

Abstract
The ability to study human cardiac development in health and disease is highly limited by the capacity to model the complexity of the human heart in vitro. Developing more efficient organ-like platforms that can model complex in vivo phenotypes, such as organoids and organs-on-a-chip, will enhance the ability to study human heart development and disease. This paper describes a protocol to generate highly complex human heart organoids (hHOs) by self-organization using human pluripotent stem cells and stepwise developmental pathway activation using small molecule inhibitors. Embryoid bodies (EBs) are generated in a 96-well plate with round-bottom, ultra-low attachment wells, facilitating suspension culture of individualized constructs.
The EBs undergo differentiation into hHOs by a three-step Wnt signaling modulation strategy, which involves an initial Wnt pathway activation to induce cardiac mesoderm fate, a second step of Wnt inhibition to create definitive cardiac lineages, and a third Wnt activation step to induce proepicardial organ tissues. These steps, carried out in a 96-well format, are highly efficient, reproducible, and produce large amounts of organoids per run. Analysis by immunofluorescence imaging from day 3 to day 11 of differentiation reveals first and second heart field specifications and at day 15, highly complex tissues inside hHOs, including myocardial tissue with regions of atrial and ventricular cardiomyocytes, as well as internal chambers lined with endocardial tissue. The organoids also exhibit an intricate vascular network throughout the structure and an external lining of epicardial tissue. From a functional standpoint, hHOs beat robustly and present normal calcium activity as determined by Fluo-4 live imaging. Overall, this protocol constitutes a solid platform for in vitro studies in human organ-like cardiac tissues.
SUMMARY:
Here, we describe a protocol to create developmentally relevant human heart organoids (hHOs) efficiently using human pluripotent stem cells by self-organization. The protocol relies on the sequential activation of developmental cues and produces highly complex, functionally relevant human heart tissues.
INTRODUCTION:
Congenital heart defects (CHDs) are the most common type of congenital defect in humans and affect approximately 1% of all live births1–3. Under most circumstances, the reasons for CHDs remain unknown. The ability to create human heart models in the lab that closely resemble the developing human heart constitutes a significant step forward to study the underlying causes of CHDs directly in humans, rather than in surrogate animal models.
The epitome of laboratory-grown tissue models are organoids, 3D cell constructs that resemble an organ of interest in cell composition and physiological function. Organoids are often derived from stem cells or progenitor cells and have been successfully used to model many organs such as the brain4,5, kidney6,7, intestine8,9, lung10,11, liver12,13, and pancreas14,15, just to name a few. Very recently, studies have emerged demonstrating the feasibility of creating self-assembling heart organoids to study heart development in vitro. These models include the use of mouse embryonic stem cells (mESCs) to model early heart development16,17 up to atrioventricular specification18 and of human pluripotent stem cells (hPSCs) to generate multi-germ layer cardiac-endoderm organoids19 and chambered cardioids20 with highly complex cellular composition.
This paper presents a novel 3-step WNT modulation protocol to generate highly complex hHOs in an efficient and cost-effective manner. Organoids are generated in 96-well plates, resulting in a high-throughput system that is scalable and can be easily automated. This method relies on creating hPSC aggregates and triggering developmental steps of cardiogenesis, including mesoderm and cardiac mesoderm formation, first and second heart field specification, proepicardial organ formation, and atrioventricular specification. After 15 days of differentiation, hHOs contain all major cell lineages found in the heart, well-defined internal chambers, atrial and ventricular chambers, and a vascular network throughout the organoid. This highly sophisticated and reproducible heart organoid system is amenable to the investigation of structural, functional, molecular and transcriptomic analyses in the study of heart development, and diseases, and pharmacological screening.
PROTOCOL:
-
hPSC culture and maintenance
NOTE: The human induced PSCs (hiPSCs) or human embryonic stem cells (hESCs) need to be cultured for at least 2 consecutive passages after thawing before being used to generate EBs for differentiation or for further cryopreservation. hPSCs are cultured in PSC medium (see the Table of Materials) on basement-membrane-extracellular matrix (BM-ECM)-coated 6-well culture plates. When performing medium changes on hPSCs in 6-well plates, add the medium directly to the inner side of the well, rather than directly on top of the cells to prevent unwanted cell detachment or stress. Users should be wary of pre-warming PSC media that should not be warmed at 37°C, in this protocol all PSC media used was thermostable.-
1.1.To coat the well-plates with the BM-ECM, thaw one aliquot of the BM-ECM (stored at −20 °C according to the manufacturer’s instructions) on ice and mix 0.5 mg of the BM-ECM with 12 mL of cold Dulbecco’s modified Eagle’s medium (DMEM)/F12 medium (stored at 4 °C). Distribute 2 mL of the DMEM/F12–BM-ECM mixture onto each well of a 6-well plate and incubate at 37 °C for at least 2 h.
-
1.2.To thaw the cells, first thaw the hPSC cryovial in a 37 °C bead or water bath for 1–2 min until only a small amount of ice is visible. Transfer the thawed cells to a centrifuge tube and slowly add 8–9 mL of the PSC medium supplemented with 2 μM of the ROCK inhibitor, thiazovivin (Thiaz), and centrifuge at 300 × g for 5 min. Remove the supernatant and resuspend the cell pallet in the PSC medium supplemented with 2 μM of Thiaz. Distribute the cells in the culture medium into 1–2 wells depending on the cryovial cell concentration and culture at 37 °C, 5% CO2 for 24 h before changing the PSC medium.
-
1.3.Change the medium on the cells at 48 h intervals. Perform washes and medium changes using DMEM/F12 (1 mL/well) and the PSC medium (2 mL/well), respectively.NOTE: Washes help remove cell waste and debris while fresh media changes provide cells with a renewed source of nutrients.
-
1.4.Passage the cells upon subconfluency (60–80% confluent) by aspirating the medium, then washing each well with 1 mL of 1x Dulbecco’s phosphate-buffered solution (no calcium, no magnesium; DPBS). Aspirate the DPBS and add 1 mL of the dissociation reagent for hPSCs (see the Table of Materials), followed by the aspiration of all but a thin film of the reagent after 10 s.
-
1.5.Incubate for 2–5 min with the thin film of the dissociation reagent for hPSCs until gaps form between cells.NOTE: The time to stop the dissociation is cell-line-dependent.
-
1.6.Add 1 mL of the PSC medium supplemented with 2 μM of Thiaz (PSC medium+Thiaz) to the well and gently tap the plate to induce cell detachment. Pipette the detached cells in the medium 1–2 times to break up any large colonies, and resuspend the cells in PSC medium+Thiaz in a 1:6 well ratio (cells from 1 well resuspended in 12 mL of culture medium). Replate the cells on BM-ECM-coated wells.
-
1.1.
- Generation of 3D self-assembling human heart organoids
-
2.1.Embryoid body (EB) formation:NOTE: It is imperative to limit observable differentiated cells prior to embryoid body formation. Two to three wells of a 6-well plate at a 60–80% confluency will yield enough cells for a single 96-well plate of organoids. All media should be aliquoted and warmed in a 37 °C bead or water bath before any medium changes to minimize temperature shock to the EBs or organoids (this does not include cell dissociation reagents). See Figure 1A, B.
-
2.1.1.Day -2
-
2.1.1.1.To create EBs, on day -2, wash sub-confluent hPSCs (60–80% confluent) with DPBS for at least 10 s to wash any cell debris, and aspirate the DPBS.
-
2.1.1.2.To detach the cells and release them into a single-cell state, add 1 mL of room-temperature cell dissociation reagent (see the Table of Materials) to each well for 3–6 min. Gently tap the plate ~5 times every minute to induce detachment while checking under the microscope. Add 1 mL of PSC medium+Thiaz to stop the reaction.
-
2.1.1.3.To collect the cells and break up any remaining aggregates, pipette the media up and down in the well 2–3 times to generate a single-cell suspension. Transfer the single-cell suspension to a centrifuge tube and spin for 5 min at 300 × g.
