

Abstract
The role of spatial attention for visual perception has been thoroughly studied in primates, but less so in mice. Several behavioral tasks in mice reveal spatial attentional effects, with similarities to observations in primates. Pairing these tasks with large-scale, cell-type specific techniques could enable deeper access to underlying mechanisms, and help define the utility and limitations of resolving attentional effects on visual perception and neural activity in mice. In this review, we evaluate behavioral and neural evidence for visual spatial attention in mice, assess how specializations of the mouse visual system and behavioral repertoire impact interpretation of spatial attentional effects, and outline how several measurement and manipulation techniques in mice could precisely test and refine models of attentional modulation across scales.
Keywords: Vision, spatial attention, mouse, visual cortex
New opportunities for study of neural circuits & spatial attention in mice
The importance of selective attention for perception of the visual world has been recognized for centuries. Pioneering studies in primates pairing rigorously controlled visual behaviors with electrophysiological recordings unveiled selective attentional effects on individual neurons. Subsequent studies revealed that selective visual spatial attention improves behavioral and neural sensitivity to stimuli at attended locations [1, 2]. This work inspired detailed models of spatial attentional effects in single neurons [3, 4], yet critical barriers remain for measuring and testing the underlying mechanisms in primates with cellular and synaptic specificity. Recently, several groups have established methods to study selective visual spatial attention in mice [5–9]. Mice provide a unique platform to study mechanisms of visual spatial processing during behavior, due to techniques allowing large-scale measurement and manipulation of neural activity with cellular and circuit specificity [10–18]. This review seeks to 1) assess the recent evidence for selective spatial attention in mice, 2) discuss advantages and constraints for measuring visuospatial attention in mice, and 3) outline how pairing multiple behavioral approaches with novel techniques in mice could address open questions about circuits and biophysical mechanisms mediating the effects of visual spatial attention.
This proposed approach enables mechanistic testing of several influential models of spatial attention. These models operate at phenomenological, circuit, and neural population levels. We highlight how these three model classes account for attentional modulations, and discuss how experiments in transgenic mice could test and integrate these models — bridging scales from cellular mechanisms, to circuits, to large-scale networks.
We begin by examining various approaches and key results from recent visuospatial attention studies in mice. We then examine limitations of these findings in comparison to primate work. Next, we consider how attentional modulation in mice compares to the effects of locomotion on visual processing. We then highlight the potential for combining multiple visuospatial attentional behaviors with advanced techniques to potentially reveal new insights for the mechanisms mediating effects of spatial attention in the mammalian brain.
We define visual spatial attention as a behavioral process that improves detectability or discriminability of stimuli in particular regions of visual space. Importantly, spatial attention selectively improves behavioral responses to stimuli in certain locations, but at the cost of diminished responses elsewhere. Throughout the review, we use the terms “attentional effects” or “attentional modulation” to describe behavioral and neural changes measured during a suitable behavior, and “neural mechanisms of attention” to describe the underlying cellular, circuit, and population level factors necessary to measure attentional effects. Divided attention is another emerging topic in studies in mice [19], but lies outside the scope of this review. Recent reviews cover neural mechanisms of attention more broadly [2, 20], and others review neural circuits and sensory processing in mice [21, 22]. A central premise of our approach is that all mammalian visual systems show orderly retinotopic and spatial organization [23–26]. Thus, examining the neuronal basis of attentional effects in the visuospatial domain facilitates comparisons across studies and species.
Behavioral tasks & evidence for spatial attention in mice
Directing spatial attention with explicit visual cues
Measuring attentional effects requires a suitably controlled behavioral task (Figure 1). Designing such tasks for mice presents several challenges. First, the locus of attention must be directed. In primates, attention is often directed using an explicit visual “cue” which indicates the relevant spatial location [27]. This approach has been extended to mice [5, 8] and provides several benefits. An explicit cue aids interpretability of ensuing changes in visual detection speed and accuracy during each trial. The location and validity of the cue are controlled to accurately predict stimulus changes on a subset of trials within a block or experiment (e.g. 80% valid vs 20% invalid). On validly cued trials, mice detect ensuing stimuli more accurately and rapidly than on invalidly cued trials [5, 8], suggesting the cue selectively improves behavior with spatial specificity. This distinguishes selective attention from general alertness that would improve behavior regardless of cue location. However, the size, strength, and timing of the cue relative to the task-relevant stimulus could produce complex facilitative or suppressive effects, and conflate purely stimulus-driven effects [28] with attentional effects. These factors could be assessed using cues and task-relevant stimuli outside of the behavioral task and measuring neural responses. Another limitation of cued attentional studies in mice is that these studies have typically presented cues at the same location for blocks of trials, or even entire sessions within each day. Fixing cues to a single position throughout sessions provides endogenous spatial context, so it is not certain that behavior is guided explicitly by the cues. To our knowledge, trial-by-trial cueing has not yet been achieved in mice. The current practice of maintaining cue location within blocks of trials reduces temporal control over the locus of attention, and hampers insights into fast timescale neural effects. Nonetheless, cueing tasks in mice provide a way for future studies to approach single trial timescales.
Figure 1: Considerations for designing spatial attentional tasks in mice.
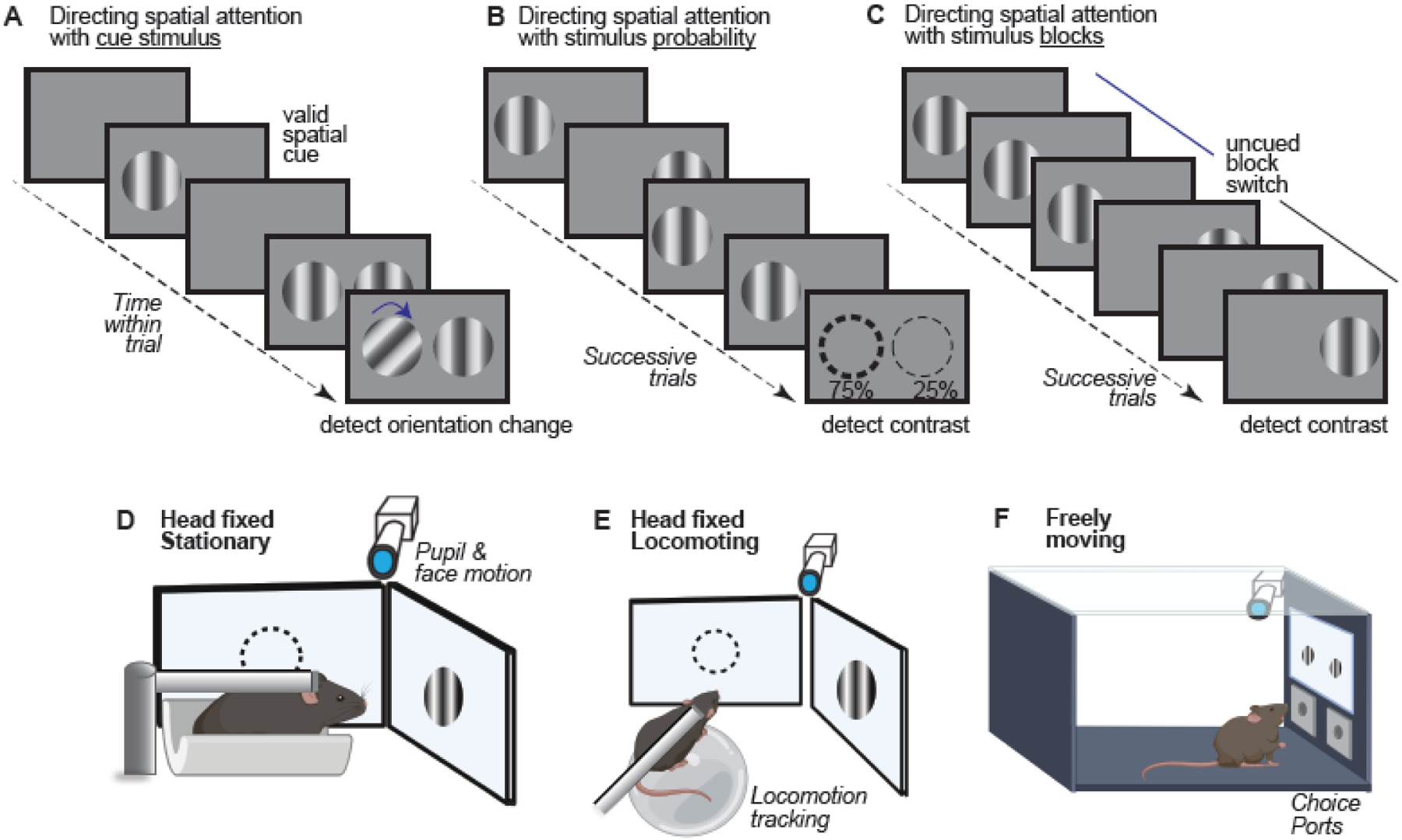
