

Abstract
目的
探讨人羊膜间充质干细胞(human amniotic mesenchymal stem cells,hAMSCs)是否具有 MSCs 特性,以及经 TGF-β1 和 VEGF 联合诱导后是否具有向韧带成纤维细胞分化的能力。
方法
取自愿捐赠的足月产妇胎盘,采用胰蛋白酶胶原酶联合消化法分离培养 hAMSCs,流式细胞术检测 hAMSCs 表型分子,免疫荧光染色检测 hAMSCs 其角蛋白-19(cytokeratin-19,CK-19)和波形蛋白表达情况。取第 3 代 hAMSCs,分别使用含 TGF-β1 和 VEGF 的 L-DMEM/F12 成韧带或纤维细胞诱导培养基(实验组)和普通 L-DMEM/F12 培养基(对照组)培养,细胞增殖-毒性检测试剂盒(cell counting kit-8,CCK-8)检测两组细胞增殖能力;培养 5、10、15 d 分别采用免疫荧光染色及实时荧光定量 PCR 检测韧带及血管生成相关特异性蛋白和基因表达。
结果
倒置相差显微镜观察示 hAMSCs 呈单层贴壁生长;流式细胞术结果示 hAMSCs 表达 MSCs 表型分子;免疫荧光染色示 hAMSCs 高表达波形蛋白、低表达 CK-19;hAMSCs 具有向成骨、成软骨及成脂细胞分化的能力。CCK-8 法检测示,7 d 时两组细胞均达增殖高峰,实验组细胞增殖能力于 7 d 后显著高于对照组(P<0.05)。免疫荧光染色示,培养 5、10、15 d 时实验组 Ⅰ 型胶原、Ⅲ 型胶原、纤维连接蛋白(Fibronectin)、细胞连接素(Tenascin-C)表达染色均较对照组增强。实时荧光定量 PCR 结果示,随时间延长实验组 Ⅰ 型胶原、Ⅲ 型胶原、Fibronectin、α-肌动蛋白(α-smooth muscle actin,α-SMA)、VEGF mRNA 相对表达量均逐渐上调(P<0.05)。除培养 5 d 两组 Ⅰ 型胶原、Ⅲ 型胶原、VEGF mRNA 相对表达量比较差异无统计学意义(P>0.05)外,其余各时间点实验组 Ⅰ 型胶原、Ⅲ 型胶原、Fibronectin、α-SMA 和 VEGF mRNA 相对表达量均显著高于对照组(P<0.05)。
结论
hAMSCs 具有 MSCs 特征,且体外增殖能力良好,可作为组织工程种子细胞来源;体外诱导后韧带成纤维细胞及血管生成相关特异性基因表达上调,韧带成纤维细胞特异性蛋白分泌增加,TGF-β1 联合 VEGF 可作为构建组织工程韧带的生长因子选择。
Keywords: 人羊膜间充质干细胞, 韧带成纤维细胞, TGF-β1, VEGF, 组织工程韧带
Abstract
Objective
To investigate whether human amniotic mesenchymal stem cells (hAMSCs) have the characteristics of mesenchymal stem cells (MSCs) and the differentiation capacity into ligament fibroblasts in vitro.
Methods
The hAMSCs were separated through trypsin and collagenase digestion from placenta, the phenotypic characteristics of hAMSCs were detected by flow cytometry, the cytokeratin-19 (CK-19) and vimentin expression of hAMSCs were tested through immunofluorescence staining. The hAMSCs at the 3rd passage were cultured with L-DMEM/F12 medium containing transforming growth factor β1 (TGF-β1) and vascular endothelial growth factor (VEGF) as the experimental group and with single L-DMEM/F12 medium as the control group. The morphology of hAMSCs was observed by inverted phase contrast microscope; the cellular activities and ability of proliferation were examined by cell counting kit-8 (CCK-8) method; the ligament fibroblasts related protein expressions including collagen type I, collagen type III, Fibronectin, and Tenascin-C were detected by immunofluorescence staining; specific mRNA expressions of ligament fibroblasts and angiogenesis including collagen type I, collagen type III, Fibronectin, α-smooth muscle actin (α-SMA), and VEGF were measured by real-time fluorescence quantitative PCR.
Results
The hAMSCs presented monolayer and adherent growth under inverted phase contrast microscope; the flow cytometry results demonstrated that hAMSCs expressed the MSCs phenotypes; the immunofluorescence staining results indicated the hAMSCs had high expression of the vimentin and low expression of CK-19; the hAMSCs possessed the differentiation ability into the osteoblasts, chondroblasts, and lipoblasts. The CCK-8 results displayed that cells reached the peak of growth curve at 7 days in each group, and the proliferation ability in the experimental group was significantly higher than that in the control group at 7 days (P<0.05). The immunofluorescence staining results showed that the expressions of collagen type I, collagen type III, Fibronectin, and Tenascin-C in the experimental group were significantly higher than those in the control group at 5, 10, and15 days after culture (P<0.05). The real-time fluorescence quantitative PCR results revealed that the mRNA relative expressions had an increasing tendency at varying degrees with time in the experimental group (P<0.05). The relative mRNA expressions of collagen type I, collagen type III, Fibronectin, α-SMA, and VEGF in the experimental group were significantly higher than those in the control group at the other time points (P<0.05), but no significant difference was found in the relative mRNA expressions of collagen type I, collagen type III, and VEGF between 2 groups at 5 days (P>0.05).
Conclusion
The hAMSCs possesses the characteristics of MSCs and good proliferation ability which could be chosen as seed cell source in tissue engineering. The expressions of ligament fibroblasts and angiogenesis related genes could be up-regulated, after induction in vitro, and the synthesis of ligament fibroblasts related proteins could be strengthened. In addition, the application of TGF-β1 and VEGF could be used as growth factors sources in constructing tissue engineered ligament.
