

Abstract
Dormancy release and reactivation of temperate-zone trees involve the temperature-modulated expression of cell-cycle genes. However, information on the detailed regulatory mechanism is limited. Here, we compared the transcriptomes of the stems of active and dormant larch trees, emphasizing the expression patterns of cell-cycle genes and transcription factors and assessed their relationships and responses to temperatures. Twelve cell-cycle genes and 31 transcription factors were strongly expressed in the active stage. Promoter analysis suggested that these 12 genes might be regulated by transcription factors from 10 families. Altogether, 73 cases of regulation between 16 transcription factors and 12 cell-cycle genes were predicted, while the regulatory interactions between LaMYB20 and LaCYCB1;1, and LaRAV1 and LaCDKB1;3 were confirmed by yeast one-hybrid and dual-luciferase assays. Last, we found that LaRAV1 and LaCDKB1;3 had almost the same expression patterns during dormancy release and reactivation induced naturally or artificially by temperature, indicating that the LaRAV1-LaCDKB1;3 module functions in the temperature-modulated dormancy release and reactivation of larch trees. These results provide new insights into the link between temperature and cell-cycle gene expression, helping to understand the temperature control of tree growth and development in the context of climate change.
Keywords: cambium, cell-cycle gene, dormancy release, larch, reactivation, temperature, transcription factor
Introduction
Dormancy release and reactivation of temperate-zone trees are important developmental processes controlled by environmental temperature (Begum et al. 2018). In the context of climate change, studying the underlying mechanism of these processes helps to predict the influence of warming on forest production and ecological systems. After dormancy release, reactivation of cell division in the meristem occurs with increasing temperature. In the secondary vascular meristem (cambium), cell division is the first step of wood formation. The seasonal division of cambium cells for many years leads to the radial growth of a tree, so the rate and duration of cambium cell division in one growing season determines the growth rate and wood yield of a forest tree, and consequentially affects the CO2 fixation and carbon sequestration (Locosselli 2018). Regular cell-cycle progression is involved in cell division, and the cell-division machinery has been reported to be the target of many external and internal signals (Li et al. 2009a). Thus, studying the regulatory mechanism of cambium cell division helps to reveal the control of wood formation and production, and understand the regular pattern of carbon sequestration in the context of climate change.
Spring temperature is an important environmental cue to induce the resumption of cambium cell division and the expression of cell-cycle genes (Li et al. 2009b, Begum et al. 2018). The timely expression of cell-cycle genes is essential for cell-cycle progression and cell division, and there is ample knowledge about the expression profiles of cell-cycle genes during tree reactivation in spring (Druart et al. 2007, Li et al. 2009a, 2009b, Qiu et al. 2013, Mishima et al. 2014, Jokipii-Lukkari et al. 2018). Especially, temperature induction of Populus tomentosa Carr. cyclin-dependent kinase B (PtoCDKB) and cyclin B (PtoCYCB) transcription has been verified and confirmed to be independent of bud-burst and the stage of dormancy (Li et al. 2009b), but little is known about the transcriptional regulation of this induction.
More interesting, in some trees in northern temperate regions it is only after the chilling requirement is satisfied in winter that the dormancy can be released, and subsequently they restart to grow in the next spring. When hybrid aspen trees are artificially induced into endodormancy, PttCDKB expression is undetectable at both the mRNA and protein levels; after transferring these endodormant trees into long-day conditions for 3 months, visible growth does not occur (Espinosa-Ruiz et al. 2004). Based on these results, we speculated that the functions of the core cell-cycle regulators may not be reactivated in these trees due to the non-fulfillment of the chilling requirement, and the induced expression of regulators such as PtoCDKB and PtoCYCB by increasing temperature in spring is based on the fulfillment of the chilling requirement in winter (Li et al. 2009a). All these data indicate that the regulation of cell-cycle progression and cambium cell division by environmental temperature is complex and involves the season-specific control of cell-cycle gene expression.
Transcriptional regulation is essential for the developmental and environmental control of cell division, and functions in cell-cycle gene expression (Berckmans and De Veylder 2009, Komaki and Sugimoto 2012, Fischer and Müller 2017, Umeda et al. 2019). In model plants, many transcription factors, such as members of the AP2/ERF (Karlberg et al. 2011, Zhou et al. 2019, Azeez et al. 2021), bHLH (Hachez et al. 2011, Han et al. 2018, Weimer et al. 2018), Dof (Skirycz et al. 2008), GRAS (Sozzani et al. 2010, Zhang et al. 2018), MYB (Ito et al. 2001, Haga et al. 2007, Kato et al. 2009, Ma et al. 2009, Xie et al. 2010, Haga et al. 2011, Vanneste et al. 2011, Yang et al. 2014, Kobayashi et al. 2015) and WOX (Forzani et al. 2014, Peng et al. 2014) families, have been found to control the expression of cell-cycle genes. Notably, EARLY BUD-BREAK 1 (EBB1), an AP2/ERF transcription factor gene, has been found to regulate the timing of bud-break in poplar via directly and negatively regulating SHORT VEGETATIVE PHASE-Like (SVL) expression (Azeez et al. 2021), because its over-expression accelerates bud-burst, whereas its down-regulation delays bud-break, and its expression rapidly increases before bud-break (Yordanov et al. 2014); a similar expression pattern also occurs in apple (Wisniewski et al. 2015), pear (Anh Tuan et al. 2016), grape and spruce (Busov et al. 2016); its pear homolog PpEBB may regulate four cyclin D genes (PpCYCD3), because PpEBB increases the promoter activity of these genes in a dual-luciferase assay using tobacco leaves (Anh Tuan et al. 2016); over-expression of its peach homolog also leads to early bud-break in peach and poplar, likely via changing the expression of genes associated with the cell cycle, hormones and the cell wall (Zhao et al. 2020). More recently, EBB3, another AP2/ERF transcription factor, whose over-expression also leads to early bud-break while its suppression causes a delay in bud-break, has been found to positively and directly regulate the CYCLIND3.1 gene downstream of EBB1 and SVL (Azeez et al. 2021). To check if these regulatory modules function in dormancy release and reactivation of larch will not only provide more information about the link between temperature and cell-cycle gene expression, but also provide new insights into the temperature control of tree growth and development.