-
2.1.1.4.To obtain the desired cell concentration, discard the supernatant and resuspend the cells in 1 mL of PSC medium+Thiaz. Count the cells using a cell counter or hemocytometer and dilute the cells in PSC medium+Thiaz to a concentration of 100,000 cells/mL.
-
2.1.1.5.To distribute the cells for EB formation, use a multichannel pipette to add 100 μL (10,000 cells) to each well of a round-bottom ultra-low attachment 96-well plate. Centrifuge the plate at 100 × g for 3 min and incubate for 24 h at 37 °C, 5% CO2.
-
2.1.1.1.
-
2.1.2.Day -1
-
2.1.2.1.Carefully remove 50 μL of medium from each well and add 200 μL of fresh PSC medium warmed to 37 °C to achieve a final volume of 250 μL per well. Incubate the cells for 24 h at 37 °C, 5% CO2.NOTE: Remove and add medium carefully on the side of the well to avoid disturbing the EBs at the bottom of the well. Due to the delicate nature of the EBs and the suspension culture, it is necessary to leave a small volume of liquid in each well when changing the medium to avoid disturbing the EBs.
-
2.1.2.1.
-
2.1.1.
-
2.2.Human Heart Organoid (hHO) Differentiation:NOTE: All media should be warmed in a 37 °C bead or water bath prior to any media changes. Remove and add medium carefully on the side of the well to avoid disturbing the developing organoids at the bottom of the well. Washes are not needed between media changes to minimize agitation and allow for gradual removal of inhibitors and growth factors. RPMI with 2% B-27 supplement was used throughout the differentiation protocol. B-27 supplement contains insulin unless specified (insulin-free in days 0–5). See Figure 1C.
-
2.2.1.Day 0
-
2.2.1.1.To initiate differentiation towards a mesoderm lineage, remove 166 μL of medium from each well (~2/3rd of total well volume) and add 166 μL of RPMI 1640 containing insulin-free B-27 supplement, 6 μM CHIR99021, 1.875 ng/mL bone morphogenetic protein 4 (BMP4), and 1.5 ng/mL Activin A for a final well concentration of 4 μM CHIR99021, 1.25 ng/mL BMP4, and 1 ng/mL Activin A. Incubate for 24 h at 37 °C, 5% CO2.
-
2.2.1.1.
-
2.2.2.Day 1
-
2.2.2.1.Remove 166 μL of medium from each well and add 166 μL of fresh RPMI 1640 with insulin-free B-27 supplement. Incubate for 24 h at 37 °C, 5% CO2.
-
2.2.2.1.
-
2.2.3.Day 2
-
2.2.3.1.To induce cardiac mesoderm specification, remove 166 μL of medium from each well and add 166 μL of RPMI 1640 containing insulin-free B-27 supplement and 3 μM Wnt-C59 for a final well concentration of 2 μM Wnt-C59. Incubate for 48 h at 37 °C, 5% CO2.
-
2.2.3.1.
-
2.2.4.Day 4
-
2.2.4.1.Remove 166 μL of medium from each well and add 166 μL of fresh RPMI 1640 with insulin-free B-27 supplement. Incubate for 48 h at 37 °C, 5% CO2.
-
2.2.4.1.
-
2.2.5.Day 6
-
2.2.5.1.Remove 166 μL of medium from each well and add 166 μL of RPMI 1640 with B-27 supplement. Incubate for 24 h at 37 °C, 5% CO2.
-
2.2.5.1.
-
2.2.6.Day 7
-
2.2.6.1.To induce proepicardial differentiation, remove 166 μL of medium from each well and add 166 μL of fresh RPMI 1640 containing B-27 supplement and 3 μM CHIR99021 for a final well concentration of 2 μM CHIR99021. Incubate for 1 h at 37 °C, 5% CO2.
-
2.2.6.2.Remove 166 μL of medium from each well and add 166 μL of fresh RPMI 1640 containing B-27 supplement. Incubate for 48 h at 37 °C, 5% CO2.NOTE: Extra caution is advised at this second medium change on day 7 as the organoids are more prone to movement because of the close proximity media changes.
-
2.2.6.3.From day 7 onwards until collection or transfer for analyses or experimentation, perform medium changes every 48 h by removing 166 μL of medium from each well and add 166 μL of fresh RPMI 1640 containing B-27 supplement.NOTE: Organoids are ready for analyses and experimentation at day 15 unless earlier developmental stages are of interest. They can be cultured past day 15 for long-term culture or maturation experiments.
-
2.2.6.1.
-
2.2.1.
-
2.1.
- Organoid analysis
-
3.1.Transferring whole organoids (live or fixed)NOTE: For live organoid transfer, ensure that pipette tips used are sterile.
-
3.1.1.Cut the tip off a P200 pipette tip 5–10 mm from the tip opening, resulting in a wide opening that is ~2–3 mm in diameter.
-
3.1.2.Insert the tip straight into the round-bottom well containing the organoid so that the pipette is completely vertical (perpendicular to the plate). Ensure that the pipette plunger is already pressed all the way before inserting the tip into the medium.
-
3.1.3.Slowly release the pipette plunger, taking up enough medium (100–200 μL) so that the organoid is collected.
-
3.1.4.Transfer the organoid in medium to the target destination (e.g., for fixing, live imaging, electrophysiology recording, new plate culture).
-
3.1.1.
-
3.2.Fixing organoidsNOTE: Fixing and staining organoids can be done either in the 96-well culture plate or in microcentrifuge tubes. Paraformaldehyde (PFA) should be handled only in a fume hood.
-
3.2.1.For fixation in microcentrifuge tubes, transfer live organoids to separate tubes with 1–8 organoids per tube.NOTE: Do not exceed 8 organoids per tube.
-
3.2.2.Carefully remove and discard as much medium from the tube as possible without touching the organoids.
-
3.2.3.Add 4% PFA to each tube or well (300–400 μL per microcentrifuge tube and 100–200 μL per well of a 96-well plate). Incubate at room temperature for 30–45 min.NOTE: Incubation times over 1 h may require antigen retrieval steps and are not recommended.
-
3.2.4.Safely discard the PFA without disturbing the organoids. Perform 3 washes with DPBS supplemented with 1.5 g/L glycine (DPBS/Gly), using the same volume used for the 4% PFA, waiting 5 min between washes. Remove DPBS/Gly and proceed to immunostaining or other analyses or add DPBS and store at 4 °C for future use for up to 2 weeks.NOTE: Storing fixed organoids for longer than 2 weeks may result in tissue degradation and contamination and is not recommended.
-
3.2.1.
-
3.3.Whole mount immunofluorescent staining
-
3.3.1.Add 100 μL of blocking/permeabilization solution (10% normal donkey serum + 0.5% bovine serum albumin (BSA) + 0.5% Triton X-100 in 1x DPBS) to each well or tube containing the fixed organoids. Incubate at room temperature overnight on a shaker.NOTE: Do not exceed 8 organoids per tube.
-
3.3.2.Carefully remove and discard as much of the blocking solution as possible without touching the organoids. Perform 3 washes with DPBS, waiting 5 min between washes.
-
3.3.3.Prepare the primary antibody solution (1% normal donkey serum + 0.5% BSA + 0.5% Triton X-100 in 1x DPBS) with the desired primary antibodies at the recommended concentrations. Incubate at 4 °C for 24 h on a shaker.
-
3.3.4.Carefully remove and discard as much of the antibody solution as possible without touching the organoids. Perform 3 washes with DPBS, waiting 5 min between washes.
-
3.3.5.Prepare secondary antibody solution (1% normal donkey serum + 0.5% BSA + 0.5% Triton X-100 in 1x DPBS) with the desired secondary antibodies at the recommended concentrations. If the antibodies are fluorescently labeled, incubate at 4 °C in the dark (e.g., covered in aluminum foil) for 24 h on a shaker.