Keywords: Human amniotic mesenchymal stem cells, ligament fibroblasts, transforming growth factor β1, vascular endothelial growth factor, tissue engineered ligament
人前交叉韧带(human anterior cruciate liga-ment,hACL)为致密结缔组织,主要功能为限制胫骨前移、膝关节过伸、内外旋等活动;由于其股骨附着点面积较大,故常发生撕裂或断裂损伤;韧带组织血供较差,损伤后自行修复能力较弱[1]。随着韧带组织工程的发展,许多新兴技术被用于动物体内实验[2]。种子细胞、细胞生长因子以及支架是构建组织工程韧带的三大要素[3-5]。其中,用于构建组织工程韧带的种子细胞主要为韧带或肌腱来源的韧带成纤维细胞,但成体细胞体外扩增速度慢,细胞分泌胶原能力不能满足组织工程韧带的需求[6-7]。另一重要的种子细胞是 MSCs,其具有增殖能力强、免疫原性低、能定向迁徙以及成骨、成软骨及成脂等多向分化能力[8-10]。目前研究已成功从各种成熟组织中分离出多种 MSCs,如 BMSCs、脂肪来源 MSCs、外周血来源 MSCs等[11-14]。人羊膜间充质干细胞(human amniotic mesenchymal stem cells,hAMSCs)取自胎盘表层羊膜,取材时对人体无额外损伤,无伦理问题,而且细胞增殖速度快[15]。研究表明,hAMSCs 不仅增殖能力较 BMSCs 强,而且具有高纯度、高丰度、高活力等特点[16-17],同时也具有低免疫原性和多向分化潜能等干细胞特性。TGF-β1 属于调节细胞生长及分化的 TGF-β 超家族,可使正常成纤维细胞表型发生转化[18],其不仅能促进成纤维细胞增殖生长,还可促进细胞外基质生成,如胶原蛋白、纤维连接蛋白(Fibronectin)的表达,抑制细胞外基质降解[19]。VEGF 是内皮细胞特异性促有丝分裂原[20],在与血管内皮细胞膜上的受体结合后,能够特异性地作用于血管内皮细胞,具有促进新生血管生成、增加血管通透性等作用,是公认的功能最强大、特异性最高的血管生成调节因子,在血管再生和软组织再生过程中起重要调节作用[21]。本研究拟采用 TGF-β1 联合 VEGF 体外诱导 hAMSCs 向韧带成纤维细胞分化,探讨 hAMSCs 作为构建组织工程韧带种子细胞的可行性。报告如下。
1. 材料与方法
1.1. 主要试剂及仪器
L-DMEM/F12 培养液(GIBCO 公司,美国);胰蛋白酶、Ⅱ 型胶原酶(HyClone 公司,美国);TGF-β1、VEGF(Protein-Tec 公司,美国);细胞角蛋白-19(cytokeratin-19,CK-19)、波形蛋白抗体(北京索莱宝科技有限公司);成骨、成软骨、成脂分化培养基以及 Ⅰ 型胶原、Ⅲ 型胶原、Fibrone-ctin、细胞连接素(Tenascin-C)抗体(Abcam 公司,美国);RNAiso Plus、转录试剂盒、荧光定量 PCR 试剂盒(Takara 公司,日本)。
FACS Calibur 流式细胞仪(Beckman 公司,美国);核酸蛋白测量 NanoDrop-1000 仪、CO2 恒温箱(Thermoscientific 公司,美国);冷冻离心机(Eppendorf 公司,德国);倒置相差显微镜、倒置荧光显微镜(Olympus 公司,日本)。
1.2. hAMSCs 分离、培养及鉴定
1.2.1 hAMSCs 分离及培养 实验所用 3 个胎盘由遵义医学院附属医院产科足月产产妇自愿捐赠,无其他基础疾病,符合遵义医学院附属医院伦理委员会要求。无菌环境下剥离胎盘胎儿面羊膜后,置于含有 D-Hank 液的广口瓶中,4℃ 条件下转至实验室,D-Hank 液反复冲洗羊膜并用钝性物轻刮羊膜表面;D-Hank 液再次冲洗羊膜,无菌剪将羊膜剪成大小为 1 mm×1 mm 的碎片,加入 2~3 倍羊膜体积的 0.05% EDTA-胰蛋白酶分别消化 40 min 和 30 min;D-Hank 液清洗后,加入与羊膜组织等体积的 0.8 g/L Ⅱ 型胶原酶,置入 37℃ 水浴箱中震荡消化 1~2 h,至肉眼观察不到羊膜碎片;300 目滤网过滤,收集细胞滤液,以离心半径 25 cm、1 500 r/min 离心 6 min。弃去上清液,加入含 10% FBS、1×105 U/L 青霉素、1×105 U/L 链霉素、1% 非必需氨基酸的 L-DMEM/F12 培养基重悬细胞,以(1~3)×108 个/mL 密度接种于 75 cm2 培养瓶中,37℃、5% CO2 及饱和湿度培养箱培养。每 2~3 天更换 1 次培养基。
待细胞融合约 80% 时加入 0.125% 胰蛋白酶,于 37℃、5% CO2 及饱和湿度细胞培养箱中消化 4 min,使贴壁细胞脱落、变圆;然后加入等体积含 10% FBS 的 L-DMEM/F12 培养基终止消化,同上法离心 6 min;弃上清液,加入 1 mL L-DMEM/F12 培养基吹打重悬细胞;最后以 1.5×108 个/mL 浓度接种于 75 cm2 培养瓶中,传至第 3 代进行以下实验。倒置相差显微镜下观察细胞生长、融合程度,判断细胞活性和增殖情况。
1.2.2 hAMSCs 鉴定 ① 流式细胞术鉴定 hAMSCs 表面标志物表达:取融合达 80% 以上的第 3 代 hAMSCs,弃培养液,PBS 清洗 2 次,加入 0.125% 胰蛋白酶-0.02% EDTA 消化液,于 37℃、5% CO2 及饱和湿度细胞培养箱内消化 5 min 后,加入等体积含 10% FBS 的 L-DMEM 培养基终止消化;将消化的细胞吹打混匀,同上法离心 6 min;弃上清,PBS 吹打重悬细胞,同上法离心 6 min,重复 2 次后调整细胞浓度至 2×109 个/mL,装入流式管中;每管加入 100 μL 细胞悬液,室温下避光分别加入鼠抗人单克隆抗体 CD44-PE、CD90-FITC、CD105-PerCP-Cy、CD73-APC 和阴性对照混合液 CD34-PE、CD19-PE、CD45-PE、CD11b-PE、HLA-DR,室温避光孵育 30 min;每管均加入 2 mL PBS,充分震荡,同上法离心 6 min;弃上清,加入 250 μL PBS 重悬细胞后行流式细胞仪鉴定,计算细胞表面抗原阳性率。
