

Abstract
Acute ethanol (EtOH) intoxication results in several maladaptive behaviors that may be attributable, in part, to the effects of EtOH on neural activity in medial prefrontal cortex (mPFC). The acute effects of EtOH on mPFC function have been largely described as inhibitory. However, translating these observations on function into a mechanism capable of delineating acute EtOH’s effects on behavior has proven difficult. This review highlights the role of acute EtOH on electrophysiological measurements of mPFC function and proposes that interpreting these changes through the lens of dynamical systems theory is critical to understand the mechanisms that mediate the effects of EtOH intoxication on behavior. Specifically, the present review posits that the effects of EtOH on mPFC N-methyl-D-aspartate (NMDA) receptors are critical for the expression of impaired behavior following EtOH consumption. This hypothesis is based on the observation that recurrent activity in cortical networks is supported by NMDA receptors, and, when disrupted, may lead to impairments in cognitive function. To evaluate this hypothesis, we discuss the representation of mPFC neural activity in low-dimensional, dynamic state spaces. This approach has proven useful for identifying the underlying computations necessary for the production of behavior. Ultimately, we hypothesize that EtOH-related alterations to NMDA receptor function produces alterations that can be effectively conceptualized as impairments in attractor dynamics and provides insight into how acute EtOH disrupts forms of cognition that rely on mPFC function.
Keywords: alcohol, prefrontal cortex, cognition, intoxication, attractor, dynamics systems theory, up-states
Introduction
Acute ethanol (EtOH) intoxication can result in impaired decision-making (George et al., 2005), heightened aggression (Massa et al., 2019), impaired bystander-intervention (Francis et al., 2019; Leone & Parrott, 2019), and an increased tendency to drive despite risks of intoxicated driving (Motschman et al., 2020). The behavioral spectrum of effects of acute EtOH intoxication begins with symptoms such as impairments in skill-requiring tasks, enhanced talkativeness, and relaxation at blood ethanol concentration (BEC) levels lower than 50 mg/dl [10.9 mmol/l]. On the extreme end, respiratory depression, coma, and death can be found at BECs higher than 400 mg/dl [86.8 mmol/l] (Vonghia et al., 2008). Collectively these impairments result in wide-ranging societal dangers. For instance, the Bureau of Transportation Statistics reported that 34% of fatal traffic accidents in the United States in 2018 involved alcohol intoxication. Within this same dataset, it was apparent that the proportion of traffic fatalities involving alcohol have remained steady (U.S. Department of Transportation). Additionally, an assessment of emergency departments across 18 different countries suggests that the alcohol-attributable fraction (AAF) of injuries was 16.4%. The AAF was particularly high in males (20.6%) compared to females (8.6%). When the AAF was broken down by specific injury, 44.1% of all assaults involved alcohol, 14.3% of falls involved alcohol, and 11.1% of traffic injuries involved alcohol (Cherpitel et al., 2015). Collectively, these data suggest that acute EtOH intoxication is a pervasive societal phenomenon that persists in causing harm. These harmful consequences of actions and behavior in social contexts during EtOH intoxication are associated with disruption of cognitive control, decision-making, and working memory, functions strongly moderated by the medial prefrontal cortex (mPFC). In this review we explore the hypothesis that the deleterious effects of EtOH on cognitive behaviors can be explained via actions on NMDA receptor function in the mPFC. Subsequently, understanding how neural activity is altered by acute EtOH can be best conceptualized with low-dimensional state-space approaches.
The mPFC is a cortical brain region involved in regulating decision-making (Malagon-Vina et al., 2018), social interaction (Franzen & Myers, 1973), and cognitive control (van Gaal et al., 2008). This makes the mPFC a promising target of investigation for understanding acute EtOH effects on behavior. The mPFC contributes to functions such as forming predictions, and behavioral selection (Alexander & Brown, 2011), which it accomplishes via its cortical and subcortical connections (Vertes, 2004). Given its highly integrated nature, neural activity within the mPFC requires regulation, typically through neuromodulation, in order to produce optimal cognitive behaviors (Moorman et al., 2015; Onos et al., 2016). Ethanol disrupts mPFC neural activity at both the acute (Tu et al., 2007; Morningstar et al., 2020) and chronic timescales (Cannady et al., 2020) which likely contributes to behavioral impairments observed across the various levels of acute ethanol intoxication.
Understanding how changes in mPFC neurophysiology result in behavioral differences is critical to understand both the acute and chronic effects of EtOH The present review will first highlight literature that investigates how acute EtOH affects mPFC mediated behaviors first in human models and then subsequently in non-human models. Along the way, electrophysiological evidence from single-unit and EEG/MEG recordings will be discussed to generate hypotheses on how EtOH may disrupt mPFC encoding during these tasks. An understanding of the biophysical alterations to mPFC neurons obtained from ex vivo electrophysiological recordings will also be discussed in order to highlight relevant neurophysiological differences that occur during acute EtOH intoxication. Following this, we will synthesize behavioral and biophysical changes into a dynamical systems theory (DST) framework. Ultimately, we aim to generate hypotheses and future directions that will aid in understanding how acute EtOH directly disrupts mPFC-function and behavior.
This review will be structured by building upwards from neuron-level biophysical changes to changes in mPFC population dynamics. Understanding how population dynamics are impaired by EtOH may provide previously unrecognized computational frameworks for understanding cognitive deficits associated with acute EtOH intoxication. These computational frameworks will be described by tracing the direct pharmacological effects of ethanol from the neuronal, biophysical level to the population and behavioral level and we will connect these levels with DST to provide dynamics-based hypotheses on how EtOH affects mPFC-mediated cognitive behaviors. To constrain the scope of the review, we will not address the changes in brain function that are observed following low doses of ethanol or the conditioned effects of ethanol, which are aptly covered in other reviews (van Skike et al., 2019).
Overview of mPFC
The majority of work examining the acute effects of EtOH on mPFC neural activity has been done in rodent models. Therefore, we will focus our neurophysiological discussion on rodent mPFC while also referring to theories and functions of primate dorsal-lateral prefrontal cortex (dlPFC) and anterior cingulate cortex (ACC) ascertained from clinical and behavioral studies. The rodent mPFC is an anterior brain region comprised of three anatomically distinct subregions, the anterior cingulate (Cg), prelimbic (PL), and infralimbic (IL) (van Eden & Uylings, 1985). The dorsal mPFC generally is involved with processing of information from the past, present, and affective states whereas the ventral mPFC is involved with processing of visceromotor responses (Vertes, 2004). The rodent mPFC is suggested to be functionally, structurally, and evolutionarily most homologous with the primate ACC (Heilbronner et al., 2016; Laubach et al., 2018).
Anatomical projections are delineated across the dorsal-ventral axis of the rodent mPFC. For example, projections from the PL are found in the insular cortex, claustrum, dorsomedial striatum, nucleus accumbens, olfactory tubercle, and raphe nuclei whereas projections from the IL are found in the basal forebrain, amygdala, hypothalamus, and brainstem (Vertes, 2004). Functional delineation is more difficult, however, and both subregions appear necessary for complex, goal-directed behaviors, and decision-making (Moorman et al., 2015; Riaz et al., 2019). Ultimately this suggests that mPFC-dependent behaviors arise from integrating information from several different brain regions. Understanding how this integration of information leads to top-down control of behavior is important to understand how neural circuits give rise to behavior.
Theories describing the computational properties of mPFC are numerous (Cavanagh et al., 2009; Alexander & Brown, 2011; Euston et al., 2012; Bekolay et al., 2014) and it is important to note that several of these theories are complementary and not mutually exclusive. One of the most prominent theories of mPFC action is in adaptive control and error monitoring. In short, the mPFC detects errors and conflict related to executed behaviors and regulates resources reflecting priorities of top-down control accordingly (Cavanagh et al., 2009). Alexander and Brown (2011) formalized the predicted response-outcome model (PRO) that explained several observed phenomena within mPFC data. The PRO presupposes that the mPFC generates expectations of action values and updates these expectations according to outcomes. Particularly, activity is highest when an expected outcome is not met (Alexander & Brown, 2011). The strength of this model is that it accounts for several disparate findings hypothesized to be functions of the mPFC, such as error and conflict monitoring, the error-related negativity signal, the N2 component of speed-accuracy tradeoffs, among others. (Alexander & Brown, 2011).
A separate but similar model of mPFC uses artificial intelligence (AI) and applies reinforcement-learning to several different tasks. Wang and colleagues (2018) sought to unify reward-prediction error (RPE) learning with mPFC related activity and developed the meta-reinforcement learning model of PFC. It posits that model-free dopaminergic (DA) projections to the PFC train the PFC to operate as a model-based and independent learning system. This suggests that disruptions to DA during initial learning phases may alter model-based cortical learning systems. Additionally, this suggests that reward-related information is first learned in a model-free context while the PFC develops internal representations of expectations that are subsequently used to drive top-down control in response to conflict and error.
Taken together, these models suggest that the computational role for the mPFC is to learn and monitor action-outcomes and their expectancies via memory processes. Action-outcomes must first be learned. Then, outcomes of those actions must be monitored in order to refine and update them if needed. Situations wherein error and conflict are detected may require more top-down control over subsequent actions whereas situations wherein the predicted outcome is observed will be more automatic. This suggests that there are at least two components to mPFC function that can go awry during acute EtOH intoxication, namely, learning and monitoring. However, a third component binds these two and allows for the integration of information across timescales – memory. More specifically, working memory, is required for high level cognitive functions and has been shown to have deficits during acute EtOH intoxication (Finn et al., 1999; Saults et al., 2007). Working memory requires sensory and perceptual information to be maintained over a delay period as a representational trace and must be actively consolidated and refreshed (Ricker et al., 2018). By doing so, working memory can maintain information over delays where external information becomes unavailable. The primate PFC has been hypothesized to be critically important for the encoding of working memory by maintaining stimulus information over delay periods via persistent activity (Goldman-Rakic, 1995). Persistent activity is a proposed mechanism by which representational traces of stimuli can be maintained, either by elevated firing rates or short-term plasticity (STP) and is discussed in more detail below.
The acute effects of alcohol on medial prefrontal cortex
Clinical Evaluations of Behavior and Neurophysiology Following Acute EtOH
A general theme underlying acute EtOH intoxication is the loss of cognitive control. Cognitive control is a broad term that refers to the ability of an agent to regulate thoughts and actions in order to attain an internally maintained goal (Botvinick et al., 2001; Braver, 2012). This phenomena is conceptualized in a way that requires several “subordinate” cognitive process required for the active maintenance and rapid updating of information (O’Reilly, 2006). Further, cognitive control has been fractionated into two processes that describe how cognitive resources are allocated, namely reactive and proactive control (Braver, 2012). Braver (2012) contends that proactive control described the allocation of cognitive resources in a stimulus independent manner and typically well in advance of the execution a goal-directed behavior. This is contrasted with reactive control that describes the allocation of cognitive resources in a stimulus-dependent manner typically close in time to the execution of and feedback from a goal-directed behavior. Below we will highlight studies that have shown EtOH interfering with both systems of control. For example, error monitoring likely engages reactive control systems whereas working memory likely engages proactive control.
The integration of present context with past experiences is essential to match goal directed, optimal behaviors to current conditions (Pezzulo et al., 2014). However, acute EtOH intoxication has been shown to reduce error-related mPFC activity – particularly in the ACC (Bartholow et al., 2012). Additionally, EtOH has been shown to reduce sustained attention at moderate (0.06 BrAC) and high doses (0.09 BrAC) while also impairing verbal memory at moderate doses in male college students (Magrys & Olmstead, 2014). Evidence for impaired stimulus response times and inappropriate error responses have additionally been found accompanied with evidence for dose-dependent (0.0%, 0.05%, 0.1% BrAC) diminishments in frontal error-signaling during a Go/No-Go task (Anderson et al., 2011). Most importantly, Bailey and colleagues (2014) showed that acute EtOH intoxication’s (0.8 g/kg EtOH dose) effects on cognitive control were most prominent following errors but not after correct choices. This was also accompanied by a reduction in frontal slow wave activity in the EtOH group following errors. Despite initial reductions in error monitoring, participants were able to adjust performance 2 trials after an initial error. This led Bailey and colleagues (2014) to suggest that acute EtOH specifically disrupts and slows performance monitoring. Consistent with this idea, EtOH (doses ranging from 0.6 – 0.8 g/kg EtOH) increases both error and/or response rates as well as decreases information processing in several cognitive tasks (Tiplady et al., 2001; Mackay et al., 2002; Tiplady et al., 2003; Scholey et al., 2012). Overall, despite the broad range of processes potentially included in the term cognitive control, EtOH reliably induces reductions in performance monitoring and error-detection that are accompanied by neurophysiological correlates. Understanding the alterations in neurophysiological processes that lead to these functional reductions is critical to understanding acute EtOH’s role in disrupting cognitive control.
Beyond cognitive control, working memory and attention are also critically disrupted following acute EtOH intoxication. Benson and colleagues (2019) recently investigated the effects of alcohol, alcohol plus an energy drink, or placebo. They administered a targeted 0.6 g/kg dose of orally consumed vodka and ran a battery of attentional and memory-related tasks. Within these tasks, they found that alcohol alone produced a typical speed-accuracy tradeoff where more rapid responding resulted in a greater number of errors across task types. There was a pronounced deficit resulting from alcohol in the attention-related composite score, but the working memory composite score did not show differences. Specific working memory tasks such as Serial Sevens did show deficits following alcohol whereas their Memory Scanning and Visuospatial Working Memory tasks showed no deficits. This suggests EtOH may be differentially altering working memory depending on sensory modalities.
Using cognitive tasks that stress speed versus accuracy, or vice versa, the effects of EtOH and benzodiazepines (BZDs) have been compared. These studies consistently implicate BZDs as reducing speed within these tasks without reducing accuracy whereas EtOH either has no effect on speed or increases speed while consistently decreasing accuracy (Tiplady et al., 2001; Tiplady et al., 2003; Schweizer et al., 2004). The speed-accuracy tradeoff may be a product of NMDA receptor antagonism within the mPFC, a feature of acute EtOH intoxication (Weitlauf & Woodward, 2008), given that BZDs act predominantly as GABA agonists. Acute EtOH prominently affects NMDA receptors in the mPFC at concentrations as low as 22 mM (corresponding to a BEC of 0.1%) whereas GABA and AMPA related receptors require significantly higher concentrations in the mPFC (> 88 mM, ~0.4% BEC) (Weitlauf & Woodward, 2008). This ultimately suggests that differences between EtOH’s and BZD’s behavioral changes in these tasks may be due to NMDA receptor antagonism in the mPFC. Additional studies, however, have produced null results during EtOH dosing on similar tasks (Simpson & Rush, 2002; Acons et al., 2006). Conflicting results across these studies may be due to a host of differences among methodology and dosing, genetic backgrounds, and EtOH history. This highlights the need for accurate and standardized pre-clinical measures of cognitive control and working memory in order to adequately control for individual differences.
