

Abstract
The integrity and permeability of the intestinal epithelial barrier are important indicators of intestinal health. Impaired intestinal epithelial barrier function and increased intestinal permeability are closely linked to the onset and progression of various intestinal diseases. Sinapic acid (SA) is a phenolic acid that has anti-inflammatory, antihyperglycemic, and antioxidant activities; meanwhile, it is also effective in the protection of inflammatory bowel disease (IBD), but the specific mechanisms remain unclear. Here, we evaluated the anti-inflammatory of SA and investigated its potential therapeutic activity in LPS-induced intestinal epithelial barrier and tight junction (TJ) protein dysfunction. SA improved cell viability; attenuated epithelial permeability; restored the protein and mRNA expression of claudin-1, ZO-1, and occludin; and reversed the redistribution of the ZO-1 and claudin-1 proteins in LPS-treated Caco-2 cells. Moreover, SA reduced the inflammatory response by downregulating the activation of the TLR4/NF-κB pathway and attenuated LPS-induced intestinal barrier dysfunction by decreasing the activation of the MLCK/MLC pathway. This study demonstrated that SA has strong anti-inflammatory activity and can alleviate the occurrence of high intercellular permeability in Caco-2 cells exposed to LPS.
1. Introduction
Intestinal epithelial cells are the key components of the epithelial lining. The intact intestinal epithelial maintains the intestinal physical barrier and plays a critical role in the body's defense functions [1]. Changes in intestinal epithelial barrier permeability incite mucosal inflammation leading to intestinal diseases, such as inflammatory bowel disease (IBD), including Crohn's disease (CD) and ulcerative colitis (UC), irritable bowel syndrome (IBS), and colon cancer (CRC) [2–4]. Intestinal epithelial barriers are formed by tight junction (TJ) proteins, including occludin, claudin, and zonula occludens (ZO), that connect the cytoskeleton and signalling molecules [5]. Intestinal inflammation has been proved to be associated with the intestinal epithelial barrier disruption [6]. Inflammatory stimulation and other endogenous cytokines directly affect the intestinal epithelial barrier by reducing the localization and expression of TJ proteins [7]. Previous studies have indicated that maintenance of the intestinal epithelial barrier and attenuated inflammatory responses is an efficient strategy for the treatment of IBD [8–11].
Lipopolysaccharide (LPS), an important risk factor of inflammation, is widely used in many researches about intestinal tight junction barriers. Stimulation of intestinal cells by LPS induces increased Toll-like receptor-4 (TLR4) expression and triggers the release of proinflammatory mediators [12–14]. What is more, studies suggest that the increase in intestinal epithelial TJ permeability is mediated by the upregulation of myosin light chain kinase (MLCK) by the TLR4/MyD88 and NF-κB signalling pathway [15, 16].
Numerous phenolic compounds of plant origin have been shown to alleviate inflammation and improve intestinal permeability due to their anti-inflammatory and antioxidant ability [17–21]. SA is a well-known phenolic acid that is found in various herbal materials, fruits, and grains, as well as in some vegetables [22, 23]. Modern pharmacological studies have reported that the SA possesses several pharmacological properties including antioxidant [24], anti-inflammatory [22, 25], antihyperglycemic, hypoglycemic [26], and anticancer activities [27]. Although SA has a potential protective role in colitis mice [25], its role in intestinal barrier remains unclear. In view of the important role of TJ proteins in the intestinal epithelial barrier [28], it is necessary for us to investigate the effect of SA on TJ proteins as well as intestinal epithelial barrier. Collectively, this study is aimed at investigating the effects of SA treatment on anti-inflammation and the localization and expression of TJ in an LPS-induced Caco-2 model of inflammation-mediated barrier dysfunction.
2. Materials and Methods
2.1. Materials and Reagents
Caco-2 cells (purchased from China National Collection of Authenticated Cell Cultures, SCSP-5027) were a gift obtained from Professor Xian-qiong Zou, School of Biotechnology, Guilin Medical University. Sinapic acid (SA) was obtained from Aladdin Biochemical Technology Co., Ltd. (Shanghai, China). Nonessential amino acid (NEAA) solution and LPS were obtained from Solarbio Life Sciences Co., Ltd. (Beijing, China). TRNzol-A+ reagent, FastQuant RT Kit, and SuperReal PreMix Plus (SYBR Green) reagent were obtained from Tiangen Biotech Co., Ltd. (Beijing, China). Triton X-100, fix solution (4% paraformaldehyde), bovine serum albumin (BSA), goat serum (C0265), and other reagents were obtained from Beyotime Biotechnology Co., Ltd. (Shanghai, China). Primary antibodies against ZO-1 (AF8394), occludin (AF7644), claudin-1 (AF6504), NF-κB (AF7569), phospho-NF-κBp65 (AN371), phospho-IκBα (Ser32) (AF5851), phospho-IKKα/β (Ser176/180) (AI139), MyD88 (AF2116), TLR4 (AF8187), and β-actin (AF0003) were purchased from Beyotime Biotechnology Co., Ltd.
2.2. Cell Culture and Treatment
Caco-2 cells were routinely cultured at 37°C in in a humidified chamber of 5% CO2 in high glucose Dulbecco's Modified Eagle's Medium (DMEM) containing 10% fetal bovine serum (FBS), 1% NEAA, and 1% penicillin/streptomycin. Then, the cells were subcultured at 80–90% confluence. In all experiments, Caco-2 cells were coincubated with LPS (10 μg/ml) in the presence or absence of SA (5, 10, or 15 μmol/l) and incubated for 24 h.
2.3. Cell Viability Assay
The MTT assay was used to evaluate the cell viability of SA. The Caco-2 cells were incubated with SA (5, 10, or 15 μmol/l) for 24 h or 48 h. After that, MTT solution (5.0 mg/ml) was added for a further 4 h. The absorbance of the MTT-formazan product was read at 490 nm after dissolving with 150 μl dimethyl sulfoxide (DMSO)/well; each sample was analyzed in quintuplicate (n = 3).