Top (A,B,C): Methods for cueing attention. There are several methods for explicitly or implicitly directing the locus of spatial attention in behavioral assays for mice. Cueing paradigms (A) make use of an explicit cue which instructs the animal about the spatial position where an ensuing stimulus is likely [5, 8]. Manipulating cue validity allows for the direct measurement of the neural and behavioral effects of attention. (B) Adjusting the relative probability that a stimulus appears in a particular location serves as an implicit attentional cue [7]. Over successive trials, the animal learns the distribution of stimulus locations, and allocates attentional resources accordingly. (C) In a blocked trial design [6], the stimulus appears in the same location with 100% probability. This allows the experimenter to track attentional effects in blocks of trials at a given spatial location. Bottom (D,E,F): Mouse positioning and freedom of movement. The position of the mouse and freedom of movement are important considerations for attentional behaviors. In head-fixed preparations [5–8] (D–E), headplates allow experimenters to restrict movement and perform repeatable acute experiments. Head-fixed preparations allow experimenters to control the location of the visual stimulus and track any changes in pupil size and eye position. In stationary head-fixed preparations [6] (D), mice are placed in an apparatus which limits body movement. In locomoting tasks [5, 7, 8] (E), mice run freely on a wheel or treadmill. (F) Freely moving preparations [9] allow the mouse to interact with the environment in a more natural way, but at the expense of some experimental control. Mouse schematics in D – F produced with BioRender.
Directing spatial attention by biasing the location of stimulus appearance
Attention can also be directed without cues by manipulating the likelihood of stimulus appearance in particular locations. When the stimulus appears in a more heavily weighted location (e.g., 80% probability in one location, 20% in another), mice show greater responsiveness at the high likelihood location [7]. However, without cues, mice must integrate information across trials to infer the relative weighting at each location. This presents difficulty in precisely identifying the time course of integration and the moment when attention shifts; further, learning weighted probabilities may obviate the need for trial-by-trial attention in multiple locations.
Directing spatial attention by presenting stimuli in different locations across blocks
A third approach to direct the locus of attention uses blocks of consecutive trials to present the task-relevant (rewarded) stimulus at a fixed location [6]. Stimuli appear at this location with 100% likelihood within a block of trials; then, without warning or explicit cueing, the stimulus switches to a new location for the next block. An advantage of this approach is that the experimenter can precisely control when the salience and positional validity of the stimulus changes (around block switches), and track if behavior shows both costs and benefits consistent with shifting the locus of attention. Another advantage is that anchoring attention to one location for the entirety of a block permits studying the effects of prolonged spatial attention on spontaneous as well as sensory-evoked neural activity at longer timescales [6, 29–32].
Directing spatial attention in freely moving mice with explicit visual cues
The preceding psychometric tasks all involve head-fixed mice that lick a reward spout, fundamental methods for study of sensory perception in rodents [33, 34]. Another recent visual spatial detection task in freely moving mice measures visually-guided nose pokes. Stimuli appear with differing location probability, and again mice responded faster and more accurately to stimuli in higher probability locations [9]. Thus, more naturalistic conditions can be used to measure spatial attentional effects, with careful consideration of effects of movement (Box 1). This study also showed another important aspect of spatial attention – spatial filtering. Mice were trained to ignore stimuli in behaviorally irrelevant parts of visual space but enhance detection at relevant locations. Filter tasks are commonly used to study attention in primates [35], and the forementioned study [9] demonstrates their utility in mice.
Box 1: Body movements and their implications for visual spatial attention tasks.
An important consideration for spatial attention tasks in mice concerns whether the mice are constrained or free to move. Many studies of mouse visual perception make use of head-fixed mice that run freely on a treadmill or floating ball [58, 59], while in other studies mice remain stationary inside of the apparatus, making licks or turning a wheel to indicate stimulus detection [33, 55, 60]. Sometimes, locomotion is required to self-initiate behavioral trials [5, 12, 56, 61], providing assurance that subjects remain motivated to continue task performance. However, locomotion has dramatic effects on both spontaneous and evoked neural activity in the visual cortex, presenting challenges for dissociating typically small, transient, spatially localized neural effects of visual spatial attention [80] from widespread locomotion or motor effects. In some cases, locomotion even hinders visual perceptual performance [62].
Nevertheless, freely moving behaviors carry ethological advantages in natural sampling of the environment, where visual, motor, and vestibular systems work together to generate neural activity and behavior shaped by evolution [63–65]. However freely moving behaviors provide greater uncertainty about stimulus position on the retina, and complicate interpretation of simultaneous neural activation when compared to head-fixed acute studies [66, 10]. High-density neural recordings during freely-moving tasks [67] present opportunities to collect ethologically relevant behavioral results during spatial attentional tasks [9]. These could then synergistically inspire more controlled experiments in head-fixed, stationary conditions.
Neural correlates of spatial attention across multiple tasks in mice
Initial insights into neural correlates of spatial attention in mice show lines of consistency with observations in primates (Table 1). First, attentional modulation engages both subcortical and cortical structures. During spatial cueing tasks in mice, for instance, optogenetic activation of the striatum facilitated behavioral responses at the contralateral cued location [8]. In another study, during blocked trial detection, primary visual cortex (V1) neural responses to the task-relevant stimulus strengthened across successive trials at a fixed location, consistent with effects of attentional allocation to this location [6]. Further, these strengthened neural responses showed lower noise correlations and fewer low frequency fluctuations. These effects were prominent in Layer 4, where activity fluctuations predict perceptual outcome [41]. All of these attentional effects were driven by a high contrast task-relevant stimulus at the receptive field location; responses to small, low contrast task-irrelevant “probe” stimuli in the receptive field were also enhanced during blocks where the task-relevant stimulus appeared at the receptive field, and when attention was presumably directed at the receptive field location by consecutive trials of stimulus detection there.
