

Abstract
Studies over the years have described a filamentous structure to mature elastin that suggests a complicated packing arrangement of tropoelastin subunits. The currently accepted mechanism for tropoelastin assembly requires microfibrils to serve as a physical extracellular scaffold for alignment of tropoelastin monomers during and before crosslinking. However, recent evidence suggests that the initial stages of tropoelastin assembly occur within the cell or at unique assembly sites on the plasma membrane where tropoelastin self assembles to form elastin aggregates. Outside the cell, elastin aggregates transfer to growing elastic fibers in the extracellular matrix where tensional forces on microfibrils generated through cell movement help shape the growing fiber. Overall, these observations challenge the widely held idea that interaction between monomeric tropoelastin and microfibrils is a requirement for elastin assembly, and point to self-assembly of tropoelastin as a driving force in elastin maturation.
Keywords: elastin, elastic fiber, microfibrils, elastin assembly, ultrastructure
Introduction
The protein composition of the elastic fiber has been the subject of intensive biochemical investigation beginning in the 1960s. That the amorphous component was cross-linked elastin was quite clear from studies of Partridge and others [1, 2], but the nature of the microfibril turned out to be a challenging puzzle that is still being solved today. Ross and Bornstein [3] were among the first to take a biochemical and ultrastructural approach to identify elastic fiber components other than elastin, and their method of using denaturing agents together with reduction and alkylation is still the basis for many current techniques for elastic fiber protein isolation. These early efforts resulted in the isolation of multiple extraction “fractions” that were characterized by amino acid analysis and established that microfibrils were heterogeneous and contained numerous components, but no specific proteins were identified [4].
In France, Ladislas (Leslie) Robert and colleagues were also using biochemical approaches to characterize elastic fiber protein [5]. They found that elastic fibers could be solubilized using base hydrolysis, producing a soluble form of cross-linked elastin (kappa-elastin) that remains a valuable reagent for biochemical and cell biological studies [6]. They also found that extracts of elastin-rich tissues contained what they had previously identified as “structural glycoproteins” [7]. Robert postulated that these unique proteins played a structural role in the matrix. He further proposed that microfibrils in elastic fibers contained members of this glycoprotein class. The specific function of these proteins was challenging to demonstrate using the techniques available at the time. Robert and colleagues persisted with attempts at isolating and characterizing members of this heterogenous group of molecules, even in the face of some skepticism as to their existence [8]. It is now clear that most matrix proteins serve some structural role and several such glycoproteins have been identified, including fibronectin among others. In hindsight, it is not surprising that Robert and others had little success in identifying specific microfibrillar components in light of their close association (if not covalent linkage) with other structural elements of the ECM. The concept of structural glycoproteins was, however, a useful stimulus to research in the field of matrix biology. Leslie’s contributions to elastin biology extend far beyond structural glycoproteins, however. His group was at the forefront of research on elastases, vascular disease, cell-matrix signaling, functional propeptides of elastin fragments, lipid biology, and, one of his favorite topics, aging. Over his research career, Leslie generated a pervasive and outstanding body of work that has withstood the test of time in terms of relevance and applicability to current connective tissue research. We acknowledge the contribution of Leslie Robert and colleagues in helping us understand the biology of the elastic fiber. This minireview is dedicated to his memory and to the significant contributions he made to extracellular matrix research.
Tropoelastin
The structure of the elastic fiber remains an enigma. In the electron microscope, elastic fibers in mature tissues consist of amorphous elastin (the cross-linked form of the monomer tropoelastin) with a thin sheath of 10–15 nm filaments named microfibrils [9, 10]. In fetal and postnatal stages of fiber organization, however, the microfibrils predominate, with elastin accumulating within the microfibrillar bed. Thus, throughout development and in tissue repair, the relative composition of the elastic fiber changes from mostly microfibrils in the immature stage to mostly elastin in the fully mature fiber.
This close structural relationship between amorphous elastin and microfibrils led to the hypothesis that microfibrils provide an extracellular scaffold for tropoelastin assembly during and before crosslinking [3, 11]. This idea has changed little since first proposed, but recent studies using tools of cell and molecular biology, as well as the power of genetic manipulation in model organisms, suggest the process is more complicated than initially suggested. A long-standing question in the field is the extent to which elastin can self-assemble without assistance from other molecules.
A single gene encodes elastin in most vertebrates. The exceptions are teleosts and frogs, which have two elastin genes (for a detailed description of elastin gene structure and evolution, see reviews by Keeley and colleagues [12, 13]). Elastin first appeared in vertebrates and, hence, evolved independently of fibrillin and several other microfibrillar proteins that are evolutionarily older. The elastin gene product, tropoelastin, is a protein of approximately 65 kDa with an unusual amino acid composition and repeating domain structure (Figure 1A). Hydrophobic domains consist of a high proportion of small side-chain, nonpolar amino acids, with glycine, alanine, proline, isoleucine, leucine, and valine accounting for close to 80 % of all amino acids. Except for human and bovine elastin and the eln-2/eln-b tropoelastins in frog and fish, negatively charged amino acids are absent from the other tropoelastins for which sequence is known, as are tryptophan, histidine, and internal methionine residues. Crosslinking domains contain the lysines destined to participate in covalent cross-link formation, predominantly in KAAK or KAAAK motifs but may be of the KP-type, consisting of KxxK motifs in which one or both of the x residues are proline. The cross-links and crosslinking process of elastin has been well described, and the interested reader should consult [14–16] for further information. A detailed description of functional domains in elastin can be found in Vindin et al. [17] and Reichheld [13] in this minireview series.
Figure 1.
