

ABSTRACT
Viruses elicit cell and organismic stress, and offset homeostasis. They trigger intrinsic, innate and adaptive immune responses, which limit infection. Viruses restore homeostasis by harnessing evolutionary conserved stress responses, such as the endoplasmic reticulum (ER) unfolded protein response (UPRER). The canonical UPRER restores homeostasis based on a cell-autonomous signalling network modulating transcriptional and translational output. The UPRER remedies cell damage, but upon severe and chronic stress leads to cell death. Signals from the UPRER flow along three branches with distinct stress sensors, the inositol requiring enzyme (Ire) 1, protein kinase R (PKR)-like ER kinase (PERK), and the activating transcription factor 6 (ATF6). This review shows how both enveloped and non-enveloped viruses use the UPRER to control cell stress and metabolic pathways, and thereby enhance infection and progeny formation, or undergo cell death. We highlight how the Ire1 axis bypasses apoptosis, boosts viral transcription and maintains dormant viral genomes during latency and persistence periods concurrent with long term survival of infected cells. These considerations open new options for oncolytic virus therapies against cancer cells where the UPRER is frequently upregulated. We conclude with a discussion of the evolutionary impact that viruses, in particular retroviruses, and anti-viral defense has on the UPRER.
Keywords: endoplasmic reticulum unfolded protein response, virus-induced cell stress, cell death, homeostasis, evolution, stress response
Although viruses trigger robust stress responses and innate and adaptive host defense, we reveal that a large spectrum of enveloped and non-enveloped viruses activate homeostasis restoring processes, especially the endoplasmic reticulum unfolded protein response, which promotes virus propagation and survival.
ABBREVIATIONS
- AdV
Adenovirus
- AARE
Amino acid response element (biding sites for ATF4 transcription factor)
- AAV
Adeno-associated virus
- AdV
Adenovirus
- ASFV
African swine fever virus
- ATF4
Activating transcription factor 4
- ATF6
Activating transcription factor 6
- Bak
Bcl-2 homologous antagonist or killer
- Bax
Bcl-2 associated X-protein
- Bcl2
B-cell lymphoma 2
- Bim
Proapoptotic protein Bcl-2 like protein 11 (Bcl2L11)
- BiP/Grp78
Binding immunoglobulin protein/Glucose regulated protein 78
- BVDV
Bovine viral diarrhea virus
- CVA16
Coxsackievirus A16
- CVB3
Coxsackievirus B3
- Cnx
Calnexin
- Crt
Calreticulin
- C/EBP
CCAAT/-enhancer-binding protein
- CD4
Cluster of differentiation 4 glycoprotein
- CHOP
CCAAT/-enhancer-binding protein homologous protein
- CMV
Cytomegalovirus
- COPII
Coatomer protein 2
- CREBH
Cyclic adenosine monophosphate (cAMP)-responsive element-binding protein H
- DENV
Dengue virus
- DDR
DNA damage response
- DnaK
Bacterial chaperone Hsp70
- EDEM
ERAD enhancing α-mannosidase-like proteins
- eIF2α
Eukaryotic translation initiation factor 2 subunit1
- ERAD
ER-associated degradation
- ERdj4
ER-localised J-protein 4
- ERSE
ER-stress response elements
- GADD34
Growth arrest and DNA damage-inducible protein 34
- HCMV
Human cytomegalovirus
- HCV
Hepatitis C virus
- HIV
Human immunodeficiency virus
- Hsp
Heat shock protein
- HSV
Herpes simplex virus
- IAV
Influenza A virus
- IFNAR1
Interferon alpha or beta receptor subunit 1
- IFNß
Interferon-ß
- IκB
Inhibitor of κB
- IKK
IκB kinase
- IL-6
Interleukin-6
- Ire1α
Inositol-requiring enzyme 1 alpha
- ISR
integrated stress response
- ISRIB
ISR inhibitor
- JeV
Japanese encephalitis virus
- JNK
c-Jun N-terminal kinases
- MHC-I
Major histocombatibility factor I
- MHV
Mouse gammaherpes virus
- NADPH
Nicotinamide adenine dinucleotide phosphate (reduced form)
- NF-κB
Nuclear factor kappa light-chain-enhancer of activated B cells
- NOX2
NADPH oxidase 2
- ORF
open-reading frame
- PERK
Protein kinase activated by double stranded RNA (PKR)-like ER kinase
- PMV
Paramyxo simian virus
- PP1
Protein phosphatase 1
- RSV
Respiratory syncytial virus
- RIDD
Regulated Ire1-dependent decay of mRNA
- ROS
Reactive oxygen species
- RPS
Ribosomal protein subunit
- SARS-CoV
Severe acute respiratory syndrome-related coronavirus
- STAT3
Signal transducer and activator of transcription 3
- TNF
Tumour necrosis factor
- TRAF2
TNF receptor associated factor 2
- UPRER
ER-unfolded protein response
- VSV
Vesicular stomatitis virus
- Xbp1s
X-box binding protein 1 spliced
- Xbp1u
X-box binding protein 1 unspliced
INTRODUCTION
An immense number of DNA and RNA viruses from bacteria and eukaryotes populate the globe, yet, most of them are harmless to humans, because they are not adapted to vertebrate cells. Several dozens of distinct viruses nevertheless enter humans, for example, through the eyes, the skin or the respiratory, gastro-intestinal and sexual tracts (Virgin, Wherry and Ahmed 2009). Such viruses cause infectious diseases, sometimes with global impact, and emerge unpredictably.
When viruses interact with cells, they perturb homeostasis, which results in their inactivation or in acute infection (Gulbahce et al. 2012; Rozenblatt-Rosen et al. 2012; Greber 2016; Greber and Flatt 2019). A number of evolutionary conserved mechanisms guard against infections. At the organismic level, anatomical barriers, such as polarized epithelial cells and mucosal secretion protect against airborne-viruses (Holt et al. 2008). Defense at the tissue level is coordinated by mucosal immunity and homing of immune cells, such as macrophages, dendritic cells, regulatory T cells, helper T cells, natural killer cells and mast cells equipped with specialized sensor proteins, including scavenger receptors and toll-like receptors (Takeda and Akira 2005; Fejer et al. 2008; Jost and Altfeld 2013; Byrne et al. 2015; Maler et al. 2017; Schmidt and Varga 2018; Stichling et al. 2018; Wang et al. 2018; Marshall, Portales-Cervantes and Leong 2019). At the cellular level, distinct biochemical processes antagonize infections and restore homeostasis. They include the DNA damage response, glycolysis, fatty acid synthesis, oxidative stress response, heat shock response, autophagy as well as the UPRER in the endoplasmic reticulum (ER) or processes in mitochondria (for reviews, see Kudchodkar and Levine 2009; Takeuchi and Akira 2009; Haynes and Ron 2010; Heaton and Randall 2011; Chan 2014; Roulin et al. 2014; Sanchez and Lagunoff 2015; Chatel-Chaix et al. 2016; Paul and Munz 2016; Khomich et al. 2018; Lotzerich et al. 2018; Weitzman and Fradet-Turcotte 2018; Hur 2019).
Acute infections arise when viral genomes replicate, and disseminate locally and systemically. Acute tissue damage is exacerbated by pathogen-associated molecular patterns (PAMPs) of viral proteins and nucleic acids triggering pattern recognition receptors (PRRs), and an immune response through the production of interferon (IFN) and proinflammatory cytokines (Haller, Kochs and Weber 2006; Rouse and Sehrawat 2010; Hoffmann, Schneider and Rice 2015).
Viruses have evolved to antagonize the inflammatory and IFN responses, and eventually restore homeostasis in a series of complex processes crucial for both the virus and the infected organism. It is notable that the failure to attenuate inflammation and IFN signalling can lead to the death of the organism, as exemplified with SARS-CoV-2, which blunts the production of IFN in the infected cells, but leaves the inflammatory response largely unaffected, a situation which results in a cytokine storm and fatal organ failure (Blanco-Melo et al. 2020). In most cases, the restoration of homeostasis involves a combined action of intrinsic, innate and adaptive immunity, and comprises microRNAs, pattern recognition receptors, antibodies and cell-based immunity (Takeuchi and Akira 2009; O'Connell et al. 2010; Pulendran, Li and Nakaya 2010). Examples of intrinsic factors are the tripartite interaction motif 5 splice variant α (TRIM5α) blocking HIV capsid uncoating, and adenosine deaminase ADAR1 balancing immune activation and self-tolerance (Colomer-Lluch et al. 2018; Lamers, van den Hoogen and Haagmans 2019). Restoration of homeostasis either clears the infection, or leads to virus persistence without obvious signs of disease and immune reactions (Virgin, Wherry and Ahmed 2009).
Here, we explore how viruses use the UPRER to restore homeostasis and virus output. For detailed reviews on the UPRER in herpesvirus and coronavirus infection, we refer the reader to recent overviews elsewhere (Fung and Liu 2019; Johnston and McCormick 2019).
THE UPRER STRESS SENSORS AND DOWNSTREAM SIGNALS
The ER has multiple functions, including the synthesis of proteins, oligosaccharides and lipids (Helenius and Aebi 2001; Ellgaard and Helenius 2003; Metcalf et al. 2020). Its lumen contains a high concentration of Ca2+ ions, and serves as both source and sink in Ca2+ signalling. The ER lumen is an oxidative environment and facilitates the formation of disulfide bonds in proteins, which is critical for the proper folding of newly synthesized proteins, together with protein- and lipid-glycosylation and molecular chaperones, such as the binding immunoglobulin protein (BiP, or glucose regulated protein 78, Grp78) (Xu et al. 2005). Newly synthesized secretory and membrane spanning proteins are properly folded in the ER, transported to intracellular organelles or secreted to the plasma membrane (Barlowe and Miller 2013). They undergo a range of modifications, including proteolytic processing, glycosylation and lipidation, interact with chaperones, isomerases, glycosyltransferases and glycosidases, and eventually exiting the ER after proper folding (Hammond and Helenius 1995; Wei et al. 2006; Braakman and Hebert 2013). In addition, the ER is a major hub for the synthesis of membrane lipids (Futerman and Riezman 2005; Maxfield and van Meer 2010; Harayama and Riezman 2018). The environment of the ER can be stressed by both physiological and pathological processes (Metcalf et al. 2020). Disturbances include the deregulation of cellular redox or the ER lipid environment, aberrant Ca2+ levels, glucose deprivation, or the accumulation of unfolded proteins in the ER.
ER-stress triggers an evolutionarily conserved response, the UPRER. UPRER is distinct from UPR in mitochondria, which is triggered by proteotoxic signals from reactive oxygen species and exacerbated by a decrease in mitochondrial membrane potential (Rolland et al. 2019). The UPRER was originally found to balance the synthesis, folding and degradation of proteins in the ER (reviewed in Ron and Walter 2007; Walter and Ron 2011). When the protein load in the ER exceeds the folding capacity, or when ER homeostasis is disturbed by ectopic cues, a set of phylogenetically conserved pathways transmits signals to relieve the condition of a stressed ER (Grootjans et al. 2016). The sensing of ER stress occurs by transmembrane stress transducers, the inositol-requiring enzyme 1 (Ire1), the protein kinase R (PKR)-like ER kinase (PERK), and the activating transcription factor 6 (ATF6). They all sense the levels of unfolded proteins by virtue of their lumenal domains in the ER, and transmit the information through their respective cytoplasmic domains to cytosolic effector pathways (Bernales, McDonald and Walter 2006). Remarkably, the Ire1 isoform alpha (Ire1α) and PERK also sense stress from saturated lipids in the ER membrane, and transduce a remedial response through their transmembrane domain (Volmer, van der Ploeg and Ron 2013; Kono, Amin-Wetzel and Ron 2017; Metcalf et al. 2020).
Ire1
Ire1 is the sensor of the most conserved branch of the UPRER. It is present in lower and higher eukaryotes. In mammals, two forms of Ire1, α and β are encoded by two separate genes ERN1 and ERN2, respectively. Ire1α is expressed ubiquitously while Ire1β is primarily expressed in gastrointestinal and respiratory tracts (Bertolotti et al. 2000; Tsuru et al. 2013). Both isoforms are type-I transmembrane proteins with an N-terminal lumenal domain and a dual-function cytoplasmic domain with Ser/Thr kinase and a ribonuclease (RNase) activities (Li et al. 2010). Ire1 has been extensively studied in yeast. However, yeast Ire1 (yIre1) is structurally different from the human Ire1 (hIre1) α (Gardner and Walter 2011). Upon accumulation of unfolded proteins in the ER, Ire1 trans-autophosphorylates and oligomerizes. This induces conformational changes in the RNase domain, which then cleaves a small intron of the transcription factor X-box binding protein (Xbp) 1 mRNA (Xbp1u), followed by ligation yielding a spliced mRNA encoding the active transcription factor Xbp1s (Aragon et al. 2009; Li et al. 2010; Jurkin et al. 2014). The role of Xbp1s in UPRER homeostasis is crucial. Along with other UPRER-induced transcription factors, Xbp1s upregulates and transactivates a repertoire of genes necessary for relieving the ER stress (Reimold et al. 2001; Acosta-Alvear et al. 2007). In addition, Xbp1s functions in cell growth, differentiation, survival and plasma cell differentiation, and immune cell development (Grootjans et al. 2016).
Although hIre1 is a sensor for protein stress in the ER, it has structurally unfavorable features for the direct binding of unfolded proteins. Hence, the question how Ire1α senses protein stress in the ER lumen, and transduces this information to the cytosol has been the subject of intense research over many years. Two main models exist for how the lumenal domain of Ire1α oligomerises and leads to the activation of the RNase function. The first model suggests that similar to yeast, unfolded proteins can bind to the core lumenal domain causing allosteric changes leading to Ire1α oligomerization (Karagoz et al. 2017). The unfolded proteins bind to an MHC-like groove of yIre1, whereas in hIre1, the helices flanking this groove are too closely placed to allow binding of unfolded proteins (Zhou et al. 2006). Dimerization and oligomerisation interfaces are separate in yIre1, whereas in hIre1, the oligomerisation interface is sterically hidden by other lumenal domains. In addition, the corresponding interface is postulated to be energetically unfavourable for oligomer formation. Yet, it is in a dynamic equilibrium between a closed- and an open-loop configuration. The open-loop structure of hIre1 can be bound and stabilised by unfolded proteins, and through allosteric changes this leads to exposure of an oligomerization interface, which promotes the formation of higher-order oligomers (Karagoz et al. 2017).
