

Abstract
Zinc-finger proteins are important transcription factors in plants, responding to adversity and regulating the growth and development of plants. However, the roles of the BBX gene family of zinc-finger proteins in wintersweet (Chimonanthus praecox) have yet to be elucidated. In this study, a group IV subfamily BBX gene, CpBBX19, was identified and isolated from wintersweet. Quantitative real-time PCR (qRT-PCR) analyses revealed that CpBBX19 was expressed in all tissues and that expression was highest in cotyledons and inner petals. CpBBX19 was also expressed in all flower development stages, with the highest expression detected in early initiating bloom, followed by late initiating bloom and bloom. In addition, the expression of CpBBX19 was induced by different abiotic stress (cold, heat, NaCl, and drought) and hormone (ABA and MeJA) treatments. Heterologous expression of CpBBX19 in Arabidopsis thaliana (Arabidopsis) enhanced the tolerance of this plant to salt and drought stress as electrolyte leakage and malondialdehyde (MDA) concentrations in transgenic Arabidopsis after stress treatments were significantly lower than those in wild-type (WT) plants. In conclusion, this research demonstrated that CpBBX19 plays a role in the abiotic stress tolerance of wintersweet. These findings lay a foundation for future studies on the BBX gene family of wintersweet and enrich understanding of the molecular mechanism of stress resistance in wintersweet.
Keywords: CpBBX19, Chimonanthus praecox, salt stress, drought, overexpression, Arabidopsis
1. Introduction
Wintersweet (Chimonanthus praecox), belonging to the Calycanthaceae family, is native to China, has a distinct flowering period (from November to March), strong aroma, and yellow flowers [1,2], and is widely used for cut flowers and as a garden plant in China [3]. In addition, wintersweet is used for aromatic oil extraction, Chinese herbal medicine, and cosmetics [1,2,4]. The unusual flowering period of wintersweet shows that this species has a strong ability to tolerate low temperatures [5]. Some genes and transcription factors have been identified as related to biotic and abiotic stress in wintersweet [6]. For instance, heterologous expression of CpSIZ1 in Arabidopsis enhanced the cold resistance of transgenic Arabidopsis (Arabidopsis thaliana) [5], while overexpression of the gene CpLEA5 increased the ability of transgenic Arabidopsis to resist low temperature and tolerate salt and alkali [7]. In addition, the amino acid sequence of CpFATB has high sequence similarity with the choroidal palmitoyl acyl carrier protein thioesterase of other plants, and overexpression of CpFATB in tobacco enhanced the drought tolerance of transgenic tobacco (Nicotiana tobaccum) [8]. Furthermore, CpCOR413 is a membrane-stable protein related to the cold tolerance of wintersweet [9], while overexpression of the gene CpTAF10 increased the germination rate of transgenic Arabidopsis seeds, and under salt stress, the taproot and lateral roots of transgenic plants showed certain growth advantages compared with WT plants [10].
In plants, transcription factors, as key elements of the expression network, participate in the regulation of expression of numerous genes and are instrumental in regulating plant growth and development and the response to the external environment, including biotic and abiotic stresses [10,11,12]. The BBX gene family, encoding zinc-finger proteins, is an important family of transcription factors in plants, with functions including response to environmental changes, and plant growth and development [13,14]. BBX proteins generally have one or two B-box domains, and some members also have VP domains and CCT domains [15]. In plants, the B-box domain contains approximately 40 amino acids and is highly conserved, but there are still some differences among different B-box motifs [16,17]. The B-box domain binds to Zn ions to stabilize its unique tertiary structure [18], which is involved in protein–protein interactions [13]. For example, BBX1 (CONSTANS), BBX4 (COL3), BBX21 (STH2), BBX22 (STH3), BBX24 (STO), and BBX25 (STH) all interact with COP1 (CONSTITUTIVE PHOTOMORPHOGENESIS 1) [16,19,20,21,22]. The CCT domain of BBX proteins comprises 42 or 43 conserved amino acids and mainly mediates transcription regulation and protein transport, but also has a function in positioning. The VP domain is adjacent to the CCT domain and mainly mediates the interaction with COP1 [17,18,23]. In Arabidopsis, the BBX protein family is divided into five structural types according to the number of B-box domains and CCT domains: the first type (BBX1-6) has two tandem B-box domains, one VP domain, and one CCT domain; the second type (BBX7-13) has two B-box domains and one CCT domain, but lacks a VP domain; the third type (BBX14-17) contains one B-box domain and one CCT domain; the fourth type (BBX18-25) only contains two tandem B-box domains, and the fifth type (BBX26-32) has only one B-box domain [17,18,23].
Under abiotic stresses, such as drought, high and low temperatures, and high salt, multiple BBX gene family members were strongly induced [17,24,25]. In Arabidopsis, the thermostability of AtBBX18-RNAi plants was improved, while the thermostability of AtBBX18 overexpression plants was decreased. In AtBBX18 overexpression plants, heat stress response genes DGD1, Hsfa2, and Hsp101 were downregulated, suggesting that the gene BBX18 plays a negative regulatory role in the heat-stress response of Arabidopsis [26]. Under drought stress, multiple MdBBX genes were upregulated in roots and leaves of apple trees (Malus domestica) [27]. Overexpression of MdBBX10 enhanced the resistance of transgenic Arabidopsis to drought, salt, and other abiotic stresses, which was closely related to the enhancement of active oxygen-scavenging ability [28]. High salt and Polyethylene glycol (PEG) treatment induced SsBBX24 gene expression and protein accumulation in Solanum sogarandinum [29]. Exogenous hormones treatment, such as abscisic acid (ABA), gibberellin (GA), ethylene, cytokinin, and auxin, also affected transcriptional levels of BBX genes [30,31,32]. The expression level of CmBBX24 was influenced by GA4/7, and suppressing the expression of CmBBX24 reduced cold and drought tolerance of chrysanthemum [33]. Under treatment with GA, Salicylic acid (SA), and Methyl Jasmonate (MeJA) hormones, expression levels of the genes OsBBX2, OsBBX7, OsBBX17, OsBBX19, and OsBBX24 were upregulated at different time points [31]. Treatment of Arabidopsis leaves with ABA resulted in significant upregulation of BBX11, BBX13, and BBX22 gene expression levels, while BBX2, BBX3, BBX16, BBX18, and BBX19 genes were downregulated by cyclic adenosine diphosphate ribose (cADPR), which is involved in ABA early signal transduction [34].
Under adversity stress, the permeability of plant cell membranes changed. The degree of plant cell membrane damage could be judged by measuring the extravasation of cell electrolytes [35]. Malondialdehyde (MDA) is an important product of membrane lipid peroxidation and is usually used as an indicator of membrane lipid peroxidation. Therefore, the changes in electrolyte leakage and MDA content reflect the degree of damage to the membrane system under adversity conditions, and were widely used in the research of plant cold resistance, drought resistance, and salt–alkali tolerance [36,37].
The BBX protein family was widely reported to be involved in the process of abiotic stress in plants [10,11,12]. The functions of BBX zinc-finger proteins have been clarified in many plants, such as Arabidopsis [26], chrysanthemum [38], and apple [28]. However, the BBX gene family has not been extensively studied in woody plants such as wintersweet. According to information from the wintersweet transcriptome database [39], expression of a new BBX gene, CpBBX19, might be different under abiotic stress. In this study, we identified and isolated a BBX gene, CpBBX19, in wintersweet, and explored the expression patterns and function of this gene. CpBBX19 was classified into the IV subfamily of BBX proteins, and significantly improved the tolerance of transgenic Arabidopsis to salt and drought.
