

Abstract
Intestinal microbiota is involved in immune response and metabolism of the host. The frequent use of anthelmintic compounds for parasite expulsion causes disturbance to the equine intestinal microbiota. However, most studies were on the effects of such treatment on the intestinal bacterial microbes; none is on the entire microbial community including archaea and eukaryotic and viral community in equine animals. This study is the first to explore the differences of the microbial community composition and structure in Przewalski's horses prior to and following anthelmintic treatment, and to determine the corresponding changes of their functional attributes based on metagenomic sequencing. Results showed that in archaea, the methanogen of Euryarchaeota was the dominant phylum. Under this phylum, anthelmintic treatment increased the Methanobrevibacter genus and decreased the Methanocorpusculum genus and two other dominant archaea species, Methanocorpusculum labreanum and Methanocorpusculum bavaricum. In bacteria, Firmicutes and Bacteroidetes were the dominant phyla. Anthelmintic treatment increased the genera of Clostridium and Eubacterium and decreased those of Bacteroides and Prevotella and dominant bacteria species. These altered genera were associated with immunity and digestion. In eukaryota, anthelmintic treatment also changed the genera related to digestion and substantially decreased the relative abundances of identified species. In virus, anthelmintic treatment increased the genus of unclassified_d__Viruses and decreased those of unclassified_f__Siphoviridae and unclassified_f__Myoviridae. Most of the identified viral species were classified into phage, which were more sensitive to anthelmintic treatment than other viruses. Furthermore, anthelmintic treatment was found to increase the number of pathogens related to some clinical diseases in horses. The COG and KEGG function analysis showed that the intestinal microbiota of Przewalski's horse mainly participated in the carbohydrate and amino acid metabolism. The anthelmintic treatment did not change their overall function; however, it displaced the population of the functional microbes involved in each function or pathway. These results provide a complete view on the changes caused by anthelmintic treatment in the intestinal microbiota of the Przewalski's horses.
Keywords: przewalski's horse, anthelmintic treatment, intestinal microbiota, metagenomic sequencing, functional annotation
Introduction
Gasterophilus spp. (horse botfly) is a common parasite in equids (1, 2). Their eggs and larvae can survive in the digestive system (e.g., stomach and intestine) of a host for 8–10 months (3, 4). Infection of horse botfly can cause serious clinical diseases, such as dysphagia, gastric and intestinal ulceration, gastric obstruction, and volvulus; it could even lead to severe risks of anemia, diarrhea, gastric rupture, peritonitis, perforating ulcers, and other complications (5–7). The horse botfly epidemic has been serious in the desert steppe of Xinjiang, China, with six species including G. haemorrhoidalis, G. inermis, G. intestinalis, G. nasalis, G. nigricornis, and G. pecorum commonly found in the local equids (8, 9). In botfly infection in Przewalski's horses, there is particular severity with a 100% infection rate, and an infection level much higher than other equine animals (10). At present, deworming is performed annually in winter through administration of anthelmintic compounds to control the infestation (9). However, the frequent use of these drugs will inevitably lead to drug resistance of parasites, which have been widely reported (11–13). It could also disturb the balance of the intestinal microbial community after removal of the parasites (14).
The equine gut hosts a complex microbial ecosystem with a variety of commensal, symbiotic, and pathogenic microbes. Disturbances to the normal intestinal microbiota could exert critical impact on the host's physiology. In horses, some disturbances are found related with colic (15, 16), diarrhea (17), obesity (18), and other clinical diseases. It is known that many factors such as nutrition and management, medication, age, disease, stress, and gender can influence equine intestinal microbiota (19, 20). However, studies on the effects of anthelmintic treatment for parasite expulsion on the intestinal microbiota of horses are still limited. Goachet et al. (21) were the first to report a reduction in cellulolytic bacteria and an increase in Lactobacilli and Streptococci in horses after anthelmintic treatment. Peachey et al. (22) found that bacterial phylum TM7 was reduced 14 days after anthelmintic treatment, while Adlercreutzia spp. were increased only 2 days after. Crotch-Harvey et al. (23) detected temporal differences of the bacterial community when horses were treated with anthelmintic drugs. Walshe et al. (24) found that the alpha and beta diversity of the bacterial community decreased at day 7 of post-anthelmintic treatment and reverted on day 14. Peachey et al. (25) confirmed again that anthelmintic treatment was associated with alteration of the relative abundances of the bacterial community. Daniels et al. (26) observed that anthelmintic treatment increased the relative abundances of Deferribacter spp. and Spirochaetes spp. For Przewalski's horses, Hu et al. (4) found that the removal of horse botflies through anthelmintic treatment decreased the alpha diversity of the gut bacterial community, increased its Firmicutes to Bacteroidetes (F/B) ratio, and increased the genera of Streptococcus and Lactobacillus and some pathogenic bacteria. Nonetheless, these studies only explored the changes of the intestinal bacterial community associated with anthelmintic treatments. There is no study available on the changes of the entire microbial community and its functional prediction due to anthelmintic treatment. In fact, fungi, viruses, and some other species also play vital roles in the physiology and immune system of the host (27); e.g., fungi of Aspergillus, Candida, Fusarium, Penicillium, and Saccharomyces and archaea of methanogens represent notable members of the intestinal microbiota (28, 29). Hu et al. (30) suggested that anthelmintic treatments on Przewalski's horses could impact the fungal communities even more than the bacterial communities. Therefore, metagenomic sequencing is used in this study to characterize the entire intestinal microbial community (archaea, bacteria, eukaryota, and virus) of Przewalski's horses prior to and following anthelmintic treatments (ivermectin), so as to identify the changes in microbial diversity and richness, as well as genes, functions, and metabolism pathways. The results will lead to a better understanding on the relationships between anthelmintic treatment and the equine intestinal microbiota.
Materials and Methods
Ethics Statement
This study was carried out in accordance with the recommendations of the Institute of Animal Care and the Ethics Committee of Beijing Forestry University. The Ethics Committee of Beijing Forestry University approved the experimental protocol. The management authority of the Kalamaili Nature Reserve (KNR) in Xinjiang approved the collection of Przewalski's horse fecal samples.
DNA Extraction and Metagenomic Sequencing
In a previous study of the present research group (4), fecal samples of seven adult Przewalski's horses (four male, three female) of similar body weight in the KNR prior to (PATPH) and following (FATPH) anthelmintic treatment of ivermectin were collected for DNA extraction and 16S rRNA sequencing. The numbers of horse botfly larvae in the fecal samples of the FATPHs were also recorded for assessing their parasitic infection status before treatment. In this study, shotgun metagenomic sequencing was conducted on the same DNA extracts used for 16S rRNA sequencing in Hu et al. (4). Out of the seven pairs of DNA samples, three pairs were chosen for this study based on their corresponding high to low and in between total fecal larva counts (FATPH3: 2,966, FATPH6: 1,928, FATPH1: 724). The concentration and purity of these six DNA samples (PATPH3, PATPH6, PATPH1; FATPH3, FATPH6, FATPH1) were tested by TBS-380 and NanoDrop 2000, respectively. DNA extract quality was checked with 1% agarose gel.
The DNA extract was fragmented to an average size of about 400 bp using Covaris M220 (Gene Company Limited, Beijing, China) for paired-end library construction with NEXTFLEX Rapid DNA-Seq (Bioo Scientific, Austin, TX, USA). Adapters containing the full complement of sequencing primer hybridization sites were ligated to the blunt end of fragments. Paired-end sequencing was performed on an Illumina sequencing platform at Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China) according to the manufacturer's instructions (www.illumina.com). Sequence data associated with this study have been deposited in the NCBI Short Read Archive database (BioProject ID: PRJNA722063).
Sequence Quality Control and Genome Assembly
Data were analyzed on the free online Majorbio Cloud Platform (www.majorbio.com). The paired-end Illumina reads were trimmed of adaptors. Low-quality reads (length <50 bp or with a quality value <20 or having N bases) were removed by fastp (31) (https://github.com/OpenGene/fastp, version 0.20.0). Reads were aligned to the Przewalski's horse genome (GenBank accession no. GCA_000696695.1) by BWA (32) (http://bio-bwa.sourceforge.net, version 0.7.9a). Any hit associated with the reads and their mated reads were removed. Metagenomics data were assembled using MEGAHIT (33) (https://github.com/voutcn/megahit, version 1.1.2), which makes use of succinct de Bruijn graphs. Contigs with the length being or over 300 bp were selected as the final assembling results; they then were used for further gene prediction and annotation.