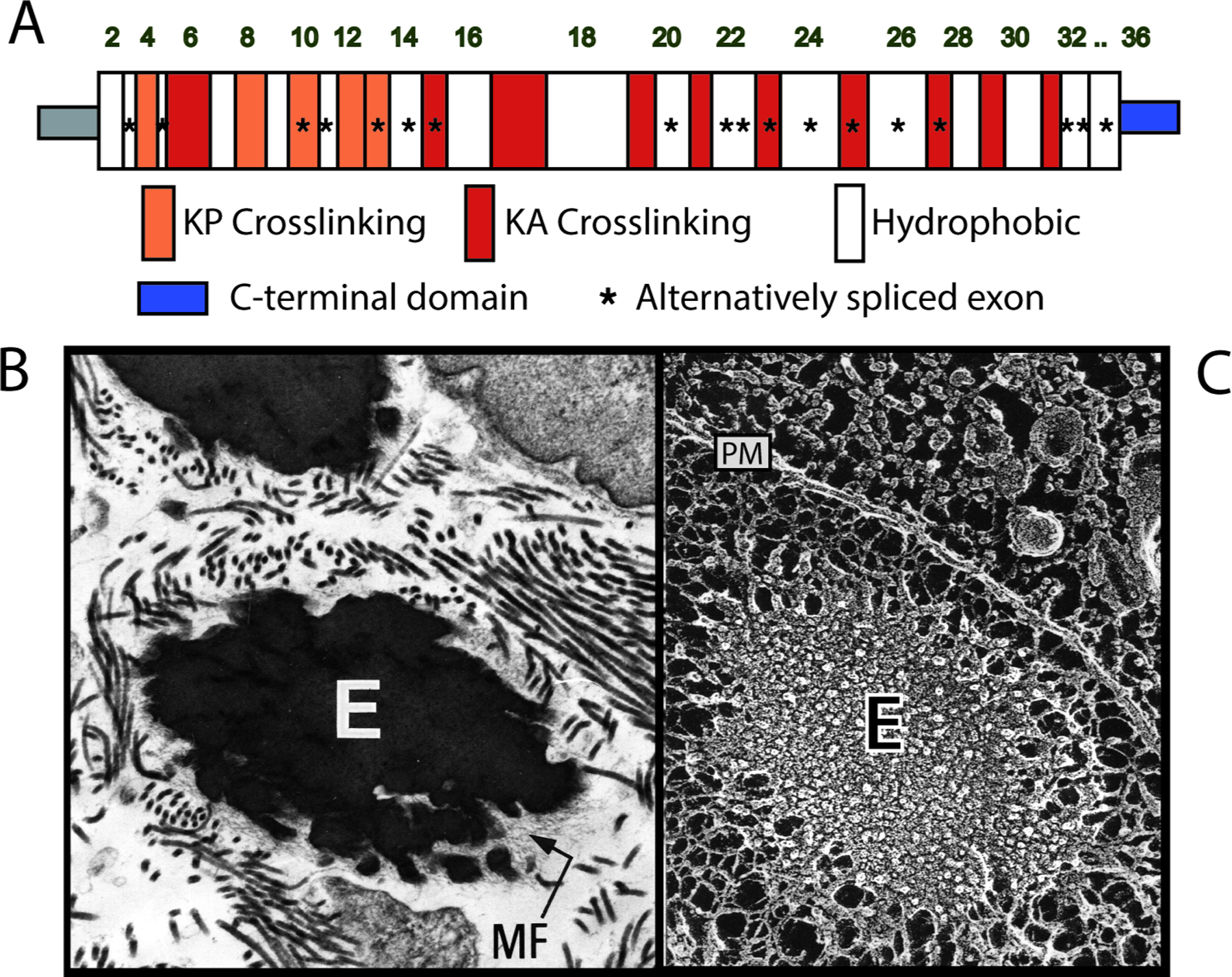
Domains of tropoelastin and structure of the elastic fiber.
A) Domain map of human tropoelastin showing the repeating hydrophobic and cross-linking domains. Domains numbering is according to encoding exons. B): Transmission electron micrograph (TEM) and C) quick freeze deep etch (QFDE) electron micrograph of an extracellular elastic fiber. The elastic fiber visualized by TEM shows an amorphous elastin core with microfibrils around the periphery. Structure within the elastin component is more detailed when visualized using QFDE. E=elastin, MF=microfibrils, PM=plasma membrane.
Elastin assembly and molecular packing
In thin sections of plastic-embedded material, elastin has an amorphous appearance when viewed by transmission electron microscopy (Figure 1B). Several laboratories, however, have reported a filamentous substructure after negative staining with uranyl acetate, especially when elastin is stretched before fixation. Fiber diameter varies widely from just over 1 nm to 4–5 nm, depending on the report [18–20]. Others have observed that purified or sonicated and negatively stained elastin consists of roughly parallel filaments, about 4–5 nm wide [21]. Tropoelastin itself forms filaments of 5 nm diameter [22], but only in the coacervated state above 40°C [22, 23]. Filaments with similar dimensions were also described in coacervates of alpha-elastin and elastin peptides [24–28]. The main criticism of these studies was that negative staining, dehydration, and heavy metal staining induced the artifactual formation of fibers from smaller molecular structures. However, when cyro methods were used to examine elastin ultrastructure in unfixed or slightly fixed tissues, filaments of diameter 5 nm were easily identified [29–32]. Optical diffraction, as well as low-angle X-ray scattering found evidence for similarly sized filaments [33].
Scanning force microscopy showed that in the relaxed fiber, cross-linked elastin molecules form a tangled network. Upon stretching, ordered longitudinal sets of beaded filaments became evident, laterally connected at regular intervals of less than 15 nm [34]. These results suggest that cross-linked elastin molecules exhibit globular domains preferentially linked in one direction to form filaments and laterally joined at regular intervals. Similar findings were obtained using freeze-fracture procedures followed by deep etching that revealed a three-dimensional network made up of globular elements of tropoelastin molecules that form discrete filaments when aggregated [31, 35, 36]. Deep-etch electron microscopy is a high-resolution imaging technique that avoids many of the artifacts inherent in standard sample preparation for electron microscopy, including the distortions produced by chemical dehydration, air-drying and embedding in plastic [37–39]. Figure 2 shows quick freeze, deep etch images of elastic fibers in unfixed tissue. What is immediately evident from the micrograph in Figure 2A is the clear structural definition of the elastin component. The primary structural feature is a densely packed, randomly arranged network of fine filaments. When viewed in three dimensions (Figure 2C), fine filaments having a mean diameter of 5–7 nm can be seen protruding from the sectioned fiber. In an unstretched fiber, the filaments have a disordered arrangement and pack so tightly that little or no etching occurs during sample preparation (Figure 2B & D).
Figure 2.
