

Abstract
Most individuals living with HIV in the USA are over 45 years old and are vulnerable to the combined effects of HIV and aging. Antiretroviral therapies reduce HIV morbidity and mortality but do not prevent HIV trans-activator of transcription (Tat) protein expression or development of HIV-associated neurocognitive disorder (HAND), which may be caused by Tat. Tat-transgenic (Tat-tg) mice are used to study Tat’s effects, typically after transgene induction with doxycycline. However, uninduced Tat-tg mice experience transgene leak and model aspects of HAND when aged, including neuroinflammation. We used in vivo 9.4-tesla proton magnetic resonance spectroscopy to compare neurochemistry in aged versus young female and male uninduced Tat-tg mice. Aged Tat-tg mice demonstrated measurable tat mRNA brain expression and had lower medial prefrontal cortex (MPFC) GABA, glutamate, and taurine levels and lower striatal GABA and taurine levels. Females had lower MPFC glutathione and taurine and lower striatal taurine levels. Brain testosterone levels were negatively correlated with age in aged males but not females. Aged mice had cortical abnormalities not previously reported in aged wild-type mice including lower MPFC GABA and taurine levels. As glutathione and taurine levels reflect inflammation and oxidative stress, our data suggest that Tat may exacerbate these processes in aged Tat-tg mice. However, additional studies in controls not expressing Tat are needed to confirm this point and to deconvolve individual effects of age and Tat expression. Sex steroid hormone supplements, which counter climacteric effects, increase taurine levels, and reduce inflammation and oxidative stress, could attenuate some of the brain abnormalities we identified in aged Tat-tg mice.
Supplementary Information
The online version contains supplementary material available at 10.1007/s11357-021-00354-w.
Keywords: Brain, Estradiol, Glutathione, HIV, Inflammation, Magnetic resonance spectroscopy, Oxidative stress, Sex difference, Tat, Taurine, Testosterone
Introduction
In 2018, nearly 25% of new HIV cases in the USA occurred in people over 45 years old, and of those currently infected, more than half are older than 50 [1]. Thus, most people in the USA living with HIV are older and potentially vulnerable to the concurrent effects of HIV and aging [2]. The use of combined antiretroviral therapy (cART) substantially reduces morbidity and mortality associated with HIV but cART does not prevent the development of HIV-associated neurocognitive disorder (HAND), which is highly prevalent and is associated with mood and cognitive impairments that reduce quality of life [3]. Reservoirs of HIV endure in the brain despite cART and continue to produce and release soluble HIV proteins, even when viral replication is nearly eliminated [3]. One such protein, trans-activator of transcription (Tat), greatly potentiates HIV replication and may be critical in the reactivation of latent HIV reservoirs [4] as Tat protein expression persists and can even increase within the brain during cART [5–7].
The importance of Tat in HIV pathophysiology is demonstrable given that its infusion into animals or its expression in Tat-transgenic (Tat-tg) mice leads to the development of mood and cognitive impairments that recapitulate aspects of HAND [8–17]. Tat protein is suspect because it induces excitotoxicity [18], oxidative stress [14, 19, 20], inflammation [6, 8, 14, 21–25], and mitochondrial dysfunction [19, 26–28], all of which are associated with HAND.
Little is known about the effects of Tat protein in the aged brain, but recent studies in aged Tat-tg mice indicate that Tat accelerates age-related hormonal, physiological, neural, and cognitive decline [15, 17, 24], effects similar to those reported in older individuals living with HIV [29–32]. Prolonged Tat transgene “leak” in Tat-tg mice leads to the accumulation of Tat protein levels that are sufficient to induce brain inflammation, synaptic and axonal abnormalities, and ventricular volume increases over the course of a year [24]. Accordingly, Tat transgene leak in Tat-tg mice may model some aspects of Tat expression occurring in HIV patients on cART and thus merits study. Further, because Tat’s deleterious effects may be enhanced by the presence of other HIV viral proteins [33, 34] or by cART medications [34–37], it is important to selectively study the effects of Tat in animal models unconfounded by other HIV proteins or cART medications, as part of a comprehensive strategy to better understand and treat HIV.
Presently, we used ultra-high-field (9.4 Tesla) in vivo proton magnetic resonance spectroscopy (MRS) to evaluate the effects of aging on cerebral metabolites in Tat-tg mice. In vivo MRS is noninvasive and thus enables longitudinal studies in animals, including in mice that are genetically similar to Tat-tg mice, to assess effects of aging [38]. MRS also facilitates within-subject longitudinal studies of the effects of potential interventions designed to target Tat or mitigate its effects. We previously used in vivo MRS to detect frontal cortex metabolite abnormalities indicative of oxidative stress in young adult male Tat-tg mice induced to express Tat protein for 7 days via doxycycline (Dox) administration [14]. In the present study, we used similar in vivo MRS methods in uninduced Tat-tg mice to examine the effects of aging and sex in frontal cortex and striatum, brain areas known to be affected by in vivo Tat exposure [14, 17, 20, 23, 39–45]. The Tat-tg mice we used contain an astrocyte-specific glial fibrillary acidic protein (GFAP) Dox-inducible promotor that controls expression of HIV-Tat1-86 protein [46, 47]. However, in the absence of Dox administration, another strain of inducible Tat transgenic mice, the rtTA strain, is exposed to Tat via Tat transgene leak and Tat mRNA and protein levels accumulate at high levels sufficient to promote the development of brain structural abnormalities as well as inflammation [24]. We hypothesized that we would detect MRS metabolite abnormalities in aged uninduced Tat-tg mice indicative of oxidative stress and inflammation.
Methods
Subjects
Male and female GFAP/Tat-tg bigenic “Tat-tg” mice backcrossed at least 7 generations to a C57BL/6J mouse line [46, 47] were bred and maintained in the Animal Care Facility at McLean Hospital. After weaning, mice were segregated by sex and were group-housed (3–4/cage). Mice were maintained on a 12:12-h light/dark cycle and were provided with ad libitum access to food and water. All experimental procedures were reviewed and approved by the McLean Hospital Institutional Animal Care and Use Committee and were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication Number 85-23). Adult mice aged 13–14 and 63–89 weeks (Table 1) were scanned once each in this cross-sectional cohort study. One female mouse (63 weeks old at the time of scanning) had been a breeder and had produced 4 litters). While breeding could have influenced her endocrine status [48], brain estradiol levels for this mouse were below the level of detection. Mice were not administered Dox or any other treatment in this study, which aimed to evaluate effects of aging and sex on MRS metabolites.
Table 1.
Study subject demographics
| Age group | ♀ (mean, weeks ± SD; N) | ♂ (mean, weeks ± SD; N) |
|---|---|---|
| Young | 13.3 ± 0.5; 10 | 13.0 ± 0.0; 10 |
| Old | 84.1 ± 8.5; 13 | 82.1 ± 8.3; 10 |
Magnetic resonance imaging and magnetic resonance spectroscopy
Anesthesia
Mice were anesthetized with 1–2% isoflurane and their vital signs, including rectal temperatures, heart-rates, respiration rates, and ECG, were continuously monitored (Small Animal Instruments, Inc., Stony Brook, NY). Body temperature and respiration rates were maintained by adjusting isoflurane levels as well as circulating water bath heating pad and warm air blower temperatures.
