

Abstract
Extracellular DNA (eDNA) is a major constituent of the extracellular matrix of Pseudomonas aeruginosa biofilms and its release is regulated via pseudomonas quinolone signal (PQS) dependent quorum sensing (QS). By screening a P. aeruginosa transposon library to identify factors required for DNA release, mutants with insertions in the twin-arginine translocation (Tat) pathway were identified as exhibiting reduced eDNA release, and defective biofilm architecture with enhanced susceptibility to tobramycin. P. aeruginosa tat mutants showed substantial reductions in pyocyanin, rhamnolipid and membrane vesicle (MV) production consistent with perturbation of PQS-dependent QS as demonstrated by changes in pqsA expression and 2-alkyl-4-quinolone (AQ) production. Provision of exogenous PQS to the tat mutants did not return pqsA, rhlA or phzA1 expression or pyocyanin production to wild type levels. However, transformation of the tat mutants with the AQ-independent pqs effector pqsE restored phzA1 expression and pyocyanin production. Since mutation or inhibition of Tat prevented PQS-driven auto-induction, we sought to identify the Tat substrate(s) responsible. A pqsA::lux fusion was introduced into each of 34 validated P. aeruginosa Tat substrate deletion mutants. Analysis of each mutant for reduced bioluminescence revealed that the primary signalling defect was associated with the Rieske iron-sulfur subunit of the cytochrome bc1 complex. In common with the parent strain, a Rieske mutant exhibited defective PQS signalling, AQ production, rhlA expression and eDNA release that could be restored by genetic complementation. This defect was also phenocopied by deletion of cytB or cytC1. Thus, either lack of the Rieske sub-unit or mutation of cytochrome bc1 genes results in the perturbation of PQS-dependent autoinduction resulting in eDNA deficient biofilms, reduced antibiotic tolerance and compromised virulence factor production.
Author summary
Pseudomonas aeruginosa is a highly adaptable human pathogen responsible for causing chronic biofilm-associated infections. Biofilms are highly refractory to host defences and antibiotics and thus difficult to eradicate. The biofilm extracellular matrix incorporates extracellular DNA (eDNA). This stabilizes biofilm architecture and helps confer tolerance to antibiotics. Since mechanisms that control eDNA release are not well understood, we screened a P. aeruginosa mutant bank for strains with defects in eDNA release and discovered a role for the twin-arginine translocation (Tat) pathway that exports folded proteins across the cytoplasmic membrane. Perturbation of the Tat pathway resulted in defective biofilms susceptible to antibiotic treatment as a consequence of perturbed pseudomonas quinolone (PQS) signalling. This resulted in the failure to produce or release biofilm components including eDNA, phenazines and rhamnolipids as well as microvesicles. Furthermore, we discovered that perturbation of PQS signalling was a consequence of the inability of tat mutants to translocate the Rieske subunit of the cytochrome bc1 complex involved in electron transfer and energy transduction. Given the importance of PQS signalling and the Tat system to virulence and biofilm maturation in P. aeruginosa, our findings underline the potential of the Tat system as a drug target for novel antimicrobial agents.
Introduction
Pseudomonas aeruginosa is an opportunistic pathogen that causes a wide range of human infections including lung, urinary tract and wound, bacteremia and infections associated with medical devices [1]. It is notorious for its tolerance to antimicrobial agents, a property that is largely a consequence of its ability to form biofilm communities [1,2]. Bacterial exoproducts including cell surface appendages, extracellular polymeric substances, biosurfactants and secondary metabolites all contribute to P. aeruginosa biofilm formation and maturation [3–7].
Apart from exopolysaccharides such as Psl, Pel and alginate, the extracellular polymeric matrix of P. aeruginosa biofilms incorporates proteins, rhamnolipids, membrane vesicles (MVs) and extracellular DNA (eDNA) [5,8–10]. Rhamnolipid biosurfactants are required during the initial stages of micro-colony formation and are involved in the migration-dependent formation of the caps of mushroom-shaped micro-colonies formed in flow-cell grown biofilms [10]. They also aid the maintenance of channels between multicellular structures within biofilms and contribute to biofilm dispersal [5]. With respect to the biofilm micro-colonies that characteristically form in flow-chambers fed with glucose minimal medium, eDNA is present at high concentrations in the outer layers of microcolonies in young biofilms. However, in mature biofilms, eDNA is primarily located in the stalks at the borders between micro-colony caps and stalks [8].
The release of eDNA occurs via the lysis of a sub-population of bacterial cells [10–14]. It is involved in attachment, aggregation and stabilization of biofilm microcolonies. eDNA can act as a nutrient source, chelate metal cations and confer tolerance to antibiotics such as the polymyxins and aminoglycosides [10,12,13]. eDNA also binds other biopolymers (exopolysaccharides and proteins) stabilizing biofilm architecture and conferring protection against adverse chemical and physical challenges [12,13]. By intercalating with eDNA, secondary metabolites such as phenazines enhance biofilm integrity [12,15]. Pyocyanin for example can contribute to DNA release through the formation of reactive oxygen species such as hydrogen peroxide that damage cell membranes [12]. Although the mechanism(s) responsible for eDNA release has not been fully elucidated both eDNA and MVs can be generated via explosive cell lysis mediated via a cryptic prophage endolysin encoded within the R- and F-pyocin gene clusters [14].
In P. aeruginosa, rhamnolipids and pyocyanin production, eDNA and MV release, and hence biofilm development, are all controlled by quorum sensing (QS) [1,8,16]. Consequently, P. aeruginosa mutants with defects in this cell-to-cell communication system form aberrant, flat undifferentiated biofilms [10]. In P. aeruginosa, the QS regulatory network consists of a hierarchical cascade incorporating the overlapping Las, Rhl and PQS pathways that employ N-acylhomoserine lactones (AHLs) and 2-alkyl-4-quinolones (AQs) as signal molecules [1,16,17]. All three QS systems contain auto-induction loops whereby activation of a dedicated transcriptional regulator by the cognate QS signal molecule induces expression of the target synthase such that QS signal molecule production can be rapidly amplified to promote co-ordination of gene expression at the population level.
P. aeruginosa produces a diverse family of AQs and AQ N-oxides [18] of which 2-heptyl-3-hydroxy-4-quinolone (the Pseudomonas Quinolone Signal, PQS) and its immediate precursor 2-heptyl-4-hydroxyquinoline (HHQ) are most closely associated with PQS signalling [17] (Fig 1). Most of the genes required for AQ biosynthesis (pqsABCDE) and response (pqsR/mvfR) are located at the same genetic locus although pqsH and pqsL are distally located [17]. The biochemical basis for AQ and AQ N-oxide biosynthesis is summarized in S1 Fig. PqsA catalyses the formation of anthraniloyl-CoA that is condensed with malonyl-CoA by PqsD to form 2-aminobenzoylacetyl-CoA (2-ABA-CoA) [19,20]. The latter is converted to 2-aminobenzoylacetate (2-ABA) via the thioesterase functionality of PqsE [21]. The PqsBC heterodimer condenses 2-ABA with octanoyl-CoA to generate HHQ [22,23]. PQS is formed through the oxidation of HHQ by PqsH [24] while formation of the AQ N-oxides such as 2-heptyl-4-hydroxyquinoline N-oxide (HQNO) requires the alternative mono-oxygenase PqsL [25]. The PqsE protein has dual functions; while it is not essential for AQ biosynthesis, it is required for the AQ-independent production of several factors that contribute to biofilm maturation including pyocyanin, rhamnolipids and lectin A [26]. While activation of RhlR-dependent genes depends on PqsE [27], the AQ-independent, thioesterase-independent mechanism by which PqsE acts has not yet been elucidated [17,21].
Fig 1. The PQS signalling pathway in P. aeruginosa.
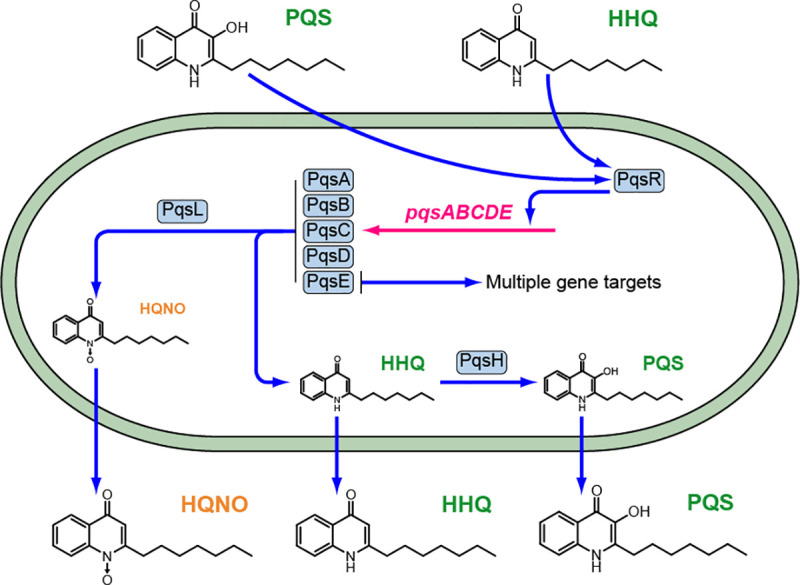
The PqsABCDE proteins synthesize HHQ, which is converted to PQS by PqsH and also HQNO in conjunction with PqsL. Both HHQ and PQS are released by the cells into the extracellular environment and are taken back up by neighboring cells. Autoinduction occurs when either HHQ or PQS binds to PqsR and amplifies expression of the pqsABCDE operon. The pqsE gene product has dual functions contributing to AQ biosynthesis as a thioesterase and also via an AQ-independent, thioesterase-independent mechanism for e.g. pyocyanin, rhamnolipid and lectin production as well as biofilm maturation. The conversion of HHQ to PQS confers additional functionalities since PQS unlike HHQ induces microvesicle formation and is a potent iron chelator.
The pqs system is subject to positive autoinduction, since the LysR-type transcriptional regulator PqsR (MvfR), binds to the promoter region of pqsABCDE (PpqsA) triggering transcription once activated by HHQ or PQS [28–30] (Fig 1). Therefore, by analogy with other QS systems, HHQ and PQS can both act as autoinducers by generating a positive feedback loop that accelerates their biosynthesis and co-ordinates a population-wide response. However, in contrast to HHQ which only regulates the pqsABCDE operon [17], PQS is a ferric iron chelator [31] that not only drives AQ biosynthesis via PqsR but also the expression of genes involved in the iron-starvation response and virulence factor production via PqsR-dependent and PqsR-independent pathways [17]. In addition, PQS can act as a cell-sensitizing pro-oxidant [32] and is required for MV production via a direct physical interaction with lipopolysaccharide (LPS) within the outer membrane [33]. The packaging of PQS within MVs also provides a means for trafficking this hydrophobic QS signal within a P. aeruginosa population [34].