Gene Prediction, Taxonomy, and Functional Annotation
Open reading frames (ORFs) from each assembled contig were predicted using MetaGene (34) (http://metagene.cb.k.u-tokyo.ac.jp/). The predicted ORFs with length 100 bp and over were retrieved and translated into amino acid sequences using the NCBI translation table (http://www.ncbi.nlm.nih.gov/Taxonomy/taxonomyhome.html/index.cgi?chapter=tgencodes#SG1). A nonredundant gene catalog was constructed using CD-HIT (35) (http://www.bioinformatics.org/cd-hit/, version 4.6.1) with 90% sequence identity and 90% coverage. Reads after quality control were mapped to the nonredundant gene catalog with 95% identity using SOAPaligner (36) (http://soap.genomics.org.cn/, version 2.21), and gene abundances in each sample were evaluated.
Representative sequences of nonredundant gene catalog were aligned to the NCBI NR database with an e-value cutoff of 1e−5 using Diamond (37) (http://www.diamondsearch.org/index.php, version 0.8.35) for taxonomic annotations. Cluster of orthologous groups of proteins (COG) annotation for the representative sequences was performed using Diamond against the eggNOG database with an e-value cutoff of 1e−5. The KEGG annotation was conducted using Diamond against the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (http://www.genome.jp/keeg/) with an e-value cutoff of 1e−5. The pathogens were predicted with the pathogen–host interactions (PHI) database (http://www.phi-base.org/, version 4.4).
Statistical Analysis
All data were checked for normality. The Wilcoxon rank-sum test in STAMP was used to seek for significant differences between groups, and the p-value was tested by Bonferroni correction. The linear discriminant analysis (LDA) effect size (LEfSe) method was used to identify bacterial taxa with significant difference among groups (http://huttenhower.sph.harvard.edu/galaxy/root?tool_id=lefse_upload). Principal component analysis (PCA) was calculated using weighted UniFrac distance metric in R software.
Results and Discussions
Anthelmintic compounds are widely used in the equine populations because of their common parasitic infections. Anthelmintic treatment will gravely harm the animal's health if left uncontrolled. Previous studies showed that the composition and structure of the intestinal microbial community in horses could be changed upon the treatment of anthelmintics (23–25, 38). However, these studies lacked a control group of horses that were free of parasites, hence making it difficult to identify if the changes were due to the administered anthelmintics or due to the removal of parasites. Then, Kunz et al. (14) conducted an experiment to investigate how the intestinal microbes of uninfected horses changed under the administration of anthelmintic compounds. Their results did not show large-scale changes in the intestinal microbial community observed in infected horses treated with anthelmintics. Thus, changes in intestinal microbes of horses following anthelmintic treatment are mainly associated with the removal of parasites.
Horse botflies are the main concern for the wild Przewalski's horses in Xinjiang. Ivermectin is administered to the horses once a year in winter to control their parasitic infestation. Hence, understanding how these regular horse botfly expulsion treatments would impact the intestinal microbiota of the horses becomes important, as the microbiota plays an important role in influencing the host metabolism, immunity, speciation, and many other functions (39–44). However, previous studies were limited to studying only the bacterial community based on 16S rRNA sequencing, thus lacking information on other members such as archaea, fungi, and virus of the microbiota, and analysis of its functions.
Differences in Composition of Intestinal Microbial Community in FATPHs and PATPHs
After quality control, a total of 437,143,618 reads (FATPHs: 222,498,384; PATPHs: 214,645,234) and 3,738,967 contigs (FATPHs: 1,932,396; PATPHs: 1,806,571) were obtained from all samples with an insert size of 565 bp and length of 150 bp (Table 1). The average values of N50 and N90 were 697 ± 73 bp and 350 ± 6 bp, respectively (Table 1). When comparing the reads with the database by the best-hit annotation method, five domains including archaea, bacteria, eukaryota, virus, and unclassified were identified. It was shown that most of the reads were annotated with bacteria, which accounted for 97.79% in FATPHs and 96.76% in PATPHs. Meanwhile, archaea, eukaryota, and virus occupied, respectively 1.07, 0.46, and 0.59% in FATPHs, and, respectively 0.87, 1.85, and 0.30% in PATPHs. Classification of the domains into each taxonomic unit showed that FATPHs had 8 kingdoms, 134 phyla, 263 classes, 579 orders, 1,052 families, 2,775 genera, and 12,677 species and correspondingly PATPHs had 8, 134, 267, 591, 1,081, 2,824, and 12,729, respectively. Both FATPHs and PATPHs had the same numbers of domains, kingdom, and phylum. Anthelmintic treatment reduced the microbial richness below the class level.
Table 1.
Sequencing information of the six samples.
| Samples | Raw reads | Raw base (bp) | Contigs | Contigs bases (bp) | N50 (bp) | N90 (bp) |
|---|---|---|---|---|---|---|
| FATPH1 | 69,471,868 | 10,490,252,068 | 548,642 | 376,741,716 | 721 | 347 |
| FATPH3 | 72,653,294 | 10,970,647,394 | 682,192 | 474,946,359 | 734 | 353 |
| FATPH6 | 72,520,072 | 10,950,530,872 | 701,562 | 513,945,911 | 791 | 358 |
| PATPH1 | 74,061,672 | 11,183,312,472 | 552,710 | 358,478,060 | 668 | 345 |
| PATPH3 | 70,389,906 | 10,628,875,806 | 624,273 | 362,638,296 | 574 | 342 |
| PATPH6 | 78,046,806 | 11,785,067,706 | 629,588 | 426,246,469 | 692 | 353 |
Archaea
In archaea, the dominant phylum was Euryarchaeota in both FATPHs and PATPHs, accounting for more than 94% (FATPHs: 95.84%; PATPHs: 94.33%). The Euryarchaeota contains most species of the archaea, including methanogens that are often found in animal guts (45, 46). The top genus in PATPHs was Methanocorpusculum (52.52%), followed by Methanobrevibacter (8.93%), Methanosarcina (4.32%), Methanomassiliicoccus (2.65%), and unclassified_p_Candidatus_Bathyarchaeota (2.32%) (Figure 1). Compared with those in FATPHs, the abundance of Methanocorpusculum showed a decreasing trend (11.15%↓, p = 0.663) and that of Methanobrevibacter showed an increasing trend (18.69%↑, p = 0.081) after anthelmintic treatment. However, other top genera did not vary much. It is known that both Methanocorpusculum and Methanobrevibacter are methanogens that promote fermentation of carbohydrates and produce methane in the gut of mammals (47, 48). Methanobrevibacter spp. were the dominant methanogens found in the guts of goats and dairy cows (49–51). The abundance of Methanocorpusculum spp. showed an increasing trend in horses fed with forage (52).
Figure 1.

Relative abundances of archaea genera ranked by one and zero orders of magnitude in PATPHs shown together with relative abundances of the corresponding genera in FATPHs.
The use of metagenomics can classify the microbes into species level (53), which 16S rRNA sequencing cannot. The present metagenomics results showed that the species of Methanocorpusculum labreanum and Methanocorpusculum bavaricum dominated in both of the FATPHs and PATPHs. Their relative abundances were 24.15% (Methanocorpusculum labreanum) and 17.13% (Methanocorpusculum bavaricum), respectively in FATPHs and 30.85 and 21.45%, respectively in PATPHs. Both species showed a decreasing trend following anthelmintic treatment (p = 0.663) (Figure 2). Furthermore, there were seven species with relative abundances at zero order of magnitude in FATPHs and PATPHs (Figure 2). Among them, Methanobrevibacter ruminantium (6.63%↑, p = 0.081) and Methanobrevibacter olleyae (5.94%↑, p = 0.081) had a significant increasing trend following anthelmintic treatment. This was consistent with the findings in sheep by Moon et al. (54), who indicated that an increase in relative abundance of Methanobrevibacter ruminantium was associated with anthelmintic treatment.
Figure 2.

Relative abundances of archaea species ranked by one and zero orders of magnitude in PATPHs shown together with relative abundances of the corresponding species in FATPHs.
Bacteria
A total of 82 bacterial phyla were detected in FATPHs and PATPHs, which is far more than the 23 phyla identified with 16S rRNA sequencing, referencing with the 16S rRNA sequencing results by Hu et al. (4), who did not find the presence of the Gemmatimonadetes phylum in FATPHs, but only a small 0.0012% in PATPHs. Results of the present metagenomic sequencing showed a successful detection of the Gemmatimonadetes in both FATPHs and PATPHs with relative abundances (percentage of read number in bacterial community) at 0.018 and 0.023%, respectively.