Table 1:
Behavioral and neural correlates of spatial attention
| Behavioral metric | Species | Refs |
|---|---|---|
| Increased Hit rate to attended stimuli | Mouse, Primate | [5–9, 29, 30, 36] |
| Improved Detection Speed | Mouse, Primate | [6, 7] |
| Improved orientation discrimination | Mouse, Primate | [5, 8, 9, 29, 30] |
| Increased perceptual sensitivity (d’) | Mouse, Primate | [5–9, 31] |
| Decreased psychometric thresholds | Mouse, Primate | [5, 6, 8, 29, 30] |
| Lowered response criterion (c) | Mouse, Primate | [5, 37] |
| Neural metric | Species | Refs |
| Decreased Noise Correlations | Mouse, Primate | [6, 30, 31] |
| Shift in contrast response curves | Mouse, Primate | [6, 36, 38] |
| Increased neural firing in response to attended stimuli | Mouse, Primate | [6, 30, 32, 36, 38–40] |
| Depolarization and increased membrane potential fluctuations | Mouse | [6] |
| Decreased Low Frequency Power | Mouse, Primate | [6, 30] |
Interpretational constraints for neural activity during visual spatial behaviors in mice
These several studies also reveal limitations in comparing mice and primates. For example, presenting blocks of trials with varying stimulus location probabilities can lead mice to withhold responses on low probability trials, even to high contrast stimuli [7]; in primates, selective attention generally leads to psychometric shifts for low and intermediate contrast stimuli at both the attended / unattended locations [36, 38]. This “binary” spatial selection strategy in mice provides efficient task performance, and illustrates ambiguity between selection and attention; Other tasks in mice do show psychometric performance effects for stimuli at multiple locations [5, 6, 8], but perceptual sensitivity and neural processing may not be symmetric across the mouse visual field [41, 42]. These constraints and interpretational challenges for spatial attentional measures and the underlying neural effects in mice might relate, in part, to fundamental differences between mice and primates. They also point to a need for refinement of the experimental paradigms. We next ask, what can be learnt by comparison to other studies of behavioral modulation in the mouse visual system?
Locomotion: a bridge to study circuits driving selective attentional effects?
Locomotion in head-fixed mice modulates visual neural activity across several scales
The discovery that locomotion increases the amplitude of visual cortical activity accelerated study of head-fixed visual perception in mice [43]. Locomotion enhances neural spatial selectivity [44, 45], preserves orientation selectivity [43], decorrelates population spiking [46, 47] and synaptic inputs [48–50], and enhances high frequency (30 – 80 Hz “gamma”) neural activity [43, 51]. These effects have been widely reproduced in mice and appear qualitatively similar to attentional effects on neural activity in primates. Some studies in mice even equivocate the effects of locomotion on spatial resolution with attention [45]. The development of mouse spatial behavioral tasks provides timely opportunity to test how effects of locomotion do or do not relate to spatial attention, at increasingly detailed circuit levels, as outlined next.
Locomotion engages cortical circuit motifs driving visual response modulation
Locomotion studies have identified several circuit motifs that modulate visual neural responses. One of the pathways relevant in this context starts in the brainstem, where locomotor areas that induce running also drive basal forebrain cholinergic neurons [52]. These neurons, in turn, project throughout cortex, and strongly activate cortical vasoactive intestinal peptide (VIP) inhibitory neurons; VIP neurons in turn suppress somatostatin (SST) inhibitory neurons, potently disinhibiting excitatory neuron activity [52]. These observations raise the question of whether attentional effects operate through these same cortical disinhibitory circuits [53, 54].
A second circuit that modulates visual responses during locomotion involves top-down glutamatergic input from secondary motor (M2) and anterior cingulate cortex (ACC) to V1; this circuit modulates V1 neural responses [15, 55] and carries efferent copy signals for visual flow processing during locomotion [56]. This circuit could convey attentional or spatial contextual signals to V1 and higher visual areas (HVAs), [57], although it remains unclear how specifically these projections align across retinotopic maps.
A third circuit involves higher order visual thalamus (LP nucleus; mouse analogue of the pulvinar); LP is anatomically positioned to broadly integrate cortical feedback, then broadcast this back to V1 and HVAs, providing both visual and contextual information, as seen during locomotion [58, 68]. Since the pulvinar in primates shows attentional effects [20], this suggests LP as another relevant target for investigation of attentional effects in mice. As discussed later, consideration of neural processing through each of these three circuits raises specific predictions for mechanisms of attentional modulation.
Locomotion causes widespread, multi-modal modulation of cortical activity
Studies of locomotion provide clear entry points for circuit-level interrogation of spatial attention in mice; nonetheless, open questions remain regarding the specificity of locomotion effects, and their utility as a surrogate model for selective attentional effects. Locomotion potently recruits multiple sources of subcortical neuromodulatory input [50, 69], as well as top-down (glutamatergic) motor cortical input [70]; these underlie widespread modulation of cortical activity by locomotion across sensory cortices [71–75], in thalamic axons imaged in cortex [68] and even retinal axons imaged in the superior colliculus [76]. Locomotion bouts simultaneously affect multiple sensory modalities [74, 77, 78] and multiple locations across the retinotopic map in V1 [74, 79], at odds with spatially localized selective attentional effects in primates [80]. Much less is known about locomotion effects on visual responses in primates, but studies have shown head and eye position strongly influences activity throughout the hippocampal formation [81, 82], suggesting visuospatial signals permeate the primate brain, even in structures driven by navigation in rodents. This illustrates potential challenges in using locomotion effects in rodents as a surrogate for visual attentional effects, since these also depend strongly on retinotopy, stimulus positioning, and control of motor variables, considered next.
Control of motor variables aides interpretability of spatial attention tasks
Isolating attentional effects in the primate visual system typically requires enforcing visual fixation and limiting eye movements (saccades) until the moment these become task-relevant. This control has proven indispensable for distinguishing spatial attentional from motor intentional effects [83]. In contrast to the saccadic eye movements observed in species with a fovea, mice performing visual perceptual tasks rarely make comparable rapid eye movements to visual stimuli (but see [84]), and accordingly, experimental paradigms are typically designed so that the animals report stimulus detection via licking, limb movements, or nose pokes, obviating the need for strict enforcement of eye fixation. Some head-fixed mouse behavioral tasks require locomotion and navigation in “closed loop” virtual reality [12, 61, 85], but variable locomotion speeds, optic flow, and differing timescales across trials present challenges for simple interpretation of neural responses. Even in head-fixed, stationary conditions without locomotion, spontaneous and task-related movements modulate neural activity [86–88]. One modification that may help better isolate attentional effects in mice could involve an enforced delay period (with penalties for movement) between the cue and task-relevant stimuli, or between task-relevant stimulus and response.
Limitations posed by specializations in neural substrates of spatial vision
Inherent differences in visual system organization provide important constraints for inferences about spatial attention in primates and mice. Perhaps most importantly, mice do not have a fovea. Foveal vision dictates how primates sample the visual field – by making saccadic eye movements that direct gaze. In animals lacking a fovea, sampling of the visual environment often involves head and full-body movements. It is likely that these structural differences starting at the retina result in unique downstream mechanisms for spatial modulation of visual signals; indeed, this may explain why locomotion in head-fixed rodents potently modulates visual neural activity, while eye movements modulate neural activity throughout the primate brain.
Differences in visual system organization between mice and primates extend throughout V1 [23] and also HVAs [26] that have been extensively studied during visual spatial attention in primates [1, 2, 20]. Although the roles of mouse HVAs in visual perception is an emerging topic [66, 89, 90], currently their precise roles in visual attentional behavior remain largely unknown. Despite clear retinotopy in mouse visual areas, the general lack of feature-specific laminar and columnar organization likely renders communication mechanisms across them [26, 91] different than those in the primate visual system [25].
