, Kv3.2 and Kv3.3. Electrophysiologically, this family differs from other voltage-gated K+ channels in terms of activation at more positive membrane potentials and rapid inactivation. In neurons, these properties mean that Kv3 channels contribute to rapid, repetitive firing of action potentials and, when mutated, to epilepsy [178]. Kv3.1 is produced as two alternately spliced variants, Kv3.1a and Kv3.1b, differing at the C-terminus. Of these, Kv3.1b is known to be modulated by oxysterols. In both 158N oligodendrocyte and BV-2 microglial cell lines, incubation with either 24(S)-HC or 7KC for 48 h decreased Kv1.3b protein abundance and function [179]. Kv1.3 is responsible for the “I-K+ current” and is transcribed at very low levels in cytotoxic T cells, but at high levels in a CD4-CD8-Thy1+ T lymphocytes from mice with autoimmune disease [180].
3.2.2. Voltage-Gated Sodium Channels
Voltage-gated Na+ channels have a canonical role in underlying action potentials in excitable tissues, but regulate cell-volume, motility, secretion and death in other cell types. They are comprised of a channel-forming, voltage-sensing α subunit (Nav1.1–1.5 encoded by SCN1A–SCN5A, Nav1.6 encoded by SCN8A, Nav1.7–1.8 encoded by SCN9A–SCN11A, and Nax encoded by SCN7A) in combination with accessory proteins, such as the β-sub-units (Navβ1–4 encoded by SCN1B–SCN4B), which influence channel gating or trafficking [181]. Canonical antagonists of these channels include the puffer-fish toxin, tetrodotoxin (TTX), and the small molecule drug, amiloride. Since voltage-gated Na+ channels are present in lymphocyte- and monocyte-derived immune cells, there is potential for small molecule modulators of these channels to be utilized as immunotherapeutic drugs. This is offset by potentially severe side-effects of such drugs on the nervous system [182].
There is strong evidence for the modulation of voltage-gated Na+ channels by oxysterols [183]. In dorsal root ganglion neurons, the TTX-resistant Nav1.8 protein is distributed in clusters, thought to be lipid rafts. This localization was disrupted by either the cholesterol-chelator methyl-β-cyclodextrin, or by the oxysterol 7KC, and was associated with a loss of neuronal excitability [184]. At micromolar concentrations, the oxysterol, cholestane-3β,5α,6β-triol (triol) reduces Na+ current density in rat hippocampal pyramidal neurons and ablated seizures in a kainate-induced mouse model of epilepsy [185]. Molecular modelling of voltage-gated Na+ channels from the bacteria Arcobacter butzleri and Magnetococcus marinus indicates that triol binds to the indole ring of tryptophan 122 within the channel pore [186].
As assessed by flow cytometry, the voltage-gated Na+ channel inhibitors batrachotoxin or veratridine depolarized a subset of human NK cells [187]. In human HLA-DR-restricted CD4+ αβ T cells, stimulation with a non-self-antigenic peptide elicited a Na+ current that could be blocked with amiloride, though not with TTX [188]. In contrast, voltage-gated Na+ currents detected in human peripheral blood mononuclear cells and in acute lymphocytic leukemia cell lines were largely inhibited by TTX and probably involved Nav1.3, Nav1.6, or Nav1.7. TTX also inhibited the invasiveness of Jurkat and MOLT-4 cell lines [189]. The expression levels of Nav1.5 underlie differences in TCR-mediated responses to Listeria antigens in two CD4+ T-cell lines [190]. Immature dendritic cells (DC) possess voltage-gated Na+ channels, mediated predominantly by the Nav1.7 subtype, which is downregulated upon differentiation [191].
Voltage-gated Na+ channels, mainly those containing Nav1.1, Nav1.5 or Nav1.6, also regulate the physiology of monocytes, macrophages and microglia [192]. Nav1.5, a subtype originally identified in the cell-surface membrane of cardiomyocytes, is located in the late endosomes of human monocytes and macrophages, where it permits Na+ efflux from these organelles and decreases their pH [193]. In macrophages, these endosomal Nav1.5 channels also regulate phagocytosis of mycobacteria [194] and initiation of innate immune responses triggered by double-stranded RNA [195]. In macrophages, alternately-spliced variants of SCN5A and SCN10A promote the expression and protein abundance of serine/threonine-protein phosphatase 1 regulatory subunit 10 (PPP1R10), a DNA repair enzyme postulated to limit damage to host cells during inflammatory responses [196]. Knockdown of voltage-gated Na+ channels using RNA interference reduced proliferation, migration and phagocytosis by the RAW264.7 macrophage cell line. This also reduced macrophage proliferation in atherosclerotic plaques [197].
3.3. Ion Channels Gated by Multiple Stimuli
Certain families of ion channels are gated by multiple stimuli. Ion channels known to be modulated by oxysterols are Ca2+ and voltage-dependent K+ channels and transient receptor potential, canonical, subtype 1 (TRPC1) cation channels.
3.3.1. Ca2+ and Voltage-Dependent K+ Channels
The opening of a subset of voltage-gated K+ channels is stimulated by increased cytoplasmic Ca2+ concentrations. A well-characterized member of this family is called KCa1.1, or Slo-1, the large-conductance K+ channel BK, or MaxiK and is encoded by the calcium-activated K+ channel subunit, subfamily M, alpha-1 (KCNMA1) gene. BK channels are formed from tetrameric assemblies of KCa1.1 protein and their electrophysiology, trafficking and interactions with ligands are modulated by associations with accessory proteins, most notably β and γ subunits [198]. A key component of KCa1.1 gating is a switch from positively charged amino acids (the “RKK ring”) binding to negatively charged glutamate residues in the protein when in a closed state, to binding oxygen atoms of membrane lipids when in an open state. This gating mechanism helps explain the modulation of BK channel gating by lipophilic modulators [199]. BK gating can be modulated by cholesterol and by bile acids that interact with the channel complex at distinct sites, including at the β1 subunits [200].
Oxysterols modulate BK channel gating. At micromolar concentrations, the side-chain oxysterols 20(S)-HC, 22(R)-HC, 24(S)-HC, 25-HC and 27-HC all inhibited K+ currents through KCa1.1, whereas the ring-modified oxysterols, 7-KC or 7α-HC did not [201]. The inhibition of BK by 24(R)-HC is distinct from that exerted by its enantiomer 24(S)-HC in terms of activation kinetics and voltage dependency. The auxiliary subunits of BK channels can also be modulated by oxysterols. Within atherosclerotic lesions, 7KC downregulates KCNB1 (the β1 subunit of BK) [202].
BK channels play important roles in the innate immune system. Monocyte adhesion to endothelial cells elicited by lysophosphatidylcholine is dependent on these K+ channels [203]. In macrophages, the blockade of BK inhibits the IκB- and NF-κB-dependent inflammatory pathways stimulated by LPS [204]. BK is not exclusively located in the cell-surface membrane. In macrophages, lysosomal BK channels promote large-particle phagocytosis, by facilitating the release of Ca2+ from endolysosomal stores [205]. In the RAW264.7 macrophage cell line, nuclear BK regulates the phosphorylation of the transcription factor cyclic AMP response element binding protein (CREBP) [206].
3.3.2. Transient Receptor Potential Channels
Transient receptor potential channels (TRPs) were originally identified in Drosophila melanogaster photoreceptors. In mammals, homologues of this trp channel form a family of over thirty genes. They are all homo- or hetero-tetrameric in structure, conducting monovalent cations and in most cases, Ca2+. In humans, there are six functional members of the canonical TRP family, TRPC1-TRPC7, with TRPC2 considered to be a pseudogene. Similar to other members of the TRP family, TRPCs are gated by multiple stimuli, often acting in concert. A canonical stimulus for TRP gating is activation of PLC, but neither of the products of this enzyme (IP3 and DAG) gate these channels. Depletion of ER Ca2+ stores via IP3R channels activates TRP channels, possibly by interaction with another class of store-operated Ca2+-entry channels, Orai. TRPC1 is also reported to be gated by mechanical stimuli and, when in heterotetrameric complexes with TRPC5, by sphingosine-1-phosphate [207].
There is limited evidence of roles for oxysterols in the regulation of TRPC1 gating. In the THP-1 monocytic cell line 7-KC stimulates translocation of TRPC1 to lipid rafts, with subsequent Ca2+-dependent inhibition of the Bcl-2 antagonist of cell death [208]. Similarly, in vascular smooth muscle cells, oxidized low-density lipoprotein (oxLDL) causes cell-death that can be inhibited by TRPC1 knockdown and which is associated with Ca2+ influx. Treatment with oxLDL causes an increase in 7KC within the caveolae of these cells, along with the translocation of TRPC1 to these membrane invaginations [209].
TRPC1 has been detected in diverse immune cells. For example, using reverse transcription PCR, in human peripheral blood leukocytes, CD3+ T cells and the Jurkat T lymphoma were found [210]. Knockout of TRPC1 in neutrophils impairs their migration and chemotaxis [211]. M1 macrophage polarization is dependent on TRPC1-mediated Ca2+ entry [212]. In addition, the inhibition of TRPC1-mediated Ca2+ entry promotes ER stress in the RAW264.7 macrophage cell line, initiating inflammatory responses [213]. In primary microglia, soluble factors from the helminth Mesocestoides corti inhibit Ca2+ influx via TRPC1, suppressing immune responses; oxysterols are candidates for these soluble molecules [214].
4. Conclusions and Perspectives
Cells of the immune system contain multiple GPCRs and ion channels that are modulated by oxysterols. However, in many cases, it is unclear if these signalling proteins are influenced by cholesterol oxidation products in vivo under physiological or pathological settings. Similarly, many of the consequences of oxysterol modulation of GPCRs and ion channels within the immune system have not been investigated. Furthermore, the molecular mechanisms by which oxysterols alter the function of many of these receptors awaits full characterization. Consequently, there is considerable scope for exploration of cell-surface receptors for oxysterols in the immune system, both for scientific discovery and for the development of new immunotherapeutic regimes.
Acknowledgments
J.M. thanks members of the European Network for Oxysterols Research for useful discussions. All authors thank the reviewers of this article, for their contributions to improving the accepted version.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/cells10082078/s1, Figure S1: Expression of receptor for oxysterols during hematopoietic differentiation and on immune cells, Figure S2: A review of the scientific literature.