② 免疫荧光检测 hAMSCs 细胞质中 CK-19 及波形蛋白表达:取密度为 1.5×105 个/mL 的第 3 代细胞悬液,接种于 6 孔板中培养,并在孔板中置入直径 25 mm 含有多聚赖氨酸的细胞爬片;培养 7 d 后取出细胞爬片,37℃ 预热的 PBS 清洗 3 次,每次 10 min;37℃、4% 多聚甲醛固定 30 min,PBS 清洗 3 次,每次 10 min;0.1% TritionX-100 37℃ 下透膜 15 min,PBS 清洗 3 次,每次 10 min;5% 牛血清白蛋白 37℃ 下封闭 30 min,甩干不清洗;加入一抗鼠抗人 CK-19 和波形蛋白单克隆抗体 100 μL,4℃ 冰箱内孵育过夜;PBS 清洗 3 次,每次 10 min;滴加 FITC 标记的羊抗鼠二抗 100 μL,37℃ 下避光孵育 1 h;D-PBS 清洗 3 次,每次 10 min;DAPI 染液复染细胞核 10 min,PBS 清洗 3 次,每次 10 min;抗荧光淬灭剂封片处理后,倒置荧光显微镜观察两种蛋白在细胞中的表达情况。
③ hAMSCs 成骨、成软骨及成脂诱导分化鉴定:取第 3 代 hAMSCs,待细胞融合率至 50% 左右时,弃去培养液及未贴壁细胞,PBS 清洗 3 次;分别更换培养液为成骨、成软骨和成脂诱导培养基,每隔 3 d 换液 1 次;培养至 14 d 时,成骨分化行茜素红染色,成软骨分化行甲苯胺蓝染色,成脂分化行油红 O 染色,倒置相差显微镜观察染色结果。
1.3. 观测指标
1.3.1 细胞增殖-毒性检测试剂盒(cell counting kit-8,CCK-8)检测细胞增殖能力 取细胞融合率为 85% 的第 3 代 hAMSCs 分为两组,实验组使用成韧带或纤维细胞诱导培养基(含 10% FBS、1×105 U/L 青霉素、100 mg/L 链霉素、10 μg/L TGF-β1 和 15 μg/L VEGF 的 L-DMEM/F12 培养基)换液,对照组使用普通培养基(含 10% FBS、1×105 U/L 青霉素、100 mg/L 链霉素的 L-DMEM/F12 培养基)换液。将细胞培养板置于 37℃、5% CO2 及饱和湿度细胞培养箱中培养 24 h 后,向孔中小心加入 10 μL CCK-8 溶液,于细胞培养箱中孵育 3 h,酶标仪测定波长为 450 nm 处的吸光度(A)值,每隔 24 h 测 1 次,连续测量 13 d,记录数据并绘制细胞增殖曲线。
1.3.2 免疫荧光染色检测细胞内蛋白表达情况 取第 3 代 hAMSCs 调整成密度为 1.5×105 个/mL 的细胞悬液,接种于 6 孔板中培养,并在孔板中置入直径 25 mm 含有多聚赖氨酸的细胞爬片。同 1.3.1 方法分组,分别培养 5、10、15 d 后取出细胞爬片,参照上述细胞免疫荧光染色步骤操作,倒置荧光显微镜观察 Ⅰ 型胶原、Ⅲ 型胶原、Fibronectin、Tenascin-C 在细胞中的表达情况。
1.3.3 实时荧光定量 PCR 检测韧带及血管生成相关基因表达 取融合率达 85%的第 3 代 hAMSCs 同 1.3.1 方法分组,分别培养 5、10、15 d 后取出细胞,PBS 洗 3 次,使用 RNAiso plus 试剂提取细胞总 RNA,反转录获得 cDNA,进行 RT-PCR 反应。按照 PCR 试剂盒说明配置 PCR 反应体系;扩增条件:预变性 95℃、30 s,变性 95℃、5 s,退火 60℃、30 s。引物由上海生工生物工程股份有限公司合成,引物序列及产物长度见表 1。以 GAPDH 作为内参,通过 2–ΔΔCt法计算 Ⅰ 型胶原、Ⅲ 型胶原、Fibronectin、α-肌动蛋白(α-smooth muscle actin,α-SMA)和 VEGF mRNA 的相对表达量。
表 1.
The primer sequence and product length for real-time fluorescence quantitative PCR
实时荧光定量 PCR 基因引物序列及产物长度
| 基因
Gene |
引物序列(5′→3′)
Primer sequence (5′→3′) |
产物长度(bp)
Product length (bp) |
| Ⅰ 型胶原
Collagen typeⅠ |
上游 GGTCCCAAAGGTGCTGATGG
Upstream |
182 |
| 下游 GACCAGCCTCACCACGGTCT
Downstream |
||
| Ⅲ 型胶原
Collagen type Ⅲ |
上游 CGAGGTGACAGAGGTGAAAGA
Upstream |
236 |
| 下游 AACCCAGTATTCTCCGCTCTT
Downstream |
||
| Fibronectin | 上游 GAGAATAAGCTGTACCATCGCAA
Upstream |
154 |
| 下游 GGGCGAGTAGGAGCAGTTG
Downstream |
||
| α-SMA | 上游 CTGACAGAGGCACCACTGAA
Upstream |
160 |
| 下游 CATCTCCAGAGTCCAGCACA
Downstream |
||
| VEGF | 上游 TTCAGCGGACTCACCAGC
Upstream |
156 |
| 下游 AACCAACCTCCTCAAACCGT
Downstream |
||
| GAPDH | 上游 CAAGGTCATCCATGACAACTTTG
Upstream |
106 |
| 下游 CAAGGTCATCCATGACAACTTTG
Downstream |
1.4. 统计学方法
采用 SPSS14.0 统计软件进行分析。数据以均数±标准差表示,组内各时间点间比较采用单因素方差分析,两两比较采用 SNK 检验;两组间比较采用独立样本 t 检验;检验水准 α=0.05。
2. 结果
2.1. hAMSCs 形态学观察及鉴定
倒置相差显微镜观察示,原代 hAMSCs 呈多角形或不规则形态,贴壁生长;传代后细胞多呈长梭形,部分可见漩涡状、鱼群状生长细胞。见图 1。
图 1.