Overall, results of the metagenomic sequencing showed that the prevalent phyla were Firmicutes (FATPHs: 63.22%; PATPHs: 46.55%) and Bacteroidetes (FATPHs: 21.04%; PATPHs: 29.33%), which are commonly found in the guts of mammals (4, 18, 55, 56). It is noted that there were 49 phyla identified as unclassified bacteria (1 phylum), candidate division (9 phyla), and Candidatus phylum (39 phyla), with a total relative abundance of 5.55% in FATPHs and 9.26% in PATPHs. Meanwhile, comparison of the relative abundances of the 33 well-classified bacterial phyla in FATPHs and PATPHs showed that anthelmintic treatment reduced the relative abundances of 27 phyla and increased those of 6 phyla (Figure 3).
Figure 3.

Relative abundances of the well classified bacterial phyla in FATPHs and PATPHs.
Moreover, there were five phyla of Epsilonbacteraeota, Kiritimatiellaeota, Patescibacteria, WPS-2, and unclassified_k__norank_d__Bacteria identified in 16S rRNA sequencing, but not found in metagenomic sequencing. At present, the main methods for studying microbial community based on high-throughput sequencing platform are marker gene amplicon (16S rRNA, 18S rRNA, ITS, etc.) and metagenomics (57). The advantages of 16S rRNA sequencing are fast, low cost, and easy result analysis, but biases associated with PCR amplification are inevitable. On the other hand, metagenomic sequencing is generally less affected by biases but more costly (58, 59). Thus, it is necessary to choose more than one sequencing method in studying one single sample.
At the genus level, the top three genera in PATPHs with relative abundance higher than 5% were Bacteroides (9.06%), Prevotella (7.39%), and Clostridium (5.52%) (Figure 4). Compared with the corresponding genera in FATPHs, Clostridium exhibited an increasing trend (FATPHs: 6.53%, p = 0.383) and Bacteroides (FATPHs: 7.07%, p = 0.663) and Prevotella (FATPHs: 6.05%, p = 0.383) showed a decreasing trend after anthelmintic treatment. Ramanan et al. (60) demonstrated that deworming treatment for mice would change their gut Clostridium and Bacteroides levels. Meanwhile, a significant abundance decrease of the Bacteroides was found in sika deer after anthelmintic treatment (30). The association between Bacteroidetes and IL-10, a key anti-inflammatory cytokine involved in the induction of immune suppression, could be established after anthelmintic treatment (61). Clostridia were known to facilitate the host immune responses due to their production of short-chain fatty acids including butyrate with anti-inflammatory properties (62, 63). Mice that were infected with Trichuris muris experienced a decline in Prevotella after the removal of the infection (64). A study indicated that the changed abundance of Prevotella could drive Th17 immune responses, which were associated with the occurrence and development of many inflammatory and autoimmune diseases (65). In addition, among the other genera identified in this study with relative abundance at zero order of magnitude, Eubacterium exhibited the largest increasing trend (3.21%↑, p = 0.081) following anthelmintic treatment. The increase of Eubacterium after deworming was consistent with a study conducted in humans infected by Opisthorchis felineus (66). Eubacterium was found related with the physiology of horses, negatively correlated with salivary cortisol levels, but positively correlated with N-butyrate production (67).
Figure 4.

Top relative abundances of the well-classified bacterial genera ranked by zero order of magnitude in PATPHs shown together with the relative abundances of the corresponding genera in FATPHs.
Indeed, disruption of the intestinal microbes by parasite infestation has the capacity to modify the host's immune regulatory system (68). For example, a type 2 immune response to parasitic infection of Nippostrongylus brasiliensis in mice was related to the altered intestinal microbial community, especially the segmented filamentous bacteria (69). Thus, it is suspected that anthelmintic treatment of the horse botfly infection could moderate the immune responses of the Przewalski's horses by shifting their relative abundances of Bacteroides, Clostridium, Eubacterium, and Prevotella. On the other hand, Bacteroides and Prevotella were associated with plant-rich diets, which played an important role in the breakdown of indigestible fibers (70). Combined with the large number of methanogen archaea identified, anthelmintic treatment may be capable of altering the digestive ability of the Przewalski's horses.
Referencing to previous studies conducted with parasite-infected horses based on 16S rRNA sequencing, anthelmintic treatment could change the relative abundances of Streptococcus, Lactobacillus (4), Adlercreutzia (22), and Acinetobacter (24). Metagenomic analysis in the present study already reveals additional effects of anthelmintic treatment on the intestinal bacterial community.
At the species level, a total of 10,453 species were identified in FATPHs, and 10,421 in PATPHs, indicating a slight increase in bacterial species following anthelmintic treatment. All annotated species had relative abundances at zero order of magnitude or lower. The top four species in PATPHs were bacterium F082 (2.96%), bacterium P3 (2.55%), Ruminococcus flavefaciens (1.43%), and unclassified_g__Bacteroides (1.12%) (Figure 5). Their corresponding relative abundances at 1.27% (p = 0.383), 1.62% (p = 0.383), 0.76% (p = 0.383), and 0.78% (p = 0.383), respectively, in FATPHs showed a decreasing trend after anthelmintic treatment.
Figure 5.

Relative abundances of top four bacterial species in PATPHs and their corresponding species in FATPHs.
Eukaryota
Eukaryota is rarely studied in animal intestinal microbes. Very limited published articles on profiling the intestinal eukaryotic community of different animals can be found. They were done on dogs (71), humans (72), shrimps (73), and sika deer (30). The present study is the first to determine the composition and structure of the eukaryotic community in equine animal.
The top eukaryota phylum with relative abundance at one order of magnitude in PATPHs was unclassified_d__Eukaryota (60.53%) (Figure 6). It displayed a decreasing trend to 30.86% in FATPHs (p = 0.190) after anthelmintic treatment. With abundance at zero order of magnitude, PATPHs had seven phyla. They were Chordata, Streptophyta, Ascomycota, Arthropoda, Apicomplexa, Basidiomycota, and Nematoda (Figure 6). Among them, Streptophyta (10.97%↑, p = 0.190), Chordata (9.56%↑, p = 0.081), and Nematoda (5.67%↑, p = 0.663) showed large increasing trends in FATPHs after anthelmintic treatment. At the genus level, the relative abundances of all annotated genera were at one order of magnitude and lower. The 12 genera in PATPHs with abundances at one and zero orders of magnitude were Oxytricha, Stylonychia, Tetrahymena, Paramecium, Pseudocohnilembus, Ichthyophthirius, Trichomonas, Triticum, Epinephelus, Danio, Entamoeba, and Aegilops (Figure 7). Of them, five genera had their relative abundances shifted by more than 5% in FATPHs after anthelmintic treatment. They were Oxytricha (8.79%↓, p = 0.190), Stylonychia (7.95%↓, p = 0.190), Tetrahymena (5.62%↓, p = 0.190), Paramecium (5.36%↓, p = 0.190), and Trichomonas (5.31%↑, p = 0.383). A study of interventional treatment with probiotics and a low-fat diet on humans showed that the levels of the abovementioned genera except Streptophyta were reduced (74). Therefore, anthelmintic treatment may affect the digestion of the treated Przewalski's horses. Note that characteristics of the other eukaryota genera identified have not yet been studied.
Figure 6.

Relative abundances of eukaryota phyla ranked by one and zero orders of magnitude in PATPHs shown together with relative abundances of the corresponding phyla in FATPHs.
Figure 7.

Relative abundances of eukaryota genera ranked by one and zero orders of magnitude in PATPHs shown together with relative abundances of the corresponding genera in FATPHs.
Finally, there were 1,139 species and 1,034 species identified in PATPHs and FATPHs, respectively. Two species, Oxytricha trifallax (12.12%) and Stylonychia lemnae (10.83%), in PATPHs had relative abundances larger than 10%. Their abundances were reduced by an order of magnitude to 3.33% (p = 0.190) and 2.88% (p = 0.190), respectively, in FATPHs after anthelmintic treatment.
Virus
So far, the intestinal viral community was just done in dogs, cats, and humans (75, 76). This is the first study to determine the composition and structure of the viral community in equines.