Subcortical structures provide an appealing testbed to identify evolutionarily conserved mechanisms of visual attention. Many studies in primates have shown spatial attentional modulation of neural activity in the superior colliculus [92] and thalamus [20]; comparable studies in mice will be important, since the retina and cortex project strongly to both the superior colliculus and visual thalamus in mice [23, 24, 26], and both show sensitivity to behavioral and perceptual state [93, 94]. In primates, the roles of the striatum in cognition and action selection have been studied for many decades, and interest in the striatum’s roles in mice has been growing (e.g. [8, 95]), suggesting another entry-point for circuit-level dissection of spatial attention versus action selection in mice.
Limitations posed by differences in cognition and attentional control
Cognitive capabilities of primates allow behavioral controls that may prove difficult to implement in mice. Multiple methods and tasks allow explicit cueing of spatial attention on a trial-by-trial basis in primates. To our knowledge, trial-by-trial cueing of spatial attention in mice has not yet been shown, and might pose a major drawback for behavioral performance. As discussed, several behavioral variants have converged on a workaround by using blocked spatial cueing. Despite their spatial specificity, blocks of successive trials sacrifice temporal specificity. It remains unresolved if single-block effects could arise without invoking attention per se (e.g., altered weighting of sensory input, selection to minimize uncertainty), nor is it known if single-trial versus single-block attentional effects engage the same neural mechanisms.
Cognitive capabilities in monkeys also allow task features that permit greater control for movement. As discussed, enforcing visual fixation limits confounding effects of eye movements in primates, while in mice locomotion, navigation [12, 61, 85], and even spontaneous and task-related movements in stationary conditions [86–88] all strongly modulate neural activity, illustrating major unresolved challenges for clear interpretation of head-fixed mouse behavioral tasks.
In light of the significant differences in neuroanatomy, cognitive capabilities, and experimental control between primates and mice, it remains an open and controversial question whether attentional effects in mice provide a meaningful basis of comparison for primate studies. In our view, there is value in continuing to define the resolvability of spatial attentional effects in mice using multiple behavioral approaches, as this will help identify if general attentional mechanisms exist in mice. A comparative approach could then help discern whether species-specific specializations provide hard constraints for inferring general neural mechanisms of visual spatial attention. There is precedence for using a comparative approach in identifying species-specific neural implementations of general neural computations [4,100,102], and our hope is that several aspects of spatial attention will be similarly approachable with new techniques in mice. Further, any commonalities and differences regarding attentional effects at both behavioral and neural levels could be further tested in visual systems of other model organisms, such as the highly specialized avian visual system [96].
Identifying and evaluating the generality of attentional effects and their underlying neural substrates across species, brain areas, and behavioral tasks necessitates multiple scales of models, discussed next.
Bridging neural scales of attentional modulation through models
The neural effects of spatial attention observed in primates, and more recently in mice, provide an outstanding challenge in linking observations from disparate scales—spanning subthreshold inputs in single neurons to large-scale, multi-area population signals. An attractive approach to bridge scales exploits simple computational models [97] (Figure 2). Many of the current models of attentional modulation rely on primate studies, and they fall broadly into three categories: 1) quantitative but phenomenological models that explain attentional modulation as an extension of changes induced by stimulus properties (e.g., contrast); 2) mechanistic models, which define plausible biophysical implementations of the phenomenological models and 3) population models that explain how attention changes neuronal population activity structure. There is now ample opportunity to expand, test, and integrate these models through studies in mice performing spatial attentional tasks. Key circuit motifs involved in locomotion effects may provide a starting point to bridge across disparate scales of spatial attentional models, outlined next.
Figure 2: Models of Visual spatial attention.

A) Phenomenological Models seek to identify computational rules that govern attentional enhancement of single neurons. These models rarely specify a source of attentional modulation (measured from excitatory or inhibitory neurons), but instead explain how stimulus-response curves change as a function of attentional state (right). Example models include contrast gain, response gain, and normalization models of attention [1–4]. B) Mechanistic Models seek to explain the neural implementation of attentional modulation. By identifying the roles of specific excitatory (E) and inhibitory neuron subtypes (PV: parvalbumin; VIP: Vasoactive intestinal peptide; SST: somatostatin) in cortex, these models can address both sources of attentional modulation, and effects on local circuits. Candidate sources of attentional signals in mice include top down (HVAs: higher visual areas; M2/ACC: secondary motor / anterior cingulate cortex)[15, 55, 56], bottom up (LGN: lateral geniculate nucleus; LP: lateral posterior nucleus)[6, 68], and neuromodulatory (ACh: acetylcholine; NE: norepinephrine) inputs [69]. Open circles, excitatory; closed circles, inhibitory. Only major circuit interactions shown for clarity, see [21]. C) Neural Population Models become essential as high-density neural recording technologies proliferate. These models identify computational operations that act on large-scale neural populations that give rise to measurable population readouts of spatial attention [31, 98, 99]. Open questions include the dimensionality of spatial attentional modulation (full rank: high-dimensional individualized modulation of neurons; low rank: low-dimensional global modulation of all neurons), the relationship of this to established population readouts of spatial attention (right), and communication across brain areas, all topics to investigate in future studies in mice.
Phenomenological models of spatial attention effects in single neurons
Phenomenological models generally correlate spatial attentional state with single neuron activity modulation. These quantitative models fall into three broad varieties—contrast gain models, response gain models, and normalization models— that generally explain how single-neuron tuning curves change as a function of attention [3, 4, 100]. The contrast gain model explains increased neural sensitivity to stimuli as a subtractive shift in the contrast response curve [36]. The response gain model explains increased sensitivity as a multiplication that increases tuning curve amplitude proportionally for all stimuli [101]. The normalization model parsimoniously captures both kinds of modulation through an interaction between an attention field and suppressive normalization dependent upon stimulus size and features [4]. These influential models adequately describe how spatial attention affects single neurons, but do not explicitly specify the circuits or sources generating the normalization [102] or attentional signals.
Mechanistic models of spatial attention effects in neural networks
Mechanistic models propose neural circuit implementations for computations described by phenomenological models. Models with recurrent excitation and broadly tuned and dominant inhibition [103] recapitulate many experimentally observed neural effects of attention [104]; other mechanistic models suggest that inhibitory neurons are key players in high-gain, low-noise attentional responses [16], echoing the role of inhibitory subtypes for locomotion effects [52–54, 58]. Some mechanistic models highlight the role of neuromodulation, and illustrate for instance how cholinergic and noradrenergic inputs may enhance neural excitability [105] and reduce noise in responses to an attended stimulus [106]; other models provide a Bayesian framework for neuromodulatory effects [107]. By taking advantage of cell-type specific experimental techniques in mice, it is now possible to directly test many of the hypotheses generated by detailed mechanistic models of attention.