Morphology observation of hAMSCs (Inverted phase contrast microscope×40) a. Primary hAMSCs; b. The 1st generation of hAMSCs; c. The 2nd generation of hAMSCs; d. The 3rd generation of hAMSCs
hAMSCs 形态学观察(倒置相差显微镜×40) a. 原代细胞;b. 第 1 代细胞;c. 第 2 代细胞;d. 第 3 代细胞
细胞鉴定:① 流式细胞术检测示,第 3 代 hAMSCs 高表达 CD73、CD90、CD105、CD44,低表达 CD34、CD19、CD45、CD11b、HLA-DR。见图 2。② 免疫荧光染色观察示,第 3 代 hAMSCs 波形蛋白表达呈阳性,CK-19 表达呈阴性,见图 3。③ 第 3 代 hAMSCs 成骨诱导分化 14 d,茜素红染色示细胞形成多个小圆形致密矿化结节,透光性差;成软骨诱导分化 14 d,甲苯胺蓝染色示细胞变短、消散,核浆比降低;成脂诱导分化 14 d,油红 O 染色示细胞排列状态紊乱,细胞由长梭形变圆并开始出现脂滴样物质。见图 4。
图 2.

Phenotype molecule expression of the 3rd generation of hAMSCs by flow cytometry a. CD90; b. CD105; c. CD73; d. CD44; e. CD45+CD19+CD34+CD11b+HLA-DR
流式细胞仪检测第 3 代 hAMSCs 表面标志物表达 a. CD90;b. CD105;c. CD73;d. CD44;e. CD45+CD19+CD34+CD11b+ HLA-DR
图 3.

Expression of vimentin and CK-19 in the 3rd generation of hAMSCs by immunofluorescence staining (Inverted fluorescence microscope×100) a. In sequence of vimentin, DAPI staining, and merge of vimentin and DAPI; b. In sequence of CK-19, DPAI, and merge of CK-19 and DAPI
免疫荧光染色检测第 3 代 hAMSCs 波形蛋白及 CK-19 表达(倒置荧光显微镜×100) a. 依次为波形蛋白、DAPI 染色及波形蛋白与 DAPI 融合;b. 依次为 CK-19、DAPI 染色及 CK-19 与 DAPI 融合
图 4.

Morphology observation of the 3rd generation of hAMSCs after osteogenic, chondrogenic, and adipogenic induction (Inverted phase contrast microscope×100) a. Alizarin red staining after osteogenic induction; b. Toluidine blue staining after chondrogenic induction; c. Oil red O staining after adipogenic induction
第 3 代 hAMSCs 成骨、成软骨及成脂诱导分化 14 d 染色观察(倒置荧光显微镜×100) a. 成骨诱导茜素红染色;b. 成软骨诱导甲苯胺蓝染色;c. 成脂诱导油红 O 染色
2.2. CCK-8 法检测细胞增殖能力
两组细胞培养 7 d 时达对数增长期,9 d 时达增殖高峰,并基本维持增殖速率。实验组培养 7 d 后各时间点 A 值均显著高于对照组,比较差异有统计学意义(P<0.05);且随时间延长,增殖能力差异更明显。见图 5。
图 5.

Proliferation ability of cells in each group by CCK-8 method
CCK-8 法检测各组细胞增殖能力
2.3. 免疫荧光染色检测细胞内蛋白表达情况
2.3.1 Ⅰ 型胶原 对照组 Ⅰ 型胶原呈细胞质内分泌,蛋白表达相对较少;实验组 Ⅰ 型胶原呈大片、网块状分布,荧光信号明显。培养 5、10 d 时,两组 Ⅰ 型胶原表达均无明显改变,15 d 与 10 d 相比,对照组表达有一定程度增强,实验组增强较明显。实验组培养 5、10、15 d 时 Ⅰ 型胶原表达染色均较对照组增强。见图 6。
图 6.

Expression of collagen type I protein in each group at each time point by immunofluorescence staining (Inverted fluorescence microscope×100) From left to right for cells cultured for 5, 10, and 15 days a. Control group; b. Experimental group
免疫荧光染色观察各时间点各组细胞 Ⅰ 型胶原蛋白表达情况(倒置荧光显微镜×100) 从左至右依次为培养 5、10、15 d a. 对照组;b. 实验组
2.3.2 Ⅲ 型胶原 对照组 Ⅲ 型胶原呈细胞质内分泌,蛋白表达水平较低;实验组 Ⅲ 型胶原分泌呈现融合状、片状及网状形态。对照组培养 10 d 时 Ⅲ 型胶原表达较 5 d 时增强,但 15 d 时无明显改变;实验组随培养时间延长,Ⅲ 型胶原表达呈逐渐递增趋势。实验组培养 5、10、15 d 时 Ⅲ 型胶原表达染色均较对照组增强。见图 7。
图 7.

Expression of collagen type III protein in each group at each time point by immunofluorescence staining (Inverted fluorescence microscope×100) From left to right for cells cultured for 5, 10, and 15 days a. Control group; b. Experimental group
免疫荧光染色观察各时间点各组细胞 Ⅲ 型胶原蛋白表达情况(倒置荧光显微镜×100) 从左至右依次为培养 5、10、15 d a. 对照组;b. 实验组
2.3.3 Fibronectin 对照组 Fibronectin 呈细胞质内分泌,少量蛋白分泌至细胞外,细胞之间蛋白连接较少;实验组则观察到融合呈片、块状及网状的蛋白。对照组培养 10 d 时可观察到 Fibronectin 表达较 5 d 时增强,但 15 d 时无明显改变;实验组随培养时间延长,Fibronectin 蛋白表达呈逐渐递增趋势。实验组培养 5、15 d 时 Fibronectin 蛋白表达染色明显较对照组增强,但培养 10 d 时两组无明显差异。见图 8。
图 8.

Expression of Fibronectin protein in each group at each time point by immunofluorescence staining (Inverted fluorescence microscope×100) From left to right for cells cultured for 5, 10, and 15 days a. Control group; b. Experimental group
免疫荧光染色观察各时间点各组细胞 Fibronectin 蛋白表达情况(倒置荧光显微镜×100) 从左至右依次为培养 5、10、15 da. 对照组;b. 实验组
2.3.4 Tenascin-C 两组细胞 Tenascin-C 均呈胞外分泌,对照组 Tenascin-C 分泌呈较小片状分布、丝状排列,且顺序紊乱,细胞间蛋白连接程度较低,荧光强度较弱;实验组 Tenascin-C 分泌呈云雾状、大片状,胞内、外均有分布。对照组培养 5、10 d Tenascin-C 表达无明显变化;15 d 较 5、10 d 表达有明显增强。实验组培养 10 d 时 Tenascin-C 表达较 5 d 时明显增强,但 15 d 时无明显变化。实验组培养 5、10、15 d 时 Tenascin-C 表达染色均较对照组增强。见图 9。
图 9.