There was only one viral phylum unclassified_d__Viruses identified in this study. Under this phylum, 70 genera were found in FATPHs and 69 in PATPHs. Eleven of the identified genera were in the unclassified group, accounting for 75.19% in FATPHs and 79.06% in PATPHs. The top three genera in PATPHs with relative abundances at one order of magnitude were unclassified_d__Viruses (26.91%), unclassified_f__Siphoviridae (25.32%), and unclassified_f__Myoviridae (17.48%). Comparing the abundances of theirs in FATPHs after anthelmintic treatment, unclassified_d__Viruses (29.33%, p = 0.081) showed an increasing trend while unclassified_f__Siphoviridae (23.76%, p = 1.000) and unclassified_f__Myoviridae (9.41%, p = 0.663) exhibited a decreasing trend. The large number of viruses annotated to unclassified indicates that there is a great opportunity to find new species in the equine gut, which is an unexplored habitat.
At the species level, 681 and 656 viruses were found in FATPHs and PATPHs, respectively. The top 10 abundant species consisted of one unidentified phage and nine uncultured viruses, making up 25.53 and 21.81% of the community in FATPHs and PATPHs, respectively. It is noted that most of viral species were classified into phage with 128 phages in FATPHs and 96 in PATPHs, occupying 71.49% of relative abundance in FATPHs and 72.98% in PATPHs, respectively. The relative abundance of phage changed more than other viral species after anthelmintic treatment.
Pathogens in FATPHs and PATPHs
There were totally 187 pathogens of 108 genera found in the samples when comparing the identified reads to the PHI database (Supplementary Table 1). The anthelmintic treatment increased the abundances of 128 pathogens and reduced those of 59 others. The top 10 pathogens based on gene abundance in PATPHs were Staphylococcus aureus (Bacteria), Salmonella enterica (Bacteria), Streptococcus pneumoniae (Bacteria), Fusarium graminearum (Eukaryota), Magnaporthe oryzae (Eukaryota), Pseudomonas aeruginosa (Bacteria), Escherichia coli (Bacteria), Cryptococcus neoformans, Listeria monocytogenes (Eukaryota), and Aspergillus fumigatus (Eukaryota) (Figure 8). Their abundances were all increased in FATPHs following anthelmintic treatment. For these pathogens, Eukaryota-related diseases have not been observed in equine animals. The methicillin-resistant Staphylococcus aureus (MRSA) is an emerging equine pathogen and is associated with a series of clinical diseases, such as septic arthritis, intravenous (jugular) catheter site infections, pneumonia cases, incisional infection, wound infection, mastitis, rhinitis, and body wall absces (77, 78). Horses infected by Salmonella enterica suffer from some clinical signs, such as fever, dehydration, diarrhea, colic, and septicemia (79, 80). Infection of Streptococcus pneumoniae will trigger immune response of the horse to accumulate leucocytes and cytokine (81). Pseudomonas aeruginosa is an opportunistic pathogen that is commonly recognized as a cause of endometritis in horses (82, 83). Escherichia coli are common commensal bacteria found in the intestinal tract of horses; they can cause diarrhea (83, 84). Casual usage of anthelmintic treatment poses a risk to the horses, known as colic (26). The common characteristics of an intestinal microbial community after antibiotic treatment include change of microbial diversity, increase of the colonization of pathogens, and development of antimicrobial resistance (85, 86). Hence, the removal of horse botflies through anthelmintic treatment raised the gene abundance of the top pathogens in the studied Przewalski's horses, which could increase their risk of sickness. However, this is limited by the small sample size used in the present study. Future repeated studies are needed for confirmation.
Figure 8.

Top 10 pathogens ranked by their gene abundances in PATPHs shown together with gene abundances of the corresponding pathogens in FATPHs.
Functional Analysis of Intestinal Microbiota Between FATPHs and PATPHs
It was shown that horse botfly expulsion by anthelmintic treatment for the studied Przewalski's horses altered their intestinal microbial community composition and structure. These changes in turn could affect the metabolism and physiology of the horses, such as microbial triggered immune responses (87), aid in regulation of energy metabolism (88, 89), and synthesis of the short-chain fatty acids or amino acids (90–92). In fact, it is increasingly recognized that the metabolism of animals is significantly affected by its intestinal microbes (93). Understanding the changes in intestinal microbial function is another key step toward clarifying the effects of anthelmintic treatment on horses. Metagenomic sequencing not only can characterize the gene content of a studied sample but also can predict the functional potential of its microbial community. The following sections present the metagenomic sequencing results on the intestinal microbial functions of the FATPHs and PATPHs.
COG Functional Annotation
A total of 4,869,449 genes with a total length of 2,194,175,671 were identified from the six samples used in this study. There were 24 kinds of COG functions associated with cellular processes and signaling, information storage and processing, metabolism, and poor characterization found in FATPHs and PATPHs. The top function was Function unknown, which accounted for 30.67% in FATPHs and 31.72% in PATPHs (Figure 9). There were five functions with their percentages of read numbers over 5%. They were replication, recombination, and repair (FATPHs: 10.09%; PATPHs: 9.55%); carbohydrate transport and metabolism (FATPHs: 7.61%; PATPHs: 7.45%); amino acid transport and metabolism (FATPHs: 6.98%; PATPHs: 6.44%); cell wall/membrane/envelope biogenesis (FATPHs: 6.56%; PATPHs: 6.86%); and translation, ribosomal structure, and biogenesis (FATPHs: 6.07%; PATPHs: 5.87%) (Figure 9). Distribution of the function catalog was consistent with that of Tang et al. (55), who studied some captive and wild Przewalski's horses without any anthelmintic treatment. Their samples were similar to those of the PATPHs in the present study.
Figure 9.

Distribution of COG functions in FATPHs and PATPHs. E ≤ 1e−5.
One important function of the intestinal microbes is metabolism, which converts carbohydrate and protein of dietary substrates to beneficial metabolites or alternative energy sources for the host (94). The intestinal microbes of the Przewalski's horses were found mainly to participate in carbohydrate and amino acid metabolism. The fermentation products of carbohydrate metabolism by microbes are short-chain fatty acids and gases. The three principal short-chain fatty acids detected in feces were acetate, butyrate, and propionate (95). Acetate plays an important role in regulating central appetite (96). Butyrate is capable of inducing the growth of cancer cells and colonic tumor cell lines, inhibiting mRNA expression and telomerase activity of cancer cells in human, enhancing memory recovery and formation, and preventing obesity in mice (97, 98). Propionate can be directly involved in portal-brain neural communication, induction of intestinal gluconeogenesis, and positive influence of the host's metabolism (99). Amino acids are key components of human and animal nutrition, which can regulate the intestinal bacterial community composition (100, 101).
