

Abstract
Objectives:
Agentic (status/independence) and communal (acceptance/connectedness) social goals are thought to shape how adolescents transact with their social environments. Despite their theoretical importance, little work has focused on the development of these higher order personality dimensions. Informed by developmental neuroscience and evolutionary psychology theoretical frameworks, the current study examined associations between pubertal status, a person’s level of pubertal development at a single point in time, and agentic and communal social goals across early to middle adolescence.
Methods:
This longitudinal study consisted of 387 (55% female) adolescents (Wave 1 M age = 12.1) who were assessed annually across three waves. Hierarchical linear modeling was used to examine growth in pubertal status and agentic and communal goals and to distinguish between- and within-person associations between pubertal status and social goals.
Results:
Within-person pubertal status was concurrently associated with higher levels of agentic and communal goals. In the cross-sectional and longitudinal models, between-person pubertal status was associated with higher levels of agentic social goals. No support was found for social goals prospectively predicting pubertal status.
Conclusions:
These findings provide support for the hypothesis that puberty, in part, may drive developmental shifts in the value adolescents place on close peer relationships and obtaining status and independence.
Keywords: agency, communion, puberty, social goals, within- and between-person effects
1 |. INTRODUCTION
Contemporary Integrative Interpersonal Theory of Personality (CIIT) is a metatheoretical framework that contends that interpersonal interactions are the building block of personality and psychopathology (Pincus, 2005). Central to CIIT is the interpersonal circumplex, which provides a structure for organizing the higher order personality dimensions of agency and communion across interpersonal functioning domains (e.g., interpersonal behaviors, goals, and problems) and multiple time frames (e.g., seconds, days, and years) (Dawood et al., 2018). Locke (2003) developed the Circumplex Scales of Interpersonal Values (CSIV) to assess individual differences in interpersonal outcomes preferences. Consistent with CIIT, the CSIV is organized around the dimensions of agency (standing out, getting ahead) and communion (getting along, fitting in). Extensive research with adults has demonstrated relationships between agency and communion with interpersonal functioning and psychopathology (for a review see Locke, 2015).
Motivated by the developmental significance of social relationships during adolescence, the CSIV was adapted for children and adolescents (e.g., the Interpersonal Goals Inventory for Children, IGI-C, Ojanen et al., 2005; the revised IGI-C, IGI-CR, Trucco et al., 2013) to assess adolescents’ values in interpersonal situations. These values are referred to as social goals and they are thought to shape how adolescents transact with their social environments (Trucco et al., 2013). Similar to the CSIV, agency refers to valuing individuality, and it is manifested in striving for power and status. Communion refers to valuing being part of and gaining acceptance from social relationships and is manifested in strivings for intimacy and connectedness with peers (Trucco et al., 2013). Both agentic and communal social goals have been associated with indicators of adjustment in childhood and adolescence, including peer acceptance (Ojanen & Findley-Van Nostrand, 2014), prosocial behavior (Ojanen et al., 2005), school connectedness (Meisel & Colder, 2017), aggression (Mayeux & Kraft, 2018), and substance use (Meisel & Colder, 2015).
Across multiple conceptualizations, including agency and communion (Ojanen et al., 2005; Trucco et al., 2013), social dominance goals (e.g., Kiefer & Ryan, 2008), autonomy and relatedness (Allen & Loeb, 2015), and approach and avoidance goals (e.g., Dickson & MacLeod, 2004), the development of social goals during adolescence is viewed as important for positive adjustment (Allen & Loeb, 2015; Baumrind, 1991). Social developmental researchers have long posited that adolescents need to develop both agency (in the form of autonomy and the ability to resist peer influence) and communion (in the form of forming close peer ties) to successfully navigate the challenges of adolescence (Allen & Loeb, 2015; Baumrind, 1991). In line with this argument, agentic and communal goals increase from early (ages 10–14) to middle adolescence (ages 14–18) (Trucco et al., 2014). Early adolescents who value both autonomy and relatedness have been found to receive the highest parent reported ratings on social, career, and emotional adjustment in early adulthood (Allen et al., 2014). Despite theory and evidence highlighting the importance of agency and communion during adolescence, surprisingly little attention has been given to factors that may contribute to the development of agentic and communal goals during early and middle adolescence. This is the primary aim of the present study.
1.1 |. Puberty and agentic and communal social goals
Puberty typically begins during late childhood and, on average, continues through middle adolescence for males and females (Walvoord, 2010). Developmental neuroscience research has suggested that physiological changes resulting from puberty may lead to increases in valuing status and dominance as well as peer acceptance (Crone & Dahl, 2012). Adolescence is a period of rapid brain development (Blakemore et al., 2010) and structural and functional brain regions implicated in socio-affective processing (e.g., prefrontal cortex, striatum) undergo particularly rapid development and restructuring (Vijayakumar et al., 2018). Hormonal changes at the onset of puberty are thought to contribute to these structural and functional changes that lead to a reorientation of social motivation (Blakemore et al., 2010; Crone & Dahl, 2012). Of note, two motivational tendencies suggested to increase are (1) stronger interest in and pursuit of peer relationships and (2) increased valuing of social status (Crone & Dahl, 2012), which align with communal and agentic social goals, respectively.
Evolutionary psychology provides a theoretical account of the development of agentic and communal social goals during adolescence (Anderson et al., 2015; Chan et al., 2018). One evolutionary model of adolescent risk taking (Ellis et al., 2012) posits that the central challenge of adolescence is to compete for a finite resource, sexual reproduction. According to this perspective, sexual reproduction is contingent on being able to form close relationships with potential mating partners as well as establish status and dominance to attract mating partners and ward off competition. It is believed that natural selection has favored physical changes, namely, puberty, to prepare adolescents to compete for this finite resource. Specifically, activation of the reproductive axis at puberty brings about increased production of androgens and estrogens, which are thought to increase the valuing of social status and dominance (agency) and group acceptance (communion) to facilitate the sociocompetitive competencies necessary for sexual reproduction (Ellis et al., 2012).
Despite developmental neuroscience and evolutionary psychology both positing that changes in pubertal status promotes the development of agentic and communal social goals, no prior work, to our knowledge, has examined the impact of pubertal status on the development of agentic and communal goals. To date, prior work had examined associations between pubertal status and interpersonal behaviors during adolescence. Steinberg (1988) found that pubertal status was prospectively associated with increased autonomy and distance in the parent-child relationship in an adolescent sample of 11 to 16-year-olds. Pubertal status and timing, both assessed using the Pubertal Development Scale (PDS; Petersen et al., 1988), have also been associated with engagement in pseudomature behaviors, such as delinquent behaviors and substance use, (Castellanos-Ryan et al., 2013), which is thought to meet adolescents’ need to appear independent from their parents and gain status with peers (Allen et al., 2014). Studies of adolescent neural responses to social interactions have found advanced pubertal status to be associated with increased sensitivity to peer rejection and emotional faces (for a review see Guyer et al., 2016). For example, Silk et al. (2013) found that advanced pubertal status was associated with increased neural reactivity to peer rejection in a chatroom interaction task. Pubertal status measured by the PDS has also been found to prospectively predict increases in friendship quality even after controlling for age and prior friendship quality (Prinstein et al., 2005). In sum, research on puberty and interpersonal functioning is consistent with the idea that pubertal status may be associated with the development of agentic and communal social goals, yet these associations have not yet been tested.
