

ABSTRACT
Cation and anion channelrhodopsins (CCRs and ACRs, respectively) primarily from two algal species, Chlamydomonas reinhardtii and Guillardia theta, have become widely used as optogenetic tools to control cell membrane potential with light. We mined algal and other protist polynucleotide sequencing projects and metagenomic samples to identify 75 channelrhodopsin homologs from four channelrhodopsin families, including one revealed in dinoflagellates in this study. We carried out electrophysiological analysis of 33 natural channelrhodopsin variants from different phylogenetic lineages and 10 metagenomic homologs in search of sequence determinants of ion selectivity, photocurrent desensitization, and spectral tuning in channelrhodopsins. Our results show that association of a reduced number of glutamates near the conductance path with anion selectivity depends on a wider protein context, because prasinophyte homologs with a glutamate pattern identical to that in cryptophyte ACRs are cation selective. Desensitization is also broadly context dependent, as in one branch of stramenopile ACRs and their metagenomic homologs, its extent roughly correlates with phylogenetic relationship of their sequences. Regarding spectral tuning, we identified two prasinophyte CCRs with red-shifted spectra to 585 nm. They exhibit a third residue pattern in their retinal-binding pockets distinctly different from those of the only two types of red-shifted channelrhodopsins known (i.e., the CCR Chrimson and RubyACRs). In cryptophyte ACRs we identified three specific residue positions in the retinal-binding pocket that define the wavelength of their spectral maxima. Lastly, we found that dinoflagellate rhodopsins with a TCP motif in the third transmembrane helix and a metagenomic homolog exhibit channel activity.
KEYWORDS: algae, channelrhodopsins, optogenetics, photosensory reception
INTRODUCTION
Channelrhodopsins (ChRs) are light-gated ion channels initially discovered in chlorophyte algae, in which they serve as photoreceptors guiding phototactic orientation (1–3). Subsequently, ChRs have also been found in the genomes/transcriptomes of cryptophyte and haptophyte algae (4, 5), the heterotrophic protists known as Labyrinthulea (5), and giant viruses that infect marine microorganisms (6, 7). Ongoing polynucleotide sequencing projects provide a rich hunting ground for further exploration of ChR diversity and taxonomic distribution.
Functionally, ChRs are divided into cation and anion ChRs (CCRs and ACRs, respectively) (8). Both ChR classes serve for photocontrol of excitable cells, such as neurons and cardiomyocytes, via a biotechnique known as optogenetics (9, 10). However, structural determinants for cation and anion selectivity in ChRs remain poorly understood. X-ray crystal structures (11–15) indicate that the ion conductance path in algal ChRs is formed by transmembrane helices 1, 2, 3 and 7 (TM1, TM2, TM3, and TM7). All so-far-known ACRs contain a noncarboxylate residue in the position of the protonated Schiff base counterion in bacteriorhodopsin (Asp85), whereas in nearly all CCRs, the carboxylate is conserved. However, this sequence feature cannot be regarded as a sole indicator of anion selectivity, because some chlorophyte CCRs also show a noncarboxylate residue in the counterion position (e.g., DsChR1 from Dunaliella salina [16]).
Most chlorophyte CCRs contain five Glu residues in TM2 and the TM2-TM3 loop (Glu82, Glu83, Glu90, Glu97, and Glu101 in ChR2 from Chlamydomonas reinhardtii [CrChR2]), whereas in all so-far-known ACRs, most or even all of the corresponding positions are occupied with noncarboxylate residues. Therefore, it has been proposed that negative electrostatic potential of the channel pore defines cation selectivity (17, 18). Indeed, mutagenetic remodeling of the pore to reduce electronegativity yielded permeability for anions in chlorophyte CCRs (17–21). However, some of the TM2 glutamates are conserved in ACRs and apparently do not interfere with their anion conductance.
Other biophysical properties of ChRs relevant for optogenetic applications are their desensitization under continuous or pulsed illumination (also called “inactivation” in the literature) and spectral sensitivity. In an earlier study, a group of ACRs discovered in the TARA marine transcriptomes demonstrated particularly rapid and strong desensitization (22). As their source organisms were not known, these proteins were named MerMAIDs (metagenomically discovered, marine, anion-conducting and intensely desensitizing channelrhodopsins). However, strong desensitization cannot serve as a characteristic of a single ChR family, because it was also observed in some bacteriorhodopsin-like CCRs (BCCRs) from cryptophytes that show very little sequence homology with MerMAIDs (23).
To gain more insight into the taxonomic distribution and structure-function relationships of ChRs, we identified 75 ChR homologs from several phylogenetic lineages and metagenomic samples and tested 27 of them along with 16 previously reported sequences by heterologous expression in cultured mammalian cells followed by patch clamp recording. We show that the same pattern of conserved Glu residues may accompany cation or anion conductance in ChRs from different taxa and that the degree of desensitization in MerMAID homologs is greater, the closer their sequences are to those of the first reported MerMAIDs. We report two prasinophyte CCRs with red-shifted spectra and confirm that three specific residues in the retinal-binding pocket are responsible for wavelength regulation in cryptophyte ACRs. Finally, we demonstrate that some dinoflagellate rhodopsins possess channel activity.
RESULTS
Prasinophyte CCRs.
Only a few of the >150 chlorophyte ChRs identified so far (Fig. 1; Data Sets S1 and S2) have been tested by heterologous expression. Both C. reinhardtii ChRs conduct cations (2, 3), so other chlorophyte ChRs were also assumed to be CCRs. However, a recent study demonstrated that two ChRs from the prasinophyte genus Pyramimonas in fact conduct anions (6), which called for a more detailed functional analysis of chlorophyte ChRs.
FIG 1.

Unrooted phylogenetic tree of ChRs. The nodes are color coded as follows: red, confirmed anion selectivity; blue, confirmed cation selectivity; gray, nonfunctional; black, ion selectivity not determined. Thicker nodes show ChRs characterized in this study. Gray circles show ultrafast bootstrap support values above 95%. A tree file in the Newick format is available as Data Set S2, and the corresponding protein alignment is presented in Data Set S3.
Newick file of the tree shown in Fig. 1. Download Data Set S2, TXT file, 0.02 MB (21KB, txt) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
An alignment of the C-truncated sequences used to construct the tree in Fig. 1. Download Data Set S3, TXT file, 0.6 MB (679.7KB, txt) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Three ChR homologs derived from the prasinophytes Crustomastix stigmatica, Mantoniella squamata, and Pyramimonas melkonianii (6) exhibit a residue pattern typical of cryptophyte ACRs; i.e., they display conserved Glu82 and Glu90 with noncarboxylate residues in the positions of Glu83, Glu97, Glu101, and Glu123 of CrChR2 (Fig. 2A). In a Cymbomonas tetramitiformis sequence (6), Glu90 and Glu97 are conserved, whereas Glu82 is replaced with Gln (Fig. 2A). We synthesized mammalian codon-adapted versions of these rhodopsin domains, fused them with C-terminal enhanced yellow fluorescent protein (EYFP), expressed them in HEK293 (human embryonic kidney) cells, and analyzed them by manual whole-cell patch clamp.
FIG 2.

Prasinophyte CCRs. (A) Amino acid residues corresponding to the indicated positions in CrChR2. ChRs characterized in this study are in bold (black, functional; gray, nonfunctional). Conserved glutamates are highlighted in red. (B) Peak photocurrent amplitudes generated at −60 mV in response to 1-s light pulses at the wavelength of the spectral maximum. (C) Desensitization of photocurrents after 1-s illumination. (D) Action spectra of photocurrents. The data points show means and SEM (n = 4 to 8 scans). (E and I) Reversal potentials of photocurrents. The bars in panels B, C, and E to I show means ± SEM; diamonds show data from individual cells.
Three of these ChRs generated photocurrents (Fig. 2B and C) in our standard buffer system (for solution compositions, see Table S1), whereas the homolog from P. melkonianii that we named PymeChR was nonelectrogenic. The action spectra of photocurrents generated by the M. squamata and C. stigmatica homologs were red shifted (the rhodopsin maxima at ∼580 and 585 nm, respectively) (Fig. 2D). Their retinal-binding pockets are nearly identical but differ from those of previously known red-shifted ChRs (Fig. S1A). Both spectra exhibited a second band at ∼520 nm that reflected a Förster resonance energy transfer (FRET) from EYFP to rhodopsin, as was earlier shown in RubyACRs from Labyrinthulea (5). The efficiency of FRET was even greater in the homolog from C. stigmatica, the rhodopsin peak of which was observed in the green spectral region and could not be accurately resolved because of the FRET contribution (Fig. 2D, olive line). To determine the peak position more accurately, we replaced EYFP with mCherry (absorption maximum, 587 nm). The action spectrum of photocurrents generated by the mCherry fusion is shown in Fig. 2B (magenta). As expected, the 520-nm band of FRET from EYFP disappeared, revealing the rhodopsin peak at ∼540 nm.