Expression of Tenascin-C protein in each group at each time point by immunofluorescence staining (Inverted fluorescence microscope×100) From left to right for cells cultured for 5, 10, and 15 days a. Control group; b. Experimental group
免疫荧光染色观察各时间点各组细胞 Tenascin-C 蛋白表达情况(倒置荧光显微镜×100) 从左至右依次为培养 5、10、15 d a. 对照组;b. 实验组
2.4. 实时荧光定量 PCR 检测韧带及血管生成相关基因表达
2.4.1 Ⅰ 型胶原 组内比较:对照组各时间点 Ⅰ 型胶原 mRNA 相对表达量比较,差异均无统计学意义(P>0.05);实验组 Ⅰ 型胶原 mRNA 相对表达量随时间延长逐渐上调,比较差异均有统计学意义(P<0.05)。组间比较:培养 5 d 时实验组 Ⅰ 型胶原 mRNA 相对表达量与对照组比较,差异无统计学意义(t=1.197,P=0.297);10、15 d 时显著高于对照组,比较差异有统计学意义(P<0.05)。见表 2。
表 2.
Comparison of relative collagen type I mRNA expressions between groups at each time point (n=3,
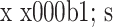 )
)
培养各时间点各组 Ⅰ 型胶原 mRNA 相对表达量比较(n=3,
 )
)
| 组别
Group |
5 d | 10 d | 15 d | 统计值
Statistic |
|
*与 5 d 时比较 P<0.05,#与 10 d 时比较 P<0.05
*Compared with the value at 5 days, P<0.05;#compared with the value at 10 days, P<0.05 | ||||
| 实验组
Experimental group |
0.877±0.346# | 1.955±0.043* | 4.643±0.466*# |
F=99.870
P= 0.000 |
| 对照组
Control group |
1.147±0.182 | 1.010±0.051 | 1.150±0.076 |
F= 1.394
P= 0.318 |
| 统计值
Statistic |
t=1.197
P=0.297 |
t=24.460
P= 0.000 |
t=12.800
P= 0.001 |
|
2.4.2 Ⅲ 型胶原 组内比较:对照组各时间点 Ⅲ 型胶原 mRNA 相对表达量比较,差异均无统计学意义(P>0.05),15 d 时相对表达量略低于 10 d 时;实验组 Ⅲ 型胶原 mRNA 相对表达量随时间延长逐渐上调,比较差异均有统计学意义(P<0.05)。组间比较:培养 5 d 时两组 Ⅲ 型胶原 mRNA 相对表达量比较,差异无统计学意义(t=0.347,P=0.746);10、15 d 时实验组明显高于对照组,比较差异有统计学意义(P<0.05)。见表 3。
表 3.
Comparison of relative collagen type III mRNA expressions between groups at each time point (n=3,
 )
)
培养各时间点各组 Ⅲ 型胶原 mRNA 相对表达量比较(n=3,
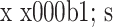 )
)
| 组别
Group |
5 d | 10 d | 15 d | 统计值
Statistic |
|
*与 5 d 时比较 P<0.05,#与 10 d 时比较 P<0.05
*Compared with the value at 5 days, P<0.05;#compared with the value at 10 days, P<0.05 | ||||
| 实验组
Experimental group |
1.037±0.209# | 2.822±0.049* | 4.010±0.920*# |
F=22.600
P= 0.002 |
| 对照组
Control group |
1.081±0.069 | 1.259±0.225 | 1.140±0.425 |
F= 0.315
P= 0.741 |
| 统计值
Statistic |
t=0.347
P=0.746 |
t=11.770
P= 0.000 |
t=4.906
P=0.008 |
|
2.4.3 Fibronectin 组内比较:对照组培养 10 d 时 Fibronectin mRNA 相对表达量较 5 d 时略有增加,15 d 时无明显变化,各时间点间比较差异均无统计学意义(P>0.05);实验组 Fibronectin mRNA 相对表达量随时间延长逐渐上调,各时间点间比较差异均有统计学意义(P<0.05)。组间比较:培养各时间点实验组 Fibronectin mRNA 相对表达量均明显高于对照组,比较差异有统计学意义(P<0.05)。见表 4。
表 4.
Comparison of relative Fibronectin mRNA expressions between groups at each time point (n=3,
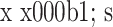 )
)
培养各时间点各组 Fibronectin mRNA 相对表达量比较(n=3,
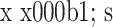 )
)
| 组别
Group |
5 d | 10 d | 15 d | 统计值
Statistic |
|
*与 5 d 时比较 P<0.05,#与 10 d 时比较 P<0.05
*Compared with the value at 5 days, P<0.05;#compared with the value at 10 days, P<0.05 | ||||
| 实验组
Experimental group |
2.068±0.110# | 4.048±0.364* | 6.123±0.779*# |
F=49.280
P= 0.000 |
| 对照组
Control group |
1.709±0.038 | 1.922±0.218 | 1.938±0.089 |
F= 2.576
P= 0.156 |
| 统计值
Statistic |
t=5.325
P=0.006 |
t=8.688
P=0.001 |
t=9.246
P=0.000 |
|
2.4.4 α-SMA 组内比较:两组 α-SMA mRNA 相对表达量随培养时间延长均逐渐上调,但对照组各时间点间比较差异无统计学意义(P>0.05),实验组各时间点间比较差异有统计学意义(P<0.05)。组间比较:培养各时间点,实验组 α-SMA mRNA 相对表达量均明显高于对照组,比较差异有统计学意义(P<0.05)。见表 5。
表 5.