With respect to biofilm development, PQS signalling is of particular interest because pqsA biosynthetic mutants fail to produce eDNA, rhamnolipids, pyocyanin and MVs, and form thin defective biofilms containing little eDNA [8, 27,33,35]. The mechanism involved in PQS-mediated DNA-release in biofilms is not understood but has been suggested to be linked to phage induction causing cell lysis [8,36–40]. Although explosive cell lysis releases eDNA in biofilms and generates MVs through vesicularization of shattered membrane fragments, pqsA mutants are not defective for explosive lysis [14] and therefore this phenomenon is unlikely to account for PQS-dependent eDNA release.
In the present study we sought to identify additional factors involved in eDNA release by screening a transposon (Tn) mutant library for eDNA-release defective mutants. Apart from pqs biosynthetic mutants, we obtained Tn insertion mutants within the twin-arginine translocation (Tat) pathway that exhibited reduced levels of eDNA release, fail to produce rhamnolipids or pyocyanin and form defective, eDNA- poor, antibiotic susceptible biofilms. Since mutation or deletion of tat resulted in altered AQ production, reduced pyocyanin, rhamnolipid and MVs, and as the tat mutants were refractory to exogenously supplied PQS, the aberrant biofilm phenotype observed could be accounted for by perturbation of PQS autoinduction. By screening a library of P. aeruginosa Tat substrate mutants, we identified the Rieske sub-unit of the cytochrome bc1 complex as the Tat substrate required for PQS-dependent QS and hence eDNA release and biofilm maturation.
Results
Transposon mutagenesis screen for P. aeruginosa mutants exhibiting reduced DNA release
To identify P. aeruginosa genes that contribute to eDNA release, a mariner Tn mutant library was generated in strain PAO1. Approximately 10,000 mutants grown in microtitre plates were assayed for reduced eDNA release using propidium iodide (PI) to quantify eDNA because it is unable to penetrate live bacteria and its fluorescence is enhanced 30-fold on binding DNA [41]. From the initial screen, 84 Tn insertion mutants were selected and re-screened to eliminate strains with double Tn insertions and to confirm their eDNA phenotype. For each of the remaining 34 mutants exhibiting reduced eDNA, the regions flanking each Tn insertion were sequenced and the corresponding genes identified. For most of the eDNA-release deficient mutants, the Tn insertions were located within genes required for AQ biosynthesis (pqsC and pqsH) or regulation (pqsR) (Fig 2) These data confirm our previous work that first uncovered a role for PQS signalling in eDNA release [8].
Fig 2. Transposon mutant screen for P. aeruginosa strains defective for eDNA release.
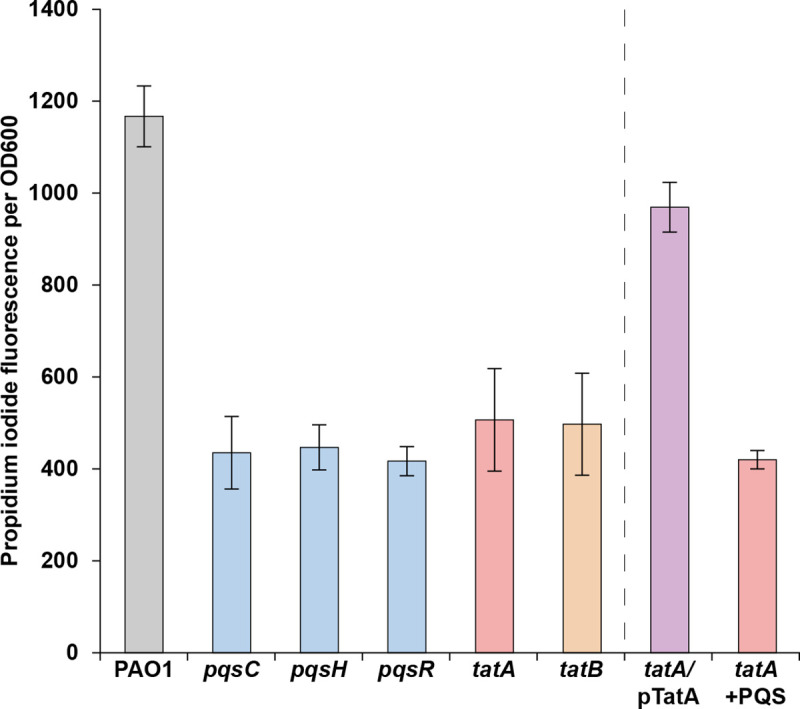
P. aeruginosa wild-type and mutant strains were grown for 24 h in 96 well microtiter plates containing ABTG medium, after which the relative levels of eDNA in the cultures were determined using a PI binding assay. The means and standard deviations of eight replicates are shown.
Apart from the pqs mutants, two mutants were obtained with insertions in the tatA and tatB genes respectively (Fig 2) that code for components of the twin-arginine translocation (Tat) system. Tat exports folded proteins out of the cytoplasm and across the cytoplasmic membrane in an ATP-independent manner [42]. It was originally named with respect to the presence of an Arg-Arg motif in the signal sequence of Tat-exported products (sometimes called Tat substrates) [42,43]. In P. aeruginosa the Tat translocase complex consists of three proteins (TatA, B and C) [43]. TatB and TatC form the receptor complex for Tat substrate precursors whereas TatA functions as the main facilitator for protein translocation across the membrane [42]. In P. aeruginosa diverse proteins involved in phosphate and iron metabolism, virulence and energy transduction are exported to the periplasm, or secreted via the Tat export system and tat mutants exhibit pleiotropic phenotypes [44,45]. Fig 2 shows that genetic complementation of the P. aeruginosa tatA mutant with a plasmid-borne copy restored eDNA release.
The Tat pathway contributes to biofilm development and tobramycin susceptibility in P. aeruginosa
Since eDNA makes an important contribution to biofilm development and architecture [10,12,13], biofilm formation by the P. aeruginosa tatA mutant in flow-chambers was investigated. After 4-days growth, the P. aeruginosa wild-type and complemented tatA/pTatA mutant formed biofilms with mushroom-shaped structures whereas the tatA mutant formed thin, flat biofilms (Fig 3A). In addition, eDNA was observed primarily in the stalks of the mushroom-shaped structures in the wild-type whereas tatA mutant biofilms contained no stalks and little extracellular DNA. Consistent with this flow-cell biofilm phenotype, exposure of the biofilms formed by each of the three strains to tobramycin showed that the tatA mutant biofilm was more sensitive to tobramycin than either the wild-type or tatA/pTatA mutant biofilm (Fig 3B). Since eDNA binds positively charged antibiotics and as exogenously provided DNA increases aminoglycoside tolerance by integrating into P. aeruginosa biofilms, the increased sensitivity to tobramycin is likely to be a consequence of the reduction in eDNA within the biofilm extracellular matrix [46,47].
Fig 3. P. aeruginosa tat mutants form defective biofilms with increased susceptibility to tobramycin.
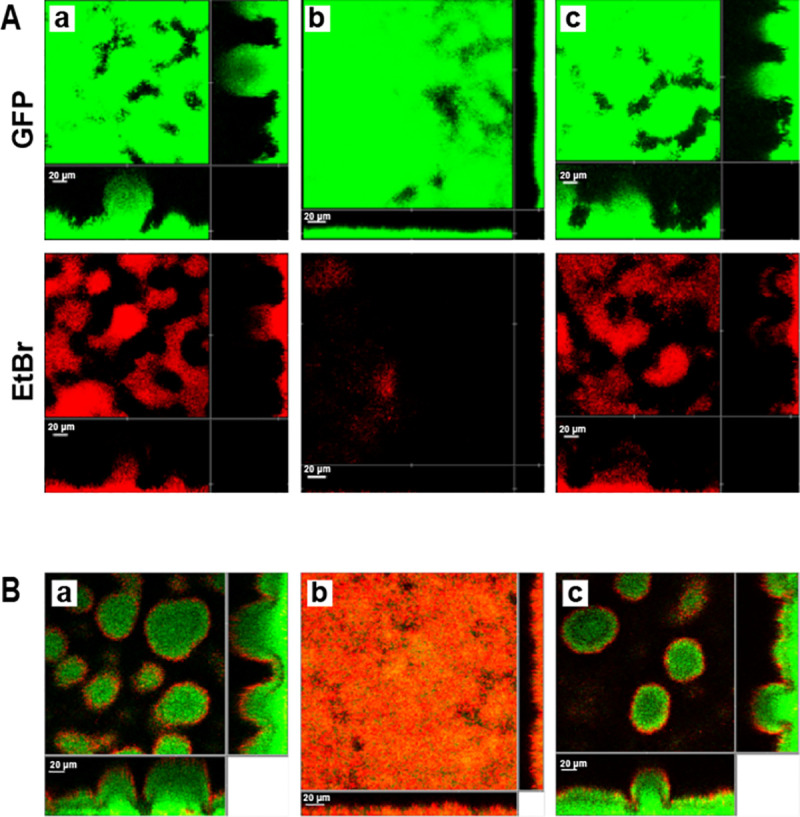
CLSM images showing four-day-old biofilms formed in flow chambers of the gfp-tagged P. aeruginosa wild-type (a), tatA mutant (b) and genetically complemented tatA mutant (c). In (A) biofilms were stained for total biomass with Syto9 (green) and for eDNA with ethidium bromide (red). In (B) biofilms were treated with tobramycin and the medium was supplemented with propidium iodide prior to CLSM such that dead cells appear red while live cells appear green. Each panel shows one horizontal optical section two flanking vertical optical sections. Bars, 20 μm.
P. aeruginosa tatA mutants are defective in the production of rhamnolipids, pyocyanin and MVs
Since rhamnolipids, pyocyanin and MVs are all important components of P. aeruginosa biofilms, their production was quantified in the tatA Tn insertion mutant and in a ΔtatABC deletion mutant. Fig 4A and 4B show that tat mutants produce substantially less pyocyanin and rhamnolipid than the parent or tatA complemented strain. Furthermore, MV levels (Fig 4C) were reduced by ~50% in the tatA mutant compared with the wild type and could be restored by genetic complementation.
Fig 4.