The second model suggests that Ire1 signalling is suppressed by the ER-resident BiP, which binds to Ire1 monomers, thereby preventing Ire1 dimerization and oligomerization (Bertolotti et al. 2000; Carrara et al. 2015). Unfolded proteins in turn also bind to BiP, and shift the equilibrium towards BiP-less Ire1 favouring Ire1 dimer and oligomer formation. Supporting this model, the co-chaperone ER-localised J-protein 4 (ERdj4) was recently shown to energetically promote the binding of BiP to Ire1α, and disrupt Ire1α dimers (Amin-Wetzel et al. 2017). It may be unlikely, however, that BiP dissociation from Ire1 provides the sole cue to Ire1 activation. A mutational study of the yIre1 lumenal domain removing the lumenal BiP binding site juxta membrane gave rise to Ire1, which remained inactive in the absence of ER stress, yet retained its stress-induced activation (Kimata et al. 2004). This opens the possibility that lumenal proteins, for example of viral origin, have evolved to directly activate Ire1α without affecting the other ER stress sensors PERK and ATF6.
Ire1 signalling has distinct downstream effects, most prominently Xbp1s, which activates a group of UPRER target genes (Acosta-Alvear et al. 2007). In parallel, specific degradation of a subset of ER-localized mRNAs has been identified and dubbed ‘regulated Ire1-dependent decay’ (RIDD) (Hollien and Weissman 2006). RIDD of ER-bound mRNAs may reduce the protein influx into the ER during ER stress. Exactly how Ire1 switches between Xbp1 splicing and RIDD is not clear, although weak activation of Ire1 gives rise to either Xbp1 splicing or RIDD. This opens a possibility for cytoplasmic viral regulators to toggle-switch between Xbp1 activation and mRNA decay.
In addition to RIDD, the cytoplasmic domain of phosphorylated Ire1α can interact with tumor necrosis factor (TNF)-receptor-associated factor (TRAF) 2, an adaptor protein coupling plasma membrane receptors to c-JUN N-terminal kinase (JNK) activation and pro-apoptotic stimulation (Urano et al. 2000). A recently discovered process by which terminally misfolded-proteins are degraded in the ER by a process termed ‘ER-associated degradation’ (ERAD) (reviewed in Smith, Ploegh and Weissman 2011). ERAD contributes to ER homeostasis by removing terminally misfolded proteins from the ER and targeting them for proteasomal degradation. Protein extraction from the ER involves the AAA+adenosine triphosphatase (ATPase) p97 (valosin-containing protein in humans) (reviewed in Metcalf et al. 2020). The ERAD is directly linked to the UPRER, as ER stress-induced Xbp1s transcriptionally upregulates components of the retrotranslocation machinery for protein transport from the ER to the cytosol (Iwakoshi, Lee and Glimcher 2003; Acosta-Alvear et al. 2007; Araki and Nagata 2012).
PERK
PERK is a type-I transmembrane protein with a lumenal stress sensing domain, and a cytoplasmic kinase domain which trans-autophosphorylates upon ER stress-induced oligomerisation. Unlike Ire1, PERK phosphorylates the alpha subunit of eukaryotic translation initiation factor 2 (eIF2α), and thereby suppresses translation initiation (Harding, Zhang and Ron 1999). PERK-mediated eIF2α phosphorylation inhibits the guanine-nucleotide exchange factor eIF2B. eIF2B accelerates the exchange of GDP for GTP in the eukaryotic initiation factor 2 (eIF2) complex (Ranu and London 1979). Although activation of PERK leads to a strong suppression of protein synthesis, some mRNAs with short inhibitory open reading frames (uORFs) in the 5’ untranslated region (5’UTR) resist translation inhibition by PERK. For example, the UPRER transcription factor ATF4 is expressed preferentially upon PERK activation (Lu et al. 2004; Vattem and Wek 2004). ATF4 enhances the pro-apoptotic C/EBP homologous protein (CHOP), which downregulates the anti-apoptotic protein Bcl-2 (B-cell lymphoma 2) and promotes cell death via cytochrome c release (Harding et al. 2000; Ma et al. 2002).
It is worth noting that ribosomal subunits are post-translationally modified in uninfected cells. For example, the induction of the UPRER induces site-specific ubiquitination on RPS3, RPS2 and RPS20 with possible functional consequences for translation (Higgins et al. 2015). Apart from a stressed ER, starvation or heme depletion can lead to phosphorylation of eIF2α. Additionally, protein kinase (PKR) is a cytoplasmic type-I IFN-induced enzyme, which can also phosphorylate eIF2α and enhance apoptosis by upregulating ATF4 and CHOP. Hence, this signalling arm is commonly referred to as the integrated stress response (ISR) (Harding et al. 2003). Recent findings suggest that ISR is persistently active in mice with traumatic brain injury leading to continuous eIF2α phosphorylation (Chou et al. 2017). The small-molecule drug-like compound ISRIB (ISR inhibitor) promotes eIF2B dimerization causing enhanced activity on its substrate eIF2 independent of upstream inhibition of eIF2α (Sekine et al. 2015; Sidrauski et al. 2015; Tsai et al. 2018; Zyryanova et al. 2018). Hence, ISRIB blunts the block on translation initiation, and was shown to enhance cognition in rodents with traumatic brain injury.
ATF6
ATF6 represents a third type of transmembrane ER stress sensor. When unfolded proteins accumulate in the ER lumen, ATF6 is transported to the Golgi apparatus in COPII vesicles (Schindler and Schekman 2009). At the Golgi, ATF6 gets processed by two proteases, S1P (site-1) and S2P (site-2) proteases, which remove the lumenal and transmembrane anchor, respectively (Haze et al. 1999; Ye et al. 2000). This gives rise to N-terminal ATF6 (ATF6-N), which acts as a potent UPRER transcription factor. In the nucleus, ATF6-N enhances UPRER gene transcription by binding to cis-acting ER stress response elements (ERSE). Prominent target genes include Xbp1 and BiP/Grp78 (Yoshida et al. 2000). ATF6-N also enhances CHOP mRNA levels and promotes pro-apoptotic signals in the terminal phase of the UPRER (Yoshida et al. 2000), together with the activating transcription factor 4 (ATF4), which is enhanced by PERK.
VIRUSES ACTIVATE AND SUPPRESS THE UPRER
The UPRER maintains homeostasis by multiple effector pathways, including a transcriptional upregulation of protein-folding enzymes (so-called chaperones), enhancement of the ERAD, and reduction of global protein synthesis. The type and duration of stress are crucial for the overall output of the UPRER. Under relatively mild ER stress, the Ire1-Xbp1s arm can remedy the detrimental effects of unfolded protein accumulation. The Ire1α activated transcription factor Xbp1s binds and enhances the expression of a subset of genes promoting cell survival, growth and differentiation, including protein biosynthesis and folding, trafficking and secretion (Acosta-Alvear et al. 2007). See Fig. 1.
Figure 1.
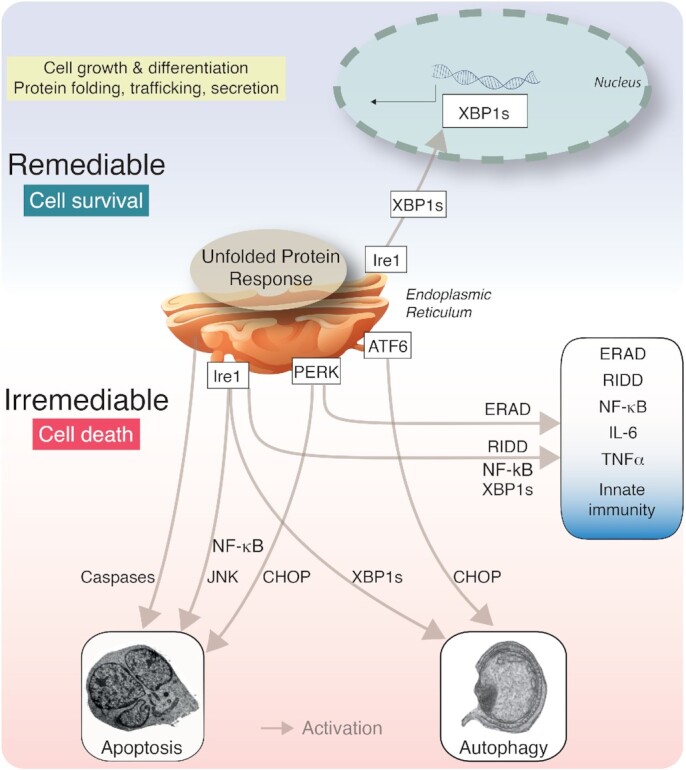
The major signalling channels of the UPRER in cell survival, death and innate immunity. Distinct signalling pathways downstream of the vertebrate UPRER sensors result in cell survival, death and innate immunity. Abbreviations: Ire1, Inositol-requiring enzyme 1; Xbp1s, X-box binding protein 1 spliced; ATF6, Activating transcription factor 6; PERK, Protein kinase activated by double stranded RNA (PKR)-like ER kinase; NF-κB, Nuclear factor kappa light-chain-enhancer of activated B cells; JNK, c-Jun N-terminal kinases; CHOP, CCAAT/-enhancer-binding protein homologous protein; RIDD, Regulated Ire1-dependent decay of mRNA; ERAD, ER-associated degradation; IL-6, Interleukin-6; TNFα, Tumour necrosis factor alpha.
Enveloped viruses commonly require large amounts of properly folded membrane glycoproteins leading to ER overload, and activation of a global UPRER involving all three sensors Ire1α, PERK and ATF6. Examples of viral proteins that bind and sequester the ER chaperone BiP/Grp78 away from the lumenal domain of Ire1α, PERK and ATF6 are listed in Fig. 2. The induction of a strong UPRER may blunt infection, for example, by attenuation of translation through PERK activation, triggering premature apoptotic cell death or immune responses (Walter and Ron 2011; Smith 2014). To balance UPRER signalling viruses have evolved strategies to either activate or inhibit particular arms of the UPRER, as depicted in Fig. 3. Below, we provide a discussion of select viruses that activate or suppress the UPRER.
Figure 2.

Viruses sequestering BiP/Grp78 in the ER lumen. Initiation of a global UPRER in virus infections can occur by the sequestration of the ER chaperone BiP/Grp78 from the lumenal domain of the UPRER sensor proteins Ire1α, PERK and ATF6. All signalling arms of the UPRER sensors are activated as a result of BiP/Grp78 removal from ER lumenal domains of the sensor. Abbreviations: BiP, Binding immunoglobulin protein; CNX, Calnexin; CRT, Calreticulin; HCV, Hepatitis C virus; PMV, Paramyxo simian virus; IAV, Influenza A virus; CMV, Cytomegalovirus; VSV, Vesicular stomatitis virus; ERAD, ER-associated segradation; Ire1, Inositol-requiring enzyme 1; PERK, Protein kinase activated by double stranded RNA (PKR)-like ER kinase; ATF6, Activating transcription factor 6.
Figure 3.

Viral proteins interfering with signal transduction along the three UPRER branches Ire1, PERK and ATF6. Examples of viruses and viral proteins that activate or inactivate specific arm of the UPRER signalling by direct or indirect interactions at the level of the sensors or downstream signal transducers. Abbreviations: BiP, Binding immunoglobulin protein; eIF2α, Eukaryotic translation initiation factor 2 subunit1; Ire1, Inositol-requiring enzyme 1; ATF6, Activating transcription factor 6; PERK, Protein kinase activated by double stranded RNA (PKR)-like ER kinase; HSV, Herpes simplex virus; HCV, Hepatitis C virus; HCMV, Human cytomegalovirus; CMV, cytomegalovirus; IAV, Influenza A virus; AdV, Adenovirus; ASFV, African swine fever virus; ATF4, Activating transcription factor 4; PP-1, Protein phosphatase 1; CHOP, CCAAT/-enhancer-binding protein homologous protein; BiM, Bcl-2 like protein 11; Bcl-2, B-cell lymphoma 2; AARE, Amino acid response element; EDEM, ERAD enhancing α-mannosidase-like proteins; Xbp1s, X-box binding protein 1 spliced; JNK, c-Jun N-terminal kinases; TRAF2, TNF receptor associated factor 2.
VIRAL ACTIVATION OF THE UPRER
The following nine enveloped viruses (in alphabetical order) induce the UPRER by ER overload or specific signals from the ER lumen.
African swine fever virus (ASFV) —ASFV infection enhanced the expression of ER chaperones, particularly calnexin and calreticulin (Galindo et al. 2012). This increase has been linked to the activation of ATF6 but a specific role of viral proteins is unknown.
Cytomegalovirus (CMV) —The CMV Us11 protein is sufficient to induce the UPRER as suggested by the upregulation of BiP levels and Xbp1 splicing (Tirosh et al. 2005). See Fig. 2. Recent studies showed that murine CMV (MCMV) early gene expression is suppressed by the unspliced form of the Xbp1 mRNA (Xbp1u) (Hinte et al. 2020). MCMV transiently activated the Ire1α-Xbp1 axis, depleted Xbp1u and relieved the transcriptional repression of the immediate early viral promoter boosting viral replication. The study also showed an unexpected role of Xbp1u as a potent repressor of both XBP1s and ATF6-mediated activation.
Dengue virus (DENV) —A generalised activation of UPRER pathways has been reported in DENV infection (Umareddy et al. 2007; Datan et al. 2016; Perera, Miller and Zitzmann 2017), without much information on viral proteins involved, although an induction of Xbp1s was reported (Yu et al. 2006).
Hepatitis C virus (HCV) —HCV encodes two envelope glycoproteins E1 and E2, which form non-covalent heterodimers and higher order oligomers. These glycoproteins bound to ER chaperones, including BiP, calnexin and calreticulin (Choukhi et al. 1998). The expression of E2 led to a generalised activation of all the UPRER sensors, and increased Xbp1s (Tardif et al. 2004; Chusri et al. 2016), phosphorylated PERK, cleaved ATF6 (Chusri et al. 2016), and an increase in BiP transcription (Liberman et al. 1999). See Fig. 2.
Influenza A (IAV) —Initially, misfolded hemagglutinin precursor protein HA0 was found to non-covalently associate with BiP (Hurtley et al. 1989) (Fig. 2). Later studies showed that IAV infection mainly activated the Ire1α pathway, whereas PERK and ATF6 activities were either unaffected or suppressed (Hassan et al. 2012). The stabilization of the UPRER by the bile component tauroursodeoxycholic acid (TUDCA) reduced Ire1α activation, activated PERK and phosphorylated eIF2α, decreased protein synthesis, and promoted the expression of ATF4 (Hassan et al. 2012; Kusaczuk 2019). Intriguingly, TUDCA suppressed IAV titers, possibly by unbalancing the UPRER network to maintain homeostasis. Alternatively, the ER stress-induced innate and adaptive immune responses, including NF-κB signalling (described in detail in section 4 and 6) and the type I IFN response may be harnessed in IAV infection to tune cell survival and virus output (Liu et al. 2012; So 2018).