Population models of the effects of spatial attention on large-scale neural populations
Developments in large-scale neural recording technologies in mice allow monitoring hundreds to thousands of single neurons simultaneously [10, 14, 108]. Interpreting these large-scale population recordings requires sophisticated analytic techniques that account for the high dimensional structure of neural activity [98]. Many of these approaches were pioneered in primate motor cortex (e.g., [109]) but are rapidly gaining traction in studies of visual perception as well [99, 110, 111]. Often it is assumed that high-dimensional neural activity is constrained to a low dimensional activity space (sometimes referred to as manifold); these manifolds vary in time and define boundaries for the complete set of single neuron spikes, even during widely different behaviors [88]. Studies along these lines could help further dissect the neural correlates of spatial attention. In particular, population models of attentional modulation often hinge on the common observation of decreased correlated variability, typically measured in pairwise correlations between neurons within a brain area [6, 29, 30]; recent measurements of multi-area activity during spatial attention suggest stimulus context can either reduce or enhance correlations across brain areas [31]. Further, low rank modulation of high dimensional, inter-areal cortical activity may underlie many cognitive functions [99, 110], including attentional effects in visual cortex [32]. These effects on multi-area population structure are now testable with cellular specificity using several recording techniques in transgenic mice engaged in spatial attentional tasks.
Techniques in mice expand capabilities to probe mechanisms of attention
The mouse provides experimenters with a suite of techniques that allow interrogation of neural activity at multiple scales during single experiments [79, 112, 116]. By combining such techniques with novel behavioral paradigms, experimenters may be able to identify key neural circuit mechanisms that explain single neuron, circuit, and population scales of attentional models (Figure 3). We briefly outline a few possibilities below.
Figure 3: Methods for identifying mechanisms of spatial attention in mice.

A) By combining cell-type and projection specific techniques in mice (left) with multi-scale activity measurement methods in cortical circuits (middle and right), experimenters can elucidate potential mechanisms of spatial attention across multiple scales. Isolating contributions of candidate attentional sources using anterograde, retrograde, or intersectional virus strategies (left) allows direct measurement of their effects on cortical circuits (middle) during multiple attentional paradigms (right), and distillation of general versus task-specific neural correlates of attentional modulation. B) Pairing multi-scale activity measures with multiple attentional spatial behaviors in mice provides new avenues to test and integrate across phenomenological, mechanistic, and population models of attention. Mouse schematic in A produced with BioRender.
Whole-cell patch-clamp recordings in awake behaving mice
Whole-cell patch-clamp recordings remain the gold standard for investigation of cellular and synaptic mechanisms underlying sensory processing [17]. This technique is feasible in awake primates [113], but has seen higher throughput in recent studies in awake mice, which have revealed synaptic mechanisms underlying effects of locomotion [49, 50, 114] and arousal [62, 103] on visual responses, surround suppression [115], population desynchronization [48], and coupling between synaptic and network levels [116]. Whole-cell recordings have been recently paired with spatial attentional behavior to reveal spatially selective subthreshold depolarization in V1 pyramidal cells [6], a long-conjectured mechanism of attentional enhancements [4]. This depolarization consisted of both tonic and transient, stimulus-specific components. One hypothesis is that tonic depolarization arises from neuromodulatory signals [50], while retinotopically localized glutamatergic inputs provide stimulus-specific depolarization; this can be tested by pairing whole-cell recordings with various chemo- or optogenetic manipulations of cholinergic inputs [117] during an attentional task. A competing hypothesis is that tonic depolarization results from a balanced increase in local synaptic excitation and inhibition [118, 119]; this could be tested by measuring subthreshold excitation and inhibition [115] in combination with neuromodulatory perturbations described above. Such experiments could begin resolving how top-down, bottom-up, and neuromodulatory inputs coalesce to produce depolarization driving enhanced spiking to stimuli at the attended location.
Targeted recordings and optogenetic manipulations with projection specificity
Experimenters can now readily optically manipulate activity of specific neural subpopulations in mice and identify their roles in modulating neuronal and behavioral responses [21, 41, 90], including during spatial attention tasks [8]. For example, feedforward sources may be more relevant in tasks with explicit spatial cues, or in those showing strong attentional modulation in V1 Layer 4 [6], whereas HVAs may integrate spatial context at longer time-scales across trials, e.g. during blocks of stimulus detection at a fixed location. These hypothesized sources of attentional modulation can be tested by manipulating neuronal subpopulations of interest. One specific possibility is that projection neurons from frontal cortical areas (M2, ACC) that mediate contextual modulation of mouse V1 [15, 55, 56] also carry attentional signals. Alternatively, HVAs might integrate perceptual and decision variables [89, 90], thereby modulating V1 activity [91]. Similar experiments could test the role of target-specific V1 projections to subcortical targets during attentional modulation [18, 94]. Moreover, by selectively expressing opsins at any of these upstream structures, and manipulating the axon terminals in downstream areas (such as V1), experimenters could isolate the contribution of each potential input to local neuronal activity changes during spatial attention tasks.
Lastly, developments in multi-channel probes and wide-field imaging enable neural activity measurements from large populations across multiple regions; these techniques can be combined in awake head-fixed mice [79], and readily adapted to spatial attentional behaviors. Advances in high-density recording techniques in mice build on decades of multi-channel recording paired with sophisticated computational analysis in the primate motor [109] and visual [110, 120] systems. However, the confluence of hardware miniaturization, scalability, and computing power allows timely pairing of these techniques with cell-type specific manipulations in transgenic mice. One hypothesis to test is that inter-areal attentional modulation flows through a low dimensional subspace, likely in specific projection neurons [121]. Using anterograde, retrograde, and intersectional virus strategies in transgenic mice [122, 123], an experimenter could tag neurons that project to an HVA or frontal area of interest, or identify neurons that project back to V1, and directly measure cell-type contributions to population communication across brain regions. Similar approaches could begin to unravel cell-type contributions to several signatures of attention within local circuits, including pairwise neuronal correlations, gain modulation, and reduction of neuronal variability.
Concluding Remarks and Future Perspectives
Multiple lines of evidence suggest that mice exhibit behavioral and neural signatures of selective spatial attention, with several main effects showing commonalities with those classically observed in primates. This suggests a path towards deeper mechanistic understanding of visual spatial attention by combining novel behavioral tasks with cell-type specific experimental techniques in mice; further, by leveraging transgenic mouse models of neurological diseases, these developments pose new avenues to discover mechanisms underlying impairments of attention. However, careful consideration is needed towards defining the inherent limitations associated with studying spatial attention in mice, stemming from the specializations of their visual system and their behavioral repertoire. The approaches we have discussed point ways forward to define the utility and limits of mice for the study of attentional mechanisms, and how these do or do not dovetail with primate studies. A key tool towards resolution will involve measurements that test and refine models of attentional modulation across scales, enabling mechanistic insights from mice to inform specific predictions testable across species. Adapting behavioral approaches pioneered in primates for mice has proven remarkably fruitful for detailed insights into sensorimotor integration, working memory, and decision making. It is hoped that emerging approaches for the study of spatial attention in mice can provide mechanistic insights into this domain as well, across multiple scales of analysis.
Outstanding Questions.
Multiple tasks for directing visual spatial attention in mice lead to behavioral improvements—do these tasks recruit overlapping or distinct brain areas?
Can individual mice perform multiple tasks and reveal general principles for behavioral improvements due to spatial attention?
Are feedforward and feedback circuits underlying attentional modulation in mice retinotopically specific?
In primates, attentional modulation appears strongest in higher visual cortical areas— is this also the case in mice?
How are spatial attentional modulations communicated across multiple visual areas in mice?
Spatial attention in primates strongly involves the oculomotor system— are analogous cortical and subcortical structures similarly recruited in mice?
Sensory and non-sensory activity in cortical excitatory neurons depends upon their downstream targets—does this shape their modulation by spatial attention?
Cortical inhibitory neurons precisely gate excitatory neurons during locomotion in mice—do these same circuits gate spatial attentional modulations?