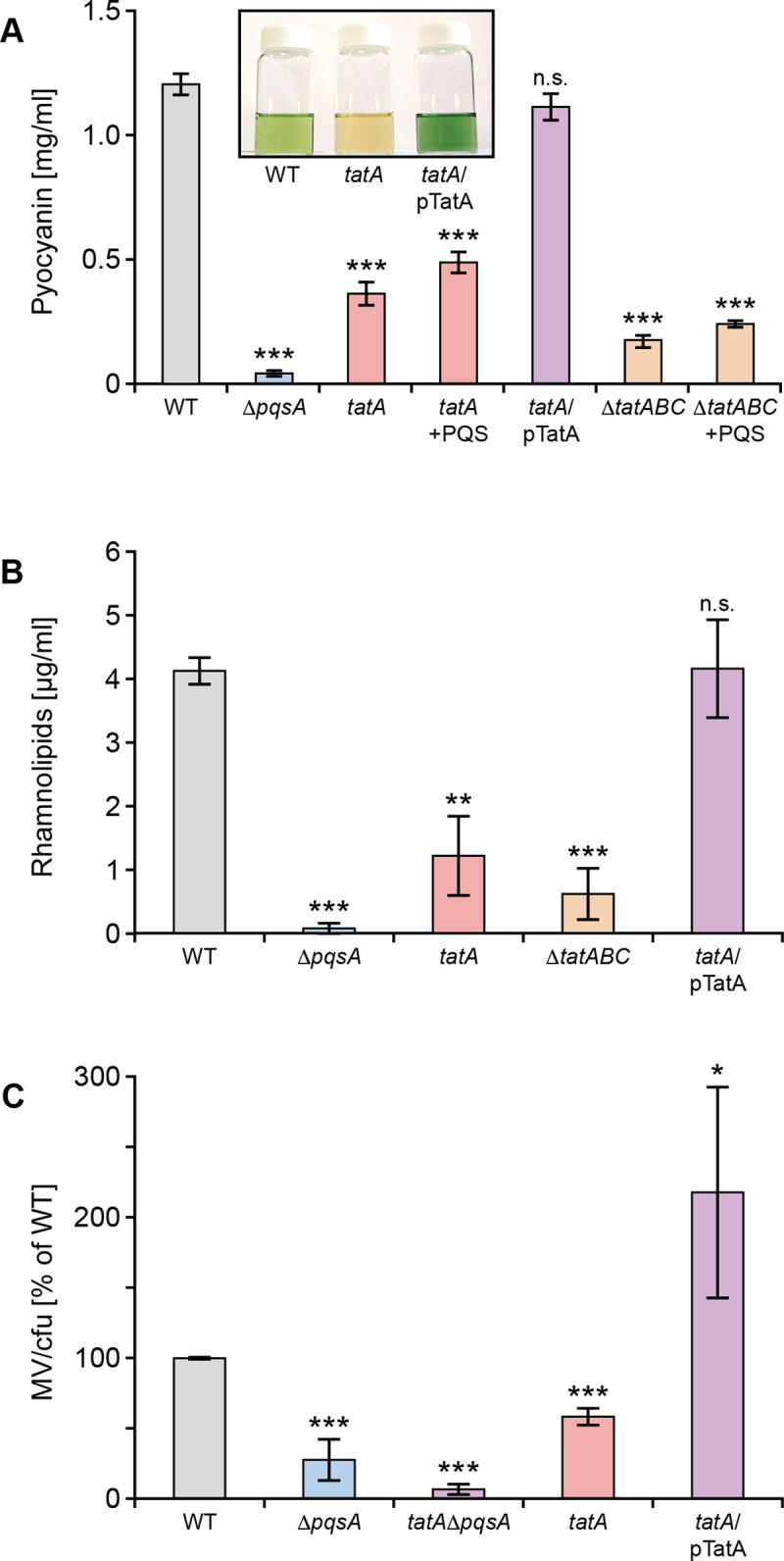
Production of pyocyanin (A), rhamnolipids (B) and MVs (C) are reduced in P. aeruginosa tat mutants. (A) Pyocyanin levels are shown in the P. aeruginosa wild type, ΔpqsA, tatA, and tatABC deletion mutants and the tatA mutant complemented with plasmid-borne tatA. The impact of exogenous PQS (40 μM) on the tatA and ΔtatABC mutants is also shown. Insert panel shows the absence of green pigment in the tatA mutant compared with the wild type and complemented tatA mutant. (B) Rhamnolipid production in the tatA and ΔtatABC mutants compared with the wild type, ΔpqsA mutant and tatA complemented with plasmid-borne tatA. (C) Comparison of MV production in the tatA mutant, complemented tatA mutant and in a double tatA ΔpqsA mutant compared with the wild type strain. Experiments were repeated in triplicate at least twice. ***p < 0.001, **p < 0.01; n.s. not significant.
Inactivation of the Tat pathway by mutation or small molecule-mediated inhibition perturbs PQS signalling
The reductions in eDNA release, rhamnolipids, pyocyanin and MVs noted in the tat mutant as well as its biofilm phenotype are comparable with those observed for P. aeruginosa strains with mutations in pqs genes such as pqsA, the first gene in the AQ biosynthetic pathway (see Figs 1 and 4 and [8, 40]). These data therefore suggested that the tat mutant biofilm phenotype was likely, at least in part, to be due to a defect in PQS signalling. To investigate the impact of the tatA mutation on the expression of pqsA, a CTX::pqsA’-lux fusion was introduced into the chromosomal CTX site of both the wild type and tatA mutant. Fig 5A shows that pqsA expression in the tatA mutant is reduced ~4 fold compared with the wild type strain and restored by genetic complementation of the mutant. In agreement with these data, the Tat inhibitor, Bayer 11–7082, identified by Vasil et al [48] reduced pqsA expression in the wild type PAO1 strain by ~4 fold consistent with the reduction noted for the CTX::pqsA’-lux fusion in the tatA mutant (Fig 5B). Bayer-11 7082 had no effect on growth or light output in P. aeruginosa expressing the lux genes from a derepressed lac promoter (S2 Fig). In addition, the concentration of PQS in whole culture extracts of P. aeruginosa after growth in LB medium as determined by LC-MS/MS was respectively ~56% lower in the tatA mutant compared with the wild type and complemented tat mutant (Fig 5C). Since pqsA expression and hence AQ production is also PqsR/MvfR-dependent, we compared the expression of pqsR in the ΔtatABC with the parent strain but found no difference (S3 Fig).
Fig 5. Mutation of tatA or exposure to the Tat inhibitor Bayer 11–7082 inhibits pqsA expression and AQ production.
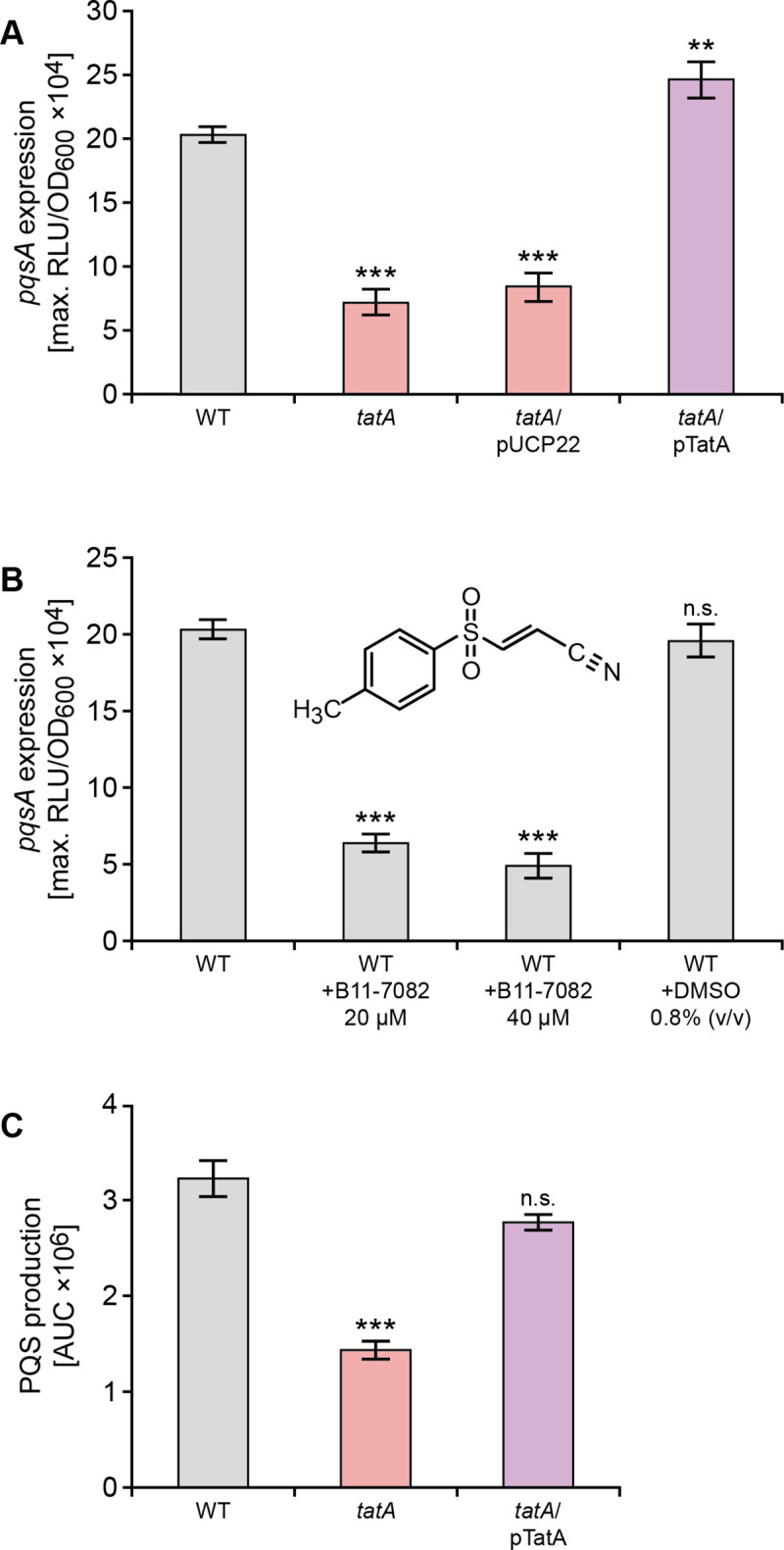
(A) Mutation of tatA or (B) treatment with Bayer 11–7082 supplied at either 20 μM or 40 μM reduces the maximal expression of a P. aeruginosa PAO1 chromosomal pqsA’-lux promoter fusion without affecting growth. (C) LC-MS/MS analysis of PQS production by P. aeruginosa PAO1 wild type compared with the tatA mutant and complemented tatA mutant. Experiments were repeated in triplicate at least twice. ***p < 0.001; n.s. not significant.