1.2 |. Agentic and communal social goals as predictors of pubertal status
Studies have identified a number of social context characteristics that impact pubertal status such as childhood rearing placement (institution versus. foster care; Johnson et al., 2018), parent-child relationship quality (Ellis et al., 2011), parental warmth (Ellis et al., 1999), family conflict (Moffitt et al., 1992), and parental divorce (Belsky et al., 2007). The peer context is also thought to be important, although it has garnered less attention in human studies (Dorn et al., 2019). Peer contextual factors have been found to impact pubertal development in nonhuman primate species (Stephens & Wallen, 2013). Across a number of nonhuman primate species, higher social rank is associated with earlier pubertal onset and emergence of secondary sex characteristics in both males and females (Alberts & Altmann, 1995; Bercovitch & Strum, 1993; Charpentier et al., 2008; Zehr et al., 2005). Nonhuman primates with greater opportunities to affiliate with members of the opposite-sex demonstrate earlier pubertal maturation (Stephens & Wallen, 2013). Greater affiliative behaviors toward peers also is associated with earlier first ovulation (Wilson et al., 2013). Together, these animal studies suggest that social status among peers may be evolutionary conserved mechanisms that regulate pubertal status (Stephens & Wallen, 2013).
This animal work is relevant in the current study because in humans, agentic, and communal social goals are associated with social status (Caravita & Cillessen, 2012; Ojanen & Findley-Van Nostrand, 2014). Specifically, agentic social goals are positively associated with perceived popularity, who adolescents view as the most popular peers in their classroom, whereas communal goals are positively associated with sociometric popularity, who adolescents like the most in their classroom (Caravita & Cillessen, 2012). The implication is that agentic and communal goals may impact social status, and in turn, pubertal status. Hence, bi-directional effects between puberty and social goals were investigated in the present study.
1.3 |. Separating between- and within- person effects
Both puberty and social goals change during early and middle adolescence (Crone & Dahl, 2012; Trucco et al., 2014). As suggested by evolutionary theory as well as neuroscience theories of adolescent development, an adolescent’s overall levels of pubertal status during early to middle adolescence should be associated with agentic and communal goals (between-person effect) (Crone & Dahl, 2012; Ellis et al., 2012; Steinberg, 1988). However, solely examining these between-person associations may not fully capture the dynamic nature of reciprocal associations between puberty and social goals (Susman et al., 2019; Wright et al., 2015).
Puberty is considered a reactive process such that the rate of progression is influenced by a variety of environmental conditions that can change across time, for example, stress-reactivity and socio-contextual factors (Belsky et al., 2007; Ellis et al., 2011). This has led to calls from puberty researchers to parse apart pubertal status levels (between-person effects) and the degree of change in puberty at the individual-level during early and middle adolescence (Susman et al., 2019). Agentic and communal social goals also ebb and flow across time as functions of the peers an adolescent interacts with, social context, topics of conversation with parents, and levels of stress an individual is experiencing in their life (Wright et al., 2015). Considering the dynamic nature of social goals and puberty during early and middle adolescence, capturing within-person associations in addition to between-person associations may be important to understand the associations between puberty and social goals. For example, according to both evolutionary psychology and developmental neuroscience perspectives, times when an adolescent is experiencing particularly rapid changes in puberty should be associated with increases in their valuing of communion and agency. Similarly, periods when an adolescent’s social goals are particularly salient, such as entering middle school or high school, or gaining entry into a high-status peer group, may be associated with subsequent increases in pubertal status.
1.4 |. Possible sex differences
The literature on adolescent social goals has consistently found females to have higher levels of communion relative to males (Mayeux & Kraft, 2018; Ojanen et al., 2005; Trucco et al., 2013). In contrast, the evidence regarding sex differences for agentic goals is mixed with some studies finding higher levels in males (Trucco et al., 2013) and other demonstrating no differences across males and females (Mayeux & Kraft, 2018). From an evolutionary biology perspective, adolescent males are proposed to have higher agency and females higher communion due to different demands to achieve reproductive success. Males engage in behaviors that establish social dominance and status over competing males in order to successfully access, attract, and retain females (Buss & Schmitt, 1993). Thus, it has been argued that evolution favors the development of agentic behaviors in males during puberty to facilitate reproductive success (Ellis et al., 2011). Relative to males, females experience greater as well as less variable reproductive success (Hammer et al., 2008). An evolutionary psychological perspective proposes that the lower need for status and dominance between females may have favored other processes that facilitate reproductive success such as forming warm relationships with potential suitors and fellow females to assist with the demands of having and raising children (e.g., have access to necessary external resources) (Buss & Schmidt, 1993).
1.5 |. Current study
Based on theoretical and empirical findings suggesting bi-directional associations between pubertal status and social goals, the current study tested the following hypotheses: (1) Agentic social goals, communal social goals, and pubertal status would demonstrate significant growth from early to middle adolescence. (2) Overall levels of pubertal status (between-person effect) as well as increases from an adolescent’s typical level of pubertal status (within-person effect) would be cross-sectionally associated with higher levels of agentic and communal goals. (3) Overall levels of pubertal status (between-person effect) as well as times when an adolescent’s pubertal status increased relative to their average level of pubertal status (within-person effect) would be prospectively associated with higher levels of agentic and communal goals. (4) Overall levels of agentic and communal goals (between-person effects) as well as increases in agentic and communal goals across waves (within-person effects) would be prospectively associated with higher levels of pubertal status. (5) At both the between- and within-person levels of analysis, associations between agency and puberty would be stronger for males and associations between communion and puberty would be stronger for females.
2 |. METHODS
2.1 |. Participants
Participants were drawn from a longitudinal study examining risk and protective factors associated with adolescent substance use. The sample included 387 families (1 caregiver and 1 child) from Erie County in western New York and was recruited from 2007 to 2009 using random-digit dialing (RDD) procedures. RDD helps to provide a representative sample through using both listed and unlisted telephone numbers. The county from which the sample was recruited is especially well suited for RDD because 98.5% of households have a landline. The sample was evenly split on sex (N = 213 females, 55%) and included non-Hispanic Caucasian (83.1%), African American (9.1%), Hispanic (2.1%), and Asian (1.0), as well as youth of mixed ethnicity (4.7%). Median family income was $70,000 (range = $1,500–$500,000), and 6.2% of the families received public income assistance. The demographic characteristics of our community sample are similar to those in Erie County from whence the sample came (see Trucco et al., 2014).
Data were collected annually, and the current study uses data from Waves 1 through 3 (spanning 2007 to 2011) because these assessments included the variables of interest and allowed us to test prospective associations. The sample at Wave (W) 1 (M age = 12.1, Range = 11–13), W2 (M age = 13.1, Range = 12–14), and W3 (M = 14.1, Range = 13–16), included 387, 373, and 370 adolescents, respectively.
Overall attrition across W1 through W3 was small (4.4%). Chi-square and analysis of variance tests indicated no significant differences (ps > .05) between targets who completed all interviews and those with missing data were found for race, sex, age, parental education, parental marital status, family income, communal goals, agentic goals, or pubertal status. The low attrition rate and lack of differences suggest that missing data did not have a substantial impact on the findings of the current study.
2.2 |. Procedures
For W1 to W3, adolescents and their parents were interviewed in university research offices. Informed consent (parents) and assent (adolescents) procedures were completed before the interviews began. Interviews took approximately 2.5 to 3 hr for each assessment. Families were compensated $75, $85 and $125 dollars for W1 to W3, respectively, and adolescents were given a small prize valued at $5 to $15 at each wave.