Japanese encephalitis virus (JeV) —JeV belongs to the genus Flavivirus, and its infection induces all branches of the UPRER, as seen directly by PERK phosphorylation, Xbp1 splicing and ATF6 cleavage (Yu et al. 2006; Sharma et al. 2017).
Paramyxo simian virus 5 (PMV SV5) —Both the unfolded and folded forms of the viral hemagglutinin-neuraminidase protein were shown to form a complex with BiP and might be causing an induction of a broad UPRER (Ng et al. 1989). A subsequent study reported an increase in the transcription of UPRER genes with ectopic HN expression and in SV5 infection (Watowich, Morimoto and Lamb 1991). See Fig. 2.
SARS Coronavirus (SARS-CoV) —The 8ab protein of SARS-CoV, which locates to the lumen of the ER, induces ER-resident chaperones and ATF6 activation, apparently without PERK or IRE1α activation, as evidenced by absence of CHOP induction or and Xbp1 splicing, respectively (Sung et al. 2009) (Fig. 2).
The expression of 8ab led to ATF6 cleavage and promoted the nuclear translocation of its transcription-active amino terminal domain ATF6-N. A subsequent study showed that an 18-amino acid long peptide of 8ab interacted with the Ire1α lumenal domain in vitro (Karagoz et al. 2017). Whether this interaction leads to activation of Ire1α in cells has remained unknown. Another study reported that SARS-CoV spike protein enhanced BiP/Grp78 expression via PERK pathway of UPRER (Chan et al. 2006).
Vesicular stomatitis virus (VSV) —A specific population of VSV G-protein that formed incomplete disulphide bonds and transiently interacted with BiP/Grp78, albeit without specific information on the UPRER induction (Machamer et al. 1990). Later studies showed eIF2a phosphorylation as an indicator of PERK activation, and other downstream UPRER genes (Connor and Lyles 2005; Liu et al. 2009). See Fig. 2.
The following three nonenveloped viruses induce the UPRER by expressing viral nonstructural proteins in the ER.
Adenovirus (AdV) —Initially, AdV infection was found to be increased by small chemical compounds enhancing the UPRER, such as Golgicide A, or RNA interfence against genes controlling ER-Golgi trafficking (Prasad et al. 2014). The enhancing effects on the early viral gene expression were dependent on the Ire1α-Xbp1 axis of the UPRER. Subsequent studies showed that AdV infection also enhanced the UPRER. In particular, the ER lumenal domain of the viral glycoprotein E3-19K formed a complex and specifically activated the Ire1α branch of the UPRER for extended periods (Prasad et al. 2020). The activation of Ire1α in the context of infection or upon expression of ER-directed E3-19K lumenal domain alone enhanced the splicing of Xbp1u to Xbp1s mRNA. In the infected cells, E3-19K promoted early viral gene expression through Xbp1s binding to the E1 and the E4 promoters, as demonstrated by chromatin immunoprecipitation and E1 promoter mutagenesis. Ire1α activation by E3-19K occurred noncanonically, that is, without increase of BiP/Grp78, RIDD or PERK and ATF6 activations. In addition, pre-existing BiP dissociated from Ire1α before Ire1α activation measured by XBP1 splicing was observed. The extended Ire1α activation promoted the long term persistence of AdV in cell cultures in the presence of IFN. AdV mutants lacking E3-19K and pharmacological interference with the Ire1α nuclease activity abrogated persistence and virus disappearance from the cultures (Cross et al. 2012; Prasad et al. 2020).
Adeno-associated virus (AAV) —Transductions of cultured cells with recombinant self-complementary AAV1 and AAV6 (or AAV2 in hepatic transductions) were shown to induce Ire1α and PERK mRNA levels, and AAV6 also induced ATF6 mRNA suggesting viral capsid dependent effects on the UPRER sensors (Balakrishnan et al. 2013). RNA interference-mediated inhibition of Ire1α and PERK, however, gave only minimal effects on AAV2 and AAV6 transduction, suggesting that the UPRER sensor induction was not a proviral response. Whether a pharmacological inhibition of the UPRER can be applied in combination with AAV transduction in clinical settings remains an open question.
Coxsackievirus (CV) —Infection with CVB3 induced the canonical UPRER, with increased BiP levels, activated Ire1α and PERK, increased ATF6-N, and enhanced expression of UPRER target genes (Zhang et al. 2010). However, interactions and direct actions of stressors on the UPRER sensors and regulators have remained unknown.
VIRAL SUPPRESSION OF THE UPRER
The production of progeny in infected cells requires the synthesis of viral structural proteins in excess over those that are actually incorporated into the particles. This is because low affinity and high avidity cooperative interactions between virion proteins themselves and the viral genome control the assembly of the particles. Low affinity/high avidity assembly gives rise to infectious particles which are able to respond to host cues for uncoating in naïve cells (reviewed in Yamauchi and Greber 2016; Greber 2019; Greber and Flatt 2019). Interestingly, virus-like particles can be evolved to package RNA synthetically, but they remain unresponsive to the uncoating cues in the target cells, as for example indicated by recent laboratory evolution of the bacterial enzyme lumazine synthase from Aquifex bacteria (Tetter et al. 2020). This highlights the importance of combinatorial evolutionary selection processes in virus biogenesis, and puts an exciting perspective for the UPRER in synthetic biology, considering the possibility of evolvable protein cages with tunable assembly and disassembly functionality (Malay et al. 2019). Viruses in nature have solved the issue by expressing an excess of virion proteins, a situation that leads to protein overload in the ER and elicits a canonical UPRER with anti-viral responses, as listed above. Both enveloped and non-enveloped viruses have evolved mechanisms to modulate the anti-viral facettes of the UPRER, as discussed below (see also Fig. 3).
Here we list five enveloped and nonenveloped viruses, which circumvent or suppress aspects of the UPRER in the course of progeny formation.
Adenovirus (AdV) —As discussed above, the canonical UPRER induction and PERK activation are absent in AdV infections (Prasad et al. 2020). Nonetheless, AdV activates double-stranded RNA activated protein kinase (PKR). PKR is a cytoplasmic type-I IFN-induced enzyme, which phosporylates eIF2α and enhances apoptosis by upregulating ATF4 and CHOP (Lee et al. 2007). Since eIF2α can be phosphorylated by both PERK and PKR, translation inhibition in AdV infection occurs even without PERK activation. The virus uses several strategies to block translation inhibition by phosphorylated eIF2α. Late in infection, global translation shutoff is prevented by viral associated RNA I (VA RNA I), which is a PKR inhibitor and acts as a decoy of double-stranded RNA which normally activates PKR (Mathews and Shenk 1991). In addition, AdV E1B-55K and E4orf6 proteins form a ubiquitin ligase complex, which inhibits the phosphorylation of eIF2α (Harada et al. 2002; Spurgeon and Ornelles 2009).
African swine fever virus (ASFV) —During ASFV infection, inhibition of CHOP expression was reported (Netherton, Parsley and Wileman 2004). The ectopically expressed ASFV protein DP71L, a homolog of protein phosphatase 1 regulatory subunit 15A, interacted with protein phosphatase 1 catalytic subunit (PP1c), recruited PP1c to eIF2α and led to eIF2α dephosphorylation (Zhang et al. 2010). The importance of eIF2α dephosphorylation was further emphasized by the finding that mutant viruses lacking DP71L still reduced eIF2α phosphorylation and CHOP levels, suggesting that virus has redundant mechanisms for eIF2α dephosphorylation (Fig. 3).
Cytomegalovirus (CMV) —Not a lot is known about PERK and eIF2α suppression in CMV infection, but the Ire1-Xbp1 arm of the UPRER was suppressed by the murine CMV M50 protein. The N-terminal region of M50 was shown to be present in a complex with Ire1α leading to a decrease in Ire1α levels via an unknown degradation pathway (Stahl et al. 2013). Similar observations were made with UL50, the human homolog of ML50. We speculate that the reduction of Ire1α levels in CMV infections is key to prevent an overreaction of the UPRER, when the viral protein load in the ER increases for virion morphogenesis.
Herpes simplex virus (HSV) —Several strategies have been reported by which HSV circumvents the global translational shutdown downstream of an activated PERK branch. For example, the ICP0 promoter of HSV1 responded to ER stress, and speculatively, may tune the UPRER early in infection (Su et al. 2016). In addition, the viral glycoprotein gB binds to PERK and reduces its phosphorylation (Mulvey, Arias and Mohr 2007). Further, two viral proteins (Us11 and ϒ134.5) targeted eIF2α phosphorylation to prevent a global translation shutdown upon PERK activation (Cheng, Feng and He 2005). HSV ϒ134.5 was expressed before viral replication and interacted with cellular phosphatase PP1a to cause eIF2α dephosphorylation (Chou et al. 1995; Mulvey et al. 2003) (Fig. 3). The ϒ134.5 gene may provide an example of evolutionary mimicry, since it has a C-terminal domain homologous to GADD34, an activator of PP1a which dephosphorylates eIF2a and prevents host cell protein synthesis shutoff (Chou and Roizman 1994; He et al. 1997).
These examples illustrate apparently redundant HSV strategies for effectively maintaining translation in the background of an ongoing UPRER. In addition to the viral modulation of PERK activity, the ectopic expression of the viral protein UL41 has been shown to suppress Ire1α RNase function and Xbp1 splicing promoted by the ER stress inducer thapsigargin, which inhibits the Ca+2 ATPase in the ER and depletes Ca+2 stores in the ER (Zhang et al. 2017). This interference may effectively blunt an overreacting Ire1α response and preserve homeostasis in the infected cell. For recent overviews on the UPR in herpesvirus and porcine reproductive and respiratory syndrome virus infections, see (Gao et al. 2019; Johnston and McCormick 2019).
Hepatitis C virus (HCV) —Although the HCV E1 and E2 glycoproteins led to the activation of all UPRER sensors, several reports show that the downstream signalling of the UPRER is inhibited by HCV. Specifically, HCV replicon-containing cells showed enhanced Xbp1 splicing but transcriptional enhancement of Xbp1 target genes of the host was rather modest (Tardif et al. 2004). This suggests that Xbp1s target genes, such as EDEM (ER degradation-enhancing alpha-mannosidase-like protein) which enhances the degradation of misfolded proteins, require additional input besides Xbp1s for robust transcription stimulation. Accordingly, Ire1α-knockout cells displayed elevated levels of HCV glycoproteins, possibly because EDEM was reduced in these cells (Tardif et al. 2004). It can be speculated that HCV suppresses the Ire1-Xbp1 axis to promote viral envelope protein expression. An additional beneficial effect for the suppression of Ire1-Xbp1 by HCV might be to reduce the activity of transcription factors, which normally increase the pro-inflammatory cytokine production in response to viral pattern recognition receptor engagement (Smith 2014) (Fig. 4). A possible evolutionary mimicry for the shut down of UPRER has also been described for HCV infection. The HCV E2 protein contains a sequence identical to the phosphorylation sites of IFN-inducible PKR and its target eIF2α. By this interaction, E2 binds and inhibits activity of PKR and neutralizes its inhibitory effect on protein synthesis, thereby contributing to the IFN resistance of HCV infections (Taylor et al. 1999).
Figure 4.
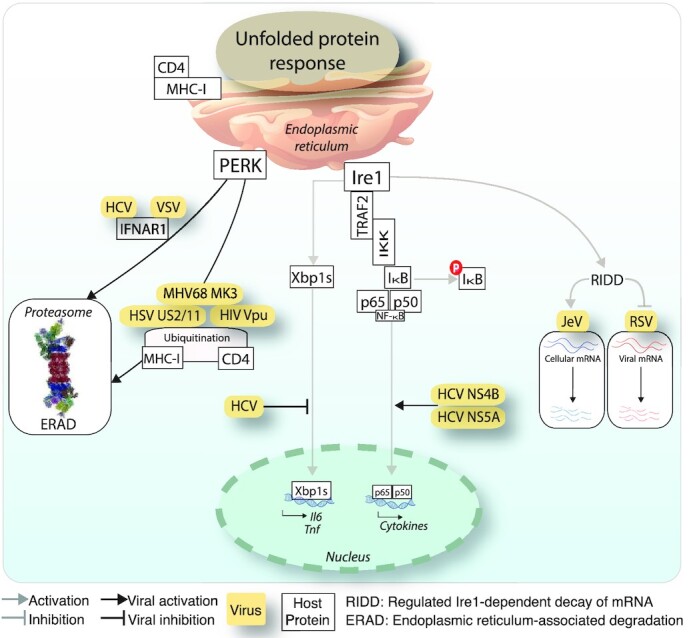
Viral modulation of innate immunity effectors interlinked with the Ire1 and PERK branches of the UPRER. Viruses intercept signalling from the Ire1-Xbp1 axis to innate immunity hubs by interfering with activated NF-κB or RIDD. This can trigger proviral effects by degradation of host mRNAs or raise susceptibility to antiviral effects, for example when RIDD degrades viral RNA. PERK signalling can be activated by viruses to target key innate immunity components, such as IFNAR1 or MHC-I for proteasomal degradation. Abbreviations: PERK, Protein kinase activated by double stranded RNA (PKR)-like ER kinase; Ire1, Inositol-requiring enzyme 1; TRAF2, TNF receptor associated factor 2; IKK, IκB kinase; CD4, Cluster of differentiation 4 glycoprotein; MHC-I, Major histocombatibility factor I; RIDD, Regulated Ire1-dependent decay of mRNA; JeV, Japanese encephalitis virus; RSV, Respiratory syncytial virus; ERAD, ER-associated degradation; IL-6, Interleukin-6; TNF, Tumour necrosis factor; HIV, Human immunodeficiency virus; HSV, Herpes Simplex Virus; MHV, Mouse gammaherpes virus; HCV, Hepatitis C virus; VSV, Vesicular stomatitis virus; IFNAR1, Interferon alpha or beta receptor subunit 1.
