

Abstract—
Prevention and management of myocardial ischemia/reperfusion (I/R) injury is a key step in coronary heart disease surgery. Luteolin is a falconoid compound that has an antioxidant effect, but its mechanism in I/R injury in vivo and in vitro is still under explored. This study attempted to reveal the role of luteolin (Lut) in I/R through mediation of the Src homology 2 domain-containing protein tyrosine phosphatase 1 (SHP-1)/Signal transducer and activator of transcription 3 (STAT3) pathway. To establish I/R rat models, the left anterior descending artery (LAD) was ligated for 30 min and re-perfused for 1 h in Lut-pretreated or nude rats. Comparisons between infarct area, cardiac dysfunction, and myocardial cell death and inflammatory reaction were performed in I/R-induced rats. Hypoxia/reoxygenation (H/R) cell models were established by stimulating H9c2 cells with 95% nitrogen and 5% carbon dioxide. Simultaneously, H/R-related cell death and inflammatory reactions were investigated following Lut treatment. The target protein of Lut was identified using western blotting. Pro-inflammatory cytokines were also measured in serum or Lut-pretreated cell culture medium. The results revealed that compared with the I/R group, Lut treatment could significantly decrease myocardial infarction (MI) area, increase left ventricular ejection fraction (LVEF), and decrease cell death and pro-inflammatory cytokines in the serum. Decreased apoptosis and inflammatory cytokines were also observed in H/R cells after Lut treatment. Lut treatment downregulated SHP-1 expression and subsequently upregulated STAT3 phosphorylation in both I/R rat heart tissue and H9c2 cells. The findings of the current study suggest that Lut can protect the heart and reduce MI area, cell apoptosis rate, and inflammatory level in I/R models.
KEY WORDS: Myocardial Ischemia;, Ischemia/Reperfusion Injury;, Hypoxia/Reoxygenation Injury;, Luteolin;, SHP-1;, STAT3.
BACKGROUND
Ischemic heart disease (IHD), a main contributor to mortality in patients with cardiovascular problems, is responsible for up to 42% of all deaths [1]. Ischemia/reperfusion injury (I/R) prevention and management are leading clinical problems associated with IHD treatment [2] as crucial factors in cardiovascular surgeries [3]. The mechanisms underlying I/R are complex and may be related to metabolic energy, free radicals, calcium overload, neutrophil infiltration, and dysfunctional vascular endothelial cell (VEC) [4], finally leading to dead or necrotic cells.
Inflammation in the myocardium is believed to be an important marker for myocardial I/R. Reduction of inflammatory reactions was found to be beneficial in the recovery of patients with I/R-induced myocardial infarction (MI) in previous research [5]. Increasing evidence shows that cell death is critical in myocardial I/R [6]. Protecting myocardial cells from I/R-induced apoptosis could principally promote the survival of myocardial cells, thereby providing current and future benefits to MI cells. Hence, myocardial inflammatory reactions and cell death may represent underlying targets for treating MI and I/R.
Luteolin (Lut), a sickle-like complex present in some medical herbs, has anti-inflammatory [7, 8], anti-oxidant [9], and anti-cancer [10–12] properties. Recently, Lut was used pharmacologically in clinical trials to prevent ischemic heart disease and I/R [13]. A recent study has also demonstrated that Lut at different dose had an inhibitory efficiency on the cardiomyocyte apoptosis in the myocardial I/R-tread rat through TLR4/NF-kappaB/NLRP3 pathway [5, 14]. However, identifying the molecular targets utilized to prevent and treat I/R requires additional research. In addition, Lut greatly increased left ventricular pseudoaneurysm and global and regional coronary flow reserve in Langendorff-perfused rabbits with recurrent MI [15].
Src homology region 2 domain-containing phosphatase-1 (SHP-1) contains an SH region 2 domain and is the non-transmembrane protein tyrosine phosphatase (PTP) primarily produced in hematopoietic progenitor cells (HPC) [16]. SHP-1, a negative-regulator of signaling transduction through receptor phosphorylation, acts as a crucial factor in cell death and/or the repression of survival factors [17]. In mouse models featuring a locally ischemic brain, SHP-1 protein was increased, reaching a peak at 7 days after cerebral ischemia and regulating the multiplication of nerve cells in the ischemic central nervous system (CNS) [18]. Notably, SHP-1 can repress the Janus kinase (JAK)/Signal transducer and activator of transcription (STAT) pathway through the dephosphorylation and inactivation of STAT3 [19], which is widely considered to be the signaling pathway of cell apoptosis and inflammatory reactions [20]. SHP-1-induced STAT3 activation was related to Y-box binding protein 1 (YB1) knockdown in inhibiting hypoxia/reoxygenation (H/R)-mediated myocardial cell death. In I/R mouse models, inactivating SHP-1-induced STAT3 was also highly correlated with improving acute myocardial infarction, reduction of MI area, myocardial cell death, and oxidative stress response.
Based on the abovementioned reports, Lut is indicated as exerting cardioprotective effects on I/R rats by promoting the inactivation of SHP-1-induced STAT3. Using an in vivo I/R model with left anterior descending artery (LAD) ligation and an in vitro model with H/R-induced H9c2 damage, we investigated the protective functions of Lut in myocardial I/R injury. Lut exerted a myocardial protective role in I/R rats and H/R-treated cells, through mediating SHP-1/STAT3 signal axis. The gain of function experiments with SHP-1 overexpression showed that SHP-1 was involved in the anti-inflammation and anti-apoptosis effect of Lut.
METHODS
Materials
In this research, Lut (pure: 99%, Sigma-Aldrich, stored at concentration of 20 mM) and ELISA kits (Nanjing KeyGEN Biotech, China) were used for detecting IL-1β, IL-18, and TNF-α.
Animals
Adult SD rats (male, 220–250 g) provided by Vitalriver (Beijing, China) were fed in animal houses using a standard configuration requirement for 5 days before this study. All rats could freely drink water and eat standard food. Every procedure was approved by the Animal Care and Use Committee of the First Affiliated Hospital of Zhengzhou University.
Drug Treatment in in I/R Models
Rats were randomly assigned to three groups (n = 10): sham group, I/R group, and I/R + Lut group. Seven-day intragastric administration was performed in the Lut group using Lut (qd, per 40 mg/kg) [21] after operation. Details on the surgery performed are mentioned above [22]. The injury models were established by 24-h myocardial reperfusion following 30-min treatment. After establishing the models, all rats were killed, blood specimens were obtained for biochemical tests, and cardiac tissue specimens were obtained for studying the morphology, biochemistry, and individual molecules.