-
3.3.6.Carefully remove and discard as much of the antibody solution as possible without touching the organoids. Perform 3 washes with DPBS, waiting 5 min between washes.
-
3.3.7.Prepare slides with beads (90–300 μm in diameter) mounted in a mounting medium (see the Table of Materials) near the edges of the slide where the coverslip with the organoids will be placed.NOTE: It is recommended to allow the mounting medium around the beads dry before proceeding; this will prevent the beads from moving around. See Figure 2.
-
3.3.8.Transfer the stained organoids using a cut pipette tip onto the slide in between the beads, ensuring spacing to avoid touching of the organoids once on the slide. Use the corner of a rolled up laboratory wipe to carefully remove excess liquid around the organoid.
-
3.3.9.Cover the organoids with mounting-clearing medium (fructose–glycerol clearing solution is 60% (vol/vol) glycerol and 2.5 M fructose)37 using 120–150 μL of the mounting-clearing medium per slide.NOTE: It is recommended to use a cut pipette tip when working with the mounting-clearing medium as it is very viscous.
-
3.3.10.Hover the coverslip over the slide with the organoids covered with mounting-clearing solution and slowly press the coverslip over the slide, ensuring the organoids are between the mounted beads.
-
3.3.11.Seal the perimeter of the coverslip on the slide using top coat nail varnish. Allow the slide to dry in the dark at room temperature for 1 h. Store at 4 °C in the dark for long-term storage.
-
3.3.1.
-
3.4.Calcium transient imaging in live heart organoidsNOTE: Fluo4-AM was reconstituted in dimethyl Sulfoxide (DMSO) according to the manufacturer’s instructions to a final stock solution concentration of 0.5 mM. Fluo4-AM was added directly to the organoid well in the 96-well plate.
-
3.4.1.Perform 2 washes on the organoids using RPMI 1640 medium.
-
3.4.1.1.Remove 166 μL of the spent medium from the well.
-
3.4.1.2.Add 166 μL of warmed RPMI 1640 medium, remove 166 μL of medium, and add 166 μL of fresh RPMI 1640 medium.NOTE: The washes are done to remove waste material and cell debris. 2/3 of the media is removed from the wells during the washes to prevent disturbance of the organoids at the bottom of the well and not agitate them before the functional assay.
-
3.4.1.1.
-
3.4.2.Add Fluo4-AM medium to the organoids.
-
3.4.2.1.Add Fluo4-AM reconstituted in DMSO to RPMI 1640 containing B-27 supplement to prepare a 1.5 μM solution.
-
3.4.2.2.Remove 166 μL of medium from the well.
-
3.4.2.3.Add 166 μL of 1.5 μM Fluo4-AM in RPMI 1640 containing B-27 supplement for a final well concentration of 1 μM. Incubate at 37 °C, 5% CO2 for 30 min.
-
3.4.2.1.
-
3.4.3.Perform 2 washes as in step 3.4.1.
-
3.4.4.Add 166 μL of RPMI 1640 containing B-27 supplement to the well.
-
3.4.5.Using a cut tip of a P200 pipette tip, transfer the organoid to a glass-bottom Petri dish (e.g., 8-well chambered cover glass with #1.5 high performance cover glass) with 100–200 μL of medium.NOTE: See section 3.1 on transferring whole organoids, above.
-
3.4.6.Image the organoids live under a microscope with a temperature- and CO2-controlled chamber at 37 °C, 5% CO2.
-
3.4.7.Record several 10–20 s videos at various locations across the organoid, showing the increase and decrease in fluorescence intensity levels as the calcium enters and exits the cells.NOTE: For high-resolution recordings, it is recommended to record at a speed of 10 fps or faster, 50 fps is recommended.
-
3.4.8.Analyze the videos using an image analysis software (e.g., ImageJ) by selecting regions of interest and measuring the levels of intensity over time.
-
3.4.9.Normalize the intensity recordings using ΔF/F0 vs. time in milliseconds and plot.
-
3.4.1.
-
3.1.
Table of Materials
| Name of Material/Equipment | Company | Catalog Number | Comments/Description |
|---|---|---|---|
| Antibodies | |||
| TNNT2 | Abcam | ab8295 | Mouse; 1:200 |
| HAND1 | Abcam | ab196622 | Rabbit; 1:200 |
| HAND2 | Abcam | ab200040 | Rabbit; 1:200 |
| WT1 | Abcam | ab89901 | Rabbit; 1:200 |
| TJP1 | Invitrogen | PA5–19090 | Goat; 1:250 |
| NFAT2 | Abcam | ab25916 | Rabbit; 1:100 |
| PECAM1 | DSHB | P2B1 | Rabbit; 1:50 |
| THY1 | Abcam | ab133350 | Rabbit; 1:200 |
| VIM | Abcam | ab11256 | Goat; 1:250 |
| Alexa Fluor 488 Donkey anti- mouse | Invitrogen | A-21202 | 1:200 |
| Alexa Fluor 488 Donkey anti- rabbit | Invitrogen | A-21206 | 1:200 |
| Alexa Fluor 594 Donkey anti- mouse | Invitrogen | A-21203 | 1:200 |
| Alexa Fluor 594 Donkey anti- rabbit | Invitrogen | A-21207 | 1:200 |
| Alexa Fluor 647 Donkey anti- goat | Invitrogen | A32849 | 1:200 |
| Media & Reagents | |||
| Essential 8 Flex Medium Kit | Gibco | A2858501 | Medium containing 1% penicillin-streptomycin |
| DMEM/F12 | Gibco | 10566016 | |
| Penicillin-Streptomycin | Gibco | 15140122 | |
| Matrigel GFR | Corning | CB40230 | |
| ReLeSR | Stem Cell Technologies | NC0729236 | |
| Accutase | Innovative Cell Technologies | NC9464543 | |
| Thiazovivin | Millipore Sigma | SML1045 | |
| Phosphate Buffer Solution | Gibco | 10010049 | |
| Phosphate Buffer Solution (10x) | Gibco | 70011044 | |
| RPMI 1640 | Gibco | 11875093 | |
| B-27 Supplement | Gibco | 17504–044 | |
| B-27 Supplement (Minus Insulin) | Gibco | A1895601 | |
| CHIR-99021 | Selleck | 442310 | |
| BMP-4 | Gibco | PHC9534 | |
| Activin A | R&D Systems | 338AC010 | |
| WNT-C59 | Selleck | NC0710557 | |
| Dimethyl Sulfoxide | Millipore Sigma | D2650 | |
| DAPI | Thermo Scientific | 62248 | 1:1000 |
| Fluo4-AM | Invitrogen | F14201 | |
| Paraformaldehyde | MP Biomedicals | IC15014601 | Powder dissolved in PBS Buffer – use at 4% |
| Glycine | Millipore Sigma | 410225 | |
| Normal Donkey Serum | Millipore Sigma | S30–100mL | |
| Triton X-100 | Millipore Sigma | T8787 | |
| Bovine Serum Albumin | Bioworld | 50253966 | |
| D-(-)-Fructose | Millipore Sigma | F0127 | |
| Glycerol | Millipore Sigma | G5516 | |
| VECTASHIELD Vibrance Antifade Mounting Medium | Vector Laboratories | H170010 | |
| Polybead Microspheres | Polysciences, Inc. | 73155 | 90 μm |
| Trypan Blue Solution | Gibco | 1525006 | |
| Other | |||
| 96-well Clear Ultra Low Attachment Microplates | Costar | 07201680 | |
| Moxi Cell Counter | Orflo Technologies | MXZ001 | |
| Moxi Z Cell Count Cassette – Type M | Orflo Technologies | MXC001 | |
| 8 Well chambered cover Glass with #1.5 high performance cover glass | Cellvis | C8–1.5H-N | |
| 15mL Falcon Tubes | Fisher Scientific | 1495970C | |
| 1.5 Microcentrifuge Tubes | Fisher Scientific | 02682002 | |
| 6-Well Flat Bottom Cell Culture Plates | Corning | 0720083 |
Figure 1: Embryoid body generation and heart organoid differentiation steps.