Evaluations of neural activity following acute EtOH intoxication in clinical populations is widely done utilizing EEG and MEG. In these studies, event-related potentials (ERPs) that are sensitive to acute EtOH have been found. Nelson and colleagues (2011) investigated feedback-related negativity (FRN) in a two-choice gambling task. They found that similarly to error-related negativity (ERN), the ERP FRN was inhibited following acute EtOH intoxication (0.087% BrAC). Similar studies have confirmed that acute EtOH intoxication inhibits error-related signals in the executive network in both Stroop (Kovacevic et al., 2012) and Flanker (Beaton et al., 2018) tasks that measure elements of cognitive control (EtOH doses ranging between 0.55 – 0.60 g/kg). It is known from both clinical and pre-clinical literature that mPFC neural activity increases as a function of task difficulty and conflict (Gill et al., 2000) and EtOH’s impact on attenuating this typical increase in activity is likely contributing to behavioral deficits within these tasks.
Working memory relies on persistent mPFC activity (Liu et al., 2014; Bolkan et al., 2017). Several clinical behavioral studies have shown deficits in working memory following acute EtOH intoxication (Weissenborn & Duka 2003 (dose of 0.8 g/kg EtOH); Gundersen et al., 2008 (two doses: 0.02% – 0.08% BrAC)). Specifically, EtOH appears to reduce individuals’ ability to inhibit responses utilizing working memory ultimately leading to more impulsive decisions and deficits in task performance (Finn et al., 1999 (targeted dose between 0.07% - 0.09% BrAC); Saults et al., 2007 (doses of 0.65 g/kg or 0.72 g/kg EtOH)). Despite the clear role acute EtOH has in disrupting working memory processes, only a few clinical electrophysiological studies have been completed. Mixed results have been seen within these studies, for instance, Sanz-Martin and colleagues (2014) found a significant reduction of fronto-parietal theta power following acute EtOH (0.5 g/kg EtOH) during task-related epochs. Additionally, there were decreases between theta and alpha synchronization. However, Boha and colleagues (2009) only found a non-significant decrease in frontal theta power during a working memory task. This may be due to task differences given that Sanz-Martin et al (2014) utilized a visuo-spatial working memory task whereas Boha et al (2009) utilized an arithmetic-based working memory task. Follow up analyses on the Boha et al (2009) dataset revealed that the synchronization likelihood of the theta band increased during task performance, however, this increase was attenuated following acute EtOH (0.2 g/kg or 0.4 g/kg EtOH) (Molnár et al., 2009). None of the three mentioned studied found behavioral deficits, however, their dosages were lower than previous studies, ranging between 0.2 g/kg and 0.5 g/kg. Ultimately the results of these three studies suggest that EtOH may be attenuating theta-related activity during working memory tasks, however, it is task and dosage dependent and does not produce clear behavioral deficits despite potential electrophysiological differences.
Preclinical Evaluations of Behavior and Neurophysiology Following Acute EtOH
In pre-clinical models, there are fewer studies of acute EtOH intoxication on behavior compared to studies of chronic EtOH treatments. This is in part due to the importance of chronic EtOH’s effects on memory and decision-making, as well as the difficulty of maintaining task-engagement in acutely intoxicated animals. Despite this, there is a set of consistent findings that can be discussed. Deficits are seen in spatial working memory (Givens, 1995; Matthews et al., 1996; Chin et al., 2011; Ketchum et al., 2016) and non-spatial working memory following doses of EtOH as low as 0.75 g/kg (Givens & McMahon, 1997). A potential explanation for the observed effects in spatial working memory is the observed deficit in hippocampal place-cell specificity (Matthews et al., 1996). An additional observed deficit is in contextually conditioned fear responses that is prominent during trace but not delay conditioning following EtOH pretreatment during the training phase (Hunt et al., 2009; Kutlu & Gould, 2016). Novel object recognition is also impaired following acute EtOH intoxication (Ryabinin et al., 2002) and can be pharmacologically rescued following enhancement of mGlu5 receptor activity using a positive allosteric modulator (Marszalek-Grabska et al., 2018). Last, acute EtOH’s effects may be directly mediated by rodent mPFC resulting in disruptions to cognitive flexibility that are not dependent on the mPFC itself (Devenport & Hale, 1989). Van Skike and colleagues (2019) extensively review behavioral, cognitive, and neurophysiological deficits associated with acute EtOH impairment far beyond the scope of what we are presently able to in this current review. It should be noted, however, that consistent effects of acute EtOH in pre-clinical models are difficult to find owing to a wide variety of experimental differences as well as nuanced differences in the cognitive systems tested and their uncertain homologies to humans (van Skike et al., 2019). Despite this, a potential common theme throughout these pre-clinical studies is the importance of maintaining clear representational traces of external stimuli as well as adequately reacting to interference events such as errors. Acute EtOH may be disrupting either the maintenance or formation of persistent activity and/or STP mechanisms which subsequently disrupts the behaviors observed.
There is a critical mass of pre-clinical and clinical data describing the deficits in behavior following acute EtOH intoxication, and it is necessary to integrate these data into a working model capable of describing how the effects of ethanol on mPFC circuits contributes to these behaviors. Tu and colleagues (2007) found dose dependent reductions in spontaneous mPFC unit activity following EtOH in both anesthetized (0.375 – 3.5 g/kg EtOH) and ex vivo electrophysiology preparations (17-100 mM, corresponding to 0.08 – 0.4% BEC). These reductions were within the context of up-down states (UDS), which are periods of sustained neural firing interspersed by periods of quiescence across populations of (typically) cortical neurons. UDS are observed in anesthetized preparations and are otherwise naturally occurring during slow-wave sleep (SWS) (Dao Duc et al., 2014). From these reductions in spontaneous activity, it was suggested that acute EtOH intoxication may be inhibiting and depressing mPFC activity. Data in awake-behaving animals treated with acute EtOH (1.0 or 2.0 g/kg EtOH) did not show a similar reduction in neural activity, however, differences in firing rate variability were observed (Morningstar et al., 2020) which is consistent with NMDA receptor antagonism (Homayoun et al., 2005). Recently, Cannady and colleagues (2020) investigated the differential excitability changes in the orbitofrontal cortex (OFC) and ACC following voluntary alcohol consumption. Briefly, mice were allowed to drink for 1 day, 1 week, 4 weeks, or 7 weeks in an intermittent alcohol access protocol. Following each group’s respective endpoint, tissue was extracted, and intrinsic excitability of neurons was established. Evoked action potentials were increased relative to water controls after 1 day in the ACC whereas no effect was observed after 1 day in the OFC. Additionally, evoked action potentials were decreased after 1 week in the ACC whereas they were increased in the OFC. At 4- and 7-weeks evoked action potentials in the ACC matched that of water controls whereas in the OFC they remained elevated. This suggests that the ACC is especially sensitive to the acute and short-term adaptations of EtOH yet remains robustly unaffected after prolonged EtOH use (Cannady et al., 2020). Additionally, and as mentioned above, acute EtOH at doses corresponding to intoxication (22 mM) has been shown to selectively affect NMDA receptor function in mPFC ex vivo preparations whereas effects on GABA and AMPA receptors required much higher concentrations of EtOH (88 mM; Weitlauf & Woodward, 2008). As such, we will critically and nearly exclusively examine the role NMDA receptors may play in the cognitive deficits associated with acute EtOH intoxication.
Beyond these few studies, little data exists looking at how EtOH influences mPFC neural activity specifically. Despite the scarcity of reports on findings in the mPFC, single-unit awake-behaving electrophysiology findings do exist for other brain regions. The main take-aways and findings will be summarized in brief. Chapin and colleagues (1986) found disrupted attentional gating in the somatosensory cortex during a treadmill task in which animals were exposed to EtOH (0.3 – 1.5 g/kg EtOH) and either forced to move, be held, or be at rest. While these behavioral conditions were active, a small paw-shock was elicited to evoke somatosensory neural activity. They ultimately found that the neural correlate of the small paw-shock in running conditions was inhibited within the somatosensory cortex whereas after EtOH treatment the neural correlate returned. Beyond the somatosensory cortex, additional work has been done within the hippocampus. EtOH has reliably been shown to disrupt spatial working memory and Matthews and colleagues (1996) found that this may be due to impairments in spatial encoding following acute EtOH treatment (2.0 g/kg EtOH). Specifically, under saline and baseline conditions they found that the hippocampus exhibited space-specific activation at different zones of a radial-arm maze. After EtOH, hippocampal activation became non-specific and diffuse across zones of a radial-arm maze. An additional hippocampal study investigated the response of CA1 neurons during a working-memory task. During the delay period of the working memory task, neural activation was observed in the saline and control conditions. After a dose of EtOH (0.75 g/kg), neural activity during the delay period was suppressed (Givens & McMahon, 1997; Givens et al., 1998). While failures to encode task-relevant information in these hippocampal and cortical regions have been reported, few reported results on mPFC-specific activity exist in response to acute EtOH during a task. These limited reports highlight a dire need in the EtOH literature to directly measure neural activity during behavior while introducing consistent EtOH manipulations (van Skike et al., 2019).
Persistent Activity and Dynamic Coding within PFC
A common theme throughout the remainder of this review is the idea that persistent activity plays a critical role for working memory, decision-making, and learning and likely serves as a consistent mechanism for disparate cognitive processes affected by EtOH. Persistent activity is a feature of recurrent neural circuits that accommodates the maintenance of relevant information despite transient external stimuli (Curtis & Lee, 2010). Historically, persistent activity has been hypothesized to be supported by single-unit activity in the PFC (Funahashi et al., 1989). However, recent work has posited that persistent activity is maintained dynamically at the network level and that select ensembles of neurons may be more informative than single units; this idea is referred within the working memory literature as dynamic coding and is an extension of persistent activity (Wimmer et al., 2014; Rajan et al., 2016; Murray et al., 2017; Cavanagh et al., 2018; Lundqvist et al., 2018). Dynamic coding is the ability of a working memory system to alter its representations over time (Murray, 2017) potentially via STP mechanisms (Barak & Tsodyks, 2007) which ultimately necessitates stable representations to be formed at the level of a population of neurons that often exhibit attractor dynamics. Specifically, Barbosa et al (2020) defines persistent activity as a feature requiring attractor dynamics in order to drive elevated levels of neural firing across a delay whereas activity-silent periods are driven by STP mechanisms. Put simply, STP mechanisms contain a blueprint for how persistent activity should unfold, and this blueprint forces population-level neural activity into a structured state often referred to in working memory as a bump attractor. A bump attractor resembles a gaussian where a subset of neurons will fire maximally for their preferred stimulus, a subset will fire half-maximally if the stimulus is similar to their preferred, and a subset will fire minimally if the stimulus is dissimilar to their preference (Compte et al., 2000; Wang et al., 2014). For example, upon reactivation of a memory, persistent activity was observed and resembled a bump-attractor facilitated by STP changes (Barbosa et al., 2020). Dynamic coding, therefore, ultimately suggests that persistent activity both influences STP and STP later influences persistent activity. Working together, these disparate modes of neural activity allow for the reactivation and reconsolidation of short-term memories (Barbosa et al., 2020).
Persistent activity may be biologically instantiated by slow, recurrent connections across cortical pyramidal populations (Compte et al., 2000) and/or STP mechanisms at the synapse that allow for temporal information to be stored (Barak & Tsodyks, 2007). As mentioned below, EtOH may be disrupting multiple processes necessary for learning, working memory, and monitoring that are dependent on persistent activity and/or features of dynamic coding such as activity-silent periods (periods without persistent activity). Persistent activity and mechanisms associated with it have long been linked with working memory; however, recent work has shown that it may also be essential to evidence accumulation and decision-making (Curtis & Lee, 2010; Mante et al., 2013). This is accomplished by constraining neural activity along a line attractor, which is an infinite set of fixed points organized along a line, that neural activity will tract onto during evidence accumulation (Mante et al., 2013). Critically, different stimuli classes are organized into their own respective line attractors and neural systems must be able to flexibly manage them (Mante et al., 2013). It has also been suggested to be an essential feature of reinforcement learning by allowing a system to temporally assign credit to an action (Curtis & Lee, 2010). Ultimately, persistent activity and dynamic coding allow for a common computational process to contribute to disparate cognitive behaviors that may be influenced by acute EtOH.
Therefore, the working hypothesis of acute EtOH intoxication’s effects on mPFC-mediated behaviors and mPFC neural activity is that the introduction of EtOH alters mPFC function by preventing both the formation of neural ensembles necessary for conflict monitoring and evaluation as well as persistent activity in ensembles related to working memory tasks and proactive control. Specifically, EtOH inhibits the rise in activity normally seen during these tasks, either in the delay period or during errors and conflict, in contrast to the prevailing view of an overall decrease in activity. This prevention of persistent, recurrent activity is likely leading to behavioral deficits observed following acute EtOH intoxication in both cognitive control and working memory or attentional domains.
mPFC NMDA Receptors Are A Critical Target of Acute EtOH
If acute EtOH prevents the formation of mPFC signals related to conflict monitoring and performance evaluation, then it is necessary to ask what are the key biophysical and pharmacological changes that EtOH induces in mPFC. For the present purposes, this review will focus on localized changes in mPFC following acute EtOH exposure with an emphasis on glutamatergic neurotransmission. The NMDA receptor is a critical target of acute EtOH’s actions on the mPFC (Weitlauf & Woodward, 2008; Badanich et al., 2013). The NMDA receptor was functionally isolated from similar glutamatergic receptors in the 1980s due to its innate property of rapid calcium-dependent desensitization (CDD) (Collingridge et al., 1983; Zorumski et al., 1989). CDD occurs following heightened intracellular calcium that in turn causes second messenger pathways to inactivate the NMDA receptor (Sibarov & Antonov, 2018). The NMDA receptor requires dual bindings of glycine and glutamate in addition to post-synaptic depolarization; once these conditions are met, the receptor pore is opened, the magnesium block is lifted, and the channel is permeable to cation flow that includes high calcium permeability, resulting in rapid depolarization of the cell (Vyklicky et al., 2013). This ultimately results in the NMDA receptor functioning as a coincidence detector between pre- and postsynaptic cells making it highly important for neural plasticity and learning (Seeburg et al., 1995). Additionally, computational work heavily implicates the NMDA receptor as being an essential feature for recurrent excitation within the mPFC and is important for the maintenance of information in cortex (Compte et al., 2000). Specifically, in simulations, enhancing NMDA receptor synaptic transmission protects against the disruptive effects of distractors (Compte et al., 2000) therefore enhancing the stability of traces in working memory.