Tissues and Serum Preparations
Blood specimens (500 μl) taken retro-orbitally were subjected to centrifugation at 1500 g for 10 min at 4 °C. Serum specimens after centrifugation were stored in Eppendorf tubes at − 80 °C for downstream analysis [23]. One part from the heart tissue samples (around 1 mm3) underwent cell fixation with 2.5% glutaraldehyde for ultrastructural examination; another part underwent cell fixation with 4% paraformaldehyde and was embedded in paraffin; the remainder underwent tissue wrapping with AF and was deeply frozen in LIN.
Detection of MI Area
Twenty-four hours after I/R, all rats were killed. The heart was evenly cut into 5 pieces below the ligation and along a line parallel to coronary sulcus after being kept at 20 °C for 15 min. Total sections were weighed again and cultured for 15 min with 2% 2,3,5-triphenyltetrazolium chloride (TTC) (Sigma, USA) at 37 °C in darkness for pathologic analysis. The MI proportion of total cardiac tissue, including non-infarct size (red, staining part) and infarct size (white/pale, non-stained part), was calculated using the IPP image software (Version 4.1, Media Cybernetics, USA).
Heart Function Test
The size and function of the heart were evaluated by M-mode echocardiography. Left ventricular end-diastolic diameter (LVEDD) and left ventricular end-diastolic and end-systolic diameter (LVESD) were obtained from the results of a left parasternal long-axis section. The mean of 5 continuous cardiac cycles is expressed in our results. Left ventricular end systolic volume (LVESV), left ventricular end-diastolic volume (LVEDV), and left ventricular ejection fraction (LVEF) were obtained using an algorithm. All analysis was blinded.
Cell Culture and H/R Injury Model Establishment
Mouse H9c2 cardiomyocytes were provided by the ATCC (Manassas, USA). H9c2 cells were stored in 10% (v/v) FBS high glucose DMEM (Gibco/Invitrogen), with 100 U/ml penicillin and 100 μg/ml streptomycin (Gibco/Invitrogen) under humid conditions including 5% carbon dioxide at 37 °C.
After H9c2 cells were cultured until 70–80% confluence, 20 mM Lut for the treatment group was solubilized in medium containing DMSO and 5% FBS to reach the final concentration of 30 μM, Lut in the control and H/R groups underwent the same operation in 5% FBS medium for 24 h. Afterward, the above operations in all groups were performed in Krebs–Ringer bicarbonate buffer; pH 7.4). After that, the cells were incubated in oxygen-deficient conditions including 95% nitrogen and 5% carbon dioxide for 6 h, then cultured in DMEM containing 10% FBS and kept in an environment containing 5% CO2 and 95% air for 5 h to induce reoxygenation damage [24].
The full-length ORF of SHP-1 was cloned into pcDNA3.1( +) to construct SHP-1 express vectors (pcDNA-SHP-1). Transfection of pcDNA-SHP-1/pcDNA-Empty was performed using Lipofectamine 2000 Reagent (Invitrogen, USA).
ELISA
The IL-1β, IL-18, and TNF-α content in serum and supernatant was measured using ELISA kits in accordance with manufacturer instructions. The absorbance at 450 nm was measured using a microplate spectrophotometer. In the end, sample content was calculated using a standard curve.
WB
Proteins were separated from evened heart tissues and cells lysed using Trizol reagent (Invitrogen, Carlsbad, Calif) in accordance with a standard Invitrogen protocol. After quantitative analysis of protein content using the improved Bradford assay (Bio-Rad Laboratories, Calif), proteins were utilized downstream for WB with primary and secondary antibodies. Spots were observed by a chemiluminescence system (Amersham Bioscience, UK). Signal quantification was performed using Image Pro Plus software (Media Cybernetics).
Q-PCR
Total RNA was separated from homogenous cardiac tissues and lysed cells using TRIzol reagent (Invitrogen, USA), then evaluated using a Nanodrop2000 (OD260). The MMLV First-Strand Kit (Invitrogen, USA) and Oligo (dT) 20 primer were utilized in reverse transcription to prepare cDNA. Q-PCR was conducted on cDNA prepared by SYBR Select Master Mix (Invitrogen, USA). mRNA was detected by Q-PCR using the corresponding kits. All operations were carried out according to the manufacturer’s instruction. PCR conditions included an initial 10-min denaturation at 95 °C s, then 40-cycles of 15-s denaturation at 95 °C and 40 s of extension at 60 °C. GAPDH mRNA was used as an internal reference. Target mRNA levels were calculated using the 2–ΔΔCT method. All tests were conducted in triplicate.
Cell Death Assay
Cell death was assessed using FC and Annexin V-FITC/PI apoptosis detection kits (BD Pharmingen™). Preparation of cell suspensions was performed with 20 µL BB, and then treatment with 10 µL Annexin V-FITC and 5 µL PI. Cell mortality rate was assessed using flow cytometry (FCMs).
Statistical Analyses
All data were analyzed USING SPSS 10.0. Data are represented as M ± SD. One-way ANOVA was applied in multi-group comparisons (significant, P < 0.05); Scheffe’s test was performed between individual groups (significant, P < 0.05).
RESULTS
Effect of Lut Treatment on MI Area and Left Ventricular Failure in I/R Rat Models
To indicate the efficiency of Lut on the I/R-induced MI and left ventricular failure in I/R rats, TTC staining and electrocardiograph were used. First, myocardial TTC staining results showed that I/R injury was closely related with the progression of I/R-induced irreversible myocardial damage. TTC staining showed no infarct existed in the sham group, and the incidence of MI area in the model group was obviously larger than in the sham group. Meanwhile, MI area expansion was significantly abrogated by Lut treatment (40 mg/kg) (Fig. 1).
Fig. 1.