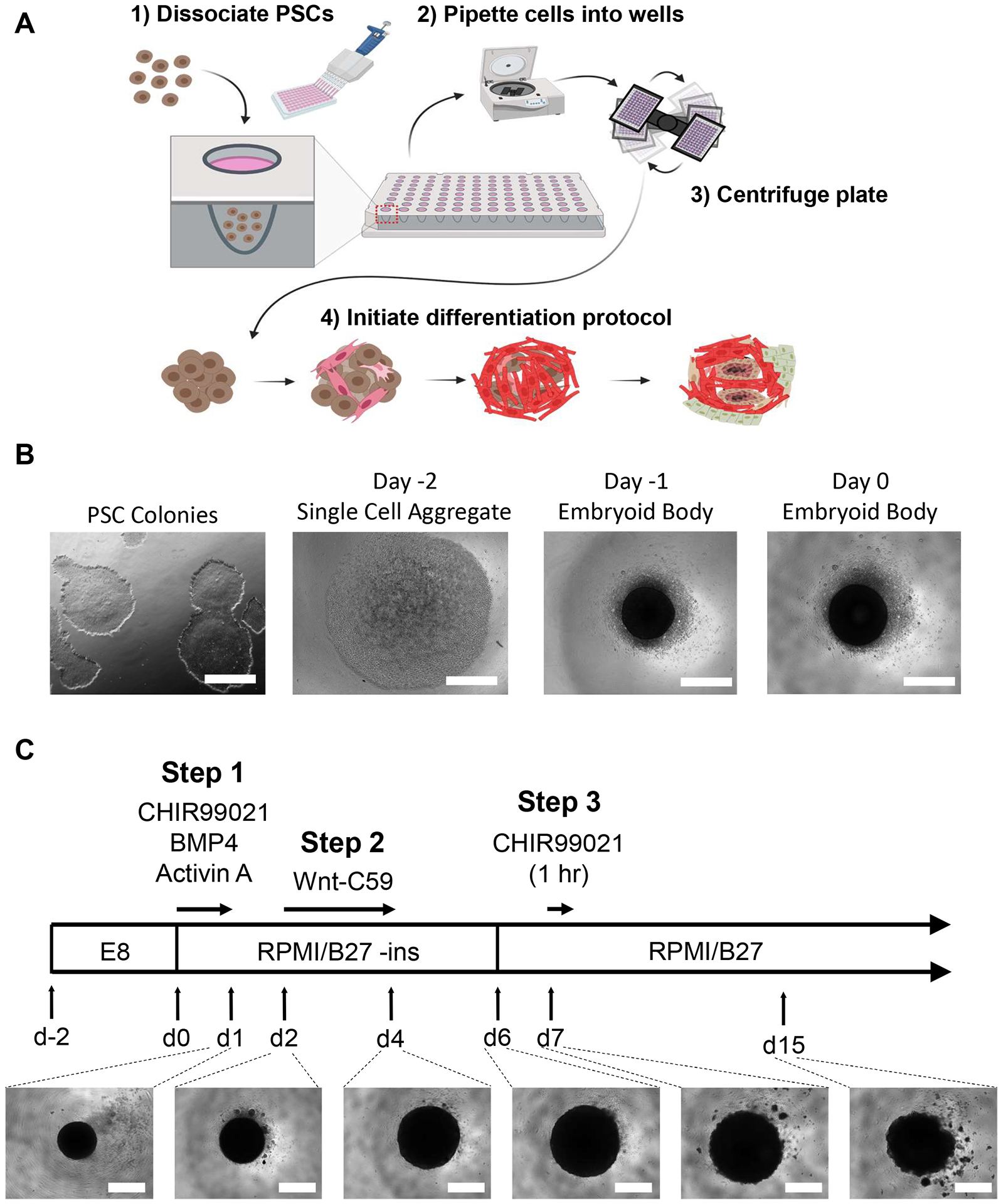
(A) (1–2) Dissociated cells are seeded into wells of a 96-well ultra-low attachment plate via a multichannel pipette. (3) The 96-well plate is then centrifuged, which allows the cells to aggregate in the center. (4) Over time, following the addition of growth factors and pathway modulators, the embryoid body begins to differentiate into several cardiac lineages and form spatially and physiologically relevant distinct cell populations surrounding internal microchambers. (B) Representative images of the progression of embryoid body generation, beginning with 2-dimensional iPSC culture (left) and ending with a Day 0 embryoid body (right); scale bar = 500 μm. (C) Summary of human heart organoid differentiation protocol, including chemical pathway modulators and inhibitors with respective timepoints, durations, and developing organoid images under light microscopy from day 1 to day 15; scale bar = 500 μm.
Figure 2: Whole organoid mounting on slides for imaging.
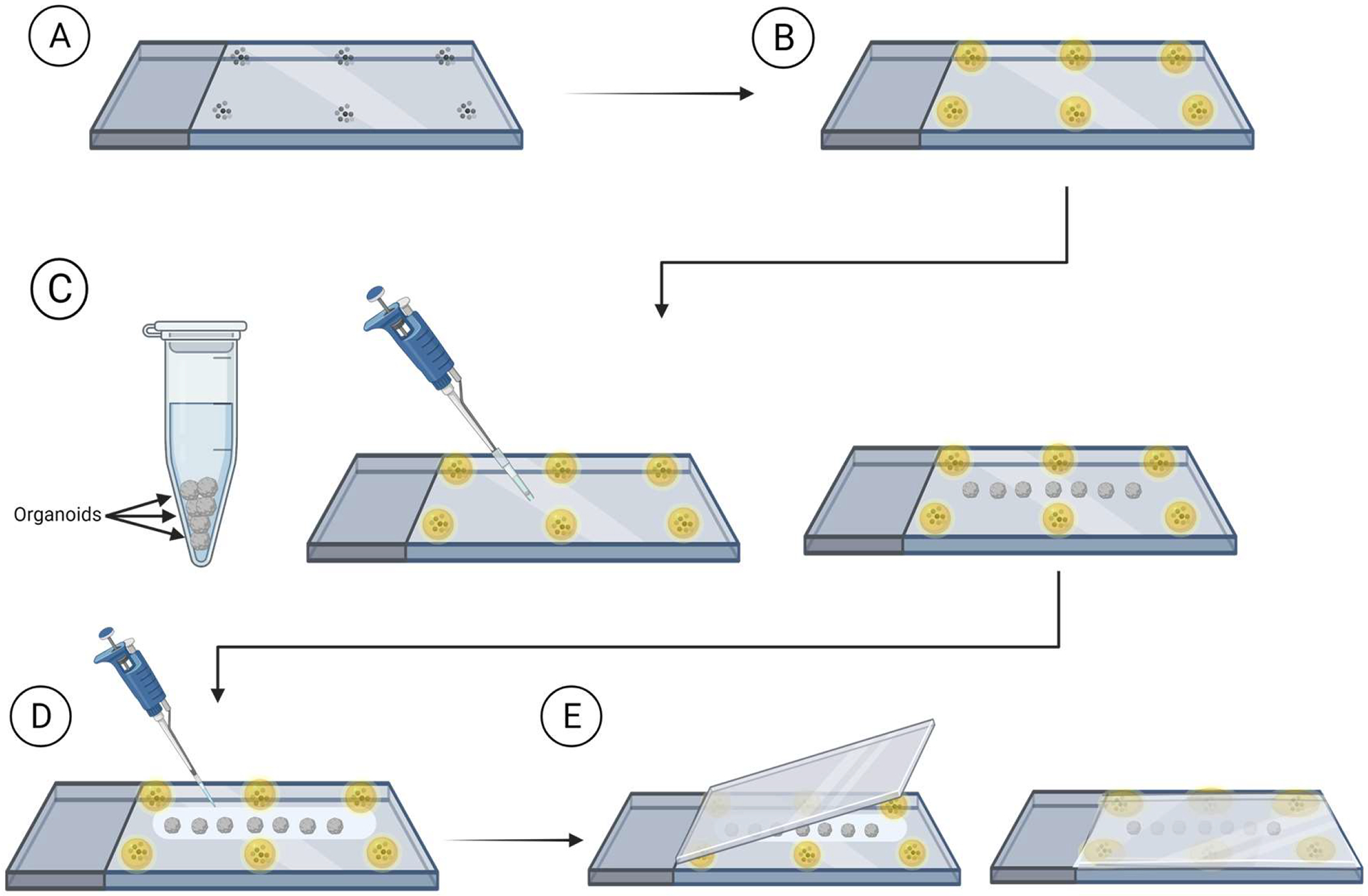
Steps for preparing slides and mounting organoids for imaging. (A) Placement of microbeads onto the periphery of a glass slide. (B) Covering the microbeads with mounting medium. (C) Transferring organoids onto the slide between the beads and removing excess liquid surrounding the organoids. (D) Covering the organoids with clearing/mounting medium. (E) Placing the coverslip on top of the slide with organoids and beads.