Exogenous PQS does not restore PQS signalling in a P. aeruginosa tatA mutant
QS systems are characteristically autoinducible such that exogenous provision of the cognate signal molecule usually induces expression of the signal synthase and hence activation of downstream target genes [49]. When the tatA mutant was provided with exogenous PQS, eDNA release did not increase (Fig 2). To investigate this finding further, either PQS or HHQ was exogenously supplied to the wild type, tatA mutant or the complemented tatA mutant strains carrying chromosomal pqsA’-lux fusions. The data presented in Fig 6A show that the response of the tatA mutant to PQS or HHQ respectively at 5 or 20 μM with respect to pqsA expression was at least 2-fold lower than the controls. Since both wild type and tatA mutant still produce AQs endogenously, the experiments were repeated in the P. aeruginosa ΔpqsA and tatA ΔpqsA mutants since no AQs are produced in these genetic backgrounds. Fig 6B shows that the response to both PQS and HHQ is substantially reduced (e.g. ~3 fold at 5 μM PQS) in the absence of tatA in the P. aeruginosa tatA ΔpqsA double mutant. This reduced response to PQS could be due either to reduced uptake or the inability to respond to the exogenous QS signal molecule.
Fig 6.
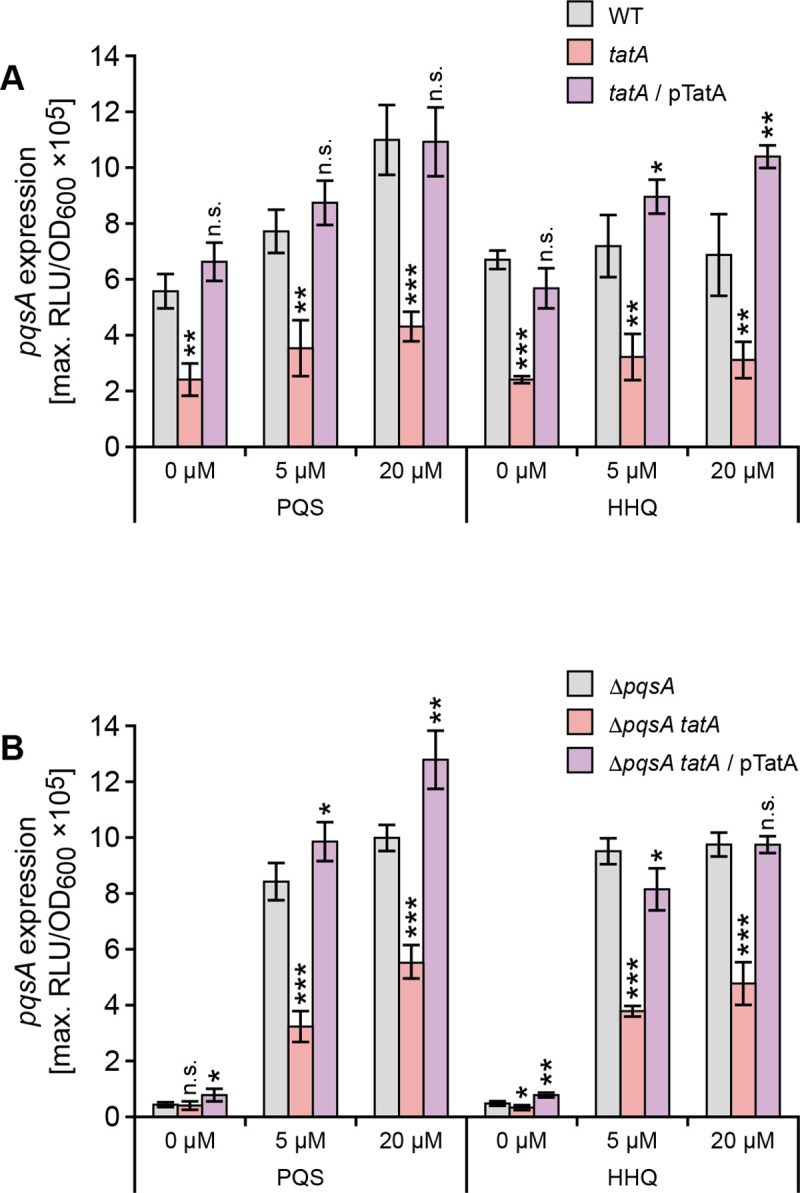
Exogenous AQs do not restore pqsA expression in P. aeruginosa tatA (A) or tatA ΔpqsA (B) mutants. Exogenous PQS or HHQ was added at 5 μM or 20 μM to (A) wild type, the tatA mutant and complemented tatA mutant or (B) ΔpqsA, ΔpqsA tatA or tatA ΔpqsA mutant complemented with tatA. Maximal light output from the chromosomal pqsA’-lux fusion was recorded as a function of growth (RLU/OD600). Experiments were repeated in triplicate at least twice. ***p < 0.001, **p < 0.01, and *p < 0.05; n.s. not significant.
To determine the consequences of perturbed PQS signalling on the expression of the rhamnolipid (rhlA) and pyocyanin biosynthetic genes (P. aeruginosa has two, almost identical redundant 7 gene phenazine biosynthetic operons termed phzA1-G1 and phzA2-G2; [50]), the corresponding miniCTX-lux promoter fusions for rhlA and phzA1 respectively were constructed and introduced onto the chromosomes of the wild type, ΔpqsA and tatA ΔpqsA mutants respectively. Fig 7A shows the expression profiles of rhlA’-lux as a function of time. Both the wild type and ΔpqsA mutant show an ~2 fold increase in rhlA expression when supplied with exogenous PQS (20 μM) and share similar profiles over the growth curve. In contrast, the rhlA’-lux fusion in the tatA mutant does not show the same expression profile or response to exogenous PQS as the wild type and ΔpqsA mutant strains. The rhlA’-lux expression profile in the tatA mutant supplied with exogenous PQS is however clearly restored when the mutation is complemented by pTatA (Fig 7A).
Fig 7. Rhamnolipid (rhlA) and pyocyanin biosynthetic (phzA1) genes show altered expression profiles in P. aeruginosa tat mutants and fail to respond to exogenous PQS.

(A) Light output from a chromosomal rhlA’-lux fusion as a function of growth (RLU/OD) over time when introduced into the wild type, pqsA and tatA mutants or complemented tatA mutant in the absence or presence of exogenous PQS (20 μM). (B) Maximal light output from a chromosomal phzA1’-lux fusion as a function of growth (RLU/OD600) when introduced into the wild type, ΔpqsA, or ΔpqsA tatA mutants in the absence or presence of exogenous PQS (10 or 40 μM) or plasmid-borne pqsE or the pUCP18 vector control. Experiments were repeated in triplicate at least twice. ***p < 0.001, **p < 0.01, and *p < 0.05; n.s. not significant.
AQ-dependent QS is required for phzA1 expression [27,50]. Exogenous PQS increased phzA1’-lux expression by ~4 fold in both wild type and ΔpqsA mutant backgrounds (Fig 7B). However, the tatA ΔpqsA double mutant responded comparatively poorly to PQS (Fig 7B).
Constitutive expression of pqsABCD does not restore AQ biosynthesis in a tat mutant
Since mutation of tatA resulted in reduced pqsA expression, it was possible that the auto-induction of PQS biosynthesis via PqsR (MvfR) is compromised. To uncouple the autoinduction of AQ production, the pqsABCD genes were introduced into the P. aeruginosa ΔpqsA and ΔpqsAΔtatA mutants respectively on a plasmid (pBBRMCS5::pqsABCD) constitutively expressing pqsABCD [51]. Fig 8 shows that PQS, HHQ and HQNO are present in the culture medium of the ΔpqsA mutant transformed with pBBRMCS5::pqsABCD. However neither the cell free supernatant (Fig 8) nor whole cells of the tatA ΔpqsA double mutant transformed with pBBRMCS5::pqsABCD contained or accumulated intracellular AQs (S4 Fig). These data suggested that in a tat mutant background, the lack of AQs is not a consequence of an AQ transport defect, but is due to the inability to fully activate AQ biosynthesis at the appropriate time/population density. In the absence of an auto-inducible pqs system in a tat mutant background, the defect in AQ biosynthesis and hence PQS signalling appears to be more severe.
Fig 8. AQ biosynthesis is not restored in a tatA ΔpqsA double mutant by plasmid-borne pqsABCD in the absence of autoinduction.

Semi-quantitative analysis by LC-MS/MS of PQS, HHQ and HQNO production by P. aeruginosa pqsA and tatA ΔpqsA mutants respectively without or with the pqsABCD biosynthetic genes provided in trans via pBBR1MCS-5::pqsABCD. Experiments were repeated in triplicate at least twice.
Reduced rhamnolipid production does not account for defective PQS signalling
In P. aeruginosa biofilms, rhamnolipids provide protective shielding against neutrophils [52,53] and contribute to the effectiveness of PQS signalling by increasing the solubility and bioactivity of PQS [54]. In Fig 4B, we showed that rhamnolipid production was substantially reduced in the P. aeruginosa tat mutant background. To determine whether the perturbation of PQS signalling in the tat mutants was a consequence of reduced rhamnolipid production, we investigated the impact of exogenous rhamnolipids on pqsA expression. S5 Fig shows that the addition of purified rhamnolipids (10 or 50 μg/ml) to the tatA ΔpqsA mutant with or without PQS (40 μM) had little effect on PQS signalling indicating that the defect in the tat mutants was not simply due to the loss of rhamnolipid production and an inability to solubilize PQS.
PqsE restores pyocyanin in the tat mutants
Although PqsE is not essential for AQ biosynthesis, it is an effector protein required for the production of pyocyanin, rhamnolipids and biofilm maturation and its function is independent of PQS, HHQ or PqsR [27,55]. Consequently, the expression of PQS -dependent exoproducts such as pyocyanin can be restored in a pqsA negative (and hence AQ-negative) mutant by expressing a plasmid-borne copy of pqsE. In the tatA ΔpqsA double mutant, phzA1 expression (Fig 7B) and pyocyanin production (Fig 9) were respectively restored by pqsE expression indicating that the tat mutation does not affect PqsE functionality.
Fig 9. PqsE restores pyocyanin production in P. aeruginosa tat mutants.