The cell cycle is divided into synthesis (S), mitotic (M) and gap phases (G1 and G2). In the active stage, cambium has cycling cells in the G1, S, G2 and M phases, whereas in the dormant stage cambium has no cells in G2 and M (Mellerowicz et al. 1989, 1992, 1995, Zhong et al. 1995, Lloyd et al. 1996). Given the expression patterns of cell-cycle genes during cell-cycle progression (Menges et al. 2005) and the response to temperature (Li et al. 2009b), we suggest that (i) cell-cycle genes are key for the G1-to-S or the G2-to-M transition, and the transcription factors positively controlling their expression are strongly expressed in the active stage of cambium, and (ii) these genes have the same expression patterns in response to temperature. Here, using Larix kaempferi (Lamb.) Carr. (Japanese larch), in which little information is available about the cell-cycle genes, we performed comparative transcriptome analysis to identify the cell-cycle genes key for cell-cycle progression and their transcriptional regulators to better understand the mechanism of the temperature-modulated dormancy release and reactivation in larch.
Materials and methods
Sample preparation and transcriptome sequencing
Branches produced in 2012 were collected from the upper crowns of 1-, 4-, 8-, 12-, 20- and 50-year-old dormant L. kaempferi trees on 10 March 2013 for transcriptome sequencing. The uppermost main stems produced in the current year had been collected from 1-, 2-, 5-, 10-, 25- and 50-year-old active L. kaempferi trees in July 2011 (Li et al. 2014), and their transcriptome sequencing was reported in our previous studies (Li et al. 2014, 2017). The trees were located in Dagujia seed orchard (42°22′ N, 124°51′ E), Liaoning Province, in Northeast China, and from young (1-, 2-, 4-, 5-, 8-, 10-, 12- and 20-year-old), middle-aged (25-year-old) and mature (50-year-old) stands. After removal of buds or needles, the samples from at least three trees from each age category were pooled, frozen in liquid nitrogen and stored at −80 °C until RNA extraction. At the same time, small blocks of vascular tissue were excised from the middle of dormant branches or the bottom of active stems and fixed in formalin–alcohol–acetic acid for anatomical observations. Transcriptome library construction and sequencing were performed for each age category as in our previous study (Li et al. 2014).
Anatomical observations of secondary vascular tissue
The small blocks fixed in formalin–alcohol–acetic acid were dehydrated in an alcohol series and embedded in Spurr’s resin (SPI, USA). Cross-sections 5-μm thick were cut on a microtome (Leitz 1512, Wetzlar, Germany), stained with toluidine blue O and observed under an Axioskop 2 Plus microscope (Zeiss, Gottingen, Germany) equipped with a computer-assisted digital camera. Images of cross-sections of larch cambium regions from the active stage had been captured in our previous study (Li et al. 2017).
Transcriptome assembly and annotation
The raw Illumina RNA sequencing reads from the dormant branches were preprocessed by discarding reads with adapters, with >5% unknown nucleotides, of low-quality (quality score < 20) or < 20 bp. Combining them with the other six Illumina RNA sequencing read sets from the active stems obtained in our previous study (Li et al. 2017), we performed de novo assembly using Trinity software (Grabherr et al. 2011) set to the default parameters.
To identify the L. kaempferi protein-coding genes, blastx (the basic local alignment tool for searching protein databases using a translated nucleotide query; Altschul et al. 1990) was used to search our assembled sequences against blastx_swissprot, protein, blastp_swissport, Pfam, SignalP, eggnog, blast2GO, Pfam2GO and Kegg with an e-value of 1e−5. Then we assigned the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) terms associated with the top hits in the nine protein databases to the annotated transcripts. After sequence clustering with trinity software (Grabherr et al. 2011), non-redundant unigenes were obtained and used for further analysis.
Identification of differentially expressed unigenes and enrichment analysis
To identify the differentially-expressed unigenes (DEUs) between the active and dormant stages in larch, 12 sets of sequencing reads were mapped to the assembled reference transcripts using Bowtie (Langmead and Salzberg 2012). RNA-Seq by expectation maximization (Li and Dewey 2011), an accurate method of transcript quantification from RNA-Seq data, was used to estimate the transcript abundance. The expression of genes was normalized with edgeR (empirical analysis of digital gene expression data in R; Nikolayeva and Robinson 2014). Fragments Per Kilobase of transcript per Million fragments was used to measure the normalized expression value.
One pair-wise comparison was performed to identify the DEUs: active versus dormant. The average normalized expression value from each group was compared. Based on this comparison, DEUs were obtained with stringent cutoffs: a false discovery rate (FDR)-corrected P-value cutoff of 1e−7. The FDR correction is designed to control the expected proportion of incorrectly rejected null hypotheses and is used in multiple-hypothesis testing to reduce type-1 errors.
To identify the biological processes in which the DEUs participated, GO enrichment analysis was performed. Using all the unigenes with assigned GO terms as the background, we identified significantly enriched biological processes with GOatools (https://github.com/tanghaibao/goatools). GOatools uses Fisher’s exact test to identify significant GO terms in one dataset compared with the background. Then, the P-value was corrected for multiple testing with the Bonferroni procedure (Hochberg 1988). The biological processes with corrected P-values ≤0.05 were considered to be enriched in the group.
Prediction of transcription factors and analysis of differences in their expression patterns between the active and dormant stages
The nucleic acid sequences of unigenes were input into a server to predict the transcription factors (http://planttfdb.cbi.pku.edu.cn/prediction.php; Jin et al. 2017), and then their expression patterns were analyzed based on the result of DEU identification.