THE UPRER IN INNATE IMMUNITY
The immune response is key to the defense of cells and organisms against pathogens, and can severely affect the ER, and the restoration of homeostasis. ER stress and the UPRER play emerging roles in immunity and exacerbate the cell sensitivity to inflammatory stimuli (Bettigole and Glimcher 2015). The UPRER shares several signalling modules with immune response pathways (see Fig. 1). For example, increase in NF-κB transcriptional activity by PERK and Ire1α is linked to pro-inflammatory cytokine production in macrophages through NF-κB signalling. This happens through increased NF-κB levels compared to the short-lived NF-κB inhibitor IκB, notably in presence of translation shutdown by eIF2α phosphorylation (Deng et al. 2004). On the other hand, Ire1α activation of NF-κB signalling is linked to Ire1α interaction with the adaptor protein TRAF2 (Hu et al. 2006). Dimers of NF-κB p65 and p50 RelA subunits are in an inactive state sequestered in the cytoplasm by ankyrin-repeats of the inhibitor of κB (IκB) (reviewed in Napetschnig and Wu 2013). Interaction of Ire1α with TRAF2 recruits IκB kinase (IKK), which phosphorylates IκB and thereby releases NF-κB for nuclear translocation, and IκB for degradation (Liu et al. 1996; Hu et al. 2006). In addition, TLR antagonists enhance the recruitment of TRAF6 to Ire1α via the NADPH oxidase protein, NOX2, thereby activating Ire1α and enhancing Xbp1 splicing (Martinon et al. 2010). This activation of Ire1α is important for cytokine production, for example interleukin-6 (IL-6), TNF and IFNß. Innate immune signalling from TNFα further induces Ire1α activity and leads to NF-κB dependent apoptotic cell death. On the contrary, the activation of glycogen synthase kinase 3ß (GSK3ß) by Ire1α together with TNF upregulation has been implicated in the down-modulation of Xbp1 transcription, and thereby fine-tunes the inflammatory response (Kim et al. 2015). Further, the survival of infected macrophages is supported by TLR activation, which favours the suppression of the ATF4/CHOP signalling in the PERK branch (Metcalf et al. 2020).
Below, we depict major overlaps between UPRER branches and innate immune responses, and discuss examples of viral down-modulation of the corresponding elements to evade cell death and clearance by the immune system.
Innate immunity and pro-inflammatory cytokines Work with flaviviruses, such as Dengue, Zika, West Nile and Tick-borne encephalitis viruses suggested that the UPRER can accelerate innate immunity by boosting the expression of IFN-stimulated genes, such as the IFN regulatory factor (IRF) 3, and thereby enhance an antiviral response (Carletti et al. 2019). HCV inhibition of Xbp1 transactivation provides an example for a viral strategy of immune evasion, which might contribute to viral persistence in hepatocytes (Tardif et al. 2004). The mechanisms are unknown, however, and it remains to be elucidated how Xbp1 down-modulation supports immune evasion of HCV.
NF-κB—The HCV proteins NS5A and NS4B were described to trigger the nuclear translocation of NF-κB, induce the UPRER, as well as reactive oxygen species (Gong et al. 2001; Li et al. 2009). Similarly, the AdV glycoprotein E3-19K causes NF-κB nuclear translocation (Pahl et al. 1996), possibly through the activation of Ire1α (Prasad et al. 2020). Interestingly, the AdV E3 promoter contains NF-κB but not Xbp1s binding motifs, giving rise to the possibility that NF-κB contributes to the maintenance of E3 levels, and thereby enhances immune defense and virus persistence (Williams et al. 1990; Lichtenstein et al. 2004; Prasad et al. 2020).
Virus-triggered ERAD of host innate immunity proteins —Several viruses were reported to use ERAD to degrade host components with anti-viral activity (for a review see Frabutt and Zheng 2016). For example, HCMV US2 and US11 proteins bound and dislocated MHC-I heavy chains for degradation in the cytoplasm (van der Wal, Kikkert and Wiertz 2002). Similarly, murine gammaherpes 68 (MHV68) encodes an E3-ubiquitin ligase MK3, which associated with a cellular E2 ubiquitin-conjugating enzyme and MHC-I, leading to the ubiquitylation and ERAD-mediated proteasomal degradation of MHC-I (Boname and Stevenson 2001). In HIV-1 infected cells the viral accessory protein Vpu promoted virion release and prevents superinfection by downregulating the virus receptor CD4 from the cell surface (Margottin et al. 1998; Magadan et al. 2010). Vpu interacted with CD4 in the ER and induced CD4 degradation depending on ERAD by forming an ion conductive membrane pore and retargeting of a E3-ubiquitin ligase to the ER (Strebel 2014). See Fig. 4. VSV infection degrades host proteins involved in innate immunity. A specific activation of the PERK pathway led to the proteasomal degradation of type-I IFN receptor (IFNAR) 1, reducing the anti-viral IFN signalling (Baltzis et al. 2004; Krishnamoorthy et al. 2008). This finding was further supported by experiments with PERK knockdown cells where the viral protein expression was reduced and IFN signalling restored (Fig. 4). A similar mechanism of virus-induced IFNAR1 degradation was shown with HCV infection (Liu et al. 2009).
Virus-triggered degradation of host mRNA by RIDD —ER stress can induce hyper-phoshorylation of UPRER effectors. Under these conditions, Ire1α catalyzed RIDD, non-specific cleavage of mRNAs (Hollien et al. 2009, Walter and Ron 2011). It remains unknown if these RNA fragments were sensed by RIG-I and thereby led to the activation of NF-κB and inflammatory cytokine production, such as IL-6 and IL-8, as suggested for bacteria (reviewed in Lencer et al. 2015). In JeV infected cells, RIDD degraded host RNAs without affecting viral RNAs (Bhattacharyya, Sen and Vrati 2014). Blunting Ire1-RIDD activity with chemical inhibitors reduced viral titers suggesting a pro-viral effect of RIDD. However, the molecular mechanisms by which the viral transcripts escape RIDD have remained unclear. In another study, Ire1 activation in respiratory syncytial virus (RSV) infection reduced number of viral transcripts and translation products, suggesting that RIDD degraded viral transcripts as part of an anti-viral response (Hassan et al. 2014). See Fig. 4.
THE UPRER IN AUTOPHAGY
Autophagy is a conserved self-eating process to encapsulate intracellular components into autophagosomes, including organelles, soluble and aggregated proteins and foreign bodies, and content degradation in lysosomes (reviewed in Yu et al. 2018). The sustained activation of the Ire1α-Xbp1 axis has been implicated in triggering an autophagic response by the transcriptional enhancement of autophagy effectors Beclin-1 and LC3 (Adolph et al. 2013; Margariti et al. 2013). In addition, the UPRER controlled transcription factors ATF4, CHOP, NF-κB and STAT3 upregulate the autophagosome machinery (reviewed in Pietrocola et al. 2013). It is conceivable that damaged ER proteins exceeding the degradation capacity of the ERAD lead to the turnover of ER segments by autophagy clearing proteins and also lipids (Houck et al. 2014). This goes along with the notion that autophagy counterbalances ER expansion during the UPRER (Bernales, McDonald and Walter 2006).
Below, we discuss examples of enveloped viruses using autophagic activity in connection with the UPRER.
Coronaviruses (CoVs) —Ire1α was found to be activated in cells infected with avian infectious bronchitis virus (IVB) and served as a survival factor by antagonizing apoptosis through modulating the phosphorylation status of proapoptotic JNK and the prosurvival through RAC-alpha serine/threonine-protein kinase (Akt) (Fung, Liao and Liu 2014). The data suggest that the UPRER constitutes a major aspect of coronavirus-host interactions.
Hepatitis C virus (HCV) —The HCV-induced UPRER activates autophagy, and thereby enhances viral RNA replication (Ke and Chen 2011). Enhanced RNA replication depended on autophagic protein degradation, and the UPRER might have been involved in suppressing IFNß activation and immune response to infection. Notably, the IFNß activation in response to HCV PAMPs occurred only in the absence of host UPRER or absence of autophagy components. In addition, HCV inhibited the AKT-TSC-MTORC1 pathway via ER stress, and may thereby contribute to the establishment of the HCV-induced autophagy (Huang et al. 2013).
Dengue virus (DENV) —DENV associated pathogenicity and viral load were shown to depend on the PERK-eIF2α and Ire1α-JNK signalling arms of the UPRER. DENV infection induced autophagy by phosphorylating Bcl-2 via the JNK signalling pathway, and promoted the dissociation of its complex with Beclin-1, thereby freeing up Beclin-1 to assemble with pre-autophagosomes (Marquez and Xu 2012). Chemical inhibition of JNK signalling reduced virus-mediated autophagy and virus titers in mice, suggesting a pro-viral role of autophagy in DENV infection (Lee et al. 2018). DENV infection also led to the degradation of host innate immune components by autophagy, akin to HCV infections (Ke and Chen 2011).
Japanese encephalopathy virus (JeV) —In contrast to HCV and DENV, ER stress induction in JeV infection enhanced autophagy, and reduced the viral titers and cell death. Silencing of the Xbp1 and ATF6 branches of the UPRER led to a suppression of JeV induced autophagy, and enhanced viral pathogenicity (Sharma et al. 2017)
Foot-and-mouth disease virus (FMDV)— Recently, FMDV-induced ER stress via PERK activation has been suggested to lead to enhanced autophagy. Knockdown of both PERK and components of the autophagy machinery reduced the viral titers and enhanced antiviral innate interferon response (Ranjitha et al. 2020).
THE UPRER IN CELL DEATH
Persistent and high intensity UPRER signalling can trigger an irremediable cell death program (see Fig. 1). This has been shown, for example, with lethal doses of the ER stress-inducing small chemicals thapsigargin or tunicamycin that trigger only transient Ire1α and ATF6 activity, but long-lasting PERK activity (Lin et al. 2007). Ire1α triggers pro-apoptotic signalling via interaction with the adaptor protein TRAF2, resulting in the activation of c-Jun N-terminal kinases (JNK) and nuclear factor kappa enhancer of B-cell (NF-κB) signalling (Urano et al. 2000; Hu et al. 2006). Binding of TRAF2 to Ire1α specifically leads to recruitment and activation cleavage of pro-caspases on the ER membrane, such as caspase-12 in mice and caspase-4 in humans (Nakagawa and Yuan 2000; Yoneda et al. 2001; Rao et al. 2002; Hitomi et al. 2004; Rosati et al. 2010). NF-κB in turn transcriptionally regulates pro-inflammatory genes with either pro-survival or apototic effects. Besides binding of TRAF2 and NF-κB, the cytosolic domain of Ire1α also binds the pro-apoptotic proteins Bcl-2 associated X-protein (Bax) and Bcl-2 homologous antagonist or killer (Bak). This binding promotes the splicing of Xbp1 mRNA, and may have a prosurvival effect, further highlighting the fine balance between UPRER signalling in cell homeostasis and death (Hetz et al. 2006).
Besides Ire1 signalling, PERK and ATF6 activations increase the levels of the proapoptotic transcription factor CHOP, which enhances the Bcl-2 family protein Bim and promotes apoptosis (McCullough et al. 2001; Puthalakath et al. 2007) (Fig. 1). UPRER-triggered cell death pathways were shown to be anti-viral, and limit virus dissemination, their effects were not investigated in detail.
Below we list examples of enveloped and non-enveloped viruses that induce UPRER associated apoptosis. For details, see also Fig. 5.
Figure 5.
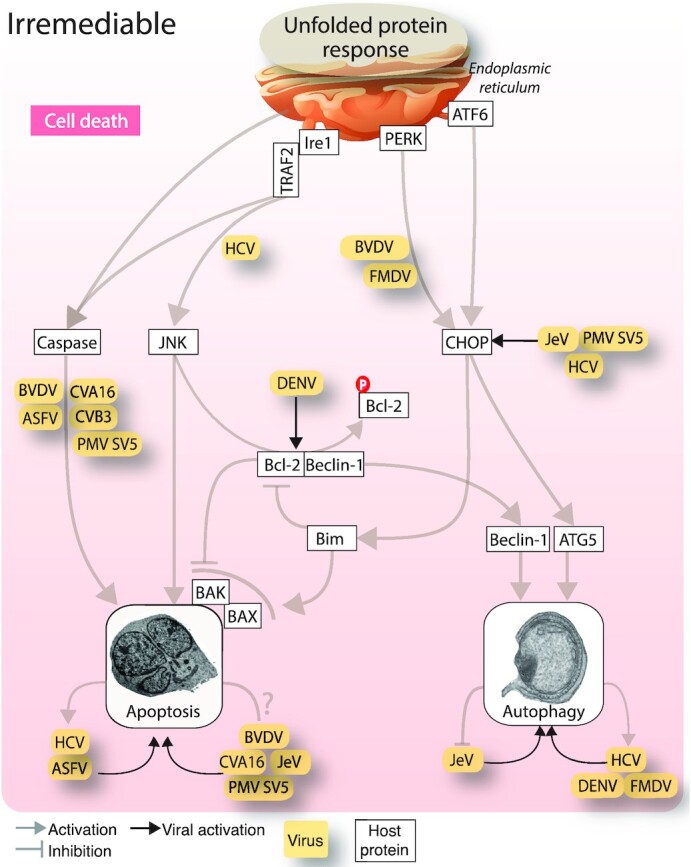
Viral modulation of UPRER controlled cell death pathways. Viruses and viral proteins use all the three UPRER sensor proteins for modulating cell death. The Ire1α-TRAF2-JNK signalling arm is used to promote apoptotic cell death. Several viruses promote ER stress dependent apoptosis via the caspase cascade. The PERK and ATF6 arms of the UPRER are targeted for the induction of autophagy via the CHOP transcription factor. Abbreviations: JNK, c-Jun N-terminal kinases; Ire1, Inositol-requiring enzyme 1; PERK, Protein kinase activated by double stranded RNA (PKR)-like ER kinase; ATF6, Activating transcription factor 6; BVDV, Bovine viral diarrhea virus; HCV, Hepatitis C virus; ASFV, African swine fever virus; CVA16, Coxsackievirus A16; CVB3, Coxsackievirus B3; JeV, Japanese encephalitis virus; PMV, Paramyxo simian virus; DENV, Dengue virus; TRAF2, TNF receptor associated factor 2; Bak, Bcl-2 homologous antagonist or killer; BAX, Bcl-2 associated X-protein; Bim, Bcl-2 like protein 11.
African swine fever virus (ASFV) —Increased caspase 3, 9 and 12 levels and apoptosis induction appear to be beneficial for the release of ASFV, as suggested by a reduction in virus egress from cells treated with caspase-3 inhibitors (Galindo et al. 2012). The enhanced caspase-12 is an ER-stress activated apoptotic response that occurs in ASFV infection primarily via ATF6 branch of the UPRER activation (Nakagawa et al. 2000). See Fig. 5.
Bovine viral diarrhea virus (BVDV) —BVDV is a positive-strand RNA virus of the genus pestivirus, and along with other members of the Flaviviridae family, uses the host ER as the primary site of replication and progeny assembly. BVDV infection is associated with pathogenicity and linked to the activation of PERK, as suggested by hyper-phosphorylation of eIF2α and caspase-12 meditated apoptotic cell death (Jordan et al. 2002).