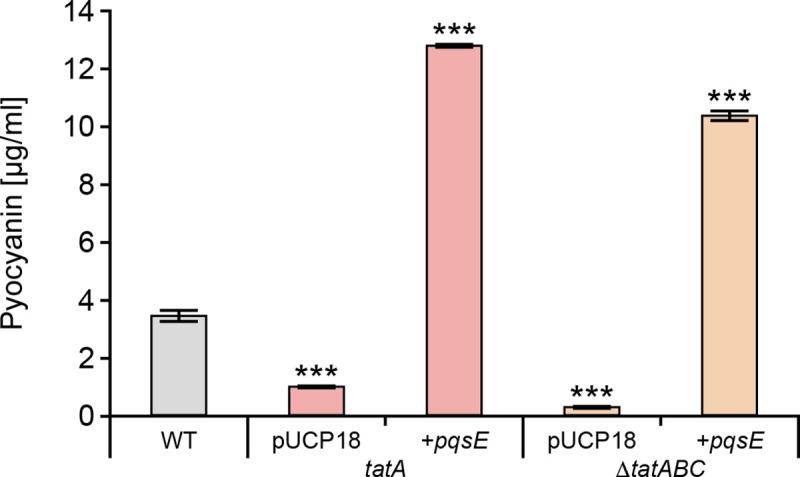
Production of pyocyanin by the wild type compared with the tatA and ΔtatABC mutants transformed with plasmid-borne pqsE or the empty vector. Experiments were repeated in triplicate at least twice. ***p < 0.001.
Identification of the Tat substrate responsible for perturbation of PQS signalling
Recently Gimenez et al [45] experimentally validated the Tat-mediated export of 34 P. aeruginosa gene products predicted to have Tat signal peptides. To determine which of the exported Tat substrates was responsible for perturbation of PQS signalling, allelic replacement mutants were constructed in P. aeruginosa strain PA14 for each substrate. Before introducing the pqsA’-lux fusion onto the chromosomal CTX site of each Tat substrate mutant, we first confirmed that PQS signalling in a PA14 ΔtatABC mutant was perturbed in a similar manner to that observed for PAO1, the genetic background used so far in the study (Fig 10A). Determination of the maximum expression of pqsA’-lux for each of the 34 Tat substrate mutants (Fig 10A) revealed that although a number of mutants exhibited reduced light output, the greatest reduction was observed for deletion of PA14_57570 (equivalent to PA4431 in PAO1, here designated petA following the nomenclature of orthologues described in other species). This gene codes for the Rieske subunit of the cytochrome bc1 complex that is involved in electron transfer and respiration under aerobic conditions and is also required for P. aeruginosa growth under anaerobic conditions in the presence of nitrite, nitric oxide or nitrous oxide [56].
Fig 10. Tat substrate screen for P. aeruginosa PA14 mutants with defects in pqs signaling uncovers a role for the cytochrome bc1 Rieske sub-unit.

(A) Comparison of peak pqsA expression in each of 34 Tat substrate mutants (S3 Table) transformed with a CTX::pqsA’-lux fusion compared with the PA14 wild type (white bar) and the ΔtatABC mutant (black bar). The bars represent mutants where pqsA expression is higher (green bar) or lower (grey bar) or the same (grey bar) as the wild type. PA14 mutant 27 (pink bar) has the lowest pqsA expression and carries a deletion in PA14-57570 (petA), the Rieske subunit of cytochrome bc1. (B) Production of PQS, HHQ and HQNO after 16 h growth by P. aeruginosa PA14, the ΔtatABC and ΔpetA mutants and the genetically complemented PA14 mutant (ΔpetA::(CTX::petA)). ***p < 0.001, **p < 0.01, and *p < 0.05; n.s. not significant.
To confirm that the ΔpetA mutant in common with the P. aeruginosa tat mutants exhibited similar defects as a consequence of perturbed PQS signalling, we compared their AQ production (Fig 10B), rhlA expression, and eDNA release in biofilms (S6 and S7 Figs). Comparison of PQS, HHQ and HQNO production in PA14 with the ΔpetA mutant and the strain with a cis complementation of the petA mutation at the CTX site confirmed the loss and increase in AQ production (Fig 10B). Similar results were obtained for pqsA expression in a PAO1 ΔpetA mutant (S8 Fig). In P. aeruginosa PA14, the rhlA expression profiles for the parent and ΔtatABC deletion mutant in the absence or presence of exogenously supplied PQS were similar to those of strain PAO1 (compare S6 Fig with Fig 7A). Confocal microscopy images of biofilm formation by P. aeruginosa PA14 under static growth conditions shows that in common with the ΔtatABC mutant, ΔpetA mutant biofilms lack the eDNA content observed in the wild type and complemented mutant (S7 Fig) demonstrating that PetA is the primary Tat substrate required for PQS-dependent eDNA release. Since PetA is one of the three electron transfer proteins that constitute the cytochrome bc1 complex we also investigated whether mutation of cytB and cytC1 genes (PA4429 and PA4430 respectively) perturbed PQS-signalling. S9 Fig shows ΔcytB and ΔcytC1 mutants also exhibited reduced pqsA expression and AQ production indicating that a fully functional cytochrome bc1 is essential for PQS-dependent QS. Given the importance of cytochrome bc1 for energy transduction and growth, we also compared aerobic growth and ATP production in the P. aeruginosa wild type with that of the tat and petA mutants. S9 Fig shows that there is a similar small reduction in growth for mutants compared with the wild type. Consistent with these observations, ATP levels were reduced by ~35% in the Δtat and ΔpetA mutants respectively compared with wild type (S10 Fig). This finding suggested that in P. aeruginosa strains carrying mutations in tat or cytochrome bc1 the energy required to synthesize secondary metabolites such as the AQs is diverted to maintain primary metabolism.
Discussion
To gain further insights into eDNA release, a P. aeruginosa Tn mutant library was screened and two groups of mutants exhibiting reduced eDNA release were identified. The first of these contained Tn insertions in pqsC, pqsH and pqsR. Mutations in pqsC or pqsR in common with pqsA both abrogate AQ biosynthesis while pqsH mutants are unable to produce 3-hydroxy-AQs such as PQS but maintain production of AQs and AQ N-oxides such as HHQ and HQNO respectively [17, 54]. These data suggest that eDNA release is likely to require PQS/HHQ rather than the effector protein PqsE. This is because although pqsE mutants in common with pqsA mutants form poor biofilms, pqsE pqsA double mutants could not be complemented by pqsE alone to restore biofilm development [26]. This contrasts with pyocyanin and lectin A for example, as production of both can be restored by PqsE in the absence of AQ biosynthesis [26]
The Tn insertions in the second group of P. aeruginosa eDNA release mutants were in tatA and tatB [42,43]. In P. aeruginosa, proteins exported by the Tat system are most frequently terminally localized in the periplasm, but some (e.g. phospholipase C) can be transported across the outer membrane by the Xcp type II secretion system, thereby becoming extracellular [43]. Using prediction algorithms for Tat substrates, 44 putative P. aeruginosa PA14 Tat signal peptides have been identified and of these, 34 confirmed experimentally [45]. These include phospholipases, and proteins involved in pyoverdine-mediated iron-uptake, respiration, osmotic stress defence, motility, and biofilm formation [44, 45]. However, none of these are known to be involved in AQ biosynthesis, transport or PQS signal transduction.
The formation of flat, eDNA deficient, tobramycin-sensitive biofilms and the reduction in rhamnolipid, pyocyanin and MV production by the P. aeruginosa tatA mutant are consistent with the pqsA mutant biofilm phenotype [8] and implied the existence of a link between Tat export and PQS signalling. Since transcriptomic studies of PQS signalling in P. aeruginosa have not provided any evidence that the Tat export system is QS controlled [17,26], we considered it likely that mutation of the tat genes resulted in perturbed PQS signalling. This observation could also account, at least in part, for the reduced virulence of P. aeruginosa tat mutants in a rat pulmonary infection model [44]. For both P. aeruginosa PAO1 and PA14 strains pqsA expression and AQ production were reduced in the respective tat mutants while exogenous provision of either PQS or HHQ failed to fully restore pqsA expression in a tatA ΔpqsA mutant. The consequences of perturbed PQS signalling are clearly apparent in the altered expression profiles of the rhlA’-lux and phzA1’-lux fusions. Thus disruption of Tat clearly impairs the ability of P. aeruginosa either to fully induce PQS-dependent QS or to respond fully to exogenous PQS.
Since pyocyanin can be produced in the absence of PQS by ectopic expression of pqsE [26], it was possible that the tat phenotype was either a consequence of the inability of PQS/HHQ to activate the pqsABCDE operon via PqsR to generate sufficient PqsE protein or because the activity of PqsE depends on a functional Tat system. However, introduction of a plasmid-borne copy of pqsE into the tat mutants fully restored pyocyanin production suggesting that the tat mutant phenotype is not due to the inability of PqsE to function but rather a failure of the PQS auto-induction circuitry to produce sufficient PQS/HHQ to efficiently activate pqsABCDE.
To uncover a direct link between Tat and PQS signalling, we screened a bank of Tat substrate mutants for reduced pqsA expression and identified PetA as primarily responsible for the PQS signalling defect. As PetA is one of the three protein sub-units that constitute the cytochrome bc1 complex, we also showed that a similar PQS signalling defect was apparent in cytB or cytC1 mutants. Since growth of both the tat and cytochrome bc1 subunit mutants was reduced compared with the wild type (S9 Fig), our working hypothesis is that shutting down secondary metabolism and virulence factor production would aid conservation of energy for primary metabolism and growth. This shutdown could be efficiently achieved by preventing PQS autoinduction since it co-ordinates the expression of multiple genes involved in secondary metabolite production [17] including the biosynthesis of over 50 different AQs [54] that are heavily-dependent on tryptophan/anthranilate and fatty acid metabolic pathways. How PQS ‘shutdown’ is achieved at the molecular level in the tat mutants is not yet apparent but given that tat ΔpqsA double mutants do not respond to exogenous PQS, it may involve uptake into the cell and/or access to the response regulator PqsR(MvfR). At present there is little information on the mechanism by which PQS gains intracellular access although PQS is known to induce MV formation via a direct interaction with LPS and to be trafficked within MVs [33,34]. This is consistent with the reduction in MV production that we observed in the tat and pqsA mutants.
As shown in Figs 1 and S1, PQS biosynthesis generates a diverse series of AQs including the N-oxide, HQNO, a well known cytochrome bc1 inhibitor [57]. HQNO binds to the quinone reduction (Qi) site of the cytochrome bc1 complex disrupting the flow of electrons resulting in the leakage of reducing equivalents to oxygen [57]. This results in the generation of reactive oxygen species that cause P. aeruginosa cell death and autolysis favouring biofilm formation and antibiotic tolerance [57]. Rieske subunit and cytochrome bc1 mutants do not undergo autolysis and are insensitive to exogenous HQNO [57]. Similarly, P. aeruginosa pqsL mutants that are unable to produce HQNO also fail to undergo autolysis [57]. Our data also show that in common with the tat mutants, the petA as well as cytB and cytC1 mutants all produce lower levels of HQNO. This suggests that lack of eDNA in the tat mutant biofilms is because the cells do not undergo limited autolysis [57] as they lack the self-poisoning mechanism that depends on both an intact cytochrome bc1 and sufficient HQNO [57]. Thus, lack of the Rieske sub-unit export is primarily responsible for the Tat-mediated perturbation of PQS-dependent QS, the loss of virulence factor production, biofilm eDNA and the tobramycin tolerance of P. aeruginosa biofilms. Given the importance of PQS signalling and the Tat system to virulence and biofilm maturation in P. aeruginosa, our findings underline the potential of the Tat system as a drug target for novel antimicrobial agents.