2.4. Epithelial Permeability Assay
The TJ permeability assays used in this study were the transepithelial electrical resistance (TEER) assay and the fluorescein isothiocyanate- (FITC-) conjugated dextran probe (FD-40) assay. For the TEER and FD-40 assays, monolayer Caco-2 cells were seeded and cultured on 12-well inserts (pore size 0.4 μm; Millipore, Bedford, MA, USA). Then, the Caco-2 cells were treated according to cell culture and treatment groups. The TEER value was determined using an ERS-2 voltohmmeter (Millipore) according to the manufacturer's instructions. For the FD-40 assay, the Caco-2 cells were washed with HBSS (pH 7.4) three times, and FD-40 (1 mg/ml) was subsequently applied to the apical side. After incubation for 2 h at 37°C, the media (100 μl) were collected from the apical to basolateral side, and the fluorescent absorbance was measured by a with ex/em 480/530 nm.
2.5. Assessment of NF-κB Nuclear Translocation
Caco-2 cells were seeded in 8-well cover slips (Thermo Fisher, USA). Then, the Caco-2 cells were treated according to cell culture and treatment groups. The slides were fixed and blocked (with 10% goat serum) at 25°C. The first antibodies NF-κBp65 were used to incubate the slides (overnight at 4°C). After being washed with PBS 3 times, the slides were incubated with fluorescent secondary antibody at 25°C for 1 h. Following PBS washing 3 times, the incubated Caco-2 cells were stained with 4′,6-diamidino-2-phenylindole (DAPI) solution to visualize nuclei. Finally, all images were obtained using a microscope (Leica Microsystems Inc., Buffalo Grove, IL, USA). Cellular characteristics were observed by a microscope with a magnification (50x).
2.6. Quantitative Reverse-Transcription- (qRT-) PCR Assay
After different treatment, total RNA was extracted from the Caco-2 cells using a TRIzol-A+ reagent. Extracted RNA reverse transcribed into cDNA according to the kit protocols (Tiangen, Beijing, China). qRT-PCR analysis reacted using QuantStudio™ 6 Flex Real-Time PCR System (Life Technologies, USA) with conditions set to 95°C for 3 min for the initial denaturation and followed by 35~50 cycles with denaturation at 95°C for 10 s, annealing at 56°C for 10 s, and extension at 72°C for 60 s. Based on the 2-ΔΔ Ct formula, we calculated relative mRNA levels. The sequences of primer are listed in Table 1.
Table 1.
Primers used to measure mRNA expression.
| Gene | Forward | Reverse |
|---|---|---|
| Gapdh | CCATTTGATGTTAGCGGGATCTC | TGGTCTACATGTTCCAGTATGACT |
| Tlr4 | GTACCTGGGGAACAACCTCTT | GCAGCTTGACTAGACTCTCCA |
| Nfκbp65 | GTGGGGACTACGACCTGAATG | GGGGCACGATTGTCAAAGATG |
| Il1β | GAATGACGCCCTCAATCAAAGT | TCATCTTGGGCAGTCACATACA |
| Il8 | CCTGAACCTTCCAAAGATGGC | TTCACCAGGCAAGTCTCCTCA |
| Mlck | CAACAGGGTCACCAACCAGC | GCCTTGCAGGTGTACTTGGC |
| Occludin | CTTCCAATGGCAAAGTGAATG | TACCACCGCTGCTGTAACGAG |
| Claudin-1 | CCAGGTACGAATTTGGTCAGG | TGGTGTTGGGTAAGAGGTTGT |
| Zo1 | GAGCCTAATCTGACCTATGAACC | TGAGGACTCGTATCTGTATGTGG |
2.7. Protein Extraction and Western Blot Analysis
Caco-2 cells were seeded in 8-well cover slips (Thermo Fisher, USA). Then, the cells were treated according to cell culture and treatment groups. Total proteins from Caco-2 cells were extracted after different treatments using cell lysis buffer for western blotting and IP reagent (Beyotime). The concentration of the extracted proteins was determined using the BCA assay, and the samples were denatured by boiling (100°C) for further study. The total protein samples (30 μg) were separated by SDS-PAGE gel for 90-120 min and then transferred to a PVDF membranes (Millipore). The membrane with separated proteins was blocked with nonfat milk (5%) for 2 h and immediately incubated with first antibodies (overnight at 4°C) TLR4, IKKα, p-IKKα, MyD88, p-NF-κBp65, IκB, and p-IκB. After washed with 3 times of PBS, all membranes were incubated with the accordingly secondary antibody for 60 min at 25°C. The blots were visualized with ECL detection reagents (7sea Biotech, Shanghai, China), and the ImageJ software (NIH, Bethesda, MD, America) was employed to numeralization for band analysis (https://imagej.nih.gov/ij/).
2.8. Immunofluorescent Localization of TJ Proteins
Localization of the TJ proteins (ZO-1 and claudin-1) was analyzed with immunofluorescence staining. For immunofluorescence staining, the non-SA-treated and SA-treated Caco-2 cells were fixed with fix solution for 15 min and then infiltrated with Triton X-100 for 15 min. Then, the cells were washed and blocked for 1 h with 5% BSA in PBS. After 1 h of blocking with 5% BSA, the cells were incubated with first antibodies (claudin-1 and ZO-1) overnight at 4°C. All cells were then incubated with the corresponding secondary antibodies and stained with DAPI solution for 3 min at room temperature. Images were obtained using a microscope (Leica Microsystems Inc., Buffalo Grove, IL, USA). Cellular characteristics were observed by a microscope with a magnification (100x).
2.9. Statistical Analysis
The mean ± standard deviation (SD) is used to describe data. The SPSS 25.0 software (SPSS Inc., Chicago, USA) was employed. One-way ANOVA and Duncan's multiple range tests were used to determine statistically significant differences between the treatments (p < 0.05). The GraphPad Prism version 5.0 statistical software package was used for the analysis.
3. Results
3.1. Toxicity Assay in SA-Treated Caco-2 Cells
The cell viability after 24 h and 48 h of SA treatment (ranging from 1 μmol/l to 20 μmol/l) is shown in Figure 1. SA treatment for 24 h showed no significant effect on cell viability at any concentration compared to the control conditions. Besides, the cell viability in Caco-2 cells was significantly inhibited by SA treatment for 48 h, which effect was in a concentration-dependent manner (p < 0.05). A high concentration of SA (20 μmol/l) showed cytotoxicity in Caco-2 cells after 48 h of treatment. Therefore, we selected 5, 10, and 15 μmol/l SA as safe concentration for further study in Caco-2 cells.