Cloning of the full-length cDNA sequences and promoter sequences
Based on the assembled transcript sequences and their annotations, special primers were designed to directly amplify the full-length cDNA sequences (see Table S1 available as Supplementary data at Tree Physiology Online). Total RNA was extracted from the active branches of 2-year-old larch trees (sampled on 9 July 2016) with the EasyPure RNA Kit (TransGen Biotech, Beijing, China) according to the manufacturer’s protocol, and then a 2.5-μg aliquot of total RNA was reverse transcribed into cDNA with TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix (TransGen Biotec Co. Ltd, Beijing, China) and subsequently diluted for sequence cloning. The PCR products were purified with a gel extraction kit (Tiangen, Beijing, China), ligated into the pEASY®-T1 simple cloning vector (TransGen Biotech) and sequenced. The deduced amino-acids were used for blast analysis with Arabidopsis thaliana homologs, and based on the results they were designated and annotated again.
The promoter sequences of the cell-cycle genes were cloned with the TADEA-PCR method (Yang et al. 2018). Genomic DNA was isolated from the active branches sampled on 9 July 2016, and the method for cloning was the same as above. The cDNA and promoter sequences were submitted to GenBank (https://www.ncbi.nlm.nih.gov/; see Table S1 available as Supplementary data at Tree Physiology Online).
Transcriptional regulation prediction of cell-cycle genes based on analysis of their promoter sequences
The promoter sequences of the cell-cycle genes were input into the PlantRegMap server (http://plantregmap.cbi.pku.edu.cn/regulation_prediction.php; Jin et al. 2017) to predict the transcription factors that might regulate their expression. The data of seven plant species were used: A. thaliana, Glycine max, Oryza sativa subsp. japonica, P. trichocarpa, Solanum lycopersicum, Triticum aestivum and Zea mays.
Yeast one-hybrid assay
Yeast one-hybrid (Y1H) assays were performed to verify the results of prediction. The open reading frame of a transcription factor was amplified by PCR and cloned into the pGADT7 vector, resulting in pGADT7-transcription factor plasmids (see Table S1 available as Supplementary data at Tree Physiology Online). The sequence fragments of cell-cycle gene promoters of different sizes were amplified and cloned into the pHIS2 vector, resulting in the pHIS2-promoter plasmids (see Table S1 available as Supplementary data at Tree Physiology Online). Then the bait and prey constructs were co-transformed into the yeast strain Y187 using the lithium acetate method, and yeast cells were plated on SD/−Leu/−Trp media and cultured for 3–5 days. The positive clones were selected and plated on SD/−Leu−Trp−His media with 30, 40 or 50 mM 3-amino-1, 2, 4-triazole, separately and cultured for 3–5 days. Possible interactions between transcription factors and promoters were determined based on the growth status of yeast colonies.
Dual-luciferase assay
Dual-luciferase assays were further used to confirm the relationships between transcription factors and cell-cycle genes verified by Y1H assays. The full-length cDNAs of LaRAV1, LaMYB84 and LaMYB20 were individually cloned into the pGreenII 0029 62-SK vector to generate effector constructs (see Table S1 available as Supplementary data at Tree Physiology Online). The LaCDKB1;3 and LaCYCB1;1 promoter fragments were individually cloned into the pGreenII 0800-LUC vector to generate reporter constructs (see Table S1 available as Supplementary data at Tree Physiology Online). Each recombinant construct was individually transformed into Agrobacterium tumefaciens strain GV3101. Nicotiana benthamiana leaves were infected with the mixed Agrobacterium strain. Fluorescence was assessed with the Dual-Luciferase Reporter Assay System (Promega, USA). The firefly luciferase (LUC) activity was normalized to Renilla luciferase (REN) activity, and the LUC/REN ratios were used to represent the activity of the promoter. For each combination, LUC/REN ratios from at least three independent transformations were determined.
Yeast two-hybrid assay
Yeast two-hybrid (Y2H) assays were performed to test the interactions of cell-cycle genes or transcription factors. Their coding sequences were cloned into the pGBKT7 and pGADT7 vectors to fuse with the binding and activation domains, respectively (see Table S1 available as Supplementary data at Tree Physiology Online). Then the bait and prey constructs were co-transformed into yeast strain AH109 using the lithium acetate method, and yeast cells were grown on SD/--Leu--Trp media for 3–5 days. The positive clones were selected and plated onto SD/−Ade/−His/−Leu/−Trp media and cultured for 3–5 days, and then positive clones were transferred onto SD/−Ade/−His/−Leu/−Trp media containing 4 mg ml−1 X-α-Gal to test possible interactions based on the growth status and blue color development in yeast colonies.
Bimolecular fluorescent complementation assay
We used the bimolecular fluorescent complementation (BiFC) assay to directly visualize protein–protein interactions in vivo. The coding sequences of LaCDKB1;3 and LaCYCB1;1 were cloned into the pSM vector to produce the nYFP-LaCDKB1;3 and LaCYCB1;1-cYFP constructs, respectively (see Table S1 available as Supplementary data at Tree Physiology Online). Each construct was individually transformed into A. tumefaciens strain GV3101. Then the mixed Agrobacterium strain was introduced into N. benthamiana leaves via agro-infiltration. After 2 days of incubation, YFP fluorescence was observed in transformed leaf epidermal cells under a laser confocal microscope (Nikon C2-ER).
Quantitative reverse transcription polymerase chain reaction
Experiment I: expression patterns in the active and dormant stages
To confirm the expression patterns of cell-cycle genes and transcription factors detected by RNA-Seq, quantitative reverse transcription PCR (qRT-PCR) was performed. Active seedlings sampled on 9 July 2016 and dormant seedlings sampled on 21 December 2016 were used. Five seedlings were sampled each time. After removal of lateral branches, buds and needles, a 10-cm segment (from the top) of the main stem of each seedling was cut, pooled, immediately frozen in liquid nitrogen and stored at −80 °C until RNA extraction.
Experiment II: expression patterns during reactivation naturally induced by temperature
To check the relationships between the increasing temperatures in early spring and the expression of cell-cycle genes and transcription factors, 1-year-old pot-grown dormant seedlings 19 cm in mean length, which were grown in Beijing (39o48′ N, 116o28′ E), were sampled from the middle of February to late March in 2017 at six time points, and from late February to late March in 2019 at four time points. Five seedlings were sampled each time. The sampling method was the same as above.