The NMDA receptor is composed of 4 subunits. Dimeric NMDA receptors contain 2 GluN1 subunits and 2 matching GluN2 subunits whereas triheteromeric NMDA receptors contain two variations of the GluN2 subunit. The GluN2 subunit has at least four different subunits (GluNR2A, GluNR2B, GluNR2C, GluNR2D) that can be expressed in different quantities across brain region and development (Sibarov & Antonov, 2018). Investigating the differences specifically between NR2A and NR2B subunits has been a large focus of research. For instance, evidence exists that different NMDA subunit compositions play differential roles in long-term potentiation (LTP), de novo long-term depression (LTD), and STP (Massey et al., 2004; Volianskis et al., 2015). NR2B subunits are localized both at the synapse and extrasynaptically whereas NR2A subunits are predominately found at the synapse (Petralia et al., 2010; Papouin et al., 2012). Additionally, the state of the receptor (desensitized vs. not) seems to play a critical role in mediating differential pharmacological effects of two clinically relevant NMDA receptor open-channel blockers—memantine and ketamine. Memantine stabilizes and prolongs CDD in NR2A-containing NMDA receptors whereas ketamine reduces desensitization in NR2B containing NMDA receptors (Glasgow et al., 2017). This highlights the complex and subunit-dependent effects of NMDA antagonists. Therefore, understanding precisely how EtOH antagonizes the NMDA receptor is necessary as it could differentially influence plasticity.
EtOH functions as an antagonist at the NMDA receptor and effectively reduces excitatory neurotransmission. Specifically, NMDA receptor conductance (Weitlauf & Woodward, 2008; Badanich et al., 2013) in the mPFC is decreased (22 – 44 mM) and CDD is enhanced (Boikov et al., 2020) in the presence of EtOH (9-30 mM). EtOH likely disrupts sodium-calcium exchange (NCX) resulting in the enhancement of CDD by prolonging heightened intracellular calcium which subsequently enhances the activation of intracellular signaling mechanisms relevant to CDD (Sibarov & Antonov, 2018; Boikov et al., 2020). Functionally this may result in EtOH decreasing the probability of opening NMDA receptor channels and decreasing the time they stay open (Wright et al., 1996). Transmembrane domain specific actions on the GluN1 and GluN2 subunits have been shown to contribute to EtOH’s antagonistic effects on NMDA receptors. TM3 and TM4 in both NR2A and NR2B subunits has been shown to be particularly affected by EtOH antagonism (Smothers & Woodward, 2016; Zhao et al., 2016). It has also been shown that EtOH reduces the duration, amplitude, and frequency of cortical up-states in both anesthetized (Tu et al., 2007; Morningstar et al., 2020) and ex vivo preparations (Tu et Al., 2007; Woodward & Pava, 2008) which suggests disruptions to spontaneous activity that may be attributable to the physiological changes mentioned above. Specifically, neurons often appear to be less able to switch from down-states to up-states, which can be seen in a dose-dependent lengthening of down-state durations (Morningstar et al., 2020). Additionally, these alterations to up-states are seen at concentrations at or below 44 mM where NMDA-mediated synaptic transmission is particularly disrupted whereas GABAergic activity and AMPA-related activity are spared (Weitlauf & Woodward, 2008). In somatosensory ex vivo preparations, Sessler and colleagues (1998) found that EtOH enhanced the rheobase of recorded cells which increases the minimum current needed for the neuron to generate an action potential. Taken together, EtOH’s antagonistic effects on NMDA receptors is necessary for alterations of mPFC neural activity. Specifically, EtOH appears to selectively target NMDA receptors in the mPFC at concentrations consistent with observed clinical deficits in cognition which may subsequently explain the blunted neurophysiological response in the mPFC during cognitive tasks. Overall, EtOFI appears to reduce NMDA conductance, enhance CDD, and ultimately disrupt channel openings. It is likely that EtOH antagonism of NMDA receptors is disrupting mechanisms necessary for recurrent excitation involved as well as mechanisms necessary for STP via prolonging CDD and the intracellular changes associated with it.
Given NMDA receptors’ role in neuronal excitability, examining their computational repertoire and contributions is critical for the present review. As discussed above, persistent activity during delayed periods or periods of conflict and error is critical for mPFC-mediated behaviors and functions such as working memory, attention, and cognitive control. Silencing or reducing NMDA receptor activity in silico and in vivo reliably decreased persistent activity as well as increased susceptibility to distractors in the environment (Compte et al., 2000; Wang et al., 2013). Specifically, Wang and colleagues (2013) probed layer 3 NMDA receptor- expressing neurons that expressed the NR2B subunit showing that this specific NMDA receptor configuration was important for persistent activity both in silico and in vivo within the primate dIPFC. While the in silico model predicted NR2B to be necessary for persistent activity, both NR2A and NR2B subunit blockade via specific subunit antagonists showed deficits in persistent activity in vivo despite differences in the subunits function and kinetics (Massey et al., 2004). Homological differences between rodent and primate may also exist given that McQuail and colleagues (2016) found that the NR2A subunit but not the NR2B subunit was necessary for rodent persistent activity within layer 2/3 mPFC neurons. Importantly, the layer 3 microcircuit has been both computationally and empirically hypothesized to be relevant for persistent activity in primates (Goldman-Rakic, 1995; O’Reilly and Frank, 2006; Wang et al., 2013) as well as rodents (McQuail et al., 2016), however, additional work determining the homology of layer 3 PFC microcircuits between rodent and primates is likely necessary (Preuss, 1995). Taken together, this set of studies heavily implicates the NMDA receptor as being a critical component of microcircuit-specific profiles of persistent activity within the mPFC of both rodents and primates.
Seung (1996) explicitly explored the relationship between NMDA receptors and persistent activity. In that work, they noted that the slow dynamics of NMDA currents (on the order of ~100 ms) may be essential for increasing the systems tolerance to small perturbations. In comparison, if AMPA receptors’ rapid timescale of ~5 ms were utilized in Seung’s (1996) model, it would greatly decrease a systems tolerance to perturbations in synaptic tuning (Seung 1996; Wang, 2001). Additionally, Wang (2001) reviewed additional components of the NMDA receptor that make it particularly well-tuned for persistent activity, namely, the slow decay time, voltage dependent gating, and slow glutamate unbinding all appear as essential candidate mechanisms for persistent activity. As mentioned above, EtOH enhances CDD in NMDA receptors which effectively increases the decay time of NMDA receptor currents in desensitized states (Boikov et al., 2020). In addition, periods of learning and plasticity such as the onset of ocular dominance in ferrets exhibit decreased NMDA receptor decay times and enhanced NR2A subunit expression (Roberts & Ramoa, 1999). Therefore, there appears to be a precise tuning of NMDA receptors that emerges as a result of learning and plasticity. If EtOH alters this precise tuning, it will ultimately lead to disruptions to persistent activity and STP by decreasing the tolerance to perturbations of the system. An extension of this hypothesis is that the degree to which a system has learned will influence the robustness of the system to perturbations, such as EtOH, via tighter tuning of synaptic properties. Therefore, EtOH may most influence behaviors that are still in the formative stages of learning, that require persistent activity, or that require subtle updates such as in error correcting, monitoring and memory reconsolidation.
Taken together, EtOH appears to have consequences on NMDA receptor conductance and duration in the desensitized state, the probability to open, the duration of opening, and the decay time of NMDA receptor currents. These appear to be essential features in EtOH’s effects on NMDA receptors and subsequently understanding how these changes in individual neurons impacts networks of neurons is necessary. Particularly, it is necessary to understand these changes evoked by EtOH in well-known dynamical motifs such as persistent activity, bump attractors, and line attractors in order to hypothesize further about acute EtOH’s role in disrupting cognitive processes.
Approximating mPFC Networks Using Recurrent Neural Networks
It is critical to characterize how networks of neurons encode information to understand how EtOH might impair this process. Recurrent neural networks (RNNs) are a generic class of artificial neural network (ANN) models that utilize feedback connections allowing for complex, time-varying signals to be recapitulated. RNNs can be expressed in the following form:
| Equation 1 |
Within Eq. 1, every bolded term represents a matrix (r, W, b, x). The instantaneous firing rate of a matrix of neurons is updated every time-step t by the output of a transfer function f that is determined by the sum of each neurons rate multiplied by its connectivity weights W, the input x, and a bias term b (Yang & Wang, 2020). ANNs in general have proven to be useful for a variety of complex tasks from classification of tumors to automated driving. RNNs can be tuned to accurately simulate time-varying data such as neural firing rates (Pandarinath et al., 2018; Schmidt et al., 2019). Briefly, they consist of an input layer, one or several hidden layers, and an output layer. The RNN then iterates over a set of training data that informs hidden layer weights and adjusts them accordingly until the output of the RNN matches the training data’s inputs. Specific architectures may vary from task to task, however, the generic formulation of RNNs is a powerful, non-linear tool in neuroscience whether they are recreating task dynamics or neural data. Indeed, they have had many successes in describing the underlying dynamics of neural data in the prefrontal cortex which will be further described below. But to what extent does the mPFC exhibit recurrent dynamics empirically?
In order to discuss the recurrent dynamics of the mPFC, a brief discussion of the essential layers is necessary. The mPFC is organized in a columnar manner with layer 1 being the most superficial relative to the surface of the brain whereas layer 5 is considered deep within the cortex. A general model posits that layer 2/3 neurons are involved in controlling the gain of an input (Quiquempoix et al., 2018) while also contextualizing and integrating that input with the organism’s present state. Additionally, it appears critical for the maintenance of persistent activity within cognitive activities such as decision-making or working memory (Goldman-Rakic, 1995; O’Reilly and Frank, 2006; Wang et al., 2013). Layer 5 neurons subsequently output the integrated information to downstream targets while also sending an output signal to layer 2/3 (Crochet, 2009; Sakata & Harris, 2009; Petersen & Crochet, 2013). Together these two layers appear critical for the maintenance of persistent activity in corticocortical loops (Kawaguchi, 2017). The anatomical distribution of neurons within these layers makes recurrent activation of mPFC possible and leads to phenomena such as the persistent activity described above, the maintenance of UDS (Sanchez-Vives & McCormick, 2000), or other rhythmic activity such as delta oscillations (Blaeser et al., 2017). The implications of these types of activity on the features of a neural system will be discussed below.
Understanding Acute EtOH Effects with Computation by Dynamics
DST is a promising tool to determine how EtOH-evoked changes in the biophysical properties of neurons are translated in altered behavioral states as linking these levels requires understanding neural activity at the population level. Neural population data is high-dimensional, noisy, and notoriously difficult to understand. DST allows high-dimensional neural data to be decomposed into low-dimensional dynamical representations in order to understand the essential algorithms a complex system is exhibiting (Sussillo & Barak, 2013). Additionally, it allows experimenters to approach highly complex neural data with tractable a priori hypotheses. Understanding the dynamical properties that neural population dynamics exhibit and how those relate to behavioral outputs is critical to converging on a hypothesis towards acute EtOH’s effects on mPFC.
DST has received vigorous attention recently with several articles reviewing their role in neuroscience (Panzeri et al., 2015; Miller, 2016; Saxena & Cunningham, 2019; Durstewitz et al., 2020; Nadeau, 2020; Vyas et al., 2020). Within this push emerged the idea that understanding the functions of particular brain regions may be gained by assessing how they perform computations via dynamics. This ultimately suggests that variance within a single unit is acceptable so long as the overlayed dynamics of the system remains in-tact. With this push towards computation through dynamics, several important concepts must be laid out. First, the generic formulation of a dynamical system requires three key variables be considered.
| Equation 2 |
Where T describes the timeseries of the system, S describes the state-space of the system, and ϕt describes the operation by which the system evolves (Beer, 2000). This simple three variable formulation can describe both linear and non-linear dynamics. Important for our purposes, DST can approximate how neural trajectories evolve over time. Given an additional generic formulation of DST:
| Equation 3 |
x can represent the change in firing rates of a population of neurons as a function of time, transformed by a generic function f with any external inputs, u (Vyas et al., 2020). Based on how the trajectory of x unfolds, we can then understand the different motifs in neural populations that emerge following behaviors. Examples of DST motifs include attractors, repellers, and saddle points (Vyas et al., 2020) that each may have their own distinct computational properties. Attractors, for example, contain a fixed point in the system that the dynamics flow towards. Repellers, in contrast, contain a point where the dynamics flow away from. Additional descriptions of these motifs can be found in Vyas et al (2020).
The DST approach has been successfully applied to neural population recordings in the motor system. Notably, Churchland and colleagues (2012) found a motif characterized by rotational dynamics that was observed in macaques performing a reaching task reliably related the behavior to the dynamics of population encoding within the motor cortex. This dataset has generated a deep understanding of the neural dynamics of the motor system and has also generated several new approaches to extracting and quantifying dynamical motifs (Pandarinath et al., 2018). One such technique seeks to utilize RNNs in order to generate approximations of low-dimension dynamical systems that the brain may utilize in vivo (Sussillo & Barak, 2013).
Within the mPFC, DST has also received interest. Lapish, Balaguer-Ballester et al (2015) investigated the effects of low and high dose amphetamine on a working memory foraging task. They found that different epochs of the task (training, delay, and test) were encoded in separate state-spaces; amphetamine increased the distance between the encoding of this epoch at the 1.0 mg/kg dose but collapsed differences at the 3.3 mg/kg dose. Additionally, these differences in encoding were reflected in the behavior of the animal. Specifically, the 3.3 mg/kg dose showed deficits in behavioral performance. Additional work has shown a similar effect of amphetamine on effort-based valuations in the mPFC (Hashemnia et al., 2020) wherein a low dose of amphetamine stabilized population dynamics whereas high doses of amphetamine disrupted both the population dynamics and behavior. Interestingly, valuation of effort was encoded within this task wherein low effort trials exhibited a smaller area within the state-space whereas high effort trials occupied a relatively larger area within the state-space. Liu and colleagues have additionally shown similar, stable neural trajectories within working memory tasks in rodents (Liu et al., 2014). Ultimately this suggests that low-dimensional structures are generated in certain task-types that can be modified by pharmacological agents.