Coxsackievirus (CV) —CVA16 infection showed an increase in caspase-3, 8 and 9 dependent apoptotic death. Apoptotic death was blunted by the chemical chaperone 4-methyl butyric acid, which increases the folding capacity of ER and reduces the UPRER (Zhu et al. 2013). The role of UPRER sensors in triggering BVDV apoptosis is unknown. In addition to the involvement of caspase-3, 8 and 9, an increase in the levels of caspase-7 and 12 and increased apoptotic cell death was observed in CVB3 infection (Zhang et al. 2010).
Hepatitis C virus (HCV) —A replicon expressing the HCV core protein in cultured cells, and the liver of transgenic HCV infected mice showed progressive depletion of ER Ca2+ stores and an increase in the pro-apoptotic UPRER induced factor CHOP, leading to an induction of apoptosis, possibly contributing to HCV-induced chronic liver disease (Benali-Furet et al. 2005). Whether PERK and ATF6 activations contributed to increased CHOP levels was not reported. Since HCV suppresses PERK signalling (Fig. 3), increased CHOP levels could at least in part be due to ATF6. Accordingly, HCV infection enhanced the UPRER via Ire1α−JNK pathway which could contribute to triggering cell death pathways (Chusri et al. 2016). See Fig. 2.
Japanese encephalitis virus (JeV) —A study reported that the UPRER induction in JeV-infected cells increased the CHOP protein levels and enhanced apoptotic death (Su et al. 2002). An overall UPRER might be responsible for this effect (Fig. 3).
Paramyxo simian virus 5 (PMV SV5)— Apoptotic cell death was induced by PMV SV5 involving an increase in the levels of ER stress activated host factors, including BiP, CHOP, Calnexin, suggesting a contribution by the virus-induced UPRER (Sun et al. 2004) (Bitko and Barik 2001). The increase in cell death during SV5 infection was linked to caspase-3 and ER-stress activated caspase-12 (Fig. 5).
THE UPRER AND ONCOLYTIC VIRUSES
Cancer cells exhibit aggravated UPRER, which is part of their program to establish homeostasis in the context of the organism (Hsu et al. 2019). This offers a potential angle to repurpose viruses towards oncolytic therapy, if the viruses of interest normally benefit from the UPRER. Several in vivo and in vitro evaluations of different oncolytic viruses in the context of the UPRER have been reported. Oncolytic viruses are engineered such that they preferentially infect and kill cancerous cells and spare the normal cells (for reviews, see Russell, Peng and Bell 2012; Alemany 2013; Gao et al. 2019; Georgi and Greber 2020; Lemos de Matos, Franco and McFadden 2020). Increased virus-induced cancer cell death via UPRER induction has been demonstrated in tissue culture. For example, the enhancement of UPRER by pharmacological agents targeting the early secretory pathway induced the UPRER and increased AdV gene expression and lytic cell death (Prasad et al. 2014). The effect occurred through the Ire1-Xbp1 axis of the UPRER. On the other hand, pharmacological inhibition of Ire1α and PERK decreased autophagy and enhanced alphavirus M1 oncolytic activity in mice (Li et al. 2018).
Akin to oncolytic viruses, chemotherapeutic drugs modulate the tumour microenvironment and can enhance the anti-tumour immune response (Grootjans et al. 2016). Combining these two anti-tumour modalities has the potential to improve anti-tumour efficacy at low toxicity in vitro and in vivo (Simpson et al. 2016). For example, a combination therapy of ERK1/2 inhibitors enhanced the oncolytic efficiency of reovirus via induction of UPRER in murine melanoma tumours (Roulstone et al. 2015). Enhancement of the UPRER by the FDA-approved proteasome inhibitor Bortezomib increased oncolytic HSV-1 replication in several cancerous cell lines and also in a murine glioma cancer model (Yoo et al. 2014). In a murine abdominal cancer model, inhibitors against Ire1α turned out to sensitise cancer cells towards caspase-2 dependent apoptosis after rhabdovirus infection (Mahoney et al. 2011). Similarly, the UPRER induction by Bortezomib showed the activation of EBV lytic switch gene ZTA in Burkitt lymphoma cells via enhanced expression of CCAAT/enhancer-binding protein (C/EBP) ß (Shirley et al. 2011). Similarly, the UPRER inducer thapsigargin, which inhibits the ER Ca+2 ATPase, appeared to trigger a lytic switch, although underlying mechanisms were not identified (Shirley et al. 2011). The fine balance between homeostasis and apoptotic induction by the UPRER, now requires more mechanistic knowledge of virus interactions with the UPRER, and drug synergy experiments, before this field is ripe for clinical applications.
EVOLUTIONARY IMPACT OF VIRUSES ON THE UPRER
UPRER sensors in the context of cell stress
So far, we have discussed evidence for virus interactions with the UPRER, and how the UPRER maintains cell homeostasis, survival and the onset of death processes in virus infections. Other interconnections of the UPRER to cell stress pathways are emerging, such as anti-viral response signalling, innate immunity through both cytokines and intrinsic factors, reactive oxygen signalling and metabolic pathways (Rouse and Sehrawat 2010; Wolfrum and Greber 2013; Thaker, Ch'ng and Christofk 2019). For example, the NLRP3-caspase-2 dependent signalling can integrate the UPRER and innate immunity and relay it to the mitochondria to promote inflammation (Bronner et al. 2015; Lencer et al. 2015). ER stress also regulates innate and adaptive immune responses, including NF-κB signalling and the type I IFN response (Liu et al. 2012). Oxidative stress further triggers the UPRER in vertebrates, and installs a feed-forward loop to remedy oxidative damage (Schwarz 1996). This is akin to the activation of Ire1-Hac1 in yeast, which upregulates genes protecting cells from oxidative stress, such as TSA1, a thioredoxin peroxidase catalyzing the reduction of peroxides and acting as a molecular chaperone (Kimata et al. 2006).
These signalling networks interlinking with the UPRER provide rich opportunities to explore evolutionary adaptation. For example, viruses toggle switch between lytic and persistent infection outcomes, as shown with herpes viruses and B-cell receptor signalling through Xbp1, which triggers the reactivation of latent KSHV into a lytic cycle (Kati et al. 2013; Johnston and McCormick 2019). Evolutionary impact of KSHV and other viruses, such as EBV, on the UPRER may have facilitated the development of vertebrate pathways to synthesize large loads of protein, for example immunoglobulins from B cells or collagen from notochord cells during development (Sun and Thorley-Lawson 2007; Ishikawa et al. 2017; Mrozek-Gorska et al. 2019). In addition, remodeling of ER membranes during virus infections to set up membranous webs or membrane zippering may further contribute to evolutionary adaptation of the ER during global UPRER (Sriburi et al. 2004; Romero-Brey and Bartenschlager 2016).
An evolutionary impact of viruses on the vertebrate UPRER may occur through sensors detecting both protein and lipid stress (Kono, Amin-Wetzel and Ron 2017; Preissler and Ron 2019). Both Ire1α and PERK are dual sensors, which may recognize specific features in pathogenic processes. For example, the AdV E3-19K glycoprotein specifically activates the Ire1α branch without inducing RIDD or cell death pathways (Prasad et al. 2020). Evolutionary tuning of the UPRER by viruses is further supported by the observation that viral proteins utilize both direct binding to the sensors or sequestration of BiP away from the sensors to trigger a UPRER response (Perera, Miller and Zitzmann 2017; Prasad et al. 2020). Such dual modality in sensor activation has recently been simulated in mathematical models, and was suggested to best account for the full activation spectrum of the UPRER (Stroberg, Eilertsen and Schnell 2019).
Yeast Ire1-mediated mobilization of retroelements is disabled in vertebrates
In vertebrates, the UPRER is a conserved three-pronged stress response that primarily restores homeostasis, where Ire1 is its most conserved arm. Lower eukaryotic cells have less complex UPRER pathways. The simple eukaryote Saccharomyces cerevisiae has just one ER stress sensor, yIre1 (Cox, Shamu and Walter 1993; Mori et al. 1993). yIre1 is thought to directly bind to unfolded proteins in the ER lumen and activate its cytosolic RNase domain leading to splicing of the precursor HAC1u to the mature HAC1i mRNA, which is translated to the functional transcription factor Hac1p (Mori et al. 2000; Ruegsegger, Leber and Walter 2001). Intriguingly, functional genetics studies in S. cerevisiae indicated that the dormancy of transposable elements can be modulated by the UPRER, and, in addition, by a variety of extrinsic and intrinsic cues, including stress signalling through mitogen-activated kinases, DNA damage, environmental signals such as temperature and nutrient availability (Carr, Bensasson and Bergman 2012).
We surmise that the large abundance of retroviruses in vertebrates has disabled a modality of the Ire1-Xbp1, such that Ire1 no longer activates genomic retrotransposons. Specifically, one of the two major retrotransposons in S. cerevisiae, Ty2 is transcriptionally upregulated by the yIre1-Hac1 pathway of the UPR, as shown in cDNA microarray analyses and knock-out strains (Kimata et al. 2006). HAC1 mutagenesis generated cells which expressed Hac1i mRNA without Ire1 activation, and these cells induced Ty2 transcription, indicating the importance of the primordial Ire1 UPRER arm in controlling retroelements (Kimata et al. 2006). Importantly, S. cerevisiae responds to the potent ER stressors dithiothreitol and tunicamycin by upregulating UPRER target genes, including those involved in ERAD, intracellular vesicle transport and lipid biosynthesis, and downregulating genes encoding proteins destined to the secretory pathway (Travers et al. 2000; Kimata et al. 2006). Although vertebrate genomes harbor large amounts of transposable elements, most of these elements are inactive and strongly suppressed, whereas yeast cells, which have only a few genome % retroelements, show high levels of retrotransposition events (Curcio, Lutz and Lesage 2015; Sotero-Caio et al. 2017). This argues for a very tight control of retroelements in vertebrate cells. In fact, retrotransposons dramatically enhance alterations in the genome by mediating chromosomal rearrangements, including deletions, segmental duplications, inversions and reciprocal and non-reciprocal translocations (Curcio, Lutz and Lesage 2015).
The Ty elements are the evolutionary progenitors of retroviruses in vertebrates. Transcription of these elements is initiated at the 5′ LTR and gives rise to an RNA from which the element-encoded proteins Gag and Gag-Pol are translated, comprising protease, integrase, reverse transcriptase and RNase H. Together with host proteins, Gag-Pol are assembled into virus-like particles, processed by the protease, and thereby serve as essential replication intermediates for the reverse transcription of the RNA into DNA and genomic insertion. The introduction of Ty2 retroelements into an ancestor of S. cerevisiae occurred rather recently as a result of horizontal transfer, whereas the Ty1 elements, which no longer respond to the UPRER, are more ancient (Kimata et al. 2006; Carr, Bensasson and Bergman 2012; Curcio, Lutz and Lesage 2015). It thus appears that the response of retrotransposons to the Ire1-Xbp1/Hac1 activation pathway declines in the course of evolution, and the decline correlates with increased genomic load of retroelements. One could argue that the high abundance of mobile genetic elements, including endogenous and exogenous retroviruses in vertebrates, selects for cells that no longer use the UPRER for boosting genomic rearrangements by their retroelements.
The proper control of retrotransposons is crucial for cell and organismic survival. For example, the impaired silencing of retrotransposons has been shown to trigger the excessive expression of retroviral env glycoproteins and thereby activate a general UPRER, causally linked to increased pro-B cell death through inactivation of the epigenetic regulator Setdb1 and an increase in histone H3-lysine 4 trimethylation (Pasquarella et al. 2016). This phenotype is exacerbated by the expression of enhanced levels of double-stranded RNA from endogenous retroviruses, and by triggering pattern-recognition receptors, such as RIG-I, and IFN (Roulois et al. 2015). We surmise that endogenous retroviruses exert evolutionary force on the cell death pathways of UPRER and synergize with interconnected innate immunity pathways to reach organismic homeostasis. This supports the possibility that the UPRER coevolved with multicellular eukaryotes, where cells of the immune system have adopted specialized functions requiring adaptations of the ER and its UPR.
CONCLUSIONS AND OUTLOOK
Viruses have a long history of inducing stress responses in their hosts. Stress responses are multifaceted and interconnected, and are accessible to tuning by the pathogens. They are evolutionary conserved, and reach back to bacterial cells, where phages increase the levels of heat shock proteins to restore homeostasis upon stress insult (Drahos and Hendrix 1982; Young 1990). Together with the pathogen, stress responses define the outcome of the infection, cytoprotective or cytotoxic. The UPRER is a significant eukaryotic stress response, controlling cell survival or death. In communicating with other signalling pathways it modulates innate immune and metabolic responses. This review illustrated how the UPRER is triggered by viruses, and how viruses overcome the antiviral effects of the UPRER. A deeper understanding of how viruses interact with the UPRER will require more mechanistic studies and also evolutionary insights. Chemical genetics, small molecule modulators of the UPRER, and the power of virology will provide the field with new opportunities not only for developing anti-viral interference but also for therapies against nonviral diseases exacerbated by the UPRER. For example, we envision that lipid homeostasis and the UPRER in viral infections will be important to understand how viruses switch between lytic and nonlytic propagation. Notably, key enzymes of lipid metabolism locate to the ER and make this organelle a primary hub for both structural and metabolic components in cell homeostasis.
ACKNOWLEDGEMENTS
We thank Dr Maarit Suomalainen for comments to the manuscript.
Contributor Information
Vibhu Prasad, Department of Molecular Life Sciences, University of Zurich, Winterthurerstrasse 190, 8057 Zurich, Switzerland.
Urs F Greber, Department of Molecular Life Sciences, University of Zurich, Winterthurerstrasse 190, 8057 Zurich, Switzerland.
FUNDING
VP acknowledges a long-term fellowship from European Molecular Biology Organization (ALTF 454–2020) and UFG funding from the Swiss National Science Foundation (31003A_179 256/1).
Conflicts of interest
None declared.