Experiment III: expression patterns during reactivation artificially induced by temperature
To further check the relationships between warm temperature and the expression of cell-cycle genes and transcription factors during tree reactivation, 1-year-old pot-grown dormant seedlings were transferred from outdoors into a dark room on 21 December 2016. Forty seedlings 19 cm in mean length were used, including 10 kept outdoors and in the dark as controls, and 25 kept in a dark room at 23 °C. Five seedlings were sampled at 2, 4, 6, 8 and 10 days after treatment; for controls, five seedlings were sampled at 0, 8 and 10 days. The sampling method was the same as above.
Experiment IV: expression patterns during dormancy release and reactivation naturally and artificially induced by temperature
To check the relationships between low temperature during dormancy release and the expression of cell-cycle genes and transcription factors during tree reactivation, forty 2-year-old pot-grown active cutting seedlings were transferred from outdoors into a greenhouse on 13 September 2017, when the seedlings had not experienced a low temperature. On 30 November 2017, when they had entered the dormant stage, 16 seedlings were transferred outside; after 6 weeks, 8 seedlings were sampled on 11 January 2018, and the remaining 8 were transferred into the greenhouse and kept for 2 weeks to determine whether dormancy was released; they were sampled on 25 January 2018. The other 24 seedlings were kept in the greenhouse as controls, 8 seedlings were sampled on 30 November 2017, and 11 and 25 January 2018 as above. The temperature in the greenhouse was >13 °C and no artificial light was supplemented. When sampling, several branches from each seedling were used, and the sampling method was the same as above.
qRT-PCR
Total RNA and cDNA were prepared with the same methods as above. Larch integral membrane protein S1 (LaSYS) was used as an internal control, based on its stable expression in the active and dormant stages, as determined by RNA-Seq. The qRT-PCR analysis was performed with the same methods as previously described (Zhang et al. 2020). The primers for qRT-PCR are listed in Table S1 available as Supplementary data at Tree Physiology Online.
Figure 1.
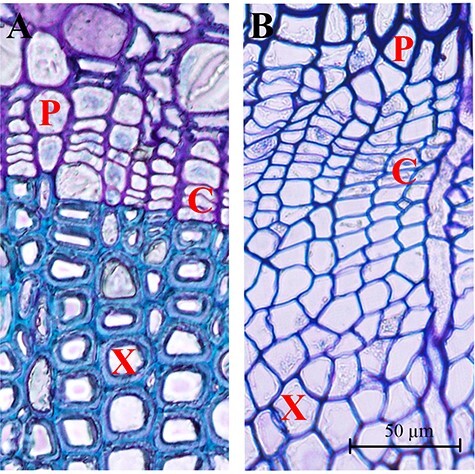
Photomicrographs of cambium regions in cross-section in the dormant stage (10 March 2013) (A) and the active stage (July 2011) (B) of Larix kaempferi. The image of the cambium region from the active stage was taken in our previous study (Li et al. 2017). P, phloem; X, xylem; C, cambium. Scale bar, 50 μm.
Relationship between temperature and gene expression
The data for average, maximum and minimum daily air temperatures near the experimental site in Beijing, China, were obtained from the National Meteorological Information Center, China Meteorological Administration (http://data.cma.cn). Pearson correlation analysis was used to analyze the relationship between temperature and gene expression with the Statistical Product and Service Solutions (SPSS Statistics 26, IBM Corp. New York, USA) program. A P-value ≤0.05 was considered to indicate a correlation.
Results
Cambium cell division occurs in the active stage
In the dormant stage, no dividing cambium cells could be found, and there were three to four cell layers between phloem and xylem cells (Figure 1A), whereas in the active stage, thin, newly-formed cell walls in the cambium regions were found, and differentiating xylem and phloem cells were also found (Li et al. 2017; Figure 1B).
Figure 2.
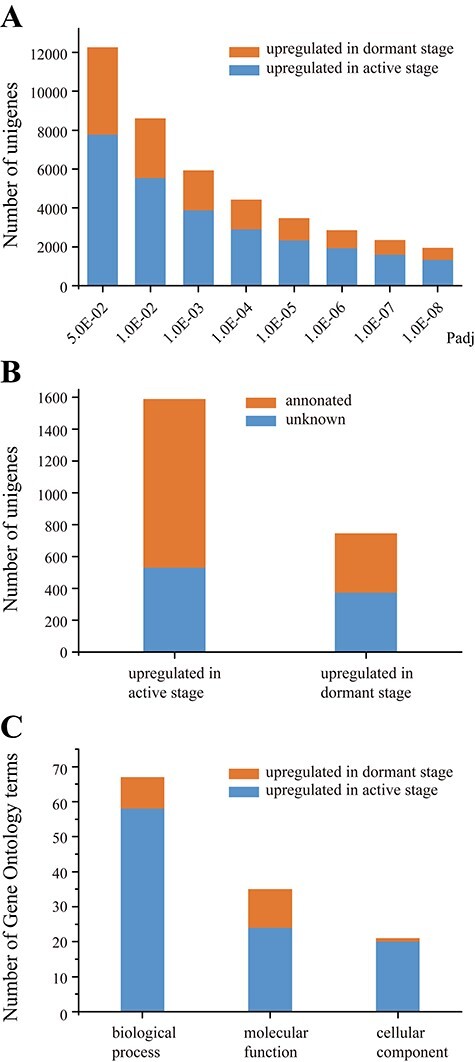
Summary of the DEUs and their enrichment analysis. (A) Number of DEUs with a given adjusted P-value (Padj). (B) Numbers of DEUs with and without annotation. (C) Numbers of Gene Ontology terms enriched in the annotated DEUs.
Transcriptome sequencing, assembly and annotation
A total of 140,691,414 reads were produced from the six age categories of dormant larch trees, with a 101-bp read length (see Table S2 available as Supplementary data at Tree Physiology Online). Together with the published Illumina RNA sequencing reads from the other six categories of active larch trees (Li et al. 2017), all reads were assembled into 128,073 transcripts, including 85,446 unigenes (see Table S3 available as Supplementary data at Tree Physiology Online). Then the 12 sets of reads were aligned to the assembled reference transcripts, and the mapping ratios ranged from 67.46 to 85.63% with an average of 76.44% (see Table S2 available as Supplementary data at Tree Physiology Online).