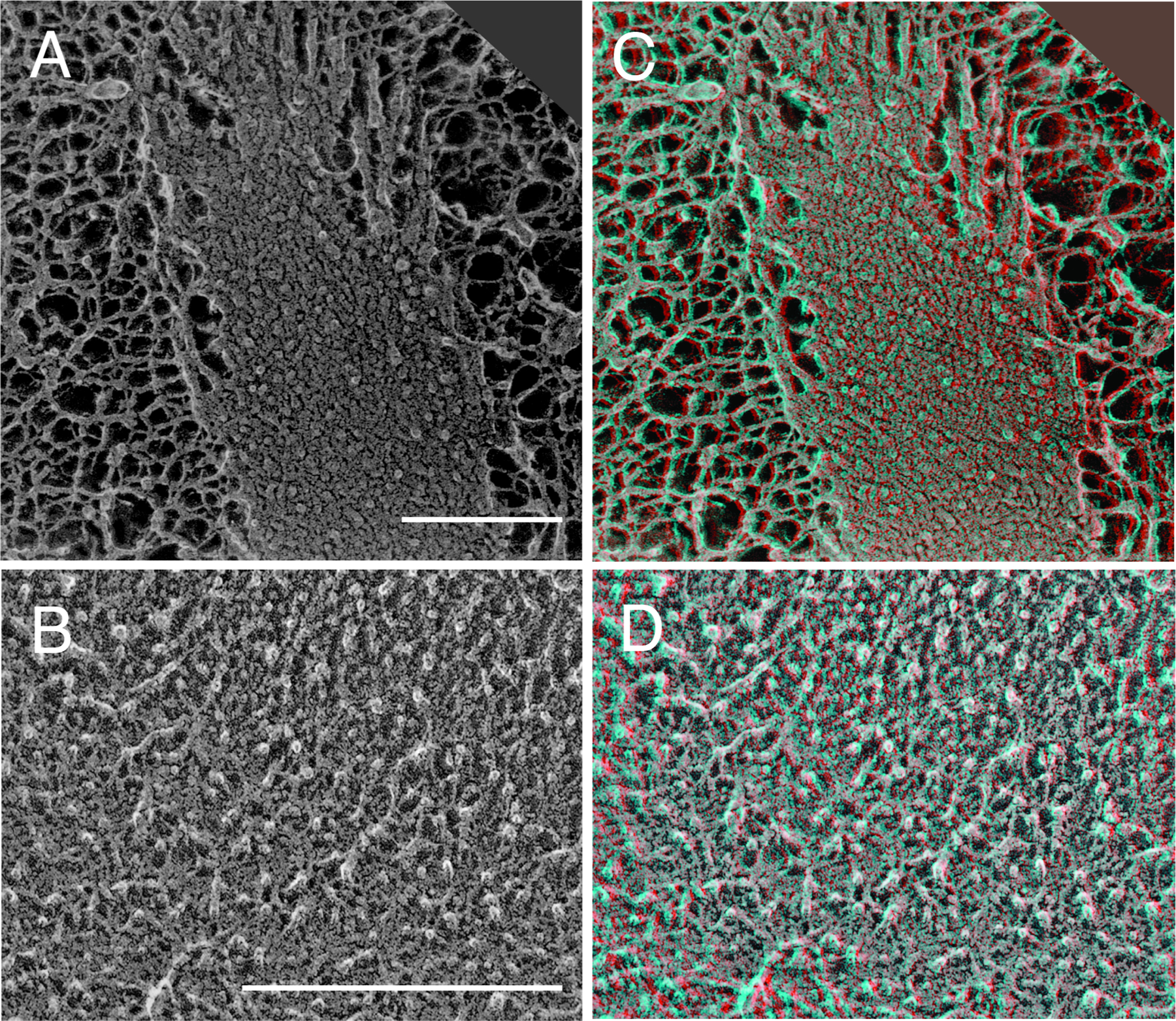
Deep etch electron microscopy images of elastic fibers in developing bovine ligamentum nuchae. A) Image showing elastin substructure where the fracture plane has cut through the elastic fiber. Filaments of 5–7nm make up the fiber structure. Microfibrils (~15 nm in diameter) are evident around the periphery. B) Higher power image showing a relatively disordered arrangement of the filamentous substructure that is so tightly packed that little or no etching occurs. Images C and D are stereo anaglyphs meant to be viewed in 3D using red-green glasses (red lens over the left eye). The greatest detail is obtained when the page is enlarged on the computer screen. A hard copy must be printed in color to retain the 3D perspective. Bar=0.2μm.
The mechanism of tropoelastin assembly during the formation of the mature fiber is still unresolved. As mentioned above, Ross and Bornstein suggested that alignment is a consequence of the interaction of tropoelastin with microfibrils, but new evidence suggests that tropoelastin assembly may begin in the absence of microfibrils. Tropoelastin, like collagen, contains in its primary sequence the information required for higher-order assembly. This assembly process is directed by the hydrophobic clusters within the protein that interact under physiological conditions to form a phase-separated coacervate [40–42]. Detailed studies suggest that during coacervation, tropoelastin rapidly self-associates in ways that facilitate ordered assembly by aligning lysine-containing domains for crosslinking by lysyl oxidase (LOX) [43, 44]. The central region, including sequences around exons 16–17 and exon 30 of tropoelastin, is of particular importance in the coacervation-assembly process [45–47].
Tropoelastin synthesis and trafficking
Cells have sophisticated mechanisms for regulating protein assembly and for transporting cargo from one location to another. Some ECM proteins, such as fibronectin, are secreted through the constitutive pathway and assemble on the cell surface with the assistance of integrins [48]. Fibrillar collagens, in contrast, self-assemble into triple-helical units within the endoplasmic reticulum (ER), are transported in unique vesicles to the Golgi [49], and then organize into larger fibers at or within specialized areas of the plasma membrane [50]. Initiation of basement membrane assembly requires a direct interaction between laminin and the plasma membrane [51, 52] and involves targeted laminin delivery to a specific membrane area. For elastin, the assembly pathway is less well characterized. Tropoelastin, like collagen, will self-assemble under physiological conditions [40], so the cell must control the rate, location, and extent of assembly to prevent the polymer from becoming too large to transport through the cell. Collagen fibril assembly is controlled by propeptides that prevent fibril formation until removal of the propeptides outside the cell. Tropoelastin lacks propeptides, so some other mechanism must regulate assembly.
Tropoelastin synthesis begins in the endoplasmic reticulum (ER), but its secretion follows an unconventional pathway involving trafficking to/through an acidic compartment before transport to the cell surface [53, 54]. Although the identity of the acidic compartment is unknown, its properties are consistent with it being an endosomal/lysosome-related organelle. These unique intracellular structures form a group of functionally diverse compartments that share features with lysosomes [55, 56]. They are, however, distinct and contain specific cargoes that usually derive from the endosomal system [57, 58].
While in the ER, tropoelastin interacts with several molecular chaperones that assist with protein folding [54, 59]. One such protein is BIP, also known as GRP-78 or heat shock 70 kDa protein 5 (HSPA5). BIP’s role is to bind newly synthesized proteins in the ER, so its association with tropoelastin is not surprising, especially given tropoelastin’s hydrophobicity. A second ER protein found to bind tropoelastin is FKBP65, an endoplasmic reticulum-resident peptidyl prolyl cis-trans isomerase (PPIase). A role for a proline isomerase in the folding of tropoelastin is attractive, based on the fact that ~12% of its amino acid residues are proline. Most prolines in tropoelastin are in beta-turns that are stabilized by a hydrogen bond that can only form if the proline is in the trans-configuration. A cis-proline could disrupt the beta-turn substructure and prevent proper folding or alter the protein’s elastomeric properties, so ensuring that all prolines are in the trans configuration is a crucial step in tropoelastin synthesis. Interestingly, Miao et al. [60] found that FKBP65 modified the kinetics of tropoelastin self-assembly in an in vitro system, consistent with a role in attenuating the premature intracellular self-aggregation of tropoelastin through a mechanism that limits the growth and maturation of aggregates.
Tropoelastin preassembly (microassembly)
Dynamic imaging of elastic fiber assembly in cultured cells has added significantly to our understanding of the assembly process. Studies using cells transfected with fluorescently labeled tropoelastin constructs found that the first step in elastic fiber formation is the appearance of small cell surface-associated elastin globules that increase in size with time [61, 62]. Using a tropoelastin fusion protein engineered with a timer construct that acts as a biological clock by changing color over time, Kozel et al. [61] found that elastin-containing globules are first detected inside the cell but rapidly become associated with the cell surface during early phases of assembly. With time, the cell surface globules frequently coalesce into larger globules until released into the ECM. The color changes associated with the timer construct suggested that tropoelastin is continuously deposited onto older tropoelastin while the globule is associated with the cell. The elastin globules eventually transfer to pre-existing elastic fibers in the extracellular matrix, where they coalesce into larger structures [61, 63].