2. Materials and Methods
2.1. Plant Materials
To analyze the expression pattern of the gene CpBBX19 in wintersweet, the roots, stems, cotyledons, and young and old leaves were collected from six-leaf wintersweet, and flowers during the full bloom period were collected from five-year-old plants. Using the wintersweet floral development stages classification method [40], flowers of germination, flower-bud, petal-display, early initiating bloom, later initiating bloom, blooming, and withering were collected, respectively. The tissues were rapidly frozen in liquid nitrogen after collection. Three biological repeats were obtained for each sample.
Wild-type Arabidopsis (ecotype Columbia−0) was used for plant transformation. After sterilizing in hypochlorite solution, Arabidopsis seeds were sown on solid Murashige and Skoog (MS) medium, vernalized at 4 °C for 3 days, and then grown in an environment of 22 ± 1 °C with a 16 h light/8 h dark photoperiod. After 2 weeks, the seedlings were transferred into a matrix of vermiculite and peat (1:1 ratio) and cultivated from MS medium under the same conditions as described above.
2.2. Gene Isolation
Total RNA was extracted from the wintersweet tissues according to the manufacturer’s instructions of the RNAprep pure plant total RNA extraction kit (Tiangen, Beijing, China). To ensure RNA quality and quantity, RNA extracts were visualized by 1% agarose gel electrophoresis, then a Nano-Drop ND-1000 spectrophotometer (Thermo Fisher Scientific, Wilmington, MA, USA) was used for quantification at optical densities of 260 and 280 nm. The cDNA samples were synthesized using a PrimeScript RT reagent kit with gDNA Eraser (TaKaRa, Otsu, Japan). Each 10 µL contained 2 µL 5 × gDNA Eraser Buffer, 1 µL gDNA Eraser, and 1μg total RNA. Then, PCR procedure was performed PCR at 42 °C for 2 min; first-strand cDNA was synthesized by reverse transcriptase in 20 μL reaction system containing 10 µL reaction product from the previous step, 1 µL PrimeScript RT Enzyme Mix I, 4 µL 5 × PrimeScript Buffer 2, and 4 µL RNase Free dH2O [5]. The process of PCR was as follows: 37 °C for 15 min, then 85 °C for 5 s, and end at 4 °C. Specific primers CpBBX19-F/R (Table S1) were designed by Primer Premier 6.0 to clone the DNA sequence of CpBBX19. The PCR procedure comprised an initial preheating step at 95 °C for 5 min, followed by 27 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 45 s, with a final extension at 72 °C for 10 min. PCR amplification products were separated via electrophoresis on a 1% agarose gel, and the target DNA fragments were recovered using an Agarose Gel DNA extraction kit (Tiangen) according to the manufacturer’s instructions. The resulting fragments were cloned into the pMD19-T vector (TakaRa, Dalian, China) and sequenced by TsingKe Company (TsingKe, Chengdu, China).
BBX19 upstream promoter sequence was cloned using genomic walking method via Universal Genome WalkerTM 2.0 User Manual kit (Clontech, Mountain View, CA, USA) by manufacturer’s description. Specific primers (SP1 and SP2) (Table S1) were designed according to the ORF sequence of CpBBX19. The genomic DNA walking library enzyme digestion template was established by the following steps: first, digest with EcoRV blunt-end enzyme, then passivate the digested product, and then ligate with T4-DNA ligase, and connect the upstream and downstream genome walking adapters in a 16 °C water bath overnight. The PCR reaction system contained 2.5 μL 10 × buffer, 0.5 μL dNTPs, 0.5 μL primer APF1/2, 0.5 μL primer SP1/2, 0.5 μL Taq DNA polymerase, 1 μL DNA, and 19.5 μL ddH2O. The reaction procedure was carried out according to the Manufacturer’s instructions. RNA was detected by 1% gel electrophoresis, the target band was recovered, and it was sequenced to determine the upstream sequence of CpBBX19.
2.3. Bioinformatics Analysis
The sequence of CpBBX19 was determined using the BLAST-Protein program in the National Center for Biotechnology Information (NCBI) database, available online https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 25 December 2019). Multiple sequence alignments were performed by BioEdit software. The online analysis website MEME (The MEME Suite 5.3.3) was used for protein motif analysis; motif setting value was six; available online https://meme-suite.org/meme/info/status?service=MEME&id=appMEME_5.4.11632023872135256190210 (accessed on 8 July 2021). To understand the distribution of Cis-acting elements upstream of the CpBBX19 promoter sequence, the 2000-bp sequence upstream of CpBBX19 [41] was analyzed by PLANTCARE, available online http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 19 March 2021).
2.4. Plant Treatments
Expression profiles of the gene CpBBX19 were determined under different abiotic and hormonal treatments using the six-leaf plant as the materials [42]. Drought and NaCl treatments involved the use of 50% PEG-6000 [43] and 300 mM NaCl [31] irrigation, respectively. Cold and heat stress comprised treatments at 4 °C and 42 °C [44], respectively. For exogenous hormone treatment, 50 µM ABA and 100 µM MeJA were sprayed [42], respectively. There were three biological replicates per treatment. Samples were collected at 0, 2, 6, 12, and 24 h after treatment and quickly frozen in liquid nitrogen.
2.5. Quantitative Real-Time PCR (qRT-PCR)
CpBBX19 expression patterns were analyzed in different tissues and flowers development stages of wintersweet. RNA was extracted from the wintersweet samples according to the method described in 2.2. The qRT-PCR analyses were performed by Bio-RAD CFX96 (Bio-RAD CFX Manager Software Version 1.6). Specific primers (RT-CpBBX19-F/R) were used for PCR amplification, and the process of qRT-PCR was performed according to the method described by Liu et al. [45]. According to the manufacturer’s instructions, Ssofast EvaGreen Supermix (50 × 20 μL reactions) includes 2× reaction buffer with dNTPs, Sso7d-fusion polymerase, MgCl2, EvaGreen dye, and stabilizers. Each 10 μL reaction mixture contained 5 μL Ssofast EvaGreen Supermix (Bio-RAD, Hercules, CA, USA), 0.5 μL each gene-specific primer, and 3.5 μL nuclease-free water. The PCR program comprised preheating at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 5 s, annealing at 60 °C for 5 s, and extension at 72 °C for 5 s. There were three biological replicates for each experiment and three technical replicates for each sample. CpActin genes were employed as the internal reference gene of wintersweet [42]. Specific primers required for amplification were designed by Primer Premier 6.0 software. The qRT-PCR product of CpBBX19 was 150 bp, and the primers (RT-CpBBX19-F/R) are shown in Table S1.
2.6. Construction of Expression Vectors
CpBBX19 was fused in-frame into the pGWB551 vector with the CaMV35S promoter by Gateway recombination reactions [43] to generate the 35S::CpBBX19 construct. Specific primers (pGWB551-CpBBX19-F/R) used in the process are shown in Table S1, and the plasmid map of the 35S::pGWB551 vector is shown in Figure S1. The resulting 35S::CpBBX19 plasmid was confirmed by sequencing and then transformed into Agrobacterium tumefaciens (strain GV3101 competent cells) via electroporation to further explore the function of CpBBX19.