2.3 |. Measures
2.3.1 |. Social goals (W1–W3)
Social goals were assessed with the Interpersonal Goals Inventory for Children Revised (IGI-CR; Trucco et al., 2013). The IGI-CR has been shown to fit a circumplex (Trucco et al., 2013) and has demonstrated strong convergent validity (Trucco et al., 2013). The IGI-CR comprised 32 items all containing the prompt, “When with your peers, in general how important is it to you that…?” Responses are on a 5-point Likert scale ranging from 0 (not at all important to me) to 4 (extremely important to me). The IGI-CR comprised 8 octants containing 4 items each. The octants are Agentic (appearing dominant, independent), Agentic-Separate (appearing to have the upper hand and getting even), Separate (appearing detached and not disclosing thoughts and feelings to others), Submissive-Separate (appearing distant and avoiding rejection from others), Submissive (going along with peers to avoid arguments or upsetting others), Submissive-Communal (putting others’ needs first, valuing approval from others), Communal (valuing solidarity with peers and belongingness), and Agentic-Communal (expressing oneself openly, being respected). Vector scores were computed to represent agentic and communal goals using formulas commonly used by social goals researchers (Locke, 2003; Ojanen et al., 2005). As shown in the formulas below, multiple octants of the interpersonal circumplex are used to calculate the communal and agentic vector scores. The combination of multiple octants allows vector scores to reflect the higher order personality dimensions of communion and agency. Both the communal (α range = .89–.92) and agentic (α range = .82–.84) vector scores demonstrated good reliability across W1-W3.
2.3.2 |. Pubertal status (W1–W3)
The PDS (Petersen et al., 1988) was used to assess pubertal status across W1–W3. The PDS assesses secondary sex characteristics for both males and females. The PDS comprised six questions. Four of these questions, which assess growth in height, body hair development, skin changes, and development relative to same-sex peers, overlap for both males and females. For males, the two remaining items assess facial hair and voice deepening. For females, the two remaining items assess breast development and menarche. Participants rated their pubertal development on a Likert-scale indicating whether a physical characteristic (1) had not yet started growing, (2) had barely started growing, (3) is definitely underway, and (4) seems complete for all but one item assessing menarche (1 = no, 2 = yes). Five of the six items, development relative to same-sex peers was excluded, were averaged to form a composite of pubertal status. The internal consistency of the PDS was adequate for females (α range = .65–.71) and males (α range = .70–.80).
2.3.3 |. Socioeconomic status
Parent reports of adjusted household income and parental education at W1 were each standardized and then summed to represent socioeconomic status (SES). Adjusted household income was calculated using the formula: Adjusted household income = Household income/(Household size)N (Garner et al., 2003). N is a number between 0 and 1, known as an equivalence scale that accounts for the fact that expenditures do not necessarily double when having two versus one child. Following prior work, N = 0.5 was used in the present study (Johnson et al., 2005). Composites of income and parental education have been argued to be more comprehensive assessments of SES (Adler et al., 2012).
2.4 |. Data analytic strategy
Hierarchical linear modeling (HLM) was used to test our hypotheses given the nested structure of the data— repeated measures (level 1) nested within individuals (level 2). HLM was implemented using Proc Mixed in SAS 9.4 (SAS Institute, 2011). Models were estimated using maximum likelihood estimation which allowed us to include subjects with some missing data. HLM allows for the disaggregation of between- and within-person effects which is critical to producing reliable estimates of lagged associations (Hamaker et al., 2015). Between-person variables were calculated as the average of pubertal status, agentic goals, and communal goals across W1–W3. Within-person variables were calculated as deviations from a subject’s overall average at each wave. Between-person variables assessed whether average levels of pubertal status, from early to middle adolescence, were associated with agentic and communal social goals, and whether average levels of agentic and communal social goals, from early to middle adolescence, were associated with pubertal status. Within-person variables assessed whether deviations at a specific wave from youths’ average pubertal status were associated with agentic and communal goals, and whether deviations at a specific wave from youths’ average agentic and communal goals were associated with pubertal status.
Unconditional growth models were used to assess growth in pubertal status, agency, and communion across W1–W3 (Hypothesis 1). To assess within-year associations between puberty and agentic and communal goals, models were arranged such that within-person pubertal status at W1 predicted agentic and communal goals at W1, within-person pubertal status at W2 predicted agentic and communal goals at W2, and within-person pubertal status at W3 predicted agentic and communal goals at W3 (Hypothesis 2). Hierarchal linear models arranged with cross-lagged associations were used to assess longitudinal associations between pubertal status and social goals (Hypothesis 3) and social goals and pubertal status (Hypothesis 4). For example, social goals were arranged such that pubertal status at W1 predicted social goals at W2, controlling for social goals at W1, W2 pubertal status predicted social goals at W3, controlling for social goals at W2 (see Figure 1). Age, biological sex, and SES were included as statistical control variables in all models (Arim et al., 2007; Ellis & Essex, 2007). Assigned biological sex at birth was binary coded (0 = male, 1 = female). Lastly, interactions with sex were tested as a separate block in all models to assess whether including interaction terms led to a significant increment in model fit (Hypothesis 5).
FIGURE 1.
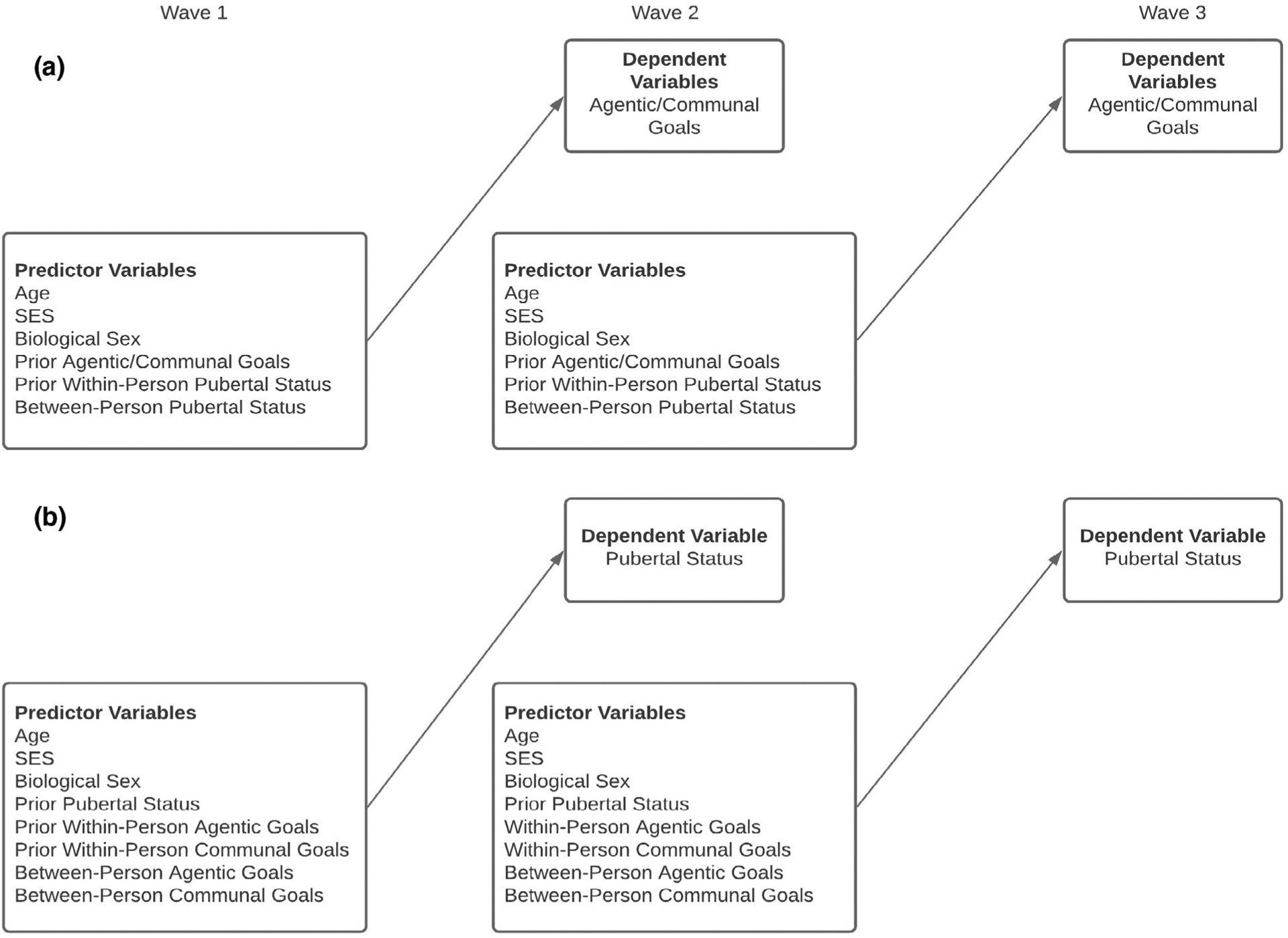
HLM conceptual model for prospective associations between pubertal status and social goals (a) and social goals and pubertal status (Panel b). Agentic/communal goals reflects that agentic goals and communal goals were independent variables in separate models prospectively predicting agentic social goals and communal social goals, respectively
3 |. RESULTS
3.1 |. Unconditional growth models
Descriptive statistics for each variable as well as zero-order correlations are presented in Supporting Information Table S1.