The globular structure of elastin seen in live-cell imaging studies suggests that tropoelastin may self-assemble before it interacts with microfibrils. Newly synthesized elastin initially localized to intracellular vesicles whose distribution, size and movement were not typical of vesicles associated with the constitutive secretory pathway. When delivered to the plasma membrane, the cargo retained the globular shape and size of the intracellular vesicle, consistent with stable polymerization of tropoelastin before vesicular release [61]. If elastin is partially assembled and crosslinked inside the vesicle that transports it to the cell surface, then other critical assembly proteins, such as lysyl oxidase, must be there as well [64, 65].
Fiber assembly outside the cell (macroassembly)
Mechanical forces associated with cell movement help shape the forming, extracellular elastic fiber network. Czirok et al. [63] used an antibody pulse-chase labeling approach to demonstrate that tropoelastin globules aggregate hierarchically, creating progressively larger fibrillar structures (macroassembly). During the assembly process, increasingly large segments of elastin are progressively connected. The aggregation process does not only create large structures, but it also results in a fiber-like, linear distribution of the constituents that can substantially elongate. The end-to-end distance within an aggregate can exhibit both reversible and permanent changes. In the case of reversible length fluctuations, the original state (length, thickness) of the fiber is restored within a few hours, even after a relative extension of 50%. The elongation of the fibers always appears to be coupled to a decrease in fiber diameter. Mapping cell and elastin displacement found that the direction of adjacent elastic fibers and cell displacement vectors were similar. As the culture matured over time, the ECM motion vectors were similar throughout the entire field, yielding additional support to the observation that matured elastin-containing ECM aggregates move as a unit. Thus, both the aggregation process and shaping the aggregates into fibrillar form is coupled to cell motion. Cells move, and thereby extend and align a network of microfibers, as well as interact with adjacent elastin-containing aggregates. The motion of non-adjacent cells became more coordinated as the physical size of elastin-containing aggregates increased, implying that the formation of elastic fibers involves the concerted action and motility of multiple cells. Explaining the mechanical coupling between elastin-containing ECM and the correlated sheet motion of cells will be vital to understanding elastin assembly in tissues.
Role of microfibrils
Tropoelastin interacts with numerous protein components of microfibrils, including fibrillin [66, 67], fibulins [68], and MAGP1 [69–71], so a direct role for microfibrils in tropoelastin assembly cannot be discounted. Mice deficient in fibrillin-1 and fibrillin-2, the backbone proteins of microfibrils, show impaired elastogenesis [72], and mutations in genes for other microfibril-associated proteins also disrupt elastin assembly [73]. Nevertheless, it is not clear whether the earliest stages of elastin assembly require microfibrils. Because tropoelastin undergoes a high degree of self-assembly, interactions with microfibrils may function to control fiber assembly by directing and mediating the coalescence of elastin aggregates outside the cell. Microfibrils could also influence elastin assembly by interacting with signaling molecules on the cell surface and thereby provide a target for tropoelastin secretion. There are many possible roles for microfibrils other than serving as an inert scaffold in the formation of an elastic fiber, but what these roles might be requires further investigation.
Influence of proteoglycans on elastin assembly
There is a long history and an extensive literature of glycosaminoglycans and proteoglycans influencing elastin production, assembly, and fiber stability. In both developing tissues [74] and mature elastin fibers [75, 76], proteoglycan components are associated with elastin. Studies have also found that proteoglycans induce conformational changes of alpha-elastin [77] through ionic and hydrophobic bonds and that these polyanions favor tropoelastin [78] and alpha-elastin fibril formation [79].
Interactions with negatively-charged proteoglycans on the cell surface may facilitate tropoelastin self-assembly by neutralizing the positive charge within the lysine-containing crosslinking domains, thereby reducing repulsive interactions so that the domains can be brought together for cross-link formation [78]. Proteoglycans may also participate in tropoelastin alignment preparatory to crosslinking through an interaction with the C-terminus of tropoelastin encoded by exon 36 – a critical and highly conserved assembly domain [80–82]. A consequence of crosslinking is a loss of positive charge on the molecule due to oxidative deamination of the lysyl ε-amino group. Hence, as crosslinking progresses, the charge on the tropoelastin aggregate is reduced such that the ionic interactions between it and negatively charged cell-associated proteoglycans diminish. This reduction of charge could facilitate the release of tropoelastin from the cell and, by neutralizing repulsive charges, assist globular fusion in the presence of microfibrils. Glycosaminoglycans may also facilitate the release of tropoelastin from a receptor/chaperone protein [83]. Two particularly important proteoglycans in this regard are versican and biglycan. Versican is a chondroitin sulfate proteoglycan found at high levels in elastin-rich tissues, such as lung and aorta (reviewed by Wight [84]). Versican forms large multimolecular complexes with hyaluronan and overexpression of versican V3, an alternatively spliced variant that lacks chondroitin sulfate chains, leads to increased tropoelastin gene expression and assembly of elastic fibers [85]. Likewise, biglycan, a small chondroitin/dermatan sulfate proteoglycan, promotes elastic fiber expression and assembly in cultured smooth muscle cells and injured arteries in animal models of experimental atherogenesis [86]. Both biglycan and the closely related small leucine-rich proteoglycan decorin bind tropoelastin with high affinity. Interestingly, binding occurs through their core protein rather than their glycosaminoglycan side chains [87].
Proteoglycans also influence tropoelastin association with existing ECM away from the cell surface. Kozel et al. [88] showed that recombinant tropoelastin added to cultured cells associated with elastic fiber components in the absence of live cells. The interaction is mediated by ionic interactions involving the lysine side chains of tropoelastin but does not depend on cross-link formation [66]. The matrix-associated tropoelastin had an initial globular appearance but became more fibrous as the matrix matured. Deposition required the C-terminal region of tropoelastin (exon 36) and correlated with the presence of preexisting elastic fibers or microfibrils. Tropoelastin incorporation was inhibited by the addition of heparan sulfate but not chondroitin sulfate sugars. Together, these results suggest that the matrix elaborated by the cell is sufficient for the initial deposition of tropoelastin in the extracellular space and that the composition of sulfated proteoglycans in the matrix influences subsequent elastin assembly.
Summary
Figure 3 summarizes possible assembly/secretory pathways for tropoelastin based on information outlined in this review. Pathway 1 is the currently-accepted model where tropoelastin, secreted as an uncrosslinked monomer through the constitutive secretory pathway, utilizes microfibrils for domain alignment before crosslinking by extracellular lysyl oxidase. Pathway 2 is similar to pathway 1, except tropoelastin is partially crosslinked by lysyl oxidase on the cell surface or outside the cell before interacting with microfibrils and other fiber building components. In pathway 3, tropoelastin is trafficked from the Golgi into an endosomal-like compartment along with lysyl oxidase and perhaps other essential assembly proteins like fibulin-4. Membrane-bound vesicles containing partially cross-linked tropoelastin are then secreted where they and their cargo associate with microfibrils and undergo further interactions to form the final fiber. Whichever pathway proves to be relevant, it is clear that tropoelastin secretion and assembly is complicated, and we have much more to learn.