Additional studies have shown that evidence accumulation during a visuo-spatial decision-making task exhibits two dynamical motifs, namely, a line attractor and selection vector (Mante et al., 2013). Specifically, macaque monkeys were trained to respond to either a color or motion stimulus depending on specific, learned context cues. From there, they would then have to saccade either towards the dominant color or the direction the motion was moving in. From this, the experimenters observed and recreated the dynamics of the PFC in this task using a RNN. Perturbations to the neural network revealed several fixed points along a line attractor. During choice, a selection vector would perturb neural activity away from the line attractor towards a decision space. It was later found (Aoi et al., 2020) from the same dataset that this decision space integrated evidence from the line attractor and formed similar rotational dynamics to Churchland et al (2012) prior to an action. This suggests the PFC can implement a range of dynamical mechanisms even in this relatively simple task, and the brain is able to flexibly switch between them (Aoi et al., 2020). Within this most recent paper, it was suggested that evidence accumulation occurs along a line attractor exclusively during the early parts of the trial whereas action-selection occurs along signed (i.e. left saccades are represented differentially from right saccades), rotational dynamics in the middle and late portions of the trial (Aoi et al., 2020). An interesting observation was made following errors in one of the monkeys; specifically, Aoi and colleagues observed that during incorrect choices in one of the animals the rotational dynamics were preserved albeit reversed in sign. This suggested that the error occurred due to an incorrect percept rather than misunderstanding the context or being off task. In the other monkey, error signals occurred when the dynamics for a given trial were shifted in time suggesting perhaps that the monkey during these trials was more impulsive or less attentive (Aoi, 2020).
An additional study completed by Márton and colleagues (2020) hypothesized the low-dimensional representations of a task with a RNN and subsequently tested their simulated results on empirical data. This resulted in their group showing both in silico and in vivo that learning of action-outcomes in the PFC is accompanied by segregation of those action-outcomes in state-space. This suggests that as an organism learns, their encoding becomes more efficient and separated in the dynamics of the system in order to minimize errors (Márton et al., 2020).
This set of studies highlights the potential of utilizing DST in identifying computational motifs via dynamics. Moving forward, it will be necessary to continue to leverage recent advances in DST techniques to move from qualitative descriptions of attractor dynamics in neural populations towards quantitative descriptions in order to rigorously describe the attracting landscape of a neural population (Schmidt et al., 2018).
To this end, work has begun on describing potential outcomes in the dynamics of a system based on connectivity features such as synaptic strength and the number of connections a node has (Mastrogiuseppe & Ostojic, 2018; Schuessler et al., 2020). This set of studies showed that low-rank random connectivity, which is a network where each neuron is only sparsely and randomly connected to other neurons, can produce low-dimensional dynamics while persevering the system’s ability to generalize. In contrast to this, the authors note that other connectivity schemes that produce low-dimensional dynamics store information in a more fixed manner. This idea connects well with the fact that mPFC multiplexes information in a contextual and generalizable manner. However, as the authors note, utilizing RNNs in this manner sacrifices biological complexity that may be important to further understanding how low-dimensional dynamics emerge given that RNN activity is continuous and rate-based and not spiking based (Mastrogiuseppe & Ostojic, 2018). Namely, excitatory and inhibitory (E/I) currents may be essential to understanding how biological neural systems are able to efficiently encode information (Denève & Machens, 2016; Sadeh & Clopath, 2020) particularly given that PFC neurons operate in high-conductance regimes (Destexhe et al., 2003) and manipulating either excitatory conductance or NMDA receptor conductance is essential to understanding how acute EtOH disrupts mPFC neural activity (Mastrogiuseppe & Ostojic, 2018).
Important to the present review is the bump attractor, a dynamical motif found during periods of persistent or intrinsic activity within a working memory task, decision-making, or head direction. This motif resembles a “bump” in that activity in a population of neurons non-linearly increases until reaching a fixed maximum and then non-linearly decreases to baseline. The location of the bump attractor itself can vary across the network of neurons, which is exhibited by transient, coordinated increases in subpopulations of neurons across the network. This allows for the encoding of continuous rather than discrete variables such as a visual target’s location in space (Wimmer et al., 2014). Poor behavioral performance can then be linked to inaccurate encoding in the network evidenced by malformed bump attractors and/or bump attractors whose firing rate is highest for the wrong stimulus. Ultimately and most importantly this allows for a tractable explanation of behavior from the low-dimensional structure of a neural population. These errors likely arise due to random fluctuations or distractors in the network (Compte et al., 2000; Wimmer et al., 2014) and future work should examine whether they can be pharmacologically induced by EtOH. This type of population dynamic is critical for persistent activity and was hypothesized (Skaggs et al., 1995) and shown to be (Zhang, 1996) an important component of head-direction related neural responses. Recently, it has been hypothesized and shown to be an important feature of spatial working memory within the primate dIPFC as well (Compte et al., 2000; Wimmer et al., 2014). Bistability of voltage in a single neural population as a result of the interplay between NMDA receptor and GABAergic conductance has been hypothesized to be an important piece of persistent activity (Sanders et al., 2013). Additionally, recurrent excitation and feedback inhibition within a network of neurons appear to be a necessary component of the bump attractor (Compte et al., 2000; Wimmer et al., 2014). A hypothesis of the present review is that NMDA receptor antagonism via acute EtOH may best be described as perturbing this bump attractor which subsequently disrupts behavioral outputs. Specifically, EtOH may disrupt the formation of the bump attractor or disrupt the precise tuning necessary to keep a specific population of orientation-tuned neurons active and robust against distractors.
A separate class of attractors critical to consider are the line attractors given their potential importance for evidence accumulation and integration in the PFC (Mante et al., 2013; Aoi et al., 2020). A line attractor is a specific class of continuous attractors that consists of an infinite set of fixed points organized along a line in the state-space. This organization allows for the accumulation of information that is robust to small perturbations in the system. A true line attractor thus exhibits properties that are essential for persistent activity. Line-attractors have been particularly successful at describing eye-movement (Seung, 1996). Specifically, Seung (1996) describes the necessity of maintaining a memory of eye-position while the eye saccades from target-to-target. They then posit that this memory is represented via persistent activity of a population of neurons that represents the position of the eye. Without perturbation, the population dynamics lie on a line attractor whereas during a saccade the dynamics move away from the line attractor and decay at some rate back to it. Seung (1996) subsequently argues that for a line attractor to form within a biological system, the system must be precisely tuned from a set of learning rules. Additionally, they argue that the persistence of a line attractor is fleeting given that it is an unstable structure and can be readily disrupted by noise and distractors. In order to maintain the line attractor, positive feedback or recurrent activity is essential. As described above, the line attractor has also been utilized to describe neural activity within the PFC related to decision-making (Mante et al., 2013; Aoi et al., 2020). In this case, the function of this particular dynamic is to integrate information about the task prior to a decision (Brody, 2016). The line attractor is particularly well suited to decision-making tasks because its dynamics suggest that it can flexibly integrate multiple streams of information and subsequently be able to maintain that information over time and contextual shifts all while maintaining robustness against perturbations (Brody et al., 2003; Mante et al., 2013; Aoi et al., 2020).
Ultimately, by establishing computational models of acute EtOH effects, we can observe how the dynamics a given network of neurons exhibits change after introducing EtOH-relevant manipulations. For example, networks of neurons with strong excitatory and inhibitory weights may exhibit discrete Hopfield attractors wherein memories are stored in basins of attraction learned by the network after several iterations. The weights for each synapse are adjusted incrementally until a strong basin of attraction is formed for each input stimulus; each input stimulus can then be stored by the system of attractors (Hopfield, 1982). An EtOH-relevant manipulation may disrupt the learning and formation of these attractors which would subsequently produce poor memory storage. Another example includes networks of neurons loosely connected, and these may exhibit transient, feed-forward dynamics (Chaudhuri & Fiete, 2016) that could be disrupted by EtOH. Particularly interesting is testing how alterations produced via acute EtOH impacts low-dimensional dynamical representations such as bump or line attractors and persistent activity. These low-dimensional dynamics likely represent an algorithmic schema that can be implemented by a neural system in order to produce behavior (Sussillo & Barak, 2013) and understanding how disruptions to those schemas arise will be an essential bridge from understanding how acute EtOH affects neural activity thus resulting in changes in behavior. While some of this work has been done previously (Compte et al., 2000), a refined focus on both acute EtOH effects and specific DST motifs will be illuminating. Importantly, this will then allow for testable hypotheses to emerge that can be challenged empirically in the future.
Demonstrating the Effects of Acute EtOH on a Population Rate Model
To illustrate the conceptual analysis performed thus far, we adapted and recreated a firing rate population model from Jercog and colleagues (2017). The model is capable of oscillating between up-down states similar to what is observed in cortex during SWS or anesthesia. Specifically, they tested the contributions of both external fluctuations and internal adaptation currents on up-down state transitions. As discussed above, Boikov et al (2020) suggests that EtOH enhances CDD of NMDA receptors. Specifically, in the presence of EtOH (22 mM), lower calcium concentrations are required to elicit CDD. A mechanism that they suggest to be responsible for this enhancement is the disruption of NCX which may lead to slower clearance of calcium from the intracellular space. This subsequently prolongs the CDD period leading to longer periods of NMDA receptor inactivation. With this in mind, we chose to alter the decay rate of the adaptation current within the chosen population rate model in order to mimic this prolonged CDD period (Jercog et al., 2017). For their population rate model, Jercog and colleagues (2017) utilized the following set of equations:
| Equation 4 |
| Equation 5 |
| Equation 6 |
| Equation 7 |
A full explanation of the parameters utilized in equations 4–7 can be found in Jercog (2017). In brief, rE represents the excitatory rates of system, rI represents the inhibitory rates of the system, and the variable a represents the adaptation current. The nonlinear function φx(x) determines the steady state firing rate for rE and rI (Equation 6; Figure 1A).
Figure 1. Reproduction of the model and results of EtOH-relevant manipulation on non-noised conditions without noise.
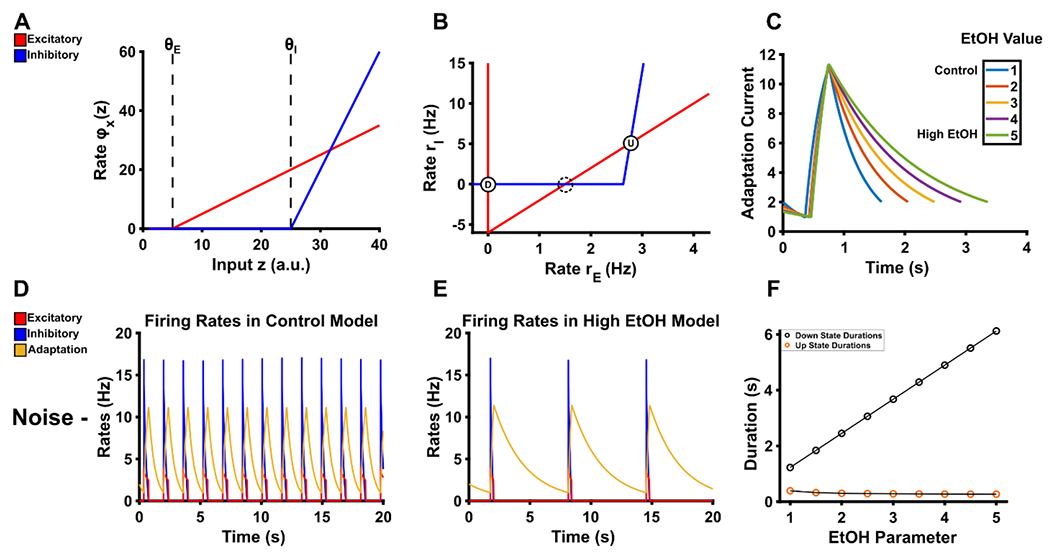
A. The response of the excitatory and inhibitory units is shown as a function of excitatory input. A non-linearity in the model leads to input values below the threshold θX to be output as 0. After the input surpasses the threshold θX, the output is a steady state firing rate of each respective population according to the gain term gE or gI. B. The phase plane of rates demonstrates three possible fixed points in the fast subsystem. The down-state is the stable fixed point where both the excitatory and inhibitory rates are at 0, which is denoted by the circle D. The up-state is the stable fixed point where both excitatory and inhibitory rates are non-zero, which is denoted by the circle U. The dashed circle at the 2nd intersection indicates an unstable fixed point. C. Adjusting the EtOH parameter produces longer decay times for the adaptation current of this model. D. In the absence of noise and with EtOH equal to 1, up-down states proceed at regular intervals. E. With EtOH equal to 5, down-states are significantly prolonged due to the longer decay times of the adaptation current. F. In the absence of noise, down-state durations linearly increase as a function of our EtOH parameter. Up-state durations are only minorly affected.
The existence and position of the attractors that correspond to the up-state and down-state are determined by the intersection of the nullclines for variables rE and rI (Figure 1B), and the nullcline for rE is dependent on the slow adaptation current a (Equations 4 and 7). Over the course of an up-state, a accumulates, and the steady state firing rate of rE adapts until the attractor that corresponds to the up-state vanishes. Thus the model transitions from up-state to down-state. During the down-state, a recovers. The steady state firing rate for rE is facilitated such that the attractor for the up-state reappears. Subsequently, a continues to recover until the steady state firing rate for rE is sufficiently facilitated, which abolishes the attractor for the down-state, and the model transitions from down-state to up-state (see Jercog et al., 2017 for details).