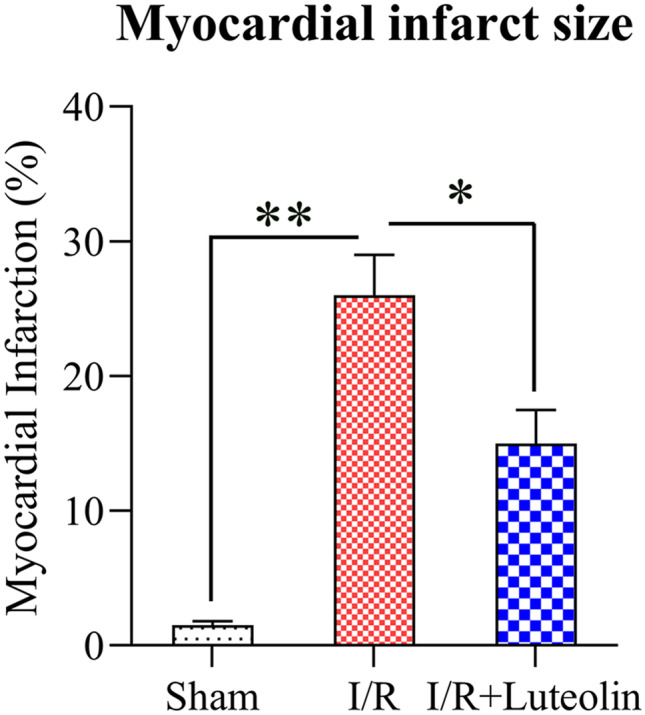
Cardiac MI area in I/R. Myocardial necrosis was evaluated by TTC staining. A Sham group, B model group, C Lut group; bar chart represents the ratio of cardiac MI area. N = 3. Data are expressed as M ± SD. Compared with indicated group, the differences were significant (p < 0.05; p < 0.01).
The electrocardiograph monitoring of hemodynamic parameters such as LVEF, LVESV, and LVEDV was performed. LVEF was evidently reduced (Fig. 2A), whereas LVESV and LVEDV were greatly increased compared to the sham group (Fig. 2B, C), indicating that the left ventricular failure in the I/R group was inhibited. However, compared to the I/R group, LVEF in the Lut group was obviously higher, while LVESV and LVEDV were obviously lower, indicating that Lut could repress left ventricular failure in I/R rats.
Fig. 2.

Lut enhances left ventricular function. A Lut treatment significantly increased LVEF, compared to the I/R group. B, C Lut treatment significantly repressed the increase in LVESV and LVEDV, compared to I/R group. LVEF left ventricular ejection fraction; LVESV left ventricular end-systolic volume; LVEDV left ventricular end-diastolic volume. N = 3. Data are expressed as M ± SD. Compared with indicated group, the difference was significant (p < 0.05).
Effect of Lut on Inflammatory Response Factors and Cell Death in I/R Rats and H/R-Mediated H9c2 Cells
Inflammatory cytokines are reported to be an important factor in I/R [25]. To verify the role of Lut on the generation of inflammatory cytokines in MI rats, serum levels of IL-1β, IL-18, and TNF-α were measured. ELISA results showed that I/R treatment could significantly increase the expression of IL-1β, IL-18, and TNF-α, while Lut administration could obviously reduce serum content of these cytokines (Fig. 3A). The level of inflammatory cytokines in the supernatant of H9c2 cells affected by H/R was measured. H/R stimulation could increase the levels of IL-1β, IL-18, and TNF-α, while Lut treatment (30 μM) [21] could obviously decrease the expression of inflammatory cytokines (Fig. 3B). The above results indicate that regulation of inflammatory cytokines may be beneficial in the protective effects of Lut on the heart.
Fig. 3.

Effects of Lut on inflammation and apoptosis in I/R-induced rats and H/R-induced cells. A Effects of Lut on serum IL-1β, IL-18 and TNF-α (ELISA assay). N = 3. B Effects of Lut on IL-1β, IL-18 and TNF-α in the supernatant of H/R treated H9c2 cells. N = 3. C Caspase-3 level and caspase-3 cleavage in heart tissue of rats (WB assay). N = 3. D Caspase-3 level and caspase-3 cleavage in H/R-induced treated H9c2 cells (WB assay). N = 3. E Cell mortality of H/R treated H9c2 cells with/without Lut treatment (FCM assay). N = 3. Data are expressed as mean ± SD. Compared with indicated group, the differences were significant (p < 0.05, p < 0.01; p < 0.001).
The mechanism of myocardial I/R is very complex, but it eventually contributes to myocardial apoptosis [26]. To assess the role of H/R-mediated apoptosis in H9c2 cells, WB and FCM assays were conducted. The cleavage level of caspase-3 in the I/R group was obviously increased compared to sham, while Lut administration greatly inhibited the expression of cleaved caspase-3 (Fig. 3C). Meanwhile, we also observed a similar phenotype for caspase-3 cleavage in H/R-induced H9c2 cells (Fig. 3D). FCM data revealed that the apoptotic cell proportion of H9c2 cells was significantly increased after H/R treatment, while Lut treatment repressed the apoptotic level of H9c2 cells (Fig. 3E).
Effects of Lut on Activation of the SHP-1/STAT3 Pathway
As previous observations indicated that the SHP-1/STAT3 pathway was highly relevant to apoptosis and inflammation [27], we therefore tried to investigate whether the SHP-1/STAT3 pathway is mediated by Lut treatment and acts as an important factor in promoting cell inflammation and viability. We measured the levels of SHP-1 and STAT3, and level of phosphorylated STAT3, in I/R-mediated heart tissue and H/R-mediated H9c2 cells. In I/R-treated heart tissue, we observed that SHP-1 was notably increased and phosphorylated STAT3 was decreased in the I/R treatment group, while Lut treatment caused a decrease in SHP-1 level and increase of STAT3 phosphorylation (Fig. 4A, B). In addition, a similar phenomenon was observed in H/R-induced H9c2 cells. Finally, STAT3 phosphorylation was decreased by H/R induction and increased after Lut administration (Fig. 4C, D).
Fig. 4.

Effects of Lut on SHP-1/STAT3 pathway activation in I/R-induced rats and H/R-induced cells. A Q-PCR detected SHP-1 mRNA level in the heart tissue of rats. N = 3. B WB analysis of SHP-1 and STAT3 levels, and STAT3 phosphorylation, in the heart tissue of rats. N = 3. C Q-PCR detected SHP-1 mRNA expression in H/R-induced H9c2 cells. N = 3. D WB analysis of SHP-1 and STAT3 levels, and STAT3 phosphorylation, in H/R-induced H9c2 cells. N = 3. Values are presented as M ± SD. Compared with indicated group, the differences were significant (p < 0.05; p < 0.01).