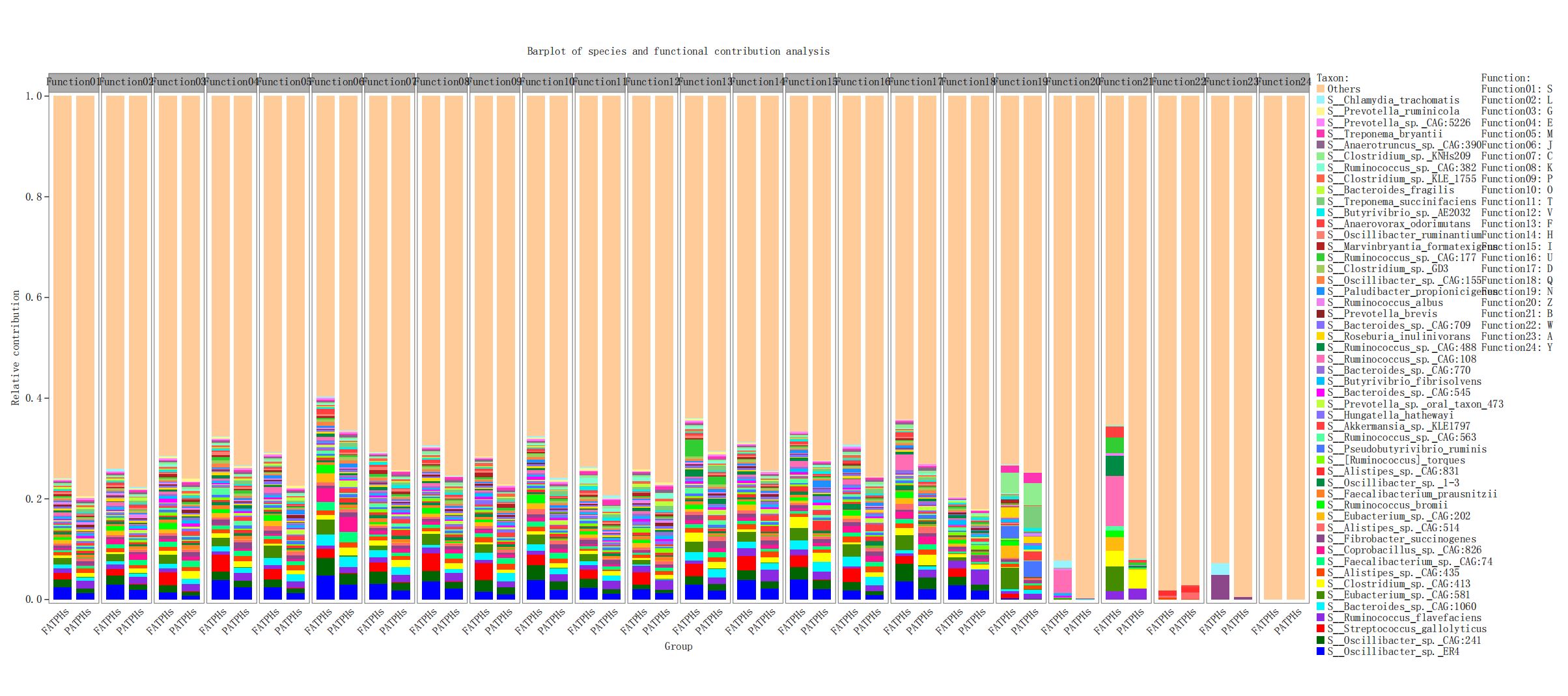
For these two important functions of the intestinal microbiota, the top five species involved in carbohydrate transport and metabolism were Ruminococcus flavefaciens, Clostridium sp.CAG:413, Faecalibacterium sp.CAG:74, Bacteroides sp.CAG:1060, and Fibrobacter succinogenes in PATPHs, and Streptococcus gallolyticus, Eubacterium sp.CAG:581, Oscillibacter sp.CAG:241, Oscillibacter sp.ER4, and Ruminococcus bromii in FATPHs (Supplementary Figure 1). The top five species involved in amino acid transport and metabolism were Oscillibacter sp.ER4, Ruminococcus flavefaciens, Fibrobacter succinogenes, Oscillibacter sp.CAG:241, and Clostridium sp.CAG:413 in PATPHs, and Oscillibacter sp.ER4, Streptococcus gallolyticus, Oscillibacter sp.CAG:241, Eubacterium sp.CAG:581, and Ruminococcus sp.CAG:563 in FATPHs (Supplementary Figure 1). Although the genes annotated with metabolism did not change significantly after anthelmintic treatment, the pattern of functional species was completely different between FATPHs and PATPHs. Anthelmintic treatment had a displacement effect on the same functional microbes.
PCA analysis showed that FATPHs and PATPHs can be separated based on the COG functional genes (Figure 10). It means that anthelmintic treatment changed the functions of the intestinal microbes. LEfSe analysis depicted that four functions showed significant differences between FATPHs and PATPHs (Figure 11). Transcription (LDA = 3.16, p = 0.05) showed significant effects on FATPHs while Function unknown (LDA = 3.97, p = 0.05), post-translational modification, protein turnover, chaperones (LDA = 3.14, p = 0.05), Nuclear structure (LDA = 3.08, p = 0.05), and intracellular trafficking, secretion, and vesicular transport (LDA = 2.85, p = 0.05) had significant effects on PATPHs. These results indicated that the functional differences of intestinal microbes resulted from some essential cellular processes rather than metabolism.
Figure 10.

PCA analysis of FATPHs and PATPHs based on calculation of the numbers of reads annotated to COG functions. Both chart axes have accounted for 99.76% of variance. Red triangles represent samples of FATPHs, while green dots represent samples of PATPHs.
Figure 11.

Cladogram showing the COG functions with significant differences between FATPHs and PATPHs. Functions with significant difference that have an LDA score >the threshold value of 3 are shown. K represents function of Transcription; S represents function of Function unknown; O represents function of Post-translational modification, protein turnover, and chaperones; Y represents function of Nuclear structure; and U represents function of Intracellular trafficking, secretion, and vesicular transport. The red bar represents samples of FATPHs, while green bars represent samples of PATPHs.
KEGG Pathway Annotation
The reads were annotated with six KEGG pathways at level 1, including Cellular processes (FATPHs: 5.45%; PATPHs: 5.96%), Environmental information processing (FATPHs: 6.28%; PATPHs: 6.41%), Genetic information processing (FATPHs: 14.67%; PATPHs: 14.00%), Human diseases (FATPHs: 4.38%; PATPHs: 5.17%), Metabolism (FATPHs: 66.02%; PATPHs: 53.84%), and Organismal systems (FATPHs: 3.20%; PATPHs: 4.62%) (Figure 12). Over 50% Level 1 KEGG pathways in FATPHs and PATPHs were Metabolism, consistent with results of COG function annotation. At level two, a total of 46 pathways were found and the top 3 belonged to Metabolism, which were Carbohydrate metabolism (FATPHs: 15.63%; PATPHs: 14.83%), Global and overview maps (FATPHs: 10.98%; PATPHs: 10.30%), and Amino acid metabolism (FATPHs: 9.14%; PATPHs: 8.76%). Carbohydrate and amino acid still represent the dominant function of the intestinal microbiota of the Przewalski's horses. In the KEGG pathway, the top five species involved in Carbohydrate metabolism were Clostridium sp.CAG:413, Ruminococcus flavefaciens, Faecalibacterium sp.CAG:74, Bacteroides sp. CAG:1060 and Oscillibacter sp. ER4 in PATPHs, and Streptococcus gallolyticus, Eubacterium sp.CAG:581, Oscillibacter sp.ER4, Oscillibacter sp.CAG:241, and Clostridium sp.CAG:413 in FATPHs (Supplementary Figure 2). For the Amino acid metabolism, the top five species contributors were Oscillibacter sp.ER4, Ruminococcus flavefaciens, Fibrobacter succinogenes, Clostridium sp.CAG:413, and Bacteroides sp.CAG:1060 in PATPHs, and Oscillibacter sp.ER4, Streptococcus gallolyticus, Eubacterium sp.CAG:581, Ruminococcus sp.CAG:563, and Oscillibacter sp.CAG:241 in FATPHs (Supplementary Figure 2). The results also suggested that anthelmintic treatment did not change the numbers of reads related to the pathways of metabolism but influenced the composition of the functional intestinal microbial community.
Figure 12.

KEGG pathways at Levels 1 and 2 found in FATPHs and PATPHs. E ≤ 1e−5.
Conclusions
In this study, the composition and structure of the complex intestinal microbial community of Przewalski's horses prior to and following anthelmintic treatment were identified by metagenomic sequencing for the first time. The pathogens were determined based on the gene information. The obtained sequences were mapped to known genes or pathways in COG and KEGG databases. The results indicated that anthelmintic treatment might have adverse effects on horses, which needs to be further confirmed; thus, optimization is suggested for this strategy of controlling parasite infections or search for alternative methods in the future.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
DH, JY, YQ, BL, and KL conceived the experiments. DH and KL undertook the sampling work and conducted the experiments. DH, KL, and KMM analyzed the results and wrote the manuscript. All the authors read and approved the final manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We sincerely thank all rangers, breeders, and staff members at the Kalamaili Nature Reserve and Przewalski's Horse Breeding Center.