We observe decreases in upstate durations and increases in down-state duration in both ex vivo and in vivo preparations (Tu et al., 2007; Morningstar et al., 2020) and it is also observed that EtOH disrupts CDD in a concentration dependent manner likely leading to longer periods of NMDA receptor inactivity (Boikov et al., 2020). To model this, we adjusted the decay rate of our adaptation current which subsequently increases the duration of down-states. We accomplished this by dividing the decay rate by the fixed constant EtOH. Larger values of EtOH slow the decay rate of the adaptation current in a linear fashion. Thus we adjusted Equation 7 such that:
| Equation 8 |
The inclusion of the unitless term EtOH allows us to vary the decay rate of the adaptation current such that down-states persist longer when EtOH takes on any value greater than 1. The effect generated from the inclusion of the EtOH term can be seen in Figure 1. Figure 1C shows the result of our manipulation on the adaptation current such that higher values of EtOH result in longer decay times. Figure 1D–F show the results of our manipulation at EtOH values equal to 1 (control model) and 5 (high EtOH model). Notably, the up-states of excitatory and inhibitory populations in the absence of noise are shortened as a function of EtOH. Figure 2A–C show the results of our manipulations in the presence of fluctuating noise while Figure 2 D and E show empirical results adapted from Morningstar et al (2020). Importantly the modeled results show a similar increase in down-state duration compared with the empirical results. Specifically, in the empirical results we saw a median down-state duration of 0.676 s in the saline condition and a median down-state duration of 0.972 s in the 1.0 g/kg EtOH condition. The modeled results show a median down-state duration of 1.024 s and 1.025 s when the EtOH parameter equals 2.5 or 3.0, respectively, whereas in the control condition the median down-state duration is 0.348 s. Ultimately this suggests that a small change in the mechanism responsible for the transitions between up-states and down-states is sufficient to produce dynamics similar to our empirical findings with 1.0 g/kg EtOH. Table 2 shows the parameter sets of each respective manipulation and additional simulation details. Our full code adapted from Jercog et al (2017) can be found at https://github.com/mdmornin/etoh_attractordynamics_review.
Figure 2. Model results during noised conditions with noise.
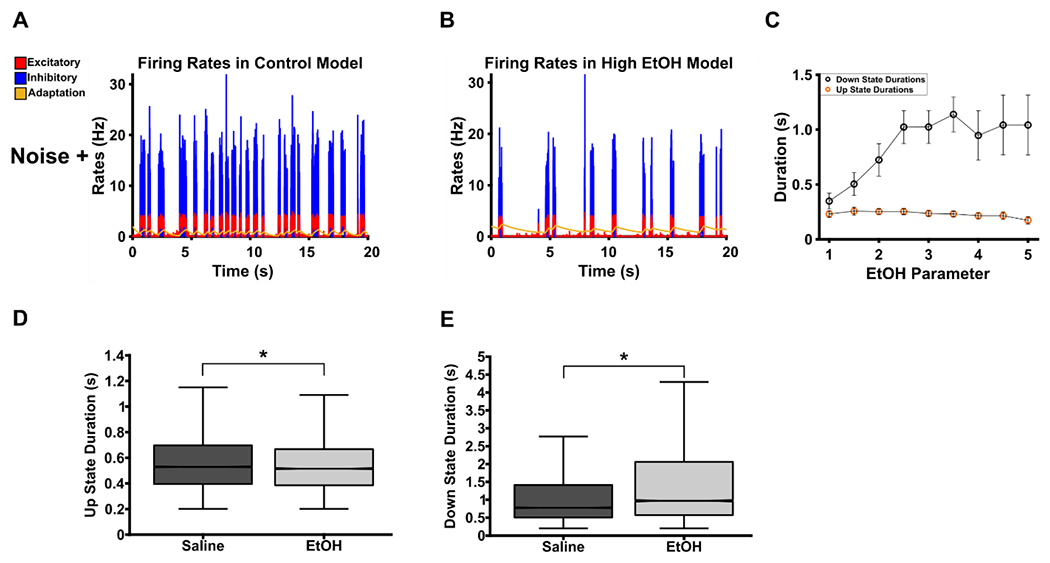
A. With noise added to the system, up-down states proceed irregularly for variable periods of time. B. With EtOH equal to 20, noise is sufficient to drive up-states that did not occur in the non-noise condition, however, down-states are on average considerably lengthened. C. Down-states increase as a function of EtOH in the noise condition. Additionally, down-state durations are considerably shorter compared to comparable EtOH values in the non-noise condition. Values depicted are median ± SEM. D and E are adaptations from Morningstar (2020). These are empirical results during a urethane anesthetized recording before and after a 1.0 g/kg EtOH injection. D shows a small, significant decrease in up-state duration following EtOH (Median Saline: 0.524 ± 0.664; Median EtOH: 0.515 ± 0.240; Wilcoxon’s rank-sum: z = −6.55, p < 0.001, Effect Size = −0.04) and E shows a larger, significant increase in down-state duration following EtOH (Median Saline: 0.676 ± 1.456; Median EtOH: 0.972 ± 2.007; Wilcoxon’s rank-sum: z = 14.69, p < 0.001, Effect Size = 0.08). Median values are followed by STD. Effect size was calculated by dividing the z-score by the square root of the total number of up or down-states tested.
Table 2.
Description of model parameters used in each instance.
| β | JEE | JEI | JIE | JII | θE | θI | gE | gI | τE | τI | τA | τN | σ | μ | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| − Noise | 7 | 5 | 1 | 10 | 0.5 | −1 | 25 | 1 | 4 | 10 | 2 | 500 | NA | NA | NA |
| + Noise | 0.7 | 5 | 1 | 10 | 0.5 | 4.8 | 25 | 1 | 4 | 10 | 2 | 500 | 1 | 3.5 | 0 |
| Phase Diagrams | NA | 5 | 1 | 10 | 0.5 | 5 | 25 | 1 | 4 | 10 | 2 | 500 | NA | NA | NA |
The parameters for both the noise and non-noise simulations are described above. The adaptation strength, β, is varied between conditions. JEE, JEI, JIE, JII describe the connection strengths between populations and does not vary. ΘE is the threshold necessary to overcome the non-linearity for the excitatory population whereas θI is the threshold necessary to overcome for the inhibitory populations, gE and gI are the gain values applied to the rates, τ values are the time constants for their respective populations (E: excitatory, I: inhibitory, A: adaptation, N: noise), σ is the variance value used for each independent Ornstein-Eulenbecker process that generated noise, μ is the mean value used to generate noise. All simulations were carried out in MATLAB2019b using a custom 4th order Runge-Kutta function with 1 ms time steps. Initial conditions were as follows: [rE 0, rI: 0, a: 2]. The parameter EtOH ranged in value from 1 to 5 with 0.5 steps.
The ability to alternate between sets of attractors is a critical feature thought to underly neural computation (Finkelstein et al., 2021). The system above presents this issue from the perspective of a relatively simple 2-attractor system: up-states and down-states. However, the critical point of the above system is that a manipulation that has physiological relevance to EtOH’s antagonistic properties on NMDA receptors can produce disruptions in the system’s ability to switch between these two attractors. Critically, if we extend similar alterations to systems that exhibit bump or line attractors, we hypothesize that similar deleterious effects on the organization and transition between attractor states. Subsequently characterizing how these bump or line attractors are disorganized should aid in understanding how acute EtOH exerts its effects on cognition. From this, one could hypothesize that during perceptual decision-making processes, where ensembles of neurons integrate information along separable line attractors (Mante et al., 2013; Aoi et al., 2020), shifting rules in the task may require more attempts due to the system being unable to flexibly alternate between attractors. Additionally, in the context of working memory, it is critical to maintain the representation of an external stimuli in the absence of said external stimuli. As mentioned above, this often results in the formation of a bump attractor wherein a population of neurons tuned to the stimulus maintain a representation of it over a delay period (Compte et al., 2000; Wimmer et al., 2014). Acute EtOH may then disrupt the precise tuning that is facilitated a bump attractor via lowering the threshold necessary for successful distractors ultimately resulting in spurious behavioral choices. Additionally, it may disrupt the ability of the system to maintain elevated population activity over delay periods resulting in shorter, easier delays being unaffected whereas longer delays would result in performance deficits (Givens & McMahon, 1997). Ultimately neither hypothesis may be correct and requires further experimentation, however, understanding neural population dynamics through DST creates a tractable, low-dimensional proving ground for neural data. Furthermore, understanding the effects of drugs of abuse and psychiatric disorders on these low-dimensional representations presents ripe clinical opportunity (Durstewitz et al., 2020).
Conclusion
Acute EtOH intoxication is a persistent hazard to society resulting in traffic accidents, injury, and violence. This may be partially due to acute EtOH’s impairing effects on mPFC function, which in turn impairs decision-making, judgement, cognitive control, and memory. The NMDA receptor appears critically and directly involved in the disruptive effects of EtOH on mPFC-related behaviors. The effects of EtOH may be mediated via disruptions to NMDA receptor conductance, channel openings, and/or decay times. These disruptions may result in an impaired ability to buffer the effects of distracting stimuli on recurrent activity in mPFC populations thus leading to impairments in working memory. Characterizing these population-level phenomena, and how they are impaired by EtOH, is facilitated by low-dimensional state space approaches.
The results of the simulations presented herein suggest that EtOH disrupts the ability of a simple 2 attractor system to alternate between up and down states. This was accomplished by examining the effects of EtOH on low dimensional dynamics in mPFC. Low-dimensional dynamics may provide a qualitative description of the algorithmic schemas implemented in the brain to guide behavior. Therefore, simulating acute EtOH by altering properties relevant to NMDA receptor function and observing the effects on low-dimensional dynamics provides insight into how changes in the biophysical properties of neurons alter computations that occur at the population level. Here, we hypothesize that the effects of EtOH on NMDA receptors in a simple 2 state attractor system assessed herein can be extended to more detailed models that are capable of more complex dynamics.
Investigating acute EtOH’s effects on emergent, low-dimensional dynamics such as line or bump attractors via altering NMDA receptor properties has the potential to provide clarity on how acute EtOH intoxication impacts the mPFC during complex, cognitive behaviors. As reviewed previously, line attractors have been suggested to be critical for the short-term temporal integration of information. We hypothesize that the effects of EtOH on NMDA prevent increases in neural firing and therefore may restrict the movement of neural activity along a line attractor. This would be expected to lead to an impaired ability to integrate evidence or deliberate options.
In bump attractors, it is hypothesized that their formation will be impaired with intoxicating doses of EtOH. Givens & McMahon (1997) shows that non-spatial working memory is impaired following a dose of 0.75 g/kg. Notably, the persistent activity observed during the delay period in the control conditions is non-existent suggesting that a bump-attractor was unable to be formed. In lower doses of EtOH, the specificity of the bump-attractors encoding may be disrupted while also increasing susceptibility to distractors as is seen in Compte et al (2000) when NMDA function is disrupted. Testing these hypotheses in silico is a necessary next step that will subsequently drive future empirical hypotheses. Specifically, a finer grained approach can be undertaken by utilizing biophysically relevant spiking networks of neurons. This will allow testing of both network effects and the effects of NMDA antagonism more directly.
The impact of EtOH on behavior is hypothesized to be particularly at its greatest during periods of learning, working memory, or where error-monitoring and performance adjustments are required (Anderson et al., 2011; Bartholow et al., 2012; Bailey et al., 2014; Benson et al., 2019). For instance, if error-monitoring provides a signal that moves neural activity from one attractor state to the next, disruptions to that signal will likely result in perseverative errors. Additionally, if memory reconsolidation is dependent on mechanisms such as STP and persistent activity (Barbosa et al., 2020), EtOH likely disrupts the persistent activity processes necessary for memory reactivation. Utilizing this framework, a dual-pronged approach between in silico and in vivo experiments can be tested. First, we can test these hypotheses immediately in silico to inspire the design of experimental studies. A specific, proposed experiment based on our modeling results would be to utilize an inter-session block schedule where animals are required to respond appropriately to stimulus X in block 1 and stimulus Y in block 2. These blocks should be pseudo-random and designed to require the animal to utilize error-monitoring signals to adjust performance. Failures to adjust performance should come after acute EtOH, resulting in more errors after block-switches but not necessarily within blocks. We can conceptualize performance in this task requiring attractor states per given behavioral rule; these attractor states must subsequently be flexibly created and destroyed in order to encode rule swaps at the beginning of each new block. EtOH likely disrupts the flexibility to swap between attractor states, as evidenced by our modeling results, subsequently resulting in the generation of errors.
It is necessary to discuss several limitations of the current model. The present review focused on localized changes in mPFC neural activity following acute EtOH. However, this is only an initial step towards a complete account capable of detailing how the effects of acute EtOH on mPFC neural activity impair behavior. Future work should focus on corticostriatal, corticothalamic, and corticocerebellar loops. The contributions of other receptor systems, such as GABA and AMPA, as well as considering the impact of neuromodulatory systems such as dopamine and norepinephrine will also be critical to consider. The present review focused heavily on NMDA receptors given that they are most directly affected in the mPFC at pharmacologically relevant levels of EtOH (Weitlauf, 2008). However, in other brain regions this is not the case. Additionally, the present review assumes a naivety to alcohol within the subject. Long-term exposure to EtOH in various patterns of consumption are certainly critical for understanding how acute EtOH results in cognitive changes given the widespread molecular and anatomical changes chronic EtOH may induce. Furthermore, the age of the subject is an important consideration given that NMDA subunit expression changes over development (Roberts & Ramoa, 1999), however, further modeling studies will be necessary to elucidate these questions using spiking neural networks of biophysically plausible units that incorporate multiple NMDA subunit types, multiple neuromodulatory populations, and plastic synapses.
Finally, while the present review focused extensively on EtOH, this approach should be applicable to understanding how other drugs of abuse influence cognitive function. Tractable, low-dimensional representations of high-dimensional neural data is essential to progress in the field of neuroscience – particularly within the domain of cognition where encoding of representations is not always a 1-to-1 relationship (Barak & Krakauer, 2021). Understanding how drugs of abuse influence cognition will require understanding how the milieu biophysical changes they evoke are aggregated en masse into changes in neural population-level dynamics. We contend that an effective way to address these questions is for future work to tightly integrate theoretical and computational neuroscience approaches with experimental neuroscience approaches.
Table 1.