(A) Residues of the retinal-binding pockets. Variants tested in this study are in bold. The residues in MsCCR and CsCCR that differ from those in Chrimson and RubyACRs, earlier known red-shifted ChRs, are highlighted red. The numbers are according to bacteriorhodopsin sequence. (B and D) The residues in the positions of the conserved glutamates (highlighted in red) in the ion conductance pathway in the indicated homologs from Chlorophyceae (B) and Chlorodendrophyceae (D). Variants tested in this study are in bold; nonfunctional variants are in gray. The numbers are according to the CrChR2 sequence. (C) Photocurrent traces recorded from C1ChR in response to 1-s illumination at −60 and 60 mV. (E) Alignment of the part of TM7 of the indicated Chlorodendrophyceae homologs. The Schiff base Lys is highlighted in blue; the upstream Glu, in red. Abbreviations: MsCCR, Mantoniella squamata cation channelrhodopsin; CsCCR, Crustomastix stigmatica cation channelrhodopsin; CrChR2, Chlamydomonas reinhardtii channelrhodopsin 2; PymeACR, Pyramimonas melkonianii anion channelrhodopsin; CnChR3, Chlamydomonas noctigama channelrhodopsin 3; CnChR4, Chlamydomonas noctigama channelrhodopsin 4; C1ChR, Chlamydomonas sp. channelrhodopsin; CsChR2, Chloromonas subdivisa channelrhodopsin 2; PsChR4, Platymonas (Tetraselmis) subcordiformis channelrhodopsin 4; TaChR, Tetraselmis astigmatica channelrhodopsin; TchChR, Tetraselmis chui channelrhodopsin; TsChR2, Tetraselmis striata channelrhodopsin 2. Download FIG S1, TIF file, 0.4 MB (386.5KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Solution compositions in AxoPatch and SyncroPatch recordings. Abbreviations: Asp, aspartate; LJP, liquid junction potential; NMDG, N-methyl-d-glucamine. All concentrations are in millimolar units. Download Table S1, DOCX file, 0.02 MB (16.4KB, docx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
To test the relative permeability of the prasinophyte homologs for H+, Na+, K+, Ca2+, and Cl−, we varied the concentration of each of these ions in the bath (for solution compositions, see Table S1), measured the current-voltage relationships, and determined the reversal potentials (Erev). CrChR2 was included in this experiment for comparison. Figure 2E to I show that under all tested conditions, Erev for all three homologs was close to the equilibrium potential of H+, indicating that they are H+-selective channels with negligible permeability for metal cations and Cl−. We named them CtCCR, MsCCR, and CsCCR. A less positive Erev of MsCCR photocurrents probed under the H+ gradient does not result from the permeability for Na+ as it does in CrChR2 and most likely reflects a contribution of intramolecular charge transfers, as previously found in other CCRs (24).
“Core” chlorophyte and streptophyte ChR homologs.
Four sequences from Chlorophyceae have only the Glu82 homolog, as do prasinophyte ACRs (Fig. S1B), but show no close sequence homology to them. Upon expression of three of these polynucleotides, small hyperpolarizing photocurrents that did not reverse at positive voltages were recorded (Fig. S1C). They likely reflect intramolecular transfer of the Schiff base proton to an outwardly located acceptor, as previously found in other ChRs (24, 25). Four sequences from Chlorodendrophyceae contain no glutamate residues in any of the six analyzed positions (Fig. S1D) and form a separate branch on the phylogenetic tree (Fig. 1). Very unusually, in this sequence group the Asp residue corresponding to Asp212 of bacteriorhodopsin is located not four residues upstream, as in most known microbial rhodopsins, but three residues upstream of the Schiff base lysine (Fig. S1E). Neither any of these proteins nor a ChR homolog from the streptophyte Coleochaete generated channel currents.
Stramenopile ACRs and their metagenomic homologs.
The first MerMAIDs reported were seven homologous ACRs identified in metagenomic samples (22). Recently, close homologs were found in unclassified stramenopile species (5, 26), which suggests that the original MerMAIDs also originate from stramenopiles. We have identified 20 additional MerMAID homologs, nine haptophyte ACR homologs, and two Labyrinthulea ACR homologs (Data Set S1) in metagenomic databases (Data Set S4). We tested EYFP fusions of five metagenomic MerMAID homologs (indicated by the designation “mg” in protein names), two closely related sequences from the unclassified stramenopile strain TOSAG23-3 (indicated by “sT” [5]), and three sequences from the bicosoecid stramenopile Cafeteria roenbergensis (indicated by “Car” to distinguish them from C. reinhardtii ChRs). In most sequences of this group, both Glu82 and Glu90 (CrChR2 numbering) are conserved, as in the previously known cryptophyte ACRs and MerMAIDs (Fig. 3A). The five tested MerMAID homologs and those from TOSAG23-3 clustered together with the first reported MerMAIDs (Fig. S2), whereas Cafeteria homologs formed a separate branch related to haptophyte ACRs (Fig. 1). Each of these homologs generated photocurrents in HEK293 cells (Fig. 3B). As shown below, these ChRs conduct anions, so we designated them ACRs.
FIG 3.

Stramenopile ACRs and their metagenomic homologs. (A) Amino acid residues in the ion conductance pathway, corresponding to the indicated positions in CrChR2. ChRs characterized in this study are in bold. Conserved glutamates are highlighted in red. (B) Peak photocurrent amplitudes generated at −60 mV in response to 1-s light pulses at the wavelength of the spectral maximum. (C) Desensitization of photocurrents after 1-s illumination. (D) Reversal potentials of photocurrents. In panels B to D, the bars show means and SEM; diamonds show data from individual cells. (E and F) Action spectra of photocurrents. The data points are means ± SEM (n = 4 to 6 scans).
A section of the phylogenetic tree from Fig. 1 redrawn in a rectangular format. Red nodes show variants with proven anion selectivity, thicker nodes show variants tested in this study. Red numbers are the values of photocurrent desensitization from Fig. 3C in the main text (mean ± SEM, n = 5 or 6 cells). Abbreviations: sTACR, stramenopile strain TOSAG23-3 anion channelrhodopsin; mgACR, metagenomic anion channelrhodopsin. Download FIG S2, TIF file, 0.5 MB (540KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
GenBank accession numbers, abbreviated protein names, source organisms, habitats, transcript names, and amino acid sequences used to construct the phylogenetic tree in Fig. 1. Note that only sequences that cover the entire N-terminal and rhodopsin domains are included. Literature references to identification and electrophysiological characterization of the sequences are also provided. The sequences identified and characterized in this study are shown in bold. Download Data Set S1, XLSX file, 0.1 MB (95.1KB, xlsx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Sequence databases searched. Download Data Set S4, XLSX file, 0.01 MB (12.4KB, xlsx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Of all tested homologs, sTACR2 is the most closely related to the first reported MerMAIDs, which exhibit nearly complete desensitization (Fig. S2). Similarly, sTACR2 photocurrents showed nearly complete desensitization (Fig. 3C, cyan), whereas photocurrents from sTACR1, the most distant homolog (Fig. S2), exhibited only ∼40% desensitization (Fig. 3C, dark yellow). The values for other homologs were intermediate (Fig. 3C). All ChRs of this group demonstrated exclusively anion permeability (Fig. 3D). The action spectra of their photocurrents are shown in Fig. 3E and F. The shape of some spectra (e.g., mgACR2 and mgACR5) indicated a contribution of FRET from EYFP.
Cryptophyte ACRs.
Cryptophytes are the taxon in which the first natural ACRs were discovered (4). To explore the diversity of cryptophyte ACRs further, we analyzed transcriptomes of 20 additional cryptophyte strains (Table S2) and identified 15 transcripts homologous to previously known cryptophyte ACRs (Data Set S1). As no species names have been assigned to their source organisms, we used the numbers 3 to 8 in the abbreviated protein names to designate different Rhodomonas strains (the numbers 1 and 2 have already been assigned to the previously analyzed strains). The Glu82 homologs is conserved in all, and the Glu90 homolog in most, of these proteins, whereas all other analyzed positions are occupied by noncarboxylate residues (Fig. 4A). Thirteen homologs generated photocurrents upon expression in HEK293 cells (Fig. 4B and C).
FIG 4.