3.1.1 |. Pubertal status
Pubertal status significantly increased from early (W1) to middle (W3) adolescence (B = 0.36, p < .001), and there was significant variability in the intercept of pubertal status at W1 (σ2 = 0.34, p < .001) and slope of pubertal status (σ2 = 0.03, p < .001). Higher initial levels of pubertal status were related to slower growth in pubertal status (covariance = −0.05, p < .001.
3.1.2 |. Social goals
Agentic social goals significantly increased from early to middle adolescence (B = 0.23, p < .001), and there was significant variability in the intercept of agency at W1 (σ2 = 1.23, p < .001) and slope of agency (σ2 = 0.10, p = .01). Higher initial levels of agency were related to slower growth in agency from W1–W3 (covariance = −0.23, p = .02). Communal goals significantly increased from early to middle adolescence (B = 0.27, p < .001) and there was significant variability in the intercept of communion at W1 (σ2 = 1.76, p < .001) and slope of communion (σ2 = 0.27, p = .001). The covariance between the intercept and slope from W1 to W3 was nonsignificant (covariance = −0.10, p = .33), such that initial levels of communal goals were unrelated to growth in communal goals.
3.2 |. Cross-sectional hierarchical linear models
Intraclass correlation coefficients (ICCs) were computed from intercept-only models to assess variability in pubertal status, agency, and communion at the between-and within-person levels of analysis. ICCs indicated that 47% of the variability in pubertal status, 37% of the variability in agentic goals, and 52% of the variability in communal goals occurred at the between-person level. These findings suggest that there was sufficient variability at the within- and between-person level of analyses for the cross-sectional and longitudinal HLMs.
The inclusion of sex interaction terms did not lead to a significant increment in model fit when predicting agentic (χ2(2, N = 386) = 0.50, p = .77) or communal social goals (χ2(2, N = 386) = 5.00, p = .08) and so these interactions were excluded from the model.
3.2.1 |. Pubertal status and agentic goals
As seen in Table 1, females had significantly lower levels of agentic social goals relative to males. Overall pubertal status across W1–W3 (between-person effect) and waves when youth had increases in their pubertal status compared to their average pubertal status across W1–W3 (within- person effect), were both associated with higher levels of agentic social goals.
TABLE 1.
Cross-sectional hierarchical linear models predicting agentic and communcal social goals
| Outcome | Agentic social goals | Communal social goals | ||||
|---|---|---|---|---|---|---|
| Predictor | β | SE | t value | β | SE | t value |
| Intercept | 0.00 | 0.04 | −0.01 | 0.00 | 0.04 | −0.11 |
| Age | 0.06 | 0.04 | 1.37 | 0.04 | 0.04 | 0.88 |
| SES | −0.05 | 0.04 | −1.23 | 0.11 | 0.04 | 2.71** |
| Biological sex | −0.22 | 0.04 | −5.20*** | 0.31 | 0.04 | 7.29*** |
| Within-person pubertal status | 0.11 | 0.02 | 4.90*** | 0.12 | 0.02 | 5.68*** |
| Between-person pubertal status | 0.15 | 0.05 | 3.25** | 0.04 | 0.05 | 0.83 |
| Pseudo R2 | 4.20% | 3.98% | ||||
p < .01;
p < .001.
3.2.2 |. Pubertal status and communal goals
Female sex and SES were both positively associated with higher levels of communal social goals. Increases in adolescents’ pubertal status compared to their average pubertal status across W1–W3 (within- person effect) were associated with higher levels of communal goals. Overall pubertal status across W1–W3 (between-person effect) was not associated with communal goals.1
3.3. Longitudinal hierarchical linear models
The inclusion of sex interaction terms did not improve model fit when predicting agentic social goals (χ2(2, N = 386) = 0.31, p = .20), communal social goals (χ2(2, N = 386) = 0.02, p = .90), or pubertal status (χ2(2, N = 386) = 4.40, p = .11) and so these interactions were not included in the final model.
3.2.3 |. Pubertal status predicting agentic goals
As seen in Table 2, females had significantly lower levels of agentic social goals relative to males. Agentic social goals at the prior wave were significantly associated with agentic social goals at the subsequent wave. Overall pubertal status across W1–W3 (between-person effect) was associated with higher agentic social goals. Within-person pubertal status was not significantly associated with agentic social goals.
TABLE 2.
Prospective within-person associations between pubertal status and social goals
| Outcome | Agentic social goals | Communal social goals | ||||
|---|---|---|---|---|---|---|
| Predictor | β | SE | t value | β | SE | t value |
| Intercept | −0.91 | 0.07 | −13.47*** | 2.55 | 0.09 | 29.33*** |
| Age | −0.03 | 0.05 | −0.67 | 0.01 | 0.06 | 0.22 |
| SES | −0.04 | 0.04 | −0.87 | 0.12 | 0.06 | 2.22* |
| Biological sex | −0.37 | 0.10 | −3.81*** | 0.75 | 0.12 | 6.08*** |
| Prior agentic social goals | 0.50 | 0.04 | 11.24*** | – | – | – |
| Prior communal social goals | – | – | – | 0.95 | 0.06 | 16.43*** |
| Prior within-person pubertal status | 0.03 | 0.04 | 0.77 | 0.02 | 0.05 | 0.42 |
| Between-person pubertal status | 0.14 | 0.05 | 2.66** | −0.02 | 0.07 | −0.35 |
| Pseudo R2 | 5.01% | 6.55% | ||||
| Outcome | Puberta status | |||||
| Predictor | β | SE | t value | |||
| Intercept | 2.67 | 0.02 | 109.40*** | |||
| Age | 0.08 | 0.02 | 4.87*** | |||
| SES | 0.01 | 0.02 | 0.53 | |||
| Biological sex | 0.25 | 0.03 | 7.04*** | |||
| Prior pubertal status | 0.41 | 0.02 | 23.22*** | |||
| Prior within-person agentic goals | −0.03 | 0.01 | −1.81 | |||
| Between-person agentic goals | 0.02 | 0.02 | 1.26 | |||
| Prior within-person communal goals | 0.01 | 0.02 | 0.91 | |||
| Communal goals between-person | 0.01 | 0.02 | 0.67 | |||
| Pseudo R2 | 39.6% | |||||
Note: “–” parameter that was not included in the model.
p < .05;
p < .01;
p < .001.