REFERENCES
- Acosta-Alvear D, Zhou Y, Blais Aet al. . XBP1 controls diverse cell type- and condition-specific transcriptional regulatory networks. Mol Cell. 2007;27:53–66. [DOI] [PubMed] [Google Scholar]
- Adolph TE, Tomczak MF, Niederreiter Let al. . Paneth cells as a site of origin for intestinal inflammation. Nature. 2013;503:272–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alemany R. Viruses in cancer treatment. Clin Transl Oncol. 2013;15:182–8. [DOI] [PubMed] [Google Scholar]
- Amin-Wetzel N, Saunders RA, Kamphuis MJet al. . A J-Protein Co-chaperone Recruits BiP to Monomerize IRE1 and Repress the Unfolded Protein Response. Cell. 2017;171:1625–1637 e1613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aragon T, van Anken E, Pincus Det al. . Messenger RNA targeting to endoplasmic reticulum stress signalling sites. Nature. 2009;457:736–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Araki K, Nagata K. Protein folding and quality control in the ER. Cold Spring Harb Perspect Biol. 2012;4:a015438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balakrishnan B, Sen D, Hareendran Set al. . Activation of the cellular unfolded protein response by recombinant adeno-associated virus vectors. PLoS One. 2013;8:e53845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baltzis D, Qu LK, Papadopoulou Set al. . Resistance to vesicular stomatitis virus infection requires a functional cross talk between the eukaryotic translation initiation factor 2alpha kinases PERK and PKR. J Virol. 2004;78:12747–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barlowe CK, Miller EA. Secretory protein biogenesis and traffic in the early secretory pathway. Genetics. 2013;193:383–410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benali-Furet NL, Chami M, Houel Let al. . Hepatitis C virus core triggers apoptosis in liver cells by inducing ER stress and ER calcium depletion. Oncogene. 2005;24:4921–33. [DOI] [PubMed] [Google Scholar]
- Bernales S, McDonald KL, Walter P. Autophagy counterbalances endoplasmic reticulum expansion during the unfolded protein response. PLoS Biol. 2006;4:e423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertolotti A, Zhang Y, Hendershot LMet al. . Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat Cell Biol. 2000;2:326–32. [DOI] [PubMed] [Google Scholar]
- Bettigole SE, Glimcher LH. Endoplasmic reticulum stress in immunity. Annu Rev Immunol. 2015;33:107–38. [DOI] [PubMed] [Google Scholar]
- Bhattacharyya S, Sen U, Vrati S. Regulated IRE1-dependent decay pathway is activated during Japanese encephalitis virus-induced unfolded protein response and benefits viral replication. J Gen Virol. 2014;95:71–79. [DOI] [PubMed] [Google Scholar]
- Bitko V, Barik S. An endoplasmic reticulum-specific stress-activated caspase (caspase-12) is implicated in the apoptosis of A549 epithelial cells by respiratory syncytial virus. J Cell Biochem. 2001;80:441–54. [DOI] [PubMed] [Google Scholar]
- Blanco-Melo D, Nilsson-Payant BE, Liu WCet al. . Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell. 2020;181:1036–1045 e1039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boname JM, Stevenson PG. MHC class I ubiquitination by a viral PHD/LAP finger protein. Immunity. 2001;15:627–36. [DOI] [PubMed] [Google Scholar]
- Braakman I, Hebert DN. Protein folding in the endoplasmic reticulum. Cold Spring Harb Perspect Biol. 2013;5:a013201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bronner DN, Abuaita BH, Chen Xet al. . Endoplasmic Reticulum Stress Activates the Inflammasome via NLRP3- and Caspase-2-Driven Mitochondrial Damage. Immunity. 2015;43:451–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Byrne AJ, Mathie SA, Gregory LGet al. . Pulmonary macrophages: key players in the innate defence of the airways. Thorax. 2015;70:1189–96. [DOI] [PubMed] [Google Scholar]
- Carletti T, Zakaria MK, Faoro Vet al. . Viral priming of cell intrinsic innate antiviral signaling by the unfolded protein response. Nat Commun. 2019;10:3889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrara M, Prischi F, Nowak PRet al. . Noncanonical binding of BiP ATPase domain to Ire1 and Perk is dissociated by unfolded protein CH1 to initiate ER stress signaling. eLife. 2015;4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr M, Bensasson D, Bergman CM. Evolutionary genomics of transposable elements in Saccharomyces cerevisiae. PLoS One. 2012;7:e50978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan CP, Siu KL, Chin KTet al. . Modulation of the unfolded protein response by the severe acute respiratory syndrome coronavirus spike protein. J Virol. 2006;80:9279–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan SW. The unfolded protein response in virus infections. Front Microbiol. 2014;5:518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chatel-Chaix L, Cortese M, Romero-Brey Iet al. . Dengue Virus Perturbs Mitochondrial Morphodynamics to Dampen Innate Immune Responses. Cell Host Microbe. 2016;20:342–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng G, Feng Z, He B. Herpes simplex virus 1 infection activates the endoplasmic reticulum resident kinase PERK and mediates eIF-2alpha dephosphorylation by the gamma(1)34.5 protein. J Virol. 2005;79:1379–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou A, Krukowski K, Jopson Tet al. . Inhibition of the integrated stress response reverses cognitive deficits after traumatic brain injury. Proc Natl Acad Sci USA. 2017;114:E6420–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou J, Chen JJ, Gross Met al. . Association of a M(r) 90,000 phosphoprotein with protein kinase PKR in cells exhibiting enhanced phosphorylation of translation initiation factor eIF-2 alpha and premature shutoff of protein synthesis after infection with gamma 134.5- mutants of herpes simplex virus 1. Proc Natl Acad Sci USA. 1995;92:10516–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou J, Roizman B. Herpes simplex virus 1 gamma(1)34.5 gene function, which blocks the host response to infection, maps in the homologous domain of the genes expressed during growth arrest and DNA damage. Proc Natl Acad Sci USA. 1994;91:5247–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choukhi A, Ung S, Wychowski Cet al. . Involvement of endoplasmic reticulum chaperones in the folding of hepatitis C virus glycoproteins. J Virol. 1998;72:3851–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chusri P, Kumthip K, Hong Jet al. . HCV induces transforming growth factor beta1 through activation of endoplasmic reticulum stress and the unfolded protein response. Sci Rep. 2016;6:22487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colomer-Lluch M, Ruiz A, Moris Aet al. . Restriction Factors: from Intrinsic Viral Restriction to Shaping Cellular Immunity Against HIV-1. Front Immunol. 2018;9:2876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connor JH, Lyles DS. Inhibition of host and viral translation during vesicular stomatitis virus infection. eIF2 is responsible for the inhibition of viral but not host translation. J Biol Chem. 2005;280:13512–9. [DOI] [PubMed] [Google Scholar]
- Cox JS, Shamu CE, Walter P. Transcriptional induction of genes encoding endoplasmic reticulum resident proteins requires a transmembrane protein kinase. Cell. 1993;73:1197–206. [DOI] [PubMed] [Google Scholar]
- Cross BC, Bond PJ, Sadowski PGet al. . The molecular basis for selective inhibition of unconventional mRNA splicing by an IRE1-binding small molecule. Proc Natl Acad Sci USA. 2012;109:E869–878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curcio MJ, Lutz S, Lesage P. The Ty1 LTR-retrotransposon of budding yeast, Saccharomyces cerevisiae. Microbiol Spectr. 2015;3:1–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Datan E, Roy SG, Germain Get al. . Dengue-induced autophagy, virus replication and protection from cell death require ER stress (PERK) pathway activation. Cell Death Dis. 2016;7:e2127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng J, Lu PD, Zhang Yet al. . Translational repression mediates activation of nuclear factor kappa B by phosphorylated translation initiation factor 2. Mol Cell Biol. 2004;24:10161–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drahos DJ, Hendrix RW. Effect of bacteriophage lambda infection on synthesis of groE protein and other Escherichia coli proteins. J Bacteriol. 1982;149:1050–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellgaard L, Helenius A. Quality control in the endoplasmic reticulum. Nat Rev Mol Cell Biol. 2003;4:181–91. [DOI] [PubMed] [Google Scholar]
- Fejer G, Drechsel L, Liese Jet al. . Key role of splenic myeloid DCs in the IFN-alphabeta response to adenoviruses in vivo. PLoS Pathog. 2008;4:e1000208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frabutt DA, Zheng YH. Arms Race between Enveloped Viruses and the Host ERAD Machinery. Viruses. 2016;8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fung TS, Liao Y, Liu DX. The endoplasmic reticulum stress sensor IRE1alpha protects cells from apoptosis induced by the coronavirus infectious bronchitis virus. J Virol. 2014;88:12752–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fung TS, Liu DX. Human Coronavirus: host-Pathogen Interaction. Annu Rev Microbiol. 2019;73:529–57. [DOI] [PubMed] [Google Scholar]
- Futerman AH, Riezman H. The ins and outs of sphingolipid synthesis. Trends Cell Biol. 2005;15:312–8. [DOI] [PubMed] [Google Scholar]
- Galindo I, Hernaez B, Munoz-Moreno Ret al. . The ATF6 branch of unfolded protein response and apoptosis are activated to promote African swine fever virus infection. Cell Death Dis. 2012;3:e341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao J, Mese K, Bunz Oet al. . State-of-the-art human adenovirus vectorology for therapeutic approaches. FEBS Lett. 2019;593:3609–22. [DOI] [PubMed] [Google Scholar]
- Gao P, Chai Y, Song Jet al. . Reprogramming the unfolded protein response for replication by porcine reproductive and respiratory syndrome virus. PLoS Pathog. 2019;15:e1008169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner BM, Walter P. Unfolded proteins are Ire1-activating ligands that directly induce the unfolded protein response. Science. 2011;333:1891–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Georgi F, Greber UF. The Adenovirus Death Protein - A small membrane protein controls cell lysis and disease. FEBS Lett. 2020. [DOI] [PubMed] [Google Scholar]
- Gong G, Waris G, Tanveer Ret al. . Human hepatitis C virus NS5A protein alters intracellular calcium levels, induces oxidative stress, and activates STAT-3 and NF-kappa B. Proc Natl Acad Sci USA. 2001;98:9599–604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greber UF, Flatt JW. Adenovirus Entry: from Infection to Immunity. Annu Rev Virol. 2019;6:177–97. [DOI] [PubMed] [Google Scholar]
- Greber UF. Editorial: physical Virology and the Nature of Virus Infections. Physical Virology Advances in Experimental Medicine and Biology.Vol. 1215, Greber UF, Cham, Switzerland: (ed). Springer: 2019, 1–11. [DOI] [PubMed] [Google Scholar]
- Greber UF. Virus and Host Mechanics Support Membrane Penetration and Cell Entry. J Virol. 2016;90:3802–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grootjans J, Kaser A, Kaufman RJet al. . The unfolded protein response in immunity and inflammation. Nat Rev Immunol. 2016;16:469–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gulbahce N, Yan H, Dricot Aet al. . Viral perturbations of host networks reflect disease etiology. PLoS Comput Biol. 2012;8:e1002531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haller O, Kochs G, Weber F. The interferon response circuit: induction and suppression by pathogenic viruses. Virology. 2006;344:119–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond C, Helenius A. Quality control in the secretory pathway. Curr Opin Cell Biol. 1995;7:523–9. [DOI] [PubMed] [Google Scholar]
- Harada JN, Shevchenko A, Shevchenko Aet al. . Analysis of the adenovirus E1B-55K-anchored proteome reveals its link to ubiquitination machinery. J Virol. 2002;76:9194–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harayama T, Riezman H. Understanding the diversity of membrane lipid composition. Nat Rev Mol Cell Biol. 2018;19:281–96. [DOI] [PubMed] [Google Scholar]
- Harding HP, Novoa I, Zhang Yet al. . Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol Cell. 2000;6:1099–108. [DOI] [PubMed] [Google Scholar]
- Harding HP, Zhang Y, Ron D. Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature. 1999;397:271–4. [DOI] [PubMed] [Google Scholar]
- Harding HP, Zhang Y, Zeng Het al. . An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell. 2003;11:619–33. [DOI] [PubMed] [Google Scholar]
- Hassan I, Gaines KS, Hottel WJet al. . Inositol-requiring enzyme 1 inhibits respiratory syncytial virus replication. J Biol Chem. 2014;289:7537–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hassan IH, Zhang MS, Powers LSet al. . Influenza A viral replication is blocked by inhibition of the inositol-requiring enzyme 1 (IRE1) stress pathway. J Biol Chem. 2012;287:4679–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes CM, Ron D. The mitochondrial UPR - protecting organelle protein homeostasis. J Cell Sci. 2010;123:3849–55. [DOI] [PubMed] [Google Scholar]
- Haze K, Yoshida H, Yanagi Het al. . Mammalian transcription factor ATF6 is synthesized as a transmembrane protein and activated by proteolysis in response to endoplasmic reticulum stress. Mol Biol Cell. 1999;10:3787–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heaton NS, Randall G. Multifaceted roles for lipids in viral infection. Trends Microbiol. 2011;19:368–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He B, Chou J, Brandimarti Ret al. . Suppression of the phenotype of gamma(1)34.5- herpes simplex virus 1: failure of activated RNA-dependent protein kinase to shut off protein synthesis is associated with a deletion in the domain of the alpha47 gene. J Virol. 1997;71:6049–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helenius A, Aebi M. Intracellular functions of N-linked glycans. Science. 2001;291:2364–9. [DOI] [PubMed] [Google Scholar]
- Hetz C, Bernasconi P, Fisher Jet al. . Proapoptotic BAX and BAK modulate the unfolded protein response by a direct interaction with IRE1alpha. Science. 2006;312:572–6. [DOI] [PubMed] [Google Scholar]
- Higgins R, Gendron JM, Rising Let al. . The Unfolded Protein Response Triggers Site-Specific Regulatory Ubiquitylation of 40S Ribosomal Proteins. Mol Cell. 2015;59:35–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinte F, van Anken E, Tirosh Bet al. . Repression of viral gene expression and replication by the unfolded protein response effector XBP1u. eLife. 2020;9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hitomi J, Katayama T, Eguchi Yet al. . Involvement of caspase-4 in endoplasmic reticulum stress-induced apoptosis and Abeta-induced cell death. J Cell Biol. 2004;165:347–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann HH, Schneider WM, Rice CM. Interferons and viruses: an evolutionary arms race of molecular interactions. Trends Immunol. 2015;36:124–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hollien J, Lin JH, Li Het al. . Regulated Ire1-dependent decay of messenger RNAs in mammalian cells. J Cell Biol. 2009;186:323–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hollien J, Weissman JS. Decay of endoplasmic reticulum-localized mRNAs during the unfolded protein response. Science. 2006;313:104–7. [DOI] [PubMed] [Google Scholar]