Cryptophyte ACRs. (A) Amino acid residues corresponding to the indicated positions in CrChR2. ChRs characterized in this study are in bold; nonfunctional homologs are in gray. Conserved glutamates are highlighted in red. (B) Peak photocurrent amplitudes generated at −60 mV in response to 1-s light pulses at the wavelength of the spectral maximum. (C) Desensitization of photocurrents after 1-s illumination. (D and E) Series of photocurrent traces recorded from R3ACR1 upon incremental voltage increase with the SyncroPatch 384i (D) and AxoPatch 200B (E) at 470-nm excitation. Note the smaller amplitude and slower kinetics of the SyncroPatch traces, as expected from the lower stimulus intensity. (F) Reversal potentials of photocurrents. In panels B, C, and F, the bars show means ± SEM; diamonds show data from individual cells.
List of cryptophyte strains analyzed. Abbreviations: BEA, Banco Español de Algas, of the Universidad of Las Palmas de Gran Canaria, Spain; CCAC, Culture Collection of Algae at the University of Cologne, Germany; SCCAP, Scandinavian Culture Collection of Algae and Protozoa at the University of Copenhagen, Denmark; CCMP, Culture Collection of Marine Phytoplankton at the Provasoli-Guillard National Center for Marine Algae and Microbiota at Woods Hole Oceanographic Institution, USA; NIES, National Institute for Environmental Studies, Tsukuba, Japan. Download Table S2, DOCX file, 0.02 MB (16.4KB, docx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
To verify permeability for anions in the three cryptophyte ACR homologs that were well expressed and generated photocurrents in the nanoampere range by manual patch clamp (Fig. 4B), we used a high-throughput automated patch clamp (APC) instrument, SyncroPatch 384i, with solutions provided by the manufacturer (for their full compositions, see Table S1). The internal solution was predominantly CsF to promote formation of gigaseals, and the external solution was predominantly NaCl. Representative series of photocurrent traces recorded from R3ACR1 under incremental voltage using the SyncroPatch 384i and AxoPatch 200B with the same solutions are shown in Fig. 4D and E, respectively. GtACR1 and CrChR2, well characterized by manual patch clamp, were included in the SyncroPatch experiment as ACR and CCR controls, respectively. With the SyncroPatch solutions, the Erev of GtACR1 photocurrents was negative, whereas that of CrChR2 was positive (Fig. 4F). In all tested homologs, the Erev was close to that of GtACR1 (Fig. 4F), which confirmed their anion selectivity.
Previously, we and others demonstrated that Cys133, Ser156, and Tyr207 in GtACR1 (absorption maximum, 515 nm) corresponding to Arg129, Gly152, and Phe203 in GtACR2 (absorption maximum, 470 nm) define the spectral difference between these two proteins (15, 27) (E. G. Govorunova, O. A. Sineshchekov, and J. L. Spudich, unpublished data). According to GtACR1 crystal structures, the side chains of Cys133 and Ser156 are located near the β-ionone ring of the chromophore (Fig. 5A), whereas the hydroxyl group of Tyr207 forms a hydrogen bond with Asp234 in the photoactive center. Comparative analysis of these positions (Fig. 5C) and action spectra of photocurrents (Fig. 5B and D) in the 13 functional ACR homologs has revealed that only proteins in which the residues match those of GtACR2 exhibit blue-shifted absorption maxima. When Cys or Met is found at position 133, or Ser or Ala at position 156, the spectrum is shifted to longer wavelengths.
FIG 5.
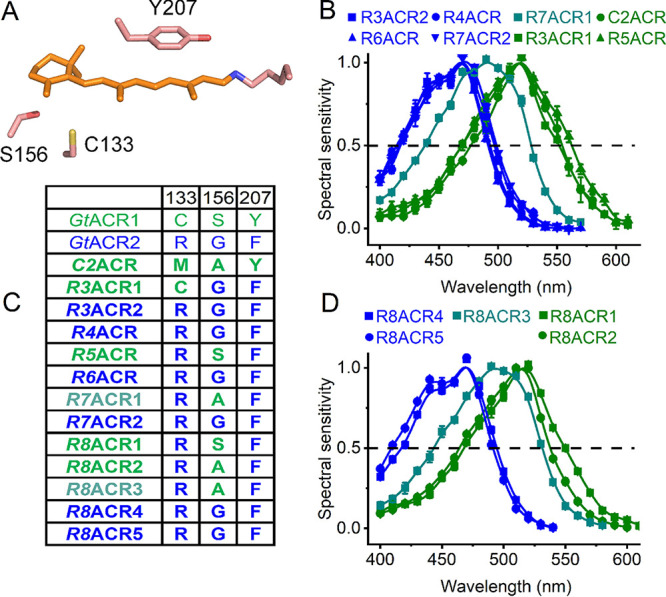
Color tuning in cryptophyte ACRs. (A) A crystal structure of GtACR1 (6EDQ) showing the three side chains that contribute to the spectral difference between GtACR1 and GtACR2. (B and D) Action spectra of photocurrents generated by the indicated cryptophyte ACRs. The data points show means ± SEM (n = 4 to 8 scans). (C) Amino acid residues involved in color tuning in the functional cryptophyte homologs. The numbering is according to the GtACR1 sequence.
Dinoflagellate ChRs and their metagenomic homologs.
Dinoflagellates exhibit genuine phototactic orientation (28–30), and their genomes encode multiple type I rhodopsins (31–33). However, to the best of our knowledge, none of these rhodopsins has so far been reported to exhibit channel function. Some rhodopsin sequences from dinoflagellates of the genera Ansanella, Pelagodinium, and Symbiodinium (the latter was recently split into several genera) (6, 34–38) contain the TCP motif in the middle of TM3 that is conserved in most so-far-known ChRs (Fig. S3A). This motif is also conserved in 17 proteins encoded by the deep-ocean TARA marine transcriptomes that cluster together with these dinoflagellate rhodopsins and form a distinct branch of the phylogenetic tree (Fig. 1). A very unusual feature of this entire sequence cluster is that Asp212 of bacteriorhodopsin, highly conserved in all ChRs known so far, is replaced with Asn or, in one homolog, Leu (Fig. S3B).
(A and B) Alignments of TM3 (A) and TM7 (B) of dinoflagellate ChRs and their metagenomic homologs. The TCP motif and the Asn residue corresponding to Asp212 of bacteriorhodopsin are highlighted in red. (C) EYFP tag fluorescence in cells expressing mgdChR1 constructs schematically shown on top of the images. TS, trafficking signal, ER, endoplasmic reticulum export motif. Abbreviations: AngChR, Ansanella granifera channelrhodopsin; PbChR, Pelagodinium bei channelrhodopsin; SA3ChR1-4, Symbiodinium clade A3 channelrhodopsins 1 to 4; SmChR1-3, Symbiodinium microadriaticum channelrhodopsins 1 to 3; SmChR1-3, Symbiodinium (Breviolum) minutum channelrhodopsins 1 to 3; mgdChR1-17, metagenomic dinoflagellate homolog channelrhodopsins 1 to 17. Download FIG S3, TIF file, 0.7 MB (765.2KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Only one of the five tested metagenomic rhodopsin domains of this group was electrogenically photoactive upon expression in HEK293 cells, producing photocurrents barely resolved from the noise level (Fig. 6A, black bar). The fusion protein formed disk-shaped fluorescent aggregates within the cells (Fig. S3C, top). The addition of the trafficking signal (TS) between rhodopsin and EYFP, and the endoplasmic reticulum export motif (ER) at the C terminus of the fusion protein (39) reduced formation of the aggregates (Fig. S3C, bottom) and significantly increased the photocurrents, although they still reached only an ∼20-pA level at best (Fig. 6A, blue bar). In our standard buffer system with nearly symmetrical ionic concentrations in the bath and pipette, the signs of the photocurrents reversed at positive voltages, indicating passive ion transport (Fig. 6B, top). We named this protein mgdChR1 (for metagenomic dinoflagellate homolog channelrhodopsin 1).
FIG 6.

ΔSmChR1 and its metagenomic homolog mgdChR1. (A) Peak photocurrent amplitudes generated at −60 mV in response to 1-s light pulses at the wavelength of the spectral maximum. The bars show means ± SEM; diamonds show data from individual cells. *, P < 0.01 by the Mann-Whitney test. “mgdChR1no” denotes the construct without TS and ER motifs. (B) Photocurrent traces recorded at −60 and 60 mV from mgdChR1 (top) and ΔSmChR1 (bottom). The ΔSmChR1 trace at 60 mV was shifted 50 ms to the right relative to the trace at −60 mV to show the fast negative peak. (C) Action spectra of photocurrents. The data points show means ± SEM (n = 10 to 12 scans).