Imaging
Magnetic resonance imaging and spectroscopy (MRI/MRS) scans were acquired with a 9.4 tesla (T) horizontal bore system equipped with a Varian VNMRJ direct-drive console (Agilent, Santa Clara, CA), using a 12-cm (inner diameter) gradient. A custom designed and built (Life Services, LLC, Minneapolis, MN) quadrature butterfly surface coil was placed over the head. Turbo fast low-angle shot (TurboFlash) scout MRI images were acquired to position medial prefrontal cortex (MPFC) and left striatal voxels (Fig. 1, inset). MPFC and striatal MRS voxel shimming and water suppression were performed using the FASTMAP and VAPOR approaches, respectively [49, 50]. Localized proton (1H) spectra (MPFC: 1.2 × 2.5 × 2.5 mm=7.5 mm3; left striatum: 2.0 × 2.0 × 1.6 mm=6.4mm3) were acquired with the LASER spectroscopy sequence [51] using a repetition time (TR)/echo time (TE) = 3000/19 ms. Bins of free induction decays (FIDs, every 8 scans) were separately acquired and stored automatically by the console and then corrected for frequency and phase shifts due to physiological or hardware variations. In total, 480 scans were acquired (24 min) for each voxel. Additionally, water-unsuppressed spectra (8 FIDS) were obtained from each voxel using identical parameters (but without VAPOR suppression), enabling metabolite concentration quantification by referencing to voxel water signals. Total scan times in each mouse were ≤ 2 h.
Fig. 1.
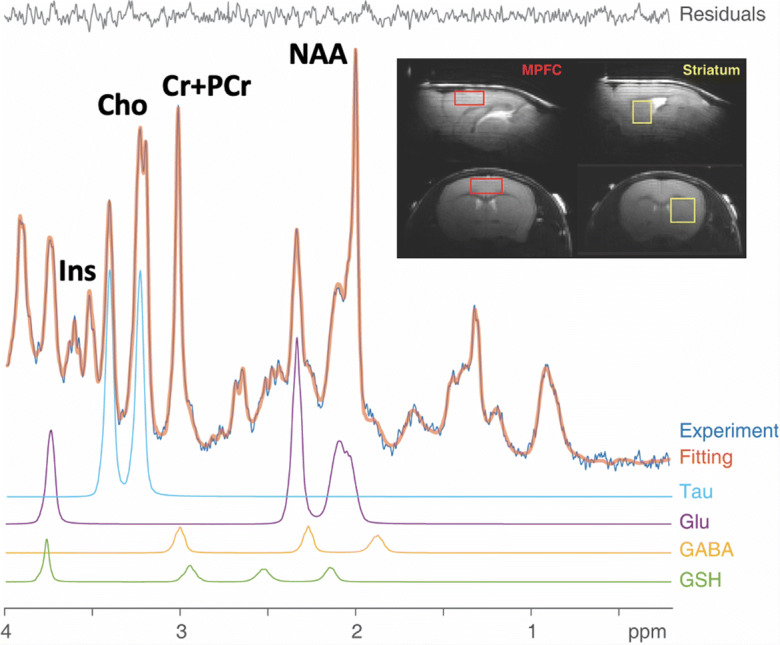
Sample medial prefrontal cortex (MPFC) MRS scan from an aged Tat-tg mouse. Shown are the original data (light blue trace), spectrum fit (brown trace for the entire spectrum, and fit residuals. To illustrate LC Model peak fitting, taurine (Tau), glutamate (Glu), GABA, and glutathione (GSH) peak fits are shown below the complete spectrum. Additional peaks that were quantifiable with high precision were N-acetylasparate (NAA), creatine + phosphocreatine (Cr+PCr), choline (Cho), and myo-inositol (Ins). Insets: MPFC and striatum MRS Voxels (red and yellow boxes, respectively) overlaid on sagittal (top row) and coronal (bottom row) MRI images (A, anterior; P,posterior; R,right; L, left)
Tissue extraction and preparation
Following imaging studies, mice were euthanized via cervical dislocation and brains were flash frozen on dry ice and stored at −80 °C. Tissues were later prepared for steroid analyses and polymerase chain reaction (PCR). All biochemical analyses were conducted in a manner blinded to condition.
Enzyme-linked immunosorbent assay for steroids
Enzyme-linked immunosorbent assay (ELISA) for testosterone and estradiol was carried out as previously described with modifications to accommodate brain tissue [15, 16, 20]. Briefly, MPFC and bilateral striatum (caudate/putamen) were grossly dissected and wet weights were collected. Dissected tissue was homogenized in a 10× volume of PBS using a Pro-200 tissue homogenizer (Pro-Scientific Inc, Oxford, CT). Homogenized tissue was incubated with ice-cold ether in glass borosilicate culture tubes, snap frozen, and evaporated to dryness. Steroid was reconstituted with kit extraction buffer at a 1:20 dilution factor and assayed for testosterone (Neogen Corp., #402510, Lexington, KY) and estradiol (Neogen Corp., #402110) per manufacturer instructions. The testosterone antibody reportedly cross-reacts with testosterone and dihydrotestosterone 100% and nominally with other steroids (< 1%). The estradiol antibody reportedly cross-reacts with 17β-estradiol 100%, testosterone 1%, and nominally with other steroids (< 1%). The intra-assay variance for testosterone was 3.3% and for estradiol was 4.4%.
Expression of tat mRNA via polymerase chain reaction
Total RNA of the remaining brain tissue was isolated with TRIzol reagent according to manufacturer’s recommendations (Life Technologies, Inc.). RNA was cleaned using the Qiagen RNeasy Mini Prep kit© (Qiagen, Germany; #74104). Synthesis of cDNA was performed with 1 μg of total RNA using a RevertAid First Strand cDNA Synthesis kit (Thermo Fisher Scientific; #K1651). Polymerase Chain Reaction (PCR) was performed with 1 μg of cDNA in a 25 μL final volume containing primers (400 nM) and PowerUp SYBR Green master mix (Thermo Fisher Scientific; #A25742), using specific primers to tat 5′-GGA ATT CAC CAT GGA GCC AGT AGA TCC T-3′ and 5′-CGG GAT CCC TAT TCC TTC GGG CCT GT-3′ and GAPDH 5′-GGA AGC TCA CTG GCA TGG C-3′ and 5′-TAG ACG GCA GGT CAG GTC CA-3′. Primers were purchased from IDT (Coralville, IA). Reactions were initiated by denaturation at 94 °C for 2 min, followed by 40 cycles of 94 °C for 30 s, 60 °C for 30 s, 72 °C for 30 s, and one cycle of extension at 72 °C for 7 min. Relative expression of tat and GAPDH mRNA was quantified via densitometry of 1.8% agarose gels (see Supplementary Information Figure S1) using Fiji [52]. The expression of tat was normalized to that of GAPDH for quantitative analyses.
Data analyses
MRS data processing was conducted in a blind manner with respect to mouse age and sex. After correcting for frequency and phase shifts, FIDs were summed for metabolite quantification. Spectra were analyzed using the LCModel frequency domain fitting method [53, 54]. The basis set for LCModel was simulated using density matrix simulations [55]. This procedure enabled quantification of metabolites of interest including glutamate (Glu), GABA, glutathione (GSH), N-acetylaspartate (NAA), myo-inositol (mI), and taurine. Metabolites with Cramer Rao Lower Bound (CRLB) fit values exceeding 20% were considered unreliable and excluded from statistical analyses. We used 2-way ANOVAs with Tukey’s HSD post hoc tests to determine effects of age and sex (Prism Version 8.4.3, GraphPad Software, LLC). Differences in the brain expression of tat and GAPDH mRNA and steroid concentrations among aged male and female mice were assessed via Student’s two-tailed t-tests. Simple linear regressions were utilized to characterize significant associations between all continuous variables. For all analyses, the threshold for statistical significance was set at p = 0.05.