After annotation, 64,808 (50.60%) transcripts had significant matches and at least one hit in the databases (see Table S3 available as Supplementary data at Tree Physiology Online). GO annotation was then performed for the 64,808 annotated transcripts in terms of ‘biological process’, ‘molecular function’ and ‘cellular component’. In total, 49,392 (76.21%) transcripts were assigned to 8116 GO terms, including 29,857 (75.86%) unigenes (see Table S3 available as Supplementary data at Tree Physiology Online).
Genes up-regulated in the active stage participate in the different processes of wood formation
After comparison, we found that the number of DEUs decreased with the decrease of P-value, and at a given P-value it was always higher for unigenes up-regulated in the active stage than those up-regulated in the dormant stage (Figure 2A). We obtained 2337 DEUs with P-values ≤1e−7 (see Table S4 available as Supplementary data at Tree Physiology Online), including 1591 (68.08%) up-regulated in the active stage and 746 (31.92%) up-regulated in the dormant stage (Figure 2B), and they were used for further analysis. Among the 1591 unigenes, 1061 (66.69%) were annotated, while among the 746 unigenes, 374 (50.13%) were annotated (Figure 2B; see Table S4 available as Supplementary data at Tree Physiology Online).
Figure 5.
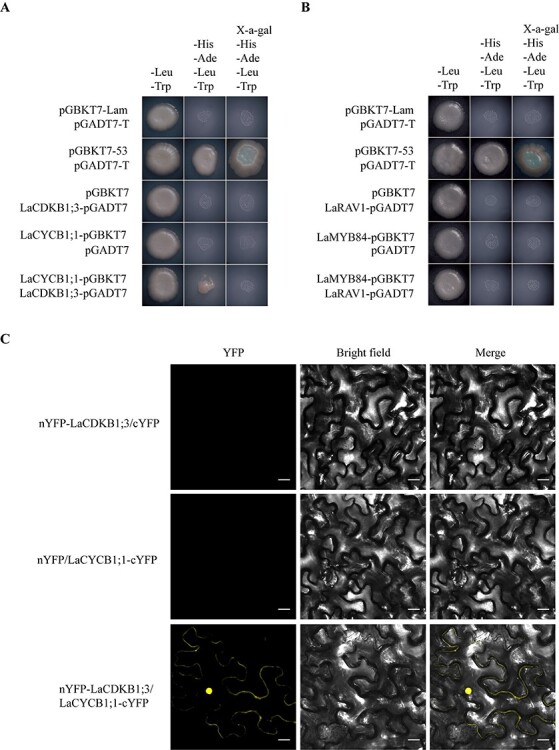
Analysis of protein interaction. Yeast two-hybrid assays show that LaCDKB1;3 interacts with LaCYCB1;1 (A), and LaMYB84 does not interact with LaRAV1 (B). Bimolecular fluorescence complementation assays further show that LaCDKB1;3 interacts with LaCYCB1;1 in tobacco leaves (C). Scale bars, 20 μm.
After analysis of DEUs, 67 GO terms in ‘biological process’ were enriched, including 58 for unigenes up-regulated in the active stage (Figure 2C). Among these 58 terms, many were related to the processes of cell division and wood formation, such as ‘mitotic nuclear division’, ‘mitotic cytokinetic process’, ‘DNA replication initiation’, ‘cytokinesis by cell plate formation’, ‘lignin catabolic process’, ‘pectin catabolic process’, ‘cellulose metabolic process’ and ‘plant-type secondary cell wall biogenesis’ (see Table S5 available as Supplementary data at Tree Physiology Online).
To study the temperature control of cambium cell division, cell-cycle genes were emphasized. Among 1435 DEUs (see Table S4 available as Supplementary data at Tree Physiology Online), 10 were annotated as cyclins and three as cyclin-dependent kinase B (CDKB), while 12 were successfully identified (Table 1). In addition, a unigene annotated as cyclin-dependent kinase A (CDKA) was also cloned and analyzed (Table 1). These 13 cell-cycle genes were designated after alignment analysis of their deduced amino-acids with their homologs from A. thaliana.
Table 1.
Expression patterns of 13 cell-cycle genes in the active (July 2011) and dormant (10 March 2013) stages of Larix kaempferi
| Family | Gene | Transcript ID | Active | Dormant | P value |
|---|---|---|---|---|---|
| CDKA | LaCDKA | TR20791|c0_g1 | 582.65 | 555.49 | 1 |
| CDKB | LaCDKB1;1 | TR22283|c0_g1 | 149.50 | 0.00 | 1.34E-21 |
| LaCDKB1;2 | TR24194|c0_g1 | 105.32 | 3.69 | 7.51E-13 | |
| LaCDKB1;3 | TR25126|c0_g2 | 266.96 | 1.41 | 7.71E-27 | |
| CYCA | LaCYCA1;1 | TR34021|c0_g1 | 67.23 | 0.00 | 4.44E-17 |
| LaCYCA2;1 | TR35988|c0_g1 | 131.85 | 0.23 | 7.44E-23 | |
| LaCYCA2;2 | TR32688|c0_g1 | 88.19 | 0.84 | 2.58E-16 | |
| CYCB | LaCYCB1;1 | TR31642|c1_g1 | 148.55 | 7.92 | 3.26E-12 |
| LaCYCB1;2 | TR36747|c2_g1 | 217.61 | 2.60 | 4.45E-22 | |
| LaCYCB2;3 | TR23547|c0_g1 | 132.71 | 2.03 | 2.66E-18 | |
| CYCD | LaCYCD1;1 | TR35481|c0_g1 | 128.59 | 4.49 | 5.28E-14 |
| LaCYCD3;1 | TR27202|c0_g1 | 448.04 | 6.05 | 7.08E-24 | |
| CYCU | LaCYCU4;1 | TR26542|c0_g1 | 357.46 | 47.23 | 5.60E-08 |
Branches produced in 2012 from the upper crowns of dormant L. kaempferi trees sampled on 10 March 2013 and the uppermost main stems produced in the current year sampled from active L. kaempferi trees in July 2011 were used for RNA extraction, transcriptome library construction, and sequencing (Li et al. 2014, 2017). Here, by comparing the transcriptomes of the dormant (10 March 2013) and active (July 2011) stages, 13 cell-cycle genes were identified. Fragments per kilobase of transcript per million fragments was used to indicate the expression value of each transcript
Identification of transcription factors controlling cell-cycle gene expression
In total, 584 transcription factors belonging to 50 families were predicted from 85,446 unigenes (see Figure S1 available as Supplementary data at Tree Physiology Online). Among 2337 DEUs, there were 43 transcription factors (1.84%), and 31 (including nine members of the MYB family) were up-regulated in the active stage and co-expressed with 12 identified cell-cycle genes (Table 2).