Figure 3.
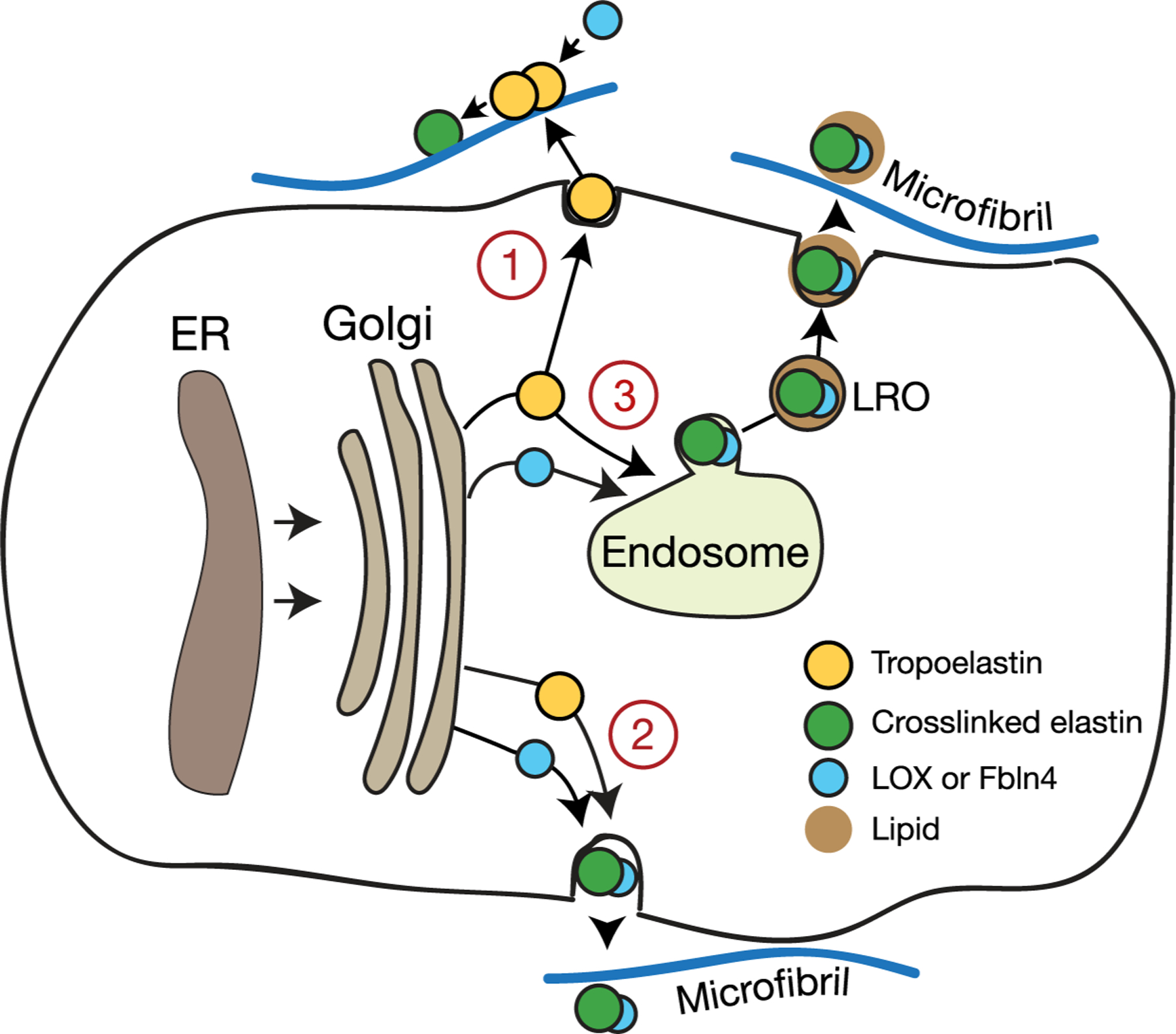
A summary of possible assembly/secretory pathways for tropoelastin based on information outlined in this review. In Pathway 1, tropoelastin is secreted through the constitutive pathway and microfibrils in the extracellular space provide a physical scaffold for tropoelastin assembly and subsequent crosslinking by lysyl oxidase. Pathway 2 is similar to pathway 1, except tropoelastin is partially crosslinked by lysyl oxidase on the cell surface or outside the cell before interacting with microfibrils. Pathway 3 proposes a mechanism where tropoelastin assembly and crosslinking occur within a membrane-limited intracellular compartment similar to an endosome or a lysosome-related organelle. Vesicle with cross-linked elastin are then secreted, associate with microfibrils, and undergo further interactions to form the final fiber.
The field of elastin biology has come a long way from the early days of boiling sodium hydroxide, guanidine extracts, and unidentifiable band on gels. Many of the elastic fiber “structural glycoproteins” initially explored by Leslie Robert and colleagues are now known. What is left is to identify their function and complete the story.
Highlights.
Elastic fibers are complex structures consisting of microfibrils and polymerized elastin.
Microfibrils are necessary for elastic fiber formation, but their exact role is unknown.
Recent evidence suggests that the initial stages of tropoelastin assembly occur within the cell or at unique assembly sites on the plasma membrane where tropoelastin self assembles to form elastin aggregates.
Microfibrils may serve to facilitate the fusion of elastin aggregates outside the cell by mediate tensional forces associated with cell movement.
These observations challenge the widely held idea that interactions between monomeric tropoelastin and microfibrils is the necessary first step in elastin polymerization.