Results
Mean ages of mice scanned for this study are shown stratified by sex in Table 1. Within each age group, there were no age differences as a function of sex. With regard to MRS metabolites, we detected effects of age on GABA and taurine levels in MPFC (GABA: F1,39=7.7, p = 0.008, taurine: F1,39=11.7, p = 0.002) and striatum (GABA: F1,39=5.3, p = 0.027, taurine: F1,39=6.4, p = 0.016), both of which were lower in older Tat-tg mice, as well as in Glu levels in MPFC (F1,39=21.6, p < 0.001), which also was lower in older Tat-tg mice (Fig. 2). We quantified the relative magnitudes of age effects on these metabolites by calculating mean Glu and GABA levels in each age group (using weighted-means to account for different numbers of mice in each group), and we found comparable MPFC reductions for Glu (−19.5%) and GABA (−15.5%) in older mice. By contrast, in striatum, we found a small age-related decrease of Glu levels (−4.9%) but an apparently larger effect of age on GABA levels (−27.7%). Because age-matched control mice not expressing Tat were not included in this study, we cannot determine the extent to which these effects are attributable to age or a combination of age plus Tat expression. We also detected an effect of sex on taurine levels in MPFC (F1,39=5.6, p = 0.023) and striatum (F1,39=4.9, p = 0.033). In MPFC, taurine and GSH were lowest in older female Tat-tg mice (Fig. 2), and we detected an age × sex interaction effect on GSH level (F1,39=4.9, p = 0.033). We did not detect effects of age or sex on levels of other MRS metabolites (Supplementary Information Table S1).
Fig. 2.
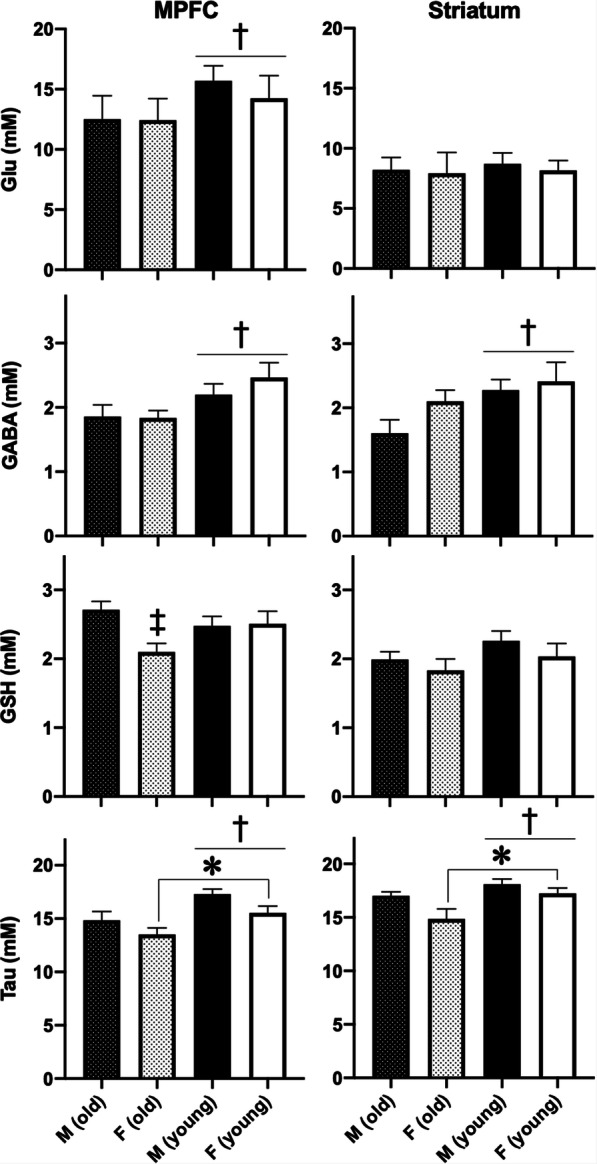
Effects of age and sex on medial prefrontal cortex (MPFC, left column) and striatal (right column) metabolite levels for glutamate (Glu), GABA, glutathione (GSH), and taurine (Tau). Shown are means + SEMs of 10-13 observations per group. GABA and taurine levels were lower in both brain regions in older mice and MPFC Glu levels were lower in older mice. MPFC GSH levels were lower in older female mice and taurine levels in both brain areas were lower in female mice. Asterisk indicates a main effect of sex. Dagger indicates a main effect of age. Double dagger indicates an interaction wherein older females differ from older males, p < 0.05
PCR confirmed the presence of tat mRNA in the brains of aged Tat-tg mice, consistent with transgene leak reported elsewhere [24] (Supplementary Information Figure S1). Expression of tat mRNA normalized to GAPDH did not significantly differ by sex (Table 2) nor did it correlate with any other continuous variable. However, GAPDH-normalized tat mRNA expression was higher in our aged Tat-tg mice (~4.75-fold greater) than the low level expression reported in uninduced young adult Tat-tg mice [46].
Table 2.
Relative expression of tat mRNA in aged male (n = 9) and female (n = 12) Tat-transgenic mice. PCR blots depicted in Supplementary Information Figure S1
| mRNA expression | Aged ♀ | Aged ♂ |
|---|---|---|
| tat relative to GAPDH (mean, tat/GAPDH ± SD) | 1.67 ± 0.35 | 1.95 ± 0.55 |
Testosterone concentrations were greater among aged males compared with aged females in MPFC [t(18) = 3.94, p = 0.001] and striatum [t(18) = 3.54, p = 0.002; Table 3]. In aged males, greater age predicted lower testosterone content in MPFC [β = −0.45, t(18) = −2.15, p < 0.04] and striatum [β = −0.53, t(18) = −2.67, p < 0.02], an effect not observed in females (Fig. 3). No significant differences were observed in brain estradiol or the estradiol/testosterone ratio (an indicator of aromatization). No additional correlations between steroid content and other study measures were observed.
Table 3.
Testosterone (ng/g) and estradiol (ng/g) content in the medial prefrontal cortex (MPFC) and striatum of aged male (n = 8) and female (n = 12) Tat-transgenic mice
| MPFC | Striatum | |||
|---|---|---|---|---|
| Aged ♀ | Aged ♂ | Aged ♀ | Aged ♂ | |
| Testosterone (mean, ng/g ± SD) | 0.13 ± 0.14 | 1.04 ± 0.80* | 0.08 ± 0.14 | 1.04 ± 0.94* |
| Estradiol (mean, ng/g ± SD) | 0.019 ± 0.014 | 0.013 ± 0.024 | 0.024 ± 0.023 | 0.019 ± 0.018 |
| Estradiol:testosterone ratio (mean ± SD) | 27.0 ± 67.9 | 1.3 ± 1.7 | 11.0 ± 15.4 | 1.9 ± 2.0 |
*Greater than females, Student’s t-tests p = 0.001–0.002
Fig. 3.

Age was inversely associated with testosterone concentration in a frontal cortex (rmale = −0.76, R2 = 0.58, p = 0.03; rfemale = −0.30, R2 = 0.09, not significant) and b striatum (rmale = −0.90, R2 = 0.82, p = 0.002; rfemale = −0.34, R2 = 0.12, not significant) of males (closed circles) but not females (open circles). Dotted lines depict simple linear regressions
Discussion
We found lower MPFC GABA, Glu, and taurine levels and lower striatal GABA and taurine levels in aged uninduced Tat-tg mice. In MPFC, we also detected effects of sex, finding lower GSH and taurine levels in female versus male mice. Similarly, in striatum, taurine levels were lower in females than males. Several of these effects parallel MRS changes identified in a longitudinal study of genetically similar C57BL/6J mice [38]. In that study, striatal GABA, Glu, GSH, and taurine declined in C57BL/6J mice aged 18 months (within the age range of the older mice we studied) as did cortical Glu [38]. Additionally, female C57BL/6J mice exhibited lower cortical and striatal taurine levels than males [38]. By contrast, we detected age effects in Tat-tg mice not reported in 18-month-old C57BL/6J mice [38] including cortical GABA, GSH, and taurine levels. While these apparent differences could result from a cohort effect in our study or from the fact that we acquired cortical MRS data from an anterior MPFC voxel versus a more posterior cortical voxel scanned in the C57BL/6J mouse study [38], it also is possible that aged Tat-tg mice experience more cortical MRS abnormalities than aged wild-type C57BL/6J mice.