Materials and methods
Bacterial strains and growth conditions
The P. aeruginosa and E. coli strains used are listed in S1 Table and were grown in LB or ABTG [8] at 37°C unless otherwise stated. P. aeruginosa biofilms were cultivated at 30°C in flow-chambers irrigated with FAB medium [58] supplemented with 0.3 mM glucose or in M9 medium with succinate (for static biofilms). Selective media were supplemented with ampicillin (Ap; 100 μg ml-1), gentamicin (Gm; 60 μg ml-1), or streptomycin (Sm; 100 μg ml-1). PQS, HHQ and 2-heptyl-4-hydroxyquinoline N-oxide (HQNO) were synthesized and characterized in house as described before [30, 59] and dissolved in methanol before being added to growth media at the appropriate concentration.
Mutant construction, screening and validation
A P. aeruginosa PAO1 Tn mutant library was constructed using the Mariner Tn vector pBT20 as previously described [60]. Transconjugants carrying Tn insertions were picked from the selective plates and inoculated into microtiter plates containing ABTG medium [61] supplemented with propidium iodide (PI) and the level of red fluorescence quantified. Mutants producing reduced levels of eDNA were selected and the sequences flanking the Tn insertion identified by arbitrary PCR essentially as described by Friedman and Kolter [62] but using the specific primers TnM1 and TnM2 (S2 Table). DNA Sequencing was performed by Macrogen, Seoul, Korea with primer TnMseq (S2 Table). An in-frame P. aeruginosa tatABC deletion mutant was generated by allelic exchange using the oligonucleotide primers Tat3D-UF, Tat3D-UR, Tat3D-DF and Tat3D-DR (S2 Table) to introduce the up- and down- stream regions of the tatABC locus into the suicide vector pME3087 to generate pME3087::tatABC (S1 Table). The latter was introduced into P. aeruginosa via conjugation with E. coli S17-1 λpir followed by enrichment for tetracycline-sensitive cells as described by Ye et al [63]. The ΔtatABC deletion in P. aeruginosa PAO1 was confirmed by PCR and sequence analysis. To generate the PA14 ΔpetA mutant, 500 bp upstream and downstream of the gene were amplified using respectively primers S.PA4431upFor/S.PA4431upRev and S.PA4431downFor/S.PA4431downRev listed in S2 Table. The PCR product was cloned in pKNG101 suicide vector by one-step sequence and ligation-independent cloning (SLIC) [64] and sequenced. The resulting plasmid, pKNGΔpetA, maintained in the E. coli CC118λpir strain, was then mobilized in P. aeruginosa strains. The mutant, in which the double recombination events occurred, was confirmed by PCR analysis. A similar strategy was used to construct allelic replacement mutants for each of 34 validated Tat substrates as well as the cytB and cytC1 mutants [45]. These strains are summarized in S3 Table and their validation will be described in detail elsewhere. For the generation of the petA cis-complemented strain, PA14ΔpetA::(CTX1::petA), the petA genes along with a 500 bp fragment corresponding to the putative promoter region for the petA gene were PCR amplified using S-4431CTXFor/S-4431CTXRev and cloned by SLIC into the mini-CTX1 vector yielding pminiCTX1-petA. Transfer of this plasmid in P. aeruginosa ΔpetA strain was carried out by triparental mating using the conjugative properties of the helper plasmid pRK2013. The recombinant clones containing the mini-CTX inserted at the attB locus on the P. aeruginosa genome were selected on tetracycline-containing PIA generating PA14ΔpetA::(attB::petA).
Construction of a tatA complementation plasmid
The tatA gene was amplified by PCR using primers FtatA and RtatA (S2 Table), introduced into pUCP22 (S1 Table) and electroporated into the P. aeruginosa tatA and tatA ΔpqsA mutants. Transformants were selected on LB plates containing 200 μg ml-1 carbenicillin.
Bioluminescent reporter assays
To investigate the impact of the tat mutation and Tat inhibitor Bayer 11–7082 [48] on PQS signalling, transcriptional fusions between the promoter regions of pqsA, pqsR, rhlA, phzA1 and phzA2 and the luxCDABE operon were constructed using the miniCTX-lux plasmid as previously described [17]. In addition, a constitutively bioluminescent reporter using a miniCTX::tac-luxCDABE promoter fusion was constructed as a control for Bayer 11–7082. Bioluminescence as a function of bacterial growth was quantified in 96 well plates using a combined luminometer- spectrometer.
Semi-quantification of cellular ATP was carried out using the BacTiter-GloTM Microbial Cell Viability Assay (Promega). Briefly, the P. aeruginosa PA14, ΔtatABC mutant and ΔpetA mutant were grown in LB broth for 8 h, diluted 1000-fold with fresh media and mixed with equal volume of BacTiter-GloTm reagent in a 96-well plate. After a 5 min incubation period, luminescence for each well was recorded in an automated plate reader.
Cultivation of biofilms
Biofilms were grown in flow-chambers with individual channel dimensions of 1 x 4 x 40 mm as described previously [65]. One hour after inoculation, with bacteria, the growth medium flow (0.2 mm/s corresponding to laminar flow with a Reynolds number of 0.02) was started. When required, eDNA in biofilms was stained with 1 μM ethidium bromide prior to microscopy. Tobramycin (10 μM) was added to the biofilm medium after 4 days of cultivation. After 24 h of tobramycin treatment, propidium iodide (10 μM) was added to the flow cells to visualize the dead cells via confocal laser scanning microscopy. P. aeruginosa PA14 and petA mutant biofilms were grown under static conditions over 48 h at 37°C on glass slides (Ibidi) incorporating 300 μl chambers. After 48 h incubation, spent medium was removed and the biofilm eDNA stained with YoYo-1 (40 μM).
Microscopy and image processing of flow cell biofilms
All images of flow-chamber-grown and static biofilms were captured with a confocal laser scanning microscope (CLSM) equipped with detectors and filter sets for monitoring green fluorescent protein, Syto9, propidium iodide, and ethidium bromide. Images were obtained using a 63x/1.4 objective or a 40x/1.3 objective. Simulated 3-D images and sections were generated using the IMARIS software package (Bitplane AG, Zürich, Switzerland).
AQ, pyocyanin, rhamnolipid and MV analysis
The AQs (PQS, HHQ and HQNO) were quantified by LC-MS/MS after extracting cell free supernatants or whole bacterial cell cultures in triplicate with acidified ethyl acetate or methanol respectively as described by Ortori et al [59]. Pyocyanin was extracted with chloroform and quantified spectrophotometrically [26]. Rhamnolipids were quantified indirectly using the orcinol method [35]. For PQS solubilization experiments, rhamnolipids were purified as described by Muller et al [66]. MVs were harvested by ultracentrifugation, the pellets resuspended in 10mM HEPES buffer and the lipid content quantified using FM4-64 essentially as described previously [67]. MV production was normalized by dividing the lipid fluorescence units by CFU values determined by dilution plating. Assays were performed in triplicate at least twice.
Statistical analysis
Significance for differences between wild type and isogenic mutants was determined by two-tailed t-tests where ****p < 0.001, ***p < 0.001, **p < 0.01, and *p < 0.05 and n.s., not significant.
Supporting information
PqsA catalyses the formation of anthraniloyl-CoA that is condensed with malonyl-CoA by PqsD to form 2-aminobenzoylacetyl-CoA (2-ABA-CoA). The latter is converted to 2-aminobenzoylacetate (2-ABA) via the thioesterase functionality of PqsE. The PqsBC heterodimer condenses 2-ABA with octanoyl-CoA to generate HHQ. PQS is formed through the oxidation of HHQ by PqsH. For AQ N-oxides such as 2-heptyl-4-hydroxyquinoline N-oxide (HQNO), 2-ABA is oxidized to 2-HABA by the alternative mono-oxygenase PqsL and then condensed with octanoyl-CoA by PqsBC to form HQNO. PqsBC can accept acyl-CoAs of different acyl chain lengths to generate diverse AQs and AQ N-oxides.
(TIF)
The co-solvent DMSO, had no effect at 0.4 or 0.8% on the lux reporter fusion Data are presented as maximal light output as a function of growth (RLU/OD600). Experiments were repeated in triplicate at least twice.
(TIF)
The data show that there are no differences in the expression of a chromosomal CTX::pqsR’-luxCDABE fusion in the P. aeruginosa wild type compared with the ΔtatABC mutant Data are presented as maximal light output as a function of growth (RLU/OD600). Experiments were repeated in triplicate at least twice.
(TIF)
Semi-quantitative analysis by LC-MS/MS of PQS (A) and HHQ (B) extracted with methanol from whole cell cultures of P. aeruginosa wild type and the tatA ΔpqsA mutant without (control) or with (+pqsABCD) the pqsABCD biosynthetic genes provided via pBBR1MCS-5::pqsABCD and harvested at 8 h and 16 h respectively. Experiments were repeated in triplicate.
(TIF)
Maximal light output as a function of growth (RLU/OD600) is presented. Experiments were repeated in triplicate at least twice.
(TIF)
Bioluminescence from a chromosomal rhlA’-lux fusion as a function of growth (RLU/OD) over time when introduced into (A) the PA14 wild type, (B) ΔtatABC and (C) ΔpetA mutants in the absence or presence of exogenous PQS (20 μM).
(TIF)
Biofilms of wild type PA14, the ΔtatABC and ΔpetA mutants and the genetically complemented PA14 mutant (ΔpetA::(CTX::petA)) were grown cultured statically and stained for eDNA with YOYO-1. (A) confocal fluorescence microscopy images and (B) eDNA quantification. Experiments were repeated in triplicate at least twice. ***p < 0.001, **p < 0.01.
(TIF)
Experiments were repeated in triplicate at least twice. ***p < 0.001, **p < 0.01, and *p < 0.05; n.s. not significant.
(TIF)
***p < 0.001, **p < 0.01, and *p < 0.05; n.s. not significant.
(TIF)
Experiments were repeated in triplicate at least twice.