Observed behavioral and neurophysiological changes associated with acute EtOH intoxication that have been presently reviewed.
| EtOH Dose (g/kg) | Neurophysiological Changes | References (Neurophysiological) | Behavioral Changes | References (Behavioral) |
|---|---|---|---|---|
|
| ||||
| 0.0 – 0.5 | Decreased theta-alpha band synchronization in working memory (WM) |
Sanz-Martin (2014)
Boha (2009) Molnar (2009) |
||
| Reduced fronto-parietal theta power in WM | Sanz-Martin (2014) | |||
|
| ||||
| 0.5 – 1.0 | Diminished frontal error-monitoring |
Nelson (2011)
Kovacevic (2012) Beaton (2018) |
Impaired WM |
Givens (1998)
Finn (1999) Weissenborn (2003) Gundersen (2008) Saults (2007) |
| NMDA antagonism |
Weitlauf (2008)
Badanich (2013) |
|||
|
Lengthening of down-states |
Tu (2007) Morningstar (2020) |
Increased speed-accuracy tradeoff |
Tiplady (2001; 2003) Schweizer (2004) |
|
| Suppressed delay period activity | Givens (1998) | Impaired error-correction |
Anderson (2011)
Bailey (2014) |
|
| Impaired stimulus-response times | Anderson (2011) | |||
| Impaired verbal memory | Magrys (2014) | |||
| Reductions in sustained attention | Magrys (2014) | |||
|
| ||||
| 1.0 – 1.5 | Disrupted attentional gating | Chapin (1986) | ||
| Decreased firing-rate (FR) variability | Morningstar (2020) | |||
|
| ||||
| 1.5 – 2.0+ | Decreased place-cell specificity | Matthews (1996) | Deficits in spatial memory | Matthews (1996) |
| Decreased FR during states of low-vigilance | Morningstar (2020) | Deficits in novel object recognition |
Ryabinin (2002)
Marszalek-Grabska (2018) |
|
| Deficits in trace conditioning | Hunt (2009) | |||
Highlights.
A mechanism is proposed for the cognitive impairing effects of acute alcohol
The acute effects of alcohol on mPFC function are hypothesized to be critical
A proof-of-concept model suggests how alcohol may alter mPFC activity from dynamical systems perspective
Acknowledgements:
This work was supported by AA023786 and P60-AA007611.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
The authors declare no competing financial interests.
References
- Acons K, Chan LS, Drummond G, & Tiplady B (2006). Effects of ethanol and promethazine on awareness of errors and judgements of performance. Journal of Psychopharmacology, 20(5), 661–669. 10.1177/0269881106060690 [DOI] [PubMed] [Google Scholar]
- Alexander WH, & Brown JW (2011). Medial prefrontal cortex as an action-outcome predictor. Nature Neuroscience, 14(10), 1338–1344. 10.1038/nn.2921 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson BM, Stevens MC, Meda SA, Jordan K, Calhoun VD, & Pearlson GD (2011). Functional Imaging of Cognitive Control During Acute Alcohol Intoxication. Alcoholism: Clinical and Experimental Research, 35(1), 156–165. 10.1111/j.1530-0277.2010.01332.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aoi MC, Mante V, & Pillow JW (2020). Prefrontal cortex exhibits multidimensional dynamic encoding during decision-making. Nature Neuroscience, 23(November). 10.1038/S41593-020-0696-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Badanich KA, Mulholland PJ, Beckley JT, Trantham-Davidson H, & Woodward JJ (2013). Ethanol reduces neuronal excitability of lateral orbitofrontal cortex neurons via a glycine receptor dependent mechanism. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology, 38(7), 1176–1188. 10.1038/npp.2013.12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey K, Bartholow BD, Saults JS, & Lust SA (2014). Give me just a little more time: Effects of alcohol on the failure and recovery of cognitive control. Journal of Abnormal Psychology, 123(1), 152–167. 10.1037/a0035662 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balaguer-Ballester E, Lapish CC, Seamans JK, & Durstewitz D (2011). Attracting states in frontal cortex networks associated with working memory and decision making. BMC Neuroscience, 12(S1), 1–2. 10.1186/1471-2202-12-s1-p8221208416 [DOI] [Google Scholar]
- Barack DL, Krakauer JW Two views on the cognitive brain. Nat Rev Neurosci (2021). 10.1038/S41583-021-00448-6 [DOI] [PubMed] [Google Scholar]
- Barak O, & Tsodyks M (2007). Persistent activity in neural networks with dynamic synapses. PLoS Computational Biology, 3(2), 0323–0332. 10.1371/journal.pcbi.0030035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbosa J, Stein H, Martinez RL, Galan-Gadea A, Li S, Dalmau J, Adam KCS, Valls-Solé J, Constantinidis C, & Compte A (2020). Interplay between persistent activity and activity-silent dynamics in the prefrontal cortex underlies serial biases in working memory. Nature Neuroscience, 23(8), 1016–1024. 10.1038/S41593-020-0644-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartholow BD, Henry EA, Lust SA, Saults JS, & Wood PK (2012). Alcohol effects on performance monitoring and adjustment: affect modulation and impairment of evaluative cognitive control. Journal of Abnormal Psychology, 121 (1), 173–186. 10.1037/a0023664 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beaton LE, Azma S, & Marinkovic K (2018). When the brain changes its mind: Oscillatory dynamics of conflict processing and response switching in a flanker task during alcohol challenge. PLoS ONE, 13( 1), 1–24. 10.1371/journal.pone.0191200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beer RD (2000). Dynamical approaches to cognitive science. Trends in Cognitive Sciences, 4(3), 91–99. 10.1016/S1364-6613(99)01440-0 [DOI] [PubMed] [Google Scholar]
- Bekolay T, Laubach M, & Eliasmith C (2014). A spiking neural integrator model of the adaptive control of action by the medial prefrontal cortex. Journal of Neuroscience, 34(5), 1892–1902. 10.1523/JNEUROSCI.2421-13.2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benson S, Tiplady B, & Scholey A (2019). Attentional and working memory performance following alcohol and energy drink: A randomised, double-blind, placebo-controlled, factorial design laboratory study. PLoS ONE, 14(1), 1–17. 10.1371/journal.pone.0209239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blaeser AS, Connors BW, & Nurmikko AV (2017). Spontaneous dynamics of neural networks in deep layers of prefrontal cortex. Journal of Neurophysiology, 117(4), 1581–1594. 10.1152/jn.00295.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boha R, Molnár M, Gaál ZA, Czigler B, Róna K, Kass K, & Klausz G (2009). The acute effect of low-dose alcohol on working memory during mental arithmetic: I. Behavioral measures and EEG theta band spectral characteristics. International journal of psychophysiology : official journal of the International Organization of Psychophysiology, 73(2), 133–137. 10.1016/j.ijpsycho.2009.02.006 [DOI] [PubMed] [Google Scholar]
- Boikov SI, Sibarov DA, & Antonov SM (2020). Ethanol inhibition of NMDA receptors in calcium-dependent and –independent modes. Biochemical and Biophysical Research Communications, 522(4), 1046–1051. 10.1016/j.bbrc.2019.12.007 [DOI] [PubMed] [Google Scholar]
- Boikov SI, Sibarov DA, & Antonov SM (2020). Ethanol inhibition of NMDA receptors in calcium-dependent and –independent modes. Biochemical and Biophysical Research Communications, 522(4), 1046–1051. 10.1016/j.bbrc.2019.12.007 [DOI] [PubMed] [Google Scholar]
- Bolkan SS, Stujenske JM, Parnaudeau S, Spellman TJ, Rauffenbart C, Abbas AI, … Kellendonk C (2017). Thalamic projections sustain prefrontal activity during working memory maintenance. Nature Neuroscience, 20(7), 987–996. 10.1038/nn.4568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, & Cohen JD (2001). Conflict monitoring and cognitive control. Psychological review, 108(3), 624–652. 10.1037/0033-295x.108.3.624 [DOI] [PubMed] [Google Scholar]
- Braver TS (2012). The variable nature of cognitive control: a dual mechanisms framework. Trends in cognitive sciences, 16(2), 106–113. 10.1016/j.tics.2011.12.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brette R, & Gerstner W (2005). Adaptive exponential integrate-and-fire model as an effective description of neuronal activity. Journal of Neurophysiology, 94(5), 3637–3642. 10.1152/jn.00686.2005 [DOI] [PubMed] [Google Scholar]
- Brody CD, & Hanks TD (2016). Neural underpinnings of the evidence accumulator. Current opinion in neurobiology, 37, 149–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brody CD, Romo R, & Kepecs A (2003). Basic mechanisms for graded persistent activity: Discrete attractors, continuous attractors, and dynamic representations. Current Opinion in Neurobiology, 13(2), 204–211. 10.1016/S0959-4388(03)00050-3 [DOI] [PubMed] [Google Scholar]
- Cannady R, Nimitvilai-Roberts S, Jennings SD, Woodward JJ, & Mulholland PJ (2020). Distinct region-and time-dependent functional cortical adaptations in C57BL/6J mice after short and prolonged alcohol drinking. ENeuro, 7(3), 1–15. 10.1523/ENEURO.0077-20.2020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanagh JF, Cohen MX, & Allen JJB (2009). Prelude to and resolution of an error: EEG phase synchrony reveals cognitive control dynamics during action monitoring. Journal of Neuroscience, 29(1), 98–105. 10.1523/JNEUROSCI.4137-08.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanagh SE, Towers JP, Wallis JD, Hunt LT, & Kennerley SW (2018). Reconciling persistent and dynamic hypotheses of working memory coding in prefrontal cortex. Nature Communications, 9(1), 1–16. 10.1038/S41467-018-05873-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chapin JK, Sorensen SM, & Woodward DJ (1986). Acute ethanol effects on sensory responses of single units in the somatosensory cortex of rats during different behavioral states. Pharmacology, Biochemistry and Behavior, 25(3), 607–614. 10.1016/0091-3057(86)90149-8 [DOI] [PubMed] [Google Scholar]
- Chaudhuri R, & Fiete I (2016). Computational principles of memory. Nature Neuroscience, 19(3), 394–403. 10.1038/nn.4237 [DOI] [PubMed] [Google Scholar]
- Cherpitel CJ, Ye Y, Bond J, Borges G, Monteiro M, Chou P, & Hao W (2015). Alcohol Attributable Fraction for Injury Morbidity from the Dose-Response Relationship of Acute Alcohol Consumption: Emergency Department Data from 18 Countries. Addiction (Abingdon, England), 110(11), 1724–1732. 10.1111/add.13031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chin VS, Van Skike CE, Berry RB, Kirk RE, Diaz-Granados J, & Matthews DB (2011). Effect of acute ethanol and acute allopregnanolone on spatial memory in adolescent and adult rats. Alcohol, 45(5), 473–483. 10.1016/j.alcohol.2011.03.001 [DOI] [PubMed] [Google Scholar]
- Churchland MM, Cunningham JP, Kaufman MT, Foster JD, Nuyujukian P, Ryu SI, … Shenoy KV (2012). Neural population dynamics during reaching. Nature, 487(7405), 51–56. 10.1038/nature11129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collingridge GL, Kehl SJ, & McLennan H (1983). Excitatory amino acids in synaptic transmission in the Schaffer collateral-commissural pathway of the rat hippocampus. The Journal of physiology, 334, 33–46. 10.1113/jphysiol.1983.sp014478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Compte A, Brunel N, Goldman-Rakic PS, & Wang XJ (2000). Synaptic mechanisms and network dynamics underlying spatial working memory in a cortical network model. Cerebral Cortex, 10(9), 910–923. 10.1093/cercor/10.9.910 [DOI] [PubMed] [Google Scholar]
- Crochet S, & Petersen CC (2009). Cortical dynamics by layers. Neuron, 64(3), 298–300. 10.1016/j.neuron.2009.10.024 [DOI] [PubMed] [Google Scholar]
- Crochet S, Lee SH, & Petersen CCH (2019). Neural Circuits for Goal-Directed Sensorimotor Transformations. Trends in Neurosciences, 42(1), 66–77. 10.1016/j.tins.2018.08.011 [DOI] [PubMed] [Google Scholar]
- Curtis CE, & Lee D (2010). Beyond working memory: The role of persistent activity in decision making. Trends in Cognitive Sciences, 14(5), 216–222. 10.1016/j.tics.2010.03.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dao Duc K, Parutto P, Chen X, Epsztein J, Konnerth A, & Holcman D (2015). Synaptic dynamics and neuronal network connectivity are reflected in the distribution of times in Up states. Frontiers in Computational Neuroscience, 9(July), 1–9. 10.3389/fncom.2015.00096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denève S, & Machens CK (2016). Efficient codes and balanced networks. Nature neuroscience, 79(3), 375–382. 10.1038/nn.4243 [DOI] [PubMed] [Google Scholar]
- Destexhe A, Rudolph M, & Paré D (2003). The high-conductance state of neocortical neurons in vivo. Nature Reviews Neuroscience, 4(9), 739–751. 10.1038/nrn1198 [DOI] [PubMed] [Google Scholar]
- Devenport LD, Hale RL Contributions of hippocampus and neocortex to the expression of ethanol effects. Psychopharmacology 99, 337–344 (1989). 10.1007/BF00445554 [DOI] [PubMed] [Google Scholar]
- Durstewitz D, Huys QJM, & Koppe G (2020). Psychiatric Illnesses as Disorders of Network Dynamics. Biological Psychiatry: Cognitive Neuroscience and Neuroimaging, Figure 1, 1–12. 10.1016/j.bpsc.2020.01.001 [DOI] [PubMed] [Google Scholar]
- Euston DR, Gruber AJ, & McNaughton BL (2012). The role of medial prefrontal cortex in memory and decision making. Neuron, 76(6), 1057–1070. 10.1016/j.neuron.2012.12.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finkelstein A, Fontolan L, Economo MN et al. Attractor dynamics gate cortical information flow during decision-making. Nat Neurosci (2021). 10.1038/S41593-021-00840-6 [DOI] [PubMed] [Google Scholar]
- Finn PR, Justus A, Mazas C, & Steinmetz JE (1999). Working memory , executive processes and the effects of alcohol on Go / No-Go learning: testing a model of behavioral regulation and impulsivity, 465–472. [DOI] [PubMed] [Google Scholar]
- Francis KB, Gummerum M, Ganis G, Howard IS, & Terbeck S (2019). Alcohol, empathy, and morality: acute effects of alcohol consumption on affective empathy and moral decision-making. Psychopharmacology, 236(12), 3477–3496. 10.1007/S00213-019-05314-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franzen EA, & Myers RE (1973). Neural control of social behavior: Prefrontal and anterior temporal cortex. Neuropsychologia, 11(2), 141–157. 10.1016/0028-3932(73)90002-X [DOI] [PubMed] [Google Scholar]