A strain difference is plausible because unlike C57BL/6J mice, Tat-tg mice experience chronic Tat transgene leak and Tat mRNA and protein exposure sufficient by 1 year of age to promote neuroinflammation, brain structural abnormalities, pre- and post-synaptic abnormalities, and axonal damage [24]. Similarly, Dox induction of Tat expression in Tat-tg mice for 1 year causes robust brain and behavioral effects including astrocyte activation in cortex and striatum, impaired long- and short-term memory, and motor dysfunction [17]. Thus, prolonged exposure to Tat protein in Tat-tg mice either by Tat transgene leak or by Dox induction affects brain structure, function, and behavior. That being said, our study lacks controls that would have allowed us to deconvolve the effects of age and Tat expression, a goal for future investigations. Below, we discuss potential implications of each of the MRS metabolite abnormalities we detected.
GABA and glutamate
Glu and GABA are the major excitatory and inhibitory neurotransmitters, respectively. Accordingly, the present findings in striatum that GABA but not Glu levels were lower in aged mice (Fig. 2) could suggest that older Tat-tg mice maintain lower striatal inhibitory tone, a possibility consistent with striatal electrophysiology abnormalities found in Tat-tg mice [41, 43]. By contrast, Glu and GABA levels both were lower in MPFC in older mice by comparable degrees, perhaps reflecting minimal change in excitatory/inhibitory balance with aging. However, Glu and GABA exist in neurotransmission and metabolic pathways [56] that cannot be distinguished with the in vivo proton MRS methods we used. Accordingly, the differential effects of age on MPFC and striatal Glu and GABA we report could reflect neurotransmission and/or metabolic abnormalities that could manifest differently in these brain regions. We suspect that abnormal metabolism may contribute to the present findings because Tat impairs mitochondrial function and metabolism [19, 26–28, 34] and thus could affect Glu and GABA levels in metabolic pools. Future studies employing in vivo proton MRS along with phosphorus (31P) MRS could help to resolve this issue by concurrently characterizing Tat’s effects on mitochondria and metabolism [57].
Glutathione
GSH is the most abundant endogenous small molecule antioxidant and it plays a key role in buffering excess oxidative stress [58]. GSH levels also are depleted by inflammatory mediators including lipopolysaccharide, IL-1β, and TNFα [59–61], which increase production of reactive oxygen species normally neutralized by GSH [62]. Accordingly, the lower MPFC GSH levels we found in females, driven mainly by lower levels in older females (Fig. 2), could reflect higher levels of oxidative stress and/or inflammation in these mice. Estrogen exerts antioxidant [63–66] and anti-inflammatory [66, 67] effects, and thus estrogen declines could result in GSH depletion, an effect reported in rats after abrupt estrogen depletion via ovariectomy [64–66, 68]. It also is plausible that chronic Tat transgene leak in older Tat-tg mice may contribute to lower GSH levels because Tat accelerates decline of circulating estrogen levels [15] and it induces inflammation, oxidative stress, and abnormal GSH levels [6, 8, 14, 21–25, 44, 69–71]. Accordingly, the combined effects of aging and Tat transgene leak in older female Tat-tg mice could more substantially deplete GSH, resulting in the age × sex interaction effect we detected for MPFC GSH levels. An MPFC GSH abnormality could have functional implications as the frontal cortex is seminally involved in cognition and behavioral inhibition, both of which are impaired by short- and long-term Tat expression [9, 15, 17, 23].
Taurine
Taurine regulates osmotic balance and cellular volume, it inhibits excitatory glutamatergic N-methyl-D-aspartate receptor signaling, it facilitates mitochondrial protein syntheses, and like GSH, taurine exerts antioxidant and anti-inflammatory effects [72–74]. In MPFC, taurine declines with aging were greater in females than in males, which could amplify inflammation and oxidative stress and contribute to the relatively large MPFC GSH decrements we found in older female Tat-tg mice. The cortical and striatal taurine declines detected presently in older mice parallel postmortem findings in rat cortex and striatum [75]. By contrast, cortical taurine levels quantified by MRS remained stable in C57BL/6J mice up to 24 months of age [38]. This apparent study difference, like that noted above for cortical GSH, may be attributable to a cohort effect, to the fact that our MRS data are from a frontal cortex voxel versus a more posterior cortex voxel scanned in the C57BL/6J mouse study [38], and/or to the combined effects of aging plus prolonged Tat transgene leak in Tat-tg mice, which could exacerbate taurine depletion. An effect of Tat transgene leak also is plausible given that induction of Tat expression induces cortical edema in young Tat-tg mice [46], that Tat likely causes edema in older female Tat-tg mice [15], and that edema triggers cellular taurine loss to counter extracellular hypoosmolality [73]. Because estrogen upregulates taurine transporter activity [76] and reduces taurine efflux in an estrogen receptor-dependent manner consequent to glutamate excitotoxicity [77], estradiol declines in older female mice also may contribute to the especially low MPFC and striatal taurine levels we found in older female Tat-tg mice (Fig. 2).
Striatal taurine declines in aged rats have been linked to abnormal striatal dopamine levels [45, 78] and to impaired spatial learning [78], the latter of which was found after Dox induction of Tat expression in young and old Tat-tg mice [9, 10, 13, 15, 17]. Given that Tat impairs mitochondrial function and induces excess oxidative stress and inflammation [6, 8, 14, 20–25, 44, 69–71] and that taurine preserves mitochondrial health [72–74], the cortical and striatal taurine declines we found in aged Tat-tg mice could exacerbate mitochondrial dysfunction, oxidative stress, and inflammation in these brain areas. Because taurine supplementation in aged mice improved spatial memory as well as Glu and GABA levels [79], age-related taurine depletion could contribute to Glu, GABA, and spatial memory impairments detected in aged Tat-tg mice as observed presently and elsewhere [15, 17].
The present findings have potential translational value based on recent demonstrations in elderly women that short-term dietary taurine supplementation reduced systemic inflammation while dietary taurine in combination with exercise improved cognition [80]. Both effects may have resulted in part from correcting low systemic taurine levels in older individuals [81]. By opposing several deleterious effects induced by Tat protein and by HIV, dietary taurine supplementation could have promise as a novel adjunct treatment for older individuals living with HIV. To our knowledge, taurine supplementation has not been tested either in Tat-tg mouse models or in individuals with HIV. Similarly, sex steroid hormones and their analogs exert pleiotropic beneficial effects and attenuate Tat-mediated neurological impairments ([20, 27, 82], possibly by inhibiting inflammation and oxidative stress and increasing cellular GSH and taurine levels [63–67, 76, 77]. The low brain testosterone levels we found in male Tat-tg mice as a function of age are consistent with reports of lower circulating sex steroid levels among HIV+ individuals [31, 32, 83, 84]; albeit, levels within the central nervous system cannot typically be assessed clinically. Accordingly, in future studies, we plan to test whether supplementation with taurine, sex steroid hormones, or sex steroid hormone analogs normalizes MRS metabolites in aged murine models of HAND. Such studies are needed given that more than half of the US population living with HIV is middle-aged or older [1] and that Tat protein expression endures and may even increase during cART [5–7]. These findings help position Tat as a potentially important therapeutic target in older individuals living with HIV.