REPRESENTATIVE RESULTS:
To achieve self-organizing hHO in vitro, we modified and combined differentiation protocols previously described for 2D monolayer differentiation of cardiomyocytes21 and epicardial cells22 using Wnt pathway modulators and for 3D precardiac organoids16 using the growth factors BMP4 and Activin A. Using the 96-well plate EB and hHO differentiation protocol described here and shown in Figure 1, the concentrations and exposure durations of the Wnt pathway activator CHIR99021 were optimized to yield highly reproducible and complex hHOs derived from human PSCs. hPSCs or hESCs cultured in PSC medium to 60–80% confluency in colony-like formation with minimal to no visible differentiation are ideal for EB generation (Figure 1B).
EBs were allowed to incubate for 48 h with a medium change after 24 h, before starting differentiation at day 0. On day 0, the EBs should appear as a spherical dark aggregate at the center of each well, under a light microscope (Figure 1B). The differentiation protocol starts on day 0 with the Wnt pathway activation and growth factor addition for exactly 24 h. This mesoderm induction followed by a cardiac mesoderm induction on day 2 using the Wnt pathway inhibitor Wnt-C59 will result in a significant enlargement of the organoid from ~200 μm in diameter to 500–800 μm in diameter at day 4 and to as much as 1 mm (organoids may experience a slight reduction in size by day 15 (Figure 1C)). The hHO will begin beating as early as day 6 (Video 1), with 100% of the organoids showing visible beating by day 10 (Video 2) (unless undergoing drug treatment or if inadequate hPSCs were used to generate the EBs). This has been observed in 5 distinct hPSC cell lines23.
To evaluate the capacity of the hHOs to represent various steps of the physiological development of the heart, we collected organoids at various time points throughout the differentiation protocol and looked for the presence and transcriptomic expression of heart field markers. Immunofluorescent staining for the first heart field (FHF) marker, HAND1, and the second heart field (SHF) marker, HAND2, revealed their nuclear presence in these cardiac progenitor cells arising at around day 3 and day 5, respectively (Figure 3A).
Figure 3: First heart field and second heart field specification in hHOs recapitulates physiological human heart development.
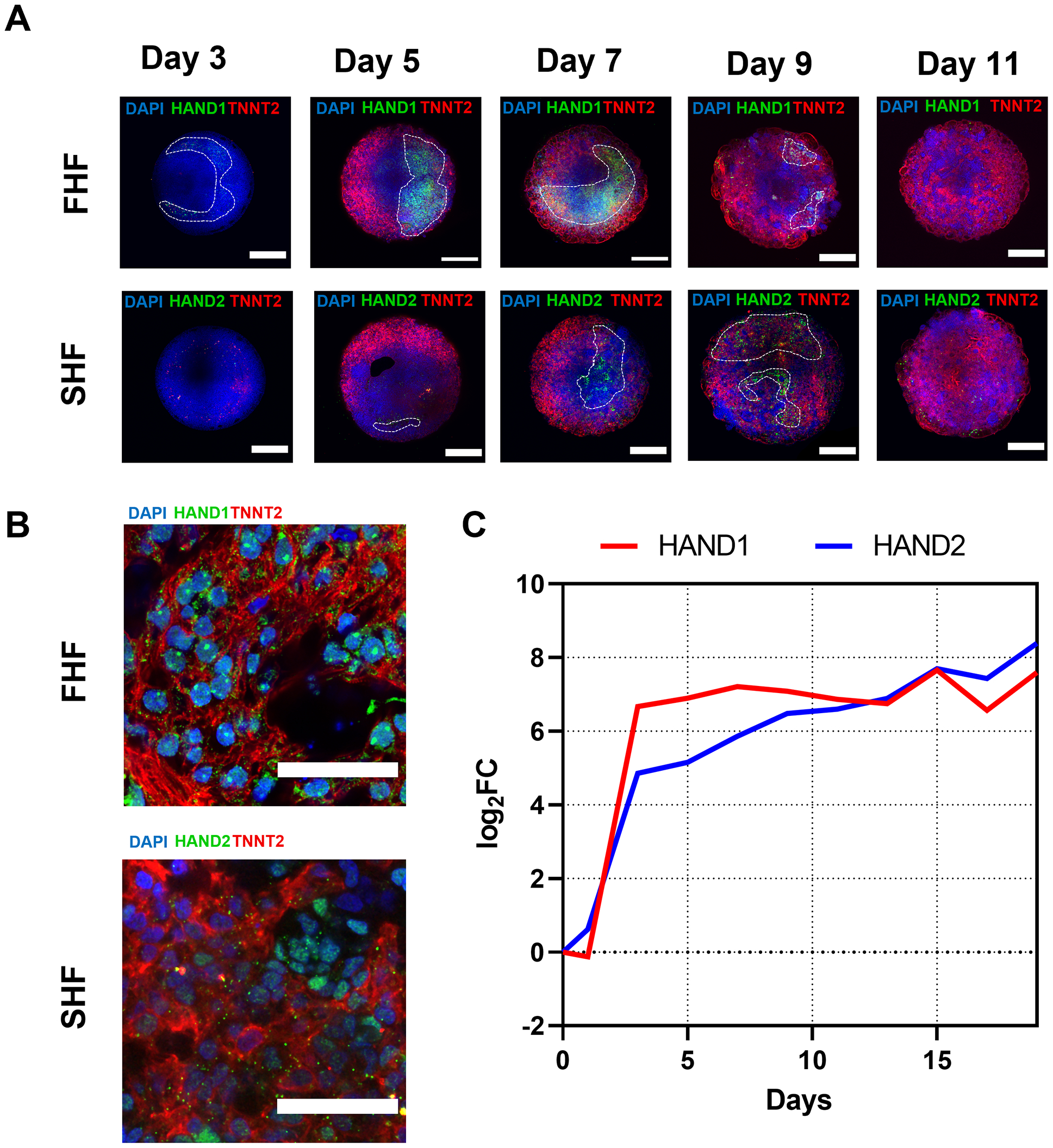
(A) Confocal immunofluorescence images of day 3 to day 11 hHOs showing the formation of FHF (HAND1, top) and SHF (HAND2, bottom), and cardiomyocytes (TNNT2) and nuclear dye DAPI; scale bars = 500 μm. (B) High-magnification images of day 7 organoids showing co-localization of HAND1 and HAND2 with the cardiomyocyte marker TNNT2; scale bars = 50 μm. (C) RNA-Seq gene expression profiles of FHF marker HAND1 (red) and SHF marker HAND2 (blue) from day 0 to day 19. Abbreviations: hHOs = human heart organoids; FHF = first heart field; SHF = second heart field; HAND = heart and neural crest derivatives expressed; TNNT2 = cardiac troponin T2; DAPI = 4’,6-diamidino-2-phenylindole.