Table 2.
Expression patterns of 43 transcription factors in the active (July 2011) and dormant (10 March 2013) stages of Larix kaempferi
| Family | Gene | Transcript ID | Active | Dormant | P value |
|---|---|---|---|---|---|
| AP2 | – | TR27925|c0_g1 | 62.25 | 2.21 | 2.29E-10 |
| ARF | – | TR40380|c3_g2 | 30.00 | 345.70 | 6.87E-11 |
| B3 | LaRAV1 | TR33337|c0_g2 | 22.31 | 0.00 | 4.80E-08 |
| LaMVI11 | TR41465|c0_g2 | 75.97 | 1.68 | 2.32E-13 | |
| BES1 | – | TR37270|c3_g1 | 59.60 | 1.51 | 4.73E-10 |
| bHLH | – | TR31737|c0_g1 | 0.45 | 134.29 | 1.62E-25 |
| LaBHLH49 | TR29122|c0_g1 | 288.87 | 2.86 | 3.02E-24 | |
| bZIP | LaBZIP34 | TR33610|c0_g1 | 210.37 | 3.49 | 6.28E-20 |
| C2H2 | – | TR35597|c1_g2 | 39.05 | 0.90 | 4.09E-09 |
| C3H | – | TR31206|c0_g1 | 256.82 | 6.17 | 1.08E-08 |
| – | TR35029|c0_g1 | 54.59 | 0.34 | 3.09E-13 | |
| Dof | – | TR38661|c0_g3 | 14.10 | 141.26 | 2.93E-08 |
| E2F/DP | LaE2FE | TR40959|c0_g1 | 102.97 | 1.56 | 2.85E-16 |
| ERF | – | TR21912|c0_g1 | 60.88 | 677.48 | 2.99E-10 |
| – | TR31180|c0_g2 | 84.70 | 1405.57 | 4.60E-14 | |
| – | TR35329|c0_g2 | 1.45 | 939.59 | 2.40E-42 | |
| – | TR40054|c1_g4 | 3.72 | 235.10 | 2.60E-20 | |
| GATA | – | TR23360|c0_g1 | 228.24 | 16.10 | 2.04E-11 |
| GRAS | LaSCL28 | TR41410|c0_g2 | 123.57 | 0.28 | 2.80E-22 |
| HD-ZIP | – | TR30886|c0_g1 | 130.93 | 0.00 | 2.98E-10 |
| – | TR3870|c0_g2 | 47.96 | 0.00 | 1.67E-09 | |
| HSF | – | TR34423|c0_g1 | 215.95 | 18.18 | 3.88E-10 |
| LBD | – | TR21818|c0_g1 | 0.79 | 991.40 | 1.22E-45 |
| MIKC_MADS | LaAGL9 | TR27522|c0_g1 | 186.99 | 0.46 | 6.11E-09 |
| MYB | LaMYB3 | TR36732|c0_g4 | 182.60 | 1.08 | 2.68E-09 |
| LaMYB16;1 | TR36192|c2_g1 | 197.60 | 8.28 | 4.22E-14 | |
| LaMYB16;2 | TR36406|c0_g2 | 33.78 | 0.00 | 2.71E-09 | |
| LaMYB20 | TR35678|c0_g1 | 50.56 | 0.87 | 2.89E-11 | |
| LaMYB46 | TR26443|c0_g1 | 231.50 | 0.49 | 2.74E-28 | |
| LaMYB61 | TR29942|c0_g1 | 374.97 | 1.55 | 1.12E-30 | |
| LaMYB84 | TR28142|c0_g1 | 237.50 | 0.52 | 8.66E-16 | |
| LaMYB93 | TR33695|c0_g1 | 91.15 | 1.01 | 6.59E-08 | |
| LaMYB103 | TR33771|c0_g1 | 187.56 | 0.00 | 6.07E-24 | |
| MYB-related | – | TR38200|c0_g2 | 40.53 | 281.93 | 9.02E-08 |
| – | TR29733|c0_g1 | 50.41 | 2.53 | 1.80E-08 | |
| NAC | – | TR30989|c0_g1 | 1.23 | 2040.82 | 9.56E-49 |
| – | TR34438|c0_g2 | 55.24 | 1748.24 | 5.38E-20 | |
| – | TR19020|c0_g5 | 291.50 | 7.15 | 5.78E-10 | |
| – | TR36333|c0_g2 | 127.01 | 4.51 | 3.71E-14 | |
| – | TR40492|c1_g2 | 127.24 | 4.75 | 1.33E-13 | |
| – | TR41700|c0_g1 | 70.69 | 0.67 | 5.08E-08 | |
| RAV | – | TR37835|c5_g1 | 9.03 | 531.09 | 2.45E-15 |
| ZF-HD | – | TR41115|c1_g4 | 50.46 | 3.30 | 6.80E-08 |
–, indicates no cloning; based on the new annotation with PlantTFDB v5.0 (http://planttfdb.gao-lab.org/aboutus.php#update), LaRAV1 belongs to RAV family
Branches produced in 2012 from the upper crowns of the dormant L. kaempferi trees sampled on 10 March 2013 and the uppermost main stems produced in the current year sampled from active L. kaempferi trees in July 2011 were used for RNA extraction, transcriptome library construction, and sequencing (Li et al. 2014, 2017). Here, by comparing the transcriptome of dormant (10 March 2013) and active (July 2011) stages, 43 transcription factors from 22 families were identified. Fragments per kilobase of transcript per million fragments was used to indicate the expression value of each transcript
To predict the transcriptional regulators of the 12 differentially expressed cell-cycle genes, we cloned their promoter sequences and found that transcription factors from 10 families might control their expression (Figure 3A and B). The 31 transcription factors that were co-expressed with the 12 cell-cycle genes belonged to 17 families, having seven families in common with the 10 predicted families (Figure 3C). In these seven common families there were 16 transcription factors. Their full-length cDNA sequences were cloned, and they were designated in the same way as cell-cycle genes (Table 2). A total of 73 predicted cases of regulation existed between these 16 transcription factors and the 12 cell-cycle genes (Figure 3D), whereas only three were confirmed by Y1H assays: regulation between LaMYB20 and LaCYCB1;1, LaMYB84 and LaCDKB1;3, and LaRAV1 and LaCDKB1;3 (Figure 4A–C). Dual-luciferase assays further confirmed that LaMYB20 and LaRAV1 increased the promoter activity of LaCYCB1;1 and LaCDKB1;3 (P ≤ 0.05), respectively (Figure 4E and G).