Effects of SHP-1 on Lut-Mediated Inflammatory Reactions and Mortality of H/R-Mediated H9c2 Cells
To assess the role of SHP-1 in Lut-ameliorated inflammation and apoptosis in H/R-treated cells, SHP-1 was overexpressed in Lut-treated H/R-mediated H9c2 cells, while empty vector was also transfected as control group. The results of Q-PCR and WB showed that transfection with pcDNA-SHP-1 could increase the intracellular content of SHP-1, compared to H/R-induced H9c2 cells (Fig. 5A, B). Meanwhile, phosphorylation of STAT3 was significantly impaired after SHP-1 overexpression as assessed by WB (Fig. 5B).
Fig. 5.

Overexpression of SHP-1 in Lut-treated H/R treated cells. H9c2 cell transfection was performed with pcDNA-SHP-1/pcDNA-Empty, followed by Lut and H/R treatment. A Q-PCR detected SHP-1 mRNA content in H/R-induced H9c2 cells. N = 3. B WB analysis of SHP-1 and STAT3 levels, and STAT3 phosphorylation, in H/R treated H9c2 cells. N = 3. Results are presented as M ± SD. Compared with indicated group, the difference was significant (p < 0.05).
ELISA was then performed to detect the secretion of inflammatory cytokines from Lut-treated H/R-induced H9c2 cells with or without SHP-1 overexpression. Our findings indicated that SHP-1 overexpression reduced the production of secreted inflammatory cytokines in the cell supernatant (Fig. 6A), suggesting that SHP-1 exerts a positive role on H/R-induced inflammation.
Fig. 6.
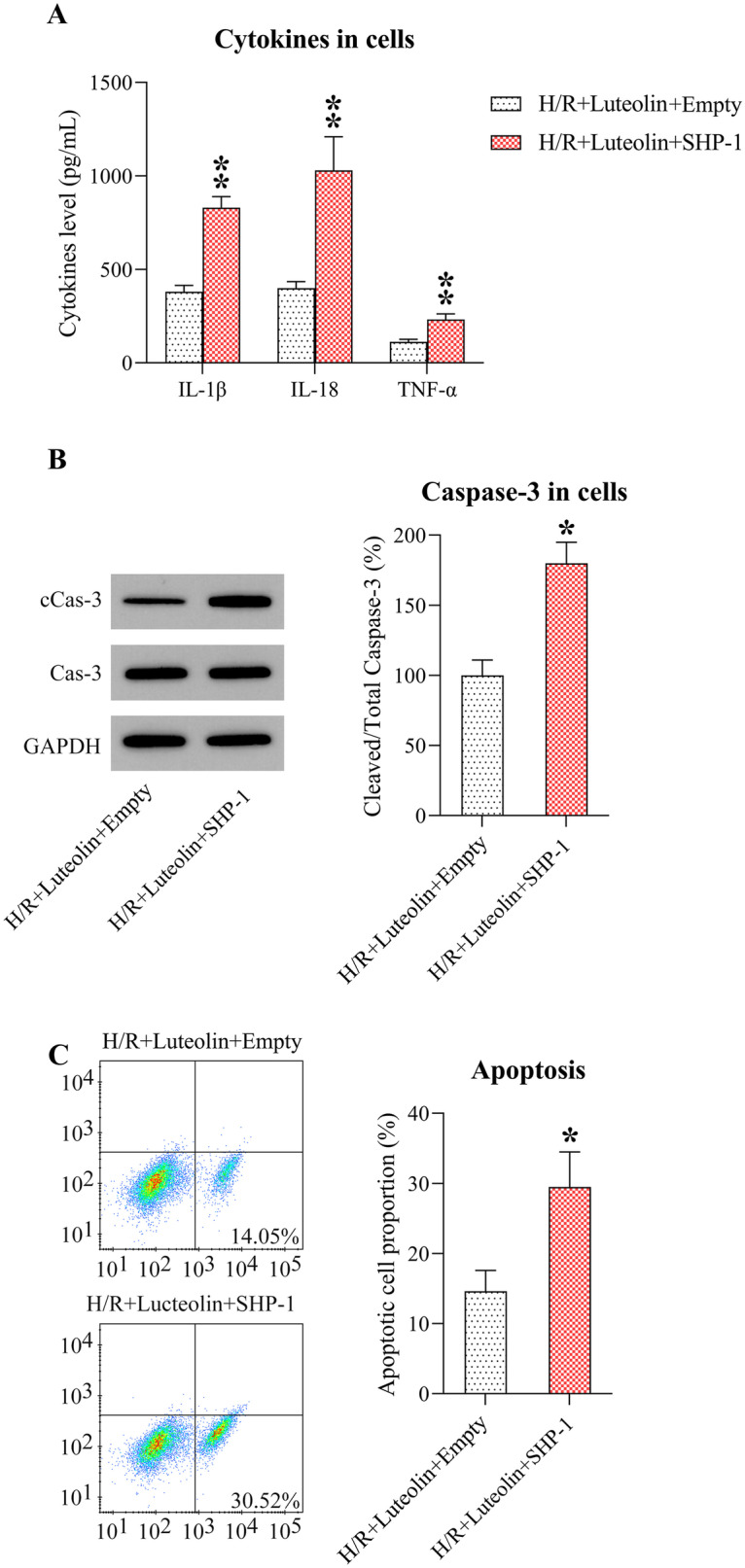
Effect of SHP-1 on inflammation and cell death in Lut-induced rats or H/R-induced cells. A Effects of Lut on IL-1β, IL-18, and TNF-α in supernatant fluid from H/R treated H9c2 cells. N = 3. B WB detection of caspase-3 level and caspase-3 cleavage in H/R-induced H9c2 cells. N = 3. C Cell mortality of H/R-induced H9c2 cells with/without Lut treatment (FCM assay). N = 3. Results are presented as M ± SD. Compared with the indicated group, the differences were significant (p < 0.05; p < 0.01).
WB then showed that cleavage of caspase-3 was obviously improved in the SHP-1 overexpression group, comparing to Lut-treated H/R-induced H9c2 cells without overexpression (Fig. 6B). FCM data further indicated an upregulation in apoptotic cell proportion after SHP-1 overexpression, indicating that SHP-1 is a key modulator of H/R-induced apoptosis (Fig. 6C).