The expression of both markers happens at regions of the organoids that diminish in size after day 7 for the FHF and after day 9 for the SHF. Interestingly, high-magnification images of day 7 organoids revealed the majority of HAND1-expressing cells were cardiomyocyte in origin (as shown by the cardiomyocyte-specific marker TNNT2), while many of the HAND2-expressing cells did not express the cardiomyocyte marker (Figure 3B). This observation is in agreement with the precardiac organoids derived from mouse ESCs demonstrating the development of non-myocyte cells from SHF progenitor cells16. It is important to note that the RNA-Sequencing data show that the RNA transcripts for both HAND1 and HAND2 were expressed from day 3 onwards, with the FHF marker being more highly expressed between days 3 and 11 and the SHF marker being more highly expressed after day 13 (Figure 3C).
Immunofluorescence staining revealed the presence of markers of various cell-type lineages that make up the human heart. Myocardial tissue (identifiable using the cardiomyocyte-specific marker TNNT2) adjacent to epicardial tissue (marked by the nuclear transcription factor WT1 and the epithelial membrane marker TJP1) (Figure 4A). Endocardial cells expressing NFATC1 were detected lining the walls of internal chamber-like structures within the organoids (Figure 4B). Endothelial cells in a vessel-like network can be seen as early as day 13 of differentiation (Figure 4C). Lastly, we report the presence of cardiac fibroblasts intermixed all throughout the organoid (Figure 4D). These cell-type markers were also observed in the RNA-Seq gene expression profiles (Figure 4E). The composition of cell types in the organoids as measured by area of the organoid they occupy, were found to be ~58% cardiomyocytes with the rest comprising of non-myocyte cardiac cells, including epicardial cells (~15%), endocardial cells (~13%), cardiac fibroblasts (~12%), and endothelial cells (~1%) (Figure 4F).
Figure 4: hHOs develop multiple cardiac lineages.
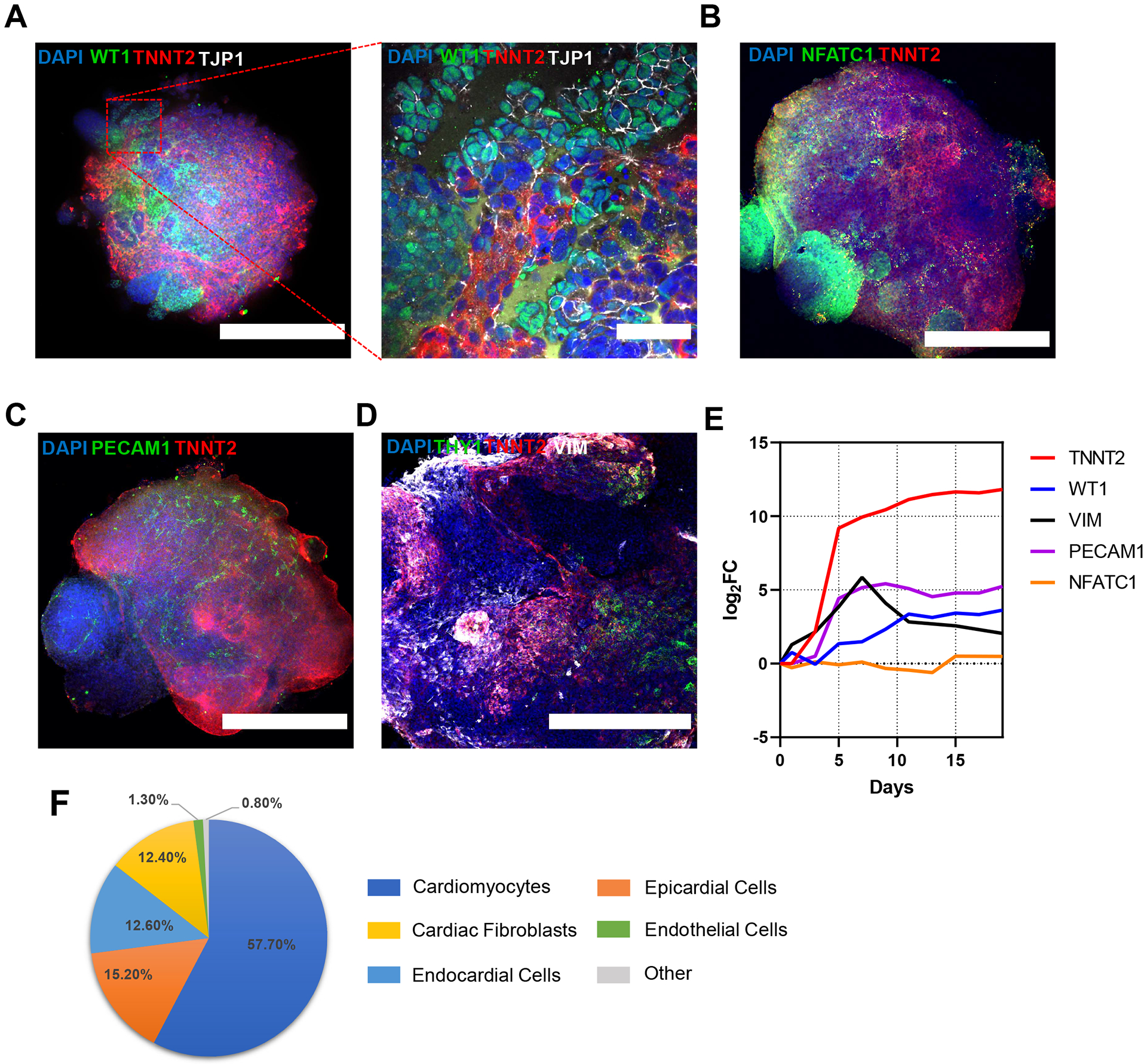
(A–D) Confocal immunofluorescence images of day 15 hHOs showing the formation of cardiomyocytes (TNNT2) and staining with nuclear dye DAPI in non-myocyte cardiac cells. (A) Whole organoid and high magnification of epicardial marker WT1 (green) and epithelial membrane marker TJP1 (white) showing epicardial cells of epithelial origin on top and adjacent to myocardial tissue; scale bar = 500 μm, inset = 50 μm. (B) Endocardial marker NFATC1 (green) expression on the lining of chambers; scale bar = 500 μm. (C) Endothelial vessel network in hHOs shown by PECAM1 (green). Scale bar = 500 μm. (D) Cardiac fibroblasts markers THY1 and VIM shown in green and white, respectively, distributed throughout the organoid. Scale bar = 500 μm. (E) RNA-Seq gene expression profiles of the major cell types present in the hHOs from days 0 to 19 of differentiation. Abbreviations: hHOs = human heart organoids; TNNT2 = cardiac troponin T2; DAPI = 4’,6-diamidino-2-phenylindole; WT1 = Wilm’s tumor-1 transcription factor; NFATC1 = cytoplasmic nuclear factor of activated T cell; PECAM1 = platelet endothelial cell adhesion molecule-1; VIM = vimentin. (F) Pie chart of average tissue type composition in hHOs, calculated as the percentage area with respective cell marker over entire organoid by nuclei staining across three z-planes throughout each organoid using ImageJ.
Electrophysiological function of the organoids was measured by live calcium imaging of individual cells in whole organoids. Fluo-4 fluorescence intensity varies over time due to calcium entry and exit from the cell, revealing regular action potentials (Figure 5A). Heatmaps showing calcium intensities over a high-magnification region of the organoid show the increased intensity because of calcium transients in individual cells (Figure 5B and Video 3).
Figure 5: Fluo-4 live calcium transient recordings in live human heart organoids.