A homologous rhodopsin domain from the coral endosymbiont Symbiodinium microadriaticum has an ∼300-residue N-terminal extension (Fig. S4), which is much longer than that found in other known ChRs, including mgdChR1. An expression construct encoding residues 1 to 600 produced no tag fluorescence. However, when the N-terminal extension was deleted and TS and ER export motifs added, fluorescence was observed, and passive photocurrents of a small amplitude, similar to that from mgdChR1, were recorded (Fig. 6B, bottom). We named this protein ΔSmChR1 to emphasize truncation of the N-terminal extension. A homologous protein, ΔSmChR2, from the same organism also generated photocurrents, but their amplitudes were even smaller. Channel currents from ΔSmChR1, but not from mgdChR1, were preceded by a fast negative current, the sign of which did not reverse at positive voltages (Fig. 6B, bottom). Such currents have previously been recorded from several other ChRs and interpreted as intramolecular charge displacement associated with isomerization of the retinal chromophore (24). The photocurrent action spectra of mgdChR1 and ΔSmChR1 peaked in the green spectral region (Fig. 6C). Small amplitudes of mgdChR1 and ΔSmChR1 photocurrents make accurate measurements of the reversal potentials problematic, so we were not able to determine their ionic selectivity.
Alignment of Symbiodinium microadriaticum ChR1 with and without the N-terminal extension. Download FIG S4, TIF file, 0.2 MB (199.9KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
DISCUSSION
We report functional testing of 43 ChR homologs from prasinophytes, stramenopiles, cryptophytes, dinoflagellates, and metagenomic samples. An unexpected result is that ACRs appear to be more widely spread among protist taxa than CCRs. Another unexpected result is that the same residue pattern comprising conserved Glu82 and Glu90 with noncarboxylate residues in the positions of Glu83, Glu97, Glu101, and Glu123 (CrChR2 numbering) found in most ACRs from stramenopiles, cryptophytes, haptophytes, and metagenomic samples is also found in the CCRs, CsCCR and MsCCR, from prasinophytes. Of five Glu residues in TM2 and the TM2-TM3 loop, Glu82 is most conserved across the entire ChR family. According to our empirical calculations using PROPKA3 (40), the pKa of the Glu82 homolog is acidic in all X-ray crystal structures of ChRs published so far, including that of GtACR1, in which it apparently does not prevent anion conductance. In CrChR2, replacement of Glu82 with Ala strongly inhibited expression in mammalian cells, as judged by the tag fluorescence and correspondingly reduced photocurrents (41), which suggests that this residue is needed for correct protein folding and/or membrane targeting.
Glu90 appears to be essential for cation conductance in CrChR2, as mutation of this residue to Lys or Arg confers permeability for anions (19). However, this Glu is conserved in most ACRs except those from prasinophytes and Labyrinthulea. In the unphotolyzed state of both CrChR2 (42) and GtACR1 (43), this residue is neutral at neutral pH. Glu90 deprotonates during the photocycle of CrChR2 (44, 45). Photoinduced protonation changes of the Glu90 homolog in GtACR1 (Glu68) have been studied by time-resolved molecular spectroscopy (25), UV-visible flash-photolysis, and electrophysiology (46), which indicate that it also deprotonates upon photoexcitation. Further research is needed to clarify the role of this residue in anion conductance.
Photocurrent desensitization in different ChR families correlates with accumulation of different intermediates of the photocycle. In MerMAID1, desensitization is correlated with the M intermediate (22), but in Rhodomonas BCCRs, it is correlated with a novel, extremely blue-shifted intermediate (23). Finally, desensitization in CrChR2 is correlated with accumulation of blue-absorbing P480 that is considered either a late intermediate in a single-branched photocycle (47) or the initial state of a parallel photocycle (45). Desensitization was reduced in the E44Q and C84T mutants of MerMAID1 (22). However, the mutated residues (corresponding, respectively, to Glu90 and Cys128 of CrChR2) are not the sole cause of strong desensitization in MerMAIDs, because they are conserved in many ChRs that do not show strong desensitization, including the closely related sTACR1 characterized here.
According to quantum mechanical/molecular mechanical calculations using the GtACR1 crystal structure, replacement of Ser156 with Gly or Ala stabilizes S0, predicting an 11- to 12-nm blue shift of the absorption maximum (48). All tested cryptophyte ACRs that contain Gly in this position exhibited blue-shifted spectra. The spectra of two ACRs that contain Ala in this position (R7ACR1 and R8ACR3) were ∼25-nm blue-shifted from that of GtACR1, whereas the spectra of the other two (C2ACR and R8ACR2) were very similar to that of GtACR1, suggesting that the expected phenotypic effect of the Ser-to-Ala substitution in these proteins was compensated for by other changed residues.
Our results and those of other groups suggest that most biophysical properties of ChRs relevant for their optogenetic applications cannot be assigned to a few individual residues but rather reflect interactions between many of them. A cumulative larger set of electrophysiological data to which our study contributes might be used in the future to train machine learning algorithms to identify sequence motifs that define ionic selectivity, desensitization, and absorption spectra. Implementation of such algorithms has already helped to improve plasma membrane targeting and light sensitivity of ChRs (49, 50).
Protein sequences of dinoflagellate ChRs and their metagenomic homologs are distantly related to ChRs from giant viruses (Fig. 1), two of which have been shown recently to passively conduct cations upon heterologous expression (7). However, Asp212 of bacteriorhodopsin is conserved in these viral CCRs, as in most other known microbial rhodopsins, whereas it is replaced with Asn in dinoflagellate ChRs. Analysis of the Symbiodiniaceae transcriptomes reveals potential latent infection by large double-stranded DNA (dsDNA) viruses (51), so a viral origin of dinoflagellate ChRs cannot be excluded.
So far, the function of ChRs as photoreceptors guiding phototaxis has been verified directly only in the chlorophyte C. reinhardtii, the model organism for which methods of gene silencing and knockdown have been developed (1, 52). Several other chlorophyte and one cryptophyte species have been shown to generate photoreceptor currents, very similar to those in C. reinhardtii and likely resulting from ChR photoexcitation (53–56). The direction of photoreceptor currents recorded in both freshwater and marine flagellates is depolarizing, which reflects cation influx or anion efflux. Both C. reinhardtii phototaxis receptors are CCRs (2, 3), but ACRs might also contribute to depolarizing photoreceptor currents even in marine flagellates, if their membrane potential is sufficiently low. To the best of our knowledge, the membrane potential has not been estimated in any ACR-containing organism, but it is −170 mV in the giant marine unicellular alga Acetabularia mediterranea (57).
Based on the action spectra of dinoflagellate phototaxis, rhodopsins have been suggested as photoreceptors that guide this behavior (58). Our demonstration of channel activity in dinoflagellate rhodopsins with the TCP motif in TM3 strongly supports this hypothesis. The spectral sensitivity of dinoflagellate ChRs matches that of phototactic accumulation observed in Symbiodinium and unclassified coral symbiotic dinoflagellates (59, 60). The latter studies suggest that coral larvae use green fluorescent protein (GFP) fluorescence to attract dinoflagellate symbionts that are necessary for their survival.
Manual patch clamp is time-consuming and requires considerable skill. We sought to test whether APC can be used for characterization of hundreds of ChR variants that evolved in various protists. The planar-array principle implemented in the SyncroPatch 384i allows seal formation on micrometer-size orifices in the glass bottom of microwell plates (chips) into which cell suspension is pipetted, thus bypassing pipette fabrication and offering the option for recording multiple cells in parallel (61). APC is mostly used for drug screening, especially cardiac safety testing, in stably transfected cell lines. However, generation of such lines for ChR screening is not practical. We found that even upon chemical transfection that yielded only 30 to 70% visibly fluorescent cells depending on the construct, using the SyncroPatch 384i considerably sped up data collection, compared to manual patch clamp.
MATERIALS AND METHODS
Bioinformatics.
To identify metagenomic homologs of MerMAIDs, haptophyte ACRs, and Labyrinthulea ACRs, we first searched selected data sets of the Integrated Microbial Genomes and Microbiomes at the Department of Energy's Joint Genome Institute (JGI) (Data Set S4) using the keyword “rhodopsin,” and then performed a BLASTp (protein-protein BLAST) search using RubyACR sequences as a query. A similar procedure was used to identify rhodopsin genes in the dinoflagellate genomes from various sources listed in Data Set S4. Cafeteria roenbergensis ChRs were identified in the National Center for Biotechnology Information (NCBI) nonredundant protein database using BLASTp and the GtACR1 sequence as a query.