(TIF)
(DOCX)
(DOCX)
(DOCX)
Acknowledgments
The authors wish to thank Alex Truman for synthesis of the AQ and AQ-N-oxide standards.
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported via grants to PW, MC and KH from the Biotechnology and Biological Sciences Research Council (BBSRC), U.K. (BB/F014392/1), BBSRC/National Biofilms Innovation Centre (BB/R012415/1), the Medical Research Council U.K. (MR/N501852/1), the Wellcome Trust (103884/Z/14/Z and 108876/Z/15/Z) and the European Union FP7 collaborative action grant (NABATIVI, 223670). BI was supported by a grant from Vaincre la Mucoviscidose and the Grégory Lemarchal associations (RF20140501138). TTN and MG were supported by grants from the Danish Strategic Research Council (9040-00023B), the Danish Council for Independent Research (09-065732), the Novo Nordisk Foundation (Biofilm2-AEEE/CPH) and the Lundbeck Foundation (2015‐486), Denmark. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Moradali MF, Ghods S, Rehm BH. Pseudomonas aeruginosa Lifestyle: A paradigm for adaptation, survival, and persistence. Front Cell Infect Microbiol. 2017; 7:39–42. doi: 10.3389/fcimb.2017.00039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science. 1999; 284:1318–22. doi: 10.1126/science.284.5418.1318 . [DOI] [PubMed] [Google Scholar]
- 3.Klausen M, Aaes-Jørgensen A, Molin S, Tolker-Nielsen T. Involvement of bacterial migration in the development of complex multicellular structures in Pseudomonas aeruginosa biofilms. Mol Microbiol. 2003; 50:61–8. doi: 10.1046/j.1365-2958.2003.03677.x . [DOI] [PubMed] [Google Scholar]
- 4.Klausen M, Heydorn A, Ragas P, Lambertsen L, Aaes-Jørgensen A, Molin S, et al. Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol Microbiol. 2003; 48:1511–24. doi: 10.1046/j.1365-2958.2003.03525.x . [DOI] [PubMed] [Google Scholar]
- 5.Pamp SJ, Tolker-Nielsen T. Multiple roles of biosurfactants in structural biofilm development by Pseudomonas aeruginosa. J Bacteriol. 2007; 189:2531–39. doi: 10.1128/JB.01515-06 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Harmsen M, Yang L, Pamp SJ, Tolker-Nielsen T. An update on Pseudomonas aeruginosa biofilm formation, tolerance, and dispersal. FEMS Immunol Med Microbiol. 2010; 59:253–68. doi: 10.1111/j.1574-695X.2010.00690.x . [DOI] [PubMed] [Google Scholar]
- 7.Yang L, Hu Y, Liu Y, Zhang J, Ulstrup J, Molin S. Distinct roles of extracellular polymeric substances in Pseudomonas aeruginosa biofilm development. Environ Microbiol. 2011; 13:1705–17. doi: 10.1111/j.1462-2920.2011.02503.x . [DOI] [PubMed] [Google Scholar]
- 8.Allesen-Holm M, Barken KB, Yang L, Klausen M, Webb JS, Kjelleberg S, et al. A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol Microbiol. 2006; 59:1114–28. doi: 10.1111/j.1365-2958.2005.05008.x . [DOI] [PubMed] [Google Scholar]
- 9.Tashiro Y, Uchiyama H, Nomura N. Multifunctional membrane vesicles in Pseudomonas aeruginosa. Environ Microbiol. 2012; 14:1349–62. doi: 10.1111/j.1462-2920.2011.02632.x . [DOI] [PubMed] [Google Scholar]
- 10.Tolker-Nielsen T. Biofilm Development. Microbiol Spectr. 20153:MB-0001–2014. doi: 10.1128/microbiolspec.MB-0001-2014 . [DOI] [PubMed] [Google Scholar]
- 11.Okshevsky M, Meyer RL. The role of extracellular DNA in the establishment, maintenance and perpetuation of bacterial biofilms. Crit Rev Microbiol. 2015;41:341–52. doi: 10.3109/1040841X.2013.841639 . [DOI] [PubMed] [Google Scholar]
- 12.Das T, Sehar S, Manefield M. The roles of extracellular DNA in the structural integrity of extracellular polymeric substance and bacterial biofilm development. Environ Microbiol Rep. 20135:778–86. doi: 10.1111/1758-2229.12085 . [DOI] [PubMed] [Google Scholar]
- 13.Wilton M, Charron-Mazenod L, Moore R, Lewenza S. Extracellular DNA acidifies biofilms and induces aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2015; 60:544–53. doi: 10.1128/AAC.01650-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Turnbull L, Toyofuku M, Hynen AL, Kurosawa M, Pessi G, Petty NK, et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat Commun. 2016; 7:11220. doi: 10.1038/ncomms11220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Das T, Kutty SK, Tavallaie R, Ibugo AI, Panchompoo J, Sehar S, et al. Phenazine virulence factor binding to extracellular DNA is important for Pseudomonas aeruginosa biofilm formation. Sci Rep. 2015; 5:8398. doi: 10.1038/srep08398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Williams P, Cámara M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: a tale of regulatory networks and multifunctional signal molecules. Curr Opin Microbiol. 2009; 12:182–91. doi: 10.1016/j.mib.2009.01.005 . [DOI] [PubMed] [Google Scholar]
- 17.Rampioni G, Falcone M, Heeb S, Frangipani E, Fletcher MP, Dubern JF, et al. Unravelling the genome-wide contributions of specific 2-alkyl-4-quinolones and PqsE to quorum sensing in Pseudomonas aeruginosa. PLoS Pathog. 2016; 12:e1006029. doi: 10.1371/journal.ppat.1006029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lépine F, Milot S, Déziel E, He J, Rahme LG. Electrospray/mass spectrometric identification and analysis of 4-hydroxy-2-alkylquinolines (HAQs) produced by Pseudomonas aeruginosa. J Am Soc Mass Spectrom. 2004; 15:862–9. doi: 10.1016/j.jasms.2004.02.012 . [DOI] [PubMed] [Google Scholar]
- 19.Coleman JP, Hudson LL, McKnight SL, Farrow JM 3rd, Calfee MW, Lindsey CA, et al. Pseudomonas aeruginosa PqsA is an anthranilate-coenzyme A ligase. J Bacteriol. 2008; 190:1247–55. doi: 10.1128/JB.01140-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhang YM, Frank MW, Zhu K, Mayasundari A, Rock CO. PqsD is responsible for the synthesis of 2,4-dihydroxyquinoline, an extracellular metabolite produced by Pseudomonas aeruginosa. J Biol Chem. 2008; 283:28788–94. doi: 10.1074/jbc.M804555200 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Drees SL, Fetzner S. PqsE of Pseudomonas aeruginosa acts as pathway-specific thioesterase in the biosynthesis of alkylquinolone signaling molecules. Chem Biol. 2015; 22:611–8. doi: 10.1016/j.chembiol.2015.04.012 . [DOI] [PubMed] [Google Scholar]
- 22.Dulcey CE, Dekimpe V, Fauvelle DA, Milot S, Groleau MC, Doucet N, et al. The end of an old hypothesis: the pseudomonas signaling molecules 4-hydroxy-2-alkylquinolines derive from fatty acids, not 3-ketofatty acids. Chem Biol. 2013; 20:1481–91. doi: 10.1016/j.chembiol.2013.09.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Drees SL, Li C, Prasetya F, Saleem M, Dreveny I, Williams P, Hennecke U, et al. PqsBC, a condensing enzyme in the biosynthesis of the Pseudomonas aeruginosa quinolone signal: crystal structure, inhibition, and reaction mechanism. J Biol Chem. 2016; 291:6610–24. doi: 10.1074/jbc.M115.708453 ; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Schertzer JW, Brown SA, Whiteley M. Oxygen levels rapidly modulate Pseudomonas aeruginosa social behaviours via substrate limitation of PqsH. Mol Microbiol. 2010; 77:1527–38. doi: 10.1111/j.1365-2958.2010.07303.x Epub 2010 Aug 17. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Drees SL, Ernst S, Belviso BD, Jagmann N, Hennecke U, Fetzner S. PqsL uses reduced flavin to produce 2-hydroxylaminobenzoylacetate, a preferred PqsBC substrate in alkyl quinolone biosynthesis in Pseudomonas aeruginosa. J Biol Chem. 2018. ’ 15: 293:9345–9357. doi: 10.1074/jbc.RA117.000789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Rampioni G, Pustelny C, Fletcher MP, Wright VJ, Bruce M, Rumbaugh KP, et al. Transcriptomic analysis reveals a global alkyl-quinolone-independent regulatory role for PqsE in facilitating the environmental adaptation of Pseudomonas aeruginosa to plant and animal hosts. Environ Microbiol. 2010; 12:1659–73. doi: 10.1111/j.1462-2920.2010.02214.x . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Groleau MC, de Oliveira Pereira T, Dekimpe V, Déziel E. PqsE Is Essential for RhlR-Dependent Quorum Sensing Regulation in Pseudomonas aeruginosa mSystems. 2020; 5:e00194–20; doi: 10.1128/mSystems.00194-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wade DS, Calfee MW, Rocha ER, Ling EA, Engstrom E, Coleman JP, et al. Regulation of pseudomonas quinolone signal synthesis in Pseudomonas aeruginosa. J Bacteriol. 2005;187:4372–80. doi: 10.1128/JB.187.13.4372-4380.2005 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Xiao G, Déziel E, He J, Lépine F, Lesic B, Castonguay MH, et al. MvfR, a key Pseudomonas aeruginosa pathogenicity LTTR-class regulatory protein, has dual ligands. Mol Microbiol. 2006; 62:1689–99. doi: 10.1111/j.1365-2958.2006.05462.x . [DOI] [PubMed] [Google Scholar]
- 30.Ilangovan A, Fletcher M, Rampioni G, Pustelny C, Rumbaugh K, Heeb S, et al. Structural basis for native agonist and synthetic inhibitor recognition by the Pseudomonas aeruginosa quorum sensing regulator PqsR (MvfR). PLoS Pathog. 2013; 9:e1003508. doi: 10.1371/journal.ppat.1003508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Diggle SP, Matthijs S, Wright VJ, Fletcher MP, Chhabra SR, Lamont IL, et al. The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem Biol. 2007;14:87–96. doi: 10.1016/j.chembiol.2006.11.014 . [DOI] [PubMed] [Google Scholar]