2.7. Generation of Transgenic Arabidopsis and Stress Treatments
Arabidopsis transgenic lines (OE lines) were obtained by transferring the 35S::CpBBX19 recombinant plasmid into wild-type Arabidopsis via the floral dip method [46]. T-0 seeds were sown on MS medium containing 25 µg/mL hygromycin for transgenic selection. The method of Khan et al. [47] was used to extract DNA from the leaves of transgenic and nontransgenic plants (WT), and PCR amplification was performed using the extracted DNA and CpBBX19 specific primers (CpBBX19-F1/R1) (Table S1). The PCR procedure comprised an initial preheating step at 95 °C for 5 min, followed by 27 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 40 s, with a final extension at 72 °C for 10 min. PCR products were detected by 1% gel electrophoresis to confirm the insertion of CpBBX19 into transgenic plants. qRT-PCR was used to identify the expression level of CpBBX19 in the transgenic lines, with the AtActin gene [43] used as the internal reference gene of Arabidopsis. The qRT-PCR product of AtActin was 200 bp. Three T3 transgenic lines (with high, medium, and low expression of CpBBX19, respectively) were selected for phenotypic investigation; wild-type Arabidopsis (WT) was used as the control.
To determine the salt tolerance of the transgenic lines, WT and transgenic Arabidopsis were irrigated every 2 days with 300 mM NaCl to ensure that the concentration of NaCl solution in the tray remained uniform. To test the drought tolerance of the transgenic lines, WT and transgenic Arabidopsis were irrigated with 30% PEG-6000. MDA content and electrolyte leakage of transgenic Arabidopsis were detected as indicators of stress tolerance. MDA content was determined as described by Heath et al. [48], while the method of electrolyte leakage was stated by Seliem et al. [49].
2.8. Data Analysis
IBM SPSS Statistics 23 software package was used for statistical analysis, and Duncan’s multiple range tests were employed to analyze the significance of differences. A p-value < 0.05 was recognized as statistically significant, and a p-value < 0.01 was classed as extremely significant.
3. Results
3.1. Isolation and Characterization of CpBBX19
Based on the wintersweet transcriptome sequence in the database [39], a 1536-bp full-length cDNA of CpBBX19 (accession number MZ740468) was obtained. The 5′ untranslated region (5′-UTR) and a 3′ untranslated region (3′-UTR) of CpBBX19 were 475 bp and 428 bp, respectively, and the open reading frame (ORF) of CpBBX19 was 633 bp, encoding 210 amino acids. The predicted molecular weight (MW) of the resulting protein was 23.47 kDa, and the deduced isoelectric point (PI) and instability index were 5.91 and 56.50, respectively. Subcellular localization prediction analysis of the CpBBX19 gene-encoded protein of wintersweet showed that the protein was predominantly distributed in the nucleus.
Multiple sequence alignment revealed that the CpBBX19 protein sequence was highly homologous to the BBX19 protein sequence of Arabidopsis, Gossypium hirsutum, Hibiscus syriacus, Ziziphus jujuba, and Hevea brasiliensis (Figure 1a), indicating that BBX19 was conserved in plants. The CpBBX19 protein contained two B-box domains, but lacked the CCT domain and VP domain at the C-terminus. A phylogenetic tree of CpBBX19 and Arabidopsis BBX proteins was reconstructed by MEGE 6 (Figure 1b) and showed the five subfamilies of the BBX protein family; CpBBX19 belonged to the IV subfamily and was closely related to AtBBX19. In addition, the promoter region 2000 bp upstream of the CpBBX19 gene was analyzed (Figure 1c and Table S2). The CpBBX19 promoter region contained one defense and stress responsive element (TC-rich), one element involved in salicylic acid responsiveness (TAC-element), one MYB binding site (MBS) involved in drought-inducibility, three elements related to abscisic acid responsiveness (ABRE), and four elements involved in MeJA responsiveness (TGACG-motif and CGTCA-motif), indicating that CpBBX19 might be related to the stress response in plants.
Figure 1.
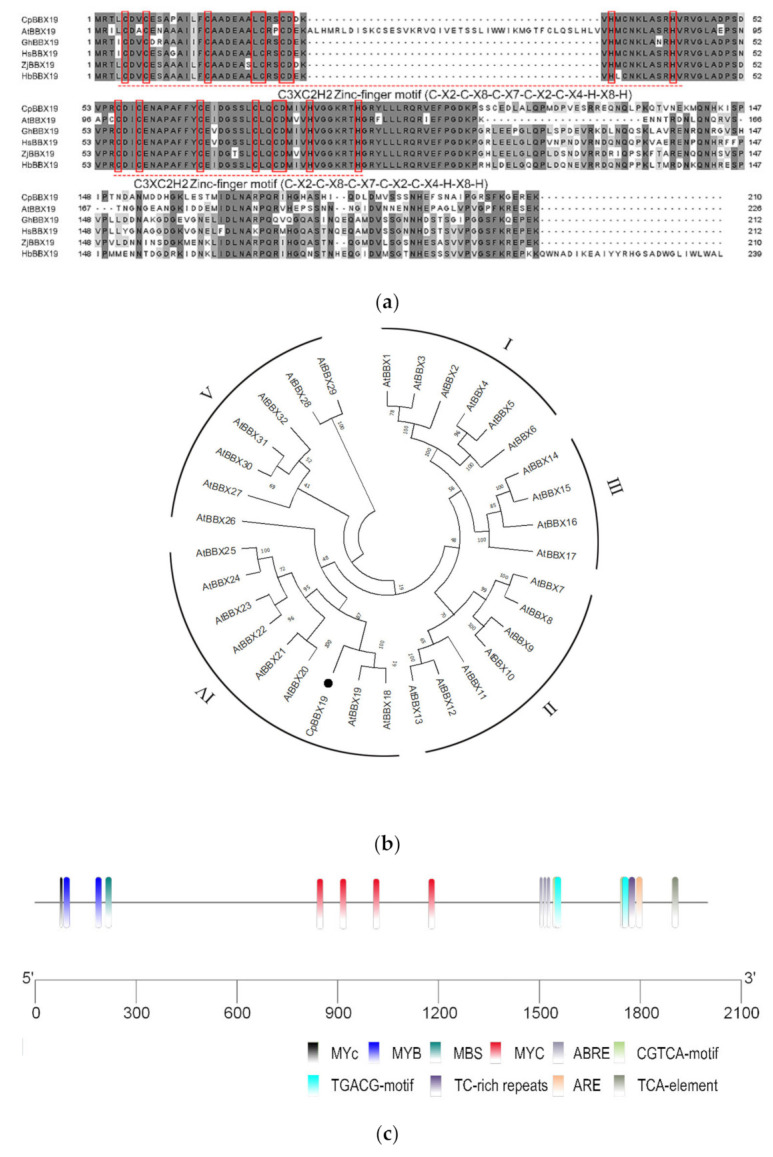
Characterization of CpBBX19. (a) Multiple sequence alignment of CpBBX19 with BBX19 proteins from Arabidopsis thaliana (accession number NP_001190958.1), Gossypium hirsutum (XP_016697629.1), Hibiscus syriacus (XP_039058672.1), Ziziphus jujube (XP_015892301.1), and Hevea brasiliensis (XP_021655083.1). Identical and similar amino acids are shaded in gray and light gray, respectively, and conserved zinc-finger motifs are marked by red boxes. (b) Phylogenetic analysis of CpBBX19 and BBX proteins from Arabidopsis. MEGA 6 software with the neighbor-joining (NJ) method (1000 bootstrap repeats) was used to reconstruct the phylogenetic tree. Accession numbers of the protein sequences used for phylogenetic analysis are shown in Table S3. (c) Cis-acting elements in the promoter of CpBBX19.