Figure 1.
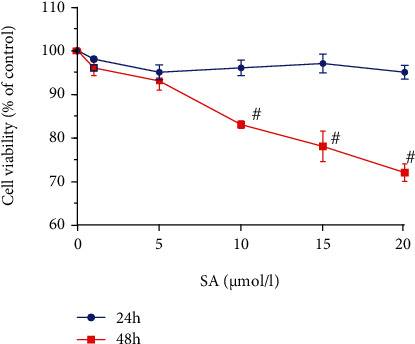
Effect of sinapic acid (SA) on the viability of Caco-2 cells. The viability of Caco-2 cells was determined after treatment with SA (1, 5, 10, 15, or 20 μmol/l) for 24 h and 48 h. The results are expressed as the mean ± SD of three independent experiments. # denotes p < 0.05 vs. the non-SA-treated cells.
3.2. Effects of SA on Epithelial Permeability in LPS-Treated Caco-2 Cells
As shown in Figure 2, LPS treatment induced a significant decrease in TEER values in Caco-2 cells (p < 0.05). Administration of SA effectively increased the TEER values in LPS-treated Caco-2 cells. However, SA concentration of 10 and 15 μmol/l led to significantly higher TEER values in Caco-2 cells than the low concentration of SA (5 μmol/l) (p < 0.05). In addition, SA induced a concentration-dependent decrease in FD-40 permeability in LPS-treated Caco-2 cells.
Figure 2.
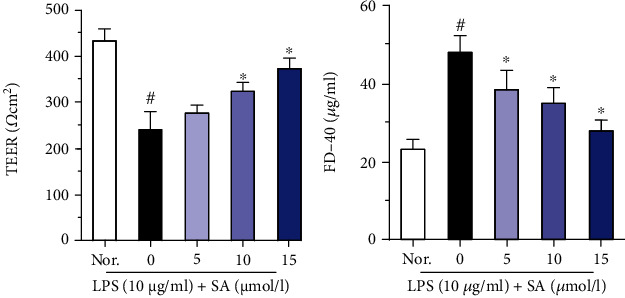
Effects of sinapic acid (SA) on the TEER and FD-40 levels in LPS-treated Caco-2 cells. The TEER and FD-40 levels were determined after treatment with SA (5, 10, or 15 μmol/l) for 24 h in LPS-treated Caco-2 cells. The results are expressed as the mean ± SD of three independent experiments. # denotes p < 0.05 vs. the normal control group (Nor.), and ∗ denotes p < 0.05 vs. the LPS group.
3.3. Effects of SA on NF-κB Nuclear Translocation in LPS-Treated Caco-2 Cells
As shown in Figure 3, NF-κBp65 was present in the cytoplasm of untreated Caco-2 cells. LPS treatment markedly increased the activation of nuclear NF-κBp65 and promoted its translocation (p < 0.05). The SA-treated group showed significantly lower translocation of nuclear NF-κBp65 than that in the LPS group (NF-κBp65 nuclear translocation rate: 45.45%), and the effect was most significant in the group treated with 15 μmol/l SA (NF-κBp65 nuclear translocation rate: 18.18%).
Figure 3.
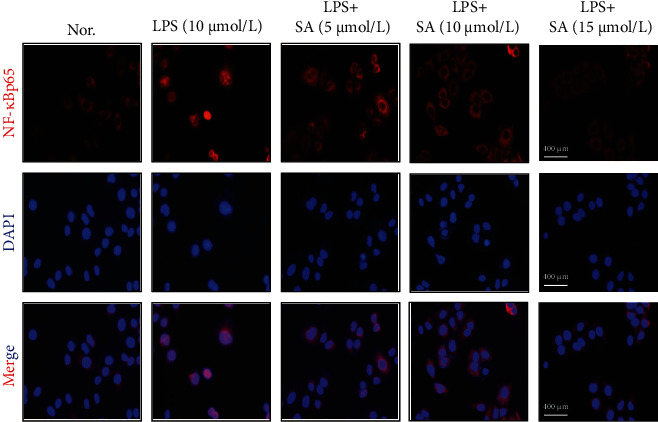
Effect of sinapic acid (SA) on NF-κBp65 translocation in LPS-treated Caco-2 cells. Caco-2 cells were incubated with LPS (10 μg/ml) and SA (5, 10, or 15 μmol/l) for 24 h and then subjected to immunofluorescence analysis (50x).
3.4. Effects of SA on the mRNA Levels of Tlr4, Nfκbp65, Il1β, and Il8 in LPS-Treated Caco-2 Cells
IL-1β and IL-8 are typical inflammatory cytokines that mediate and promote the inflammatory response. The mRNA levels of Tlr4, Nfκbp65, Il1β, and Il8 in Caco-2 cells were analyzed by qRT-PCR assay. Treatment with LPS significantly increased the mRNA expression of Tlr4, Nfκbp65, Il1β, and Il8 compared with no treatment (p < 0.05; Figure 4). However, this effect was mitigated by treatment with different concentrations of SA (p < 0.05). This finding suggests that SA can inhibit the activation of the inflammatory cascade and may decrease the mRNA levels of TLR4/NF-κB signalling pathway components in LPS-treated Caco-2 cells.
Figure 4.
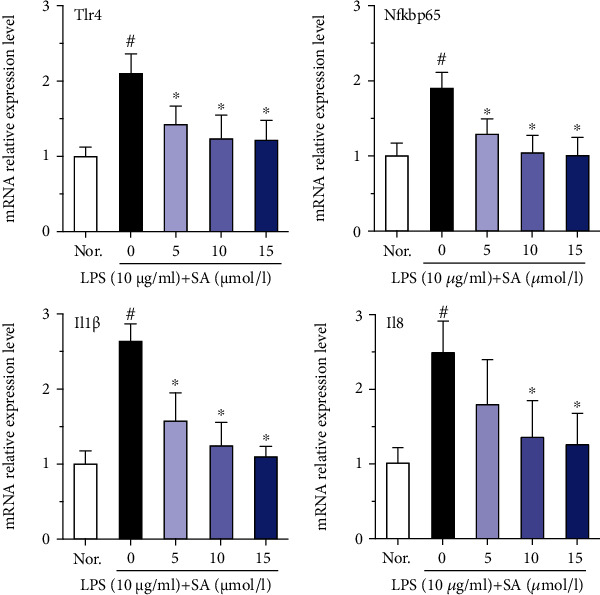
Effects of sinapic acid (SA) on the mRNA levels of Tlr4, Nfκbp65, Il1β, and Il8 in LPS-treated Caco-2 cells. Caco-2 cells were incubated with LPS (10 μg/ml) and SA (5, 10, or 15 μmol/l) for 24 h and then subjected to qRT-PCR analysis. The results are expressed as the mean ± SD of three independent experiments. # denotes p < 0.05 vs. the normal control group (Nor.), and ∗ denotes p < 0.05 vs. the LPS group.