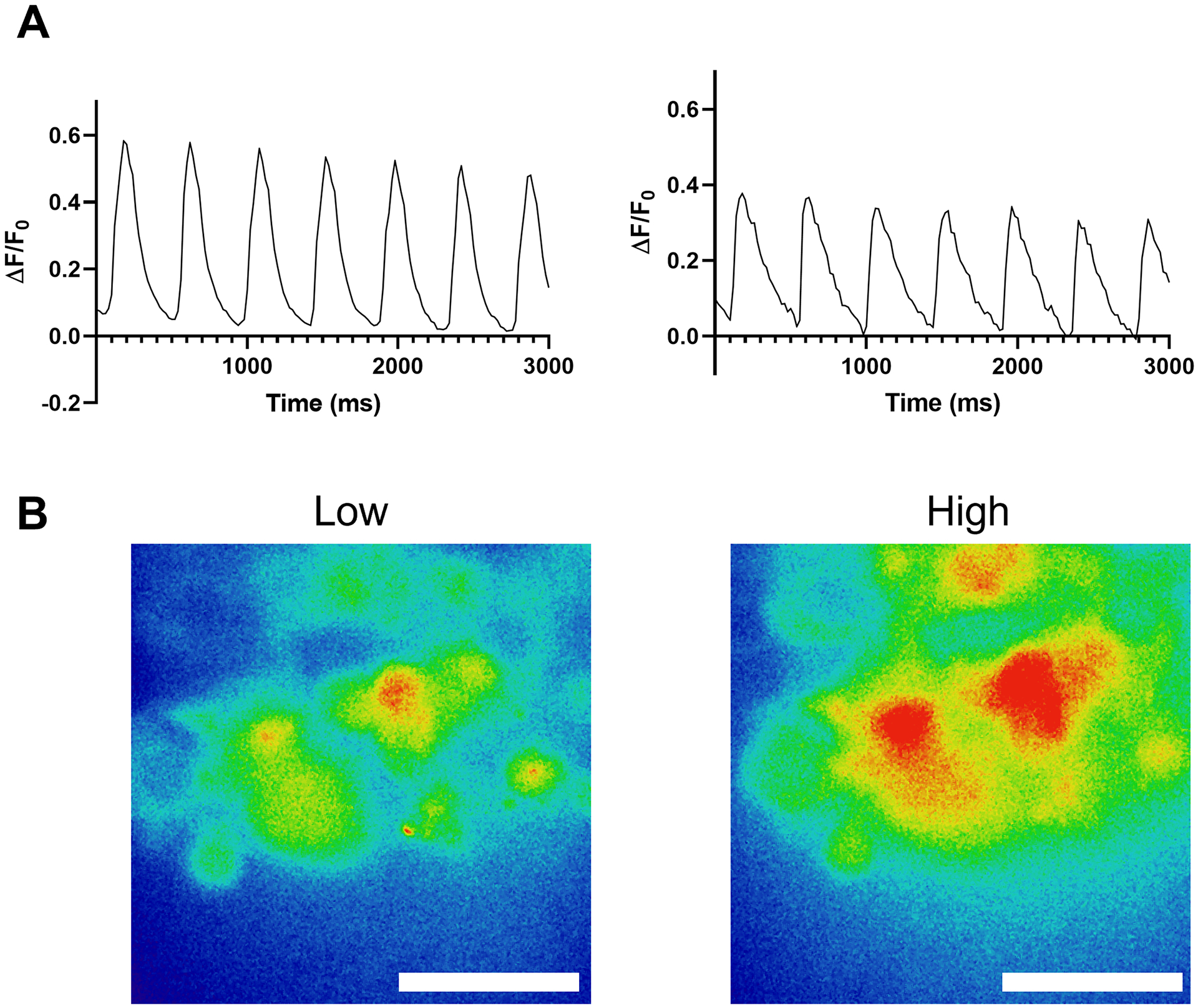
(A) Representative calcium transient recordings of individual cardiomyocytes within whole organoids. (B) Heatmap showing low and peak calcium levels between action potentials as determined by Fluo-4 intensity; scale bars = 10 μm.
DISCUSSION:
Recent advances in human stem cell-derived cardiomyocytes and other cells of cardiac origin have been used to model human heart development22,24,25 and disease26–28 and as tools to screen therapeutics29,30 and toxic agents31,32. Here, we report an easy-to-implement, highly reproducible protocol to generate and differentiate EBs into highly complex hHOs. This protocol has been successful in multiple cell lines, including hPSCs and hESCs23, showing consistent beating frequencies and cell type organization. This protocol draws aspects from previously described protocols for cardiomyocyte differentiation24, epicardial cell differentiation22, and precardiac organoids derived from mouse ESCs16 and optimizes the step-wise modulation of canonical WNT signaling using chemical inhibitors and growth factors in a fully defined medium. Several optimization methodologies were employed in the generation of this protocol.
First, the chemical inhibitor concentrations and exposure durations as well as the addition of growth factors have been optimized for the 3D environment and are discussed in previous work23. These were optimized to elucidate structures with both physiological complexity and representation of the in vivo human heart, with physiological composition and ratios of cardiomyocytes to non-myocyte cardiac cell types (epicardial cells, cardiac fibroblasts). Second, the two-thirds medium change strategy allows minimal agitation of the EBs/organoids, as they sit in suspension near the bottom of the well, while also facilitating a gradient exposure to chemical inhibitors and growth factors when the medium is refreshed. The combination of cardiac mesoderm differentiation through Wnt pathway activation, followed by inhibition24, and the subsequent induction of proepicardial specification via a second Wnt pathway activation22, allows for a single protocol to yield highly complex hHOs. The organoids grow up to 1 mm after 15 days of differentiation and can be easily transferred for live or fixed analyses and assays. Third, given the large size of the organoids, the use of microbeads or other similar structures to maintain space between the slide and coverslip were found to better preserve the 3D structure of the organoids and improve the imaging process.
This developing human heart model allows access to otherwise inaccessible stages of heart development, such as early first and second heart field specification—observed between days 3 and 9 of differentiation—and organization into cardiac progenitor cells that give rise to heart tissues, including the myocardium, endocardium, epicardium, endothelial vasculature, and supporting cardiac fibroblasts, which were observed on day 15 of differentiation. The tissue types present in the heart organoids derived from this protocol are highly representative of the human fetal heart in both composition33 and transcriptomic profile23,34 and can therefore facilitate tissue-tissue and cell-cell higher order interactions resembling that of the in vivo heart. This protocol was found to be highly efficient and reproducible across experiments and cell lines, yielding organoids that comprise of mostly of cardiomyocytes and included non-myocyte cardiac cells, such as epicardial cells endocardial cells, cardiac fibroblasts, and endothelial cells, representing physiological composition23,33,35.
Analyses of the ultrastructure of the forming cardiomyocytes via transmission electron microscopy and the development of chambers and a vascular network via optical coherence tomography and confocal imaging are discussed in detail in previous work23. A great advantage of this heart organoid protocol over other existing protocols recently published17–20,36,37 is the robust formation of an endothelial network throughout the organoid, allowing the ability to investigate vascular development and disease in the early human heart, without the need of further external inductions to the protocol. Lastly, functional analysis of the heart organoids is achievable through various approaches, including the use of a calcium-sensitive dye to track the calcium transients in the cardiomyocytes across the organoid. Using high-resolution microscopy, we recorded the fluorescence intensity of calcium entering and exiting cells and observed highly representative action potentials. Other possible functional analyses methods include the use of a transgenic line with a calcium-sensitive indicator or direct recording using a microelectrode array23.
The heart organoids described here are recapitulative of the developing human fetal heart, yet are limited in demonstrating more mature, adult-like features. Future protocols may build on the protocol described here to induce maturation in these organoids and yield constructs that better model the adult heart. Moreover, this protocol is designed to create miniature models of the human heart and is limited to research studies of heart development and disease or for pharmaceuticals screening and may not be suitable as a means of clinical intervention such as replacement of heart tissue via transplantation. Overall, we describe here an easy-to-follow and cost-effective protocol to generate highly reproducible and sophisticated human heart organoids that can facilitate research studies in human heart development, disease etiology, and pharmacological screening.