3.2.4 |. Pubertal status predicting communal goals
Female sex and SES were both positively associated with higher levels of communal social goals. Communal goals at the prior wave were also significantly associated with communal goals at the subsequent wave. Neither within- nor between-person pubertal status was prospectively associated with communal social goals.
3.2.5 |. Social goals predict pubertal status
Female sex was associated with higher levels of pubertal status and pubertal status at the prior wave was significantly associated with pubertal status at the subsequent wave. Agentic and communal social goals were unrelated to pubertal status at both the within- and between-person level of analysis.
3.3 |. Summary of cross-sectional and longitudinal hierarchical linear model
Our analysis provided the ability to distinguish concurrent between- and within-person associations, as well as prospective within-person associations. Consistent with Hypothesis 1, pubertal status, agentic, and communal social goals increased from early to middle adolescence. In line with Hypothesis 2, overall levels of pubertal status from early to middle adolescence as well as times when youth had increases in their pubertal status compared to their average pubertal status across W1–W3 were concurrently associated with agentic goals. Only within-person pubertal status was concurrently associated communal goals. Prospective analyses partially supported Hypothesis 3 such that overall pubertal status across early to middle adolescence was associated with higher levels of agentic social goals (between-person effect). Pubertal status was unrelated to communal goals across waves. Within-person agentic and communal goals were not significantly prospectively associated with pubertal status (Hypothesis 4) and no sex differences were detected in relationships between social goals and pubertal status (Hypothesis 5). Of note, although no interactions with sex were observed, females consistently had higher levels of communal goals and males had higher levels of agentic social goals.
4 |. DISCUSSION
The social goals of agency and communion, rooted in CIIT, are thought to shape how adolescents transact with their social environments and are related to indicators of adjustment and psychopathology during adolescence (Meisel & Colder, 2017; Trucco et al., 2013). Theoretical frameworks from developmental neuroscience and evolutionary psychology suggest that pubertal status may be associated with increases in the valuing of independence and status as well as close peer relationships (Chan et al., 2018; Crone & Dahl, 2012). Further, human and animal studies demonstrating the impact of social context on pubertal development also raise the possibility that agentic and communal goals may impact pubertal development (Moffitt et al., 1992; Stephens & Wallen, 2013). Despite these claims, no prior studies, to our knowledge, have examined whether overall levels of pubertal status during early to middle adolescence (between-person effect) as well as yearly deviations from average pubertal status (within-person effect) are associated with the value adolescents place on independence/dominance (agentic goals) and closeness/intimacy (communal goals), and whether within- or between-person agentic and communal social goals predict pubertal status. The current study assessed growth in pubertal status and social goals across early to middle adolescence (Hypothesis 1) as well as cross-sectional and prospective associations between pubertal status and social goals (Hypotheses 2 and 3) and social goals and pubertal status (Hypotheses 4 and 5). Levels of pubertal status, agentic social goals, and communal social goals all increased from early to middle adolescence. Cross-sectional HLMs indicated that increases in an adolescent’s pubertal status, relative to their average pubertal status during early to middle adolescence, were associated with high levels of both agentic and communal social goals. Further, average levels of pubertal status across the three assessments were positively associated with agentic social goals. Although pubertal status was associated with social goals, these effects were generally small in magnitude (see Tables 1 and 2).
Despite pubertal status demonstrating strong zero-order correlations with communal goals compared to agentic goals, between-person pubertal status was not significantly associated with communal social goals in our HLMs. This was a function of the strong associations between communal goals and biological sex. When removing sex from our models, average levels of pubertal status from early to middle adolescence were significantly associated with communal social goals in the cross-sectional (β = .20, p < .001) and longitudinal models (β = .12, p = .04). Nonsignificant longitudinal associations between within- person pubertal status and communal social goals were partially a function of strong stabilities for communal social goals across waves. When stabilities for communal goals were removed from the longitudinal HLM, within-person pubertal status was significantly associated with communal goals (β = .09, p = .04).
Importantly, all models controlled for the effects of age suggesting the cross-sectional associations between pubertal status and social goals were not simply age-related effects. These finding build on animal studies that have found associations between puberty and social behaviors in the peer context (Schulz & Sisk, 2006). Further, they lend support to evolutionary arguments for the valuing of agency and communion that contend that these social goals facilitate the acquisition of finite resources, favors, and mates (Chan et al., 2018; Ellis et al., 2012). These findings similarly support developmental cognitive neuroscience perspectives that argue that the social motivational tendencies of pursuing close peer relationships and valuing social status results from physiological changes resulting from puberty (Crone & Dahl, 2012).
An important avenue for future research is to assess relevant mechanisms that may link pubertal status with agentic and communal social goals. Developmental cognitive neuroscience and evolutionary psychology frameworks hypothesize that puberty is linked to increases in valuing close relationships and social status through changes in reproductive hormones (Crone & Dahl, 2012). Reproductive hormones, particularly androgens and estrogens, have been associated with status seeking, social dominance, and affiliative behaviors in human and animal studies (Balzer et al., 2015; Ervin et al., 2015; Rowe et al., 2004; Schaal et al., 1996; Tarter et al., 2007; Terburg & Van Honk, 2013). For example, testosterone levels have been positively associated with agentic goals in male undergraduate students (Turan et al., 2014). Reproductive hormones have already been found to mediate associations between pubertal status and sensation seeking, an additional personality trait discussed in these evolutionary psychology and developmental cognitive neuroscience frameworks (Crone & Dahl, 2012; Ellis et al., 2012). Specifically, Harden and colleagues (2018) found testosterone mediated the association between pubertal status and sensation seeking in males between the ages of 13–20.
Further, this mediational chain from pubertal status to social goals through reproductive hormones may be moderated by complex interactions with other hormones, individual difference factors, and social contexts (Hamilton et al., 2015). For example, the dual-hormone hypothesis suggests that testosterone should only be associated with agentic behaviors in the context of low cortisol (Mehta & Josephs, 2010). Indeed, there is evidence that testosterone is associated with status relevant outcomes at low but not high levels of cortisol and that this effect is particularly evident in males (for a review see Dekkers et al., 2019). In sum, these findings point to the importance of future work including assessments of reproductive hormones such as testosterone and estradiol as well as cortisol to clarify the relationship between pubertal status and agentic and communal social goals during adolescence. This future work will help clarify the extent to which the higher-order personality dimensions of agency and communion may be biologically rooted.
Within-person pubertal status was not prospectively associated with either agentic or communal social goals in the prospective models. The timing of the assessments in the current study may have contributed to these null findings. Puberty is a non-linear process whereby there may be periods of rapid development followed by more gradual development (Aylwin et al., 2019; Belsky et al., 2007). Capturing these fluctuations was a central goal in the current study and motivated the parsing of within- and between-person pubertal effects (Dorn et al., 2019; Susman et al., 2019). With that said, the best timing of measurements to capture these changes remains uncertain. To date, the majority of longitudinal studies examining changes in pubertal status employ large time gaps between assessments ranging from months to years (e.g., Mendle et al., 2010). If an adolescent experiences rapid changes in pubertal status over the course of weeks or months, followed by more protracted development the remainder of the year, this may obfuscate true prospective associations between puberty and agentic and social goal development.