3.5. Effects of SA on the Protein Levels of TLR4, MyD88, p-NF-κB, p-IKKα, and p-IκB in LPS-Treated Caco-2 Cells
The protein expression levels of TLR4, MyD88, p-NF-κBp65, p-IKKα, and p-IκB were analyzed using a protein blotting assay, and the effect of SA on the TLR4/NF-κB pathway was evaluated. As shown in Figure 5, the protein levels of MyD88, p-NF-κBp65, TLR4, p-IKKα, and p-IκB were increased following treatment with LPS (p < 0.05). However, administration of different concentrations of SA modulated the expression levels of these proteins (p < 0.05). These results clearly indicate that SA inhibits the TLR4/NF-κB signalling pathway in LPS-treated Caco-2 cells.
Figure 5.
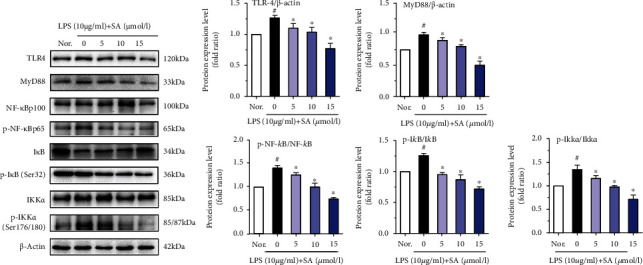
The effects of sinapic acid (SA) on the protein levels of NF-κB-related factors in LPS-treated Caco-2 cells. Caco-2 cells were incubated with LPS (10 μg/ml) and SA (5, 10, or 15 μmol/l) for 24 h and then subjected to western blot analysis. The results are expressed as the protein expression level (normalized to β-actin) relative to that in unstimulated cells and are shown as the mean ± SD of three independent experiments. # denotes p < 0.05 vs. the normal control group (Nor.), and ∗ denotes p < 0.05 vs. the LPS group.
3.6. Effects of SA on the Protein Levels of MLCK and MLC in LPS-Treated Caco-2 Cells
MLCK regulates the spatial conformation and function of the cytoskeleton and TJ of intestinal epithelial barrier cells. LPS treatment significantly induced the activation of MLCK and MLC (p < 0.05; Figure 6). In addition, treatment with different concentrations of SA significantly reduced the activation of MLCK and MLC in LPS-treated Caco-2 cells (p < 0.05).
Figure 6.
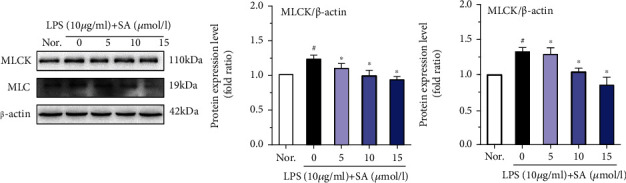
The effects of sinapic acid (SA) on the protein levels of MLCK and MLC in LPS-treated Caco-2 cells. Caco-2 cells were incubated with LPS (10 μg/ml) and SA (5, 10, or 15 μmol/l) for 24 h and then used for western blot analysis. The results are expressed as the protein expression level (normalized to β-actin) relative to that in unstimulated cells and are shown as the mean ± SD of three independent experiments. # denotes p < 0.05 vs. the normal control group (Nor.), and ∗ denotes p < 0.05 vs. the LPS group.
3.7. Effects of SA on the mRNA Levels of ZO-1, Claudin-1, and Occludin in LPS-Treated Caco-2 Cells
Normal levels of ZO-1, claudin-1, and occludin, the main members of the TJ protein family, act as a vital role in maintaining normal intestinal barrier function. qRT-PCR analysis showed that LPS treatment significant decreases in the mRNA levels of Zo1, claudin-1, and occludin in Caco-2 cells (Figure 7). However, administration of SA increased the mRNA levels of Zo1, claudin-1, and occludin in LPS-treated Caco-2 cells. At a concentration of 15 μmol/l, SA markedly increased the mRNA expression of these TJ factors (including Zo1, claudin-1, and occludin) (p < 0.05).
Figure 7.
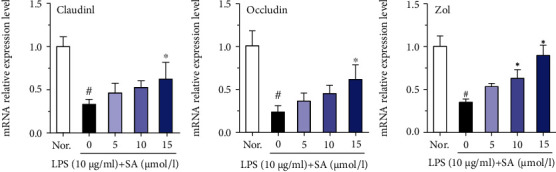
Effects of sinapic acid (SA) on tight junction mRNA levels in LPS-treated Caco-2 cells. Caco-2 cells were incubated with LPS (10 μg/ml) and SA (5, 10, or 15 μmol/l) for 24 h and then subjected to qRT-PCR analysis. The results are expressed as the mean ± SD of three independent experiments. # denotes p < 0.05 vs. the normal control group (Nor.), and ∗ denotes p < 0.05 vs. the LPS group.
3.8. Effects of SA on the Protein Levels of Occludin, Claudin-1, and ZO-1 in LPS-Treated Caco-2 Cells
LPS reduced the protein levels of ZO-1, claudin-1, and occludin in Caco-2 cells (p < 0.05; Figure 8). However, treatment with SA (15 μmol/l) markedly attenuated the downregulation of these TJ proteins (p < 0.05).
Figure 8.