As shown in Figure 2, the six conserved motifs in CpBBX19 and the IV subfamily of Arabidopsis, identified by MEME software, comprised between 8 and 47 amino acids. Sequence analysis revealed that motif 1 contained the complete structure C-X2-C-X8-C-X7-C-X2-C-X4-H-X8-H of B-Box1. Motif 2 contained the C-X2-C-X8-C part of B-box2, while motifs 3 and 4 contained the last histidine (H) of B-box2 and a more conserved amino acid sequence. Motifs 2, 3, and 4 constituted a relatively complete conserved domain of B-Box2.
Figure 2.

Motif distribution in CpBBX19 and the IV subfamily of the BBX family of Arabidopsis. (a) Motif locations. (b) The width and amino acid sequence of conservative motifs. CpBBX19 is marked by a red point.
3.2. Tissue-Specific Expression of CpBBX19
qRT-PCR revealed that CpBBX19 was expressed in the root, stem, leaf, and flower tissues of wintersweet, but the expression levels varied with location. Expression levels of CpBBX19 were relatively high in cotyledons and inner petals of wintersweet, and in other floral organs (Figure 3a). The expression patterns of CpBBX19 were also explored during the different developmental stages of wintersweet flowers (Figure 3b). The highest level of CpBBX19 expression was detected in the early initiating bloom stage, followed by late initiating bloom and bloom, and the lowest levels of expression were present in the stages of germination, flower-bud, petal-display, and withering.
Figure 3.

Expression pattern analysis of CpBBX19 in wintersweet by qRT-PCR. (a) Expression of CpBBX19 in different tissues of wintersweet. (b) Expression of CpBBX19 in different floral development stages of wintersweet. CpActin was used as the internal reference gene. Data represent the mean of three biological repeats ± SD. Error bars indicate standard deviation. Different lowercase letters (a–e, cd) on bars indicate significant differences (p < 0.05).
3.3. Expression Profiles of CpBBX19 under Abiotic Stresses and Hormone Treatments
To explore the effect of stresses and exogenous hormones on CpBBX17 expression, qRT-PCR was used to determine the transcription level of CpBBX19 in wintersweet under different abiotic stresses (drought, salt, cold, and heat) and hormone treatments (ABA and MeJA). These treatments were selected based on the cis-acting elements in the CpBBX19 promoter region (Table S1). For the ABA, MeJA, and NaCl treatments, expression of CpBBX19 was induced and peaked at 2 h (Figure 4a,b,e). For heat treatment, CpBBX19 expression increased and reached a peak at 2 h, then gradually decreased and was lower than that of the control (untreated; CK) at 24 h. (Figure 4c). For cold treatment, the expression of CpBBX19 was significantly higher than that of the untreated control at 6 h and 12 h (Figure 4d). Under PEG-6000 treatment, expression of CpBBX19 was induced and increased between 2 h and 6 h, then decreased from 6 h to 12 h, before increasing again and peaking at 24 h (Figure 4f).
Figure 4.

Expression profiles of CpBBX19 in response to abiotic stress and hormone treatments. Six-leaf stage wintersweet was exposed to (a) 50 µM ABA, (b) 100 µM MeJA, (c) 42 °C, (d) 4 °C, (e) 300 mM NaCl, and (f) 50% PEG-6000 treatments. RNA was extracted from the leaves at 0, 2, 6, 12, and 24 h after treatment. CpActin was used as the internal reference gene. Data represent the mean of three biological repeats ± SD. Error bars indicate standard deviation. *p < 0.05, ** p < 0.01.
3.4. Overexpression of CpBBX19 Improves Tolerance to Salt and Drought Stress in Arabidopsis
To further study the function of CpBBX19, the expression vector pGWB551-CpBBX19 was constructed and then transferred into wild-type Arabidopsis thaliana (WT). Using the leaves DNA of the T2 lines as templates, the CpBBX19 overexpression lines were identified by PCR (Figure S2), and WT was used as control. Three lines with high, medium, and low expression levels of CpBBX19 (namely, OE-2, OE-5, and OE-9; Figure 5) were selected for further functional analysis. The relative expression levels of OE-2, OE-5, and OE-9 were 1.104 ± 0.080, 0.460 ± 0.019, and 0.211 ± 0.029, respectively. Wild-type plants (WT) were used as a control.
Figure 5.

qRT-PCR identification of CpBBX19 transcript levels in the leaves of nine overexpressing Arabidopsis lines and WT plants. AtActin was used as the internal reference gene. Bars indicate the SE of the mean from three technical replicates and three biological replicates.
To determine the response of CpBBX19 overexpression to NaCl stress, the three transgenic lines and WT plants were irrigated with 300 mM NaCl solution. After 4 days, WT and transgenic lines all showed obvious wilting, but the growth status of the transgenic lines was slightly better than that of WT (Figure 6a). Electrolyte leakage and malondialdehyde (MDA) are reference indicators of the degree of plant damage [48,49]. Electrolyte leakage of the three transgenic lines was 65.24%, 68.76%, and 69.35%, respectively, all significantly lower than that of WT (85.27%) (Figure 6b). In addition, MDA molar concentrations of the transgenic lines (0.409, 0.6785, and 0.701 μmol/g∙Fw, respectively) were markedly lower than that of WT (0.929 μmol/g∙Fw) (Figure 6c). Therefore, transgenic lines have better resistance to damage under salt stress compared with WT plants.
Figure 6.

Heterologous expression of CpBBX19 enhanced salt tolerance of transgenic Arabidopsis plants. (a) Performance of Arabidopsis overexpressing CpBBX19 (OE lines) under NaCl treatment. Bar denotes 1 cm. (b) Electrolyte leakage of WT and OE lines under NaCl treatment. (c) MDA content of WT and OE lines under NaCl treatment. Data represent the mean of three biological repeats ± SD. Error bars indicate standard deviation. Different lowercase letters (a, b, c) on bars indicate significant differences (p < 0.05).
To explore the role of CpBBX19 in drought stress, WT plants and the three transgenic lines were irrigated with 30% PEG. After 36 h, WT plants were markedly wilted compared with transgenic plants (Figure 7a). Measurement of electrolyte leakage showed that WT was the most damaged (67.37% electrolyte leakage) among the analyzed plants, and this measurement was significantly different from that of the three transgenic lines (31.81%, 48.87%, and 62.30%, respectively) (Figure 7b). MDA content in WT plants (0.678 μmol/g∙Fw) was almost three times that of OE-2 plants (0.226 μmol/g∙Fw), and twice that of OE-5 (0.360 μmol/g∙Fw) and OE-9 (0.430 μmol/g∙Fw) plants (Figure 7c). In summary, overexpression of CpBBX19 improved the drought tolerance of transgenic Arabidopsis.
Figure 7.