Author Contributions
This review was conceptualized by the corresponding authors, M.R. and J.M. The original draft of this review was prepared by L.R., M.R. and J.M., with all authors contributing to the refinement, revision and final preparation of this work. C.-C.H. and J.H. investigated, curated, analysed and visualised transcriptomic data. All authors contributed to literature review, critical assessment published work and its synthesis. L.R., C.-C.H. and J.M. generated the figures. J.M. contributed article publication charges. All authors have read and agreed to the published version of the manuscript.
Funding
C.-C.H. and J.H. are supported by the German Research Foundation (FOR 2149). For this project, J.M. was supported by the Department of Physiology, University College Cork, funded by the Higher Education Authority of Ireland.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Luu W., Sharpe L.J., Capell-Hattam I., Gelissen I.C., Brown A.J. Oxysterols: Old Tale, New Twists. Annu. Rev. Pharm. Toxicol. 2016;56:447–467. doi: 10.1146/annurev-pharmtox-010715-103233. [DOI] [PubMed] [Google Scholar]
- 2.Kloudova A., Guengerich F.P., Soucek P. The Role of Oxysterols in Human Cancer. Trends Endocrinol. Metab. 2017;28:485–496. doi: 10.1016/j.tem.2017.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zmysłowski A., Szterk A. Current knowledge on the mechanism of atherosclerosis and pro-atherosclerotic properties of oxysterols. Lipids Health Dis. 2017;16:188. doi: 10.1186/s12944-017-0579-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Choi C., Finlay D.K. Diverse Immunoregulatory Roles of Oxysterols—The Oxidized Cholesterol Metabolites. Metabolites. 2020;10:384. doi: 10.3390/metabo10100384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Duc D., Vigne S., Pot C. Oxysterols in Autoimmunity. Int. J. Mol. Sci. 2019;20:4522. doi: 10.3390/ijms20184522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Spann N.J., Glass C.K. Sterols and oxysterols in immune cell function. Nat. Immunol. 2013;14:893–900. doi: 10.1038/ni.2681. [DOI] [PubMed] [Google Scholar]
- 7.Massey J.B. Membrane and protein interactions of oxysterols. Curr. Opin. Lipidol. 2006;17:296–301. doi: 10.1097/01.mol.0000226123.17629.ab. [DOI] [PubMed] [Google Scholar]
- 8.Zang R., Case J.B., Yutuc E., Ma X., Shen S., Castro M.F.G., Liu Z., Zeng Q., Zhao H., Son J., et al. Cholesterol 25-hydroxylase suppresses SARS-CoV-2 replication by blocking membrane fusion. Proc. Natl. Acad. Sci. USA. 2020;117:32105–32113. doi: 10.1073/pnas.2012197117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Domingues M., Gomes B., Hollmann A., Santos N. 25-Hydroxycholesterol Effect on Membrane Structure and Mechanical Properties. Int. J. Mol. Sci. 2021;22:2574. doi: 10.3390/ijms22052574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ghzaiel I., Sassi K., Zarrouk A., Nury T., Ksila M., Leoni V., Bouhaouala-Zahar B., Hammami S., Hammami M., Mackrill J.J., et al. 7-Ketocholesterol: Effects on viral infections and hypothetical contribution in COVID-19. J. Steroid Biochem. Mol. Biol. 2021;212:105939. doi: 10.1016/j.jsbmb.2021.105939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Abrams M.E., Johnson K., Perelman S.S., Zhang L.-S., Endapally S., Mar K.B., Thompson B.M., McDonald J.G., Schoggins J.W., Radhakrishnan A., et al. Oxysterols provide innate immunity to bacterial infection by mobilizing cell surface accessible cholesterol. Nat. Microbiol. 2020;5:929–942. doi: 10.1038/s41564-020-0701-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Antonny B., Bigay J., Mesmin B. The Oxysterol-Binding Protein Cycle: Burning Off PI(4)P to Transport Cholesterol. Annu. Rev. Biochem. 2018;87:809–837. doi: 10.1146/annurev-biochem-061516-044924. [DOI] [PubMed] [Google Scholar]
- 13.Pietrangelo A., Ridgway N.D. Bridging the molecular and biological functions of the oxysterol-binding protein family. Cell. Mol. Life Sci. 2018;75:3079–3098. doi: 10.1007/s00018-018-2795-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Olkkonen V.M. OSBP-Related Protein Family in Lipid Transport over Membrane Contact Sites. Lipid Insights. 2015;8s1:LPI-S31726. doi: 10.4137/LPI.S31726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mackrill J.J. Oxysterols and calcium signal transduction. Chem. Phys. Lipids. 2011;164:488–495. doi: 10.1016/j.chemphyslip.2011.04.001. [DOI] [PubMed] [Google Scholar]
- 16.Zhong W., Yi Q., Xu B., Li S., Wang T., Liu F., Zhu B., Hoffmann P.R., Ji G., Lei P., et al. ORP4L is essential for T-cell acute lymphoblastic leukemia cell survival. Nat. Commun. 2016;7:12702. doi: 10.1038/ncomms12702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cao X., Chen J., Li D., Xie P., Xu M., Lin W., Li S., Pan G., Tang Y., Xu J., et al. ORP4L couples IP3to ITPR1 in control of endoplasmic reticulum calcium release. FASEB J. 2019;33:13852–13865. doi: 10.1096/fj.201900933RR. [DOI] [PubMed] [Google Scholar]
- 18.Lessmann E., Ngo M., Leitges M., Minguet S., Ridgway N.D., Huber M. Oxysterol-binding protein-related protein (ORP) 9 is a PDK-2 substrate and regulates Akt phosphorylation. Cell. Signal. 2007;19:384–392. doi: 10.1016/j.cellsig.2006.07.009. [DOI] [PubMed] [Google Scholar]
- 19.Strating J., Van Der Linden L., Albulescu L., Bigay J., Arita M., Delang L., Leyssen P., Van Der Schaar H.M., Lanke K.H., Thibaut H.J., et al. Itraconazole Inhibits Enterovirus Replication by Targeting the Oxysterol-Binding Protein. Cell Rep. 2015;10:600–615. doi: 10.1016/j.celrep.2014.12.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ma L., Nelson E.R. Oxysterols and nuclear receptors. Mol. Cell. Endocrinol. 2019;484:42–51. doi: 10.1016/j.mce.2019.01.016. [DOI] [PubMed] [Google Scholar]
- 21.Korf H., Beken S.V., Romano M., Steffensen K., Stijlemans B., Gustafsson J., Grooten J., Huygen K. Liver X receptors contribute to the protective immune response against Mycobacterium tuberculosis in mice. J. Clin. Investig. 2009;119:1626–1637. doi: 10.1172/JCI35288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.A DeBose-Boyd R., Brown M.S., Li W.-P., Nohturfft A., Goldstein J.L., Espenshade P.J. Transport-Dependent Proteolysis of SREBP: Relocation of Site-1 Protease from Golgi to ER Obviates the Need for SREBP Transport to Golgi. Cell. 1999;99:703–712. doi: 10.1016/S0092-8674(00)81668-2. [DOI] [PubMed] [Google Scholar]
- 23.Gong Y., Lee J.N., Brown M.S., Goldstein J.L., Ye J. Juxtamembranous aspartic acid in Insig-1 and Insig-2 is required for cholesterol homeostasis. Proc. Natl. Acad. Sci. USA. 2006;103:6154–6159. doi: 10.1073/pnas.0601923103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang Y., Kumar N., Crumbley C., Griffin P.R., Burris T.P. A second class of nuclear receptors for oxysterols: Regulation of RORα and RORγ activity by 24S-hydroxycholesterol (cerebrosterol) Mol. Cell Biol. Lipids. 2010;1801:917–923. doi: 10.1016/j.bbalip.2010.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.DuSell C.D., Umetani M., Shaul P.W., Mangelsdorf D., McDonnell D.P. 27-Hydroxycholesterol Is an Endogenous Selective Estrogen Receptor Modulator. Mol. Endocrinol. 2008;22:65–77. doi: 10.1210/me.2007-0383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Umetani M., Domoto H., Gormley A.K., Yuhanna I.S., Cummins C., Javitt N.B., Korach K., Shaul P.W., Mangelsdorf D. 27-Hydroxycholesterol is an endogenous SERM that inhibits the cardiovascular effects of estrogen. Nat. Med. 2007;13:1185–1192. doi: 10.1038/nm1641. [DOI] [PubMed] [Google Scholar]
- 27.Wang Y., Chen L., Pandak W.M., Heuman D., Hylemon P.B., Ren S. High Glucose Induces Lipid Accumulation via 25-Hydroxycholesterol DNA-CpG Methylation. iScience. 2020;23:101102. doi: 10.1016/j.isci.2020.101102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bauman D.R., Bitmansour A.D., McDonald J.G., Thompson B., Liang G., Russell D.W. 25-Hydroxycholesterol secreted by macrophages in response to Toll-like receptor activation suppresses immunoglobulin A production. Proc. Natl. Acad. Sci. USA. 2009;106:16764–16769. doi: 10.1073/pnas.0909142106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sanchez L.D., Pontini L., Marinozzi M., Sanchez-Aranguren L.C., Reis A., Dias I.H. Cholesterol and oxysterol sulfates: Pathophysiological roles and analytical challenges. Br. J. Pharm. 2020;178:3327–3341. doi: 10.1111/bph.15227. [DOI] [PubMed] [Google Scholar]
- 30.Wang Y., Lin W., Brown J.E., Chen L., Pandak W.M., Hylemon P.B., Ren S. 25-Hydroxycholesterol 3-sulfate is an endogenous ligand of DNA methyltransferases in hepatocytes. J. Lipid Res. 2021;62:100063. doi: 10.1016/j.jlr.2021.100063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang Y., Li X., Ren S. Cholesterol Metabolites 25-Hydroxycholesterol and 25-Hydroxycholesterol 3-Sulfate Are Potent Paired Regulators: From Discovery to Clinical Usage. Metabolites. 2020;11:9. doi: 10.3390/metabo11010009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bottemanne P., Paquot A., Ameraoui H., Guillemot-Legris O., Alhouayek M., Muccioli G.G. 25-Hydroxycholesterol metabolism is altered by lung inflammation, and its local administration modulates lung inflammation in mice. FASEB J. 2021;35:e21514. doi: 10.1096/fj.202002555R. [DOI] [PubMed] [Google Scholar]