DISCUSSION
Lut, a naturally occurring polyphenol flavonoid, has demonstrated to exert myocardial protection effects. Due to the newly emerged COVID-19 threat, Lut has been found as a dietary molecular to inhibit the SARS-COV-2 [28]: corticosteroid dexamethasone can reduce mortality of severe COVID-19 patient through limiting the production and damaging effect of the cytokines and inhibiting the protective function of T cells and block B cells from making antibodies [29]. Dexamethasone could be given together with the Lut due to its antiviral and anti-inflammatory properties, especially its ability to inhibit Mast cell activation syndrome, which are the main source of cytokines in the lungs of COVID-19 patients [29, 30]. The in vitro study in the present study demonstrated that myocardial I/R injury activated myocardial SHP-1/STAT3 signal transduction pathways and induced inflammatory reactions and apoptotic death, but Lut treatment decreased inflammatory cytokines and apoptotic cell proportion and affected the SHP-1/STAT3 pathway. In vivo experiments confirmed that Lut administration ameliorated the I/R-induced MI lesion and left ventricular failure through mediating SHP-1/STAT3 pathway. In brief, the protection provided by Lut in I/R injury may be due to inhibiting inflammatory reactions and cell apoptosis by modulating the SHP-1/STAT3 pathway.
A major indicator of I/R is the large production of inflammatory cytokines. Ischemic damage is generated by temporary insufficiency of blood supply in the tissues, but reperfusion damage is caused by rapid inflammation after blood reperfusion [31]. The production of inflammatory cytokines is an important process for the progression of inflammatory reactions in the heart [32]. For instance, TNF-α had a negative inotropic effect on myocardium contraction and blood pressure control. TNF-α could also expand the MI area during I/R injury and is involved in adverse ventricular remodeling [33]. IL-18 also participated in the pathophysiological process of I/R-associated inflammatory diseases by abrogating myocyte necrosis and immunity reactions [34]. IL-1β is an important and early inflammatory mediator in I/R injury [35]. Our results confirmed that IL-1β, IL-18, TNF-α, and other inflammatory cytokines were obviously increased in I/R models and H/R-treated H9c2 cells. However, significant inhibition of the cardiac inflammatory response in vivo and in vitro was found after Lut treatment, due to the inhibition of these three inflammatory cytokines in I/R-mediated rats and H/R-mediated H9c2 cells.
Cardiomyocyte apoptosis is another important representation in heart failure during MI. Inhibition of cell death represses damage in contractile cells, reduces I/R injury-induced heart injuries, and thus attenuates the incidence of heart failure [36, 37]. A recent study has demonstrated that Lut at different dose had an inhibitory efficiency on the cardiomyocyte apoptosis in the myocardial I/R-tread rat [14]. The present study indicates an elevation in I/R-induced cell death in the I/R treated rats, which was evidenced by the cleavage of caspase-3. In vitro results also showed that H/R obviously promoted the apoptotic death of H9c2 cells. Apoptosis in both in vivo and in vitro model was inhibited in the Lut treatment.
The non-receptor PTPs, SHP-1 (aka PTPN6), and SHP-2 (aka PTPN11) play crucial regulatory roles in basic cellular processes, including cell proliferation, cell death, cell migration, cytodifferentiation, inflammatory reactions, and intermediate metabolisms [38]. The expression of SHP-1 in hematopoietic and epithelial cells is extensively considered to play a role in the negative regulation of inflammatory reactions [39]. SHP-1 is also closely related to cell death and can bind with tumor necrosis factor receptor 1, Fas receptor, and other death receptors to block anti-apoptosis signals in neutrophils. Therefore, SHP-1 is believed to act as a crucial factor in cell death and/or the blocking of survival-related factors. STAT3 is an oncogenic transcription factor that could play a significant role through the phosphorylation dimerization of tyrosine residues and translocation to the nucleus [40, 41]. Constitutive activation of STAT3 in various malignancies is closely related to disease progress [42, 43]. Phosphorylated STAT3 was involved in inflammatory reactions and cell death in rat models with Cd-induced testis toxicity [44]. Various evidences have shown that SHP-1 protein level is upregulated in rats with cerebral ischemia and MI, resulting in the increase of cerebral ischemia and MI [45]. The mechanism regarding how SHP-1 regulated STAT3 phosphorylation is well-documented in many previous studies [35, 46–51]; however, the influence and mechanism of SHP-1/STAT3 axis in MI was not elucidated. As far as we know, this is the first attempt to assess the role of SHP-1/STAT3 axis in MI. In this study, our findings showed that the expression of SHP-1 in cardiac tissue and cells was upregulated in rats after I/R and H9c2 cells with H/R treatment, with deactivated STAT3 sensor. Lut treatment significantly reduced SHP-1/STAT3 signal transduction by inhibiting the expression of SHP-1. SHP-1 overexpression restored both inflammation and apoptosis which were downregulated accompanying SHP-1 upregulation in Lut-treated and H/R-induced H9c2 cells. These data suggest that the SHP-1/STAT3 axis is highly associated with Lut-mediated inflammation and apoptotic death in an H/R cell model, as well as I/R rats.
CONCLUSIONS
In conclusion, this research indicates that Lut provides myocardial protection during I/R by improving myocardial contraction function and inhibiting inflammatory reactions and cell death. Meanwhile, activating the SHP-1/STAT3 signaling pathway is related to the protective effects of Lut on the MI-injured heart tissue.
AUTHOR CONTRIBUTION
Study design/planning: Donghai Liu, Hong Luo, Chenhui Qiao. Data collection/entry: Donghai Liu, Hong Luo, Chenhui Qiao. Data analysis/statistics: Donghai Liu, Hong Luo, Chenhui Qiao. Data interpretation: Donghai. Liu, Chenhui Qiao. Preparation of manuscript: Chenhui Qiao. Literature analysis/search: Donghai Liu, Hong. Luo, Chenhui Qiao.
DECLARATIONS
Ethics Approval
Every procedure was approved by the Animal Care and Use Committee of the First Affiliated Hospital of Zhengzhou University.
Informed Consent
Informed consent was obtained from all individual participants included in the study.
Conflict of Interest
The authors declare no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Donghai Liu, Email: ldy10101@163.com.
Hong Luo, Email: superluohong@tom.com.
Chenhui Qiao, Email: qchenhui@yeah.net.