To explore the diversity of cryptophyte ACRs, we analyzed transcriptomes of 20 cryptophyte strains each sequenced on the Illumina HiSeq 2000 platform and assembled with the Bridger algorithm (62). Using a hidden Markov model (HMM) (63) based on known cryptophyte ACRs, we identified 15 novel transcripts for experimental characterization. We also analyzed 136 deep-ocean metatranscriptomic libraries from the TARA Oceans Expedition (64) assembled with the Plass protein-level algorithm (65). Four distinct HMMs were built using previously known sequences of cryptophyte ACRs, cryptophyte BCCRs, chlorophyte CCRs, and MerMAIDs. While many transcripts could be uniquely assigned to one of these four HMMs, some aligned weakly but equally well to two or more HMMs and could not be assigned unambiguously. Remarkably, 17 of these ambiguous sequences turned out to be close homologs of dinoflagellate ChRs that were not included among our HMMs.
Rhodopsin sequences from Data Set S1 were aligned using MUSCLE with default parameters as implemented in MegAlign Pro software v. 17.1.1 (DNASTAR Lasergene, Madison, WI) and truncated after the end of TM7. Phylogeny was analyzed with IQ-TREE v. 2.1.2 (66) using automatic model selection and ultrafast bootstrap approximation (1,000 replicates) (67). The best tree was visualized and annotated with iTOL v. 5.7 (68).
Molecular biology and HEK293 transfection.
DNA polynucleotides encoding the opsin domains optimized for human codon usage were synthesized and cloned by GenScript (Piscataway, NJ) into the mammalian expression vector pcDNA3.1 (Life Technologies, Grand Island, NY) in frame with an EYFP tag for expression in HEK293 cells. The cells were transfected using the ScreenFectA transfection reagent (Wako Chemicals USA, Richmond, VA). All-trans-retinal (Sigma) was added at the final concentration of 3 μM immediately after transfection.
Manual patch clamp recording.
Photocurrents were recorded 48 to 96 h after transfection in whole-cell voltage clamp mode with an AxoPatch 200B amplifier and digitized with a Digidata 1440A using pClamp 10 software (all from Molecular Devices, Union City, CA). Patch pipettes with resistances of 2 to 4 MΩ were fabricated from borosilicate glass. The ionic compositions of the bath and pipette solutions are shown in Table S1. For determination of Erev, K+ in the pipette solution was replaced with Na+ to minimize the number of ionic species in the system, and the holding voltages were corrected for liquid junction potentials calculated using the Clampex built-in calculator. Continuous light pulses were provided by a Polychrome V (T.I.L.L. Photonics GMBH, Grafelfing, Germany) in combination with a mechanical shutter (Uniblitz model LS6; Vincent Associates, Rochester, NY; half-opening time, 0.5 ms). The maximal photon density at the focal plane of the 40× objective was 5.2 to 8.5 mW mm−2 depending on the wavelength. The action spectra were constructed by calculation of the initial slope of photocurrent and corrected for the photon density measured at each wavelength (5). Further analysis was performed using Origin Pro software (OriginLab Corporation, Northampton, MA). The images were taken with a CoolSNAP HQ2 monochrome camera (Photometrics, Tucson, AZ).
Automated patch clamp recording.
Automated patch clamp recording was conducted with a SyncroPatch 384i (Nanion Technologies) using planar borosilicate glass medium-resistance chips in a 384-well microtiter plate format with one or four holes per well and Nanion Standard solutions (for their composition, see Table S1). Transfected cells were dissociated using TrypLE Express, diluted with CHO-S-SFM-II medium (both from Thermo Fisher) and resuspended in external physiological solution (Nanion Technologies) at 1 × 105 to 4 × 105 cells ml−1. Each well was filled with 30 μl Chip Fill solution, to which 20 μl of the cell suspension was added. Seal formation was enhanced by the addition of 40 μl of NMDG (N-methyl-d-glucamine) 60 solution with 10 mM CaCl2 (final concentration). After capturing the cells, 50 μl of the external solution was replaced with 40 μl of NMDG 60 solution, and 40 μl of the mixture was removed. For data acquisition and analysis, respectively, PatchControl384 and DataControl384 software v. 1.9.0 were used (both Nanion Technologies). Illumination was provided with Luxeon Z blue LEDs, LXZ1-PB01 (470 ± 20 nm), controlled by custom-built software.
Statistics.
Descriptive statistics was used as implemented in Origin software. The data are presented as means and standard errors of the means (SEM); the data from individual replicates are also shown when appropriate. The sample size was estimated from previous experience and published work on similar subjects, as recommended by the NIH guidelines. No normal distribution of the data was assumed; when a specific statistics hypothesis was tested, nonparametric tests were used.
Data availability.
The polynucleotide sequences of ChR homologs reported in this study have been deposited in GenBank (accession numbers MW557552 to MW557594).
ACKNOWLEDGMENTS
We thank Tim Strassmaier, Leo Angelo Morada, Stephan Holzhauser, and Rasmus Gönner (Nanion Technologies) for their expert help with the SyncroPatch 384i.
This work was supported by National Institutes of Health grants R35GM140838 and U01NS118288 and Endowed Chair AU-0009 from the Robert A. Welch Foundation to J.L.S. and by Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant RGPIN-2018-04397 to L.S.B. A.P. was supported by a NSERC USRA scholarship, and S.C. was supported by the Agricultural Science and Technology Innovation Program (ASTIP). The access to the SyncroPatch 384i was provided by a research grant from Nanion Technologies. The work conducted by the U.S. Department of Energy Joint Genome Institute, a DOE Office of Science User Facility, is supported by the Office of Science of the U.S. Department of Energy under contract DE-AC02-05CH11231.
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
We declare no conflict of interest.
Footnotes
This article is a direct contribution from John L. Spudich, a Fellow of the American Academy of Microbiology, who arranged for and secured reviews by Edward Boyden, Massachusetts Institute of Technology, and Arthur Grossman, Carnegie Institute at Stanford University.
Citation Govorunova EG, Sineshchekov OA, Li H, Wang Y, Brown LS, Palmateer A, Melkonian M, Cheng S, Carpenter E, Patterson J, Wong GKS, Spudich JL. 2021. Cation and anion channelrhodopsins: sequence motifs and taxonomic distribution. mBio 12:e01656-21. https://doi.org/10.1128/mBio.01656-21.
Contributor Information
John L. Spudich, Email: john.l.spudich@uth.tmc.edu.