- 33.Jiang W., Miyamoto T., Kakizawa T., Nishio S.-I., Oiwa A., Takeda T., Suzuki S., Hashizume K. Inhibition of LXRα signaling by vitamin D receptor: Possible role of VDR in bile acid synthesis. Biochem. Biophys. Res. Commun. 2006;351:176–184. doi: 10.1016/j.bbrc.2006.10.027. [DOI] [PubMed] [Google Scholar]
- 34.Wamil M., Andrew R., Chapman K., Street J., Morton N.M., Seckl J.R. 7-Oxysterols Modulate Glucocorticoid Activity in Adipocytes through Competition for 11β-Hydroxysteroid Dehydrogenase Type. Endocrinology. 2008;149:5909–5918. doi: 10.1210/en.2008-0420. [DOI] [PubMed] [Google Scholar]
- 35.Repa J., Liang G., Ou J., Bashmakov Y., Lobaccaro J.-M.A., Shimomura I., Shan B., Brown M.S., Goldstein J.L., Mangelsdorf D. Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRalpha and LXRbeta. Genes Dev. 2000;14:2819–2830. doi: 10.1101/gad.844900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Liang G., Yang J., Horton J.D., Hammer R.E., Goldstein J.L., Brown M.S. Diminished Hepatic Response to Fasting/Refeeding and Liver X Receptor Agonists in Mice with Selective Deficiency of Sterol Regulatory Element-binding Protein-1c. J. Biol. Chem. 2002;277:9520–9528. doi: 10.1074/jbc.M111421200. [DOI] [PubMed] [Google Scholar]
- 37.Maïga A., Lemieux S., Pabst C., Lavallée V.-P., Bouvier M., Sauvageau G., Hébert J. Transcriptome analysis of G protein-coupled receptors in distinct genetic subgroups of acute myeloid leukemia: Identification of potential disease-specific targets. Blood Cancer J. 2016;6:e431. doi: 10.1038/bcj.2016.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Schmiedel B.J., Singh D., Madrigal A., Valdovino-Gonzalez A.G., White B.M., Zapardiel-Gonzalo J., Ha B., Altay G., Greenbaum J.A., McVicker G., et al. Impact of Genetic Polymorphisms on Human Immune Cell Gene Expression. Cell. 2018;175:1701–1715. doi: 10.1016/j.cell.2018.10.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Villani A.-C., Satija R., Reynolds G., Sarkizova S., Shekhar K., Fletcher J., Griesbeck M., Butler A., Zheng S., Lazo S., et al. Single-cell RNA-seq reveals new types of human blood dendritic cells, monocytes, and progenitors. Science. 2017;356:eaah4573. doi: 10.1126/science.aah4573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rosenkilde M.M., Benned-Jensen T., Andersen H., Holst P.J., Kledal T.N., Luttichau H.R., Larsen J.K., Christensen J., Schwartz T.W. Molecular Pharmacological Phenotyping of EBI2. J. Biol. Chem. 2006;281:13199–13208. doi: 10.1074/jbc.M602245200. [DOI] [PubMed] [Google Scholar]
- 41.Yi T., Wang X., Kelly L.M., An J., Xu Y., Sailer A., Gustafsson J.-A., Russell D., Cyster J.G. Oxysterol Gradient Generation by Lymphoid Stromal Cells Guides Activated B Cell Movement during Humoral Responses. Immunity. 2012;37:535–548. doi: 10.1016/j.immuni.2012.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kelly L.M., Pereira J., Yi T., Xu Y., Cyster J.G. EBI2 Guides Serial Movements of Activated B Cells and Ligand Activity Is Detectable in Lymphoid and Nonlymphoid Tissues. J. Immunol. 2011;187:3026–3032. doi: 10.4049/jimmunol.1101262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gatto M., Wood K., Brink R. EBI2 Operates Independently of but in Cooperation with CXCR5 and CCR7 To Direct B Cell Migration and Organization in Follicles and the Germinal Center. J. Immunol. 2011;187:4621–4628. doi: 10.4049/jimmunol.1101542. [DOI] [PubMed] [Google Scholar]
- 44.Pereira J., Kelly L.M., Xu Y., Cyster J.G. EBI2 mediates B cell segregation between the outer and centre follicle. Nat. Cell Biol. 2009;460:1122–1126. doi: 10.1038/nature08226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kim S.T., Choi J.-Y., Lainez B., Schulz V.P., Karas D.E., Baum E.D., Setlur J., Gallagher P.G., Craft J. Human Extrafollicular CD4+ Th Cells Help Memory B Cells Produce Igs. J. Immunol. 2018;201:1359–1372. doi: 10.4049/jimmunol.1701217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ki S., Thyagarajan H.M., Hu Z., Lancaster J.N., Ehrlich L.I. EBI2 contributes to the induction of thymic central tolerance in mice by promoting rapid motility of medullary thymocytes. Eur. J. Immunol. 2017;47:1906–1917. doi: 10.1002/eji.201747020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Baptista A.P., Gola A., Huang Y., Milanez-Almeida P., Torabi-Parizi P., Urban J., Shapiro V.S., Gerner M.Y., Germain R.N. The Chemoattractant Receptor Ebi2 Drives Intranodal Naive CD4+ T Cell Peripheralization to Promote Effective Adaptive Immunity. Immunity. 2019;50:1188–1201. doi: 10.1016/j.immuni.2019.04.001. [DOI] [PubMed] [Google Scholar]
- 48.Li J., Lu E., Yi T., Cyster J.G. EBI2 augments Tfh cell fate by promoting interaction with IL-2-quenching dendritic cells. Nat. Cell Biol. 2016;533:110–114. doi: 10.1038/nature17947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yi T., Cyster J.G. EBI2-mediated bridging channel positioning supports splenic dendritic cell homeostasis and particulate antigen capture. eLife. 2013;2:e00757. doi: 10.7554/eLife.00757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lu E., Dang E.V., McDonald J.G., Cyster J.G. Distinct oxysterol requirements for positioning naïve and activated dendritic cells in the spleen. Sci. Immunol. 2017;2:eaal5237. doi: 10.1126/sciimmunol.aal5237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Chiang E.Y., Johnston R.J., Grogan J.L. EBI2 Is a Negative Regulator of Type I Interferons in Plasmacytoid and Myeloid Dendritic Cells. PLoS ONE. 2013;8:e83457. doi: 10.1371/journal.pone.0083457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Shen Z.-J., Hu J., Kashi V.P., Kelly E., Denlinger L.C., Lutchman K., McDonald J.G., Jarjour N.N., Malter J.S. Epstein-Barr Virus–induced Gene 2 Mediates Allergen-induced Leukocyte Migration into Airways. Am. J. Respir. Crit. Care Med. 2017;195:1576–1585. doi: 10.1164/rccm.201608-1580OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Emgård J., Kammoun H., García-Cassani B., Chesné J., Parigi S.M., Jacob J.-M., Cheng H.-W., Evren E., Das S., Czarnewski P., et al. Oxysterol Sensing through the Receptor GPR183 Promotes the Lymphoid-Tissue-Inducing Function of Innate Lymphoid Cells and Colonic Inflammation. Immunity. 2018;48:120–132. doi: 10.1016/j.immuni.2017.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wyss A., Raselli T., Perkins N., Ruiz F., Schmelczer G., Klinke G., Moncsek A., Roth R., Spalinger M.R., Hering L., et al. The EBI2-oxysterol axis promotes the development of intestinal lymphoid structures and colitis. Mucosal Immunol. 2019;12:733–745. doi: 10.1038/s41385-019-0140-x. [DOI] [PubMed] [Google Scholar]
- 55.Mutemberezi V., Buisseret B., Masquelier J., Guillemot-Legris O., Alhouayek M., Muccioli G.G. Oxysterol levels and metabolism in the course of neuroinflammation: Insights from in vitro and in vivo models. J. Neuroinflamm. 2018;15:1–16. doi: 10.1186/s12974-018-1114-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rutkowska A., O’Sullivan S.A., Christen I., Zhang J., Sailer A., Dev K.K. The EBI2 signalling pathway plays a role in cellular crosstalk between astrocytes and macrophages. Sci. Rep. 2016;6:25520. doi: 10.1038/srep25520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Barington L., Wanke F., Arfelt K.N., Holst P.J., Kurschus F.C., Rosenkilde M.M. EBI2 in splenic and local immune responses and in autoimmunity. J. Leukoc. Biol. 2018;104:313–322. doi: 10.1002/JLB.2VMR1217-510R. [DOI] [PubMed] [Google Scholar]
- 58.Van Der Poel M., Ulas T., Mizee M.R., Hsiao C.-C., Miedema S., Adelia, Schuurman K.G., Helder B., Tas S.W., Schultze J.L., et al. Transcriptional profiling of human microglia reveals grey–white matter heterogeneity and multiple sclerosis-associated changes. Nat. Commun. 2019;10:1139. doi: 10.1038/s41467-019-08976-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Braden K., Giancotti L.A., Chen Z., DeLeon C., Latzo N., Boehm T., D’Cunha N., Thompson B.M., Doyle T.M., McDonald J.G., et al. GPR183-Oxysterol Axis in Spinal Cord Contributes to Neuropathic Pain. J. Pharm. Exp. 2020;375:349–357. doi: 10.1124/jpet.120.000105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wanke F., Moos S., Croxford A.L., Heinen A.P., Gräf S., Kalt B., Tischner D., Zhang J., Christen I., Bruttger J., et al. EBI2 Is Highly Expressed in Multiple Sclerosis Lesions and Promotes Early CNS Migration of Encephalitogenic CD4 T Cells. Cell Rep. 2017;18:1270–1284. doi: 10.1016/j.celrep.2017.01.020. [DOI] [PubMed] [Google Scholar]
- 61.Clottu A.S., Mathias A., Sailer A.W., Schluep M., Seebach J., Du Pasquier R., Pot C. EBI2 Expression and Function: Robust in Memory Lymphocytes and Increased by Natalizumab in Multiple Sclerosis. Cell Rep. 2017;18:213–224. doi: 10.1016/j.celrep.2016.12.006. [DOI] [PubMed] [Google Scholar]