References
- 1.Mendis, Shanthi, Pekka, Puska, Bo, Norrving, and World Health Organization. 2011. Global atlas on cardiovascular disease prevention and control. World Health Organization.
- 2.Rodriguez-Porcel Martin, Zhu Xiang-Yang, Chade Alejandro R, Amores-Arriaga Beatriz, Caplice Noel M, Ritman Erik L, Lerman Amir, Lerman Lilach O. Functional and structural remodeling of the myocardial microvasculature in early experimental hypertension. American Journal of Physiology-Heart and Circulatory Physiology. 2006;290(3):H978–H984. doi: 10.1152/ajpheart.00538.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Monassier, Jean Pierre. 2008. Reperfusion injury in acute myocardial infarction. From bench to cath lab. Part I: Basic considerations. Archives of cardiovascular diseases 101 (7–8): 491–500. [DOI] [PubMed]
- 4.Turer Aslan T, Hill Joseph A. Pathogenesis of myocardial ischemia-reperfusion injury and rationale for therapy. The American journal of cardiology. 2010;106(3):360–368. doi: 10.1016/j.amjcard.2010.03.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhang Xu, Qianming Du, Yang Yan, Wang Jianing, Dou Shuai, Liu Chao, Duan Junguo. The protective effect of Luteolin on myocardial ischemia/reperfusion (I/R) injury through TLR4/NF-κB/NLRP3 inflammasome pathway. Biomedicine & Pharmacotherapy. 2017;91:1042–1052. doi: 10.1016/j.biopha.2017.05.033. [DOI] [PubMed] [Google Scholar]
- 6.Lee Young Soo, Kang Young Jin, Kim Hye Jung, Park Min Kyu, Seo Han Geuk, Lee Jae Heun, Yun-Choi Hye Sook, Chang Ki Churl. Higenamine reduces apoptotic cell death by induction of heme oxygenase-1 in rat myocardial ischemia-reperfusion injury. Apoptosis. 2006;11(7):1091–1100. doi: 10.1007/s10495-006-7110-y. [DOI] [PubMed] [Google Scholar]
- 7.Palombo Ramona, Savini Isabella, Luciana Avigliano S, Madonna Andrea Cavani, Albanesi C, Alessandro Mauriello G, Melino, and A Terrinoni. Luteolin-7-glucoside inhibits IL-22/STAT3 pathway, reducing proliferation, acanthosis, and inflammation in keratinocytes and in mouse psoriatic model. Cell death & disease. 2016;7(8):e2344–e2344. doi: 10.1038/cddis.2016.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Taliou Anilia, Zintzaras Elias, Lykouras Lefteris, Francis Kostantinos. An open-label pilot study of a formulation containing the anti-inflammatory flavonoid luteolin and its effects on behavior in children with autism spectrum disorders. Clinical therapeutics. 2013;35(5):592–602. doi: 10.1016/j.clinthera.2013.04.006. [DOI] [PubMed] [Google Scholar]
- 9.Manju VAIYAPURI, Balasubramaniyan VAIRAPPAN, Nalini NAMASIVAYAM. Rat colonic lipid peroxidation and antioxidant status: The effects of dietary luteolin on 1, 2-dimethylhydrazine challenge. Cellular and Molecular Biology Letters. 2005;10(3):535. [PubMed] [Google Scholar]
- 10.Stefano De, Alessandro Sabrina Caporali, Di Daniele Nicola, Rovella Valentina, Cardillo Carmine, Schinzari Francesca, Minieri Marilena, Pieri Massimo, Candi Eleonora, Bernardini Sergio. Anti-Inflammatory and Proliferative Properties of Luteolin-7-O-Glucoside. International journal of molecular sciences. 2021;22(3):1321. doi: 10.3390/ijms22031321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lin Yong, Shi Ranxin, Wang Xia, Shen Han-Ming. Luteolin, a flavonoid with potential for cancer prevention and therapy. Current cancer drug targets. 2008;8(7):634–646. doi: 10.2174/156800908786241050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Palombo Ramona, Caporali Sabrina, Falconi Mattia, Iacovelli Federico, Rocca Blasco Morozzo Della, Surdo Alessandro Lo, Campione Elena, Candi Eleonora, Melino Gerry, Bernardini Sergio. Luteolin-7-O-β-d-Glucoside Inhibits Cellular Energy Production Interacting with HEK2 in Keratinocytes. International journal of molecular sciences. 2019;20(11):2689. doi: 10.3390/ijms20112689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Qi Lingling, Pan Huanjun, Li Dongye, Fang Fang, Chen Dan, Sun Hong. Luteolin improves contractile function and attenuates apoptosis following ischemia–reperfusion in adult rat cardiomyocytes. European journal of pharmacology. 2011;668(1–2):201–207. doi: 10.1016/j.ejphar.2011.06.020. [DOI] [PubMed] [Google Scholar]
- 14.Li, J.P., R. Yan, P.L. Ma, P. Fu, H.T. Tian, and L.L. Wang. 2020. Effects of luteolin in different doses on the cardiomyocyte apoptosis in rats with myocardial ischemia reperfusion. Journal of Biological Regulators and Homeostatic Agents 34 (6). [DOI] [PubMed]
- 15.Rump AFE, Schüssler M, Acar D, Cordes A, Theisohn M, Rösen R, Klaus W, Fricke U. Functional and antiischemic effects of luteolin-7-glucoside in isolated rabbit hearts. General Pharmacology: The Vascular System. 1994;25(6):1137–1142. doi: 10.1016/0306-3623(94)90129-5. [DOI] [PubMed] [Google Scholar]
- 16.Wlodarski, Pawel, Qian, Zhang, Xiaobin, Liu, Monika, Kasprzycka, Michal, Marzec, and Mariusz A. Wasik. 2007. PU. 1 activates transcription of SHP-1 gene in hematopoietic cells. Journal of Biological Chemistry 282 (9): 6316–6323. [DOI] [PubMed]
- 17.Daigle Isabelle, Yousefi Shida, Colonna Marco, Green Douglas R, Simon Hans-Uwe. Death receptors bind SHP-1 and block cytokine-induced anti-apoptotic signaling in neutrophils. Nature medicine. 2002;8(1):61–67. doi: 10.1038/nm0102-61. [DOI] [PubMed] [Google Scholar]