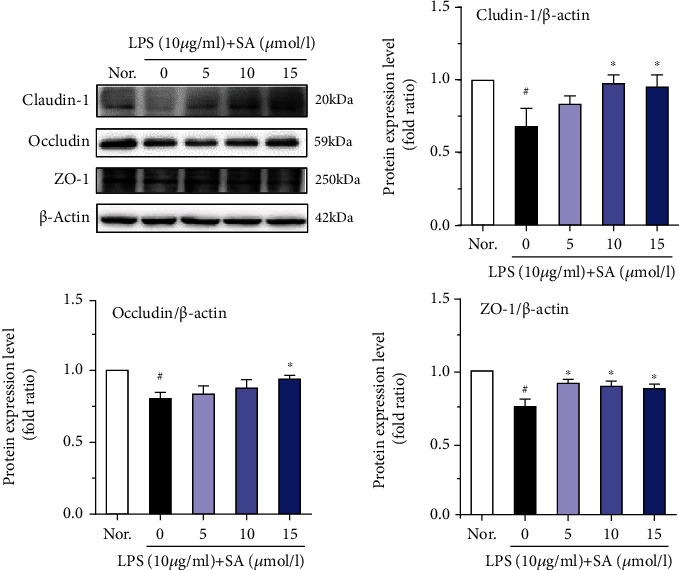
Effects of sinapic acid (SA) on tight junction protein levels in LPS-treated Caco-2 cells. Caco-2 cells were incubated with LPS (10 μg/ml) and SA (5, 10, or 15 μmol/l) for 24 h and then used for western blot analysis. The results are expressed as the protein expression level (normalized to β-actin) relative to that in unstimulated cells and are shown as the mean ± SD of three independent experiments. # denotes p < 0.05 vs. the normal control group (Nor.), and ∗ denotes p < 0.05 vs. the LPS group.
3.9. Effects of SA on Claudin-1 and ZO-1 Localization and Distribution in LPS-Treated Caco-2 Cells
As shown in Figure 9, claudin-1 and ZO-1 were appropriately localized to their respective intercellular junctions and were connected without damage in untreated Caco-2 cells. However, local claudin-1 and ZO-1 staining in the pericellular was discontinuous in the cells treated with LPS. These discontinuous pericellular expressions of both claudin-1 and ZO-1 were counteracted by SA treatment, and a strong fluorescence intensity was observed at the periphery of the cells.
Figure 9.
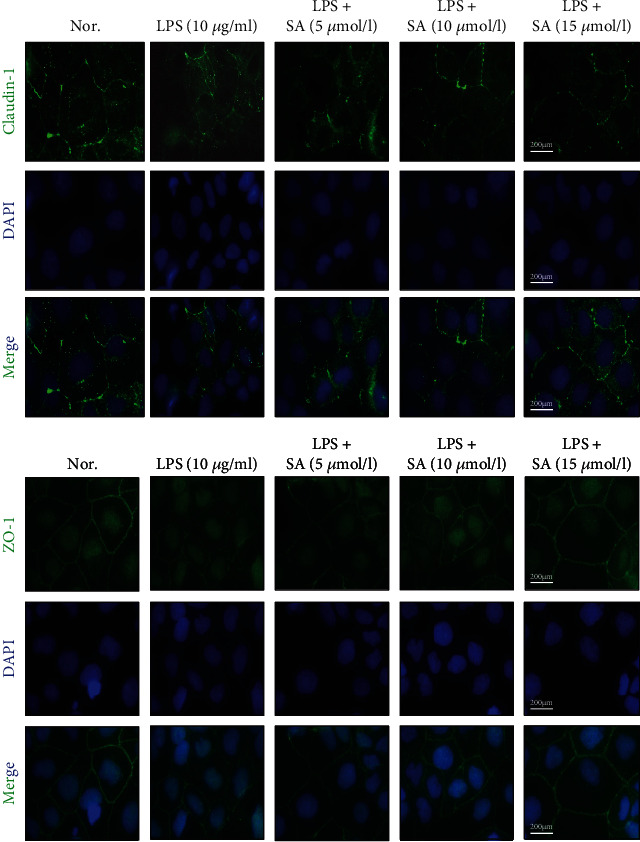
Effects of sinapic acid (SA) on claudin-1 and ZO-1 localization and distribution in LPS-treated Caco-2 cells. Caco-2 cells were incubated with LPS (10 μg/ml) and SA (5, 10, or 15 μmol/l) for 24 h and then subjected to immunofluorescence analysis (100x).
4. Discussion
Plant-derived phenolic compounds are a type of organic acid that contains either a benzoic or cinnamic acid skeleton with phenol as the basic framework and a relatively simple structure. Recent studies have suggested that SA has anti-inflammatory effects on intestinal inflammation and can regulate the intestinal microbiota and improve the redox state [25, 29, 30]. In the current investigation, we attempted to appraise the effects of SA on intestinal inflammation and permeability. Our results clearly confirmed that SA reduced epithelial permeability, increased the expression of the TJ proteins, and attenuated LPS-induced inflammation by modulating the TLR4/NF-κB pathway.
The intestinal epithelial barrier can effectively prevent pathogenic microorganisms, antigens, and toxic substances from entering the body from the gut [31, 32]. Epithelial barrier function is mediated by intercellular junctions [33]. TJ formation and assembly involve a complex of proteins. The TJ proteins (such as ZO-1, occludin, and claudin-1) are generally known as the main event during the change course of intestinal permeability [34]. Therefore, our investigation focused on the expression and distribution of ZO-1, occludin, and claudin-1.
Numerous studies have shown that LPS increases intestinal barrier permeability regulates the expression of TJ proteins and eventually lead to intestinal barrier dysfunction [35, 36]. On the other hand, the excessive accumulation of proinflammatory cytokines, including IL-1β and IL-8, is associated with promoting the generation of inflammation and ultimately causes the destruction of the intestinal epithelial barrier. LPS decreases TJ proteins in intestinal act as a centrical role in the cellular mechanisms of intestinal barrier defects [32]. In our study, LPS (10 μg/ml) was used to successfully establish the inflammation model. Here, our results suggested that the LPS-induced increases in the IL-1β and IL-8 expression and decreases in the protein levels of ZO-1, occludin, and claudin-1 were alleviated by SA.