Limitations
A major limitation to this study is that we did not scan age-matched wild-type (C57BL/6J) mice to determine whether the MRS effects we detected as a function of age and sex in Tat-tg mice differ from effects of age and sex in wild-type mice or in other controls (e.g., Tat(-)-tg mice). Also, we did not directly measure oxidative stress or inflammation. Thus, we cannot deconvolve individual effects of age or tat mRNA expression on outcomes and we cannot definitively conclude that MRS metabolite abnormalities reflect oxidative stress or inflammation. Further, we did not quantify sex steroid hormone levels or tat mRNA levels in young Tat-tg mice in this this study so we cannot determine whether the MRS differences we detected as a function of age could be driven by age-related sex steroid abnormalities, Tat expression increases, or by a combination of these effects. Yet, prior studies in older Tat-tg mice indicate that nearly two-thirds of 18-month-old female mice are peri- or post-estropausal and exhibit sex steroid hormone imbalances [15] and that Tat transgene leak is considerable by 1 year of age [24]. Together, these findings support the possibility that sex steroid abnormalities and Tat transgene leak contribute to the present MRS findings. These issues will be addressed directly in our future studies. Notwithstanding these limitations, the present data indicate that age and sex differences in several MRS metabolites are apparent in uninduced Tat-tg mice including for metabolites that reflect oxidative stress and inflammation (GSH and taurine), neural excitation, inhibition, and metabolism (Glu and GABA), and osmolality (taurine). These MRS metabolites may serve as useful noninvasive biomarkers in future prospective studies of aged Tat-tg mice including those evaluating novel treatments for HIV.
Supplementary Information
{kind=link}
Expression of tat mRNA via polymerase chain reaction. Expression of GAPDH and tat mRNA via PCR among aged male and female Tat-transgenic mice. In cases 1-19, the left band is GAPDH and the right band is tat in each case. In cases 20 and 21, the first two bands are GAPDH in each case and the second bands are tat in each case. (PNG 1166 kb)
(PDF 59 kb)
Availability of data and material
Data will be made available to qualified researchers upon request.
Author contributions
Drs. Paris and Kaufman conceptualized the study.
Drs. Chen, Du, and Ms. Qrareya, Mahdi, and Mr. Anderson acquired the data.
Drs. Paris, Chen, Du, and Kaufman, and Ms. Qrareya, Mahdi, and Mr. Anderson analyzed the data.
Drs. Paris, Du, and Kaufman interpreted the data.
Drs. Paris and Kaufman drafted the manuscript.
All authors reviewed, provided edits, and approved of the manuscript final version.
Funding
This work was supported in part by National Institutes of Health grants S10RR019356 (MJK), R01DA039044 (MJK and JPM), R00DA039791 (JJP), R01DA052851 (JJP), an administrative supplement from award P30GM122733 (JJP and MJK), and by the Counterdrug Technology Assessment Center, an office within the Office of National Drug Control Policy, via Contract No. DBK39-03-C-0075 (MJK), awarded by the Army Contracting Agency. The content of the information does not necessarily reflect the position or the policy of the U.S. Government, and no official endorsement should be inferred.
Declarations
Conflict of interest
The authors declare no competing interests.
Ethics approval
The animal research included in this study was conducted after review and approval by the McLean Hospital Institutional and Animal Care Use Committee (IACUC).
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Centers for Disease Control and Prevention. Estimated HIV incidence and prevalence in the United States, 2014–2018. HIV Surveillance Supplemental Report. 2020:2020.
- 2.Mackiewicz MM, Overk C, Achim CL, Masliah E. Pathogenesis of age-related HIV neurodegeneration. J Neurovirol. 2019;25:622–633. doi: 10.1007/s13365-019-00728-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Saylor D, Dickens AM, Sacktor N, Haughey N, Slusher B, Pletnikov M, Mankowski JL, Brown A, Volsky DJ, McArthur JC. HIV-associated neurocognitive disorder--pathogenesis and prospects for treatment. Nat Rev Neurol. 2016;12:234–248. doi: 10.1038/nrneurol.2016.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mediouni S, Chinthalapudi K, Ekka MK, Usui I, Jablonski JA, Clementz MA, et al. Didehydro-Cortistatin A Inhibits HIV-1 by Specifically Binding to the Unstructured Basic Region of Tat. MBio. 2019;10. 10.1128/mBio.02662-18. [DOI] [PMC free article] [PubMed]
- 5.Mediouni S, Darque A, Baillat G, Ravaux I, Dhiver C, Tissot-Dupont H, Mokhtari M, Moreau H, Tamalet C, Brunet C, Paul P, Dignat-George F, Stein A, Brouqui P, A. Spector S, R. Campbell G, P. Loret E. Antiretroviral therapy does not block the secretion of the human immunodeficiency virus tat protein. Infect Disord Drug Targets. 2012;12:81–86. doi: 10.2174/187152612798994939. [DOI] [PubMed] [Google Scholar]
- 6.Johnson TP, Patel K, Johnson KR, Maric D, Calabresi PA, Hasbun R, Nath A. Induction of IL-17 and nonclassical T-cell activation by HIV-Tat protein. Proc Natl Acad Sci U S A. 2013;110:13588–13593. doi: 10.1073/pnas.1308673110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Henderson LJ, Johnson TP, Smith BR, Reoma LB, Santamaria UA, Bachani M, Demarino C, Barclay RA, Snow J, Sacktor N, Mcarthur J, Letendre S, Steiner J, Kashanchi F, Nath A. Presence of Tat and transactivation response element in spinal fluid despite antiretroviral therapy. AIDS. 2019;33(Suppl 2):S145–S157. doi: 10.1097/QAD.0000000000002268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lawson MA, Kelley KW, Dantzer R. Intracerebroventricular administration of HIV-1 Tat induces brain cytokine and indoleamine 2,3-dioxygenase expression: a possible mechanism for AIDS comorbid depression. Brain Behav Immun. 2011;25:1569–1575. doi: 10.1016/j.bbi.2011.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Carey AN, Sypek EI, Singh HD, Kaufman MJ, McLaughlin JP. Expression of HIV-Tat protein is associated with learning and memory deficits in the mouse. Behav Brain Res. 2012;229:48–56. doi: 10.1016/j.bbr.2011.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fitting S, Ignatowska-Jankowska BM, Bull C, Skoff RP, Lichtman AH, Wise LE, Fox MA, Su J, Medina AE, Krahe TE, Knapp PE, Guido W, Hauser KF. Synaptic dysfunction in the hippocampus accompanies learning and memory deficits in human immunodeficiency virus type-1 Tat transgenic mice. Biol Psychiatry. 2013;73:443–453. doi: 10.1016/j.biopsych.2012.09.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Paris JJ, Singh HD, Ganno ML, Jackson P, McLaughlin JP. Anxiety-like behavior of mice produced by conditional central expression of the HIV-1 regulatory protein, Tat. Psychopharmacology. 2014;231:2349–2360. doi: 10.1007/s00213-013-3385-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kesby JP, Markou A, Semenova S. The effects of HIV-1 regulatory TAT protein expression on brain reward function, response to psychostimulants and delay-dependent memory in mice. Neuropharmacology. 