- 18.Wishcamper Celine A, Brooks Diane M, Douglas Coffin J, Lurie Diana I. Focal cerebral ischemia upregulates SHP-1 in reactive astrocytes in juvenile mice. Brain research. 2003;974(1–2):88–98. doi: 10.1016/S0006-8993(03)02564-2. [DOI] [PubMed] [Google Scholar]
- 19.Han Yajun, Amin Hesham M, Franko Bevin, Frantz Christine, Shi Xinzhe, Lai Raymond. Loss of SHP1 enhances JAK3/STAT3 signaling and decreases proteosome degradation of JAK3 and NPM-ALK in ALK+ anaplastic large-cell lymphoma. Blood. 2006;108(8):2796–2803. doi: 10.1182/blood-2006-04-017434. [DOI] [PubMed] [Google Scholar]
- 20.Chen Kuen-Feng, Jung-Chen Su, Liu Chun-Yu, Huang Jui-Wen, Chen Kuei-Chiu, Chen Wei-Lin, Tai Wei-Tien, Shiau Chung-Wai. A novel obatoclax derivative, SC-2001, induces apoptosis in hepatocellular carcinoma cells through SHP-1-dependent STAT3 inactivation. Cancer letters. 2012;321(1):27–35. doi: 10.1016/j.canlet.2012.03.023. [DOI] [PubMed] [Google Scholar]
- 21.Yu Dongsheng, Li Mengwen, Tian Youqing, Liu Jun, Shang Jing. Luteolin inhibits ROS-activated MAPK pathway in myocardial ischemia/reperfusion injury. Life Sciences. 2015;122:15–25. doi: 10.1016/j.lfs.2014.11.014. [DOI] [PubMed] [Google Scholar]
- 22.Bhindi Ravinay, Witting Paul K, McMahon Aisling C, Khachigian Levon M, Lowe Harry C. Rat models of myocardial infarction. Thrombosis and haemostasis. 2006;96(11):602–610. doi: 10.1160/TH05-07-0514. [DOI] [PubMed] [Google Scholar]
- 23.Wu, Hua-Li, Si-Lin, Pang, Qiong-Zhen, Liu, Qian, Wang, Min-Xuan, Cai, and Jing Shang. 2014. 5-HT1A/1B receptors as targets for optimizing pigmentary responses in C57BL/6 mouse skin to stress. PloS one 9 (2): e89663. [DOI] [PMC free article] [PubMed]
- 24.Chiu Po Yee, Luk Ka Fai, Leung Hoi Yan, Ng Ka Ming, Ko Kam Ming. Schisandrin B stereoisomers protect against hypoxia/reoxygenation-induced apoptosis and inhibit associated changes in Ca2+-induced mitochondrial permeability transition and mitochondrial membrane potential in H9c2 cardiomyocytes. Life sciences. 2008;82(21–22):1092–1101. doi: 10.1016/j.lfs.2008.03.006. [DOI] [PubMed] [Google Scholar]
- 25.Sivarajah Ahila, Collino Massimo, Yasin Mohammed, Benetti Elisa, Gallicchio Margherita, Mazzon Emanuela, Cuzzocrea Salvatore, Fantozzi Roberto, Thiemermann Christoph. Anti-apoptotic and anti-inflammatory effects of hydrogen sulfide in a rat model of regional myocardial I/R. Shock. 2009;31(3):267–274. doi: 10.1097/SHK.0b013e318180ff89. [DOI] [PubMed] [Google Scholar]
- 26.Zhao Zhi-Qing, Nakamura Masanori, Wang Ning-Ping, Wilcox Josiah N, Shearer Steven, Ronson Russell S, Guyton Robert A, Vinten-Johansen Jakob. Reperfusion induces myocardial apoptotic cell death. Cardiovascular research. 2000;45(3):651–660. doi: 10.1016/S0008-6363(99)00354-5. [DOI] [PubMed] [Google Scholar]
- 27.Hodge David R, Hurt Elaine M, Farrar William L. The role of IL-6 and STAT3 in inflammation and cancer. European journal of cancer. 2005;41(16):2502–2512. doi: 10.1016/j.ejca.2005.08.016. [DOI] [PubMed] [Google Scholar]
- 28.Shawan Mohammad Mahfuz, Khan Ali, Halder Sajal Kumar, Hasan Md Ashraful. Luteolin and abyssinone II as potential inhibitors of SARS-CoV-2: An in silico molecular modeling approach in battling the COVID-19 outbreak. Bulletin of the National Research Centre. 2021;45(1):1–21. doi: 10.1186/s42269-020-00479-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Theoharides, TC, and P Conti. 2020. Dexamethasone for COVID-19? Not so fast. J Biol Regul Homeost Agents 34 (3): 10.23812. [DOI] [PubMed]
- 30.Theoharides, TC, and P Conti. 2020. COVID-19 and multisystem inflammatory syndrome, or is it mast cell activation syndrome? Journal of Biological Regulators and Homeostatic Agents 34 (5). [DOI] [PubMed]
- 31.Kaisho Tsuneyasu, Akira Shizuo. Toll-like receptor function and signaling. Journal of Allergy and Clinical Immunology. 2006;117(5):979–987. doi: 10.1016/j.jaci.2006.02.023. [DOI] [PubMed] [Google Scholar]
- 32.He He, Chang Xiayun, Gao Jin, Zhu Lingpeng, Miao Mingxing, Yan Tianhua. Salidroside mitigates sepsis-induced myocarditis in rats by regulating IGF-1/PI3K/Akt/GSK-3β signaling. Inflammation. 2015;38(6):2178–2184. doi: 10.1007/s10753-015-0200-7. [DOI] [PubMed] [Google Scholar]
- 33.Tsai, Hsin-Ju, Shiang-Suo Huang, Meng-Ting Tsou, Hsiao-Ting Wang, and Jen-Hwey Chiu. 2015. Role of opioid receptors signaling in remote electrostimulation-induced protection against ischemia/reperfusion injury in rat hearts. PloS one 10 (10): e0138108. [DOI] [PMC free article] [PubMed]
- 34.Gu Haifeng, Xie Minghua, Liqian Xu, Zheng Xiaojun, Yang Yunmei, Lv Xueying. The protective role of interleukin-18 binding protein in a murine model of cardiac ischemia/reperfusion injury. Transplant International. 2015;28(12):1436–1444. doi: 10.1111/tri.12683. [DOI] [PubMed] [Google Scholar]
- 35.Li BL, Zhao DY, Du PL, Wang XT, Yang Q, Cai YR. Luteolin alleviates ulcerative colitis through SHP-1/STAT3 pathway. Inflammation Research. 2021 doi: 10.1007/s00011-021-01468-9. [DOI] [PubMed] [Google Scholar]