Supplementary Material
Video 1: Live imaging of representative organoid derived from hPSCs at day 6 of differentiation under light microscopy at room temperature. Abbreviation: hPSC = human pluripotent stem cell.
Video 2: Live imaging of representative organoid derived from hPSCs at day 15 of differentiation under light microscopy at room temperature. Abbreviation: hPSC = human pluripotent stem cell.
Video 3: Live recording of day 10 organoid showing heatmap of calcium transients under fluorescence microscope.
ACKNOWLEDGMENTS:
This work was supported by the National Heart, Lung, and Blood Institute of the National Institutes of Health under award numbers K01HL135464 and R01HL151505 and by the American Heart Association under award number 19IPLOI34660342. We wish to thank the MSU Advanced Microscopy Core and Dr. William Jackson at the MSU Department of Pharmacology and Toxicology for access to confocal microscopes, the IQ Microscopy Core, and the MSU Genomics Core for sequencing services. We also wish to thank all members of the Aguirre Lab for their valuable comments and advice.
Footnotes
DISCLOSURES:
The authors have no conflicts of interest to declare.
REFERENCES:
- 1.Hoffman JIE & Kaplan S The incidence of congenital heart disease. J. Am. Coll. Cardiol 39, 1890–1900 (2002). [DOI] [PubMed] [Google Scholar]
- 2.Wu W, He J & Shao X Incidence and mortality trend of congenital heart disease at the global, regional, and national level, 1990–2017. Medicine (Baltimore). 99, e20593 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Fahed AC, Gelb BD, Seidman JG & Seidman CE Genetics of congenital heart disease: The glass half empty. Circ. Res 112, 707–720 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lancaster MA et al. Cerebral organoids model human brain development and microcephaly. Nature 501, 373–379 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mansour AA et al. An in vivo model of functional and vascularized human brain organoids. Nat. Biotechnol 36, 432–441 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Homan KA et al. Flow-enhanced vascularization and maturation of kidney organoids in vitro. Nat. Methods 16, 255–262 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Uchimura K, Wu H, Yoshimura Y & Humphreys BD Human Pluripotent Stem Cell-Derived Kidney Organoids with Improved Collecting Duct Maturation and Injury Modeling. Cell Rep. 33, 108514 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Serra D et al. Self-organization and symmetry breaking in intestinal organoid development. Nature 569, 66–72 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mithal A et al. Generation of mesenchyme free intestinal organoids from human induced pluripotent stem cells. Nat. Commun 11, 1–15 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Porotto M et al. Authentic Modeling of Human Respiratory Virus Infection in Human Pluripotent Stem Cell-Derived Lung Organoids. MBio 10, e00723–19 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dye BR et al. In vitro generation of human pluripotent stem cell derived lung organoids. Elife 4, e05098 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mun SJ et al. Generation of expandable human pluripotent stem cell-derived hepatocyte-like liver organoids. J. Hepatol 71, 970–985 (2019). [DOI] [PubMed] [Google Scholar]
- 13.Vyas D et al. Self-assembled liver organoids recapitulate hepatobiliary organogenesis in vitro. Hepatology 67, 750–761 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dossena M et al. Standardized GMP-compliant scalable production of human pancreas organoids. Stem Cell Res. Ther 11, 94 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Georgakopoulos N et al. Long-term expansion, genomic stability and in vivo safety of adult human pancreas organoids. BMC Dev. Biol 20, 4 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Andersen P et al. Precardiac organoids form two heart fields via Bmp/Wnt signaling. Nat. Commun 9, 1–13 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rossi G et al. Capturing Cardiogenesis in Gastruloids. Cell Stem Cell 1–11 (2020) doi: 10.1016/j.stem.2020.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lee J et al. In vitro generation of functional murine heart organoids via FGF4 and extracellular matrix. Nat. Commun 11, 4283 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Drakhlis L et al. Human heart-forming organoids recapitulate early heart and foregut development. Nat. Biotechnol 1–10 doi: 10.1038/s41587-021-00815-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hofbauer P et al. Cardioids reveal self-organizing principles of human cardiogenesis. Cell (2021) doi: 10.1016/j.cell.2021.04.034. [DOI] [PubMed] [Google Scholar]
- 21.Bao X et al. Directed differentiation and long-term maintenance of epicardial cells derived from human pluripotent stem cells under fully defined conditions. Nat. Protoc 12, 1890–1900 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bao X et al. Long-term self-renewing human epicardial cells generated from pluripotent stem cells under defined xeno-free conditions. Nat. Biomed. Eng 1, 1–12 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lewis-Israeli YR et al. Self-assembling human heart organoids for the modeling of cardiac development and congenital heart disease. Nat. Commun 12, 5142 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lian X et al. Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc. Natl. Acad. Sci. U. S. A 109, E1848–E1857 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Burridge PW, Keller G, Gold JD & Wu JC Production of de novo cardiomyocytes: Human pluripotent stem cell differentiation and direct reprogramming. Cell Stem Cell 10, 16–28 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hashem SI et al. Impaired mitophagy facilitates mitochondrial damage in Danon disease. J. Mol. Cell. Cardiol 108, 86–94 (2017). [DOI] [PubMed] [Google Scholar]
- 27.Sun N et al. Patient-specific induced pluripotent stem cells as a model for familial dilated cardiomyopathy. Sci. Transl. Med 4, 130ra47–130ra47 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Stroud MJ et al. Luma is not essential for murine cardiac development and function. Cardiovasc. Res 114, 378–388 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Liang P et al. Drug screening using a library of human induced pluripotent stem cell-derived cardiomyocytes reveals disease-specific patterns of cardiotoxicity. Circulation 127, 1677–1691 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mills RJ et al. Functional screening in human cardiac organoids reveals a metabolic mechanism for cardiomyocyte cell cycle arrest. Proc. Natl. Acad. Sci. U. S. A 114, E8372–E8381 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Braam SR et al. Prediction of drug-induced cardiotoxicity using human embryonic stem cell-derived cardiomyocytes. Stem Cell Res. 4, 107–116 (2010). [DOI] [PubMed] [Google Scholar]
- 32.Burridge PW et al. Human induced pluripotent stem cell–derived cardiomyocytes recapitulate the predilection of breast cancer patients to doxorubicin-induced cardiotoxicity. Nat. Med 22, 547–556 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Pinto AR et al. Revisiting Cardiac Cellular Composition Alexander. Circ Res. 118, 400–409 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bertero A et al. Dynamics of genome reorganization during human cardiogenesis reveal an RBM20-dependent splicing factory. Nat. Commun 10, 1538 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Scott F. Gilbert, M. J. FB Lateral Plate Mesoderm: Heart and Circulatory System. in Developmental Biology 591–610 (Sinauer Associates is an imprint of Oxford University Press, 2019). [Google Scholar]
- 36.Richards DJ et al. Human cardiac organoids for the modelling of myocardial infarction and drug cardiotoxicity. Nat. Biomed. Eng 4, 446–462 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lewis-Israeli YR, Wasserman AH & Aguirre A Heart Organoids and Engineered Heart Tissues: Novel Tools for Modeling Human Cardiac Biology and Disease. Biomolecules 11, 1277 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Video 1: Live imaging of representative organoid derived from hPSCs at day 6 of differentiation under light microscopy at room temperature. Abbreviation: hPSC = human pluripotent stem cell.
Video 2: Live imaging of representative organoid derived from hPSCs at day 15 of differentiation under light microscopy at room temperature. Abbreviation: hPSC = human pluripotent stem cell.
Video 3: Live recording of day 10 organoid showing heatmap of calcium transients under fluorescence microscope.