Comparison of relative α-SMA mRNA expressions between groups at each time point (n=3,
 )
)
培养各时间点各组 α-SMA mRNA 相对表达量比较(n=3,
 )
)
| 组别
Group |
5 d | 10 d | 15 d | 统计值
Statistic |
|
*与 5 d 时比较 P<0.05,#与 10 d 时比较 P<0.05
*Compared with the value at 5 days, P<0.05;#compared with the value at 10 days, P<0.05 | ||||
| 实验组
Experimental group |
2.139±0.090# | 3.867±1.081* | 4.077±0.164*# |
F=8.453
P=0.018 |
| 对照组
Control group |
1.203±0.096 | 1.295±0.108 | 1.311±0.210 |
F=0.474
P=0.644 |
| 统计值
Statistic |
t=12.350
P= 0.000 |
t=4.022
P=0.015 |
t=24.570
P= 0.000 |
|
2.4.5 VEGF 组内比较:对照组各时间点 VEGF mRNA 相对表达量比较,差异均无统计学意义(P>0.05);实验组 VEGF mRNA 相对表达量随培养时间延长逐渐上调,各时间点间比较差异均有统计学意义(P<0.05)。组间比较:培养 5 d 时,对照组 VEGF mRNA 相对表达量高于实验组,但差异无统计学意义(t=0.421,P=0.696);但 10、15 d 时实验组 VEGF mRNA 相对表达量显著高于对照组,比较差异有统计学意义(P<0.05)。见表 6。
表 6.
Comparison of relative VEGF mRNA expressions between groups at each time point (n=3,
 )
)
培养各时间点各组 VEGF mRNA 相对表达量比较(n=3,
 )
)
| 组别
Group |
5 d | 10 d | 15 d | 统计值
Statistic |
|
*与 5 d 时比较 P<0.05,#与 10 d 时比较 P<0.05
*Compared with the value at 5 days, P<0.05;#compared with the value at 10 days, P<0.05 | ||||
| 实验组
Experimental group |
1.505±0.348# | 3.007±0.467* | 4.747±0.421*# |
F=45.871
P= 0.000 |
| 对照组
Control group |
1.593±0.105 | 1.592±0.230 | 1.634±0.093 |
F= 0.071
P= 0.933 |
| 统计值
Statistic |
t=0.421
P=0.696 |
t=4.996
P=0.001 |
t=11.390
P= 0.000 |
|
3. 讨论
膝关节韧带组织工程是以生物材料与再生医学为基础,目的在于种子细胞经生长因子作用后具有相应的生物活性及组织相容性,促进 ACL 的修复与再生,为膝关节 ACL 损伤提供了新型治疗模式。构建组织工程韧带的首要任务是筛选种子细胞[22],目前种子细胞来源广泛,其中成体干细胞中的 MSCs,尤其以 BMSCs 为代表,具有体外增殖能力强、经诱导后可向多种细胞分化的功能[23-24],是目前极具有临床应用前景的组织工程种子细胞[25]。hAMSCs 来源于中胚层,取自废弃的羊膜,通过酶消化法获得,hAMSCs 与 BMSCs 均具有类似的生物学特性[26-27]。获取 hAMSCs 的方法对人体无额外创伤,不存在伦理道德方面争议;既往研究显示,hAMSCs 的增殖能力优于 BMSCs[28]。目前,hAMSCs 已普遍应用于人体结缔组织相关损伤,如软骨、骨、肌腱、韧带、神经系统疾病及多种软组织损伤疾病的体内外研究[29-32]。本研究按照国际细胞治疗协会的 MSCs 鉴定标准[33],对 hAMSCs 流式表型分子进行分析,结果显示第 3 代 hAMSCs 高表达 CD44、CD90、CD105、CD73,且不表达 CD34、CD19、CD45、CD11b 及 HLA-DR,说明 hAMSCs 具有 MSCs 的表型特征。此外,hAMSCs 低表达免疫因子 CD14 及主要组织相容性 Ⅱ 类抗原 HLA-DR,说明 hAMSCs 免疫原性较低,不引起同种或异种淋巴细胞增殖反应,作为组织工程细胞来源或用于组织、器官损伤修复时引起免疫排斥反应风险低。由于在原代细胞获取过程中容易混杂人羊膜上皮细胞,CK-19 和波形蛋白分别作为人羊膜上皮细胞和 hAMSCs 的特异性表达分子,所以我们选择对 CK-19 和波形蛋白表达进行检测,以评价分离培养细胞的纯度。本研究结果显示,第 3 代 hAMSCs 高表达波形蛋白,低表达 CK-19,说明培养的 hAMSCs 符合实验对细胞的纯度要求。
本研究结果提示,经 TGF-β1、VEGF 联合诱导后,实验组细胞增殖以及韧带相关特异性蛋白 Ⅰ 型胶原、Ⅲ 型胶原、Fibronectin、Tenascin-C 的表达较对照组明显增强;说明 TGF-β1 与 VEGF 联合使用可诱导体外 hAMSCs 向韧带成纤维细胞分化,并且在诱导分化过程中,TGF-β1 和 VEGF 对细胞形态、蛋白分泌方面起促进作用。近年研究显示,TGF-β1 是一种属于 TGF 家族的生长性多肽分子,除了在细胞增殖、凋亡和分化方面起调节作用,还可增加细胞外基质合成[34-37]。Hara 等[38]发现经活化后的 TGF-β1 可通过 Smad 信号通路刺激并动员 MSCs 向肌腱细胞分化。Farhat 等[39]研究表明 TGF-β1 促进屈肌肌腱愈合,它是通过诱导纤溶酶原激活物抑制剂 1,抑制纤溶酶介导的基质金属蛋白酶 2 活性,来降低瘢痕组织粘连,促进瘢痕组织的修复和屈肌肌腱的再生。前交叉韧带重建术后患者通常需要长时间恢复,由于支配韧带的血管丛较少,因此 VEGF 在移植后血管重建中起重要作用。Chen 等[40]研究观察了透明质酸钠载 VEGF165 药物对兔膝关节前交叉韧带移植重建术后血管重建的作用,结果显示使用 VEGF 组重建术后韧带微血管密度明显高于对照组,并且韧带最大拉伸负荷也显著升高。说明 VEGF 可提高前交叉韧带重建术后早期血运重建及生物力学特性的恢复。本研究实时荧光定量 PCR 结果显示,实验组培养 10、15 d 时 Ⅰ 型胶原、Ⅲ 型胶原、Fibronectin、α-SMA 和 VEGF mRNA 相对表达量均高于对照组(P<0.05)。因此我们认为 hAMSCs 经 TGF-β1 和 VEGF 联合诱导后可向韧带成纤维细胞分化,且在 VEGF 作用下,韧带细胞的血管修复能力得到一定程度提高。
综上述,hAMSCs 表达 MSCs 特异性分子,具有低免疫原性和较高增殖能力,可作为组织工程韧带种子细胞来源。经 TGF-β1 联合 VEGF 体外诱导后,韧带成纤维细胞相关特异性基因表达上调、蛋白合成增高,同时血管生成相关基因表达也显著增加。但 hAMSCs 搭载于支架材料后的生物相容性以及植入体内后的生物力学、成韧带能力等,有待进一步实验研究。
Funding Statement
贵州省科学技术基金资助项目(黔科合SY字[2010]3091、黔科合LH字[2016]7477)
Science and Technology Foundation of Guizhou Province (SY[2010]3091, LH[2016]7477)
References
- 1.张承昊, 金瑛, 范青洪, 等 rBMSC 和 rLFs 做为韧带组织工程种子细胞的特性比较. 遵义医学院学报. 2015;38(1):54–59. [Google Scholar]
- 2.Smith SE, White RA, Grant DA, et al Gold and Hydroxyapatite Nano-Composite Scaffolds for Anterior Cruciate Ligament Reconstruction: In Vitro Characterization . J Nanosci Nanotechnol. 2016;16(1):1160–1169. doi: 10.1166/jnn.2016.10861. [DOI] [PubMed] [Google Scholar]
- 3.Zhi Y, Liu W, Zhang P, et al Erratum to: Electrospun silk fibroin mat enhances tendon-bone healing in a rabbit extra-articular model. Biotechnol Letters. 2016;38(10):1837–1838. doi: 10.1007/s10529-016-2171-7. [DOI] [PubMed] [Google Scholar]
- 4.Cooper JA Jr, Bailey LO, Carter JN, et al Evaluation of the anterior cruciate ligament, medial collateral ligament, achilles tendon and patellar tendon as cell sources for tissue-engineered ligament. Biomaterials. 2006;27(13):2747–2754. doi: 10.1016/j.biomaterials.2005.12.013. [DOI] [PubMed] [Google Scholar]