TLR4, one of the best characterized pattern recognition receptors, is activated by LPS, leading to the activation of NF-κB and subsequently inducing the production of proinflammatory mediators. Recently, many studies have indicated that NF-κB is a key factor in inflammatory gene expression [37–39]. Under normal conditions, NF-κB binds with IκB to form an inactivation complex. NF-κB is released through IκB kinase (IKK), leading to IκB-α phosphorylation and degradation. Some stimulatory factors, such as proinflammatory cytokines, antigen receptors, growth factors, and LPS, may activate the IκB kinase (IKK) complex, which phosphorylates IκB. The phosphorylation of IκB causes its ubiquitination and proteasomal degradation, releasing NF-κB from the complex. In addition, NF-κB subunit p65 is transferred from the cytoplasm into the nucleus and initiates an inflammatory response [40]. A study reported that cinnamic acid downregulated the protein and mRNA expression of p-NF-κB and p-IKKα/β, which exerted anti-inflammatory effects in LPS-treated Caco-2 and RAW264.7 coculture systems [41]. Similar results were also found that ferulic acid treatment decreased the phosphorylation of IκB and NF-κBp65 in LPS-treated bovine endometrial epithelial cells [42]. Here, these results suggested that that SA treatment significantly diminished the activation of NF-κBp65; decreased the mRNA or protein expression of TLR4, MyD88, p-NF-κB, p-IKKα, and p-IκB; and inhibited LPS-induced IL-1β and IL-8 expression via modulation of the TLR4/NF-κB pathway. Moreover, the effects of SA treatment were concentration-dependent. These results suggest that SA has strong anti-inflammatory activity and can inhibit the LPS-induced activation of TLR4/NF-κB signalling pathway.
Activation of the MLCK-MLC pathway mainly regulates the spatial conformation and function of the cytoskeleton and plays a key role in regulating the tight junctions of intestinal cells. MLCK is a Ca2+/calmodulin-dependent kinase. Under physiological or pathological conditions, the tight junctions of cells can be regulated, and the cytoskeleton can be remodeled by catalyzing the phosphorylation of MLC, thus affecting the permeability of the intestinal epithelium [43]. It has been reported that overexpression of MLCK leads to increase intestinal epithelial TJ permeability [15, 44]. Many studies have reported that increased MLCK activity phosphorylates MLC and subsequently leads to the centripetal contraction of the TJ complex, eventually result in the opening of the intestinal epithelial barrier [45–47]. The results from our study clearly indicate that LPS promoted the expression of MLCK and MLC and diminished the expression of key sealing TJ proteins, such as ZO-1, occludin, and claudin-1, causing varying degrees of displacement of the claudin-1 and ZO-1 proteins. In contrast, SA inhibited the overactivation of MLCK and partially restored the expression and localization of related TJ proteins, reflecting that SA may alleviate injury to the intestinal epithelial barrier by inhibiting the MLCK/MLC pathway.
5. Conclusions
This study demonstrated that SA could ameliorate damage to the intestinal epithelial barrier and inhibit inflammation in LPS-stimulated intestinal epithelial cells. SA restored tight junction protein expression and protein localization via inhibition of the TLR4/NF-κB/MLCK-MLC pathway. Future research will reveal in more detail the potential of SA in the treatment or prevention of inflammation-induced intestinal barrier defects. In addition, SA also has the potential to be a possible alternative for the treatment of gastrointestinal diseases.
Acknowledgments
This research was funded by the National Natural Science Foundation of China (Nos. 81560530, 81760589, and 81960590 to J.-L. S.), the Guangxi Natural Science Foundation of China (Nos. 2016GXNSFBA380172 and 2020GXNSFAA159160 to J.-L. S.), the Funding Scheme for High-level Overseas Chinese Students' Return from the Ministry of Human Resources and Social Security of China (No. RSTH [2019]160 to J.-L. S.), the Foundation for University Key Teachers by the Department of Education of Guangxi Zhuang Autonomous Region (No. GRJ [2018]18 to J.-L. S.), and the Scientific Research Foundation for Returned Scholars of Guilin Medical University (04010150001 to J.-L. S.).
Data Availability
All generated and analyzed data used to support the findings of this study are included within the article.
Conflicts of Interest
The authors declare no conflict of interest.
Authors' Contributions
Huan Lan, Lu-Ying Zhang, and Wan-Ying Li performed the experiments. Zhen Zeng, Bo Qian, and Chengqiang Wang analyzed the results and drew the pictures. Jia-Le Song designed the experiments. Lu-Ying Zhang and Wen He wrote the manuscript. Jia-Le Song supervised the manuscript. Lu-Ying Zhang, Huan Lan, and Wen He contributed equally to this work.
References
- 1.Otani S., Coopersmith C. M. Gut integrity in critical illness. Journal of Intensive Care. 2019;7(1):p. 17. doi: 10.1186/s40560-019-0372-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mileo A. M., Nisticò P., Miccadei S. Polyphenols: immunomodulatory and therapeutic implication in colorectal cancer. Frontiers in Immunology. 2019;10:p. 729. doi: 10.3389/fimmu.2019.00729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Landy J., Ronde E., English N., et al. Tight junctions in inflammatory bowel diseases and inflammatory bowel disease associated colorectal cancer. World Journal of Gastroenterology. 2016;22(11):3117–3126. doi: 10.3748/wjg.v22.i11.3117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chang J., Leong R. W., Wasinger V. C., Ip M., Yang M., Phan T. G. Impaired intestinal permeability contributes to ongoing bowel symptoms in patients with inflammatory bowel disease and mucosal healing. Gastroenterology. 2017;153(3):723–731.e1. doi: 10.1053/j.gastro.2017.05.056. [DOI] [PubMed] [Google Scholar]