Figure 9.

Expression patterns of two cell-cycle genes (LaCDKB1;3 and LaCYCB1;1) and three transcription factors (LaMYB20, LaMYB84 and LaRAV1) during reactivation artificially induced by temperature. One-year-old pot-grown dormant Larix kaempferi seedlings were transferred from outdoors into a dark room on 21 December 2016. Forty seedlings 19 cm in mean length were used: 10 were kept outdoors and in the dark as controls and 25 were kept in a dark room at 23 °C. Five seedlings were sampled at 2, 4, 6, 8 and 10 days after treatment; for controls five seedlings were sampled at 0, 8 and 10 days. (A) Climatic daily temperatures from 21 to 31 December 2016. Variation of LaCDKB1;3 (B), LaCYCB1;1 (C), LaMYB20 (D), LaMYB84 (E) and LaRAV1 (F) expression assayed by qRT-PCR with LaSYS as the internal control. The P-values for LaCDKB1;3 and LaRAV1 were generated between samples harvested at 2 and 4 days after treatment, at 0 and 10 days for the control seedlings and for LaCYCB1;1 at 4 and 6 days after treatment.
Figure 10.
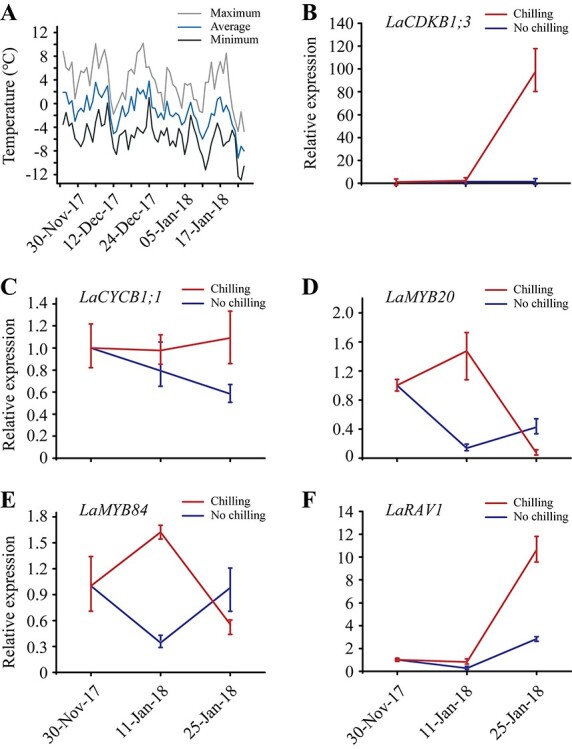
Expression patterns of two cell-cycle genes (LaCDKB1;3 and LaCYCB1;1) and three transcription factors (LaMYB20, LaMYB84 and LaRAV1) during dormancy release and reactivation naturally or artificially induced by temperature. Forty 2-year-old pot-grown active Larix kaempferi cutting seedlings were transferred from outdoors into a greenhouse on 13 September 2017, when they had not experienced low temperatures. On 30 November 2017, when they had entered the dormant stage, 16 seedlings were transferred outside; after 6 weeks, eight seedlings were sampled on 11 January 2018 and the remaining eight were transferred into the greenhouse and kept for 2 weeks to determine whether dormancy was released; they were sampled on 25 January 2018. The other 24 seedlings were kept in the greenhouse as controls, and eight seedlings were sampled on 30 November 2017 and 11 and 25 January 2018. (A) Climatic daily temperatures from 25 November 2017 to 25 January 2018. Variation of LaCDKB1;3 (B), LaCYCB1;1 (C), LaMYB20 (D), LaMYB84 (E) and LaRAV1 (F) expression assayed by qRT-PCR with LaSYS as the internal control. The P-values for LaCDKB1;3 and LaRAV1 were generated between samples harvested on 11 and 25 January 2018 with natural chilling, and on 25 January 2018 with or without natural chilling.
Figure 4.