Locomotion strongly recruits neuromodulatory systems—are these also recruited by spatial attention, independent of locomotion or movement?
Can circuit motifs identified in mice help define general mechanisms of attentional modulation, despite differences in anatomical, functional, and molecular properties of neurons across brain areas and species?
Does locomotion enhance visual processing in primates? What can this reveal about general versus species-specific mechanisms that modulate visual processing?
Spatial attentional effects in mice elicit both baseline and stimulus-evoked depolarization of V1 neurons. Do these two components result from distinct mechanisms?
How can models of brain area communication be tested using large-scale neural recordings with cell-type specific manipulations in behaving mice?
Can insights gained from mouse studies be used to refine established models of spatial attentional modulation?
Highlights.
Several tasks now enable study of visual spatial attentional effects in mice
These tasks manipulate visual cues, stimulus location probabilities, or stimulus location across consecutive trial blocks, with distinct advantages and limitations for resolving attentional effects
Behavioral and neural correlates of spatial attention during these tasks show both consistency and discrepancies with primate work
Visual system organization, behavioral capabilities, and motor variables all provide important caveats for comparisons of mouse and primate studies
Probing circuits and cell-types underlying spatial attention in mice can follow a roadmap provided by detailed studies of locomotion effects on vision
Large-scale recordings, cell-type specific perturbations, and patch clamp recordings in transgenic mice provide new ways to test models of spatial attention across multiple scales
Acknowledgements
We thank members of the Haider Lab and the anonymous reviewers for constructive comments. We apologize to colleagues whose work we were unable to cite due to space limits. B.H. was funded by the Whitehall Foundation, Sloan Foundation, Simons Foundation (SFARI 600343), NIH NINDS (NS107968), and NIH BRAIN Initiative (NS109978).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Declaration of Interests
The authors declare no competing interests.
References
- 1.Reynolds JH and Chelazzi L, Attentional modulation of visual processing. Annu Rev Neurosci, 2004. 27: p. 611–47. [DOI] [PubMed] [Google Scholar]
- 2.Maunsell JHR, Neuronal Mechanisms of Visual Attention. Annu Rev Vis Sci, 2015. 1: p. 373–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lee J and Maunsell JH, A normalization model of attentional modulation of single unit responses. PLoS One, 2009. 4(2): p. e4651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Reynolds JH and Heeger DJ, The Normalization Model of Attention. Neuron, 2009. 61: p. 168–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang L and Krauzlis RJ, Visual Selective Attention in Mice. Current Biology, 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Speed A, et al. , Spatial attention enhances network, cellular and subthreshold responses in mouse visual cortex. Nature Communications, 2020. 11(1): p. 505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.McBride EG, Lee S-YJ, and Callaway EM, Local and Global Influences of Visual Spatial Selection and Locomotion in Mouse Primary Visual Cortex. Current Biology, 2019. 29(10): p. 1592–1605.e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wang L and Krauzlis RJ, Involvement of Striatal Direct Pathway in Visual Spatial Attention in Mice. Current Biology, 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.You WK and Mysore SP, Endogenous and exogenous control of visuospatial selective attention in freely behaving mice. Nature Communications, 2020. 11(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Steinmetz NA, et al. , Challenges and opportunities for large-scale electrophysiology with Neuropixels probes. Curr Opin Neurobiol, 2018. 50: p. 92–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Voigts J and Harnett MT, Somatic and Dendritic Encoding of Spatial Variables in Retrosplenial Cortex Differs during 2D Navigation. Neuron, 2020. 105(2): p. 237–245 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Saleem AB, et al. , Coherent encoding of subjective spatial position in visual cortex and hippocampus. Nature, 2018. 562(7725): p. 124–127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chettih SN and Harvey CD, Single-neuron perturbations reveal feature-specific competition in V1. Nature, 2019. 567(7748): p. 334–340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rumyantsev OI, et al. , Fundamental bounds on the fidelity of sensory cortical coding. Nature, 2020. 580(7801): p. 100–105. [DOI] [PubMed] [Google Scholar]
- 15.Zhang S, et al. , Selective attention. Long-range and local circuits for top-down modulation of visual cortex processing. Science, 2014. 345(6197): p. 660–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kanashiro T, et al. , Attentional modulation of neuronal variability in circuit models of cortex. Elife, 2017. 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Petersen CCH, Whole-Cell Recording of Neuronal Membrane Potential during Behavior. Neuron, 2017. 95(6): p. 1266–1281. [DOI] [PubMed] [Google Scholar]
- 18.Zingg B, et al. , AAV-Mediated Anterograde Transsynaptic Tagging: Mapping Corticocollicular Input-Defined Neural Pathways for Defense Behaviors. Neuron, 2017. 93(1): p. 33–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wimmer RD, et al. , Thalamic control of sensory selection in divided attention. Nature, 2015. 526(7575): p. 705–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Buschman TJ and Kastner S, From Behavior to Neural Dynamics: An Integrated Theory of Attention. Neuron, 2015. 88(1): p. 127–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ferguson KA and Cardin JA, Mechanisms underlying gain modulation in the cortex. Nat Rev Neurosci, 2020. 21(2): p. 80–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Harris KD and Shepherd GM, The neocortical circuit: themes and variations. Nat Neurosci, 2015. 18(2): p. 170–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Niell CM, Cell types, circuits, and receptive fields in the mouse visual cortex. Annu Rev Neurosci, 2015. 38: p. 413–31. [DOI] [PubMed] [Google Scholar]
- 24.Seabrook TA, et al. , Architecture, Function, and Assembly of the Mouse Visual System. Annu Rev Neurosci, 2017. 40: p. 499–538. [DOI] [PubMed] [Google Scholar]
- 25.Nassi JJ and Callaway EM, Parallel processing strategies of the primate visual system. Nat Rev Neurosci, 2009. 10(5): p. 360–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Froudarakis E, et al. , The Visual Cortex in Context. Annual Review of Vision Science, 2019. 5(1): p. 317–339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Posner MI, Snyder CRR, and Davidson BJ. Attention and the Detection of Signals. in Journal of Experimental Psychology: General. 1980. [PubMed] [Google Scholar]
- 28.Funayama K, et al. , Neocortical Rebound Depolarization Enhances Visual Perception. PLoS Biol, 2015. 13(8): p. e1002231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mitchell JF, Sundberg KA, and Reynolds JH, Spatial attention decorrelates intrinsic activity fluctuations in macaque area V4. Neuron, 2009. 63(6): p. 879–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cohen MR and Maunsell JH, Attention improves performance primarily by reducing interneuronal correlations. Nat Neurosci, 2009. 12(12): p. 1594–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ruff DA and Cohen MR, Simultaneous multi-area recordings suggest that attention improves performance by reshaping stimulus representations. Nat Neurosci, 2019. 22(10): p. 1669–1676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ruff DA, et al. , Low rank mechanisms underlying flexible visual representations. Proc Natl Acad Sci U S A, 2020. 117(47): p. 29321–29329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Carandini M and Churchland AK, Probing perceptual decisions in rodents. Nat Neurosci, 2013. 16(7): p. 824–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Stüttgen M, Schwarz C, and Jäkel F, Mapping Spikes to Sensations. Frontiers in Neuroscience, 2011. 5(125). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Everling S, et al. , Filtering of neural signals by focused attention in the monkey prefrontal cortex. Nat Neurosci, 2002. 5(7): p. 671–6. [DOI] [PubMed] [Google Scholar]