Igor B. Zhulin, The Ohio State University
REFERENCES
- 1.Sineshchekov OA, Jung K-H, Spudich JL. 2002. Two rhodopsins mediate phototaxis to low- and high-intensity light in Chlamydomonas reinhardtii. Proc Natl Acad Sci U S A 99:8689–8694. doi: 10.1073/pnas.122243399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nagel G, Ollig D, Fuhrmann M, Kateriya S, Musti AM, Bamberg E, Hegemann P. 2002. Channelrhodopsin-1: a light-gated proton channel in green algae. Science 296:2395–2398. doi: 10.1126/science.1072068. [DOI] [PubMed] [Google Scholar]
- 3.Nagel G, Szellas T, Huhn W, Kateriya S, Adeishvili N, Berthold P, Ollig D, Hegemann P, Bamberg E. 2003. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc Natl Acad Sci U S A 100:13940–13945. doi: 10.1073/pnas.1936192100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Govorunova EG, Sineshchekov OA, Liu X, Janz R, Spudich JL. 2015. Natural light-gated anion channels: a family of microbial rhodopsins for advanced optogenetics. Science 349:647–650. doi: 10.1126/science.aaa7484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Govorunova EG, Sineshchekov OA, Li H, Wang Y, Brown LS, Spudich JL. 2020. RubyACRs, non-algal anion channelrhodopsins with highly red-shifted absorption. Proc Natl Acad Sci U S A 117:22833–22840. doi: 10.1073/pnas.2005981117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rozenberg A, Oppermann J, Wietek J, Fernandez Lahore RG, Sandaa RA, Bratbak G, Hegemann P, Béjà O. 2020. Lateral gene transfer of anion-conducting channelrhodopsins between green algae and giant viruses. Curr Biol 30:4910–4920. doi: 10.1016/j.cub.2020.09.056. [DOI] [PubMed] [Google Scholar]
- 7.Zabelskii D, Alekseev A, Kovalev K, Rankovic V, Balandin T, Soloviov D, Bratanov D, Savelyeva E, Podolyak E, Volkov D, Vaganova S, Astashkin R, Chizhov I, Yutin N, Rulev M, Popov A, Eria-Oliveira AS, Rokitskaya T, Mager T, Antonenko Y, Rosselli R, Armeev G, Shaitan K, Vivaudou M, Buldt G, Rogachev A, Rodriguez-Valera F, Kirpichnikov M, Moser T, Offenhausser A, Willbold D, Koonin E, Bamberg E, Gordeliy V. 2020. Viral rhodopsins 1 are an unique family of light-gated cation channels. Nat Commun 11:5707. doi: 10.1038/s41467-020-19457-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Govorunova EG, Sineshchekov OA, Li H, Spudich JL. 2017. Microbial rhodopsins: diversity, mechanisms, and optogenetic applications. Annu Rev Biochem 86:845–872. doi: 10.1146/annurev-biochem-101910-144233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Deisseroth K. 2015. Optogenetics: 10 years of microbial opsins in neuroscience. Nat Neurosci 18:1213–1225. doi: 10.1038/nn.4091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sasse P, Funken M, Beiert T, Bruegmann T. 2019. Optogenetic termination of cardiac arrhythmia: mechanistic enlightenment and therapeutic application? Front Physiol 10:675. doi: 10.3389/fphys.2019.00675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kato HE, Zhang F, Yizhar O, Ramakrishnan C, Nishizawa T, Hirata K, Ito J, Aita Y, Tsukazaki T, Hayashi S, Hegemann P, Maturana AD, Ishitani R, Deisseroth K, Nureki O. 2012. Crystal structure of the channelrhodopsin light-gated cation channel. Nature 482:369–374. doi: 10.1038/nature10870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Volkov O, Kovalev K, Polovinkin V, Borshchevskiy V, Bamann C, Astashkin R, Marin E, Popov A, Balandin T, Willbold D, Buldt G, Bamberg E, Gordeliy V. 2017. Structural insights into ion conduction by channelrhodopsin 2. Science 358:eaan8862. doi: 10.1126/science.aan8862. [DOI] [PubMed] [Google Scholar]
- 13.Oda K, Vierock J, Oishi S, Rodriguez-Rozada S, Taniguchi R, Yamashita K, Wiegert JS, Nishizawa T, Hegemann P, Nureki O. 2018. Crystal structure of the red light-activated channelrhodopsin Chrimson. Nat Commun 9:3949. doi: 10.1038/s41467-018-06421-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kim YS, Kato HE, Yamashita K, Ito S, Inoue K, Ramakrishnan C, Fenno LE, Evans KE, Paggi JM, Dror RO, Kandori H, Kobilka BK, Deisseroth K. 2018. Crystal structure of the natural anion-conducting channelrhodopsin GtACR1. Nature 561:343–348. doi: 10.1038/s41586-018-0511-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li H, Huang CY, Govorunova EG, Schafer CT, Sineshchekov OA, Wang M, Zheng L, Spudich JL. 2019. Crystal structure of a natural light-gated anion channelrhodopsin. Elife 8:e41741. doi: 10.7554/eLife.41741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhang F, Vierock J, Yizhar O, Fenno LE, Tsunoda S, Kianianmomeni A, Prigge M, Berndt A, Cushman J, Polle J, Magnuson J, Hegemann P, Deisseroth K. 2011. The microbial opsin family of optogenetic tools. Cell 147:1446–1457. doi: 10.1016/j.cell.2011.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Berndt A, Lee SY, Ramakrishnan C, Deisseroth K. 2014. Structure-guided transformation of channelrhodopsin into a light-activated chloride channel. Science 344:420–424. doi: 10.1126/science.1252367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wietek J, Beltramo R, Scanziani M, Hegemann P, Oertner TG, Wiegert JS. 2015. An improved chloride-conducting channelrhodopsin for light-induced inhibition of neuronal activity in vivo. Sci Rep 5:14807. doi: 10.1038/srep14807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wietek J, Wiegert JS, Adeishvili N, Schneider F, Watanabe H, Tsunoda SP, Vogt A, Elstner M, Oertner TG, Hegemann P. 2014. Conversion of channelrhodopsin into a light-gated chloride channel. Science 344:409–412. doi: 10.1126/science.1249375. [DOI] [PubMed] [Google Scholar]
- 20.Berndt A, Lee SY, Wietek J, Ramakrishnan C, Steinberg EE, Rashid AJ, Kim H, Park S, Santoro A, Frankland PW, Iyer SM, Pak S, Ahrlund-Richter S, Delp SL, Malenka RC, Josselyn SA, Carlen M, Hegemann P, Deisseroth K. 2016. Structural foundations of optogenetics: determinants of channelrhodopsin ion selectivity. Proc Natl Acad Sci U S A 113:822–829. doi: 10.1073/pnas.1523341113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wietek J, Rodriguez-Rozada S, Tutas J, Tenedini F, Grimm C, Oertner TG, Soba P, Hegemann P, Wiegert JS. 2017. Anion-conducting channelrhodopsins with tuned spectra and modified kinetics engineered for optogenetic manipulation of behavior. Sci Rep 7:14957. doi: 10.1038/s41598-017-14330-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Oppermann J, Fischer P, Silapetere A, Liepe B, Rodriguez-Rozada S, Flores-Uribe J, Peter E, Keidel A, Vierock J, Kaufmann J, Broser M, Luck M, Bartl F, Hildebrandt P, Simon Wiegert J, Beja O, Hegemann P, Wietek J. 2019. MerMAIDs: a family of metagenomically discovered marine anion-conducting and intensely desensitizing channelrhodopsins. Nat Commun 10:3315. doi: 10.1038/s41467-019-11322-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sineshchekov OA, Govorunova EG, Li H, Wang Y, Melkonian M, Wong GK-S, Brown LS, Spudich JL. 2020. Conductance mechanisms of rapidly desensitizing cation channelrhodopsins from cryptophyte algae. mBio 11:e00657-20. doi: 10.1128/mBio.00657-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sineshchekov OA, Govorunova EG, Wang J, Li H, Spudich JL. 2013. Intramolecular proton transfer in channelrhodopsins. Biophys J 104:807–817. doi: 10.1016/j.bpj.2013.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dreier MA, Althoff P, Norahan MJ, Tennigkeit SA, El-Mashtoly SF, Lübben M, Kötting C, Rudack T, Gerwert K. 2021. Time-resolved spectroscopic and electrophysiological data reveal insights in the gating mechanism of anion channelrhodopsin. Commun Biol 4:578. doi: 10.1038/s42003-021-02101-5. [DOI] [PMC free article] [PubMed]
- 26.Labarre A, Lopez-Escardo D, Latorre F, Leonard G, Bucchini F, Obiol A, Cruaud C, Sieracki ME, Jaillon O, Wincker P, Vandepoele K, Logares R, Massana R. 2021. Comparative genomics reveals new functional insights in uncultured MAST species. ISME J 15:1767–1781. doi: 10.1038/s41396-020-00885-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kojima K, Miyoshi N, Shibukawa A, Chowdhury S, Tsujimura M, Noji T, Ishikita H, Yamanaka A, Sudo Y. 2020. Green-sensitive, long-lived, step-functional anion channelrhodopsin-2 variant as a high-potential neural silencing tool. J Phys Chem Lett 11:6214–6218. doi: 10.1021/acs.jpclett.0c01406. [DOI] [PubMed] [Google Scholar]