2016;109:205–215. doi: 10.1016/j.neuropharm.2016.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Marks WD, Paris JJ, Schier CJ, Denton MD, Fitting S, McQuiston AR, et al. HIV-1 Tat causes cognitive deficits and selective loss of parvalbumin, somatostatin, and neuronal nitric oxide synthase expressing hippocampal CA1 interneuron subpopulations. J Neurovirol. 2016;22:747–762. doi: 10.1007/s13365-016-0447-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.McLaughlin JP, Paris JJ, Mintzopoulos D, Hymel KA, Kim JK, Cirino TJ, Gillis TE, Eans SO, Vitaliano GD, Medina JM, Krapf RC, Stacy HM, Kaufman MJ. Conditional Human Immunodeficiency Virus Transactivator of Transcription Protein Expression Induces Depression-like Effects and Oxidative Stress. Biol Psychiatry Cogn Neurosci Neuroimaging. 2017;2:599–609. doi: 10.1016/j.bpsc.2017.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Qrareya AN, Mahdi F, Kaufman MJ, Ashpole NM, Paris JJ. HIV-1 Tat promotes age-related cognitive, anxiety-like, and antinociceptive impairments in female mice that are moderated by aging and endocrine status. Geroscience. 2020. 10.1007/s11357-020-00268-z. [DOI] [PMC free article] [PubMed]
- 16.Salahuddin MF, Qrareya AN, Mahdi F, Jackson D, Foster M, Vujanovic T, Box JG, Paris JJ. Combined HIV-1 Tat and oxycodone activate the hypothalamic-pituitary-adrenal and -gonadal axes and promote psychomotor, affective, and cognitive dysfunction in female mice. Horm Behav. 2020;119:104649. doi: 10.1016/j.yhbeh.2019.104649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhao X, Fan Y, Vann PH, Wong JM, Sumien N, He JJ. Long-term HIV-1 Tat Expression in the Brain Led to Neurobehavioral, Pathological, and Epigenetic Changes Reminiscent of Accelerated Aging. Aging Dis. 2020;11:93–107. doi: 10.14336/AD.2019.0323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Eugenin EA, King JE, Nath A, Calderon TM, Zukin RS, Bennett MVL, Berman JW. HIV-tat induces formation of an LRP-PSD-95- NMDAR-nNOS complex that promotes apoptosis in neurons and astrocytes. Proc Natl Acad Sci U S A. 2007;104:3438–3443. doi: 10.1073/pnas.0611699104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kruman II, Nath A, Mattson MP. HIV-1 protein Tat induces apoptosis of hippocampal neurons by a mechanism involving caspase activation, calcium overload, and oxidative stress. Exp Neurol. 1998;154:276–288. doi: 10.1006/exnr.1998.6958. [DOI] [PubMed] [Google Scholar]
- 20.Paris JJ, Zou S, Hahn YK, Knapp PE, Hauser KF. 5α-reduced progestogens ameliorate mood-related behavioral pathology, neurotoxicity, and microgliosis associated with exposure to HIV-1 Tat. Brain Behav Immun. 2016;55:202–214. doi: 10.1016/j.bbi.2016.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Flora G, Pu H, Hennig B, Toborek M. Cyclooxygenase-2 is involved in HIV-1 Tat-induced inflammatory responses in the brain. Neuromolecular Med. 2006;8:337–352. doi: 10.1385/NMM:8:3:337. [DOI] [PubMed] [Google Scholar]
- 22.Mediouni S, Jablonski J, Paris JJ, Clementz MA, Thenin-Houssier S, McLaughlin JP, Valente S. Didehydro-cortistatin A inhibits HIV-1 Tat mediated neuroinflammation and prevents potentiation of cocaine reward in Tat transgenic mice. Curr HIV Res. 2015;13:64–79. doi: 10.2174/1570162X13666150121111548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Paris JJ, Singh HD, Carey AN, McLaughlin JP. Exposure to HIV-1 Tat in brain impairs sensorimotor gating and activates microglia in limbic and extralimbic brain regions of male mice. Behav Brain Res. 2015;291:209–218. doi: 10.1016/j.bbr.2015.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dickens AM, Yoo SW, Chin AC, Xu J, Johnson TP, Trout AL, Hauser KF, Haughey NJ. Chronic low-level expression of HIV-1 Tat promotes a neurodegenerative phenotype with aging. Sci Rep. 2017;7:7748. doi: 10.1038/s41598-017-07570-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gonek M, McLane VD, Stevens DL, Lippold K, Akbarali HI, Knapp PE, et al. CCR5 mediates HIV-1 Tat-induced neuroinflammation and influences morphine tolerance, dependence, and reward. Brain Behav Immun. 2018;69:124–138. doi: 10.1016/j.bbi.2017.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Norman JP, Perry SW, Kasischke KA, Volsky DJ, Gelbard HA. HIV-1 trans activator of transcription protein elicits mitochondrial hyperpolarization and respiratory deficit, with dysregulation of complex IV and nicotinamide adenine dinucleotide homeostasis in cortical neurons. J Immunol. 2007;178:869–876. doi: 10.4049/jimmunol.178.2.869. [DOI] [PubMed] [Google Scholar]
- 27.Paris JJ, Liere P, Kim S, Mahdi F, Buchanan ME, Nass SR, Qrareya AN, Salahuddin MF, Pianos A, Fernandez N, Shariat-Madar Z, Knapp PE, Schumacher M, Hauser KF. Pregnane steroidogenesis is altered by HIV-1 Tat and morphine: Physiological allopregnanolone is protective against neurotoxic and psychomotor effects. Neurobiol Stress. 2020;12:100211. doi: 10.1016/j.ynstr.2020.100211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sivalingam K, Cirino TJ, McLaughlin JP, Samikkannu T. HIV-Tat and Cocaine Impact Brain Energy Metabolism: Redox Modification and Mitochondrial Biogenesis Influence NRF Transcription-Mediated Neurodegeneration. Mol Neurobiol. 2020;58:490–504. doi: 10.1007/s12035-020-02131-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schoenbaum EE, Hartel D, Lo Y, Howard AA, Floris-Moore M, Arnsten JH, Santoro N. HIV infection, drug use, and onset of natural menopause. Clin Infect Dis. 2005;41:1517–1524. doi: 10.1086/497270. [DOI] [PubMed] [Google Scholar]
- 30.Fan MD, Maslow B-S, Santoro N, Schoenbaum E. HIV and the menopause. Menopause Int. 2008;14:163–168. doi: 10.1258/mi.2008.008027. [DOI] [PubMed] [Google Scholar]
- 31.Karim R, Mack WJ, Kono N, Tien PC, Anastos K, Lazar J, Young M, Cohen M, Golub E, Greenblatt RM, Kaplan RC, Hodis HN. Gonadotropin and sex steroid levels in HIV-infected premenopausal women and their association with subclinical atherosclerosis in HIV-infected and -uninfected women in the women’s interagency HIV study (WIHS) J Clin Endocrinol Metab. 2013;98:E610–E618. doi: 10.1210/jc.2012-3195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Slama L, Jacobson LP, Li X, Palella FJ, Jr, Margolick JB, Kingsley LA, et al. Longitudinal Changes Over 10 Years in Free Testosterone Among HIV-Infected and HIV-Uninfected Men. J Acquir Immune Defic Syndr. 2016;71:57–64. doi: 10.1097/QAI.0000000000000821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ivanov AV, Valuev-Elliston VT, Ivanova ON, Kochetkov SN, Starodubova ES, Bartosch B, et al. Oxidative Stress during HIV Infection: Mechanisms and Consequences. Oxid Med Cell Longev. 2016;2016:8910396. doi: 10.1155/2016/8910396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fields JA, Ellis RJ. HIV in the cART era and the mitochondrial: immune interface in the CNS. Int Rev Neurobiol. 2019;145:29–65. doi: 10.1016/bs.irn.2019.04.003. [DOI] [PubMed] [Google Scholar]