- 32.Häussler S, Becker T. The pseudomonas quinolone signal (PQS) balances life and death in Pseudomonas aeruginosa populations. PLoS Pathog. 2008; 4:e1000166. doi: 10.1371/journal.ppat.1000166; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mashburn-Warren L, Howe J, Garidel P, Richter W, Steiniger F, Roessle M, et al. Interaction of quorum signals with outer membrane lipids: insights into prokaryotic membrane vesicle formation. Mol Microbiol. 200869:491–502. doi: 10.1111/j.1365-2958.2008.06302.x . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mashburn LM, Whiteley M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature. 2005; 437:422–5. doi: 10.1038/nature03925 [DOI] [PubMed] [Google Scholar]
- 35.Pustelny C, Albers A, Büldt-Karentzopoulos K, Parschat K, Chhabra SR, Cámara M, et al. Dioxygenase-mediated quenching of quinolone-dependent quorum sensing in Pseudomonas aeruginosa. Chem Biol. 2009; 16:1259–67. doi: 10.1016/j.chembiol.2009.11.013 . [DOI] [PubMed] [Google Scholar]
- 36.D’Argenio DA, Calfee MW, Rainey PB, Pesci EC. Autolysis and autoaggregation in Pseudomonas aeruginosa colony morphology mutants. J Bacteriol. 2002; 184:6481–9. doi: 10.1128/JB.184.23.6481-6489.2002 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Webb JS, Thompson LS, James S, Charlton T, Tolker-Nielsen T, Koch B, et al. Cell death in Pseudomonas aeruginosa biofilm development. J Bacteriol. 2003; 185:4585–92. doi: 10.1128/JB.185.15.4585-4592.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Spoering AL, Gilmore MS. Quorum sensing and DNA release in bacterial biofilms. Curr Opin Microbiol. 2006; 9:133–7. doi: 10.1016/j.mib.2006.02.004 . [DOI] [PubMed] [Google Scholar]
- 39.Yang L, Barken KB, Skindersoe ME, Christensen AB, Givskov M, Tolker-Nielsen T. et al. Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa. Microbiology. 2007;153:1318–28. doi: 10.1099/mic.0.2006/004911-0 . [DOI] [PubMed] [Google Scholar]
- 40.Yang L, Nilsson M, Gjermansen M, Givskov M, Tolker-Nielsen T. Pyoverdine and PQS mediated subpopulation interactions involved in Pseudomonas aeruginosa biofilm formation. Mol Microbiol. 2009;74:1380–92. doi: 10.1111/j.1365-2958.2009.06934.x . [DOI] [PubMed] [Google Scholar]
- 41.Suzuki T, Fujikura K, Higashiyama T, Takata K. DNA staining for fluorescence and laser confocal microscopy. J Histochem Cytochem. 1997; 45:49–53. doi: 10.1177/002215549704500107 . [DOI] [PubMed] [Google Scholar]
- 42.Frain KM, Robinson C, van Dijl JM. Transport of folded proteins by the tat system. Protein J. 2019; 38:377–388. doi: 10.1007/s10930-019-09859-y . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Voulhoux R, Ball G, Ize B, Vasil ML, Lazdunski A, Wu LF, et al. Involvement of the twin-arginine translocation system in protein secretion via the type II pathway. EMBO J. 2001; 20: 6735–6741. doi: 10.1093/emboj/20.23.6735 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ochsner UA, Snyder A, Vasil AI, Vasil ML. Effects of the twin-arginine translocase on secretion of virulence factors, stress response, and pathogenesis. Proc Natl Acad Sci U S A. 2002; 99:8312–8317. doi: 10.1073/pnas.082238299 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gimenez MR, Chandra G, Van Overvelt P, Voulhoux R, Ize B, Bleves S. et al. Genome wide identification and experimental validation of Pseudomonas aeruginosa Tat substrates. Sci Rep 2018; 11950. doi: 10.1038/s41598-018-30393-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Calfee MW, Shelton JG, McCubrey JA, Pesci EC. Solubility and bioactivity of the Pseudomonas quinolone signal are increased by a Pseudomonas aeruginosa-produced surfactant. Infect Immun. 2005; 73:878–82. doi: 10.1128/IAI.73.2.878-882.2005 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chiang WC, Nilsson M, Jensen PØ, Høiby N, Nielsen TE, Givskov M, et al. Extracellular DNA shields against aminoglycosides in Pseudomonas aeruginosa biofilms. Antimicrob Agents Chemother. 2013; 57:2352–61. doi: 10.1128/AAC.00001-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Vasil ML, Tomaras AP, Pritchard AE. Identification and evaluation of twin-arginine translocase inhibitors. Antimicrob Agents Chemother. 2012; 56:6223–34. doi: 10.1128/AAC.01575-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Williams P, Winzer K, Chan WC, Cámara M. Look who’s talking: communication and quorum sensing in the bacterial world. Philos Trans R Soc Lond B Biol Sci. 2007; 362:1119–34. doi: 10.1098/rstb.2007.2039 ; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Recinos DA, Sekedat MD, Hernandez A, Cohen TS, Sakhtah H, Prince AS, et al. Redundant phenazine operons in Pseudomonas aeruginosa exhibit environment-dependent expression and differential roles in pathogenicity. Proc Natl Acad Sci U S A. 2012; 109:19420–5. doi: 10.1073/pnas.1213901109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Niewerth H, Bergander K, Chhabra SR, Williams P, Fetzner S. Synthesis and biotransformation of 2-alkyl-4(1H)-quinolones by recombinant Pseudomonas putida KT2440. Appl Microbiol Biotechnol. 2011; 91:1399–408. doi: 10.1007/s00253-011-3378-0 . [DOI] [PubMed] [Google Scholar]
- 52.Van Gennip M, Christensen LD, Alhede M, Phipps R, Jensen PØ, Christophersen L, et al. Inactivation of the rhlA gene in Pseudomonas aeruginosa prevents rhamnolipid production, disabling the protection against polymorphonuclear leukocytes. APMIS. 2009117:537–46. doi: 10.1111/j.1600-0463.2009.02466.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Alhede M, Bjarnsholt T, Jensen PØ, Phipps RK, Moser C, Christophersen L, et al. Pseudomonas aeruginosa recognizes and responds aggressively to the presence of polymorphonuclear leukocytes. Microbiology (Reading). 2009155:3500–3508. doi: 10.1099/mic.0.031443-0 . [DOI] [PubMed] [Google Scholar]
- 54.Déziel E, Lépine F, Milot S, He J, Mindrinos MN, Tompkins RG, et al. Analysis of Pseudomonas aeruginosa 4-hydroxy-2-alkylquinolines (HAQs) reveals a role for 4-hydroxy-2-heptylquinoline in cell-to-cell communication. Proc Natl Acad Sci U S A. 2004; 101:1339–44. doi: 10.1073/pnas.0307694100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Farrow JM 3rd, Sund ZM, Ellison ML, Wade DS, Coleman JP, Pesci EC. et al. PqsE functions independently of PqsR-Pseudomonas quinolone signal and enhances the rhl quorum-sensing system. J Bacteriol. 2008; 190:7043–51. doi: 10.1128/JB.00753-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Williams HD, Zlosnik JE, Ryall B. Oxygen, cyanide and energy generation in the cystic fibrosis pathogen Pseudomonas aeruginosa. Adv Microb Physiol. 2007;52:1–71. doi: 10.1016/S0065-2911(06)52001-6 . [DOI] [PubMed] [Google Scholar]
- 57.Hazan R, Que YA, Maura D, Strobel B, Majcherczyk PA, Hopper LR, et al. Auto poisoning of the respiratory chain by a quorum-sensing-regulated molecule favors biofilm formation and antibiotic tolerance. Curr Biol. 2016; 26:195–206. doi: 10.1016/j.cub.2015.11.056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Heydorn A, Nielsen AT, Hentzer M, Sternberg C, Givskov M, Ersbøll BK, et al. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology. 2000;146:2395–407. doi: 10.1099/00221287-146-10-2395 . [DOI] [PubMed] [Google Scholar]
- 59.Ortori CA, Dubern JF, Chhabra SR, Cámara M, Hardie K, Williams P, et al. Simultaneous quantitative profiling of N-acyl-L-homoserine lactone and 2-alkyl-4(1H)-quinolone families of quorum-sensing signaling molecules using LC-MS/MS. Anal Bioanal Chem. 2011; 399:839–50. doi: 10.1007/s00216-010-4341-0 . [DOI] [PubMed] [Google Scholar]
- 60.Kulasekara HD, Ventre I, Kulasekara BR, Lazdunski A, Filloux A, Lory S. et al. A novel two-component system controls the expression of Pseudomonas aeruginosa fimbrial cup genes. Mol Microbiol. 2005Jan;55(2):368–80. doi: 10.1111/j.1365-2958.2004.04402.x . [DOI] [PubMed] [Google Scholar]
- 61.Chua SL, Tan SY, Rybtke MT, Chen Y, Rice SA, Kjelleberg S, et al. Bis-(3’-5’)-cyclic dimeric GMP regulates antimicrobial peptide resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2013; 57:2066–75. doi: 10.1128/AAC.02499-12 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Friedman L, Kolter R. Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Mol Microbiol. 2004; 51:675–90. doi: 10.1046/j.1365-2958.2003.03877.x . [DOI] [PubMed] [Google Scholar]
- 63.Ye RWl, Haas D, Ka JO, Krishnapillai V, Zimmermann A, Baird C, et al. Anaerobic activation of the entire denitrification pathway in Pseudomonas aeruginosa requires Anr, an analog of Fnr. J Bacteriol. 1995; 177:3606–9. doi: 10.1128/jb.177.12.3606-3609.1995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Jeong JY, Yim HS, Ryu JY, Lee HS, Lee JH, Seen DS, et al. One-step sequence- and ligation-independent cloning as a rapid and versatile cloning method for functional genomics studies. Appl Environ Microbiol. 2012; 78:5440–3. doi: 10.1128/AEM.00844-12 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Crusz SA, Popat R, Rybtke MT, Cámara M, Givskov M, Tolker-Nielsen T, et al. Bursting the bubble on bacterial biofilms: a flow cell methodology. Biofouling. 2012; 28:835–42. doi: 10.1080/08927014.2012.716044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Müller MM, Hörmann B, Syldatk C, Hausmann R. Pseudomonas aeruginosa PAO1 as a model for rhamnolipid production in bioreactor systems. Appl Microbiol Biotechnol. 2010; 87:167–74. doi: 10.1007/s00253-010-2513-7 . [DOI] [PubMed] [Google Scholar]
- 67.McBroom AJ, Johnson AP, Vemulapalli S, Kuehn MJ. Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J Bacteriol. 2006; 188:5385–92. doi: 10.1128/JB.00498-06 . [DOI] [PMC free article] [PubMed] [Google Scholar]