Within- and between-level social goals were also not prospectively associated with pubertal status. Considering peer relationships are dynamic during adolescence, the yearlong gap between assessments may have contributed to the null findings between social goals and changes in pubertal status (Hartl et al., 2015). Another possibility is that the IGI-CR assessed what youth value in interpersonal relationships and not how youth behave in interpersonal relationships. Salient communal-related behaviors such as peer rejection and social exclusion as well as agentic-related behaviors such as gaining peer status have been associated with cortisol levels (Kornienko et al., 2013; Stroud et al., 2009). Agentic and communal behaviors, rather than agentic and communal goals, may be related to changes in pubertal status during these developmentally salient social events through altering hypothalamic-pituitary-adrenal (HPA) axis activation of cortisol. Future work that can capture salient agentic and communal behaviors may help clarify associations between agentic and communal social goals and changes in pubertal status.
4.1 |. Limitations
The current study should be understood within the context of certain limitations. Our sample only spanned early adolescence to the beginning of middle adolescence. Pubertal development typically begins in late childhood, thus limiting our ability to generalize our results to the initial period of pubertal development. Moreover, considering males go through puberty later than females, including measures through the end of middle adolescence may have led to the emergence of sex differences in the relationship between pubertal status and agentic goals. Although sample demographics matched those of the county from whence the sample was recruited, families were predominantly non-Hispanic Caucasian and the majority of households were middle class. Considering environmental factors such as social disadvantage and stress are associated with earlier pubertal development, future replication efforts with more diverse samples may benefit from recruiting younger more diverse samples than the current study. From an early age, children are socialized differently based on their gender to value different social behaviors through implicit and explicit parental practices (Mesman & Groeneveld, 2018). Thus differences in agency and communion may be attributable to evolution (biological sex) or socialization (gender identity). Our measures in early and middle adolescence only assessed biological sex and thus, we were unable to examine gender identity as a covariate in our models. Lastly, although our measures of social goals and pubertal status have been shown to have strong psychometric properties, both measures were reliant on self-reports. Future work may benefit from including observer reports of social goals considering observer reports of social goals can provide incremental predictive power and would attenuate concerns regarding reporter bias.
4.2 |. Conclusions
To our knowledge, the current study was the first to formally assess suggestions of developmental neuroscience and evolutionary psychological models of social development that pubertal status is associated with the development of agentic and communal social goals. Support for within-person increases in pubertal status being concurrently associated with higher levels of agentic and communal social goals as well as support significant associations for between-person pubertal status and agentic goals provide initial evidence that puberty contributes to the development of social goals during adolescence. These findings highlight the importance of future work assessing the biological mechanisms, such as reproductive hormones, that may mediate these relationships.
Supplementary Material
ACKNOWLEDGMENTS
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Preparation of this manuscript was supported by grants AA025521 and AA028414 from the National Institute on Alcohol Abuse and Alcoholism and grant R01DA019631 from the National Institute on Drug Abuse.
Footnotes
CONFLICT OF INTEREST
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Given the cross-sectional nature of the within-wave analyses, temporal precedence could not be established. Thus, the cross-sectional model could have depicted pubertal status predicting social goals, or vice versa. When running within-wave HLM where social goals predicted pubertal status, the same pattern of results emerged. Specifically, greater levels of agentic (β = 0.10, p < .001) and communal social goals (β = 0.12, p < .001) compared to their average levels of social goals from W1–W3 were positively associated with pubertal status and overall levels of agentic social goals from early to middle adolescence were also positively associated with pubertal status (β = 0.11, p = .001).
SUPPORTING INFORMATION
Additional Supporting Information may be found online in the Supporting Information section.
REFERENCES
- Adler N, Bush NR, & Pantell MS (2012). Rigor, vigor, and the study of health disparities. Proceedings of the National Academy of Sciences, 109(Suppl. 2), 17154–17159. 10.1073/pnas.1121399109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alberts SC, & Altmann J (1995). Preparation and activation: Determinants of age at reproductive maturity in male baboons. Behavioral Ecology and Sociobiology, 36(6), 397–406. 10.1007/BF00177335 [DOI] [Google Scholar]
- Allen JP, Chango J, & Szwedo D (2014). The adolescent relational dialectic and the peerroots of adult social functioning. Child Development, 85(1), 192–204. 10.1111/cdev.12106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen JP, & Loeb EL (2015). The autonomy-connection challenge in adolescent–peer relationships. Child Development Perspectives, 9(2), 101–105. 10.1111/cdep.12111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen JP, Schad MM, Oudekerk B, & Chango J (2014). What ever happened to the “cool” kids? Long-term sequelae of early adolescent pseudomature behavior. Child Development, 85(5), 1866–1880. 10.1111/cdev.12250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson C, Hildreth JAD, & Howland L (2015). Is the desire for status a fundamental human motive? A review of the empirical literature. Psychological Bulletin, 141(3), 574–601. 10.1037/a0038781 [DOI] [PubMed] [Google Scholar]
- Arim RG, Shapka JD, Dahinten VS, & Willms JD (2007). Patterns and correlates of pubertal development in Canadian youth. Canadian Journal of Public Health, 98(2), 91–96. 10.1007/BF03404316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aylwin CF, Toro CA, Shirtcliff E, & Lominiczi A (2019). Emerging genetic and epigenetic mechanisms underlying pubertal maturation in adolescence. Journal of Research on Adolescence, 29, 54–79. 10.1111/jora.12385 [DOI] [PubMed] [Google Scholar]
- Balzer BW, Duke SA, Hawke CI, & Steinbeck KS (2015). The effects of estradiol onmood and behavior in human female adolescents: A systematic review. European Journal of Pediatrics, 174(3), 289–298. 10.1007/s00431-014-2475-3 [DOI] [PubMed] [Google Scholar]
- Baumrind D (1991). The influence of parenting style on adolescent competence and substance use. The Journal of Early Adolescence, 11, 56–95. 10.1177/0272431691111004 [DOI] [Google Scholar]
- Belsky J, Steinberg LD, Houts RM, Friedman SL, DeHart G, Cauffman E, Roisman GI, Halpern-Felsher BL, Susman E, & NICHD Early Child Care Research Network. (2007). Family rearing antecedents of pubertal timing. Child Development, 78(4), 1302–1321. 10.1111/j.14678624.2007.01067.x [DOI] [PubMed] [Google Scholar]
- Bercovitch FB, & Strum SC (1993). Dominance rank, resource availability, and reproductive maturation in female savanna baboons. Behavioral Ecology and Sociobiology, 33(5), 313–318. 10.1007/BF00172929 [DOI] [Google Scholar]