Footnotes
Funding. This project was funded by the China Postdoctoral Science Foundation (No. 2020TQ0047) and National Natural Science Foundation of China (NSFC No. 31670538).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2021.708512/full#supplementary-material
{kind=link}
{kind=link}
References
- 1.Liu S, Hu D, Li K. Parasites observed in the proximal alimentary tract of a Przewalski's horse in China. Equine Vet Educ. (2016) 30:20–3. 10.1111/eve.12593 [DOI] [Google Scholar]
- 2.Zhang K, Huang H, Zhou R, Zhang B, Wang C, Ente M, et al. The impact of temperature on the life cycle of Gasterophilus pecorum in northwest China. Parasites Vect. (2021) 14:129. 10.1186/s13071-021-04623-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Coles G, Pearson G. Gasterophilus nasalis infection: prevalence and pathological changes in equids in south-west England. Vet Rec. (2000) 146:222–3. 10.1136/vr.146.8.222 [DOI] [PubMed] [Google Scholar]
- 4.Hu D, Chao Y, Zhang B, Wang C, Qi Y, Ente M, et al. Effects of Gasterophilus pecorum infestation on the intestinal microbiota of the rewilded Przewalski's horses in China. PLoS One. (2021) 16:e0251512. 10.1371/journal.pone.0251512 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Soulsby E. Helminths, Arthropods and Protozoa of Domesticated Animals, 7th Edn. London: Bailliere Tindall; (1982). [Google Scholar]
- 6.Sequeira J, Tostes R, Oliveira-Sequeira T. Prevalence and macro- and microscopic lesions produced by Gasterophilus nasalis (Diptera: Oestridae) in the Botucatu Region, SP, Brazil. Vet Parasitol. (2001) 102:261–6. 10.1016/S0304-4017(01)00536-2 [DOI] [PubMed] [Google Scholar]
- 7.Wang W, Zhang D, Hu D, Chu H, Cao J, Ente M, et al. Population genetic structure of Gasterophilus pecorum in the Kalamaili Nature Reserve, Xinjiang, based on mitochondrial cytochrome oxidase (COI) gene sequence. Med Vet Entomol. (2014) 28:75–82. 10.1111/mve.12073 [DOI] [PubMed] [Google Scholar]
- 8.Li K, Wu Z, Hu DF, Cao J, Wang C. A report on new causative agent (Gasterophilus spp.) of the myiasis of Przewalski's horse occurred in China. Chin J Anim Vet Sci. (2007) 38:837–40 (in Chinese). [Google Scholar]
- 9.Huang H, Zhang B, Chu H, Zhang D, Li K. Gasterophilus (Diptera, gasterophilidae) infestation of equids in the Kalamaili Nature Reserve, China. Parasite. (2016) 23:36. 10.1051/parasite/2016036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Liu S, Li K, Hu D. The incidence and species composition of Gasterophilus (Diptera, Gasterophilidae) causing equine myiasis in northern Xinjiang, China. Vet Parasitol. (2016) 217:36–8. 10.1016/j.vetpar.2015.12.028 [DOI] [PubMed] [Google Scholar]
- 11.Nielsen MK, Reinemeyer CR, Donecker JM, Leathwick DM, Marchiondo AA, Kaplan RM. Anthelmintic resistance in equine parasites-current evidence and knowledge gaps. Vet Parasitol. (2014) 204:55–63. 10.1016/j.vetpar.2013.11.030 [DOI] [PubMed] [Google Scholar]
- 12.Peregrine AS, Molento MB, Kaplan RM, Nielsen MK. Anthelmintic resistance in important parasites of horses: does it really matter? Vet Parasitol. (2014) 201:1–8. 10.1016/j.vetpar.2014.01.004 [DOI] [PubMed] [Google Scholar]
- 13.Relf VE, Lester HE, Morgan ER, Hodgkinson JE, Matthews JB. Anthelmintic efficacy on UK Thoroughbred stud farms. Int J Parasitol. (2014) 44:507–14. 10.1016/j.ijpara.2014.03.006 [DOI] [PubMed] [Google Scholar]
- 14.Kunz IGZ, Reed KJ, Metcalf JL, Hassel DM, Coleman RJ, Hess TM, et al. Equine fecal microbiota changes associated with anthelmintic administration. J Equine Vet Sci. (2019) 77:98–106. 10.1016/j.jevs.2019.01.018 [DOI] [PubMed] [Google Scholar]
- 15.Dicks LMT, Botha M, Dicks E, Botes M. The equine gastro-intestinal tract: an overview of the microbiota, disease and treatment. Livest Sci. (2014) 160:69–81. 10.1016/j.livsci.2013.11.025 [DOI] [Google Scholar]
- 16.Stewart HL, Southwood LL, Indugu N, Vecchiarelli B, Engiles JB, Pitta D. Differences in the equine faecal microbiota between horses presenting to a tertiary referral hospital for colic compared to an elective surgical procedure. Equine Vet J. (2018) 51:336–42. 10.1111/evj.13010 [DOI] [PubMed] [Google Scholar]
- 17.Schoster A, Staempfli HR, Guardabassi LG, Jalali M, Weese JS. Comparison of the fecal bacterial microbiota of healthy and diarrheic foals at two and four weeks of life. BMC Vet Res. (2017) 13:1. 10.1186/s12917-017-1064-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Biddle A, Tomb J-F, Fan Z. Microbiome and blood analyte differences point to community and metabolic signatures in lean and obese horses. Front Vet Sci. (2018) 5:225. 10.3389/fvets.2018.00225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Garber A, Hastie P, Murray J-A. Factors influencing equine gut microbiota: current knowledge. J Equine Vet Sci. (2020) 88:102943. 10.1016/j.jevs.2020.102943 [DOI] [PubMed] [Google Scholar]
- 20.Hu D, Chao Y, Li Y, Peng X, Wang C, Wang Z, Zhang D, Li K. Effect of gender bias on equine fecal microbiota. J Equine Vet Sci. (2020) 97:103355. 10.1016/j.jevs.2020.103355 [DOI] [PubMed] [Google Scholar]
- 21.Goachet AG, Ricard JM, Jacotot E, Varloud M, Julliand V. Effect of oral administration of anthelmintics on colonic microflora of horses. In: Proceedings of French Equine Veterinary Association. Pau (2004). [Google Scholar]
- 22.Peachey LE, Molena RA, Jenkins TP, Cesare AD, Traversa D, Hodgkinson JE, et al. The relationships between faecal egg counts and gut microbial composition in UK Thoroughbreds infected by cyathostomins. Int J Parasitol. (2018) 48:403–12. 10.1016/j.ijpara.2017.11.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Crotch-Harvey L, Thomas L-A, Worgan HJ, Douglas J-L, Gilby DE, Mcewan NR. The effect of administration of fenbendazole on the microbial hindgut population of the horse. J Equine Sci. (2018) 29:47–51. 10.1294/jes.29.47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Walshe N, Duggan V, Cabrera-Rubio R, Crispie F, Cotter P, Feehan O, et al. Removal of adult cyathostomins alters faecal microbiota and promotes an inflammatory phenotype in horses. Int J Parasitol. (2019) 49:489–500. 10.1016/j.ijpara.2019.02.003 [DOI] [PubMed] [Google Scholar]
- 25.Peachey LE, Castro C, Molena RA, Jenkins TP, Griffin JL, Cantacessi C. Dysbiosis associated with acute helminth infections in herbivorous youngstock - observations and implications. Sci Rep. (2019) 9:11121. 10.1038/s41598-019-47204-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Daniels SP, Leng J, Swann JR, Proudman CJ. Bugs and drugs: a systems biology approach to characterizing the effect of moxidectin on the horse's feecal microbiome. Anim Microbiome. (2020) 2:38. 10.1186/s42523-020-00056-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Iliev ID, Cadwell K. Effects of intestinal fungi and viruses on immune responses and inflammatory bowel diseases. Gastroenterology. (2020) 160:1050–66. 10.1053/j.gastro.2020.06.100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gaci N, Borrel G, Tottey W, O'Toole PW, Brugère J-F. Archaea and the human gut: new beginning of an old story. World J Gastroenterol. (2014) 20:16062–78. 10.3748/wjg.v20.i43.16062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Witherden E, Moyes DL. Mycobiome and gut inflammation. In: Chatterjee S, Jungraithmayr W, Bagchi D, editors. Immunity and Inflammation in Health and Disease. Amsterdam: Elsevier; (2018). p. 271–80. [Google Scholar]
- 30.Hu X, Xu Y, Liu G, Hu D, Wang Y, Zhang W, et al. The impact of anthelmintic treatment on gut bacterial and fungal communities in diagnosed parasite-free sika deer Cervus nippon. Appl Microbiol Biotechnol. (2020) 104:9239–50. 10.1007/s00253-020-10838-y [DOI] [PubMed] [Google Scholar]
- 31.Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. (2018) 34:884–90. 10.1093/bioinformatics/bty560 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li H, Durbin R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics. (2009) 25:1754–60. 10.1093/bioinformatics/btp324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Li D, Liu CM, Luo R, Sadakane K, Lam TW. MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics. (2015) 31:1674–6. 10.1093/bioinformatics/btv033 [DOI] [PubMed] [Google Scholar]