Acknowledgements
The original work reported in this review was support by National Institutes of Health grants HL-53325 and HL-105314. Funds were also provided by the Ines Mandl Research Foundation, New York, New York. We thank Dr. John Heuser and Robyn Roth for the electron microscopy images.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Declarations of interest: none
References
- [1].Partridge SM, Elastin, Adv. Prot. Chem 17 (1962) 227–302. [Google Scholar]
- [2].Gotte L, Stern P, Elsden DF, Partridge SM, The chemistry of connective tissues. 8. The composition of elastin from three bovine tissues, Biochem. J 87 (1963) 344–351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Ross R, Bornstein P, The elastic fiber. I. The separation and partial characterization of its macromolecular components., J Cell Biol 40(2) (1969) 366–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Ross R, The elastic fiber, J. Histochem. Cytochem 21 (1973) 199–208. [DOI] [PubMed] [Google Scholar]
- [5].Robert B, Szigeti M, Derouette JC, Robert L, Studies on the nature of the “microfibrillar” component of elastic fibers, Eur J Biochem 21(4) (1971) 507–16. [DOI] [PubMed] [Google Scholar]
- [6].Moczar M, Moczar E, Robert L, Peptides obtained from elastin by hydrolysis with aqueous ethanolic potassium hydroxide., Connect Tissue Res 6(4) (1979) 207–13. [DOI] [PubMed] [Google Scholar]
- [7].Moczar M, Moczar E, Robert L, Composition of glycopeptides obtained by proteolytic digestion of the media of porcine aorta, Atherosclerosis 12(1) (1970) 31–40. [DOI] [PubMed] [Google Scholar]
- [8].Bach PR, Bentley JP, Structural glycoprotein, fact or artefact, Connect Tissue Res 7(3) (1980) 185–96. [DOI] [PubMed] [Google Scholar]
- [9].Haust MD, Fine fibrils of extracellular space (microfibrils). Their structure and role in connective tissue organization, Am J Pathol 47(6) (1965) 1113–37. [PMC free article] [PubMed] [Google Scholar]
- [10].Karrer HE, An electron microscope study of the aorta in young and aging mice, J. Ultrastruct. Res 5 (1961) 1–27. [DOI] [PubMed] [Google Scholar]
- [11].Fahrenbach WH, Sandberg LB, Cleary EG, Ultrastructural studies on early elastogenesis, Anat. Rec 155 (1966) 563–576. [Google Scholar]
- [12].Keeley FW, The Evolution of Elastin, in: Keeley FW, Mecham RP (Eds.), Evolution of Extracellular Matrix, Springer-Verlag, Berlin Heidelberg, 2013, pp. 73–119. [Google Scholar]
- [13].Reichheld SE, Muiznieks LD, Lu R, Sharpe S, Keeley FW, Sequence variants of human tropoelastin affecting assembly, structural characteristics and functional properties of polymeric elastin in health and disease, Matrix Biol (2019). [DOI] [PubMed] [Google Scholar]
- [14].Kozel BA, mecham RP, Rosenbloom J, Elastin, in: Mecham RP (Ed.), Biology of Extracellular Matrix, Springer-Verlag, Berlin-Heidelberg, 2011, pp. 267–301. [Google Scholar]
- [15].Schrader CU, Heinz A, Majovsky P, Karaman B, Brinckmann J, Sippl W, Schmelzer CEH, Elastin is heterogeneously cross-linked, J Biol Chem (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Mithieux SM, Wise SG, Raftery MJ, Starcher B, Weiss AS, A model two-component system for studying the architecture of elastin assembly in vitro, J. Struct. Biol 149(3) (2005) 282–289. [DOI] [PubMed] [Google Scholar]
- [17].Vindin H, Mithieux SM, Weiss AS, Elastin architecture, Matrix Biol (2019). [DOI] [PubMed] [Google Scholar]
- [18].Gotte L, Volpin D, Horne RW, Mammi M, Electron microscopy and optical diffraction of elastin, Micron 7 (1976) 95–102. [Google Scholar]
- [19].Cleary E, Cliff W, The substructure of elastin., Exp Mol Pathol 28(2) (1978) 227–46. [DOI] [PubMed] [Google Scholar]
- [20].Serafini-Fracassini A, Field J, Hinnie J, The primary filament of bovine elastin., J Ultrastruct Res 65(2) (1978) 190–3. [DOI] [PubMed] [Google Scholar]
- [21].Serafini-Fracassini A, Field J, Spina M, The macromolecular organization of the elastin fibril., J Mol Biol 100(1) (1976) 73–84. [DOI] [PubMed] [Google Scholar]
- [22].Bressan GM, Castellani I, Giro MG, Volpin D, Fornieri C, Pasquali-Ronchetti I, Banded fibers in tropoelastin coacervates at physiological temperatures, J. Ultrastruc. Res 82 (1983) 335–340. [DOI] [PubMed] [Google Scholar]
- [23].Bressan GM, Pasquali-Ronchetti I, Fornieri C, Mattioli F, Castellani I, Volpin D, Relevance of aggregation properties of tropoelastin to the assembly and structure of elastic fibers, J. Ultrastruct. Mol. Struct. Res 94(3) (1986) 209–16. [DOI] [PubMed] [Google Scholar]
- [24].Cox B, Starcher B, Urry D, Coacervation of alpha-elastin results in fiber formation., Biochim Biophys Acta 317(1) (1973) 209–13. [DOI] [PubMed] [Google Scholar]
- [25].Volpin D, Pasquali-Ronchetti I, Urry D, Gotte L, Banded figers in high temperature coacervates of elastin peptides., J Biol Chem 251(21) (1976) 6871–3. [PubMed] [Google Scholar]
- [26].Volpin D, Urry DW, Pasquali Ronchetti I, Gotte L, Studies by electron microscopy on the structure of coacervates of synthetic polypeptides of tropoelastin, Micron 7 (1976) 193–198. [Google Scholar]
- [27].Volpin D, Pasquali-Ronchetti I, The ultrastructure of high-temperature coacervates from elastin, J. Ultra. Res 61 (1977) 295–302. [DOI] [PubMed] [Google Scholar]
- [28].Volpin D, Pasquali-Ronchetti I, The ultrastruct of high-temperature coacervates from elastin, J Ultrastruct Res 61(3) (1977) 295–302. [DOI] [PubMed] [Google Scholar]
- [29].Pasquali-Ronchetti I, Fornieri C, Baccarani-Contri M, Volpin D, The ultrastructure of elastin revealed by freeze-fracture electron microscopy, Micron 10 (1979) 89–99. [Google Scholar]
- [30].Fornieri C, Ronchetti I, Edman A, Sjostrom M, Contribution of cryotechniques to the study of elastin ultrastructure., J Microsc 126(Pt 1) (1982) 87–93. [DOI] [PubMed] [Google Scholar]
- [31].Nakamura H, Ooyama T, Ultrastructure of elastic fibers in the rat aorta as revealed by the deep-etching technique., J Electron Microsc (Tokyo) 37(3) (1988) 167–70. [PubMed] [Google Scholar]
- [32].Mecham R, Heuser J, Three-dimensional organization of extracellular matrix in elastic cartilage as viewed by quick freeze, deep etch electron microscopy., Connect. Tissue Res 24(2) (1990) 83–93. [DOI] [PubMed] [Google Scholar]