- 62.Chalmin F., Rochemont V., Lippens C., Clottu A., Sailer A., Merkler D., Hugues S., Pot C. Oxysterols regulate encephalitogenic CD4+ T cell trafficking during central nervous system autoimmunity. J. Autoimmun. 2015;56:45–55. doi: 10.1016/j.jaut.2014.10.001. [DOI] [PubMed] [Google Scholar]
- 63.Nevius E., Pinho F., Dhodapkar M., Jin H., Nadrah K., Horowitz M.C., Kikuta J., Ishii M., Pereira J.P. Oxysterols and EBI2 promote osteoclast precursor migration to bone surfaces and regulate bone mass homeostasis. J. Exp. Med. 2015;212:1931–1946. doi: 10.1084/jem.20150088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hannedouche S., Zhang J., Yi T., Shen W., Nguyen D., Pereira J., Guerini D., Baumgarten B.U., Roggo S., Wen B., et al. Oxysterols direct immune cell migration via EBI2. Nat. Cell Biol. 2011;475:524–527. doi: 10.1038/nature10280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Liu C., Yang X.V., Wu J., Kuei C., Mani N.S., Zhang L., Yu J., Sutton S.W., Qin N., Banie H., et al. Oxysterols direct B-cell migration through EBI2. Nat. Cell Biol. 2011;475:519–523. doi: 10.1038/nature10226. [DOI] [PubMed] [Google Scholar]
- 66.Honda A., Miyazaki T., Ikegami T., Iwamoto J., Maeda T., Hirayama T., Saito Y., Teramoto T., Matsuzaki Y. Cholesterol 25-hydroxylation activity of CYP3A. J. Lipid Res. 2011;52:1509–1516. doi: 10.1194/jlr.M014084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Griffiths W.J., Wang Y. Oxysterols as lipid mediators: Their biosynthetic genes, enzymes and metabolites. Prostaglandins Other Lipid Mediat. 2019;147:106381. doi: 10.1016/j.prostaglandins.2019.106381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Park K., Scott A.L. Cholesterol 25-hydroxylase production by dendritic cells and macrophages is regulated by type I interferons. J. Leukoc. Biol. 2010;88:1081–1087. doi: 10.1189/jlb.0610318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Diczfalusy U., Olofsson K.E., Carlsson A.-M., Gong M., Golenbock D.T., Rooyackers O., Fläring U., Björkbacka H. Marked upregulation of cholesterol 25-hydroxylase expression by lipopolysaccharide. J. Lipid Res. 2009;50:2258–2264. doi: 10.1194/jlr.M900107-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Preuss I., Ludwig M.-G., Baumgarten B., Bassilana F., Gessier F., Seuwen K., Sailer A.W. Transcriptional regulation and functional characterization of the oxysterol/EBI2 system in primary human macrophages. Biochem. Biophys. Res. Commun. 2014;446:663–668. doi: 10.1016/j.bbrc.2014.01.069. [DOI] [PubMed] [Google Scholar]
- 71.Madenspacher J.H., Morrell E.D., Gowdy K.M., McDonald J.G., Thompson B.M., Muse G.W., Martinez J., Thomas S.Y., Mikacenic C., Nick J.A., et al. Cholesterol-25-hydroxylase promotes efferocytosis and resolution of lung inflammation. JCI Insight. 2020;5 doi: 10.1172/jci.insight.137189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Jia J., Conlon T.M., Sarker R.S., Taşdemir D., Smirnova N., Srivastava B., Verleden S., Güneş G., Wu X., Prehn C., et al. Cholesterol metabolism promotes B-cell positioning during immune pathogenesis of chronic obstructive pulmonary disease. EMBO Mol. Med. 2018;10:e8349. doi: 10.15252/emmm.201708349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Li-Hawkins J., Lund E.G., Turley S.D., Russell D.W. Disruption of the Oxysterol 7α-Hydroxylase Gene in Mice. J. Biol. Chem. 2000;275:16536–16542. doi: 10.1074/jbc.M001811200. [DOI] [PubMed] [Google Scholar]
- 74.Dulos J., Van Der Vleuten M.A.J., Kavelaars A., Heijnen C.J., Boots A.M. CYP7B expression and activity in fibroblast-like synoviocytes from patients with rheumatoid arthritis: Regulation by proinflammatory cytokines. Arthritis Rheum. 2005;52:770–778. doi: 10.1002/art.20950. [DOI] [PubMed] [Google Scholar]
- 75.Stoilov I., Krueger W., Mankowski D., Guernsey L., Kaur A., Glynn J., Thrall R.S. The cytochromes P450 (CYP) response to allergic inflammation of the lung. Arch. Biochem. Biophys. 2006;456:30–38. doi: 10.1016/j.abb.2006.09.029. [DOI] [PubMed] [Google Scholar]
- 76.Raselli T., Hearn T., Wyss A., Atrott K., Peter A., Frey-Wagner I., Spalinger M.R., Maggio E.M., Sailer A.W., Schmitt J., et al. Elevated oxysterol levels in human and mouse livers reflect nonalcoholic steatohepatitis. J. Lipid Res. 2019;60:1270–1283. doi: 10.1194/jlr.M093229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kakiyama G., Marques D., Martin R., Takei H., Rodriguez-Agudo D., LaSalle S.A., Hashiguchi T., Liu X., Green R., Erickson S., et al. Insulin resistance dysregulates CYP7B1 leading to oxysterol accumulation: A pathway for NAFL to NASH transition. J. Lipid Res. 2020;61:1629–1644. doi: 10.1194/jlr.RA120000924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Shi S.-Z., Lee E.-J., Lin Y.-J., Chen L., Zheng H.-Y., He X.-Q., Peng J.-Y., Noonepalle S.K., Shull A.Y., Pei F.C., et al. Recruitment of monocytes and epigenetic silencing of intratumoral CYP7B1 primarily contribute to the accumulation of 27-hydroxycholesterol in breast cancer. Am. J. Cancer Res. 2019;9:2194–2208. [PMC free article] [PubMed] [Google Scholar]
- 79.Benned-Jensen T., Rosenkilde M.M. Structural Motifs of Importance for the Constitutive Activity of the Orphan 7TM Receptor EBI2: Analysis of Receptor Activation in the Absence of an Agonist. Mol. Pharm. 2008;74:1008–1021. doi: 10.1124/mol.108.049676. [DOI] [PubMed] [Google Scholar]
- 80.Benned-Jensen T., Rosenkilde M.M. Distinct expression and ligand-binding profiles of two constitutively active GPR17 splice variants. Br. J. Pharm. 2010;159:1092–1105. doi: 10.1111/j.1476-5381.2009.00633.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Zhang L., Shih A.Y., Yang X.V., Kuei C., Wu J., Deng X., Mani N.S., Mirzadegan T., Sun S., Lovenberg T.W., et al. Identification of Structural Motifs Critical for Epstein-Barr Virus-Induced Molecule 2 Function and Homology Modeling of the Ligand Docking Site. Mol. Pharm. 2012;82:1094–1103. doi: 10.1124/mol.112.080275. [DOI] [PubMed] [Google Scholar]
- 82.Benned-Jensen T., Norn C., Laurent S., Madsen C.M., Larsen H.M., Arfelt K.N., Wolf R.M., Frimurer T., Sailer A., Rosenkilde M.M. Molecular Characterization of Oxysterol Binding to the Epstein-Barr Virus-induced Gene 2 (GPR183) J. Biol. Chem. 2012;287:35470–35483. doi: 10.1074/jbc.M112.387894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sensi C., Daniele S., Parravicini C., Zappelli E., Russo V., Trincavelli M.L., Martini C., Abbracchio M.P., Eberini I. Oxysterols act as promiscuous ligands of class-A GPCRs: In silico molecular modeling and in vitro validation. Cell. Signal. 2014;26:2614–2620. doi: 10.1016/j.cellsig.2014.08.003. [DOI] [PubMed] [Google Scholar]
- 84.Daugvilaite V., Madsen C.M., Lückmann M., Echeverria C.C., Sailer A.W., Frimurer T.M., Rosenkilde M.M., Benned-Jensen T. Biased agonism and allosteric modulation of G protein-coupled receptor 183—A 7TM receptor also known as Epstein-Barr virus-induced gene 2. Br. J. Pharm. 2017;174:2031–2042. doi: 10.1111/bph.13801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Thal D.M., Glukhova A., Sexton P., Christopoulos A. Structural insights into G-protein-coupled receptor allostery. Nat. Cell Biol. 2018;559:45–53. doi: 10.1038/s41586-018-0259-z. [DOI] [PubMed] [Google Scholar]
- 86.Wang Y., Yu Z., Xiao W., Lu S., Zhang J. Allosteric binding sites at the receptor–lipid bilayer interface: Novel targets for GPCR drug discovery. Drug Discov. Today. 2020;26:690–703. doi: 10.1016/j.drudis.2020.12.001. [DOI] [PubMed] [Google Scholar]
- 87.Guixà-González R., Albasanz J.L., Rodriguez-Espigares I., Pastor M., Sanz F., Marti-Solano M., Manna M., Martinez-Seara H., Hildebrand P.W., Martín M., et al. Membrane cholesterol access into a G-protein-coupled receptor. Nat. Commun. 2017;8:14505. doi: 10.1038/ncomms14505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Coffelt S., Kersten K., Doornebal C.W., Weiden J., Vrijland K., Hau C.-S., Verstegen N., Ciampricotti M., Hawinkels L., Jonkers J., et al. IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nat. Cell Biol. 2015;522:345–348. doi: 10.1038/nature14282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Baek A.E., Yu Y.-R.A., He S., Wardell S.E., Chang C.-Y., Kwon S., Pillai R.V., McDowell H.B., Thompson J.W., Dubois L.G., et al. The cholesterol metabolite 27 hydroxycholesterol facilitates breast cancer metastasis through its actions on immune cells. Nat. Commun. 2017;8:864. doi: 10.1038/s41467-017-00910-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Addison C.L., Daniel T.O., Burdick M.D., Liu H., Ehlert J.E., Xue Y.Y., Buechi L., Walz A., Richmond A., Strieter R.M. The CXC Chemokine Receptor 2, CXCR2, Is the Putative Receptor for ELR + CXC Chemokine-Induced Angiogenic Activity. J. Immunol. 2000;165:5269–5277. doi: 10.4049/jimmunol.165.9.5269. [DOI] [PubMed] [Google Scholar]