- 34.Noguchi H, Park J, Takagi T. MetaGene: prokaryotic gene finding from environmental genome shotgun sequences. Nucleic Acids Res. (2006) 34:5623–30. 10.1093/nar/gkl723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fu L, Niu B, Zhu Z, Wu S, Li W. CD-HIT: accelerated for clustering the next-generation sequencing data. Bioinformatics. (2012) 28:3150–2. 10.1093/bioinformatics/bts565 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Li R, Li Y, Kristiansen K, Wang J. SOAP: short oligonucleotide alignment program. Bioinformatics. (2008) 24:713–4. 10.1093/bioinformatics/btn025 [DOI] [PubMed] [Google Scholar]
- 37.Buchfink B, Xie C, Huson DH. Fast and sensitive protein alignment using DIAMOND. Nat Methods. (2015) 12:59–60. 10.1038/nmeth.3176 [DOI] [PubMed] [Google Scholar]
- 38.Peachey LE, Jenkins TP, Cantacessi C. This gut ain't big enough for both of us. Or is it? Helminth-microbiota interactions in veterinary species. Trends Parasitol. (2017) 33:619–32. 10.1016/j.pt.2017.04.004 [DOI] [PubMed] [Google Scholar]
- 39.Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci U S A. (2011) 108:16050–5. 10.1073/pnas.1102999108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bravo JA, Julio-Pieper M, Forsythe P, Kunze W, Dinan TG, Bienenstock J, et al. Communication between gastrointestinal bacteria and the nervous system. Curr Opin Pharmacol. (2012) 12:667–72. 10.1016/j.coph.2012.09.010 [DOI] [PubMed] [Google Scholar]
- 41.Brucker RM, Bordenstein SR. Speciation by symbiosis. Trends Ecol Evol. (2012) 27:443–51. 10.1016/j.tree.2012.03.011 [DOI] [PubMed] [Google Scholar]
- 42.Ezenwa VO, Gerardo NM, Inouye DW, Medina M, Xavier JB. Animal behavior and the microbiome. Science. (2012) 338:198–9. 10.1126/science.1227412 [DOI] [PubMed] [Google Scholar]
- 43.Aron-Wisnewsky J, Warmbrunn MV, Nieuwdorp M, Clement K. Metabolism and metabolic disorders and the microbiome: the intestinal microbiota associated with obesity, lipid metabolism and metabolic health: pathophysiology and therapeutic strategies. Gastroenterology. (2021) 160:573–99. 10.1053/j.gastro.2020.10.057 [DOI] [PubMed] [Google Scholar]
- 44.Yang W, Cong Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol Immunol. (2021) 18:866–77. 10.1038/s41423-021-00661-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Janssen PH, Kirs M. (2008). Structure of the archaeal community of the rumen. Applied and environmental microbiology, (2008). 74, 3619. 10.1128/AEM.02812-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Guindo CO, Davoust B, Drancourt M, Grine G. Diversity of methanogens in animals' gut. Microorganisms. (2020) 9:13. 10.3390/microorganisms9010013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Bapteste É, Brochier C, Boucher Y. Higher-level classification of the archaea: evolution of methanogenesis and methanogens. Archaea. (2005) 1:353–63. 10.1155/2005/859728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li Z, Wang X, Zhang T, Si H, Xu C, Wright A-DG, et al. Heterogeneous development of methanogens and the correlation with bacteria in the rumen and cecum of sika deer (Cervus nippon) during early life suggest different ecology relevance. BMC Microbiol. (2019) 19:129. 10.1186/s12866-019-1504-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guzman CE, Bereza-Malcolm LT, De Groef B, Franks AE. Presence of selected methanogens, fibrolytic bacteria, and proteobacteria in the gastrointestinal tract of neonatal dairy calves from birth to 72 hours. PLoS One. (2016) 10:e0133048. 10.1371/journal.pone.0133048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Dill-McFarland KA, Breaker JD, Suen G. Microbial succession in the gastrointestinal tract of dairy cows from 2 weeks to first lactation. Sci Rep. (2017) 7:40864. 10.1038/srep40864 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang Z, Elekwachi CO, Jiao J, Wang M, Tang S, Zhou C, et al. Investigation and manipulation of metabolically active methanogen community composition during rumen development in black goats. Sci Rep. (2017) 7:422. 10.1038/s41598-017-00500-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Fernandes KA, Kittelmann S, Rogers CW, Gee EK, Bolwell CF, Bermingham EN, et al. Faecal microbiota of forage-fed horses in New Zealand and the population dynamics of microbial communities following dietary change. PLoS One. (2014) 9:e112846. 10.1371/journal.pone.0112846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Scholz M, Ward D, Pasolli E, Tolio T, Asnicar F, Truong DT, et al. Strain-level microbial epidemiology and population genomics from shotgun metagenomics. Nat Methods. (2016) 13:435–8. 10.1038/nmeth.3802 [DOI] [PubMed] [Google Scholar]
- 54.Moon CD, Carvalho L, Kirk MR, Mcculloch AF, Leathwick DM. Effects of long-acting, broad spectra anthelmintic treatments on the rumen microbial community compositions of grazing sheep. Sci Rep. (2021) 11:3836. 10.1038/s41598-021-82815-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Tang L, Li Y, Srivathsan A, Guo Y, Li K, Hu D, et al. Gut microbiomes of endangered Przewalski's horse populations in short-and long-term captivity: implication for species reintroduction based on the soft-release strategy. Front Microbiol. (2020) 11:363. 10.3389/fmicb.2020.00363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Li Y, Zhang K, Liu Y, Li K, Hu D, Wronski T. Community composition and diversity of intestinal microbiota in captive and reintroduced Przewalski's horse (Equus ferus przewalskii). Front Microbiol. (2019) 10:1821. 10.3389/fmicb.2019.01821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Perez-Cobas AE, Gomez-Valero L, Buchrieser C. Metagenomic approaches in microbial ecology: an update on whole-genome and marker gene sequencing analysis. Microb Genomics. (2020) 6:8. 10.1099/mgen.0.000409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Walker AW, Martin JC, Scott P, Parkhill J, Flint HJ, Scott KP. 16S rRNA gene-based profiling of the human infant gut microbiota is strongly influenced by sample processing and PCR primer choice. Microbiome. (2015) 3:1–11. 10.1186/s40168-015-0087-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Knight R, Vrbanac A, Taylor BC, Aksenov A, Callewaert C, Debelius J, et al. Best practices for analysing microbiomes. Nat Rev Microbiol. (2018) 16:410–22. 10.1038/s41579-018-0029-9 [DOI] [PubMed] [Google Scholar]
- 60.Ramanan D, Bowcutt R, Lee SC, Tang MS, Kurtz ZD, Ding Y, et al. Helminth infection promotes colonization resistance via type 2 immunity. Science. (2016) 352:608–12. 10.1126/science.aaf3229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Martin I, Kaisar MMM, Wiria AE, Hamid F, Djuardi Y, Sartono E, et al. The effect of gut microbiome composition on human immune responses: an exploration of interference by helminth infections. Front Genet. (2019) 10:1028. 10.3389/fgene.2019.01028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G, Takahashi D, et al. Commensal microbe-derived butyrate induces the diferentiation of colonic regulatory T cells. Nature. (2013) 504:446–50. 10.1038/nature12721 [DOI] [PubMed] [Google Scholar]
- 63.Zaiss MM, Rapin A, Lebon L, Dubey LK, Mosconi I, Sarter K, et al. The intestinal microbiota contributes to the ability of helminths to modulate allergic infammation. Immunity. (2015) 43:998–1010. 10.1016/j.immuni.2015.09.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Houlden A, Hayes KS, Bancroft AJ, Worthington JJ, Wang P, Grencis RK, et al. Chronic Trichuris muris infection in C57BL/6 mice causes significant changes in host microbiota and metabolome: effects reversed by pathogen clearance. PLoS One. (2015) 10:e0125945. 10.1371/journal.pone.0125945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Larsen JM. Te immune response to Prevotella bacteria in chronic infammatory disease. Immunology. (2017) 151:363–74. 10.1111/imm.12760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Sokoloca TS, Petrov VA, Saltykova IV, Dorofeeva YB, Fedorova OS. The impact of Opisthorchis felineus infection and praziquantel treatment on the intestinal microbiota in children. Acta Trop. (2021) 217:105835. 10.1016/j.actatropica.2021.105835 [DOI] [PubMed] [Google Scholar]