- 36.Fu Jiajia, Huang Heqing, Liu Jiajun, Pi Rongbiao, Chen Jianwen, Liu Peiqing. Tanshinone IIA protects cardiac myocytes against oxidative stress-triggered damage and apoptosis. European journal of pharmacology. 2007;568(1–3):213–221. doi: 10.1016/j.ejphar.2007.04.031. [DOI] [PubMed] [Google Scholar]
- 37.Song Jun-Qiu, Teng Xu, Cai Yan, Tang Chao-Shu, Qi Yong-Fen. Activation of Akt/GSK-3β signaling pathway is involved in intermedin 1–53 protection against myocardial apoptosis induced by ischemia/reperfusion. Apoptosis. 2009;14(11):1299. doi: 10.1007/s10495-009-0398-7. [DOI] [PubMed] [Google Scholar]
- 38.Chong ZZ, Maiese Kenneth. The Src homology 2 domain tyrosine phosphatases SHP-1 and SHP-2: Diversified control of cell growth, inflammation, and injury. Histology and histopathology. 2007;22(11):1251. doi: 10.14670/hh-22.1251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Fish Jason E, Santoro Massimo M, Morton Sarah U, Sangho Yu, Yeh Ru-Fang, Wythe Joshua D, Ivey Kathryn N, Bruneau Benoit G, Stainier Didier YR, Srivastava Deepak. miR-126 regulates angiogenic signaling and vascular integrity. Developmental cell. 2008;15(2):272–284. doi: 10.1016/j.devcel.2008.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lim Cheh Peng, Cao Xinmin. Structure, function, and regulation of STAT proteins. Molecular biosystems. 2006;2(11):536–550. doi: 10.1039/b606246f. [DOI] [PubMed] [Google Scholar]
- 41.Schindler Christian, Levy David E, Decker Thomas. JAK-STAT signaling: From interferons to cytokines. Journal of Biological Chemistry. 2007;282(28):20059–20063. doi: 10.1074/jbc.R700016200. [DOI] [PubMed] [Google Scholar]
- 42.Santoni Matteo, Massari Francesco, Del Re Marzia, Ciccarese Chiara, Piva Francesco, Principato Giovanni, Montironi Rodolfo, Santini Daniele, Danesi Romano, Tortora Giampaolo. Investigational therapies targeting signal transducer and activator of transcription 3 for the treatment of cancer. Expert opinion on investigational drugs. 2015;24(6):809–824. doi: 10.1517/13543784.2015.1020370. [DOI] [PubMed] [Google Scholar]
- 43.Subramaniam, Aruljothi, Muthu K. Shanmugam, Ekambaram, Perumal, Feng Li, Alamelu, Nachiyappan, Xiaoyun, Dai, Shivananju Nanjunda Swamy, Kwang Seok Ahn, Alan Prem Kumar, and Benny KH Tan. 2013. Potential role of signal transducer and activator of transcription (STAT) 3 signaling pathway in inflammation, survival, proliferation and invasion of hepatocellular carcinoma. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer 1835 (1): 46–60. [DOI] [PubMed]
- 44.Andreadou Ioanna, Efentakis Panagiotis, Balafas Evangelos, Togliatto Gabriele, Davos Constantinos H, Varela Aimilia, Dimitriou Constantinos A, Nikolaou Panagiota-Efstathia, Maratou Eirini, Lambadiari Vaia. Empagliflozin limits myocardial infarction in vivo and cell death in vitro: Role of STAT3, mitochondria, and redox aspects. Frontiers in Physiology. 2017;8:1077. doi: 10.3389/fphys.2017.01077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sugano Masahiro, Tsuchida Keiko, Hata Tomoji, Makino Naoki. RNA interference targeting SHP-1 attenuates myocardial infarction in rats. The FASEB journal. 2005;19(14):2054–2056. doi: 10.1096/fj.05-4020fje. [DOI] [PubMed] [Google Scholar]
- 46.Bian Y, Yuan L, Yang X, Weng L, Zhang Y, Bai H, Chen J. SMURF1-mediated ubiquitylation of SHP-1 promotes cell proliferation and invasion of endometrial stromal cells in endometriosis. Ann Transl Med. 2021;9(5):362. doi: 10.21037/atm-20-2897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ding X, Lu D, Fan J. A natural product phillygenin suppresses osteosarcoma growth and metastasis by regulating the SHP-1/JAK2/STAT3 signaling. Bioscience, Biotechnology, and Biochemistry. 2021;85(2):307–314. doi: 10.1093/bbb/zbaa007. [DOI] [PubMed] [Google Scholar]
- 48.Han Y, Zhang J, Pang Y, Wang Y, Zhang X, Zhang H. The role of Src homology region 2 domain-containing phosphatase-1 hypermethylation in the classification of patients with myelodysplastic syndromes and its association with signal transducer and activator of transcription 3 phosphorylation in skm-1 cells. Journal of International Medical Research. 2021;49(3):300060521999550. doi: 10.1177/0300060521999550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Teng R, Wang Y, Lv N, Zhang D, Williamson RA, Lei L, Chen P, et al. Corrigendum to "Hypoxia Impairs NK Cell Cytotoxicity through SHP-1-Mediated Attenuation of STAT3 and ERK Signaling Pathways". Journal of Immunology Research. 2021;2021:3625084. doi: 10.1155/2021/3625084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Teng R, Wang Y, Lv N, Zhang D, Williamson RA, Lei L, Chen P, et al. Hypoxia Impairs NK Cell Cytotoxicity through SHP-1-Mediated Attenuation of STAT3 and ERK Signaling Pathways. Journal of Immunology Research. 2020;2020:4598476. doi: 10.1155/2020/4598476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yang, M.H., I.J. Ha, J.Y. Um, and K.S. Ahn. 2021. Albendazole Exhibits Anti-Neoplastic Actions against Gastric Cancer Cells by Affecting STAT3 and STAT5 Activation by Pleiotropic Mechanism(s). Biomedicines 9 (4). 10.3390/biomedicines9040362. [DOI] [PMC free article] [PubMed]