Heterologous expression of CpBBX19 enhanced drought tolerance of transgenic Arabidopsis. (a) Performance of Arabidopsis overexpressing CpBBX19 (OE lines) under 30% PEG-6000 treatment. Bar denotes 1 cm. (b) Electrolyte leakage of WT and OE lines under 30% PEG-6000 treatment. (c) MDA content of WT and OE lines under 30% PEG-6000 treatment. Data represent the mean of three biological repeats ± SD. Error bars indicate standard deviation. Different lowercase letters (a, b, c) on bars indicate significant differences (p < 0.05).
4. Discussion
Transcription factor regulation is a crucial link in gene expression. B-box-like zinc-finger proteins in animal cells are involved in many cellular processes, such as apoptosis, cell cycle regulation, virus response, and other intracellular physiological processes [50,51]. Research on the structure and function of B-box-like proteins in plants has also made some progress. BBX family members have been identified in Arabidopsis [16], rice [30], apple [27], Toona Sinens [24], wheat [52], pear [25], and Petunia hybrida [53], with the number of BBX members varying among different species. Wintersweet is an ornamental plant that blossoms in winter in China. However, there are no reports on BBX transcription factors of wintersweet. In this study, a novel BBX transcription factor gene, CpBBX19, was isolated based on information from the wintersweet transcriptome database [39]. Multiple sequence alignment revealed that CpBBX19 protein shared a high sequence similarity with the BBX19 protein sequences of other plants. Two B-box domain motifs were found in the CpBBX19 sequence (Figure 1a), which was consistent with the characteristics of group IV members of BBX proteins. Phylogenetic analysis showed that CpBBX19 clustered with group IV BBX proteins and was closely related to AtBBX19 (Figure 1b). Prediction of the subcellular localization of protein CpBBX19 indicated that it was located in the nucleus, and it was speculated that this transcription factor might be involved in the transcription level of other genes.
Gene function can be reflected by the expression patterns of a gene to a certain degree [42]. OsBBX13 had the highest expression in leaves, then in roots, and lower expression in stems and young panicles [44]. CmBBX19 had higher transcript contents in leaves, flowers, and stems, but lower levels in roots [54]. PuBBX24 was expressed in the roots, stems, leaves, flowers, and fruits of Pyrus ussuriensis, but expression levels were highest in stems and lowest in leaves [55]. In the current study, CpBBX19 was expressed in the stems, roots, leaves, and flowers of wintersweet, with highest expression found in cotyledons and inner petals and lower expression detected in roots, stems, and old leaves. The expression patterns of PuBBX24, CmBBX19, and OsBBX13 were slightly different from that of CpBBX19, but all four genes exhibited higher expression in leaves and flowers and lower expression in roots. We speculated that the gene CpBBX19 may be involved in leaf morphogenesis and may play a role in the inner petal formation of wintersweet flowers. At the stage of floral development, the highest expression of CpBBX19 was in the early flowering stage, followed by late initiating bloom and bloom, and the lowest level of expression was detected in the stages of germination, flower-bud, petal-display, and withering. As wintersweet blossoms in cold winter and high expression of CpBBX19 was detected in the early initiating bloom stage, this gene may be related to the stress-resistant protection mechanism of wintersweet against adverse environments. CpBBX19 may be involved in initiating the relevant regulatory network to enhance the stress resistance of floral organs.
Cis-acting elements play a key role in gene transcription and expression [41]. Most of the cis-acting elements with biological functions are predominantly active 50 bp upstream of the transcription start site (TSS), and most of the transcription factor binding sites (TFBSs) are in the region from −1000 bp to +200 bp with respect to the TSS [56]. Analysis of the promoter of CpBBX19 revealed the presence of a TATA-box, which is a core promoter element usually located 25–35 bp upstream of the TSS [57,58,59,60]. The promoter of CpBBX19 also contained a CAAT-box, which also commonly exists in promoters and is usually located 75 bp upstream of TSS [57,58,59,60]. Many cis-acting elements of abiotic stresses and hormone responses, such as ABRE, ARE, MBS, MYB, MYC, TC-rich, TCA-element, and TGACG-motif, were also present. These observations indicated that CpBBX19 may be involved in abiotic stress and hormone responses. Expression of the BBX gene family members SlBBX7, SlBBX9, and SlBBX20 was induced by cold stress in tomato [61], and there were differences in the expression of some GhBBX genes after ABA treatment in Gossypium hirsutum [62]. PavBBX6 and PavBBX9 showed strong induction under ABA, GA, and BR treatment in sweet cherry fruit [63]. Furthermore, abiotic stress such as drought, low temperature, and salt stress strongly induced expression of OsBBX1, OsBBX2, OsBBX8, OsBBX19, and OsBBX24 [31], and expression of OsBBX2, OsBBX7, OsBBX17, OsBBX19, and OsBBX24 was upregulated by GA, SA, and MeJA hormones at different time points [31]. These results are consistent with those of CpBBX19, with the expression of CpBBX19 being induced by abiotic stress (drought, salt, cold, and heat) and exogenous hormones (ABA and MeJA) (Figure 4) in the current study. Therefore, CpBBX19 may be involved in the response of wintersweet to drought, salt, cold, heat, ABA, and MeJA. However, the expression pattern of BBX genes in some plants was opposite to the above pattern. For example, expression of CmBBX19 was downregulated by drought stress and ABA treatment [54], and the expression of GhBBX5, GhBBX23, and GhBBX28 in cotton was continuously downregulated after PEG treatment for 1 h, reaching the lowest expression point at 12 h, and then increased at 24 h [62]. These differences suggested that BBX genes have different expression patterns in different plants.
Electrolyte leakage and MDA content, as an important index of plant stress resistance, can reflect the degree of damage of the plant cell membrane system under adversity [64]. The increase of electrolyte leakage and MDA content indicates the cell membrane structure damage and intensified membrane permeability [65]. To verify our hypothesis and facilitate understanding of the functions of CpBBX19, Arabidopsis plants overexpressing CpBBX19 were obtained. Growth of the transgenic lines was better than that of WT lines under drought stress, and the electrolyte leakage and MDA content of these transgenic lines were much lower than those of WT lines. Under salt stress, all plants (WT and OE lines) were obviously withering, but the electrolyte leakage and MDA content of OE lines were significantly lower than those of WT. This indicated that the transgenic Arabidopsis plants had enhanced salt and drought resistance compared with the WT plants. Some functions of BBX genes have been verified. For example, CmBBX24 transgenic Arabidopsis was more tolerant to low temperature and drought stress than the WT lines were [66]; transgenic CmBBX22 in Arabidopsis improved the drought resistance of Arabidopsis [38]; heterologous expression of MdBBX10 enhanced the tolerance of transgenic Arabidopsis to salt and drought stress [28], and compared with WT plants, overexpression of AtBBX24 could improve the tolerance of Arabidopsis to salt stress [67]. However, because of the inclusion of an EAR motif in CmBBX19, the drought tolerance of CmBBX19-overexpressing (CmBBX19-OX) lines was lower than that of WT [54]. In plants, proteins containing an EAR motif have been proven to be transcriptional repressors and are instrumental in many biological processes, including responses to hormones and biotic and abiotic stresses, floral transition, and meristem maintenance [68,69]. In general, different BBX transcription factors have complex roles in plant responses to environmental stresses.