- 67.Mach N, Foury A, Kittelmann S, Reigner F, Moroldo M, Ballester M, et al. The effects of weaning methods on gut microbiota composition and horse physiology. Front Physiol. (2017) 8:535. 10.3389/fphys.2017.00535 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Brosschot TP, Reynolds LA. The impact of a helminth-modifed microbiome on host immunity. Mucosal Immunol. (2018) 11:1039–46. 10.1038/s41385-018-0008-5 [DOI] [PubMed] [Google Scholar]
- 69.Fricke WF, Song Y, Wang A-J, Smith A, Grinchuk V, Pei C, et al. Type 2 immunity-dependent reduction of segmented flamentous bacteria in mice infected with the helminthic parasite Nippostrongylus brasiliensis. Microbiome. (2015) 3:40. 10.1186/s40168-015-0142-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ley R. Prevotella in the gut: choose carefully. Nat Rev Gastroenterol Hepatol. (2016) 13:69–70. 10.1038/nrgastro.2016.4 [DOI] [PubMed] [Google Scholar]
- 71.Heitlinger E, Ferreira SCM, Thierer D, Hofer H, East ML. The intestinal eukaryotic and bacterial biome of spotted hyenas: the impact of social status and age on diversity and composition. Front Cell Infect Microbiol. (2017) 7:262. 10.3389/fcimb.2017.00262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lukes J, Stensvold CR, Jirku-Pomajbíková K, Wegener PL. Are human intestinal eukaryotes beneficial or commensals? PLoS Pathog. (2015) 11:e1005039. 10.1371/journal.ppat.1005039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Dai W, Yu W, Zhang J, Zhu J, Tan Z, Xiong J. The gut eukaryotic microbiota influences the growth performance among cohabitating shrimp. Appl Microbiol Biotechnol. (2017) 101:6447–57. 10.1007/s00253-017-8388-0 [DOI] [PubMed] [Google Scholar]
- 74.Qian L, Huang J, Qin H. Probiotics and dietary intervention modulate the colonic mucosa-associated microbiota in high-fat diet populations. Turk J Gastroenterol. (2020) 31:295–304. 10.5152/tjg.2020.19013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Suchodolski JS. Intestinal microbiota of dogs and cats: a bigger world than we thought. Vet Clin N Am Small Anim Pract. (2011) 41:261–72. 10.1016/j.cvsm.2010.12.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Matijašić M, Meštrović T, Cipčić Paljetak H, Perić M, Barešić A, Verbanac D. Gut microbiota beyond bacteria-mycobiome, virome, archaeome, and eukaryotic parasites in IBD. Int J Mol Sci. (2020) 21:2668. 10.3390/ijms21082668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Weese JS, Rousseau J, Willey BM, Archambault M, McGeer A, Low DE. Methicillin-resistant Staphylococcus aureus in horses at a veterinary teaching hospital: frequency, characterization, and association with clinical disease. J Vet Intern Med. (2006) 20:182–6. 10.1111/j.1939-1676.2006.tb02839.x [DOI] [PubMed] [Google Scholar]
- 78.Park S, Ronholm J. Staphylococcus aureus in agriculture: lessons in evolution from a multispecies pathogen. Clin Microbiol Rev. (2021) 34:e00182–20. 10.1128/CMR.00182-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hernandez JA, Long MT, Traub-Dargatz JL, Besser TE. “Salmonellosis” - Equine Infectious Diseases. Amsterdam: Elsevier; (2014). p. 321–333. [Google Scholar]
- 80.Cummings KJ, Perkins GA, Khatibzadeh SM, Warnick LD, Aprea VA, Altier C. Antimicrobial resistance trends among Salmonella isolates obtained from horses in the northeastern United States (2001-2013). Am J Vet Res. (2016) 77:505–13. 10.2460/ajvr.77.5.505 [DOI] [PubMed] [Google Scholar]
- 81.Timoney J. The pathogenic equine streptococci. Vet Res. (2004) 35:397–409. 10.1051/vetres:2004025 [DOI] [PubMed] [Google Scholar]
- 82.Allen JL, Begg AP, Browning GF. Outbreak of equine endometritis caused by a genotypically identical strain of Pseudomonas aeruginosa. J Vet Diagn Invest. (2011) 23:1236–9. 10.1177/1040638711425589 [DOI] [PubMed] [Google Scholar]
- 83.Leon A, Castagnet S, Maillard K, Paillot R, Giard J-C. Evolution of in vitro antimicrobial susceptibility of equine clinical isolates in France between 2016 and 2019. Animals. (2020) 10:812. 10.3390/ani10050812 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Van Duijkeren E, van Asten AJAM, Gaastra W. Characterization of Escherichia coli isolated from adult horses with and without enteritis. Vet Q. (2020) 3:162–6. 10.1080/01652176.2000.9695048 [DOI] [PubMed] [Google Scholar]
- 85.Robinson CJ, Young VB. Antibiotic administration alters the community structure of the gastrointestinal microbiota. Gut Microbes. (2010) 1:279–84. 10.4161/gmic.1.4.12614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Schubert AM, Sinani H, Schloss PD. Antibiotic-induced alterations of the murine gut microbiota and subsequent effects on colonization resistance against Clostridium difficile. MBio. (2015) 6:e00974. 10.1128/mBio.00974-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Mackos AR, Maltz R, Bailey MT. The role of the commensal microbiota in adaptive and maladaptive stressor-induced immunomodulation. Horm Behav. (2017) 88:70–8. 10.1016/j.yhbeh.2016.10.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Cani PD, Delzenne NM. The role of the gut microbiota in energy metabolism and metabolic disease. Curr Pharm Des. (2019) 15:1546–58. 10.2174/138161209788168164 [DOI] [PubMed] [Google Scholar]
- 89.Zhang L, Liu C, Jiang Q, Yin Y. Butyrate in energy metabolism: there is still more to learn. Trends Endocrinol Metab. (2021) 3:159–69. 10.1016/j.tem.2020.12.003 [DOI] [PubMed] [Google Scholar]
- 90.Carey HV, Walters WA, Knight R. Seasonal restructuring of the ground squirrel gut microbiota over the annual hibernation cycle. Am J Physiol. (2013) 304:33–42. 10.1152/ajpregu.00387.2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud D, Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. (2013) 54:2325–40. 10.1194/jlr.R036012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Carey R.A, Montag D. (2021). Exploring the relationship between gut microbiota and exercise: short-chain fatty acids and their role in metabolism. BMJ Open Sport Exerc Med. 7:e000930. 10.1136/bmjsem-2020-000930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Lindsay EC, Metacalfe NB, Llewellyn MS. The potential role of the gut microbiota in shaping host energetics and metabolic rate. J Anim Ecol. (2020) 89:2415–26. 10.1111/1365-2656.13327 [DOI] [PubMed] [Google Scholar]
- 94.Rowland I, Gibson G, Heinken A, Scott K, Swann J, Thiele I, et al. Gut microbiota functions: metabolism of nutrients and other food components. Eur J Nutr. (2018) 57:1–24. 10.1007/s00394-017-1445-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Ivashkin V, Zolnikova O, Potskherashvili N, Trukhmanov A, Kokina N, Dzhakhaya N, et al. Metabolic activity of intestinal microflora in patients with bronchial asthma. Clin Pract. (2019) 9:18–22. 10.4081/cp.2019.1126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Frost G, Sleeth ML, Sahuri-Arisoylu M, Lizarbe B, Cerdan S, Brody L, et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun. (2014) 5:3611. 10.1038/ncomms4611 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Steliou K, Boosalis MS, Perrine SP, Sangerman J, Faller DV. Butyrate histone deacetylase inhibitors. Biores Open Access. (2012) 1:192–8. 10.1089/biores.2012.0223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Klepinina L, Klepinin A, Truu L, Chekulayev V, Vija H, Kuus K, et al. Colon cancer cell differentiation by sodium butyrate modulates metabolic plasticity of Caco-2 cells via alteration of phosphotransfer network. PLoS One. (2021) 16:e0245348. 10.1371/journal.pone.0245348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.De Vadder F, Kovatcheva-Datchary P, Goncalves D, Vinera J, Zitoun C, Duchampt A, et al. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell. (2014) 156:84–96. 10.1016/j.cell.2013.12.016 [DOI] [PubMed] [Google Scholar]
- 100.Dai ZL, Li XL, Xi PB, Zhang J, Wu G, Zhu WY. L-Glutamine regulates amino acid utilization by intestinal bacteria. Amino Acids. (2013) 45:501–12. 10.1007/s00726-012-1264-4 [DOI] [PubMed] [Google Scholar]
- 101.Karau A, Grayson I. Amino acids in human and animal nutrition. Adv Biochem Eng Biotechnol. (2014) 143:189–228. 10.1007/10_2014_269 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.