- Blakemore SJ, Burnett S, & Dahl RE (2010). The role of puberty in the developing adolescent brain. Human Brain Mapping, 31(6), 926–933. 10.1002/hbm.21052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buss DM, & Schmitt DP (1993). Sexual strategies theory: An evolutionary perspective on human mating. Psychological Review, 100(2), 204–232. 10.1037/0033-295X.100.2.204 [DOI] [PubMed] [Google Scholar]
- Caravita SC, & Cillessen AH (2012). Agentic or communal? Associations between interpersonal goals, popularity, and bullying in middle childhood and early adolescence. Social Development, 21(2), 376–395. 10.1111/j.1467-9507.2011.00632.x [DOI] [Google Scholar]
- Castellanos-Ryan N, Parent S, Vitaro F, Tremblay RE, & Séguin JR (2013). Pubertal development, personality, and substance use: A 10-year longitudinal study from childhood to adolescence. Journal of Abnormal Psychology, 122(3), 782–796. 10.1037/a0033133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan T, Wang I, & Ybarra O (2018). Connect and strive to survive and thrive: The evolutionary meaning of communion and agency. In Abele AE & Wojciszke B (Eds.), London, UK: Agency and communion in social psychology (pp. 13–24). Routledge. [Google Scholar]
- Charpentier MJE, Tung J, Altmann J, & Alberts SC (2008). Age at maturity in wild baboons: Genetic, environmental and demographic influences. Molecular Ecology, 17(8), 2026–2040. 10.1111/j.1365-294X.2008.03724.x [DOI] [PubMed] [Google Scholar]
- Crone EA, & Dahl RE (2012). Understanding adolescence as a period of social–affective engagement and goal flexibility. Nature Reviews Neuroscience, 13(9), 636–650. 10.1038/nrn3313 [DOI] [PubMed] [Google Scholar]
- Dawood S, Dowgwillo EA, Wu LZ, & Pincus AL (2018). Contemporary integrative interpersonal theory of personality. The SAGE Handbook of Personality and Individual Differences, 1, 171–202. [Google Scholar]
- Dekkers TJ, van Rentergem JAA, Meijer B, Popma A, Wagemaker E, & Huizenga HM (2019). A meta-analytical evaluation of the dual-hormone hypothesis: Does cortisol moderate the relationship between testosterone and status, dominance, risk taking, aggression, and psychopathy? Neuroscience and Biobehavioral Reviews, 96, 250–271. 10.1016/j.neubiorev.2018.12.004 [DOI] [PubMed] [Google Scholar]
- Dickson JM, & MacLeod AK (2004). Approach and avoidance goals and plans: Their relationship to anxiety and depression. Cognitive Therapy and Research, 28(3), 415–432. 10.1023/B:COTR.0000031809.20488.ee [DOI] [Google Scholar]
- Dorn LD, Hostinar CE, Susman EJ, & Pervanidou P (2019). Conceptualizing puberty as a window of opportunity for impacting health and well-being across the life span. Journal of Research on Adolescence, 29(1), 155–176. 10.1111/jora.12431 [DOI] [PubMed] [Google Scholar]
- Ellis BJ, Del Giudice M, Dishion TJ, Figueredo AJ, Gray P, Griskevicius V, Hawley PH, Jacobs WJ, James J, Volk AA, & Wilson DS (2012). The evolutionary basis of risky adolescent behavior: Implications for science, policy, and practice. Developmental Psychology, 48(3), 598–623. 10.1037/a0026220 [DOI] [PubMed] [Google Scholar]
- Ellis BJ, & Essex MJ (2007). Family environments, adrenarche, and sexual maturation: A longitudinal test of a life history model. Child Development, 78(6), 1799–1817. 10.1111/j.1467-8624.2007.01092.x [DOI] [PubMed] [Google Scholar]
- Ellis BJ, McFadyen-Ketchum S, Dodge KA, Pettit GS, & Bates JE (1999). Quality of early family relationships and individual differences in the timing of pubertal maturation in girls: A longitudinal test of an evolutionary model. Journal of Personality and Social Psychology, 77(2), 387–401. 10.1037/0022-3514.77.2.387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis BJ, Shirtcliff EA, Boyce WT, Deardorff J, & Essex MJ (2011). Quality of early family relationships and the timing and tempo of puberty: Effects depend on biological sensitivity to context. Development and Psychopathology, 23(1), 85–99. 10.1017/S0954579410000660 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ervin KS, Lymer JM, Matta R, Clipperton-Allen AE, Kavaliers M, & Choleris E (2015). Estrogen involvement in social behavior in rodents: Rapid and long-term actions. Hormones and Behavior, 74, 53–76. 10.1016/j.yhbeh.2015.05.023 [DOI] [PubMed] [Google Scholar]
- Garner TI, Ruiz-Castillo J, & Sastre M (2003). The influence of demographics and household-specific price indices on consumption-based inequality and welfare: Acomparison of Spain and the United States. Southern Economic Journal, 70, 22–48. 10.2307/1061630 [DOI] [Google Scholar]
- Guyer AE, Silk JS, & Nelson EE (2016). The neurobiology of the emotional adolescent: From the inside out. Neuroscience & Biobehavioral Reviews, 70, 74–85. 10.1016/j.neubiorev.2016.07.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamaker EL, Kuiper RM, & Grasman RP (2015). A critique of the cross-lagged panelmodel. Psychological Methods, 20(1), 102–116. 10.1037/a0038889 [DOI] [PubMed] [Google Scholar]
- Hamilton LD, Carré JM, Mehta PH, Olmstead N, & Whitaker JD (2015). Social neuroendocrinology of status: A review and future directions. Adaptive Human Behavior and Physiology, 1(2), 202–230. 10.1007/s40750-015-0025-5 [DOI] [Google Scholar]
- Hammer MF, Mendez FL, Cox MP, Woerner AE, & Wall JD (2008). Sex-biased evolutionary forces shape genomic patterns of human diversity. PLoS Genetics, 4(9), e1000202. 10.1371/journal.pgen.1000202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harden KP, Mann FD, Grotzinger AD, Patterson MW, Steinberg L, Tackett JL, & Tucker-Drob EM (2018). Developmental differences in reward sensitivity and sensation seeking in adolescence: Testing sex-specific associations with gonadal hormones and pubertal development. Journal of Personality and Social Psychology, 115(1), 161–168. 10.1037/pspp0000172 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartl AC, Laursen B, & Cillessen AH (2015). A survival analysis of adolescent friendships: The downside of dissimilarity. Psychological Science, 26(8), 1304–1315. 10.1177/0956797615588751 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson DS, Smeeding TM, & Torrey BB (2005). Economic inequality through the prisms of income and consumption. Monthly Labor Review, 128, 11–24. [Google Scholar]
- Johnson DE, Tang A, Almas AN, Degnan KA, McLaughlin KA, Nelson CA, Fox NA, Zeanah CH, & Drury SS (2018). Caregiving disruptions affect growth and pubertal development in early adolescence in institutionalized and fostered Romanian children: A randomized clinical trial. The Journal of Pediatrics, 203, 345–353. 10.1016/j.jpeds.2018.07.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiefer SM, & Ryan AM (2008). Striving for social dominance over peers: The implications for academic adjustment during early adolescence. Journal of Educational Psychology, 100(2), 417–428. 10.1037/0022-0663.100.2.417 [DOI] [Google Scholar]
- Kornienko O, Clemans KH, Out D, & Granger DA (2013). Friendship network position and salivary cortisol levels. Social Neuroscience, 8(4), 385–396. 10.1080/17470919.2013.795500 [DOI] [PubMed] [Google Scholar]
- Locke K (2003). Status and solidarity in social comparison: Agentic and communal values and vertical and horizontal directions. Journal of Personality and Social Psychology, 84, 619–631. 10.1037/0022-3514.84.3.619 [DOI] [PubMed] [Google Scholar]
- Locke KD (2015). Agentic and communal social motives. Social and Personality Psychology Compass, 9(10), 525–538. 10.1111/spc3.12201 [DOI] [Google Scholar]
- Mayeux L, & Kraft C (2018). Social goals moderate the associations between peer status and behavior in middle school. Social Development, 27(4), 699–714. 10.1111/sode.12298 [DOI] [Google Scholar]