- Funahashi S, Bruce CJ, & Goldman-Rakic PS (1989). Mnemonic coding of visual space in the monkey’s dorsolateral prefrontal cortex. Journal of Neurophysiology, 61(2), 331–349. 10.1152/jn.1989.61.2.331 [DOI] [PubMed] [Google Scholar]
- George S, Rogers RD, & Duka T (2005). The acute effect of alcohol on decision making in social drinkers. Psychopharmacology, 182(1), 160–169. 10.1007/s00213-005-0057-9 [DOI] [PubMed] [Google Scholar]
- Gill TM, Sarter M, & Givens B (2000). Sustained visual attention performance-associated prefrontal neuronal activity: Evidence for cholinergic modulation. Journal of Neuroscience, 20(12), 4745–4757. 10.1523/jneurosci.20-12-04745.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Givens B (1995). Low Doses of Ethanol Impair Spatial Working Memory and Reduce Hippocampal Theta Activity. Alcoholism: Clinical and Experimental Research, 19(3), 763–767. 10.1111/j.1530-0277.1995.tb01580.x [DOI] [PubMed] [Google Scholar]
- Givens B, & McMahon K (1997). Effects of ethanol on nonspatial working memory and attention in rats. Behavioral neuroscience, 111(2), 275–282. 10.1037//0735-7044.111.2.275 [DOI] [PubMed] [Google Scholar]
- Givens B, Williams J, & Gill TM (1998). Cognitive correlates of single neuron activity in task-performing animals: application to ethanol research. Alcoholism, Clinical and Experimental Research, 22(1), 23–31. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9514282 [PubMed] [Google Scholar]
- Glasgow NG, Povysheva NV, Azofeifa AM, & Johnson JW (2017). Memantine and ketamine differentially alter NMDA receptor desensitization. Journal of Neuroscience, 37(40), 9686–9704. 10.1523/JNEUROSCI.1173-17.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman-Rakic PS (1995). Cellular basis of working memory. Neuron, 14(3), 477–485. 10.1016/0896-6273(95)90304-6 [DOI] [PubMed] [Google Scholar]
- Gonda S, Giesen J, Sieberath A, West F, Buchholz R, Klatt O, Ziebarth T, Räk A, Kleinhubbert S, Riedel C, Hollmann M, Hamad MIK, Reiner A, & Wahle P (2020). GluN2B but Not GluN2A for Basal Dendritic Growth of Cortical Pyramidal Neurons. Frontiers in Neuroanatomy, 14(November), 1–18. 10.3389/fnana.2020.571351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gundersen H, Specht K, Grüner R, Ersland L, & Hugdahl K (2008). Separating the effects of alcohol and expectancy on brain activation: An fMRI working memory study. NeuroImage, 42(4), 1587–1596. 10.1016/j.neuroimage.2008.05.037 [DOI] [PubMed] [Google Scholar]
- Hashemnia S, Euston DR, & Gruber AJ (2020). Amphetamine reduces reward encoding and stabilizes neural dynamics in rat anterior cingulate cortex. ELife, 9, 1–22. 10.7554/ELIFE.56755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heilbronner SR, Rodriguez-Romaguera J, Quirk GJ, Groenewegen HJ, & Haber SN (2016). Circuit-Based Corticostriatal Homologies Between Rat and Primate. Biological psychiatry, 80(7), 509–521. 10.1016/j.biopsych.2016.05.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heilbronner SR, Rodriguez-Romaguera J, Quirk GJ, Groenewegen HJ, & Haber SN (2016). Circuit-Based Corticostriatal Homologies Between Rat and Primate. Biological psychiatry, 80(7), 509–521. 10.1016/j.biopsych.2016.05.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Homayoun H, Jackson ME, & Moghaddam B (2005). Activation of metabotropic glutamate 2/3 receptors reverses the effects of NMDA receptor hypofunction on prefrontal cortex unit activity in awake rats. Journal of Neurophysiology, 93(4), 1989–2001. 10.1152/jn.00875.2004 [DOI] [PubMed] [Google Scholar]
- Hopfield JJ (1982). Neural networks and physical systems with emergent collective computational abilities. Proceedings of the National Academy of Sciences of the United States of America, 79(8), 2554–2558. 10.1073/pnas.79.8.2554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt PS, Levillain ME, Spector BM, & Kostelnik LA (2009). Neurobiology of Learning and Memory Post-training ethanol disrupts trace conditioned fear in rats: Effects of timing of ethanol, dose and trace interval duration. Neurobiology of Learning and Memory, 91(1), 73–80. 10.1016/j.nlm.2008.10.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawaguchi Y (2017). Pyramidal cell subtypes and their synaptic connections in layer 5 of rat frontal cortex. Cerebral Cortex, 27(12), 5755–5771. 10.1093/cercor/bhx252 [DOI] [PubMed] [Google Scholar]
- Ketchum MJ, Weyand TG, Weed PF, & Winsauer PJ (2016). Learning by subtraction: Hippocampal activity and effects of ethanol during the acquisition and performance of response sequences. Hippocampus, 26(5), 601–622. 10.1002/hipo.22545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovacevic S, Azma S, Irimia A, Sherfey J, Halgren E, & Marinkovic K (2012). Theta oscillations are sensitive to both early and late conflict processing stages: Effects of alcohol intoxication. PLoS ONE, 7(8), 1–11. 10.1371/journal.pone.0043957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutlu MG, & Gould TJ (2016). Effects of drugs of abuse on hippocampal plasticity and hippocampus-dependent learning and memory: contributions to development and maintenance of addiction. Learning & Memory, 23(10), 515–533. 10.1101/lm.042192.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lapish CC, Balaguer-Ballester E, Seamans JK, Phillips AG, & Durstewitz D (2015). Amphetamine exerts dose-dependent changes in prefrontal cortex attractor dynamics during working memory. Journal of Neuroscience, 35(28), 10172–10187. 10.1523/JNEUROSCI.2421-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lara AH, & Wallis JD (2015). The role of prefrontal cortex in working memory: A mini review. Frontiers in Systems Neuroscience, 9(DEC), 1–7. 10.3389/fnsys.2015.00173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laubach M, Amarante LM, Swanson K, & White SR (2018). What, if anything, is rodent prefrontal cortex? ENeuro, 5(5). 10.1523/ENEURO.0315-18.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leone RM, & Parrott DJ (2019). Acute Alcohol Intoxication Inhibits Bystander Intervention Behavior for Sexual Aggression Among Men with High Intent to Help. Alcoholism, clinical and experimental research, 43(1), 170–179. 10.1111/acer.13920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu D, Gu X, Zhu J, Zhang X, Han Z, Yan W, … Li CT (2014). Medial prefrontal activity during delay period contributes to learning of a working memory task. Science, 346(6208), 458–463. 10.1126/science.1256573 [DOI] [PubMed] [Google Scholar]
- Lundqvist M, Herman P, & Miller EK (2018). Working memory: Delay activity, yes! persistent activity? maybe not. Journal of Neuroscience, 38(32), 7013–7019. 10.1523/JNEUROSCI.2485-17.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay M, Tiplady B, & Scholey AB (2002). Interactions between alcohol and caffeine in relation to psychomotor speed and accuracy. Human psychopharmacology, 17(3), 151–156. 10.1002/hup.371 [DOI] [PubMed] [Google Scholar]
- Magrys SA, & Olmstead MC (2014). Alcohol intoxication alters cognitive skills mediated by frontal and temporal brain regions. Brain and Cognition, 85(1), 271–276. 10.1016/j.bandc.2013.12.010 [DOI] [PubMed] [Google Scholar]
- Malagon-Vina H, Ciocchi S, Passecker J, Dorffner G, & Klausberger T (2018). Fluid network dynamics in the prefrontal cortex during multiple strategy switching. Nature Communications, 9(1), 1–13. 10.1038/s41467-017-02764-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mante V, Sussillo D, Shenoy KV, & Newsome WT (2013). Context-dependent computation by recurrent dynamics in prefrontal cortex. Nature, 503(7474), 78–84. 10.1038/nature12742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marszalek-grabska M, Gibula-bruzda E, Bodzon-kulakowska A, Suder P, & Gawel K (2018). Effects of the Positive Allosteric Modulator of Metabotropic Glutamate Receptor 5 , VU-29 , on Impairment of Novel Object Recognition Induced by Acute Ethanol and Ethanol Withdrawal in Rats, 607–620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Márton CD, Schultz SR, & Averbeck BB (2020). Learning to select actions shapes recurrent dynamics in the corticostriatal system. Neural Networks, 132, 375–393. 10.1016/j.neunet.2020.09.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massa AA, Subramani OS, Eckhardt CI, & Parrott DJ (2019). Problematic alcohol use and acute intoxication predict anger-related attentional biases: A test of the alcohol myopia theory. Psychology of Addictive Behaviors, 33(2), 139–143. 10.1037/adb0000426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massey PV, Johnson BE, Moult PR, Auberson YP, Brown MW, Molnar E, … Bashir ZI (2004). Differential roles of NR2A and NR2B-containing NMDA receptors in cortical long-term potentiation and long-term depression. Journal of Neuroscience, 24(36), 7821–7828. 10.1523/JNEUROSCI.1697-04.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mastrogiuseppe F, & Ostojic S (2018). Linking Connectivity, Dynamics, and Computations in Low-Rank Recurrent Neural Networks. Neuron, 99(3), 609–623.e29. 10.1016/j.neuron.2018.07.003 [DOI] [PubMed] [Google Scholar]
- Matthews DB, Sirnson PE, & Best PJ (1996). Ethanol Alters Spatial Processing of Hippocampal Place Cells: A Mechanism for Impaired Navigation When Intoxicated, 20(2), 404–407. [DOI] [PubMed] [Google Scholar]
- McQuail JA, Beas BS, Kelly KB, Simpson KL, Frazier CJ, Setlow B, & Bizon JL (2016). NR2A-containing NMDARs in the prefrontal cortex are required for working memory and associated with age-related cognitive decline. Journal of Neuroscience, 36(50), 12537–12548. 10.1523/JNEUROSCI.2332-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendoza-Halliday D, Torres S, & Martinez-Trujillo JC (2014). Sharp emergence of feature-selective sustained activity along the dorsal visual pathway. Nature Neuroscience, 17(9), 1255–1262. 10.1038/nn.3785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller EK, Lundqvist M, & Bastos AM (2018). Working Memory 2.0. Neuron, 100(2), 463–475. 10.1016/j.neuron.2018.09.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller P (2016). Dynamical systems, attractors, and neural circuits [version 1; referees: 3 approved], F1000Research, 5, 1–18. 10.12688/F1000RESEARCH.7698.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molnár M, Boha R, Czigler B, Gaál ZA, Benyovszky M, Róna K, & Klausz G (2009). The acute effect of low-dose alcohol on working memory during mental arithmetic: II. Changes of nonlinear and linear EEG-complexity in the theta band, heart rate and electrodermal activity. International journal of psychophysiology: official journal of the International Organization of Psychophysiology, 73(2), 138–142. 10.1016/j.ijpsycho.2009.02.007 [DOI] [PubMed] [Google Scholar]
- Moorman DE, James MH, Mcglinchey EM, & Aston-jones G (2015). Differential roles of medial prefrontal subregions in the regulation of drug seeking. Brain Research, 1628, 130–146. 10.1016/j.brainres.2014.12.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morningstar MD, Linsenbardt DN, & Lapish CC (2020). Ethanol Alters Variability, But Not Rate, of Firing in Medial Prefrontal Cortex Neurons of Awake-Behaving Rats. Alcoholism: Clinical and Experimental Research. 10.1111/acer.14463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motschman CA, Hatz LE, McCarty KN, Merkle EC, Trull TJ, & McCarthy DM (2020). Event-Level Predictors of Alcohol-Impaired Driving Intentions. Journal of studies on alcohol and drugs, 81(5), 647–654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray JD, Jaramillo J, & Wang XJ (2017). Working memory and decision-making in a frontoparietal circuit model. Journal of Neuroscience, 37(50), 12167–12186. 10.1523/JNEUROSCI.0343-17.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadeau SE (2020). Neural Population Dynamics and Cognitive Function. Frontiers in Human Neuroscience, 14(March), 1–21. 10.3389/fnhum.2020.00050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naud R, Marcille N, Clopath C, & Gerstner W (2008). Firing patterns in the adaptive exponential integrate-and-fire model. Biological Cybernetics, 99(4–5), 335–347. 10.1007/s00422-008-0264-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson LD, Patrick CJ, Collins P, Lang AR, & Bernat EM (2011). Alcohol impairs brain reactivity to explicit loss feedback. Psychopharmacology, 218(2), 419–428. 10.1007/s00213-011-2323-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onos KD, Francoeur MJ, Wormwood BA, Miller RLA, Gibson BM, & Mair RG (2016). Prefrontal neurons encode actions and outcomes in conjunction with spatial location in rats performing a dynamic delayed non-Match to position task. PLoS ONE, 11(2), 1–23. 10.1371/journal.pone.0149019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Reilly RC (2006). Biologically based computational models of high-level cognition. Science (New York, N.Y.), 314(5796), 91–94. 10.1126/science.1127242 [DOI] [PubMed] [Google Scholar]
- Pandarinath C, O’Shea DJ, Collins J, Jozefowicz R, Stavisky SD, Kao JC, … Sussillo D (2018). Inferring single-trial neural population dynamics using sequential auto-encoders. Nature Methods, 15(10), 805–815. 10.1038/s41592-018-0109-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pandarinath C, O’Shea DJ, Collins J, Jozefowicz R, Stavisky SD, Kao JC, … Sussillo D (2018). Inferring single-trial neural population dynamics using sequential auto-encoders. Nature Methods, 15(10), 805–815. 10.1038/s41592-018-0109-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panzeri S, Macke JH, Gross J, & Kayser C (2015). Neural population coding: Combining insights from microscopic and mass signals. Trends in Cognitive Sciences, 19(3), 162–172. 10.1016/j.tics.2015.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papouin T, Ladépêche L, Ruel J, Sacchi S, Labasque M, Hanini M, Groc L, Pollegioni L, Mothet JP, & Oliet SHR (2012). Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell, 150(3), 633–646. 10.1016/j.cell.2012.06.029 [DOI] [PubMed] [Google Scholar]