- Holt PG, Strickland DH, Wikstrom MEet al. . Regulation of immunological homeostasis in the respiratory tract. Nat Rev Immunol. 2008;8:142–52. [DOI] [PubMed] [Google Scholar]
- Houck SA, Ren HY, Madden VJet al. . Quality control autophagy degrades soluble ERAD-resistant conformers of the misfolded membrane protein GnRHR. Mol Cell. 2014;54:166–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu SK, Chiu CC, Dahms HUet al. . Unfolded Protein Response (UPR) in Survival, Dormancy, Immunosuppression, Metastasis, and Treatments of Cancer Cells. Int J Mol Sci. 2019;20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang H, Kang R, Wang Jet al. . Hepatitis C virus inhibits AKT-tuberous sclerosis complex (TSC), the mechanistic target of rapamycin (MTOR) pathway, through endoplasmic reticulum stress to induce autophagy. Autophagy. 2013;9:175–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu P, Han Z, Couvillon ADet al. . Autocrine tumor necrosis factor alpha links endoplasmic reticulum stress to the membrane death receptor pathway through IRE1alpha-mediated NF-kappaB activation and down-regulation of TRAF2 expression. Mol Cell Biol. 2006;26:3071–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hur S. Double-Stranded RNA Sensors and Modulators in Innate Immunity. Annu Rev Immunol. 2019;37:349–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurtley SM, Bole DG, Hoover-Litty Het al. . Interactions of misfolded influenza virus hemagglutinin with binding protein (BiP). J Cell Biol. 1989;108:2117–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishikawa T, Toyama T, Nakamura Yet al. . UPR transducer BBF2H7 allows export of type II collagen in a cargo- and developmental stage-specific manner. J Cell Biol. 2017;216:1761–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwakoshi NN, Lee AH, Glimcher LH. The X-box binding protein-1 transcription factor is required for plasma cell differentiation and the unfolded protein response. Immunol Rev. 2003;194:29–38. [DOI] [PubMed] [Google Scholar]
- Johnston BP, McCormick C. Herpesviruses and the Unfolded Protein Response. Viruses. 2019;12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan R, Wang L, Graczyk TMet al. . Replication of a cytopathic strain of bovine viral diarrhea virus activates PERK and induces endoplasmic reticulum stress-mediated apoptosis of MDBK cells. J Virol. 2002;76:9588–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jost S, Altfeld M. Control of human viral infections by natural killer cells. Annu Rev Immunol. 2013;31:163–94. [DOI] [PubMed] [Google Scholar]
- Jurkin J, Henkel T, Nielsen AFet al. . The mammalian tRNA ligase complex mediates splicing of XBP1 mRNA and controls antibody secretion in plasma cells. EMBO J. 2014;33:2922–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karagoz GE, Acosta-Alvear D, Nguyen HTet al. . An unfolded protein-induced conformational switch activates mammalian IRE1. eLife. 2017;6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kati S, Tsao EH, Gunther Tet al. . Activation of the B cell antigen receptor triggers reactivation of latent Kaposi's sarcoma-associated herpesvirus in B cells. J Virol. 2013;87:8004–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ke PY, Chen SS. Activation of the unfolded protein response and autophagy after hepatitis C virus infection suppresses innate antiviral immunity in vitro. J Clin Invest. 2011;121:37–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khomich OA, Kochetkov SN, Bartosch Bet al. . Redox Biology of Respiratory Viral Infections. Viruses. 2018;10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimata Y, Ishiwata-Kimata Y, Yamada Set al. . Yeast unfolded protein response pathway regulates expression of genes for anti-oxidative stress and for cell surface proteins. Genes Cells. 2006;11:59–69. [DOI] [PubMed] [Google Scholar]
- Kimata Y, Oikawa D, Shimizu Yet al. . A role for BiP as an adjustor for the endoplasmic reticulum stress-sensing protein Ire1. J Cell Biol. 2004;167:445–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S, Joe Y, Kim HJet al. . Endoplasmic reticulum stress-induced IRE1alpha activation mediates cross-talk of GSK-3beta and XBP-1 to regulate inflammatory cytokine production. J Immunol. 2015;194:4498–506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kono N, Amin-Wetzel N, Ron D. Generic membrane-spanning features endow IRE1alpha with responsiveness to membrane aberrancy. Mol Biol Cell. 2017;28:2318–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishnamoorthy J, Mounir Z, Raven JFet al. . The eIF2alpha kinases inhibit vesicular stomatitis virus replication independently of eIF2alpha phosphorylation. Cell Cycle. 2008;7:2346–51. [DOI] [PubMed] [Google Scholar]
- Kudchodkar SB, Levine B. Viruses and autophagy. Rev Med Virol. 2009;19:359–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kusaczuk M. Tauroursodeoxycholate-Bile Acid with Chaperoning Activity: molecular and Cellular Effects and Therapeutic Perspectives. Cells. 2019;8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamers MM, van den Hoogen BG, Haagmans BL. ADAR1: “Editor-in-Chief” of Cytoplasmic Innate Immunity. Front Immunol. 2019;10:1763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee ES, Yoon CH, Kim YSet al. . The double-strand RNA-dependent protein kinase PKR plays a significant role in a sustained ER stress-induced apoptosis. FEBS Lett. 2007;581:4325–32. [DOI] [PubMed] [Google Scholar]
- Lee YR, Kuo SH, Lin CYet al. . Dengue virus-induced ER stress is required for autophagy activation, viral replication, and pathogenesis both in vitro and in vivo. Sci Rep. 2018;8:489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemos de Matos A, Franco LS, McFadden G. Oncolytic Viruses and the Immune System: the Dynamic Duo. Mol Ther Methods Clin Dev. 2020;17:349–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lencer WI, DeLuca H, Grey MJet al. . Innate immunity at mucosal surfaces: the IRE1-RIDD-RIG-I pathway. Trends Immunol. 2015;36:401–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liberman E, Fong YL, Selby MJet al. . Activation of the grp78 and grp94 promoters by hepatitis C virus E2 envelope protein. J Virol. 1999;73:3718–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lichtenstein DL, Toth K, Doronin Ket al. . Functions and mechanisms of action of the adenovirus E3 proteins. Int Rev Immunol. 2004;23:75–111. [DOI] [PubMed] [Google Scholar]
- Li H, Korennykh AV, Behrman SLet al. . Mammalian endoplasmic reticulum stress sensor IRE1 signals by dynamic clustering. Proc Natl Acad Sci U S A. 2010;107:16113–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li K, Hu C, Xing Fet al. . Deficiency of the IRE1alpha-Autophagy Axis Enhances the Antitumor Effects of the Oncolytic Virus M1. J Virol. 2018;92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin JH, Li H, Yasumura Det al. . IRE1 signaling affects cell fate during the unfolded protein response. Science. 2007;318:944–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Ye L, Yu Xet al. . Hepatitis C virus NS4B induces unfolded protein response and endoplasmic reticulum overload response-dependent NF-kappaB activation. Virology. 2009;391:257–64. [DOI] [PubMed] [Google Scholar]
- Liu CJ, Chuang WL, Lee CMet al. . Peginterferon alfa-2a plus ribavirin for the treatment of dual chronic infection with hepatitis B and C viruses. Gastroenterology. 2009;136:496–504 e493. [DOI] [PubMed] [Google Scholar]
- Liu J, HuangFu WC, Kumar KGet al. . Virus-induced unfolded protein response attenuates antiviral defenses via phosphorylation-dependent degradation of the type I interferon receptor. Cell Host Microbe. 2009;5:72–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu YP, Zeng L, Tian Aet al. . Endoplasmic reticulum stress regulates the innate immunity critical transcription factor IRF3. J Immunol. 2012;189:4630–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu ZG, Hsu H, Goeddel DVet al. . Dissection of TNF receptor 1 effector functions: JNK activation is not linked to apoptosis while NF-kappaB activation prevents cell death. Cell. 1996;87:565–76. [DOI] [PubMed] [Google Scholar]
- Lotzerich M, Roulin PS, Boucke Ket al. . Rhinovirus 3C protease suppresses apoptosis and triggers caspase-independent cell death. Cell Death Dis. 2018;9:272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu PD, Harding HP, Ron D. Translation reinitiation at alternative open reading frames regulates gene expression in an integrated stress response. J Cell Biol. 2004;167:27–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machamer CE, Doms RW, Bole DGet al. . Heavy chain binding protein recognizes incompletely disulfide-bonded forms of vesicular stomatitis virus G protein. J Biol Chem. 1990;265:6879–83. [PubMed] [Google Scholar]
- Magadan JG, Perez-Victoria FJ, Sougrat Ret al. . Multilayered mechanism of CD4 downregulation by HIV-1 Vpu involving distinct ER retention and ERAD targeting steps. PLoS Pathog. 2010;6:e1000869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahoney DJ, Lefebvre C, Allan Ket al. . Virus-tumor interactome screen reveals ER stress response can reprogram resistant cancers for oncolytic virus-triggered caspase-2 cell death. Cancer Cell. 2011;20:443–56. [DOI] [PubMed] [Google Scholar]
- Malay AD, Miyazaki N, Biela Aet al. . An ultra-stable gold-coordinated protein cage displaying reversible assembly. Nature. 2019;569:438–42. [DOI] [PubMed] [Google Scholar]
- Maler MD, Nielsen PJ, Stichling Net al. . Key Role of the Scavenger Receptor MARCO in Mediating Adenovirus Infection and Subsequent Innate Responses of Macrophages. mBio. 2017;8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Margariti A, Li H, Chen Tet al. . XBP1 mRNA splicing triggers an autophagic response in endothelial cells through BECLIN-1 transcriptional activation. J Biol Chem. 2013;288:859–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Margottin F, Bour SP, Durand Het al. . A novel human WD protein, h-beta TrCp, that interacts with HIV-1 Vpu connects CD4 to the ER degradation pathway through an F-box motif. Mol Cell. 1998;1:565–74. [DOI] [PubMed] [Google Scholar]
- Marquez RT, Xu L. Bcl-2:beclin 1 complex: multiple, mechanisms regulating autophagy/apoptosis toggle switch. Am J Cancer Res. 2012;2:214–21. [PMC free article] [PubMed] [Google Scholar]
- Marshall JS, Portales-Cervantes L, Leong E. Mast Cell Responses to Viruses and Pathogen Products. Int J Mol Sci. 2019;20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinon F, Chen X, Lee AHet al. . TLR activation of the transcription factor XBP1 regulates innate immune responses in macrophages. Nat Immunol. 2010;11:411–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathews MB, Shenk T. Adenovirus virus-associated RNA and translation control. J Virol. 1991;65:5657–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maxfield FR, van Meer G. Cholesterol, the central lipid of mammalian cells. Curr Opin Cell Biol. 2010;22:422–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma Y, Brewer JW, Diehl JAet al. . Two distinct stress signaling pathways converge upon the CHOP promoter during the mammalian unfolded protein response. J Mol Biol. 2002;318:1351–65. [DOI] [PubMed] [Google Scholar]
- McCullough KD, Martindale JL, Klotz LOet al. . Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Mol Cell Biol. 2001;21:1249–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metcalf MG, Higuchi-Sanabria R, Garcia Get al. . Beyond the cell factory: homeostatic regulation of and by the UPR(ER). Sci Adv. 2020;6:eabb9614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mori K, Ma W, Gething MJet al. . A transmembrane protein with a cdc2+/CDC28-related kinase activity is required for signaling from the ER to the nucleus. Cell. 1993;74:743–56. [DOI] [PubMed] [Google Scholar]
- Mori K, Ogawa N, Kawahara Tet al. . mRNA splicing-mediated C-terminal replacement of transcription factor Hac1p is required for efficient activation of the unfolded protein response. Proc Natl Acad Sci USA. 2000;97:4660–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mrozek-Gorska P, Buschle A, Pich Det al. . Epstein-Barr virus reprograms human B lymphocytes immediately in the prelatent phase of infection. Proc Natl Acad Sci USA. 2019;116:16046–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulvey M, Arias C, Mohr I. Maintenance of endoplasmic reticulum (ER) homeostasis in herpes simplex virus type 1-infected cells through the association of a viral glycoprotein with PERK, a cellular ER stress sensor. J Virol. 2007;81:3377–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulvey M, Poppers J, Sternberg Det al. . Regulation of eIF2alpha phosphorylation by different functions that act during discrete phases in the herpes simplex virus type 1 life cycle. J Virol. 2003;77:10917–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakagawa T, Yuan J. Cross-talk between two cysteine protease families. Activation of caspase-12 by calpain in apoptosis. J Cell Biol. 2000;150:887–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakagawa T, Zhu H, Morishima Net al. . Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-beta. Nature. 2000;403:98–103. [DOI] [PubMed] [Google Scholar]
- Napetschnig J, Wu H. Molecular basis of NF-kappaB signaling. Annu Rev Biophys. 2013;42:443–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Netherton CL, Parsley JC, Wileman T. African swine fever virus inhibits induction of the stress-induced proapoptotic transcription factor CHOP/GADD153. J Virol. 2004;78:10825–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng DT, Randall RE, Lamb RA. Intracellular maturation and transport of the SV5 type II glycoprotein hemagglutinin-neuraminidase: specific and transient association with GRP78-BiP in the endoplasmic reticulum and extensive internalization from the cell surface. J Cell Biol. 1989;109:3273–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Connell RM, Rao DS, Chaudhuri AAet al. . Physiological and pathological roles for microRNAs in the immune system. Nat Rev Immunol. 2010;10:111–22. [DOI] [PubMed] [Google Scholar]
- Pahl HL, Sester M, Burgert HGet al. . Activation of transcription factor NF-kappaB by the adenovirus E3/19K protein requires its ER retention. J Cell Biol. 1996;132:511–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasquarella A, Ebert A, Pereira de Almeida Get al. . Retrotransposon derepression leads to activation of the unfolded protein response and apoptosis in pro-B cells. Development. 2016;143:1788–99. [DOI] [PubMed] [Google Scholar]
- Paul P, Munz C. Autophagy and Mammalian Viruses: roles in Immune Response, Viral Replication, and Beyond. Adv Virus Res. 2016;95:149–95. [DOI] [PubMed] [Google Scholar]
- Perera N, Miller JL, Zitzmann N. The role of the unfolded protein response in dengue virus pathogenesis. Cell Microbiol. 2017;19. [DOI] [PubMed] [Google Scholar]
- Pietrocola F, Izzo V, Niso-Santano Met al. . Regulation of autophagy by stress-responsive transcription factors. Semin Cancer Biol. 2013;23:310–22. [DOI] [PubMed] [Google Scholar]
- Prasad V, Suomalainen M, Jasiqi Yet al. . The UPR sensor IRE1alpha and the adenovirus E3-19K glycoprotein sustain persistent and lytic infections. Nat Commun. 2020;11:1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasad V, Suomalainen M, Pennauer Met al. . Chemical Induction of Unfolded Protein Response Enhances Cancer Cell Killing through Lytic Virus Infection. J Virol. 2014;88:13086–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preissler S, Ron D. Early Events in the Endoplasmic Reticulum Unfolded Protein Response. Cold Spring Harb Perspect Biol. 2019;11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pulendran B, Li S, Nakaya HI. Systems vaccinology. Immunity. 2010;33:516–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puthalakath H, O'Reilly LA, Gunn Pet al. . ER stress triggers apoptosis by activating BH3-only protein Bim. Cell. 2007;129:1337–49. [DOI] [PubMed] [Google Scholar]