- 28.Forward RB. 1974. Phototaxis by the dinoflagellate Gymnodinium splendens Lebour. J Protozool 21:312–315. doi: 10.1111/j.1550-7408.1974.tb03659.x. [DOI] [PubMed] [Google Scholar]
- 29.Liu S-M, Häder D-P, Ullrich W. 1990. Photoorientation in the freshwater dinoflagellate, Peridinium gatunense Nygaard. FEMS Microbiol Lett 73:91–102. doi: 10.1111/j.1574-6968.1990.tb03929.x. [DOI] [Google Scholar]
- 30.Horiguchi T, Kawai H, Kubota M, Takahashi T, Watanabe M. 1999. Phototactic responses of four marine dinoflagellates with different types of eyespot and chloroplast. Phycol Res 47:101–107. doi: 10.1046/j.1440-1835.1999.47220158.x. [DOI] [Google Scholar]
- 31.Slamovits CH, Okamoto N, Burri L, James ER, Keeling PJ. 2011. A bacterial proteorhodopsin proton pump in marine eukaryotes. Nat Commun 2:183. doi: 10.1038/ncomms1188. [DOI] [PubMed] [Google Scholar]
- 32.Marchetti A, Schruth DM, Durkin CA, Parker MS, Kodner RB, Berthiaume CT, Morales R, Allen AE, Armbrust EV. 2012. Comparative metatranscriptomics identifies molecular bases for the physiological responses of phytoplankton to varying iron availability. Proc Natl Acad Sci U S A 109:E317–E325. doi: 10.1073/pnas.1118408109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Coesel SN, Durham BP, Groussman RD, Hu SK, Caron DA, Morales RL, Ribalet F, Armbrust EV. 2021. Diel transcriptional oscillations of light-sensitive regulatory elements in open-ocean eukaryotic plankton communities. Proc Natl Acad Sci U S A 118:e2011038118. doi: 10.1073/pnas.2011038118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shoguchi E, Shinzato C, Kawashima T, Gyoja F, Mungpakdee S, Koyanagi R, Takeuchi T, Hisata K, Tanaka M, Fujiwara M, Hamada M, Seidi A, Fujie M, Usami T, Goto H, Yamasaki S, Arakaki N, Suzuki Y, Sugano S, Toyoda A, Kuroki Y, Fujiyama A, Medina M, Coffroth MA, Bhattacharya D, Satoh N. 2013. Draft assembly of the Symbiodinium minutum nuclear genome reveals dinoflagellate gene structure. Curr Biol 23:1399–1408. doi: 10.1016/j.cub.2013.05.062. [DOI] [PubMed] [Google Scholar]
- 35.Aranda M, Li Y, Liew YJ, Baumgarten S, Simakov O, Wilson MC, Piel J, Ashoor H, Bougouffa S, Bajic VB, Ryu T, Ravasi T, Bayer T, Micklem G, Kim H, Bhak J, LaJeunesse TC, Voolstra CR. 2016. Genomes of coral dinoflagellate symbionts highlight evolutionary adaptations conducive to a symbiotic lifestyle. Sci Rep 6:39734. doi: 10.1038/srep39734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shoguchi E, Beedessee G, Tada I, Hisata K, Kawashima T, Takeuchi T, Arakaki N, Fujie M, Koyanagi R, Roy MC, Kawachi M, Hidaka M, Satoh N, Shinzato C. 2018. Two divergent Symbiodinium genomes reveal conservation of a gene cluster for sunscreen biosynthesis and recently lost genes. BMC Genomics 19:458. doi: 10.1186/s12864-018-4857-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.LaJeunesse TC, Parkinson JE, Gabrielson PW, Jeong HJ, Reimer JD, Voolstra CR, Santos SR. 2018. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr Biol 28:2570–2580.E6. doi: 10.1016/j.cub.2018.07.008. [DOI] [PubMed] [Google Scholar]
- 38.Grigoriev IV, Hayes RD, Calhoun S, Kamel B, Wang A, Ahrendt S, Dusheyko S, Nikitin R, Mondo SJ, Salamov A, Shabalov I, Kuo A. 2021. PhycoCosm, a comparative algal genomics resource. Nucleic Acids Res 49:D1004–D1011. doi: 10.1093/nar/gkaa898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gradinaru V, Zhang F, Ramakrishnan C, Mattis J, Prakash R, Diester I, Goshen I, Thompson KR, Deisseroth K. 2010. Molecular and cellular approaches for diversifying and extending optogenetics. Cell 141:154–165. doi: 10.1016/j.cell.2010.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Olsson MHM, Sondergaard CR, Rostkowski M, Jensen JH. 2011. PROPKA3: consistent treatment of internal and surface residues in empirical pK(a) predictions. J Chem Theory Comput 7:525–537. doi: 10.1021/ct100578z. [DOI] [PubMed] [Google Scholar]
- 41.Sugiyama Y, Wang H, Hikima T, Sato M, Kuroda J, Takahashi T, Ishizuka T, Yawo H. 2009. Photocurrent attenuation by a single polar-to-nonpolar point mutation of channelrhodopsin-2. Photochem Photobiol Sci 8:328–336. doi: 10.1039/b815762f. [DOI] [PubMed] [Google Scholar]
- 42.Ritter E, Stehfest K, Berndt A, Hegemann P, Bartl FJ. 2008. Monitoring light-induced structural changes of channelrhodopsin-2 by UV-visible and Fourier transform infrared spectroscopy. J Biol Chem 283:35033–35041. doi: 10.1074/jbc.M806353200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Yi A, Mamaeva NV, Li H, Spudich JL, Rothschild KJ. 2016. Resonance Raman study of an anion channelrhodopsin: effects of mutations near the retinylidene Schiff base. Biochemistry 55:2371–2380. doi: 10.1021/acs.biochem.6b00104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lorenz-Fonfria VA, Resler T, Krause N, Nack M, Gossing M, Fischer von Mollard G, Bamann C, Bamberg E, Schlesinger R, Heberle J. 2013. Transient protonation changes in channelrhodopsin-2 and their relevance to channel gating. Proc Natl Acad Sci U S A 110:E1273–E1281. doi: 10.1073/pnas.1219502110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kuhne J, Vierock J, Tennigkeit SA, Dreier MA, Wietek J, Petersen D, Gavriljuk K, El-Mashtoly SF, Hegemann P, Gerwert K. 2019. Unifying photocycle model for light adaptation and temporal evolution of cation conductance in channelrhodopsin-2. Proc Natl Acad Sci U S A 116:9380–9389. doi: 10.1073/pnas.1818707116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sineshchekov OA, Li H, Govorunova EG, Spudich JL. 2016. Photochemical reaction cycle transitions during anion channelrhodopsin gating. Proc Natl Acad Sci U S A 113:E1993–E2000. doi: 10.1073/pnas.1525269113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lorenz-Fonfria VA, Heberle J. 2014. Channelrhodopsin unchained: structure and mechanism of a light-gated cation channel. Biochim Biophys Acta 1837:626–642. doi: 10.1016/j.bbabio.2013.10.014. [DOI] [PubMed] [Google Scholar]
- 48.Tsujimura M, Noji T, Saito K, Kojima K, Sudo Y, Ishikita H. 2021. Mechanism of absorption wavelength shifts in anion channelrhodopsin-1 mutants. Biochim Biophys Acta Bioenerg 1862:148349. doi: 10.1016/j.bbabio.2020.148349. [DOI] [PubMed] [Google Scholar]
- 49.Bedbrook CN, Yang KK, Rice AJ, Gradinaru V, Arnold FH. 2017. Machine learning to design integral membrane channelrhodopsins for efficient eukaryotic expression and plasma membrane localization. PLoS Comput Biol 13:e1005786. doi: 10.1371/journal.pcbi.1005786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bedbrook CN, Yang KK, Robinson JE, Mackey ED, Gradinaru V, Arnold FH. 2019. Machine learning-guided channelrhodopsin engineering enables minimally invasive optogenetics. Nat Methods 16:1176–1184. doi: 10.1038/s41592-019-0583-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lawrence SA, Floge SA, Davy JE, Davy SK, Wilson WH. 2017. Exploratory analysis of Symbiodinium transcriptomes reveals potential latent infection by large dsDNA viruses. Environ Microbiol 19:3909–3919. doi: 10.1111/1462-2920.13782. [DOI] [PubMed] [Google Scholar]
- 52.Greiner A, Kelterborn S, Evers H, Kreimer G, Sizova I, Hegemann P. 2017. Targeting of photoreceptor genes in Chlamydomonas reinhardtii via zinc-finger nucleases and CRISPR/Cas9. Plant Cell 29:2498–2518. doi: 10.1105/tpc.17.00659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Litvin FF, Sineshchekov OA, Sineshchekov VA. 1978. Photoreceptor electric potential in the phototaxis of the alga Haematococcus pluvialis. Nature 271:476–478. doi: 10.1038/271476a0. [DOI] [PubMed] [Google Scholar]
- 54.Sineshchekov OA, Govorunova EG, Jung K-H, Zauner S, Maier U-G, Spudich JL. 2005. Rhodopsin-mediated photoreception in cryptophyte flagellates. Biophys J 89:4310–4319. doi: 10.1529/biophysj.105.070920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Govorunova EG, Spudich EN, Lane CE, Sineshchekov OA, Spudich JL. 2011. New channelrhodopsin with a red-shifted spectrum and rapid kinetics from Mesostigma viride. mBio 2:e00115-11. doi: 10.1128/mBio.00115-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Govorunova EG, Sineshchekov OA, Li H, Janz R, Spudich JL. 2013. Characterization of a highly efficient blue-shifted channelrhodopsin from the marine alga Platymonas subcordiformis. J Biol Chem 288:29911–29922. doi: 10.1074/jbc.M113.505495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Saddler HDW. 1970. The membrane potential of Acetabularia mediterranea. J Gen Physiol 55:802–821. doi: 10.1085/jgp.55.6.802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Foster K-W, Smyth RD. 1980. Light antennas in phototactic algae. Microbiol Rev 44:572–630. doi: 10.1128/mr.44.4.572-630.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hollingsworth LL, Kinzie RA, Lewis TD, Krupp DA, Leong JAC. 2005. Phototaxis of motile zooxanthellae to green light may facilitate symbiont capture by coral larvae. Coral Reefs 24:523–523. doi: 10.1007/s00338-005-0063-8. [DOI] [Google Scholar]