- 35.Mandas A, Iorio EL, Congiu MG, Balestrieri C, Mereu A, Cau D, et al. Oxidative imbalance in HIV-1 infected patients treated with antiretroviral therapy. J Biomed Biotechnol. 2009;2009:749575. doi: 10.1155/2009/749575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Manda KR, Banerjee A, Banks WA, Ercal N. Highly active antiretroviral therapy drug combination induces oxidative stress and mitochondrial dysfunction in immortalized human blood-brain barrier endothelial cells. Free Radic Biol Med. 2011;50:801–810. doi: 10.1016/j.freeradbiomed.2010.12.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Asundi A, Robles Y, Starr T, Landay A, Kinslow J, Ladner J, White L, Plank RM, Melbourne K, Weisholtz D, Bennett M, Pan H, Stern E, Lin A, Kuritzkes DR, Lin NH. Immunological and Neurometabolite Changes Associated With Switch From Efavirenz to an Integrase Inhibitor. J Acquir Immune Defic Syndr. 2019;81:585–593. doi: 10.1097/QAI.0000000000002079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Duarte JMN, Do KQ, Gruetter R. Longitudinal neurochemical modifications in the aging mouse brain measured in vivo by 1H magnetic resonance spectroscopy. Neurobiol Aging. 2014;35:1660–1668. doi: 10.1016/j.neurobiolaging.2014.01.135. [DOI] [PubMed] [Google Scholar]
- 39.Carey AN, Liu X, Mintzopoulos D, Paris JJ, McLaughlin JP, Kaufman MJ. Conditional Tat protein brain expression in the GT-tg bigenic mouse induces cerebral fractional anisotropy abnormalities. Curr HIV Res. 2015;13:3–9. doi: 10.2174/1570162X13666150126125244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hahn YK, Podhaizer EM, Farris SP, Miles MF, Hauser KF, Knapp PE. Effects of chronic HIV-1 Tat exposure in the CNS: heightened vulnerability of males versus females to changes in cell numbers, synaptic integrity, and behavior. Brain Struct Funct. 2015;220:605–623. doi: 10.1007/s00429-013-0676-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schier CJ, Marks WD, Paris JJ, Barbour AJ, McLane VD, Maragos WF, et al. Selective Vulnerability of Striatal D2 versus D1 Dopamine Receptor-Expressing Medium Spiny Neurons in HIV-1 Tat Transgenic Male Mice. J Neurosci. 2017;37:5758–5769. doi: 10.1523/JNEUROSCI.0622-17.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zou S, Balinang JM, Paris JJ, Hauser KF, Fuss B, Knapp PE. Effects of HIV-1 Tat on oligodendrocyte viability are mediated by CaMKIIβ-GSK3β interactions. J Neurochem. 2019;149:98–110. doi: 10.1111/jnc.14668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cirino TJ, Harden SW, McLaughlin JP, Frazier CJ. Region-specific effects of HIV-1 Tat on intrinsic electrophysiological properties of pyramidal neurons in mouse prefrontal cortex and hippocampus. J Neurophysiol. 2020;123:1332–1341. doi: 10.1152/jn.00029.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nass SR, Hahn YK, McLane VD, Varshneya NB, Damaj MI, Knapp PE, et al. Chronic HIV-1 Tat exposure alters anterior cingulate cortico-basal ganglia-thalamocortical synaptic circuitry, associated behavioral control, and immune regulation in male mice. Brain, Behavior, & Immunity - Health. 2020;5:100077. doi: 10.1016/j.bbih.2020.100077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Strauss M, O’Donovan B, Ma Y, Xiao Z, Lin S, Bardo MT, Ortinski PI, McLaughlin JP, Zhu J. [3H]Dopamine Uptake through the Dopamine and Norepinephrine Transporters is Decreased in the Prefrontal Cortex of Transgenic Mice Expressing HIV-1 Transactivator of Transcription Protein. J Pharmacol Exp Ther. 2020;374:241–251. doi: 10.1124/jpet.120.266023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kim BO, Liu Y, Ruan Y, Xu ZC, Schantz L, He JJ. Neuropathologies in transgenic mice expressing human immunodeficiency virus type 1 Tat protein under the regulation of the astrocyte-specific glial fibrillary acidic protein promoter and doxycycline. Am J Pathol. 2003;162:1693–1707. doi: 10.1016/S0002-9440(10)64304-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Langford D, Oh Kim B, Zou W, Fan Y, Rahimain P, Liu Y, et al. Doxycycline-inducible and astrocyte-specific HIV-1 Tat transgenic mice (iTat) as an HIV/neuroAIDS model. J Neurovirol. 2018;24:168–179. doi: 10.1007/s13365-017-0598-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Paris JJ, Walf AA, Frye CA. II. Cognitive performance of middle-aged female rats is influenced by capacity to metabolize progesterone in the prefrontal cortex and hippocampus. Brain Res. 2011;1379:149–163. doi: 10.1016/j.brainres.2010.10.099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gruetter R. Automatic, localized in vivo adjustment of all first- and second-order shim coils. Magn Reson Med. 1993;29:804–811. doi: 10.1002/mrm.1910290613. [DOI] [PubMed] [Google Scholar]
- 50.Tkac I, Starcuk Z, Choi I-Y, Gruetter R. In Vivo 1H NMR Spectroscopy of Rat Brain at 1 ms Echo Time. Magn Reson Med. 1999;41:649–656. doi: 10.1002/(SICI)1522-2594(199904)41:4<649::AID-MRM2>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 51.Deelchand DK, Van de Moortele P-F, Adriany G, Iltis I, Andersen P, Strupp JP, et al. In vivo 1H NMR spectroscopy of the human brain at 9.4 T: initial results. J Magn Reson. 2010;206:74–80. doi: 10.1016/j.jmr.2010.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White DJ, Hartenstein V, Eliceiri K, Tomancak P, Cardona A. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012;9:676–682. doi: 10.1038/nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Provencher SW. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn Reson Med. 1993;30:672–679. doi: 10.1002/mrm.1910300604. [DOI] [PubMed] [Google Scholar]
- 54.Provencher SW. Automatic quantitation of localized in vivo 1H spectra with LCModel. NMR Biomed. 2001;14:260–264. doi: 10.1002/nbm.698. [DOI] [PubMed] [Google Scholar]
- 55.Henry P-G, Marjanska M, Walls JD, Valette J, Gruetter R, Ugurbil K. Proton-observed carbon-edited NMR spectroscopy in strongly coupled second-order spin systems. Magn Reson Med. 2006;55:250–257. doi: 10.1002/mrm.20764. [DOI] [PubMed] [Google Scholar]
- 56.Walls AB, Waagepetersen HS, Bak LK, Schousboe A, Sonnewald U. The glutamine-glutamate/GABA cycle: function, regional differences in glutamate and GABA production and effects of interference with GABA metabolism. Neurochem Res. 2015;40:402–409. doi: 10.1007/s11064-014-1473-1. [DOI] [PubMed] [Google Scholar]
- 57.Liu Y, Gu Y, Yu X. Assessing tissue metabolism by phosphorous-31 magnetic resonance spectroscopy and imaging: a methodology review. Quant Imaging Med Surg. 2017;7:707–726. doi: 10.21037/qims.2017.11.03. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Forman HJ, Zhang H, Rinna A. Glutathione: overview of its protective roles, measurement, and biosynthesis. Mol Aspects Med. 2009;30:1–12. doi: 10.1016/j.mam.2008.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Noble F, Rubira E, Boulanouar M, Palmier B, Plotkine M, Warnet J-M, Marchand-Leroux C, Massicot F. Acute systemic inflammation induces central mitochondrial damage and mnesic deficit in adult Swiss mice. Neurosci Lett. 2007;424:106–110. doi: 10.1016/j.neulet.2007.07.005. [DOI] [PubMed] [Google Scholar]