- Petersen CCH, & Crochet S (2013). Review Synaptic Computation and Sensory Processing. Neuron, 78(1), 28–48. 10.1016/j.neuron.2013.03.020 [DOI] [PubMed] [Google Scholar]
- Petralia RS, Wang YX, Hua F, Yi Z, Zhou A, Ge L, Stephenson FA, & Wenthold RJ (2010). Organization of NMDA receptors at extrasynaptic locations. Neuroscience, 167(1), 68–87. 10.1016/j.neuroscience.2010.01.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pezzulo G, Verschure PFMJ, Balkenius C, & Pennartz CMA (2014). The principles of goal-directed decision-making: From neural mechanisms to computation and robotics. Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1655). 10.1098/rstb.2013.0470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preuss TM (1995). Do rats have prefrontal cortex? The rose-woolsey-akert program reconsidered. Journal of cognitive neuroscience, 7(1), 1–24. 10.1162/jocn.1995.7.1.1 [DOI] [PubMed] [Google Scholar]
- Quiquempoix M, Fayad SL, Boutourlinsky K, Leresche N, Lambert C, & Bessaih T (2018). Layer 2 / 3 Pyramidal Neurons Control the Gain of Cortical Output. 10.1016/j.celrep.2018.08.038 [DOI] [PubMed] [Google Scholar]
- Rajan K, Harvey CDD, & Tank DWW (2016). Recurrent Network Models of Sequence Generation and Memory. Neuron, 90(1), 128–142. 10.1016/j.neuron.2016.02.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reilly RCO, & Frank MJ (2006). Making Working Memory Work: A Computational Model of Learning in the Prefrontal Cortex and Basal Ganglia, 328, 283–328. [DOI] [PubMed] [Google Scholar]
- Riaz S, Puveendrakumaran P, Khan D, Yoon S, Hamel L, & Ito R (2019). Prelimbic and infralimbic cortical inactivations attenuate contextually driven discriminative responding for reward, (August 2018), 1–13. 10.1038/S41598-019-40532-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ricker TJ, Nieuwenstein MR, Bayliss DM, & Barrouillet P (2018). Working memory consolidation: Insights from studies on attention and working memory. Annals of the New York Academy of Sciences, 1424(1), 8–18. 10.1111/nyas.13633 [DOI] [PubMed] [Google Scholar]
- Roberts EB, & Ramoa AS (1999). Enhanced NR2A subunit expression and decreased NMDA receptor decay time at the onset of ocular dominance plasticity in the ferret. Journal of Neurophysiology, 81(5), 2587–2591. 10.1152/jn.1999.81.5.2587 [DOI] [PubMed] [Google Scholar]
- Ryabinin AE, Miller MN, & Durrant S (2002). Effects of acute alcohol administration on object recognition learning in C57BL / 6J mice, 71, 307–312. [DOI] [PubMed] [Google Scholar]
- Sadeh S, & Clopath C (2020). Inhibitory stabilization and cortical computation. Nature Reviews Neuroscience. 10.1038/s41583-020-00390-z [DOI] [PubMed] [Google Scholar]
- Sakata S, & Harris KD (2009). Laminar Structure of Spontaneous and Sensory-Evoked Population Activity in Auditory Cortex. Neuron, 64(3), 404–418. 10.1016/j.neuron.2009.09.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez-Vives MV, & McCormick DA (2000). Cellular and network mechanisms of rhythmic recurrent activity in neocortex. Nature neuroscience, 3(10), 1027–1034. 10.1038/79848 [DOI] [PubMed] [Google Scholar]
- Sanders H, Berends M, Major G, Goldman MS, & Lisman JE (2013). NMDA and GABAB (KIR) conductances: The “perfect couple” for bistability. Journal of Neuroscience, 33(2), 424–429. 10.1523/JNEUROSCI.1854-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanz-Martin A, Hernández-González M, Guevara MÁ, Santana G, & Gumá-Díaz E (2014). Effect of alcohol on electrical organisation in the brain during a visuospatial working memory task and its relationship with the menstrual cycle. Revista de neurologia, 58(3), 103–112. [PubMed] [Google Scholar]
- Saults JS, Cowan N, Sher KJ, & Moreno MV (2007). Differential effects of alcohol on working memory: distinguishing multiple processes. Experimental and clinical psychopharmacology, 15(6), 576–587. doi: 10.1037/1064-1297.15.6.576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saxena S, & Cunningham JP (2019). Towards the neural population doctrine. Current Opinion in Neurobiology, 55, 103–111. 10.1016/j.conb.2019.02.002 [DOI] [PubMed] [Google Scholar]
- Schmidt D, Koppe G, Beutelspacher M, & Durstewitz D (2019). Inferring Dynamical Systems with Long-Range Dependencies through Line Attractor Regularization, 1–19. Retrieved from http://arxiv.org/abs/1910.03471 [Google Scholar]
- Scholey AB, Benson S, Neale C, Owen L, & Tiplady B (2012). Neurocognitive and mood effects of alcohol in a naturalistic setting. Human psychopharmacology, 27(5), 514–516. 10.1002/hup.2245 [DOI] [PubMed] [Google Scholar]
- Schuessler F, Dubreuil A, Mastrogiuseppe F, Ostojic S, & Barak O (2019). Dynamics of random recurrent networks with correlated low-rank structure. ArXiv, 2(1), 13111. 10.1103/physrevresearch.2.013111 [DOI] [Google Scholar]
- Schweizer TA, Jolicœur P, Vogel-Sprott M, & Dixon MJ (2004). Fast, but Error-Prone, Responses during Acute Alcohol Intoxication: Effects of Stimulus-Response Mapping Complexity. Alcoholism: Clinical and Experimental Research, 28(4), 643–649. 10.1097/01.ALC.0000121652.84754.30 [DOI] [PubMed] [Google Scholar]
- Seeburg PH, Burnashev N, Köhr G, Kuner T, Sprengel R, & Monyer H (1995). The NMDA receptor channel: molecular design of a coincidence detector. Recent progress in hormone research, 50, 19–34. 10.1016/b978-0-12-571150-0.50006-8 [DOI] [PubMed] [Google Scholar]
- Sessler FM, Hsu F, Felder TN, Zhai J, Lin RC, Wieland SJ, & Kosobud AEK (1998). Effects of ethanol on rat somatosensory cortical neurons, 266–274. [DOI] [PubMed] [Google Scholar]
- Seung HS (1996). How the brain keeps the eyes still. Proceedings of the National Academy of Sciences of the United States of America, 93(23), 13339–13344. 10.1073/pnas.93.23.13339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sibarov DA, & Antonov SM (2018). Calcium-Dependent Desensitization of NMDA Receptors. Biochemistry (Moscow), 83(10), 1173–1183. 10.1134/S0006297918100036 [DOI] [PubMed] [Google Scholar]
- Simpson CA, & Rush CR (2002). Acute performance-impairing and subject-rated effects of triazolam and temazepam, alone and in combination with ethanol, in humans. Journal of Psychopharmacology, 16(1), 23–34. 10.1177/026988110201600102 [DOI] [PubMed] [Google Scholar]
- Skaggs WE, Knierim JJ, Kudrimoti HS, & McNaughton BL (1995). A model of the neural basis of the rat’s sense of direction. Advances in Neural Information Processing Systems, 7(1984), 173–180. [PubMed] [Google Scholar]
- Smothers CT, & Woodward JJ (2016). Differential effects of TM4 tryptophan mutations on inhibition of N-methyl-d-aspartate receptors by ethanol and toluene. Alcohol (Fayetteville, N.Y.), 56, 15–19. 10.1016/j.alcohol.2016.10.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sussillo D, & Barak O (2013). Opening the black box: Low-dimensional dynamics in high-dimensional recurrent neural networks. Neural Computation, 25(3), 626–649. 10.1162/NECO_a_00409 [DOI] [PubMed] [Google Scholar]
- Tiplady B, Drummond GB, Cameron E, Gray E, Hendry J, Sinclair W, & Wright P (2001). Ethanol, errors, and the speed-accuracy trade-off. Pharmacology, biochemistry, and behavior, 69(3-4), 635–641. 10.1016/s0091-3057(01)00551-2 [DOI] [PubMed] [Google Scholar]
- Tiplady B, Hiroz J, Holmes L, & Drummond G (2003). Errors in performance testing: A comparison of ethanol and temazepam. Journal of Psychopharmacology, 77(1), 41–49. 10.1177/0269881103017001691 [DOI] [PubMed] [Google Scholar]
- Touboul J, & Brette R (2008). Dynamics and bifurcations of the adaptive exponential integrate-and-fire model. Biological Cybernetics, 99(4-5), 319–334. 10.1007/s00422-008-0267-4 [DOI] [PubMed] [Google Scholar]
- Tu Y, Kroener S, Abernathy K, Lapish C, Seamans J, Chandler LJ, & Woodward JJ (2007). Ethanol Inhibits Persistent Activity in Prefrontal Cortical Neurons. Journal of Neuroscience, 27(17), 4765–4775. 10.1523/JNEUROSCI.5378-06.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- U.S. Department of Transportation, National Highway Traffic Safety Administration, National Center for Statistics and Analysis, personal communications, Oct. 16, 2018 and Nov. 6, 2019. [Google Scholar]
- van Eden CG, & Uylings HBM (1985). Cytoarchitectonic development of the prefrontal cortex in the rat. Journal of Comparative Neurology, 241(3), 253–267. 10.1002/cne.902410302 [DOI] [PubMed] [Google Scholar]
- van Gaal S, Ridderinkhof KR, Fahrenfort JJ, Scholte HS, & Lamme VAF (2008). Frontal Cortex Mediates Unconsciously Triggered Inhibitory Control. Journal of Neuroscience, 28(32), 8053–8062. 10.1523/JNEUROSCI.1278-08.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Skike CE, Goodlett C, & Matthews DB (2019). Acute alcohol and cognition: Remembering what it causes us to forget. Alcohol, 79, 105–125. 10.1016/J.ALCOHOL.2019.03.006 [DOI] [PubMed] [Google Scholar]
- Vertes RP (2004). Differential projections of the infralimbic and prelimbic cortex in the rat. Synapse, 51(1), 32–58. 10.1002/syn.10279 [DOI] [PubMed] [Google Scholar]
- Volianskis A, France G, Jensen MS, Bortolotto ZA, Jane DE, & Collingridge GL (2015). Long-term potentiation and the role of N-methyl-D-aspartate receptors. Brain Research, 1621,5–16. 10.1016/j.brainres.2015.01.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vonghia L, Leggio L, Ferrulli A, Bertini M, Gasbarrini G, Addolorato G, … Fontana G (2008). Acute alcohol intoxication. European Journal of Internal Medicine, 19(8), 561–567. 10.1016/j.ejim.2007.06.033 [DOI] [PubMed] [Google Scholar]
- Vyas S, Golub MD, Sussillo D, & Shenoy KV (2020). Computation through Neural Population Dynamics. Annual Review of Neuroscience, 43, 249–275. 10.1146/annurev-neuro-092619-094115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vyklicky V, Korinek M, Smejkalova T, Balik A, Krausova B, Kaniakova M, … Vyklicky L (2014). Structure, function, and pharmacology of NMDA receptor channels. Physiological Research, 63(SUPPL.). 10.33549/physiolres.932678 [DOI] [PubMed] [Google Scholar]
- Wang JX, Kurth-Nelson Z, Kumaran D et al. Prefrontal cortex as a meta-reinforcement learning system. Nat Neurosci 21, 860–868 (2018). 10.1038/s41593-018-0147-8 [DOI] [PubMed] [Google Scholar]
- Wang M, Yang Y, Wang CJ, Gamo NJ, Jin LE, Mazer JA, … Arnsten AFT (2013). NMDA Receptors Subserve Persistent Neuronal Firing during Working Memory in Dorsolateral Prefrontal Cortex. Neuron, 77(4), 736–749. 10.1016/j.neuron.2012.12.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang XJ (2001). Synaptic reverberation underlying mnemonic persistent activity. Trends in Neurosciences, 24(8), 455–463. 10.1016/S0166-2236(00)01868-3 [DOI] [PubMed] [Google Scholar]
- Weissenborn R, & Duka T (2003). Acute alcohol effects on cognitive function in social drinkers: their relationship to drinking habits, 306–312. 10.1007/s00213-002-1281-1 [DOI] [PubMed] [Google Scholar]
- Weitlauf C, & Woodward JJ (2008). Ethanol selectively attenuates nmdar-mediated synaptic transmission in the prefrontal cortex. Alcoholism: Clinical and Experimental Research, 32(4), 690–698. 10.1111/j.1530-0277.2008.00625.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wimmer K, Nykamp DQ, Constantinidis C, & Compte A (2014). Bump attractor dynamics in prefrontal cortex explains behavioral precision in spatial working memory. Nature Neuroscience, 17(3), 431–439. 10.1038/nn.3645 [DOI] [PubMed] [Google Scholar]
- Woodward JJ, & Pava MJ (2009). Effects of ethanol on persistent activity and up-states in excitatory and inhibitory neurons in prefrontal cortex. Alcoholism: Clinical and Experimental Research, 33(12), 2134–2140. 10.1111/j.1530-0277.2009.01059.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright JM, Peoples RW, & Weight FF (1996). Single-channel and whole-cell analysis of ethanol inhibition of NMDA-activated currents in cultured mouse cortical and hippocampal neurons. Brain research, 738(2), 249–256. 10.1016/s0006-8993(96)00780-9 [DOI] [PubMed] [Google Scholar]
- Yang GR, & Wang XJ (2020). Artificial Neural Networks for Neuroscientists: A Primer. Neuron, 107(6), 1048–1070. 10.1016/j.neuron.2020.09.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zamudio PA, Smothers TC, Homanics GE, & Woodward JJ (2020). Knock-in Mice Expressing an Ethanol-Resistant GluN2A NMDA Receptor Subunit Show Altered Responses to Ethanol. Alcoholism: Clinical and Experimental Research, 44(2), 479–491. 10.1111/acer.14273 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K (1996). Representation of spatial orientation by the intrinsic dynamics of the head-direction cell ensemble: A theory. Journal of Neuroscience, 16(6), 2112–2126. 10.1523/jneurosci.16-06-02112.1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Yan W, Wang W, Fan H, Hou R, Chen Y, … Li CT (2019). Active information maintenance in working memory by a sensory cortex. ELife, 8, 1–29. 10.7554/eLife.43191.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Y, Ren H, & Peoples RW (2016). Intersubunit interactions at putative sites of ethanol action in the M3 and M4 domains of the NMDA receptor GluN1 and GluN2B subunits. British Journal of Pharmacology, 173(12), 1950–1965. 10.1111/bph.13487 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zorumski CF, Yang J, & Fischbach GD (1989). Calcium-dependent, slow desensitization distinguishes different types of glutamate receptors. Cellular and Molecular Neurobiology, 9(1), 95–104. 10.1007/BF00711446 [DOI] [PMC free article] [PubMed] [Google Scholar]