- 5.Leroy A, Nottelet B, Bony C, et al PLA-poloxamer/poloxamine copolymers for ligament tissue engineering: sound macromolecular design for degradable scaffolds and MSC differentiation. Biomater Sci. 2015;3(4):617–626. doi: 10.1039/c4bm00433g. [DOI] [PubMed] [Google Scholar]
- 6.付维力, 陈刚, 唐新, 等 BMP-12 重组腺病毒载体转染外周血 MSCs 向肌腱/韧带细胞分化研究. 中国修复重建外科杂志. 2015;29(4):472–476. [PubMed] [Google Scholar]
- 7.Caplan AI New era of cell-based orthopedic therapies. Tissue Eng Part B Rev. 2009;15(2):195–200. doi: 10.1089/ten.teb.2008.0515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Maidhof R, Rafiuddin A, Chowdhury F, et al Timing of mesenchymal stem cell delivery impacts the fate and therapeutic potential in intervertebral disc repair. J Orthop Res. 2017;35(1):32–40. doi: 10.1002/jor.23350. [DOI] [PubMed] [Google Scholar]
- 9.Nogami M, Kimura T, Seki S, et al A Human Amnion-Derived Extracellular Matrix-Coated Cell-Free Scaffold for Cartilage Repair: In Vitro and In Vivo Studies . Tissue Eng Part A. 2016;22(7-8):680–688. doi: 10.1089/ten.TEA.2015.0285. [DOI] [PubMed] [Google Scholar]
- 10.洪佳琼, 高雅, 宋洁, 等 人羊膜间充质干细胞与骨髓间充质干细胞的生物学特性及免疫抑制作用的比较. 中国实验血液学杂志. 2016;24(3):858–864. doi: 10.7534/j.issn.1009-2137.2016.03.041. [DOI] [PubMed] [Google Scholar]
- 11.Kim SH, Bang SH, Kang SY, et al Human amniotic membrane-derived stromal cells (hAMSC) interact depending on breast cancer cell type through secreted molecules. Tissue Cell. 2015;47(1):10–16. doi: 10.1016/j.tice.2014.10.003. [DOI] [PubMed] [Google Scholar]
- 12.Cortes Y, Ojeda M, Araya D, et al Isolation and multilineage differentiation of bone marrow mesenchymal stem cells from abattoir-derived bovine fetuses. BMC Vet Res. 2013;9:133. doi: 10.1186/1746-6148-9-133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vargas D, Rosales W, Lizcano F Modifications of Human Subcutaneous ADMSC after PPARgamma Activation and Cold Exposition. Stem Cells Int. 2015;2015:196348. doi: 10.1155/2015/196348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Fu Q, Tang NN, Zhang Q, et al Preclinical Study of Cell Therapy for Osteonecrosis of the Femoral Head with Allogenic Peripheral Blood-Derived Mesenchymal Stem Cells. Yonsei Med J. 2016;57(4):1006–1015. doi: 10.3349/ymj.2016.57.4.1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.金瑛, 李豫皖, 张承昊, 等 体外诱导人羊膜间充质干细胞向韧带细胞分化的实验研究. 中国修复重建外科杂志. 2016;30(2):237–244. [PubMed] [Google Scholar]
- 16.Filioli Uranio M, Dell’Aquila ME, Caira M, et al Characterization and in vitro differentiation potency of early-passage canine amnion- and umbilical cord-derived mesenchymal stem cells as related to gestational age . Mol Reprod Dev. 2014;81(6):539–551. doi: 10.1002/mrd.22322. [DOI] [PubMed] [Google Scholar]
- 17.丁志, 杨松林 间充质干细胞生物学特性及其分化潜能. 中国组织工程研究与临床康复. 2011;15(1):147–150. [Google Scholar]
- 18.Ghebes CA, van Lente J, Post JN, et al High-Throughput Screening Assay Identifies Small Molecules Capable of Modulating the BMP-2 and TGF-beta1 Signaling Pathway. SLAS Discov. 2017;22(1):40–50. doi: 10.1177/1087057116669346. [DOI] [PubMed] [Google Scholar]
- 19.Lee SB, Lim AR, Rah DK, et al Modulation of heat shock protein 90 affects TGF-beta-induced collagen synthesis in human dermal fibroblast cells. Tissue Cell. 2016;48(6):616–623. doi: 10.1016/j.tice.2016.09.002. [DOI] [PubMed] [Google Scholar]
- 20.Sasaki K, Kuroda R, Ishida K, et al Enhancement of tendon-bone osteointegration of anterior cruciate ligament graft using granulocyte colony-stimulating factor. Am J Sports Med. 2008;36(8):1519–1527. doi: 10.1177/0363546508316282. [DOI] [PubMed] [Google Scholar]
- 21.Bissell L, Tibrewal S, Sahni V, et al Growth factors and platelet rich plasma in anterior cruciate ligament reconstruction. Curr Stem Cell Res Ther. 2015;10(1):19–25. doi: 10.2174/1574888x09666140710102002. [DOI] [PubMed] [Google Scholar]
- 22.刘毅, 张承昊, 范青洪, 等 TGF-β1 和 bFGF 对单层共培养 BMSCs 和韧带成纤维细胞韧带特异性的影响 . 中国修复重建外科杂志. 2014;28(11):1406–1412. [PubMed] [Google Scholar]
- 23.王佃亮 种子细胞——组织工程连载之三. 中国生物工程杂志. 2014;34(7):108–113. [Google Scholar]