- 91.Khaw Y.M., Cunningham C., Tierney A., Sivaguru M., Inoue M. Neutrophil-selective deletion of Cxcr2 protects against CNS neurodegeneration in a mouse model of multiple sclerosis. J. Neuroinflamm. 2020;17:49. doi: 10.1186/s12974-020-1730-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Carlson T., Kroenke M., Rao P., Lane T.E., Segal B. The Th17–ELR + CXC chemokine pathway is essential for the development of central nervous system autoimmune disease. J. Exp. Med. 2008;205:811–823. doi: 10.1084/jem.20072404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Steele C.W., A Karim S., Foth M., Rishi L., Leach J., Porter R., Nixon C., Evans T.J., Carter C.R., Nibbs R., et al. CXCR2 inhibition suppresses acute and chronic pancreatic inflammation. J. Pathol. 2015;237:85–97. doi: 10.1002/path.4555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Farooq S.M., Stillie R., Svensson M., Svanborg C., Strieter R.M., Stadnyk A.W. Therapeutic Effect of Blocking CXCR2 on Neutrophil Recruitment and Dextran Sodium Sulfate-Induced Colitis. J. Pharm. Exp. 2009;329:123–129. doi: 10.1124/jpet.108.145862. [DOI] [PubMed] [Google Scholar]
- 95.Zhu F., He H., Fan L., Ma C., Xu Z., Xue Y., Wang Y., Zhang C., Zhou G. Blockade of CXCR2 suppresses proinflammatory activities of neutrophils in ulcerative colitis. Am. J. Transl. Res. 2020;12:5237–5251. [PMC free article] [PubMed] [Google Scholar]
- 96.Sinclair A., Park L., Shah M., Drotar M., Calaminus S., Hopcroft L.E.M., Kinstrie R., Guitart A., Dunn K., Abraham S., et al. CXCR2 and CXCL4 regulate survival and self-renewal of hematopoietic stem/progenitor cells. Blood. 2016;128:371–383. doi: 10.1182/blood-2015-08-661785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Eash K.J., Greenbaum A., Gopalan P.K., Link D.C. CXCR2 and CXCR4 antagonistically regulate neutrophil trafficking from murine bone marrow. J. Clin. Investig. 2010;120:2423–2431. doi: 10.1172/JCI41649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Han X., Shi H., Sun Y., Shang C., Luan T., Wang D., Ba X., Zeng X. CXCR2 expression on granulocyte and macrophage progenitors under tumor conditions contributes to mo-MDSC generation via SAP18/ERK/STAT3. Cell Death Dis. 2019;10:598. doi: 10.1038/s41419-019-1837-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ahuja S.K., Murphy P.M. The CXC Chemokines Growth-regulated Oncogene (GRO)α, GROβ, GROγ, Neutrophil-activating Peptide-2, and Epithelial Cell-derived Neutrophil-activating Peptide-78 Are Potent Agonists for the Type B, but Not the Type A, Human Interleukin-8 Receptor. J. Biol. Chem. 1996;271:20545–20550. doi: 10.1074/jbc.271.34.20545. [DOI] [PubMed] [Google Scholar]
- 100.Das S.T., Rajagopalan L., Guerrero-Plata A., Sai J., Richmond A., Garofalo R.P., Rajarathnam K. Monomeric and Dimeric CXCL8 Are Both Essential for In Vivo Neutrophil Recruitment. PLoS ONE. 2010;5:e11754. doi: 10.1371/journal.pone.0011754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Shaik S.S., Soltau T.D., Chaturvedi G., Totapally B., Hagood J.S., Andrews W.W., Athar M., Voitenok N.N., Killingsworth C.R., Patel R., et al. Low Intensity Shear Stress Increases Endothelial ELR + CXC Chemokine Production via a Focal Adhesion Kinase-p38β MAPK-NF-κB Pathway. J. Biol. Chem. 2009;284:5945–5955. doi: 10.1074/jbc.M807205200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Raccosta L., Fontana R., Traversari C., Russo V. Oxysterols recruit tumor-supporting neutrophils within the tumor microenvironment. OncoImmunology. 2013;2:e26469. doi: 10.4161/onci.26469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Raccosta L., Fontana R., Maggioni D., Lanterna C., Villablanca E., Paniccia A., Musumeci A., Chiricozzi E., Trincavelli M.L., Daniele S., et al. The oxysterol–CXCR2 axis plays a key role in the recruitment of tumor-promoting neutrophils. J. Exp. Med. 2013;210:1711–1728. doi: 10.1084/jem.20130440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Liu K., Wu L., Yuan S., Wu M., Xu Y., Sun Q., Li S., Zhao S., Hua T., Liu Z.-J. Structural basis of CXC chemokine receptor 2 activation and signalling. Nat. Cell Biol. 2020;585:135–140. doi: 10.1038/s41586-020-2492-5. [DOI] [PubMed] [Google Scholar]
- 105.Catusse J., Liotard A., Loillier B., Pruneau D., Paquet J.-L. Characterization of the molecular interactions of interleukin-8 (CXCL8), growth related oncogen α (CXCL1) and a non-peptide antagonist (SB 225002) with the human CXCR2. Biochem. Pharm. 2003;65:813–821. doi: 10.1016/S0006-2952(02)01619-2. [DOI] [PubMed] [Google Scholar]
- 106.Nicholls D.J., Tomkinson N.P., Wiley K.E., Brammall A., Bowers L., Grahames C., Gaw A., Meghani P., Shelton P., Wright T.J., et al. Identification of a Putative Intracellular Allosteric Antagonist Binding-Site in the CXC Chemokine Receptors 1 and 2. Mol. Pharm. 2008;74:1193–1202. doi: 10.1124/mol.107.044610. [DOI] [PubMed] [Google Scholar]
- 107.De Kruijf P., Van Heteren J., Lim H.D., Conti P.G., Van Der Lee M.M.C., Bosch L., Ho K.-K., Auld U., Ohlmeyer M., Smit M.J., et al. Nonpeptidergic Allosteric Antagonists Differentially Bind to the CXCR2 Chemokine Receptor. J. Pharm. Exp. 2009;329:783–790. doi: 10.1124/jpet.108.148387. [DOI] [PubMed] [Google Scholar]
- 108.Salchow K., Bond M., Evans S., Press N., Charlton S., Hunt P., Bradley M. A common intracellular allosteric binding site for antagonists of the CXCR2 receptor. Br. J. Pharm. 2010;159:1429–1439. doi: 10.1111/j.1476-5381.2009.00623.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Baugher P.J., Richmond A. The Carboxyl-terminal PDZ Ligand Motif of Chemokine Receptor CXCR2 Modulates Post-endocytic Sorting and Cellular Chemotaxis. J. Biol. Chem. 2008;283:30868–30878. doi: 10.1074/jbc.M804054200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Wu Y., Wang S., Farooq S.M., Castelvetere M.P., Hou Y., Gao J.-L., Navarro J., Oupicky D., Sun F., Li C. A Chemokine Receptor CXCR2 Macromolecular Complex Regulates Neutrophil Functions in Inflammatory Diseases. J. Biol. Chem. 2012;287:5744–5755. doi: 10.1074/jbc.M111.315762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Legler D.F., Matti C., Laufer J.M., Jakobs B.D., Purvanov V., Allmen E.U.-V., Thelen M. Modulation of Chemokine Receptor Function by Cholesterol: New Prospects for Pharmacological Intervention. Mol. Pharm. 2017;91:331–338. doi: 10.1124/mol.116.107151. [DOI] [PubMed] [Google Scholar]
- 112.Chen J., Taipale J., Young K.E., Maiti T., Beachy P.A. Small molecule modulation of Smoothened activity. Proc. Natl. Acad. Sci. USA. 2002;99:14071–14076. doi: 10.1073/pnas.182542899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Frank-Kamenetsky M., Zhang X.M., Bottega S., Guicherit O., Wichterle H., Dudek H., Bumcrot D., Wang F.Y., Jones S., Shulok J., et al. Small-molecule modulators of Hedgehog signaling: Identification and characterization of Smoothened agonists and antagonists. J. Biol. 2002;1:10. doi: 10.1186/1475-4924-1-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Rohatgi R., Milenkovic L., Corcoran R.B., Scott M.P. Hedgehog signal transduction by Smoothened: Pharmacologic evidence for a 2-step activation process. Proc. Natl. Acad. Sci. USA. 2009;106:3196–3201. doi: 10.1073/pnas.0813373106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Outram S.V., Varas A., Pepicelli C.V., Crompton T. Hedgehog Signaling Regulates Differentiation from Double-Negative to Double-Positive Thymocyte. Immunity. 2000;13:187–197. doi: 10.1016/S1074-7613(00)00019-4. [DOI] [PubMed] [Google Scholar]
- 116.Shah D.K., Hager-Theodorides A., Outram S.V., Ross S.E., Varas A., Crompton T. Reduced Thymocyte Development in Sonic Hedgehog Knockout Embryos. J. Immunol. 2004;172:2296–2306. doi: 10.4049/jimmunol.172.4.2296. [DOI] [PubMed] [Google Scholar]
- 117.Rowbotham N.J., Hager-Theodorides A.L., Cebecauer M., Shah D.K., Drakopoulou E., Dyson J., Outram S.V., Crompton T. Activation of the Hedgehog signaling pathway in T-lineage cells inhibits TCR repertoire selection in the thymus and peripheral T-cell activation. Blood. 2007;109:3757–3766. doi: 10.1182/blood-2006-07-037655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Rowbotham N.J., Furmanski A.L., Hager-Theodorides A.L., Ross S.E., Drakopoulou E., Koufaris C., Outram S.V., Crompton T. Repression of Hedgehog signal transduction in T-lineage cells increases TCR-induced activation and proliferation. Cell Cycle. 2008;7:904–908. doi: 10.4161/cc.7.7.5628. [DOI] [PubMed] [Google Scholar]