- [33].Serafini-Fracassini A, Field JM, X-ray analysis of enzymically purified elastin from bovine ligamentum nuchae, Adv Exp Med Biol 79 (1977) 679–83. [DOI] [PubMed] [Google Scholar]
- [34].Pasquali Ronchetti I, Alessandrini A, Baccarani Contri M, Fornieri C, Mori G, Quaglino DJ, Valdre U, Study of elastic fiber organization by scanning force microscopy., Matrix Biol 17(1) (1998) 75–83. [DOI] [PubMed] [Google Scholar]
- [35].Morocutti M, Raspanti M, Govoni P, Kadar A, Ruggeri A, Ultrastructural aspects of freeze-fractured and etched elastin., Connect Tissue Res 18(1) (1988) 55–64. [DOI] [PubMed] [Google Scholar]
- [36].Mecham RP, Heuser JE, The elastic fiber, in: Hay ED (Ed.), Cell Biology of Extracellular Matrix, 2nd Edition, Plenum Press, Inc., New York, 1991, pp. 79–109. [Google Scholar]
- [37].Heuser J, The origins and evolution of freeze-etch electron microscopy., J Electron Microsc (Tokyo) 60 Suppl 1 (2011) S3–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Heuser JE, Quick-freeze, deep etch preparation of samples for 3-D electron microscopy, Trends Biochem. Sci 24(12) (1981) 64–68. [Google Scholar]
- [39].Heuser J, Preparing biological samples for stereomicroscopy by the quick-freeze, deep-etch, rotary-replication technique., Methods Cell Biol. 22 (1981) 97–122. [DOI] [PubMed] [Google Scholar]
- [40].Cox BA, Starcher BC, Urry DW, Coacervation of tropoelastin results in fiber formation, J. Biol. Chem 249 (1974) 997–998. [PubMed] [Google Scholar]
- [41].Vrhovski B, Jensen S, Weiss AS, Coacervation characteristics of recombinant human tropoelastin, Eur. J. Biochem 250 (1997) 92–98. [DOI] [PubMed] [Google Scholar]
- [42].Bellingham C, Lillie M, Gosline J, Wright G, Starcher B, Bailey A, Woodhouse K, Keeley F, Recombinant human elastin polypeptides self-assemble into biomaterials with elastin-like properties., Biopolymers 70(4) (2003) 445–455. [DOI] [PubMed] [Google Scholar]
- [43].Clarke AW, Arnspang EC, Mithieux SM, Korkmaz E, Braet F, Weiss AS, Tropoelastin massively associates during coacervation to form quantized protein spheres, Biochemistry 45(33) (2006) 9989–9996. [DOI] [PubMed] [Google Scholar]
- [44].Muiznieks L, Cirulis J, van der Horst A, Reinhardt D, Wuite G, Pomes R, Keeley F, Modulated growth, stability and interactions of liquid-like coacervate assemblies of elastin., Matrix Biol (2014). [DOI] [PubMed] [Google Scholar]
- [45].Wachi H, Sato F, Nakazawa J, Nonaka R, Szabo Z, Urban Z, Yasunaga T, Maeda I, Okamoto K, Starcher BC, Li DY, Mecham RP, Seyama Y, Domains 16 and 17 of tropoelastin in elastic fibre formation, Biochem. J 402(1) (2007) 63–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Dyksterhuis L, Baldock C, Lammie D, Wess T, Weiss A, Domains 17–27 of tropoelastin contain key regions of contact for coacervation and contain an unusual turn-containing crosslinking domain., Matrix Biol. 26(2) (2007) 125–135. [DOI] [PubMed] [Google Scholar]
- [47].Wise SG, Mithieux SM, Raftery MJ, Weiss AS, Specificity in the coacervation of tropoelastin: solvent exposed lysines, J. Struct. Biol 149(3) (2005) 273–281. [DOI] [PubMed] [Google Scholar]
- [48].Schwarzbauer JE, DeSimone DW, Fibronectins, their fibrillogenesis, and in vivo functions, Cold Spring Harb Perspect Biol 3(7) (2011) 3:a005041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Wilson D, Phamluong K, Li L, Sun M, Cao T, Liu P, Modrusan Z, Sandoval W, Rangell L, Carano R, Peterson A, Solloway M, Global defects in collagen secretion in a Mia3/TANGO1 knockout mouse., J Cell Biol 193(5) (2011) 935–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Kalson N, Starborg T, Lu Y, Mironov A, Humphries S, Holmes D, Kadler K, Nonmuscle myosin II powered transport of newly formed collagen fibrils at the plasma membrane., Proc Natl Acad Sci U S A 110(49) (2013) E4743–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Hohenester E, Yurchenco P, Laminins in basement membrane assembly., Cell Adh Migr 7(1) (2013) 56–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Yurchenco PD, Amenta PS, Patton BL, Basement membrane assembly, stability and activities observed through a developmental lens, Matrix Biol. 22(7) (2004) 521–38. [DOI] [PubMed] [Google Scholar]
- [53].Davis E, Mecham R, Intracellular trafficking of tropoelastin., Matrix Biol. 17(4) (1998) 245–254. [DOI] [PubMed] [Google Scholar]
- [54].Davis E, Broekelmann T, Ozawa Y, Mecham R, Identification of tropoelastin as a ligand for the 65-kD FK506-binding protein, FKBP65, in the secretory pathway., J. Cell Biol 140(2) (1998) 295–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Marks M, Heijnen H, Raposo G, Lysosome-related organelles: unusual compartments become mainstream., Curr Opin Cell Biol 25(4) (2013) 495–505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Delevoye C, Marks MS, Raposo G, Lysosome-related organelles as functional adaptations of the endolysosomal system, Curr Opin Cell Biol 59 (2019) 147–158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Raposo G, Stoorvogel W, Extracellular vesicles: exosomes, microvesicles, and friends., J Cell Biol 200(4) (2013) 373–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Bultema J, Di Pietro S, Cell type-specific Rab32 and Rab38 cooperate with the ubiquitous lysosome biogenesis machinery to synthesize specialized lysosome-related organelles., Small GTPases 4(1) (2013) 16–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Patterson C, Schaub T, Coleman E, Davis E, Developmental regulation of FKBP65. An ER-localized extracellular matrix binding-protein., Mol. Biol. Cell 11(11) (2000) 3925–3935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Miao M, Reichheld S, Muiznieks L, Huang Y, Keeley F, Elastin binding protein and FKBP65 modulate in vitro self-assembly of human tropoelastin., Biochemistry 52(44) (2013) 7731–41. [DOI] [PubMed] [Google Scholar]
- [61].Kozel BA, Rongish BJ, Czirok A, Zach J, Little CD, Davis EC, Knutsen RH, Wagenseil JE, Levy MA, Mecham RP, Elastic fiber formation: A dynamic view of extracellular matrix assembly using timer reporters, J. Cell. Physiol 207(1) (2006) 87–96. [DOI] [PubMed] [Google Scholar]
- [62].Wagenseil J, Mecham R, New insights into elastic fiber assembly., Birth Defects Res C Embryo Today 81(4) (2007) 229–40. [DOI] [PubMed] [Google Scholar]