- 60.Gavillet M, Allaman I, Magistretti PJ. Modulation of astrocytic metabolic phenotype by proinflammatory cytokines. Glia. 2008;56:975–989. doi: 10.1002/glia.20671. [DOI] [PubMed] [Google Scholar]
- 61.Chowdhury T, Allen MF, Thorn TL, He Y, Hewett SJ. Interleukin-1β Protects Neurons against Oxidant-Induced Injury via the Promotion of Astrocyte Glutathione Production. Antioxidants (Basel). 2018;7. 10.3390/antiox7080100. [DOI] [PMC free article] [PubMed]
- 62.Muralidharan S, Mandrekar P. Cellular stress response and innate immune signaling: integrating pathways in host defense and inflammation. J Leukoc Biol. 2013;94:1167–1184. doi: 10.1189/jlb.0313153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Moosmann B, Behl C. The antioxidant neuroprotective effects of estrogens and phenolic compounds are independent from their estrogenic properties. Proc Natl Acad Sci U S A. 1999;96:8867–8872. doi: 10.1073/pnas.96.16.8867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Dilek M, Naziroğlu M, Baha Oral H, Suat Ovey I, Küçükayaz M, Mungan MT, et al. Melatonin modulates hippocampus NMDA receptors, blood and brain oxidative stress levels in ovariectomized rats. J Membr Biol. 2010;233:135–142. doi: 10.1007/s00232-010-9233-x. [DOI] [PubMed] [Google Scholar]
- 65.Da Silva MM, Schnorr CE, Behr GA, Gasparotto J, Bortolin RC, Moresco KS, et al. Oral administration of curcumin relieves behavioral alterations and oxidative stress in the frontal cortex, hippocampus, and striatum of ovariectomized Wistar rats. J Nutr Biochem. 2016;32:181–188. doi: 10.1016/j.jnutbio.2016.03.010. [DOI] [PubMed] [Google Scholar]
- 66.Yazğan B, Yazğan Y, Övey İS, Nazıroğlu M. Raloxifene and Tamoxifen Reduce PARP Activity, Cytokine and Oxidative Stress Levels in the Brain and Blood of Ovariectomized Rats. J Mol Neurosci. 2016;60:214–222. doi: 10.1007/s12031-016-0785-9. [DOI] [PubMed] [Google Scholar]
- 67.Sárvári M, Hrabovszky E, Kalló I, Solymosi N, Tóth K, Likó I, Széles J, Mahó S, Molnár B, Liposits Z. Estrogens regulate neuroinflammatory genes via estrogen receptors α and β in the frontal cortex of middle-aged female rats. J Neuroinflammation. 2011;8:82. doi: 10.1186/1742-2094-8-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Abbas AM, Elsamanoudy AZ. Effects of 17β-estradiol and antioxidant administration on oxidative stress and insulin resistance in ovariectomized rats. Can J Physiol Pharmacol. 2011;89:497–504. doi: 10.1139/y11-053. [DOI] [PubMed] [Google Scholar]
- 69.Flora G, Lee YW, Nath A, Hennig B, Maragos W, Toborek M. Methamphetamine potentiates HIV-1 Tat protein-mediated activation of redox-sensitive pathways in discrete regions of the brain. Exp Neurol. 2003;179:60–70. doi: 10.1006/exnr.2002.8048. [DOI] [PubMed] [Google Scholar]
- 70.Banerjee A, Zhang X, Manda KR, Banks WA, Ercal N. HIV proteins (gp120 and Tat) and methamphetamine in oxidative stress-induced damage in the brain: potential role of the thiol antioxidant N-acetylcysteine amide. Free Radic Biol Med. 2010;48:1388–1398. doi: 10.1016/j.freeradbiomed.2010.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chivero ET, Guo M-L, Periyasamy P, Liao K, Callen SE, Buch S. HIV-1 Tat Primes and Activates Microglial NLRP3 Inflammasome-Mediated Neuroinflammation. J Neurosci. 2017;37:3599–3609. doi: 10.1523/JNEUROSCI.3045-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Menzie J, Pan C, Prentice H, Wu J-Y. Taurine and central nervous system disorders. Amino Acids. 2014;46:31–46. doi: 10.1007/s00726-012-1382-z. [DOI] [PubMed] [Google Scholar]
- 73.Pasantes-Morales H. Taurine Homeostasis and Volume Control. Adv Neurobiol. 2017;16:33–53. doi: 10.1007/978-3-319-55769-4_3. [DOI] [PubMed] [Google Scholar]
- 74.Jakaria M, Azam S, Haque ME, Jo S-H, Uddin MS, Kim I-S, Choi DK. Taurine and its analogs in neurological disorders: Focus on therapeutic potential and molecular mechanisms. Redox Biol. 2019;24:101223. doi: 10.1016/j.redox.2019.101223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Benedetti MS, Russo A, Marrari P, Dostert P. Effects of ageing on the content in sulfur-containing amino acids in rat brain. J Neural Transm Gen Sect. 1991;86:191–203. doi: 10.1007/BF01250705. [DOI] [PubMed] [Google Scholar]
- 76.Shennan DB, Thomson J. Estrogen regulation and ion dependence of taurine uptake by MCF-7 human breast cancer cells. Cell Mol Biol Lett. 2007;12:396–406. doi: 10.2478/s11658-007-0011-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ritz M-F, Schmidt P, Mendelowitsch A. 17beta-estradiol effect on the extracellular concentration of amino acids in the glutamate excitotoxicity model in the rat. Neurochem Res. 2002;27:1677–1683. doi: 10.1023/A:1021695213099. [DOI] [PubMed] [Google Scholar]
- 78.Dawson R, Jr, Pelleymounter MA, Cullen MJ, Gollub M, Liu S. An age-related decline in striatal taurine is correlated with a loss of dopaminergic markers. Brain Res Bull. 1999;48:319–324. doi: 10.1016/S0361-9230(99)00003-9. [DOI] [PubMed] [Google Scholar]
- 79.El Idrissi A, Shen CH, L’amoreaux WJ. Neuroprotective role of taurine during aging. Amino Acids. 2013;45:735–750. doi: 10.1007/s00726-013-1544-7. [DOI] [PubMed] [Google Scholar]
- 80.Chupel MU, Minuzzi LG, Furtado G, Santos ML, Hogervorst E, Filaire E, Teixeira AM. Exercise and taurine in inflammation, cognition, and peripheral markers of blood-brain barrier integrity in older women. Appl Physiol Nutr Metab. 2018;43:733–741. doi: 10.1139/apnm-2017-0775. [DOI] [PubMed] [Google Scholar]
- 81.Jeevanandam M, Young DH, Ramias L, Schiller WR. Effect of major trauma on plasma free amino acid concentrations in geriatric patients. Am J Clin Nutr. 1990;51:1040–1045. doi: 10.1093/ajcn/51.6.1040. [DOI] [PubMed] [Google Scholar]
- 82.Paris JJ, Fenwick J, McLaughlin JP. Progesterone protects normative anxiety-like responding among ovariectomized female mice that conditionally express the HIV-1 regulatory protein, Tat, in the CNS. Horm Behav. 2014;65:445–453. doi: 10.1016/j.yhbeh.2014.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Yoshino Y, Koga I, Wakabayashi Y, Kitazawa T, Ota Y. Prevalence of and Risk Factors for Low Free Testosterone Levels in Japanese Men with Well-controlled Human Immunodeficiency Virus Infection. Curr HIV Res. 2020;18:381–386. doi: 10.2174/1570162X18666200720000344. [DOI] [PubMed] [Google Scholar]
- 84.De Vincentis S, Decaroli MC, Fanelli F, Diazzi C, Mezzullo M, Morini F, et al. Health status is related to testosterone, estrone and body fat: moving to functional hypogonadism in adult men with HIV. Eur J Endocrinol. 2021;184:107–122. doi: 10.1530/EJE-20-0855. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Expression of tat mRNA via polymerase chain reaction. Expression of GAPDH and tat mRNA via PCR among aged male and female Tat-transgenic mice. In cases 1-19, the left band is GAPDH and the right band is tat in each case. In cases 20 and 21, the first two bands are GAPDH in each case and the second bands are tat in each case. (PNG 1166 kb)
(PDF 59 kb)