- 119.Mengrelis K., Lau C.-I., Rowell J., Solanki A., Norris S., Ross S., Ono M., Outram S., Crompton T. Sonic Hedgehog Is a Determinant of γδ T-Cell Differentiation in the Thymus. Front. Immunol. 2019;10:1629. doi: 10.3389/fimmu.2019.01629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Saldana J.I., Solanki A., Lau C.-I., Sahni H., Ross S., Furmanski A.L., Ono M., Holländer G., Crompton T. Sonic Hedgehog regulates thymic epithelial cell differentiation. J. Autoimmun. 2016;68:86–97. doi: 10.1016/j.jaut.2015.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Furmanski A.L., Saldana J.I., Ono M., Sahni H., Paschalidis N., D’Acquisto F., Crompton T. Tissue-Derived Hedgehog Proteins Modulate Th Differentiation and Disease. J. Immunol. 2013;190:2641–2649. doi: 10.4049/jimmunol.1202541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.de la Roche M., Ritter A.T., Angus K.L., Dinsmore C., Earnshaw C.H., Reiter J.F., Griffiths G.M. Hedgehog Signaling Controls T Cell Killing at the Immunological Synapse. Science. 2013;342:1247–1250. doi: 10.1126/science.1244689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Syn W.-K., Witek R.P., Curbishley S.M., Jung Y., Choi S.S., Enrich B., Omenetti A., Agboola K.M., Fearing C.M., Tilg H., et al. Role for hedgehog pathway in regulating growth and function of invariant NKT cells. Eur. J. Immunol. 2009;39:1879–1892. doi: 10.1002/eji.200838890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Solanki A., Lau C.I., Saldaña J.I., Ross S., Crompton T. The transcription factor Gli3 promotes B cell development in fetal liver through repression of Shh. J. Exp. Med. 2017;214:2041–2058. doi: 10.1084/jem.20160852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Solanki A., Yanez D.C., Ross S., Lau C.I., Papaioannou E., Li J., Saldaña J.I., Crompton T. In the fetal thymus, Gli3 in thymic epithelial cells promotes thymocyte positive selection and differentiation by repression of Shh. Development. 2018;145 doi: 10.1242/dev.146910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Papaioannou E., Yánez D.C., Ross S., Lau C.-I., Solanki A., Chawda M.M., Virasami A., Ranz I., Ono M., O’Shaughnessy R.F.L., et al. Sonic Hedgehog signaling limits atopic dermatitis via Gli2-driven immune regulation. J. Clin. Investig. 2019;129:3153–3170. doi: 10.1172/JCI125170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Myers B.R., Sever N., Chong Y.C., Kim J., Belani J.D., Rychnovsky S., Bazan J.F., Beachy P.A. Hedgehog Pathway Modulation by Multiple Lipid Binding Sites on the Smoothened Effector of Signal Response. Dev. Cell. 2013;26:346–357. doi: 10.1016/j.devcel.2013.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Cooper M.K., Wassif C., Krakowiak P.A., Taipale J., Gong R., Kelley R.I., Porter F.D., Beachy P.A. A defective response to Hedgehog signaling in disorders of cholesterol biosynthesis. Nat. Genet. 2003;33:508–513. doi: 10.1038/ng1134. [DOI] [PubMed] [Google Scholar]
- 129.Xiao X., Tang J.-J., Peng C., Wang Y., Fu L., Qiu Z.-P., Xiong Y., Yang L.-F., Cui H.-W., He X.-L., et al. Cholesterol Modification of Smoothened Is Required for Hedgehog Signaling. Mol. Cell. 2017;66:154–162.e10. doi: 10.1016/j.molcel.2017.02.015. [DOI] [PubMed] [Google Scholar]
- 130.Raleigh D.R., Sever N., Choksi P.K., Sigg M., Hines K., Thompson B., Elnatan D., Jaishankar P., Bisignano P., Garcia-Gonzalo F., et al. Cilia-Associated Oxysterols Activate Smoothened. Mol. Cell. 2018;72:316–327.e5. doi: 10.1016/j.molcel.2018.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Huang P., Nedelcu D., Watanabe M., Jao C., Kim Y., Liu J., Salic A. Cellular Cholesterol Directly Activates Smoothened in Hedgehog Signaling. Cell. 2016;166:1176–1187. doi: 10.1016/j.cell.2016.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Zhang X., Zhao F., Wu Y., Yang J., Han G.W., Zhao S., Ishchenko A., Ye L., Lin X., Ding K., et al. Crystal structure of a multi-domain human smoothened receptor in complex with a super stabilizing ligand. Nat. Commun. 2017;8:15383. doi: 10.1038/ncomms15383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Huang P., Zheng S., Wierbowski B., Kim Y., Nedelcu D., Aravena L., Liu J., Kruse A.C., Salic A. Structural Basis of Smoothened Activation in Hedgehog Signaling. Cell. 2018;174:312–324. doi: 10.1016/j.cell.2018.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Deshpande I., Liang J., Hedeen D., Roberts K.J., Zhang Y., Ha B., Latorraca N.R., Faust B., Dror R.O., Beachy P.A., et al. Smoothened stimulation by membrane sterols drives Hedgehog pathway activity. Nat. Cell Biol. 2019;571:284–288. doi: 10.1038/s41586-019-1355-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Qi X., Friedberg L., De Bose-Boyd R., Long T., Li X. Sterols in an intramolecular channel of Smoothened mediate Hedgehog signaling. Nat. Chem. Biol. 2020;16:1368–1375. doi: 10.1038/s41589-020-0646-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Phelan K., Mahler H. Acute Exposure to 25-Hydroxy-cholesterol Selectively Reduces GABAb and Not GABAa Receptor-Mediated Synaptic Inhibition. Biochem. Biophys. Res. Commun. 1997;237:68–73. doi: 10.1006/bbrc.1997.7070. [DOI] [PubMed] [Google Scholar]
- 137.A Kuhn S., van Landeghem F., Zacharias R., Färber K., Rappert A., Pavlovic S., Hoffmann A., Nolte C., Kettenmann H. Microglia express GABAB receptors to modulate interleukin release. Mol. Cell. Neurosci. 2004;25:312–322. doi: 10.1016/j.mcn.2003.10.023. [DOI] [PubMed] [Google Scholar]
- 138.Huang S., Mao J., Wei B., Pei G. The anti-spasticity drug baclofen alleviates collagen-induced arthritis and regulates dendritic cells. J. Cell. Physiol. 2015;230:1438–1447. doi: 10.1002/jcp.24884. [DOI] [PubMed] [Google Scholar]
- 139.Hammoud Y., Rice T., Mackrill J.J. Oxysterols modulate calcium signalling in the A7r5 aortic smooth muscle cell-line. Biochimie. 2013;95:568–577. doi: 10.1016/j.biochi.2012.08.003. [DOI] [PubMed] [Google Scholar]
- 140.Yan Z., Li S., Liang Z., Tomić M., Stojilkovic S.S. The P2X7 Receptor Channel Pore Dilates under Physiological Ion Conditions. J. Gen. Physiol. 2008;132:563–573. doi: 10.1085/jgp.200810059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.McCarthy A.E., Yoshioka C., Mansoor S.E. Full-Length P2X7 Structures Reveal How Palmitoylation Prevents Channel Desensitization. Cell. 2019;179:659–670. doi: 10.1016/j.cell.2019.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Robinson L.E., Shridar M., Smith P., Murrell-Lagnado R.D. Plasma Membrane Cholesterol as a Regulator of Human and Rodent P2X7 Receptor Activation and Sensitization. J. Biol. Chem. 2014;289:31983–31994. doi: 10.1074/jbc.M114.574699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Lajdova I., Chorvát D., Chorvatova A. Rapid effects of 1α,25(OH)2D3 in resting human peripheral blood mononuclear cells. Eur. J. Pharm. 2008;586:14–23. doi: 10.1016/j.ejphar.2008.02.004. [DOI] [PubMed] [Google Scholar]
- 144.Olivier E., Dutot M., Regazzetti A., Laprévote O., Rat P. 25-Hydroxycholesterol induces both P2X7-dependent pyroptosis and caspase-dependent apoptosis in human skin model: New insights into degenerative pathways. Chem. Phys. Lipids. 2017;207:171–178. doi: 10.1016/j.chemphyslip.2017.06.001. [DOI] [PubMed] [Google Scholar]
- 145.Adinolfi E., Giuliani A.L., De Marchi E., Pegoraro A., Orioli E., Di Virgilio F. The P2X7 receptor: A main player in inflammation. Biochem. Pharm. 2018;151:234–244. doi: 10.1016/j.bcp.2017.12.021. [DOI] [PubMed] [Google Scholar]
- 146.Olivier E., Dutot M., Regazzetti A., Leguillier T., Dargère D., Auzeil N., Laprévote O., Rat P. P2X7-pannexin-1 and amyloid β-induced oxysterol input in human retinal cell: Role in age-related macular degeneration? Biochimie. 2016;127:70–78. doi: 10.1016/j.biochi.2016.04.014. [DOI] [PubMed] [Google Scholar]
- 147.A Grahames C.B., Michel A.D., Chessell I.P., A Humphrey P.P. Pharmacological characterization of ATP- and LPS-induced IL-1β release in human monocytes. Br. J. Pharm. 1999;127:1915–1921. doi: 10.1038/sj.bjp.0702732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Martinez-García J.J., Banaclocha H.M., Angosto D., De Torre-Minguela C., Baroja-Mazo A., Alarcón-Vila C., Martinez-Alarcon L., Amores-Iniesta J., Martín-Sánchez F., Ercole G.A., et al. P2X7 receptor induces mitochondrial failure in monocytes and compromises NLRP3 inflammasome activation during sepsis. Nat. Commun. 2019;10:2711. doi: 10.1038/s41467-019-10626-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Grassi F. The P2X7 Receptor as Regulator of T Cell Development and Function. Front. Immunol. 2020;11:1179. doi: 10.3389/fimmu.2020.01179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Tsukimoto M., Tokunaga A., Harada H., Kojima S. Blockade of murine T cell activation by antagonists of P2Y6 and P2X7 receptors. Biochem. Biophys. Res. Commun. 2009;384:512–518. doi: 10.1016/j.bbrc.2009.05.011. [DOI] [PubMed] [Google Scholar]
- 151.Schenk U., Frascoli M., Proietti M., Geffers R., Traggiai E., Buer J., Ricordi C., Westendorf A.M., Grassi F. ATP Inhibits the Generation and Function of Regulatory T Cells Through the Activation of Purinergic P2X Receptors. Sci. Signal. 2011;4:ra12. doi: 10.1126/scisignal.2001270. [DOI] [PubMed] [Google Scholar]
- 152.Frascoli M., Marcandalli J., Schenk U., Grassi F. Purinergic P2X7 Receptor Drives T Cell Lineage Choice and Shapes Peripheral γδ Cells. J. Immunol. 2012;189:174–180. doi: 10.4049/jimmunol.1101582. [DOI] [PubMed] [Google Scholar]