- 60.Aihara Y, Maruyama S, Baird AH, Iguchi A, Takahashi S, Minagawa J. 2019. Green fluorescence from cnidarian hosts attracts symbiotic algae. Proc Natl Acad Sci U S A 116:2118–2123. doi: 10.1073/pnas.1812257116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Obergrussberger A, Friis S, Bruggemann A, Fertig N. 2021. Automated patch clamp in drug discovery: major breakthroughs and innovation in the last decade. Expert Opin Drug Discov 16:1–5. doi: 10.1080/17460441.2020.1791079. [DOI] [PubMed] [Google Scholar]
- 62.Chang Z, Li G, Liu J, Zhang Y, Ashby C, Liu D, Cramer CL, Huang X. 2015. Bridger: a new framework for de novo transcriptome assembly using RNA-seq data. Genome Biol 16:30. doi: 10.1186/s13059-015-0596-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Eddy SR. 2011. Accelerated profile HMM searches. PLoS Comput Biol 7:e1002195. doi: 10.1371/journal.pcbi.1002195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Carradec Q, Pelletier E, Da Silva C, Alberti A, Seeleuthner Y, Blanc-Mathieu R, Lima-Mendez G, Rocha F, Tirichine L, Labadie K, Kirilovsky A, Bertrand A, Engelen S, Madoui MA, Meheust R, Poulain J, Romac S, Richter DJ, Yoshikawa G, Dimier C, Kandels-Lewis S, Picheral M, Searson S, Tara Oceans C, Jaillon O, Aury JM, Karsenti E, Sullivan MB, Sunagawa S, Bork P, Not F, Hingamp P, Raes J, Guidi L, Ogata H, de Vargas C, Iudicone D, Bowler C, Wincker P, Tara Oceans Coordinators . 2018. A global ocean atlas of eukaryotic genes. Nat Commun 9:373. doi: 10.1038/s41467-017-02342-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Steinegger M, Mirdita M, Söding J. 2019. Protein-level assembly increases protein sequence recovery from metagenomic samples manyfold. Nat Methods 16:603–606. doi: 10.1038/s41592-019-0437-4. [DOI] [PubMed] [Google Scholar]
- 66.Minh BQ, Schmidt HA, Chernomor O, Schrempf D, Woodhams MD, von Haeseler A, Lanfear R. 2020. IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol Biol Evol 37:1530–1534. doi: 10.1093/molbev/msaa015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hoang DT, Chernomor O, von Haeseler A, Minh BQ, Vinh LS. 2018. UFBoot2: improving the ultrafast bootstrap approximation. Mol Biol Evol 35:518–522. doi: 10.1093/molbev/msx281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Letunic I, Bork P. 2019. Interactive Tree Of Life (iTOL) v4: recent updates and new developments. Nucleic Acids Res 47:W256–W259. doi: 10.1093/nar/gkz239. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Newick file of the tree shown in Fig. 1. Download Data Set S2, TXT file, 0.02 MB (21KB, txt) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
An alignment of the C-truncated sequences used to construct the tree in Fig. 1. Download Data Set S3, TXT file, 0.6 MB (679.7KB, txt) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
(A) Residues of the retinal-binding pockets. Variants tested in this study are in bold. The residues in MsCCR and CsCCR that differ from those in Chrimson and RubyACRs, earlier known red-shifted ChRs, are highlighted red. The numbers are according to bacteriorhodopsin sequence. (B and D) The residues in the positions of the conserved glutamates (highlighted in red) in the ion conductance pathway in the indicated homologs from Chlorophyceae (B) and Chlorodendrophyceae (D). Variants tested in this study are in bold; nonfunctional variants are in gray. The numbers are according to the CrChR2 sequence. (C) Photocurrent traces recorded from C1ChR in response to 1-s illumination at −60 and 60 mV. (E) Alignment of the part of TM7 of the indicated Chlorodendrophyceae homologs. The Schiff base Lys is highlighted in blue; the upstream Glu, in red. Abbreviations: MsCCR, Mantoniella squamata cation channelrhodopsin; CsCCR, Crustomastix stigmatica cation channelrhodopsin; CrChR2, Chlamydomonas reinhardtii channelrhodopsin 2; PymeACR, Pyramimonas melkonianii anion channelrhodopsin; CnChR3, Chlamydomonas noctigama channelrhodopsin 3; CnChR4, Chlamydomonas noctigama channelrhodopsin 4; C1ChR, Chlamydomonas sp. channelrhodopsin; CsChR2, Chloromonas subdivisa channelrhodopsin 2; PsChR4, Platymonas (Tetraselmis) subcordiformis channelrhodopsin 4; TaChR, Tetraselmis astigmatica channelrhodopsin; TchChR, Tetraselmis chui channelrhodopsin; TsChR2, Tetraselmis striata channelrhodopsin 2. Download FIG S1, TIF file, 0.4 MB (386.5KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Solution compositions in AxoPatch and SyncroPatch recordings. Abbreviations: Asp, aspartate; LJP, liquid junction potential; NMDG, N-methyl-d-glucamine. All concentrations are in millimolar units. Download Table S1, DOCX file, 0.02 MB (16.4KB, docx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
A section of the phylogenetic tree from Fig. 1 redrawn in a rectangular format. Red nodes show variants with proven anion selectivity, thicker nodes show variants tested in this study. Red numbers are the values of photocurrent desensitization from Fig. 3C in the main text (mean ± SEM, n = 5 or 6 cells). Abbreviations: sTACR, stramenopile strain TOSAG23-3 anion channelrhodopsin; mgACR, metagenomic anion channelrhodopsin. Download FIG S2, TIF file, 0.5 MB (540KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
GenBank accession numbers, abbreviated protein names, source organisms, habitats, transcript names, and amino acid sequences used to construct the phylogenetic tree in Fig. 1. Note that only sequences that cover the entire N-terminal and rhodopsin domains are included. Literature references to identification and electrophysiological characterization of the sequences are also provided. The sequences identified and characterized in this study are shown in bold. Download Data Set S1, XLSX file, 0.1 MB (95.1KB, xlsx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Sequence databases searched. Download Data Set S4, XLSX file, 0.01 MB (12.4KB, xlsx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
List of cryptophyte strains analyzed. Abbreviations: BEA, Banco Español de Algas, of the Universidad of Las Palmas de Gran Canaria, Spain; CCAC, Culture Collection of Algae at the University of Cologne, Germany; SCCAP, Scandinavian Culture Collection of Algae and Protozoa at the University of Copenhagen, Denmark; CCMP, Culture Collection of Marine Phytoplankton at the Provasoli-Guillard National Center for Marine Algae and Microbiota at Woods Hole Oceanographic Institution, USA; NIES, National Institute for Environmental Studies, Tsukuba, Japan. Download Table S2, DOCX file, 0.02 MB (16.4KB, docx) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
(A and B) Alignments of TM3 (A) and TM7 (B) of dinoflagellate ChRs and their metagenomic homologs. The TCP motif and the Asn residue corresponding to Asp212 of bacteriorhodopsin are highlighted in red. (C) EYFP tag fluorescence in cells expressing mgdChR1 constructs schematically shown on top of the images. TS, trafficking signal, ER, endoplasmic reticulum export motif. Abbreviations: AngChR, Ansanella granifera channelrhodopsin; PbChR, Pelagodinium bei channelrhodopsin; SA3ChR1-4, Symbiodinium clade A3 channelrhodopsins 1 to 4; SmChR1-3, Symbiodinium microadriaticum channelrhodopsins 1 to 3; SmChR1-3, Symbiodinium (Breviolum) minutum channelrhodopsins 1 to 3; mgdChR1-17, metagenomic dinoflagellate homolog channelrhodopsins 1 to 17. Download FIG S3, TIF file, 0.7 MB (765.2KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Alignment of Symbiodinium microadriaticum ChR1 with and without the N-terminal extension. Download FIG S4, TIF file, 0.2 MB (199.9KB, tif) .
Copyright © 2021 Govorunova et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Data Availability Statement
The polynucleotide sequences of ChR homologs reported in this study have been deposited in GenBank (accession numbers MW557552 to MW557594).