5. Conclusions
To date, there are no reports on the BBX gene family in wintersweet. In this study, CpBBX19 from wintersweet was isolated and its expression pattern and functional characterization were investigated. Expression of CpBBX19 was induced by multiple abiotic stresses (drought, salt, cold, and heat), as well as by hormone treatments (ABA and MeJA). Heterologous expression of CpBBX19 in Arabidopsis enhanced tolerance to drought and salt stresses. These findings indicate that CpBBX19 plays a role in drought and salt stress responses. Moreover, this study also lays a strong foundation for further investigations into BBX genes of wintersweet and enriches understanding of the molecular mechanism of stress resistance in wintersweet.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/genes12091456/s1. Table S1: List of primers used in this study; Table S2: Analysis of cis-acting elements of promoters; Table S3: Names and accession numbers used for phylogenetic analysis; Figure S1. The simple map of the pGWB551 vector.
Author Contributions
Conceptualization, H.W. and X.W.; Data curation, H.W., X.W. and Y.C.; Funding acquisition, H.L. and S.S.; Methodology, H.Z. and R.H.; Software, H.W. and X.W.; Writing—original draft, H.W. and X.W.; Writing—review and editing, S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant number 31971711), the Chongqing Science and Technology Bureau (cstc2018jscx-mszdX0004), and Natural Science Foundation Project of Chongqing (No. cstc2019jcyj-msxmX0687).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Hu H., Chai N., Zhu H., Li R., Huang R., Wang X., Liu D., Li M., Song X., Sui S. Factors affecting vegetative propagation of wintersweet (Chimonanthus praecox) by softwood cuttings. HortScience. 2020;55:1853–1860. doi: 10.21273/HORTSCI15289-20. [DOI] [Google Scholar]
- 2.Tian J., Ma Z., Zhao K., Zhang J., Xiang L., Chen L. Transcriptomic and proteomic approaches to explore the differences in monoterpene and benzenoid biosynthesis between scented and unscented genotypes of wintersweet. Physiol. Plant. 2019;166:478–493. doi: 10.1111/ppl.12828. [DOI] [PubMed] [Google Scholar]
- 3.Kamran H.M., Hussain S.B., Junzhong S., Xiang L., Chen L.-Q. Identification and molecular characterization of geranyl diphosphate synthase (GPPS) genes in wintersweet flower. Plants. 2020;9:666. doi: 10.3390/plants9050666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Shang J., Tian J., Cheng H., Yan Q., Li L., Jamal A., Xu Z., Xiang L., Saski C.A., Jin S., et al. The chromosome-level wintersweet (Chimonanthus praecox) genome provides insights into floral scent biosynthesis and flowering in winter. Genome Biol. 2020;21:1–28. doi: 10.1186/s13059-020-02088-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Li R., Ma J., Liu H., Wang X., Li J., Li Z., Li M., Sui S., Liu D. Overexpression of CpSIZ1, a SIZ/PIAS-Type SUMO E3 ligase from wintersweet (Chimonanthus praecox), delays flowering, accelerates leaf senescence and enhances cold tolerance in Arabidopsis. Plant Mol. Biol. Rep. 2021:1–16. doi: 10.1007/s11105-020-01250-x. [DOI] [Google Scholar]
- 6.Sui S., Luo J., Liu D., Ma J., Men W., Fan L., Bai Y., Li M. Effects of hormone treatments on cut flower opening and senescence in wintersweet (Chimonanthus praecox) HortScience. 2015;50:1365–1369. doi: 10.21273/HORTSCI.50.9.1365. [DOI] [Google Scholar]
- 7.Liu Y., Xie L., Liang X., Zhang S. CpLEA5, the late embryogenesis abundant protein gene from Chimonanthus praecox, possesses low temperature and osmotic resistances in prokaryote and eukaryotes. Int. J. Mol. Sci. 2015;16:26978–26990. doi: 10.3390/ijms161126006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang L.-H., Jia B., Zhuo R.-Y., Liu J.-L., Pan H.-Y., Baldwin T.C., Zhang S.-H. An acyl–acyl carrier protein thioesterase gene isolated from wintersweet (Chimonanthus praecox), CpFATB, enhances drought tolerance in transgenic tobacco (Nicotiana tobaccum) Plant Mol. Biol. Rep. 2011;30:433–442. doi: 10.1007/s11105-011-0359-5. [DOI] [Google Scholar]
- 9.Qin H., Sui S., Li M., Lei X., Yu G. Molecular characterization and expression analysis of a now COR413 gene from win-tersweet [Chimonanthus praecox (L.) link] Chin. J. Biochem. Mol. Biol. 2006;22:547–552. doi: 10.13865/j.cnki.cjbmb.2006.07.006. [DOI] [Google Scholar]
- 10.Liu D.F., Wang X., Dai Y., Yang J.F., Ma J., Li M.Y., Sui S.Z. Cloning and function analysis of CpTAF10 from wintersweet (Chimonanthus praecox) Sci. Silvae Sin. 2019;55:176–183. doi: 10.11707/j.1001-7488.20190621. [DOI] [Google Scholar]
- 11.Ahmad A., Qamarunnisa S., Jamil I., Raza S., Khatoon K. In-silico and phylogenetic analysis of DREB transcription factor in Solanum melongena L. Pak. J. Bot. 2021;53:1271–1279. doi: 10.30848/PJB2021-4(2). [DOI] [Google Scholar]
- 12.Akdemir H. Evaluation of transcription factor and aquaporin gene expressions in response to Al2O3 and ZnO nanoparticles during barley germination. Plant Physiol. Biochem. 2021;166:466–476. doi: 10.1016/j.plaphy.2021.06.018. [DOI] [PubMed] [Google Scholar]
- 13.Robson F., Costa M.M., Hepworth S., Vizir I., Piñeiro M., Reeves P., Putterill J., Coupland G. Functional importance of conserved domains in the flowering-time gene CONSTANS demonstrated by analysis of mutant alleles and transgenic plants. Plant J. 2002;28:619–631. doi: 10.1046/j.1365-313x.2001.01163.x. [DOI] [PubMed] [Google Scholar]
- 14.Lira B.S., Oliveira M.J., Shiose L., Wu R.T.A., Rosado D., Lupi A.C.D., Freschi L., Rossi M. Light and ripening-regulated BBX protein-encoding genes in Solanum lycopersicum. Sci. Rep. 2020;10:1–14. doi: 10.1038/s41598-020-76131-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Talar U., Kiełbowicz-Matuk A. Beyond Arabidopsis: BBX regulators in crop plants. Int. J. Mol. Sci. 2021;22:2906. doi: 10.3390/ijms22062906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Holtan H.E., Bandong S., Marion C.M., Adam L., Tiwari S., Shen Y., Maloof J., Maszle D.R., Ohto M.-A., Preuss S., et al. BBX32, an Arabidopsis B-box protein, functions in light signaling by suppressing HY5-regulated gene expression and interacting with STH2/BBX21. Plant Physiol. 2011;156:2109–2123. doi: 10.1104/pp.111.177139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gangappa S.N., Botto J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014;19:460–470. doi: 10.1016/j.tplants.2014.01.010. [DOI] [PubMed] [Google Scholar]