- 36.Reynolds JH, Pasternak T, and Desimone R, Attention increases sensitivity of V4 neurons. Neuron, 2000. [DOI] [PubMed] [Google Scholar]
- 37.Luo TZ and Maunsell JHR, Attention can be subdivided into neurobiological components corresponding to distinct behavioral effects. Proceedings of the National Academy of Sciences, 2019. 116(52): p. 26187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sundberg KA, Mitchell JF, and Reynolds JH, Spatial attention modulates center-surround interactions in macaque visual area v4. Neuron, 2009. 61(6): p. 952–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Moran J and Desimone R, Selective attention gates visual processing in the extrastriate cortex. Science, 1985. 229(4715): p. 782–4. [DOI] [PubMed] [Google Scholar]
- 40.Reynolds JH, Chelazzi L, and Desimone R, Competitive mechanisms subserve attention in macaque areas V2 and V4. J Neurosci, 1999. 19(5): p. 1736–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Speed A, et al. , Cortical State Fluctuations across Layers of V1 during Visual Spatial Perception. Cell Rep, 2019. 26(11): p. 2868–2874 e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Williams B, et al. , Spatial modulation of dark versus bright stimulus responses in mouse visual cortex. Curr Biol, 2021311–8, DOI: 10.1016/j.cub.2021.06.094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Niell CM and Stryker MP, Modulation of Visual Responses by Behavioral State in Mouse Visual Cortex. Neuron, 2010. 65: p. 472–479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ayaz A, et al. , Locomotion controls spatial integration in mouse visual cortex. Curr Biol, 2013. 23(10): p. 890–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mineault PJ, et al. , Enhanced Spatial Resolution During Locomotion and Heightened Attention in Mouse Primary Visual Cortex. J Neurosci, 2016. 36(24): p. 6382–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Dadarlat MC and Stryker MP, Locomotion Enhances Neural Encoding of Visual Stimuli in Mouse V1. J Neurosci, 2017. 37(14): p. 3764–3775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Erisken S, et al. , Effects of locomotion extend throughout the mouse early visual system. Curr Biol, 2014. 24(24): p. 2899–907. [DOI] [PubMed] [Google Scholar]
- 48.Arroyo S, Bennett C, and Hestrin S, Correlation of Synaptic Inputs in the Visual Cortex of Awake, Behaving Mice . Neuron, 2018. 99(6): p. 1289–1301 e2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bennett C, Arroyo S, and Hestrin S, Subthreshold mechanisms underlying state-dependent modulation of visual responses. Neuron, 2013. 80(2): p. 350–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Polack PO, Friedman J, and Golshani P, Cellular mechanisms of brain state-dependent gain modulation in visual cortex. Nat Neurosci, 2013. 16(9): p. 1331–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Saleem AB, et al. , Subcortical Source and Modulation of the Narrowband Gamma Oscillation in Mouse Visual Cortex. Neuron, 2017. 93(2): p. 315–322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Fu Y, et al. , A cortical circuit for gain control by behavioral state. Cell, 2014. 156(6): p. 1139–1152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Dipoppa M, et al. , Vision and Locomotion Shape the Interactions between Neuron Types in Mouse Visual Cortex. Neuron, 2018. 98(3): p. 602–615 e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pakan JM, et al. , Behavioral-state modulation of inhibition is context-dependent and cell type specific in mouse visual cortex. Elife, 2016. 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Huda R, et al. , Distinct prefrontal top-down circuits differentially modulate sensorimotor behavior. Nat Commun, 2020. 11(1): p. 6007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Leinweber M, et al. , A Sensorimotor Circuit in Mouse Cortex for Visual Flow Predictions. Neuron, 2017. 95(6): p. 1420–1432 e5. [DOI] [PubMed] [Google Scholar]
- 57.Saleem AB, Two stream hypothesis of visual processing for navigation in mouse. Curr Opin Neurobiol, 2020. 64: p. 70–78. [DOI] [PubMed] [Google Scholar]
- 58.Khan AG and Hofer SB, Contextual signals in visual cortex. Curr Opin Neurobiol, 2018. 52: p. 131–138. [DOI] [PubMed] [Google Scholar]
- 59.Pakan JM, Francioni V, and Rochefort NL, Action and learning shape the activity of neuronal circuits in the visual cortex. Curr Opin Neurobiol, 2018. 52: p. 88–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Burgess CP, et al. , High-Yield Methods for Accurate Two-Alternative Visual Psychophysics in Head-Fixed Mice. Cell Rep, 2017. 20(10): p. 2513–2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Poort J, et al. , Learning Enhances Sensory and Multiple Non-sensory Representations in Primary Visual Cortex. Neuron, 2015. 86(6): p. 1478–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Neske GT, et al. , Distinct Waking States for Strong Evoked Responses in Primary Visual Cortex and Optimal Visual Detection Performance. J Neurosci, 2019. 39(50): p. 10044–10059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hoy JL, et al. , Vision Drives Accurate Approach Behavior during Prey Capture in Laboratory Mice. Curr Biol, 2016. 26(22): p. 3046–3052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hoy JL, Bishop HI, and Niell CM, Defined Cell Types in Superior Colliculus Make Distinct Contributions to Prey Capture Behavior in the Mouse. Curr Biol, 2019. 29(23): p. 4130–4138 e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.De Franceschi G, et al. , Vision Guides Selection of Freeze or Flight Defense Strategies in Mice. Curr Biol, 2016. 26(16): p. 2150–4. [DOI] [PubMed] [Google Scholar]
- 66.Siegle JH, et al. , Survey of spiking in the mouse visual system reveals functional hierarchy. Nature, 2021. 592(7852): p. 86–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Juavinett AL, Bekheet G, and Churchland AK, Chronically implanted Neuropixels probes enable high-yield recordings in freely moving mice. Elife, 2019. 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Roth MM, et al. , Thalamic nuclei convey diverse contextual information to layer 1 of visual cortex. Nat Neurosci, 2016. 19(2): p. 299–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Reimer J, et al. , Pupil fluctuations track rapid changes in adrenergic and cholinergic activity in cortex. Nat Commun, 2016. 7: p. 13289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zagha E and McCormick DA, Neural control of brain state. Curr Opin Neurobiol, 2014. 29: p. 178–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ayaz A, et al. , Layer-specific integration of locomotion and sensory information in mouse barrel cortex. Nat Commun, 2019. 10(1): p. 2585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.McGinley MJ, et al. , Waking State: Rapid Variations Modulate Neural and Behavioral Responses. Neuron, 2015. 87(6): p. 1143–1161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Schneider DM, Nelson A, and Mooney R, A synaptic and circuit basis for corollary discharge in the auditory cortex. Nature, 2014. 513(7517): p. 189–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Shimaoka D, Harris KD, and Carandini M, Effects of Arousal on Mouse Sensory Cortex Depend on Modality. Cell Rep, 2018. 22(12): p. 3160–3167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Diamanti EM, et al. , Spatial modulation of visual responses arises in cortex with active navigation. Elife, 2021. 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Schroder S, et al. , Arousal Modulates Retinal Output. Neuron, 2020. 107(3): p. 487–495 e9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Busse L, et al. , Sensation during Active Behaviors. The Journal of Neuroscience, 2017. 37(45): p. 10826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Händel BF and Schölvinck ML, The brain during free movement - What can we learn from the animal model. Brain research, 2019. 1716: p. 3–15. [DOI] [PubMed] [Google Scholar]