- Mehta PH, & Josephs RA (2010). Testosterone and cortisol jointly regulate dominance: Evidence for a dual-hormone hypothesis. Hormones and Behavior, 58(5), 898–906. 10.1016/j.yhbeh.2010.08.020 [DOI] [PubMed] [Google Scholar]
- Meisel SN, & Colder CR (2015). Social goals and grade as moderators of social normative influences on adolescent alcohol use. Alcoholism: Clinical and Experimental Research, 39(12), 2455–2462. 10.1111/acer.12906 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meisel SN, & Colder CR (2017). Social goals impact adolescent substance use through influencing adolescents’ connectedness to their schools. Journal of Youth and Adolescence, 46(9), 2015–2027. 10.1007/s10964-017-0655-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendle J, Harden KP, Brooks-Gunn J, & Graber JA (2010). Development’s tortoise and hare: Pubertal timing, pubertal tempo, and depressive symptoms in boys and girls. Developmental Psychology, 46(5), 1341–1353. 10.1037/a0020205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mesman J, & Groeneveld MG (2018). Gendered parenting in early childhood: Subtle but unmistakable if you know where to look. Child Development Perspectives, 12(1), 22–27. 10.1111/cdep.12250 [DOI] [Google Scholar]
- Moffitt TE, Caspi A, Belsky J, & Silva PA (1992). Childhood experience and the onset of menarche: A test of a sociobiological model. Child Development, 63(1), 47–58. 10.1111/j.1467-8624.1992.tb03594.x [DOI] [PubMed] [Google Scholar]
- Ojanen T, & Findley-Van Nostrand D (2014). Social goals, aggression, peer preference, and popularity: Longitudinal links during middle school. Developmental Psychology, 50(8), 2134–2143. 10.1037/a0037137 [DOI] [PubMed] [Google Scholar]
- Ojanen T, Grönroos M, & Salmivalli C (2005). An interpersonal circumplex model of children’s social goals: Links with peer-reported behavior and sociometric status. Developmental Psychology, 41(5), 699–710. 10.1037/00121649.41.5.699 [DOI] [PubMed] [Google Scholar]
- Petersen AC, Crockett L, Richards M, & Boxer A (1988). A self-report measure of pubertal status: Reliability, validity, and initial norms. Journal of Youth and Adolescence, 17(2), 117–133. 10.1007/BF01537962 [DOI] [PubMed] [Google Scholar]
- Pincus AL (2005). A contemporary integrative interpersonal theory of personality disorders. In Clarkin J & Lenzenweger M (Eds.), Major theories of personality disorder (2nd ed., pp. 282–331). Guilford Press. [Google Scholar]
- Prinstein MJ, Borelli JL, Cheah CS, Simon VA, & Aikins JW (2005). Adolescentgirls’ interpersonal vulnerability to depressive symptoms: A longitudinal examination of reassurance-seeking and peer relationships. Journal of Abnormal Psychology, 114(4), 676–688. 10.1037/0021-843X.114.4.676 [DOI] [PubMed] [Google Scholar]
- Rowe R, Maughan B, Worthman CM, Costello EJ, & Angold A (2004). Testosterone, antisocial behavior, and social dominance in boys: Pubertal development and biosocial interaction. Biological Psychiatry, 55(5), 546–552. 10.1016/j.biopsych.2003.10.010 [DOI] [PubMed] [Google Scholar]
- SAS Institute. (2011). SAS (Version 9.3) [Computer software]. Author. Schaal B, Tremblay RE, Soussignan R, & Susman EJ (1996). Male testosterone linked to high social dominance but low physical aggression in early adolescence. Journal of the American Academy of Child & Adolescent Psychiatry, 35(10), 1322–1330. 10.1097/00004583-199610000-00019 [DOI] [PubMed] [Google Scholar]
- Schulz KM, & Sisk CL (2006). Pubertal hormones, the adolescent brain, and the maturation of social behaviors: Lessons from the Syrian hamster. Molecular and Cellular Endocrinology, 254, 120–126. 10.1016/j.mce.2006.04.025 [DOI] [PubMed] [Google Scholar]
- Silk JS, Siegle GJ, Lee KH, Nelson EE, Stroud LR, & Dahl RE (2013). Increased neural response to peer rejection associated with adolescent depression and pubertal development. Social Cognitive and Affective Neuroscience, 9(11), 1798–1807. 10.1093/scan/nst175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinberg L (1988). Reciprocal relation between parent-child distance and pubertal maturation. Developmental Psychology, 24(1), 122–129. 10.1037/00121649.24.1.122 [DOI] [Google Scholar]
- Stephens SB, & Wallen K (2013). Environmental and social influences on neuroendocrine puberty and behavior in macaques and other nonhuman primates. Hormones and Behavior, 64(2), 226–239. 10.1016/j.yhbeh.2013.05.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stroud LR, Foster E, Papandonatos GD, Handwerger K, Granger DA, Kivlighan KT, & Niaura R (2009). Stress response and the adolescent transition: Performance versus peer rejection stressors. Development and Psychopathology, 21(1), 47. 10.1017/S0954579409000042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Susman EJ, Marceau K, Dockray S, & Ram N (2019). Interdisciplinary work is essential for research on puberty: Complexity and dynamism in action. Journal of Research on Adolescence, 29(1), 115–132. 10.1111/jora.12420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarter RE, Kirisci L, Kirillova GP, Gavaler J, Giancola P, & Vanyukov MM (2007). Social dominance mediates the association of testosterone and neurobehavioral disinhibition with risk for substance use disorder. Psychology of Addictive Behaviors, 21(4), 462–468. 10.1037/0893-164X.21.4.462 [DOI] [PubMed] [Google Scholar]
- Terburg D, & van Honk J (2013). Approach–avoidance versus dominance–submissiveness: Amultilevel neural framework on how testosterone promotes social status. Emotion Review, 5(3), 296–302. 10.1177/1754073913477510 [DOI] [Google Scholar]
- Trucco EM, Colder CR, Wieczorek WF, Lengua LJ, & Hawk LW (2014). Early adolescent alcohol use in context: How neighborhoods, parents, and peers impact youth. Development and Psychopathology, 26, 425–436. 10.1017/S0954579414000042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trucco EM, Wright AG, & Colder CR (2013). A revised interpersonal circumplex inventory of children’s social goals. Assessment, 20(1), 98–113. 10.1177/1073191111411672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trucco EM, Wright AG, & Colder CR (2014). Stability and change of social goals in adolescence. Journal of Personality, 82(5), 379–389. 10.1111/jopy.12069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turan B, Guo J, Boggiano MM, & Bedgood D (2014). Dominant, cold, avoidant, and lonely: Basal testosterone as a biological marker for an interpersonal style. Journal of Research in Personality, 50, 84–89. 10.1016/j.jrp.2014.03.008 [DOI] [Google Scholar]
- Vijayakumar N, de Macks ZO, Shirtcliff EA, & Pfeifer JH (2018). Puberty and the human brain: Insights into adolescent development. Neuroscience & Biobehavioral Reviews, 92, 417–436. 10.1016/j.neubiorev.2018.06.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walvoord EC (2010). The timing of puberty: Is it changing? Does it matter? Journal of Adolescent Health, 47(5), 433–439. 10.1016/j.jadohealth.2010.05.018 [DOI] [PubMed] [Google Scholar]
- Wilson ME, Bounar S, Godfrey J, Michopoulos V, Higgins M, & Sanchez M (2013). Social and emotional predictors of the tempo of puberty in female rhesusmonkeys. Psychoneuroendocrinology, 38(1), 67–83. 10.1016/j.psyneuen.2012.04.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright AG, Hopwood CJ, & Simms LJ (2015). Daily interpersonal and affective dynamics in personality disorder. Journal of Personality Disorders, 29(4), 503–525. 10.1521/pedi.2015.29.4.503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zehr JL, Van Meter PE, & Wallen K (2005). Factors regulating the timing of puberty onset in female rhesus monkeys (Macaca mulatta): Role of prenatal androgens, social rank, and adolescent body weight. Biology of Reproduction, 72(5), 1087–1094. 10.1095/biolreprod.104.027755 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.