- 18.Khanna R., Kronmiller B., Maszle D.R., Coupland G., Holm M., Mizuno T., Wu S.-H. The Arabidopsis B-box zinc finger family. Plant Cell. 2009;21:3416–3420. doi: 10.1105/tpc.109.069088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Datta S., Hettiarachchi G., Deng X.-W., Holm M. Arabidopsis CONSTANS-LIKE3 is a positive regulator of red light signaling and root growth. Plant Cell. 2005;18:70–84. doi: 10.1105/tpc.105.038182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Datta S., Hettiarachchi C., Johansson H., Holm M. SALT TOLERANCE HOMOLOG2, a B-box protein in Arabidopsis that activates transcription and positively regulates light-mediated development. Plant Cell. 2007;19:3242–3255. doi: 10.1105/tpc.107.054791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Liu L.-J., Zhang Y.-C., Li Q.-H., Sang Y., Mao J., Lian H.-L., Wang L., Yang H.-Q. COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis. Plant Cell. 2008;20:292–306. doi: 10.1105/tpc.107.057281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yan H., Marquardt K., Indorf M., Jutt D., Kircher S., Neuhaus G., Rodríguez-Franco M. Nuclear localization and interaction with COP1 are required for STO/BBX24 function during photomorphogenesis. Plant Physiol. 2011;156:1772–1782. doi: 10.1104/pp.111.180208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Crocco C.D., Botto J. BBX proteins in green plants: Insights into their evolution, structure, feature and functional diversification. Gene. 2013;531:44–52. doi: 10.1016/j.gene.2013.08.037. [DOI] [PubMed] [Google Scholar]
- 24.Liping R., Jinbo Z., Xiaohan C., Wenyang W., Dandan Y., Xiaohui S. Transcriptome-wide identification and functional characterization of BBX transcription factor family in Toona sinensis. Pak. J. Bot. 2021;53:915–921. doi: 10.30848/PJB2021-3(8). [DOI] [Google Scholar]
- 25.Zou Z., Wang R., Wang R., Yang S., Yang Y. Genome-wide identification, phylogenetic analysis, and expression profiling of the BBX family genes in pear. J. Hortic. Sci. Biotechnol. 2017;93:37–50. doi: 10.1080/14620316.2017.1338927. [DOI] [Google Scholar]
- 26.Wang Q., Tu X., Zhang J., Chen X., Rao L. Heat stress-induced BBX18 negatively regulates the thermotolerance in Arabidopsis. Mol. Biol. Rep. 2012;40:2679–2688. doi: 10.1007/s11033-012-2354-9. [DOI] [PubMed] [Google Scholar]
- 27.Liu X., Li R., Dai Y., Chen X., Wang X. Genome-wide identification and expression analysis of the B-box gene family in the Apple (Malus domestica Borkh.) genome. Mol. Genet. Genom. 2017;293:303–315. doi: 10.1007/s00438-017-1386-1. [DOI] [PubMed] [Google Scholar]
- 28.Liu X., Li R., Dai Y., Yuan L., Sun Q., Zhang S., Wang X. A B-box zinc finger protein, Md BBX10, enhanced salt and drought stresses tolerance in Arabidopsis. Plant Mol. Biol. 2019;99:437–447. doi: 10.1007/s11103-019-00828-8. [DOI] [PubMed] [Google Scholar]
- 29.Kiełbowicz-Matuk A., Rey P., Rorat T. Interplay between circadian rhythm, time of the day and osmotic stress constraints in the regulation of the expression of a solanum double B-box gene. Ann. Bot. 2014;113:831–842. doi: 10.1093/aob/mct303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huang J., Zhao X., Weng X., Wang L., Xie W. The rice B-box zinc finger gene family: Genomic identification, characterization, expression profiling and diurnal analysis. PLoS ONE. 2012;7:e48242. doi: 10.1371/journal.pone.0048242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Shalmani A., Jing X.-Q., Shi Y., Muhammad I., Zhou M.-R., Wei X.-Y., Chen Q.-Q., Li W.-Q., Liu W.-T., Chen K.-M. Characterization of B-box gene family and their expression profiles under hormonal, abiotic and metal stresses in Poaceae plants. BMC Genom. 2019;20:27. doi: 10.1186/s12864-018-5336-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Min J.-H., Chung J.-S., Lee K.-H., Kim C.S. The CONSTANS-like 4 transcription factor, AtCOL4, positively regulates abiotic stress tolerance through an abscisic acid-dependent manner in Arabidopsis. J. Integr. Plant Biol. 2015;57:313–324. doi: 10.1111/jipb.12246. [DOI] [PubMed] [Google Scholar]
- 33.Yang Y., Ma C., Xu Y., Wei Q., Imtiaz M., Lan H., Gao S., Cheng L., Wang M., Fei Z., et al. A zinc finger protein regulates flowering time and abiotic stress tolerance in chrysanthemum by modulating gibberellin biosynthesis. Plant Cell. 2014;26:2038–2054. doi: 10.1105/tpc.114.124867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sánchez J.-P., Duque P., Chua N.-H. ABA activates ADPR cyclase and cADPR induces a subset of ABA-responsive genes in Arabidopsis. Plant J. 2004;38:381–395. doi: 10.1111/j.1365-313X.2004.02055.x. [DOI] [PubMed] [Google Scholar]
- 35.Li P., Li X., Jiang M. CRISPR/Cas9-mediated mutagenesis of WRKY3 and WRKY4 function decreases salt and Me-JA stress tolerance in Arabidopsis thaliana. Mol. Biol. Rep. 2021;48:5821–5832. doi: 10.1007/s11033-021-06541-4. [DOI] [PubMed] [Google Scholar]
- 36.Hu Q., Cui H., Ma C., Li Y., Yang C., Wang K., Sun Y. Lipidomic metabolism associated with acetic acid priming-induced salt tolerance in Carex rigescens. Plant Physiol. Biochem. 2021;167:665–677. doi: 10.1016/j.plaphy.2021.08.045. [DOI] [PubMed] [Google Scholar]
- 37.Zhong J.Y., Zhang Y.L. Effect of low temperature on MDA content and relative conductivity of different almond trees branch. Tianjin Agricultural Sciences. 2013;19:93–96. doi: 10.3969/j.issn.1006-6500.2013.05.025. [DOI] [Google Scholar]
- 38.Liu Y., Chen H., Ping Q., Zhang Z., Guan Z., Fang W., Chen S., Chen F., Jiang J., Zhang F. The heterologous expression of CmBBX22 delays leaf senescence and improves drought tolerance in Arabidopsis. Plant Cell Rep. 2018;38:15–24. doi: 10.1007/s00299-018-2345-y. [DOI] [PubMed] [Google Scholar]
- 39.Liu D., Sui S., Ma J., Li Z., Guo Y., Luo D., Yang J., Li M. Transcriptomic analysis of flower development in wintersweet (Chimonanthus praecox) PLoS ONE. 2014;9:e86976. doi: 10.1371/journal.pone.0086976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wu C.L., Hu N.Z. Studies on the flower form and blooming characteristics of the wintersweet. Acta Hortic. Sin. 1995;22:277–282. [Google Scholar]