- 5.Rowart P., Wu J., Caplan M. J., Jouret F. Implications of AMPK in the Formation of Epithelial Tight Junctions. International Journal of Molecular Sciences. 2018;19(7):p. 2040. doi: 10.3390/ijms19072040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Deng J., Zeng L., Lai X., et al. Metformin protects against intestinal barrier dysfunctionviaAMPKα1-dependent inhibition of JNK signalling activation. Journal of Cellular and Molecular Medicine. 2018;22(1):546–557. doi: 10.1111/jcmm.13342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cui W., Li L. X., Sun C. M., et al. Tumor necrosis factor alpha increases epithelial barrier permeability by disrupting tight junctions in Caco-2 cells. Brazilian Journal of Medical and Biological Research. 2010;43(4):330–337. doi: 10.1590/S0100-879X2010007500020. [DOI] [PubMed] [Google Scholar]
- 8.Mennigen R., Nolte K., Rijcken E., et al. Probiotic mixture VSL#3 protects the epithelial barrier by maintaining tight junction protein expression and preventing apoptosis in a murine model of colitis. American Journal of Physiology: Gastrointestinal and Liver Physiology. 2009;296(5):G1140–G1149. doi: 10.1152/ajpgi.90534.2008. [DOI] [PubMed] [Google Scholar]
- 9.Zhang R., Xu J., Zhao J., Chen Y. Genistein improves inflammatory response and colonic function through NF-κB signal in DSS-induced colonic injury. Oncotarget. 2017;8(37):61385–61392. doi: 10.18632/oncotarget.18219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cocetta V., Governa P., Borgonetti V., et al. Cannabidiol isolated from Cannabis sativa L. protects intestinal barrier from in vitro inflammation and oxidative stress. Frontiers in Pharmacology. 2021;12:p. 641210. doi: 10.3389/fphar.2021.641210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang H., Wang Y., Su Y., Fang X., Guo W. The Alleviating Effect and Mechanism of Bilobalide on Ulcerative Colitis. Food & Function. 2021;12(14):6226–6239. doi: 10.1039/D1FO01266E. [DOI] [PubMed] [Google Scholar]
- 12.Cao S. G., Chen R., Wang H., Lin L. M., Xia X. P. Cryptotanshinone inhibits prostaglandin E2 production and COX-2 expression via suppression of TLR4/NF-κB signaling pathway in LPS-stimulated Caco-2 cells. Microbial Pathogenesis. 2018;116:313–317. doi: 10.1016/j.micpath.2017.12.027. [DOI] [PubMed] [Google Scholar]
- 13.Chelakkot C., Ghim J., Ryu S. H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Experimental and Molecular Medicine. 2018;50(8):p. 103. doi: 10.1038/s12276-018-0126-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Stephens M., von der Weid P. Y. Lipopolysaccharides modulate intestinal epithelial permeability and inflammation in a species-specific manner. Gut Microbes. 2020;11(3):421–432. doi: 10.1080/19490976.2019.1629235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nighot M., al-Sadi R., Guo S., et al. Lipopolysaccharide-induced increase in intestinal epithelial tight permeability is mediated by toll-like receptor 4/myeloid differentiation primary response 88 (MyD88) activation of myosin light chain kinase expression. American Journal of Pathology. 2017;187(12):2698–2710. doi: 10.1016/j.ajpath.2017.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hou Q., Zhu S., Zhang C., et al. Berberine improves intestinal epithelial tight junctions by upregulating A20 expression in IBS-D mice. Biomedicine and Pharmacotherapy. 2019;118, article 109206 doi: 10.1016/j.biopha.2019.109206. [DOI] [PubMed] [Google Scholar]
- 17.Attri S., Goel G. Influence of polyphenol rich seabuckthorn berries juice on release of polyphenols and colonic microbiota on exposure to simulated human digestion model. Food Research International. 2018;111:314–323. doi: 10.1016/j.foodres.2018.05.045. [DOI] [PubMed] [Google Scholar]
- 18.Roberts K. M., Grainger E. M., Thomas-Ahner J. M., et al. Application of a low polyphenol or low ellagitannin dietary intervention and its impact on ellagitannin metabolism in men. Molecular Nutrition & Food Research. 2017;61(3, article 1600224) doi: 10.1002/mnfr.201600224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vetrani C., Vitale M., Bozzetto L., et al. Association between different dietary polyphenol subclasses and the improvement in cardiometabolic risk factors: evidence from a randomized controlled clinical trial. Acta Diabetologica. 2018;55(2):149–153. doi: 10.1007/s00592-017-1075-x. [DOI] [PubMed] [Google Scholar]
- 20.Ganesan K., Xu B. Polyphenol-rich dry common beans (Phaseolus vulgaris L.) and their health benefits. International Journal of Molecular Sciences. 2017;18(11):2331–2357. doi: 10.3390/ijms18112331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Awika J. M., Duodu K. G. Bioactive polyphenols and peptides in cowpea (Vigna unguiculata) and their health promoting properties: a review. Journal of Functional Foods. 2017;38:686–697. doi: 10.1016/j.jff.2016.12.002. [DOI] [Google Scholar]
- 22.Bin Jardan Y. A., Ansari M. A., Raish M., et al. Sinapic Acid Ameliorates Oxidative Stress, Inflammation, and Apoptosis in Acute Doxorubicin-Induced Cardiotoxicity via the NF-κB-Mediated Pathway. BioMed Research International. 2020;2020:10. doi: 10.1155/2020/3921796.3921796 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zare K., Eidi A., Roghani M., Rohani A. H. The neuroprotective potential of sinapic acid in the 6-hydroxydopamine-induced hemi-parkinsonian rat. Metabolic Brain Disease. 2015;30(1):205–213. doi: 10.1007/s11011-014-9604-6. [DOI] [PubMed] [Google Scholar]
- 24.Roy S. J., Mainzen Prince P. S. Protective effects of sinapic acid on cardiac hypertrophy, dyslipidaemia and altered electrocardiogram in isoproterenol-induced myocardial infarcted rats. European Journal of Pharmacology. 2013;699(1-3):213–218. doi: 10.1016/j.ejphar.2012.11.012. [DOI] [PubMed] [Google Scholar]
- 25.Lee J. Y. Anti-inflammatory effects of sinapic acid on 2,4,6-trinitrobenzenesulfonic acid-induced colitis in mice. Archives of Pharmacal Research. 2018;41(2):243–250. doi: 10.1007/s12272-018-1006-6. [DOI] [PubMed] [Google Scholar]
- 26.Kanchana G., Shyni W. J., Rajadurai M., Periasamy R. Evaluation of antihyperglycemic effect of sinapic acid in normal and streptozotocin-induced diabetes in albino rats. Global Journal of Pharmacology. 2011;5(1) [Google Scholar]