- 24.Montali M, Barachini S, Panvini FM, et al Growth Factor Content in Human Sera Affects the Isolation of Mesangiogenic Progenitor Cells (MPCs) from Human Bone Marrow. Front Cell Dev Biol. 2016;4:114. doi: 10.3389/fcell.2016.00114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kalathur M, Baiguera S, Macchiarini P Translating tissue-engineered tracheal replacement from bench to bedside. Cell Mol Life Sci. 2010;67(24):4185–4196. doi: 10.1007/s00018-010-0499-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Niwa H, Masui S, Chambers I, et al Phenotypic complementation establishes requirements for specific POU domain and generic transactivation function of Oct-3/4 in embryonic stem cells. Mol Cell Biol. 2002;22(5):1526–1536. doi: 10.1128/mcb.22.5.1526-1536.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chehelcheraghi F, Eimani H, Homayoonsadraie S, et al Effects of Acellular Amniotic Membrane Matrix and Bone Marrow-Derived Mesenchymal Stem Cells in Improving Random Skin Flap Survival in Rats. IranRed Crescent Med J. 2016;18(6):e25588. doi: 10.5812/ircmj.25588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.方宁, 张路, 宋秀军, 等 人羊膜间充质干细胞的分离、培养及鉴定. 遵义医学院学报. 2009;32(3):234–236. [Google Scholar]
- 29.Bonomi A, Silini A, Vertua E, et al Human amniotic mesenchymal stromal cells (hAMSCs) as potential vehicles for drug delivery in cancer therapy: an in vitro study . Stem Cell Res Ther. 2015;6:155. doi: 10.1186/s13287-015-0140-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kostjuk S, Loseva P, Chvartatskaya O, et al Extracellular GC-rich DNA activates TLR9- and NF-kB-dependent signaling pathways in human adipose-derived mesenchymal stem cells (haMSCs) Expert Opin Biol Ther. 2012;12(Suppl 1):S99–111. doi: 10.1517/14712598.2012.690028. [DOI] [PubMed] [Google Scholar]
- 31.Miranda-Sayago JM, Fernandez-Arcas N, Reyes-Engel A, et al Changes in CDKN2D, TP53, and miR125a expression: potential role in the evaluation of human amniotic fluid-derived mesenchymal stromal cell fitness. Genes Cells. 2012;17(8):673–687. doi: 10.1111/j.1365-2443.2012.01623.x. [DOI] [PubMed] [Google Scholar]
- 32.Rutka JT Expression of Concern: Dual regeneration of muscle and nerve by intravenous administration of human amniotic fluid-derived mesenchymal stem cells regulated by stromal cell-derived factor-1alpha in a sciatic nerve injury model. J Neurosurg. 2015;123(6):1605. doi: 10.3171/2015.7.JNS111360r. [DOI] [PubMed] [Google Scholar]
- 33.Dominici M, Le Blanc K, Mueller I, et al Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8(4):315–317. doi: 10.1080/14653240600855905. [DOI] [PubMed] [Google Scholar]
- 34.Crane JL, Xian L, Cao X Role of TGF-beta Signaling in Coupling Bone Remodeling. Methods Mol Biol. 2016;1344:287–300. doi: 10.1007/978-1-4939-2966-5_18. [DOI] [PubMed] [Google Scholar]
- 35.Sulaiman W, Nguyen DH Transforming growth factor beta 1, a cytokine with regenerative functions. Neural Regen Res. 2016;11(10):1549–1552. doi: 10.4103/1673-5374.193223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Utsunomiya T, Ishibazawa A, Nagaoka T, et al Transforming Growth Factor-beta Signaling Cascade Induced by Mechanical Stimulation of Fluid Shear Stress in Cultured Corneal Epithelial Cells. Invest Ophthalmol Vis Sci. 2016;57(14):6382–6388. doi: 10.1167/iovs.16-20638. [DOI] [PubMed] [Google Scholar]
- 37.Chou PH, Wang ST, Ma HL, et al Development of a two-step protocol for culture expansion of human annulus fibrosus cells with TGF-beta1 and FGF-2. Stem Cell Res Ther. 2016;7(1):89. doi: 10.1186/s13287-016-0332-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hara T, Yoshida E, Shinkai Y, et al Biglycan Intensifies ALK5-Smad2/3 Signaling by TGF-beta1 and Downregulates Syndecan-4 in Cultured Vascular Endothelial Cells. J Cell Biochem. 2017;118(5):1087–1096. doi: 10.1002/jcb.25721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Farhat YM, Al-Maliki AA, Easa A, et al TGF-beta1 Suppresses Plasmin and MMP Activity in Flexor Tendon Cells via PAI-1: Implications for Scarless Flexor Tendon Repair. J Cell Physiol. 2015;230(2):318–326. doi: 10.1002/jcp.24707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chen J, Yang L, Guo L, et al Sodium hyaluronate as a drug-release system for VEGF 165 improves graft revascularization in anterior cruciate ligament reconstruction in a rabbit model. Exp Ther Med. 2012;4(3):430–434. doi: 10.3892/etm.2012.629. [DOI] [PMC free article] [PubMed] [Google Scholar]