- 79.Clancy KB, Orsolic I, and Mrsic-Flogel TD, Locomotion-dependent remapping of distributed cortical networks. Nat Neurosci, 2019. 22(5): p. 778–786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Engel TA, et al. , Selective modulation of cortical state during spatial attention. Science, 2016. 354(6316): p. 1140. [DOI] [PubMed] [Google Scholar]
- 81.Mao D, et al. , Spatial modulation of hippocampal activity in freely moving macaques. bioRxiv, 10.1101/2020.10.03.324848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Killian NJ, Jutras MJ, and Buffalo EA, A map of visual space in the primate entorhinal cortex. Nature, 2012. 491(7426): p. 761–764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Jonikaitis D and Moore T, The interdependence of attention, working memory and gaze control: behavior and neural circuitry. Curr Opin Psychol, 2019. 29: p. 126–134. [DOI] [PubMed] [Google Scholar]
- 84.Itokazu T, et al. , Streamlined sensory motor communication through cortical reciprocal connectivity in a visually guided eye movement task. Nat Commun, 2018. 9(1): p. 338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Leinweber M, et al. , A Sensorimotor Circuit in Mouse Cortex for Visual Flow Predictions. Neuron, 2017. 96(5): p. 1204. [DOI] [PubMed] [Google Scholar]
- 86.Musall S, et al. , Single-trial neural dynamics are dominated by richly varied movements. Nat Neurosci, 2019. 22(10): p. 1677–1686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Salkoff DB, et al. , Movement and Performance Explain Widespread Cortical Activity in a Visual Detection Task. Cereb Cortex, 2020. 30(1): p. 421–437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Stringer C, et al. , Spontaneous behaviors drive multidimensional, brainwide activity. Science, 2019. 364(6437): p. 255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Jin M and Glickfeld LL, Mouse Higher Visual Areas Provide Both Distributed and Specialized Contributions to Visually Guided Behaviors. Curr Biol, 2020. 30(23): p. 4682–4692 e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Goldbach HC, et al. , Performance in even a simple perceptual task depends on mouse secondary visual areas. Elife, 2021. 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.D’Souza RD and Burkhalter A, A Laminar Organization for Selective Cortico-Cortical Communication. Frontiers in Neuroanatomy, 2017. 11(71). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Krauzlis RJ, Lovejoy LP, and Zenon A, Superior colliculus and visual spatial attention. Annu Rev Neurosci, 2013. 36: p. 165–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Nestvogel DB and McCormick DA, Visual Thalamocortical Mechanisms of Waking State Dependent Activity and Alpha Oscillations. 10.1101/2021.04.14.439865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ruediger S and Scanziani M, Learning speed and detection sensitivity controlled by distinct cortico-fugal neurons in visual cortex. Elife, 2020. 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Peters AJ, et al. , Striatal activity topographically reflects cortical activity. Nature, 2021. 591(7850): p. 420–425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Knudsen EI, Neural Circuits That Mediate Selective Attention: A Comparative Perspective. Trends Neurosci, 2018. 41(11): p. 789–805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Carandini M, From circuits to behavior: a bridge too far? Nat Neurosci, 2012. 15(4): p. 507–9. [DOI] [PubMed] [Google Scholar]
- 98.Cunningham JP and Yu BM, Dimensionality reduction for large-scale neural recordings. Nat Neurosci, 2014. 17(11): p. 1500–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Kohn A, et al. , Principles of Corticocortical Communication: Proposed Schemes and Design Considerations. Trends Neurosci, 2020. 43(9): p. 725–737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Carandini M and Heeger DJ, Normalization as a canonical neural computation. Nat Rev Neurosci, 2011. 13(1): p. 51–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.McAdams CJ and Maunsell JH, Effects of attention on orientation-tuning functions of single neurons in macaque cortical area V4. J Neurosci, 1999. 19(1): p. 431–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Sato TK, et al. , An excitatory basis for divisive normalization in visual cortex. Nat Neurosci, 2016. 19(4): p. 568–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Haider B, Hausser M, and Carandini M, Inhibition dominates sensory responses in the awake cortex. Nature, 2013. 493(7430): p. 97–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Ardid S, Wang XJ, and Compte A, An integrated microcircuit model of attentional processing in the neocortex. J Neurosci, 2007. 27(32): p. 8486–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Herrero JL, et al. , Acetylcholine contributes through muscarinic receptors to attentional modulation in V1. Nature, 2008. 454(7208): p. 1110–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Deco G and Thiele A, Cholinergic control of cortical network interactions enables feedback-mediated attentional modulation. Eur J Neurosci, 2011. 34(1): p. 146–57. [DOI] [PubMed] [Google Scholar]
- 107.Yu AJ and Dayan P, Uncertainty, neuromodulation, and attention. Neuron, 2005. 46(4): p. 681–92. [DOI] [PubMed] [Google Scholar]
- 108.Steinmetz NA, et al. , Neuropixels 2.0: A miniaturized high-density probe for stable, long-term brain recordings. Science, 2021. 372(6539). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kaufman MT, et al. , Cortical activity in the null space: permitting preparation without movement. Nat Neurosci, 2014. 17(3): p. 440–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Semedo JD, et al. , Cortical Areas Interact through a Communication Subspace. Neuron, 2019. 102(1): p. 249–259 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Steinmetz NA, et al. , Distributed coding of choice, action and engagement across the mouse brain. Nature, 2019. 576(7786): p. 266–273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Cardin JA, Crair MC, and Higley MJ, Mesoscopic Imaging: Shining a Wide Light on Large-Scale Neural Dynamics. Neuron, 2020. 108(1): p. 33–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Li B, et al. , Voltage-Gated Intrinsic Conductances Shape the Input-Output Relationship of Cortical Neurons in Behaving Primate V1. Neuron, 2020. 107(1): p. 185–196 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Einstein MC, et al. , Visually Evoked 3–5 Hz Membrane Potential Oscillations Reduce the Responsiveness of Visual Cortex Neurons in Awake Behaving Mice. J Neurosci, 2017. 37(20): p. 5084–5098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Adesnik H, Synaptic Mechanisms of Feature Coding in the Visual Cortex of Awake Mice. Neuron, 2017. 95(5): p. 1147–1159 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Haider B, et al. , Millisecond Coupling of Local Field Potentials to Synaptic Currents in the Awake Visual Cortex. Neuron, 2016. 90(1): p. 35–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Gasselin C, et al. , Cell-type-specific nicotinic input disinhibits mouse barrel cortex during active sensing. Neuron, 2021. 109(5): p. 778–787 e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Haider B and McCormick DA, Rapid neocortical dynamics: cellular and network mechanisms. Neuron, 2009. 62(2): p. 171–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Silver RA, Neuronal arithmetic. Nat Rev Neurosci, 2010. 11(7): p. 474–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Davis ZW, et al. , Spontaneous travelling cortical waves gate perception in behaving primates. Nature, 2020. 587(7834): p. 432–436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Lur G, et al. , Projection-Specific Visual Feature Encoding by Layer 5 Cortical Subnetworks . Cell Rep, 2016. 14(11): p. 2538–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Yetman MJ, et al. , Intersectional monosynaptic tracing for dissecting subtype-specific organization of GABAergic interneuron inputs. Nat Neurosci, 2019. 22(3): p. 492–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Luo L, Callaway EM, and Svoboda K, Genetic dissection of neural circuits. Neuron, 2008. 57(5): p. 634–60. [DOI] [PMC free article] [PubMed] [Google Scholar]