- Ranjitha HB, Ammanathan V, Guleria Net al. . Foot-and-mouth disease virus induces PERK-mediated autophagy to suppress the antiviral interferon response. J Cell Sci. 2020;134. [DOI] [PubMed] [Google Scholar]
- Ranu RS, London IM. Regulation of protein synthesis in rabbit reticulocyte lysates: additional initiation factor required for formation of ternary complex (eIF-2.GTP.Met-tRNAf) and demonstration of inhibitory effect of heme-regulated protein kinase. Proc Natl Acad Sci USA. 1979;76:1079–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao RV, Castro-Obregon S, Frankowski Het al. . Coupling endoplasmic reticulum stress to the cell death program. An Apaf-1-independent intrinsic pathway. J Biol Chem. 2002;277:21836–42. [DOI] [PubMed] [Google Scholar]
- Reimold AM, Iwakoshi NN, Manis Jet al. . Plasma cell differentiation requires the transcription factor XBP-1. Nature. 2001;412:300–7. [DOI] [PubMed] [Google Scholar]
- Rolland SG, Schneid S, Schwarz Met al. . Compromised Mitochondrial Protein Import Acts as a Signal for UPR(mt). Cell Rep. 2019;28:1659–1669 e1655. [DOI] [PubMed] [Google Scholar]
- Romero-Brey I, Bartenschlager R. Endoplasmic Reticulum: the Favorite Intracellular Niche for Viral Replication and Assembly. Viruses. 2016;8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol. 2007;8:519–29. [DOI] [PubMed] [Google Scholar]
- Rosati E, Sabatini R, Rampino Get al. . Novel targets for endoplasmic reticulum stress-induced apoptosis in B-CLL. Blood. 2010;116:2713–23. [DOI] [PubMed] [Google Scholar]
- Roulin PS, Lotzerich M, Torta Fet al. . Rhinovirus Uses a Phosphatidylinositol 4-Phosphate/Cholesterol Counter-Current for the Formation of Replication Compartments at the ER-Golgi Interface. Cell Host Microbe. 2014;16:677–90. [DOI] [PubMed] [Google Scholar]
- Roulois D, Loo Yau H, Singhania Ret al. . DNA-Demethylating Agents Target Colorectal Cancer Cells by Inducing Viral Mimicry by Endogenous Transcripts. Cell. 2015;162:961–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roulstone V, Pedersen M, Kyula Jet al. . BRAF- and MEK-Targeted Small Molecule Inhibitors Exert Enhanced Antimelanoma Effects in Combination With Oncolytic Reovirus Through ER Stress. Mol Ther. 2015;23:931–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rouse BT, Sehrawat S. Immunity and immunopathology to viruses: what decides the outcome?. Nat Rev Immunol. 2010;10:514–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rozenblatt-Rosen O, Deo RC, Padi Met al. . Interpreting cancer genomes using systematic host network perturbations by tumour virus proteins. Nature. 2012;487:491–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruegsegger U, Leber JH, Walter P. Block of HAC1 mRNA translation by long-range base pairing is released by cytoplasmic splicing upon induction of the unfolded protein response. Cell. 2001;107:103–14. [DOI] [PubMed] [Google Scholar]
- Russell SJ, Peng KW, Bell JC. Oncolytic virotherapy. Nat Biotechnol. 2012;30:658–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez EL, Lagunoff M. Viral activation of cellular metabolism. Virology. 2015;479–480:609–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schindler AJ, Schekman R. In vitro reconstitution of ER-stress induced ATF6 transport in COPII vesicles. Proc Natl Acad Sci USA. 2009;106:17775–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt ME, Varga SM. The CD8 T Cell Response to Respiratory Virus Infections. Front Immunol. 2018;9:678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarz KB. Oxidative stress during viral infection: a review. Free Radic Biol Med. 1996;21:641–9. [DOI] [PubMed] [Google Scholar]
- Sekine Y, Zyryanova A, Crespillo-Casado Aet al. . Stress responses. Mutations in a translation initiation factor identify the target of a memory-enhancing compound. Science. 2015;348:1027–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma M, Bhattacharyya S, Sharma KBet al. . Japanese encephalitis virus activates autophagy through XBP1 and ATF6 ER stress sensors in neuronal cells. J Gen Virol. 2017;98:1027–39. [DOI] [PubMed] [Google Scholar]
- Shirley CM, Chen J, Shamay Met al. . Bortezomib induction of C/EBPbeta mediates Epstein-Barr virus lytic activation in Burkitt lymphoma. Blood. 2011;117:6297–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidrauski C, McGeachy AM, Ingolia NTet al. . The small molecule ISRIB reverses the effects of eIF2alpha phosphorylation on translation and stress granule assembly. eLife. 2015;4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson GR, Relph K, Harrington Ket al. . Cancer immunotherapy via combining oncolytic virotherapy with chemotherapy: recent advances. Oncolytic Virother. 2016;5:1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JA. A new paradigm: innate immune sensing of viruses via the unfolded protein response. Frontiers in microbiology. 2014;5:222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith MH, Ploegh HL, Weissman JS. Road to ruin: targeting proteins for degradation in the endoplasmic reticulum. Science. 2011;334:1086–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- So JS. Roles of Endoplasmic Reticulum Stress in Immune Responses. Mol Cells. 2018;41:705–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sotero-Caio CG, Platt RN 2nd, Suh Aet al. . Evolution and Diversity of Transposable Elements in Vertebrate Genomes. Genome Biol Evol. 2017;9:161–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spurgeon ME, Ornelles DA. The adenovirus E1B 55-kilodalton and E4 open reading frame 6 proteins limit phosphorylation of eIF2alpha during the late phase of infection. J Virol. 2009;83:9970–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sriburi R, Jackowski S, Mori Ket al. . XBP1: a link between the unfolded protein response, lipid biosynthesis, and biogenesis of the endoplasmic reticulum. J Cell Biol. 2004;167:35–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stahl S, Burkhart JM, Hinte Fet al. . Cytomegalovirus downregulates IRE1 to repress the unfolded protein response. PLoS Pathog. 2013;9:e1003544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stichling N, Suomalainen M, Flatt JWet al. . Lung macrophage scavenger receptor SR-A6 (MARCO) is an adenovirus type-specific virus entry receptor. PLoS Pathog. 2018;14:e1006914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strebel K. HIV-1 Vpu - an ion channel in search of a job. Biochim Biophys Acta. 2014;1838:1074–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stroberg W, Eilertsen J, Schnell S. Information processing by endoplasmic reticulum stress sensors. J R Soc Interface. 2019;16:20190288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su C, Zhan G, Zheng C. Evasion of host antiviral innate immunity by HSV-1, an update. Virol J. 2016;13:38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su HL, Liao CL, Lin YL. Japanese encephalitis virus infection initiates endoplasmic reticulum stress and an unfolded protein response. J Virol. 2002;76:4162–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun CC, Thorley-Lawson DA. Plasma cell-specific transcription factor XBP-1s binds to and transactivates the Epstein-Barr virus BZLF1 promoter. J Virol. 2007;81:13566–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sung SC, Chao CY, Jeng KSet al. . The 8ab protein of SARS-CoV is a luminal ER membrane-associated protein and induces the activation of ATF6. Virology. 2009;387:402–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun M, Rothermel TA, Shuman Let al. . Conserved cysteine-rich domain of paramyxovirus simian virus 5 V protein plays an important role in blocking apoptosis. J Virol. 2004;78:5068–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeda K, Akira S. Toll-like receptors in innate immunity. Int Immunol. 2005;17:1–14. [DOI] [PubMed] [Google Scholar]
- Takeuchi O, Akira S. Innate immunity to virus infection. Immunol Rev. 2009;227:75–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tardif KD, Mori K, Kaufman RJet al. . Hepatitis C virus suppresses the IRE1-XBP1 pathway of the unfolded protein response. J Biol Chem. 2004;279:17158–64. [DOI] [PubMed] [Google Scholar]
- Taylor DR, Shi ST, Romano PRet al. . Inhibition of the interferon-inducible protein kinase PKR by HCV E2 protein. Science. 1999;285:107–10. [DOI] [PubMed] [Google Scholar]
- Tetter S, Terasaka N, Steinauer Aet al. . Evolution of a virus-like architecture and packaging mechanism in a repurposed bacterial protein. bioRxiv. 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thaker SK, Ch'ng J, Christofk HR. Viral hijacking of cellular metabolism. BMC Biol. 2019;17:59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tirosh B, Iwakoshi NN, Lilley BNet al. . Human cytomegalovirus protein US11 provokes an unfolded protein response that may facilitate the degradation of class I major histocompatibility complex products. J Virol. 2005;79:2768–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Travers KJ, Patil CK, Wodicka Let al. . Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell. 2000;101:249–58. [DOI] [PubMed] [Google Scholar]
- Tsai JC, Miller-Vedam LE, Anand AAet al. . Structure of the nucleotide exchange factor eIF2B reveals mechanism of memory-enhancing molecule. Science. 2018;359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuru A, Fujimoto N, Takahashi Set al. . Negative feedback by IRE1beta optimizes mucin production in goblet cells. Proc Natl Acad Sci U S A. 2013;110:2864–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umareddy I, Pluquet O, Wang QYet al. . Dengue virus serotype infection specifies the activation of the unfolded protein response. Virol J. 2007;4:91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urano F, Wang X, Bertolotti Aet al. . Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science. 2000;287:664–6. [DOI] [PubMed] [Google Scholar]
- van der Wal FJ, Kikkert M, Wiertz E. The HCMV gene products US2 and US11 target MHC class I molecules for degradation in the cytosol. Curr Top Microbiol Immunol. 2002;269:37–55. [DOI] [PubMed] [Google Scholar]
- Vattem KM, Wek RC. Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc Natl Acad Sci USA. 2004;101:11269–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Virgin HW, Wherry EJ, Ahmed R. Redefining chronic viral infection. Cell. 2009;138:30–50. [DOI] [PubMed] [Google Scholar]
- Volmer R, van der Ploeg K, Ron D. Membrane lipid saturation activates endoplasmic reticulum unfolded protein response transducers through their transmembrane domains. Proc Natl Acad Sci USA. 2013;110:4628–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation. Science. 2011;334:1081–6. [DOI] [PubMed] [Google Scholar]
- Wang R, Moniruzzaman M, Shuffle Eet al. . Immune regulation of the unfolded protein response at the mucosal barrier in viral infection. Clin Transl Immunology. 2018;7:e1014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watowich SS, Morimoto RI, Lamb RA. Flux of the paramyxovirus hemagglutinin-neuraminidase glycoprotein through the endoplasmic reticulum activates transcription of the GRP78-BiP gene. J Virol. 1991;65:3590–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weitzman MD, Fradet-Turcotte A. Virus DNA Replication and the Host DNA Damage Response. Annu Rev Virol. 2018;5:141–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei Y, Wang D, Topczewski Fet al. . Saturated fatty acids induce endoplasmic reticulum stress and apoptosis independently of ceramide in liver cells. Am J Physiol Endocrinol Metab. 2006;291:E275–281. [DOI] [PubMed] [Google Scholar]
- Williams JL, Garcia J, Harrich Det al. . Lymphoid specific gene expression of the adenovirus early region 3 promoter is mediated by NF-kappa B binding motifs. EMBO J. 1990;9:4435–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfrum N, Greber UF. Adenovirus signalling in entry. Cell Microbiol. 2013;15:53–62. [DOI] [PubMed] [Google Scholar]
- Xu C, Bailly-Maitre B, Reed JC. Endoplasmic reticulum stress: cell life and death decisions. J Clin Invest. 2005;115:2656–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamauchi Y, Greber UF. Principles of Virus Uncoating: cues and the Snooker Ball. Traffic. 2016;17:569–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye J, Rawson RB, Komuro Ret al. . ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Mol Cell. 2000;6:1355–64. [DOI] [PubMed] [Google Scholar]
- Yoneda T, Imaizumi K, Oono Ket al. . Activation of caspase-12, an endoplastic reticulum (ER) resident caspase, through tumor necrosis factor receptor-associated factor 2-dependent mechanism in response to the ER stress. J Biol Chem. 2001;276:13935–40. [DOI] [PubMed] [Google Scholar]
- Yoo JY, Hurwitz BS, Bolyard Cet al. . Bortezomib-induced unfolded protein response increases oncolytic HSV-1 replication resulting in synergistic antitumor effects. Clin Cancer Res. 2014;20:3787–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida H, Okada T, Haze Ket al. . ATF6 activated by proteolysis binds in the presence of NF-Y (CBF) directly to the cis-acting element responsible for the mammalian unfolded protein response. Mol Cell Biol. 2000;20:6755–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young RA. Stress proteins and immunology. Annu Rev Immunol. 1990;8:401–20. [DOI] [PubMed] [Google Scholar]
- Yu CY, Hsu YW, Liao CLet al. . Flavivirus infection activates the XBP1 pathway of the unfolded protein response to cope with endoplasmic reticulum stress. J Virol. 2006;80:11868–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu L, Chen Y, Tooze SA. Autophagy pathway: cellular and molecular mechanisms. Autophagy. 2018;14:207–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang F, Moon A, Childs Ket al. . The African swine fever virus DP71L protein recruits the protein phosphatase 1 catalytic subunit to dephosphorylate eIF2alpha and inhibits CHOP induction but is dispensable for these activities during virus infection. J Virol. 2010;84:10681–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang HM, Ye X, Su Yet al. . Coxsackievirus B3 infection activates the unfolded protein response and induces apoptosis through downregulation of p58IPK and activation of CHOP and SREBP1. J Virol. 2010;84:8446–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang P, Su C, Jiang Zet al. . Herpes Simplex Virus 1 UL41 Protein Suppresses the IRE1/XBP1 Signal Pathway of the Unfolded Protein Response via Its RNase Activity. J Virol. 2017;91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou J, Liu CY, Back SHet al. . The crystal structure of human IRE1 luminal domain reveals a conserved dimerization interface required for activation of the unfolded protein response. Proc Natl Acad Sci USA. 2006;103:14343–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu G, Zheng Y, Zhang Let al. . Coxsackievirus A16 infection triggers apoptosis in RD cells by inducing ER stress. Biochem Biophys Res Commun. 2013;441:856–61. [DOI] [PubMed] [Google Scholar]
- Zyryanova AF, Weis F, Faille Aet al. . Binding of ISRIB reveals a regulatory site in the nucleotide exchange factor eIF2B. Science. 2018;359:1533–6. [DOI] [PMC free article] [PubMed] [Google Scholar]