- 27.Fresco P., Borges F., Diniz C., Marques M. P. New insights on the anticancer properties of dietary polyphenols. Medicinal Research Reviews. 2006;26(6):747–766. doi: 10.1002/med.20060. [DOI] [PubMed] [Google Scholar]
- 28.Turner J. R., Buschmann M. M., Romero-Calvo I., Sailer A., Shen L. The role of molecular remodeling in differential regulation of tight junction permeability. Seminars in Cell & Developmental Biology. 2014;36:204–212. doi: 10.1016/j.semcdb.2014.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Yang C., Deng Q., Xu J., et al. Sinapic acid and resveratrol alleviate oxidative stress with modulation of gut microbiota in high-fat diet-fed rats. Food Research International. 2019;116:1202–1211. doi: 10.1016/j.foodres.2018.10.003. [DOI] [PubMed] [Google Scholar]
- 30.Qian B., Wang C., Zeng Z., Ren Y., Li D., Song J. L. Ameliorative effect of sinapic acid on dextran sodium sulfate- (DSS-) induced ulcerative colitis in Kunming (KM) mice. Oxidative Medicine and Cellular Longevity. 2020;2020:13. doi: 10.1155/2020/8393504.8393504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang B., Wu Z., Ji Y., Sun K., Dai Z., Wu G. L-Glutamine enhances tight junction integrity by activating CaMK kinase 2-AMP-activated protein kinase signaling in intestinal porcine epithelial cells. Journal of Nutrition. 2016;146(3):501–508. doi: 10.3945/jn.115.224857. [DOI] [PubMed] [Google Scholar]
- 32.Chen M., Liu Y., Xiong S., et al. Dietary l-tryptophan alleviated LPS-induced intestinal barrier injury by regulating tight junctions in a Caco-2 cell monolayer model. Food & Function. 2019;10(5):2390–2398. doi: 10.1039/C9FO00123A. [DOI] [PubMed] [Google Scholar]
- 33.Zhang J., Lu Y., Li S., et al. Co-infection with porcine bocavirus and porcine circovirus 2 affects inflammatory cytokine production and tight junctions of IPEC-J2 cells. Virus Genes. 2018;54(5):684–693. doi: 10.1007/s11262-018-1596-6. [DOI] [PubMed] [Google Scholar]
- 34.Oshima T., Miwa H. Gastrointestinal mucosal barrier function and diseases. Journal of Gastroenterology. 2016;51(8):768–778. doi: 10.1007/s00535-016-1207-z. [DOI] [PubMed] [Google Scholar]
- 35.He C., Deng J., Hu X., et al. Vitamin A inhibits the action of LPS on the intestinal epithelial barrier function and tight junction proteins. Food & Function. 2019;10(2):1235–1242. doi: 10.1039/C8FO01123K. [DOI] [PubMed] [Google Scholar]
- 36.Wang J., Ghosh S. S., Ghosh S. Curcumin improves intestinal barrier function: modulation of intracellular signaling, and organization of tight junctions. American Journal of Physiology: Cell Physiology. 2017;312(4):C438–c445. doi: 10.1152/ajpcell.00235.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yue B., Ren J., Yu Z., et al. Pinocembrin alleviates ulcerative colitis in mice via regulating gut microbiota, suppressing TLR4/MD2/NF-κB pathway and promoting intestinal barrier. Bioscience Report. 2020;40(7) doi: 10.1042/BSR20200986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rashidian A., Rashki A., Abdollahi A., et al. Dapsone reduced acetic acid-induced inflammatory response in rat colon tissue through inhibition of NF-kB signaling pathway. Immunopharmacology and Immunotoxicology. 2019;41(6):607–613. doi: 10.1080/08923973.2019.1678635. [DOI] [PubMed] [Google Scholar]
- 39.Serra G., Incani A., Serreli G., et al. Olive oil polyphenols reduce oxysterols -induced redox imbalance and pro- inflammatory response in intestinal cells. Redox Biology. 2018;17:348–354. doi: 10.1016/j.redox.2018.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Vermeulen L., De Wilde G., Notebaert S., Vanden Berghe W., Haegeman G. Regulation of the transcriptional activity of the nuclear factor-κB p65 subunit. Biochemical Pharmacology. 2002;64(5-6):963–970. doi: 10.1016/S0006-2952(02)01161-9. [DOI] [PubMed] [Google Scholar]
- 41.Kim M. S., Kim J. Y. Cinnamon subcritical water extract attenuates intestinal inflammation and enhances intestinal tight junction in a Caco-2 and RAW264.7 co-culture model. Food & Function. 2019;10(7):4350–4360. doi: 10.1039/C9FO00302A. [DOI] [PubMed] [Google Scholar]
- 42.Yin P., Zhang Z., Li J., et al. Ferulic acid inhibits bovine endometrial epithelial cells against LPS-induced inflammation via suppressing NK-κB and MAPK pathway. Research in Veterinary Science. 2019;126:164–169. doi: 10.1016/j.rvsc.2019.08.018. [DOI] [PubMed] [Google Scholar]
- 43.Yang X., Zou D., Tang S., et al. Ameliorative effect of melatonin against increased intestinal permeability in diabetic rats: possible involvement of MLCK-dependent MLC phosphorylation. Molecular and Cellular Biochemistry. 2016;416(1-2):23–32. doi: 10.1007/s11010-016-2691-4. [DOI] [PubMed] [Google Scholar]
- 44.Clayburgh D. R., Rosen S., Witkowski E. D., et al. A Differentiation-dependent Splice Variant of Myosin Light Chain Kinase, MLCK1, Regulates Epithelial Tight Junction Permeability. Journal of Biological Chemistry. 2004;279(53):55506–55513. doi: 10.1074/jbc.M408822200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Weber C. R., Raleigh D. R., Su L., et al. Epithelial Myosin Light Chain Kinase Activation Induces Mucosal Interleukin-13 Expression to Alter Tight Junction Ion Selectivity. Journal of Biological Chemistry. 2010;285(16):12037–12046. doi: 10.1074/jbc.M109.064808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zhai Z., Wang J., Huang B., Yin S. Low-fat yogurt alleviates the pro-inflammatory cytokine IL-1β-induced intestinal epithelial barrier dysfunction. Journal of Dairy Science. 2019;102(2):976–984. doi: 10.3168/jds.2018-15226. [DOI] [PubMed] [Google Scholar]
- 47.Chen S. W., Zhu J., Zuo S., et al. Protective effect of hydrogen sulfide on TNF-α and IFN-γ-induced injury of intestinal epithelial barrier function in Caco-2 monolayers. Inflammation Research. 2015;64(10):789–797. doi: 10.1007/s00011-015-0862-5. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All generated and analyzed data used to support the findings of this study are included within the article.