- 153.Beynon V., Quintana F.J., Weiner H.L. Activated Human CD4+CD45RO+ Memory T-Cells Indirectly Inhibit NLRP3 Inflammasome Activation through Downregulation of P2X7R Signalling. PLoS ONE. 2012;7:e39576. doi: 10.1371/journal.pone.0039576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.De Salles M., Menezes M., Siqueira R., Da Silva H.B., Amaral E.P., Castillo-Méndez S.I., Cunha I., Cassado A.D.A., Vieira F.S., Olivieri D.N., et al. P2X7 receptor drives Th1 cell differentiation and controls the follicular helper T cell population to protect against Plasmodium chabaudi malaria. Plos Pathog. 2017;13:e1006595. doi: 10.1371/journal.ppat.1006595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Da Silva H.B., Beura L.K., Wang H., Hanse E.A., Gore R., Scott M.C., Walsh D.A., Block K.E., Fonseca R., Yan Y., et al. The purinergic receptor P2RX7 directs metabolic fitness of long-lived memory CD8+ T cells. Nat. Cell Biol. 2018;559:264–268. doi: 10.1038/s41586-018-0282-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Dubyak G.R. P2X7 receptor regulation of non-classical secretion from immune effector cells. Cell. Microbiol. 2012;14:1697–1706. doi: 10.1111/cmi.12001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Baroja-Mazo A., Barberà-Cremades M., Pelegrín P. P2X7 Receptor Activation Impairs Exogenous MHC Class I Oligopeptides Presentation in Antigen Presenting Cells. PLoS ONE. 2013;8:e70577. doi: 10.1371/journal.pone.0070577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Figliuolo V.R., Savio L.E., Safya H., Nanini H.F., Bernardazzi C., Abalo A., de Souza H.S., Kanellopoulos J., Bobé P., Coutinho C.M., et al. P2X7 receptor promotes intestinal inflammation in chemically induced colitis and triggers death of mucosal regulatory T cells. Mol. Basis Dis. 2017;1863:1183–1194. doi: 10.1016/j.bbadis.2017.03.004. [DOI] [PubMed] [Google Scholar]
- 159.Salussolia C.L., Prodromou M.L., Borker P., Wollmuth L.P. Arrangement of Subunits in Functional NMDA Receptors. J. Neurosci. 2011;31:11295–11304. doi: 10.1523/JNEUROSCI.5612-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Zhu S., Gouaux E. Structure and symmetry inform gating principles of ionotropic glutamate receptors. Neuropharmacology. 2016;112:11–15. doi: 10.1016/j.neuropharm.2016.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Pérez-Otaño I., Larsen R., Wesseling I.P.-O.J.F. Emerging roles of GluN3-containing NMDA receptors in the CNS. Nat. Rev. Neurosci. 2016;17:623–635. doi: 10.1038/nrn.2016.92. [DOI] [PubMed] [Google Scholar]
- 162.Hogan-Cann A.D., Anderson C.M. Physiological Roles of Non-Neuronal NMDA Receptors. Trends Pharm. Sci. 2016;37:750–767. doi: 10.1016/j.tips.2016.05.012. [DOI] [PubMed] [Google Scholar]
- 163.Ganor Y., Levite M. The neurotransmitter glutamate and human T cells: Glutamate receptors and glutamate-induced direct and potent effects on normal human T cells, cancerous human leukemia and lymphoma T cells, and autoimmune human T cells. J. Neural Transm. 2014;121:983–1006. doi: 10.1007/s00702-014-1167-5. [DOI] [PubMed] [Google Scholar]
- 164.Paul S.M., Doherty J.J., Robichaud A.J., Belfort G.M., Chow B.Y., Hammond R.S., Crawford D., Linsenbardt A.J., Shu H.-J., Izumi Y., et al. The Major Brain Cholesterol Metabolite 24(S)-Hydroxycholesterol Is a Potent Allosteric Modulator of N-Methyl-d-Aspartate Receptors. J. Neurosci. 2013;33:17290–17300. doi: 10.1523/JNEUROSCI.2619-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Emnett C.M., Eisenman L.N., Mohan J., A Taylor A., Doherty J.J., Paul S.M., Zorumski C.F., Mennerick S. Interaction between positive allosteric modulators and trapping blockers of the NMDA receptor channel. Br. J. Pharm. 2015;172:1333–1347. doi: 10.1111/bph.13007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Sun M.-Y., Izumi Y., Benz A.M., Zorumski C.F., Mennerick S. Endogenous 24S-hydroxycholesterol modulates NMDAR-mediated function in hippocampal slices. J. Neurophysiol. 2016;115:1263–1272. doi: 10.1152/jn.00890.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Wilding T.J., Lopez M.N., Huettner J.E. Chimeric Glutamate Receptor Subunits Reveal the Transmembrane Domain Is Sufficient for NMDA Receptor Pore Properties but Some Positive Allosteric Modulators Require Additional Domains. J. Neurosci. 2016;36:8815–8825. doi: 10.1523/JNEUROSCI.0345-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Wei X., Nishi T., Kondou S., Kimura H., Mody I. Preferential enhancement of GluN2B-containing native NMDA receptors by the endogenous modulator 24S-hydroxycholesterol in hippocampal neurons. Neuropharmacology. 2018;148:11–20. doi: 10.1016/j.neuropharm.2018.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Sun M.-Y., Taylor A., Zorumski C.F., Mennerick S. 24S-hydroxycholesterol and 25-hydroxycholesterol differentially impact hippocampal neuronal survival following oxygen-glucose deprivation. PLoS ONE. 2017;12:e0174416. doi: 10.1371/journal.pone.0174416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Mateos L., Akterin S., Gil-Bea F., Spulber S., Rahman A., Björkhem I., Schultzberg M., Flores-Morales A., Cedazo-Mínguez A. Activity-Regulated Cytoskeleton-Associated Protein in Rodent Brain is Down-Regulated by High Fat Dietin vivoand by 27-Hydroxycholesterolin vitro. Brain Pathol. 2008;19:69–80. doi: 10.1111/j.1750-3639.2008.00174.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Boldyrev A.A., Carpenter D.O., Johnson P. Emerging evidence for a similar role of glutamate receptors in the nervous and immune systems. J. Neurochem. 2005;95:913–918. doi: 10.1111/j.1471-4159.2005.03456.x. [DOI] [PubMed] [Google Scholar]
- 172.Mashkina A.P., Tyulina O.V., Solovyova T.I., Kovalenko E., Kanevski L.M., Johnson P., Boldyrev A.A. The excitotoxic effect of NMDA on human lymphocyte immune function. Neurochem. Int. 2007;51:356–360. doi: 10.1016/j.neuint.2007.04.009. [DOI] [PubMed] [Google Scholar]
- 173.Kahlfuß S., Simma N., Mankiewicz J., Bose T., Lowinus T., Klein-Hessling S., Sprengel R., Schraven B., Heine M., Bommhardt U. Immunosuppression by N-Methyl-d-Aspartate Receptor Antagonists Is Mediated through Inhibition of Kv1.3 and KCa3.1 Channels in T Cells. Mol. Cell. Biol. 2013;34:820–831. doi: 10.1128/MCB.01273-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Simma N., Bose T., Kahlfuß S., Mankiewicz J., Lowinus T., Lühder F., Schüler T., Schraven B., Heine M., Bommhardt U. NMDA-receptor antagonists block B-cell function but foster IL-10 production in BCR/CD40-activated B cells. Cell Commun. Signal. 2014;12:75. doi: 10.1186/s12964-014-0075-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Miller P.S., Aricescu A.R. Crystal structure of a human GABAA receptor. Nat. Cell Biol. 2014;512:270–275. doi: 10.1038/nature13293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Alvarez L.D., Pecci A., Estrin D.A. In Search of GABAA Receptor’s Neurosteroid Binding Sites. J. Med. Chem. 2018;62:5250–5260. doi: 10.1021/acs.jmedchem.8b01400. [DOI] [PubMed] [Google Scholar]
- 177.Barragan A., Weidner J.M., Jin Z., Korpi E.R., Birnir B. GABAergic signalling in the immune system. Acta Physiol. 2015;213:819–827. doi: 10.1111/apha.12467. [DOI] [PubMed] [Google Scholar]
- 178.Rudy B., McBain C.J. Kv3 channels: Voltage-gated K+ channels designed for high-frequency repetitive firing. Trends Neurosci. 2001;24:517–526. doi: 10.1016/S0166-2236(00)01892-0. [DOI] [PubMed] [Google Scholar]
- 179.Bezine M., Maatoug S., Ben Khalifa R., Debbabi M., Zarrouk A., Wang Y., Griffiths W.J., Nury T., Samadi M., Vejux A., et al. Modulation of Kv3.1b potassium channel level and intracellular potassium concentration in 158N murine oligodendrocytes and BV-2 murine microglial cells treated with 7-ketocholesterol, 24S-hydroxycholesterol or tetracosanoic acid (C24:0) Biochimie. 2018;153:56–69. doi: 10.1016/j.biochi.2018.02.008. [DOI] [PubMed] [Google Scholar]
- 180.Grissmer S., Ghanshani S., Dethlefs B., McPherson J.D., Wasmuth J.J., Gutman G.A., Cahalan M.D., Chandy K.G. The Shaw-related potassium channel gene, Kv3.1, on human chromosome 11, encodes the type l K+ channel in T cells. J. Biol. Chem. 1992;267:20971–20979. doi: 10.1016/S0021-9258(19)36784-5. [DOI] [PubMed] [Google Scholar]
- 181.Black J.A., Waxman S.G. Noncanonical Roles of Voltage-Gated Sodium Channels. Neuron. 2013;80:280–291. doi: 10.1016/j.neuron.2013.09.012. [DOI] [PubMed] [Google Scholar]
- 182.Roselli F., Livrea P., Jirillo E. Voltage-Gated Sodium Channel Blockers as Immunomodulators. Recent Pat. CNS Drug Discov. 2006;1:83–91. doi: 10.2174/157488906775245255. [DOI] [PubMed] [Google Scholar]
- 183.Bezine M., Namsi A., Sghaier R., Ben Khalifa R., Hamdouni H., Brahmi F., Badreddine I., Mihoubi W., Nury T., Vejux A., et al. The effect of oxysterols on nerve impulses. Biochimie. 2018;153:46–51. doi: 10.1016/j.biochi.2018.04.013. [DOI] [PubMed] [Google Scholar]