- [63].Czirok A, Zach J, Kozel BA, Mecham RP, Davis EC, Rongish BJ, Elastic fiber macro-assembly is a hierarchical, cell motion-mediated process, J. Cell. Physiol 207 (2006) 97–106. [DOI] [PubMed] [Google Scholar]
- [64].Kagan HM, Vaccaro CA, Bronson RE, Tang SS, Brody JS, Ultrastructural immunolocalization of lysyl oxidase in vascular connective tissue, J. Cell Biol 103 (1986) 1121–1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Sato F, Seino-Sudo R, Okada M, Sakai H, Yumoto T, Wachi H, Lysyl Oxidase Enhances the Deposition of Tropoelastin through the Catalysis of Tropoelastin Molecules on the Cell Surface, Biol Pharm Bull 40(10) (2017) 1646–1653. [DOI] [PubMed] [Google Scholar]
- [66].Trask TM, Crippes Trask B, Ritty TM, Abrams WR, Rosenbloom J, Mecham RP, Interaction of tropoelastin with the amino-terminal domains of fibrillin-1 and fibrillin-2 suggests a role for the fibrillins in elastic fiber assembly, J. Biol. Chem 275 (2000) 24400–24406. [DOI] [PubMed] [Google Scholar]
- [67].Jensen SA, Reinhardt DP, Gibson MA, Weiss AS, Protein interaction studies of MAGP-1 with tropoelastin and fibrillin-1, J. Biol. Chem 276 (2001) 39661–39666. [DOI] [PubMed] [Google Scholar]
- [68].Roark EF, Keene DR, Haudenschild CC, Godyna S, Little CD, Argraves WS, The association of human fibulin-1 with elastic fibers: an immunohistological, ultrastructural, and RNA study, J Histochem Cytochem 43(4) (1995) 401–11. [DOI] [PubMed] [Google Scholar]
- [69].Brown-Augsburger P, Broekelmann T, Mecham L, Mercer R, Gibson MA, Cleary EG, Abrams WR, Rosenbloom J, Mecham RP, Microfibril-associated glycoprotein (MAGP) binds to the carboxy-terminal domain of tropoelastin and is a substrate for transglutaminase, J. Biol. Chem 269 (1994) 28443–28449. [PubMed] [Google Scholar]
- [70].Brown-Augsburger PB, Broekelmann T, Rosenbloom J, Mecham RP, Functional domains on elastin and MAGP involved in elastic fiber assembly, Biochem. J 318 (1996) 149–155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Gibson MA, Hatzinikolas G, Kumaratilake JS, Sandberg LB, Nicholl JK, Sutherland GR, Cleary EG, Further characterization of proteins associated with elastic fiber microfibrils including the molecular cloning of MAGP-2 (MP25), J. Biol. Chem 271 (1996) 1096–1103. [DOI] [PubMed] [Google Scholar]
- [72].Carta L, Pereira L, Arteaga-Soli E, Lee-Arteaga SY, Lenart B, Starcher B, Merkel CA, Sukoyan M, Kerkis A, Hazeki N, Keene DR, Sakai LY, Ramirez F, Fibrillins 1 and 2 perform partially overlapping functions during aortic development, J. Biol. Chem 281 (2006) 8016–8023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].McLaughlin PJ, Chen Q, Horiguchi M, Starcher BC, Stanton JB, Broekelmann TJ, Marmorstein AD, McKay B, Mecham R, Nakamura T, Marmorstein LY, Targeted disruption of fibulin-4 abolishes elastogenesis and causes perinatal lethality in mice, Mol. Cell Biol 26(5) (2006) 1700–1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Kadar A, Gardner DL, Bush V, Glycosaminoglycans in developing chick-embryo aorta revealed by ruthenium red: an electron-microscope study, J Pathol 108(4) (1972) 275–80. [DOI] [PubMed] [Google Scholar]
- [75].Pasquali-Ronchetti I, Bressan GM, Fornieri C, Baccarani-Contri M, Castellani I, Volpin D, Elastin fiber-associated glycosaminoglycans in beta-aminopropionitrile-induced lathyrism, Exp. Molec. Pathol 40 (1984) 235–245. [DOI] [PubMed] [Google Scholar]
- [76].Contri MB, Fornieri CC, Ronchetti IP, Elastin-proteoglycans association revealed by cytochemical methods, Connect. Tissue Res 13(3) (1985) 237–49. [DOI] [PubMed] [Google Scholar]
- [77].Podrazky V, Stokrova S, Fric I, Elastin--proteoglycan interaction. Conformational changes of alpha-elastin induced by the interaction, Connect Tissue Res 4(1) (1975) 51–4. [DOI] [PubMed] [Google Scholar]
- [78].Adam M, Podrazky V, Fibrillation of tropoelastin induced by proteoglycan, Experientia 32(4) (1976) 430–432. [DOI] [PubMed] [Google Scholar]
- [79].Podrazky V, Adam M, Fibrillation of alpha-elastin induced by proteoglycan, Experientia 31(5) (1975) 523–4. [DOI] [PubMed] [Google Scholar]
- [80].Broekelmann TJ, Kozel BA, Ishibashi H, Werneck CC, Keeley FW, Zhang L, Mecham RP, Tropoelastin interacts with cell-surface glycosaminoglycans via its C-terminal domain, J. Biol. Chem 280 (2005) 40939–40947. [DOI] [PubMed] [Google Scholar]
- [81].Kozel BA, Wachi H, Davis EC, Mecham RP, Domains in tropoelastin that mediate elastin deposition in vitro and in vivo, J. Biol. Chem 278 (2003) 18491–18498. [DOI] [PubMed] [Google Scholar]
- [82].Nonaka R, Sato F, Wachi H, Domain 36 of tropoelastin in elastic fiber formation., Biol Pharm Bull 37(4) (2014) 698–702. [DOI] [PubMed] [Google Scholar]
- [83].Privitera S, Prody CA, Callahan JW, Hinek A, The 67-kDa enzymatically inactive alternatively spliced variant of β-galactosidase is identical to the elastin/laminin binding protein, J. Biol. Chem 273 (1998) 6319–6326. [DOI] [PubMed] [Google Scholar]
- [84].Wight TN, A role for proteoglycans in vascular disease, Matrix Biol 71–72 (2018) 396–420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Merrilees MJ, Lemire JM, Fischer JW, Kinsella MG, Braun KR, Clowes AW, Wight TN, Retrovirally mediated overexpression of versican v3 by arterial smooth muscle cells induces tropoelastin synthesis and elastic fiber formation in vitro and in neointima after vascular injury, Circ. Res 90(4) (2002) 481–487. [DOI] [PubMed] [Google Scholar]
- [86].Hwang J, Johnson P, Braun K, Hinek A, Fischer J, O’Brien K, Starcher B, Clowes A, Merrilees M, Wight T, Retrovirally mediated overexpression of glycosaminoglycan-deficient biglycan in arterial smooth muscle cells induces tropoelastin synthesis and elastic fiber formation in vitro and in neointimae after vascular injury., Am J Pathol 173(6) (2008) 1919–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Reinboth B, Hanssen E, Cleary EG, Gibson MA, Molecular interactions of biglycan and decorin with elastic fiber components: biglycan forms a ternary complex with tropoelastin and microfibril-associated glycoprotein 1, J. Biol. Chem 277(6) (2002) 3950–3957. [DOI] [PubMed] [Google Scholar]
- [88].Kozel BA, Ciliberto CH, Mecham RP, Deposition of tropoelastin into the extracellular matrix requires a competent elastic fiber scaffold but not live cells, Matrix Biol. 23 (2004) 23–34. [DOI] [PubMed] [Google Scholar]