- 41.Wang D., Chen Q., Chen W., Liu X., Xia Y., Guo Q., Jing D., Liang G. A WRKY transcription factor, EjWRKY17, from Eriobotrya japonica enhances drought tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2021;22:5593. doi: 10.3390/ijms22115593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Huang R., Liu D., Ma J., Li M., Sui S. CpWRKY71, a WRKY transcription factor gene of wintersweet (Chimonanthus praecox), promotes flowering and leaf senescence in Arabidopsis. Int. J. Mol. Sci. 2019;20:5325. doi: 10.3390/ijms20215325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lin J., Liu D., Wang X., Ahmed S., Li M., Kovinich N., Sui S. Transgene CpNAC68 from wintersweet (Chimonanthus praecox) improves Arabidopsis survival of multiple abiotic stresses. Plants. 2021;10:1403. doi: 10.3390/plants10071403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Li N., Zhang C., Wang W.P. Cloning, expression and bioinformatics analysis of zinc finger protein gene OsBBX13 in rice. Mol. Plant Breed. 2019;17:1393–1400. doi: 10.13271/j.mpb.017.001393. [DOI] [Google Scholar]
- 45.Liu H., Huang R., Ma J., Sui S., Guo Y., Liu D., Li Z., Lin Y., Li M. Two C3H type zinc finger protein genes, CpCZF1 and CpCZF2, from Chimonanthus praecox affect stamen development in Arabidopsis. Genes. 2017;8:199. doi: 10.3390/genes8080199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Clough S.J., Bent A.F. Floral dip: A simplified method for agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16:735–743. doi: 10.1046/j.1365-313x.1998.00343.x. [DOI] [PubMed] [Google Scholar]
- 47.Khan S., Qureshi M.I., Alam T., Abdin M.Z. Protocol for isolation of genomic DNA from dry and fresh roots of medicinal plants suitable for RAPD and restriction digestion. Afr. J. Biotechnol. 2007;6:175–178. doi: 10.5897/AJB06.612. [DOI] [Google Scholar]
- 48.Heath R.L., Packer L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968;125:189–198. doi: 10.1016/0003-9861(68)90654-1. [DOI] [PubMed] [Google Scholar]
- 49.Seliem M., El-Mahrouk M., El-Banna A., Hafez Y., Dewir Y. Micropropagation of Philodendron selloum: Influence of copper sulfate on endophytic bacterial contamination, antioxidant enzyme activity, electrolyte leakage, and plant survival. S. Afr. J. Bot. 2021;139:230–240. doi: 10.1016/j.sajb.2021.01.024. [DOI] [Google Scholar]
- 50.Meroni G., Diez-Roux G. TRIM/RBCC, a novel class of ‘single protein RING finger’ E3 ubiquitin ligases. Bioessays. 2005;27:1147–1157. doi: 10.1002/bies.20304. [DOI] [PubMed] [Google Scholar]
- 51.Torok M., Etkin L.D. Two B or not two B? Overview of the rapidly expanding B-box family of proteins. Differentation. 2001;67:63–71. doi: 10.1046/j.1432-0436.2001.067003063.x. [DOI] [PubMed] [Google Scholar]
- 52.Chen S., Jiang W., Yin J., Wang S., Fang Z., Ma D., Gao D. Genome-wide mining of wheat B-BOX zinc finger (BBX) gene family provides new insights into light stress responses. Crop. Pasture Sci. 2021;72:17. doi: 10.1071/CP20342. [DOI] [Google Scholar]
- 53.Wen S., Zhang Y., Deng Y., Chen G., Yu Y., Wei Q. Genomic identification and expression analysis of the BBX transcription factor gene family in Petunia hybrida. Mol. Biol. Rep. 2020;47:6027–6041. doi: 10.1007/s11033-020-05678-y. [DOI] [PubMed] [Google Scholar]
- 54.Xu Y., Zhao X., Aiwaili P., Mu X., Zhao M., Zhao J., Cheng L., Ma C., Gao J., Hong B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. 2020;103:1783–1795. doi: 10.1111/tpj.14863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wang R.H., Song M.Y., Wang R., Yang Y.J. Expression pattern and function in juvenile regulation of B-box gene PuBBX24 in Pyrus ussuriensis. Acta Hortic. Sin. 2019;46:1458–1472. doi: 10.16420/j.issn.0513-353x.2018-1020. [DOI] [Google Scholar]
- 56.Yu C.-P., Lin J.-J., Li W.-H. Positional distribution of transcription factor binding sites in Arabidopsis thaliana. Sci. Rep. 2016;6:25164. doi: 10.1038/srep25164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Molina C., Grotewold E. Genome wide analysis of Arabidopsis core promoters. BMC Genom. 2005;6:25. doi: 10.1186/1471-2164-6-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhang X.H., Qi Y.X. Analysis on TATA-box, GC-box and CAAT-box in eukaryotic promoters. J. Anhui Agr. Sci. 2008;36:1380–1381, 1395. doi: 10.13989/j.cnki.0517-6611.2008.04.008. [DOI] [Google Scholar]
- 59.Shahmuradov I.A. PlantProm: A database of plant promoter sequences. Nucleic Acids Res. 2003;31:114–117. doi: 10.1093/nar/gkg041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Florquin K. Large-scale structural analysis of the core promoter in mammalian and plant genomes. Nucleic Acids Res. 2005;33:4255–4264. doi: 10.1093/nar/gki737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bu X., Wang X., Yan J., Zhang Y., Zhou S., Sun X., Yang Y., Ahammed G.J., Liu Y., Qi M., et al. Genome-wide characterization of B-box gene family and its roles in responses to light quality and cold stress in tomato. Front. Plant Sci. 2021;12:1–18. doi: 10.3389/fpls.2021.698525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Feng Z., Li M., Li Y., Yang X., Wei H., Fu X., Ma L., Lu J., Wang H., Yu S. Comprehensive identification and expression analysis of B-box genes in cotton. BMC Genom. 2021;22:1–16. doi: 10.1186/s12864-021-07770-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang Y., Zhai Z., Sun Y., Feng C., Peng X., Zhang X., Xiao Y., Zhou X., Wang W., Jiao J., et al. Genome-wide identification of the B-box genes that respond to multiple ripening related signals in sweet cherry fruit. Int. J. Mol. Sci. 2021;22:1622. doi: 10.3390/ijms22041622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lu J.X., Jiang H.Y., Li W. Effects of low temperature stress on the cold resistance of rootstock and branch of wine grapes. J. Fruit Sci. 2012;29:1040–1046. doi: 10.13925/j.cnki.gsxb.2012.06.020. [DOI] [Google Scholar]
- 65.Li H.Y., Guo X.W. Influence of NaCl on activities of protective enzymes and MDA content in grape rootstock leaves. J. Fruit Sci. 2008;25:240–243. doi: 10.13925/j.cnki.gsxb.2008.02.021. [DOI] [Google Scholar]
- 66.Imtiaz M., Yang Y., Liu R., Xu Y., Khan M.A., Wei Q., Gao J., Hong B. Identification and functional characterization of the BBX24 promoter and gene from chrysanthemum in Arabidopsis. Plant Mol. Biol. 2015;89:1–19. doi: 10.1007/s11103-015-0347-5. [DOI] [PubMed] [Google Scholar]
- 67.Nagaoka S. Salt tolerance-related protein STO binds to a Myb transcription factor homologue and confers salt tolerance in Arabidopsis. J. Exp. Bot. 2003;54:2231–2237. doi: 10.1093/jxb/erg241. [DOI] [PubMed] [Google Scholar]
- 68.Kagale S., Rozwadowski K. EAR motif-mediated transcriptional repression in plants. Epigenetics. 2011;6:141–146. doi: 10.4161/epi.6.2.13627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kazan K. Negative regulation of defence and stress genes by EAR-motif-containing repressors. Trends Plant Sci. 2006;11:109–112. doi: 10.1016/j.tplants.2006.01.004. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Not applicable.