- 184.Pristerà A., Baker M.D., Okuse K. Association between Tetrodotoxin Resistant Channels and Lipid Rafts Regulates Sensory Neuron Excitability. PLoS ONE. 2012;7:e40079. doi: 10.1371/journal.pone.0040079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Tang L., Wang Y., Leng T., Sun H., Zhou Y., Zhu W., Qiu P., Zhang J., Lu B., Yan M., et al. Cholesterol metabolite cholestane-3β,5α,6β-triol suppresses epileptic seizures by negative modulation of voltage-gated sodium channels. Steroids. 2015;98:166–172. doi: 10.1016/j.steroids.2014.12.025. [DOI] [PubMed] [Google Scholar]
- 186.Tang L., Yan M., Leng T., Yin W., Cai S., Duan S., Zhu W., Lin S., Huang J., Yan G., et al. Cholestane-3β, 5α, 6β-triol suppresses neuronal hyperexcitability via binding to voltage-gated sodium channels. Biochem. Biophys. Res. Commun. 2018;496:95–100. doi: 10.1016/j.bbrc.2018.01.004. [DOI] [PubMed] [Google Scholar]
- 187.Mandler R.N., Seamer L.C., Whitlinger D., Lennon M., Rosenberg E., Bankhurst A.D. Human natural killer cells express Na+ channels. A pharmacologic flow cytometric study. J. Immunol. 1990;144:2365–2370. [PubMed] [Google Scholar]
- 188.Lai Z.-F., Chen Y.-Z., Nishimura Y., Nishi K. An amiloride-sensitive and voltage-dependent Na+ channel in an HLA-DR-restricted human T cell clone. J. Immunol. 2000;165:83–90. doi: 10.4049/jimmunol.165.1.83. [DOI] [PubMed] [Google Scholar]
- 189.Huang W., Lu C., Wu Y., Ouyang S., Chen Y. Identification and functional characterization of voltage-gated sodium channels in lymphocytes. Biochem. Biophys. Res. Commun. 2015;458:294–299. doi: 10.1016/j.bbrc.2015.01.103. [DOI] [PubMed] [Google Scholar]
- 190.Milam A.A.V., Bartleson J.M., Donermeyer D.L., Horvath S., Durai V., Raju S., Yu H., Redmann V., Zinselmeyer B., White J.M., et al. Tuning T Cell Signaling Sensitivity Alters the Behavior of CD4+ T Cells during an Immune Response. J. Immunol. 2018;200:3429–3437. doi: 10.4049/jimmunol.1701422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Zsiros E., Kis-Toth K., Hajdu P., Gaspar R., Bielanska J., Felipe A., Rajnavolgyi E., Panyi G. Developmental Switch of the Expression of Ion Channels in Human Dendritic Cells. J. Immunol. 2009;183:4483–4492. doi: 10.4049/jimmunol.0803003. [DOI] [PubMed] [Google Scholar]
- 192.Black J.A., Waxman S.G. Sodium channels and microglial function. Exp. Neurol. 2011;234:302–315. doi: 10.1016/j.expneurol.2011.09.030. [DOI] [PubMed] [Google Scholar]
- 193.Carrithers M.D., Dib-Hajj S., Carrithers L.M., Tokmoulina G., Pypaert M., Jonas E.A., Waxman S.G. Expression of the Voltage-Gated Sodium Channel NaV1.5 in the Macrophage Late Endosome Regulates Endosomal Acidification. J. Immunol. 2007;178:7822–7832. doi: 10.4049/jimmunol.178.12.7822. [DOI] [PubMed] [Google Scholar]
- 194.Carrithers L.M., Hulseberg P., Sandor M., Carrithers M.D. The human macrophage sodium channel NaV1.5 regulates mycobacteria processing through organelle polarization and localized calcium oscillations. FEMS Immunol. Med. Microbiol. 2011;63:319–327. doi: 10.1111/j.1574-695X.2011.00853.x. [DOI] [PubMed] [Google Scholar]
- 195.Jones A., Kainz D., Khan F., Lee C., Carrithers M.D. Human Macrophage SCN5A Activates an Innate Immune Signaling Pathway for Antiviral Host Defense. J. Biol. Chem. 2014;289:35326–35340. doi: 10.1074/jbc.M114.611962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.White C.R., Dungan M., Carrithers M.D. Activation of human macrophage sodium channels regulates RNA processing to increase expression of the DNA repair protein PPP1R10. Immunobiology. 2018;224:80–93. doi: 10.1016/j.imbio.2018.10.005. [DOI] [PubMed] [Google Scholar]
- 197.Sun H., Jiang J., Gong L., Li X., Yang Y., Luo Y., Guo Z., Lu R., Li H., Li J., et al. Voltage-gated sodium channel inhibitor reduces atherosclerosis by modulating monocyte/macrophage subsets and suppressing macrophage proliferation. Biomed. Pharm. 2019;120:109352. doi: 10.1016/j.biopha.2019.109352. [DOI] [PubMed] [Google Scholar]
- 198.Gonzalez-Perez V., Lingle C.J. Regulation of BK Channels by Beta and Gamma Subunits. Annu. Rev. Physiol. 2019;81:113–137. doi: 10.1146/annurev-physiol-022516-034038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Tian Y., Heinemann S.H., Hoshi T. Large-conductance Ca2+- and voltage-gated K+channels form and break interactions with membrane lipids during each gating cycle. Proc. Natl. Acad. Sci. USA. 2019;116:8591–8596. doi: 10.1073/pnas.1901381116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Dopico A.M., Bukiya A.N. Regulation of Ca2+-Sensitive K+ Channels by Cholesterol and Bile Acids via Distinct Channel Subunits and Sites. Curr. Top. Membr. 2017;80:53–93. doi: 10.1016/bs.ctm.2017.07.001. [DOI] [PubMed] [Google Scholar]
- 201.Tajima N., Xiaoyan L., Taniguchi M., Kato N. 24S-hydroxycholesterol alters activity of large-conductance Ca2+-dependent K+ (slo1 BK) channel through intercalation into plasma membrane. Mol. Cell Biol. Lipids. 2019;1864:1525–1535. doi: 10.1016/j.bbalip.2019.05.010. [DOI] [PubMed] [Google Scholar]
- 202.Son Y., Chun W., Ahn Y.-T., Kim K., Lee C.-W., Kim J.-M., Lee C., An W.G. 7-Ketocholesterol induces the reduction of KCNMB1 in atherosclerotic blood vessels. Biochem. Biophys. Res. Commun. 2015;457:324–327. doi: 10.1016/j.bbrc.2014.12.109. [DOI] [PubMed] [Google Scholar]
- 203.Erdogan A., Schaefer M.B., Kuhlmann C.R.W., Most A., Hartmann M., Mayer K., Renner F.C., Schaefer C., Abdallah Y., Hoelschermann H., et al. Activation of Ca2+-activated potassium channels is involved in lysophosphatidylcholine-induced monocyte adhesion to endothelial cells. Atherosclerosis. 2007;190:100–105. doi: 10.1016/j.atherosclerosis.2006.02.035. [DOI] [PubMed] [Google Scholar]
- 204.Papavlassopoulos M., Stamme C., Thon L., Adam D., Hillemann D., Seydel U., Schromm A.B. MaxiK Blockade Selectively Inhibits the Lipopolysaccharide-Induced IκB-α/NF-κB Signaling Pathway in Macrophages. J. Immunol. 2006;177:4086–4093. doi: 10.4049/jimmunol.177.6.4086. [DOI] [PubMed] [Google Scholar]
- 205.Sun X., Xu M., Cao Q., Huang P., Zhu X., Dong X.-P. A lysosomal K+ channel regulates large particle phagocytosis by facilitating lysosome Ca2+ release. Sci. Rep. 2020;10:1038. doi: 10.1038/s41598-020-57874-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Selezneva A., Yoshida M., Gibb A., Willis D. Nuclear BK channels regulate CREB phosphorylation in RAW264.7 macrophages. Pharm. Rep. 2021;73:881–890. doi: 10.1007/s43440-021-00229-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Chen X., Sooch G., Demaree I., White F., Obukhov A. Transient Receptor Potential Canonical (TRPC) Channels: Then and Now. Cells. 2020;9:1983. doi: 10.3390/cells9091983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Berthier A., Lemaire-Ewing S., Prunet C., Monier S., Athias A., Bessède G., De Barros J.-P.P., Laubriet A., Gambert P., Lizard G., et al. Involvement of a calcium-dependent dephosphorylation of BAD associated with the localization of Trpc-1 within lipid rafts in 7-ketocholesterol-induced THP-1 cell apoptosis. Cell Death Differ. 2004;11:897–905. doi: 10.1038/sj.cdd.4401434. [DOI] [PubMed] [Google Scholar]
- 209.Ingueneau C., Huynh-Do U., Marcheix B., Athias A., Gambert P., Nègre-Salvayre A., Salvayre R., Vindis C. TRPC1 is regulated by caveolin-1 and is involved in oxidized LDL-induced apoptosis of vascular smooth muscle cells. J. Cell. Mol. Med. 2008;13:1620–1631. doi: 10.1111/j.1582-4934.2008.00593.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Wenning A.S., Neblung K., Strauß B., Wolfs M.-J., Sappok A., Hoth M., Schwarz E.C. TRP expression pattern and the functional importance of TRPC3 in primary human T-cells. Mol. Cell Res. 2011;1813:412–423. doi: 10.1016/j.bbamcr.2010.12.022. [DOI] [PubMed] [Google Scholar]
- 211.Lindemann O., Strodthoff C., Horstmann M., Nielsen N., Jung F., Schimmelpfennig S., Heitzmann M., Schwab A. TRPC1 regulates fMLP-stimulated migration and chemotaxis of neutrophil granulocytes. Mol. Cell Res. 2015;1853:2122–2130. doi: 10.1016/j.bbamcr.2014.12.037. [DOI] [PubMed] [Google Scholar]
- 212.Chauhan A., Sun Y., Sukumaran P., Zangbede F.O.Q., Jondle C.N., Sharma A., Evans D.L., Chauhan P., Szlabick R.E., Aaland M.O., et al. M1 Macrophage Polarization Is Dependent on TRPC1-Mediated Calcium Entry. iScience. 2018;8:85–102. doi: 10.1016/j.isci.2018.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Da Conceicao V.N., Sun Y., Zboril E.K., De la Chapa J.J., Singh B.B. Loss of Ca2+ entry via Orai-TRPC1 induces ER stress that initiates immune activation in macrophage cells. J. Cell Sci. 2019;133 doi: 10.1242/jcs.237610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Sun Y., Chauhan A., Sukumaran P., Sharma J., Singh B.B., Mishra B.B. Inhibition of store-operated calcium entry in microglia by helminth factors: Implications for immune suppression in neurocysticercosis. J. Neuroinflamm. 2014;11:210. doi: 10.1186/s12974-014-0210-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.