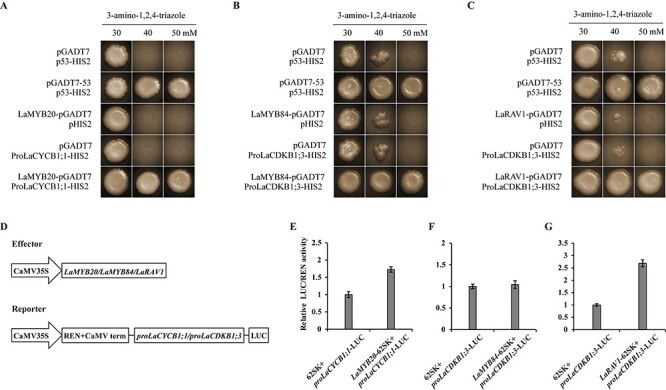
Analysis of interactions between transcription factors and cell-cycle genes. Yeast one-hybrid assays show that LaMYB20 binds to the promoter of LaCYCB1;1 (ProLaCYCB1;1) (A), and both LaMYB84 (B) and LaRAV1 (C) bind to the promoter of LaCDKB1;3 (ProLaCDKB1;3). Dual-luciferase assays show that LaMYB20 (E) and LaRAV1 (G) increase the promoter activity of LaCYCB1;1 (ProLaCYCB1;1) and LaCDKB1;3 (ProLaCDKB1;3; P ≤ 0.05), respectively, and LaMYB84 (F) does not increase the promoter activity of LaCDKB1;3 (ProLaCDKB1;3). Schematic diagrams of the effector and reporter vector used in dual-luciferase assays is shown in (D) [values are the ratio of firefly luciferase (LUC) to Renilla luciferase (REN) activity]. Data represent the mean ± SD of three biological replications. Error bars represent standard error. Statistical significance was determined using Student’s t-test.
Based on the results of Y1H assays, both LaMYB84 and LaRAV1 might regulate LaCDKB1;3 expression. When Y2H assay was performed to check if they constitute a complex and work together, no interaction between them was found (Figure 5B). Notably, an interaction existed between LaCDKB1;3 and LaCYCB1;1, as verified by Y2H and BiFC assays (Figure 5A and C).
Figure 11.
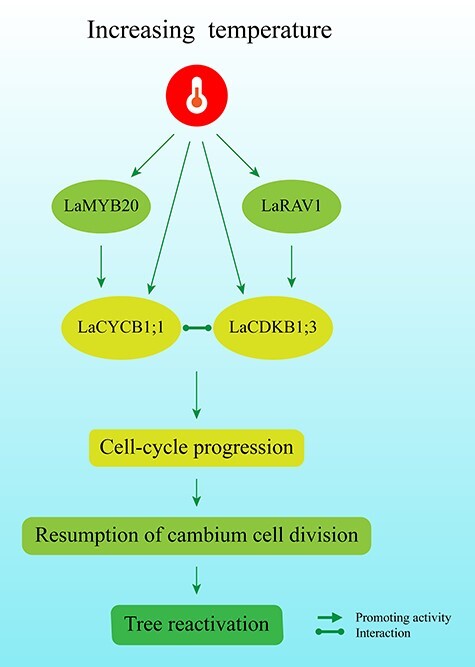
Regulatory network model for Larix kaempferi dormancy release and reactivation based on the expression patterns of cell-cycle genes and transcription factors and their relationships and responses to temperature. After fulfillment of the chilling requirement in winter, the increasing spring temperatures induce LaCDKB1;3, LaCYCB1;1, LaRAV1 and LaMYB20 expression; meanwhile, LaRAV1 and LaMYB20 promote LaCDKB1;3 and LaCYCB1;1 expression, respectively, and LaCDKB1;3 interacts with LaCYCB1;1, together promoting the resumption of cell-cycle progression and cambium cell division.
Variations of LaCDKB1;3, LaCYCB1;1, LaMYB20, LaMYB84 and LaRAV1 expression in response to temperature
To confirm the expression patterns revealed by RNA-Seq, we measured the variations of LaCDKB1;3, LaCYCB1;1, LaMYB20, LaMYB84 and LaRAV1 expression between the active and dormant stages using qRT-PCR. The results showed that they were all strongly expressed in the active stage and weakly in the dormant stage, showing almost the same patterns as those revealed by RNA-Seq (Figure 6).
Figure 6.
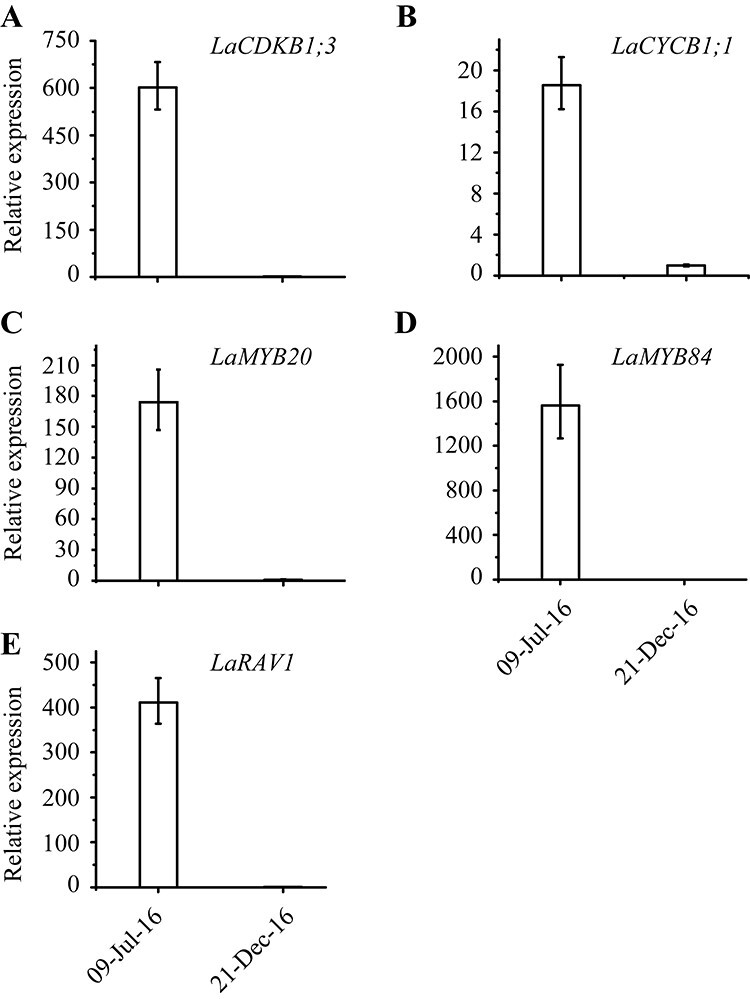
Expression patterns of two cell-cycle genes [LaCDKB1;3 (A) and LaCYCB1;1 (B)] and three transcription factors [LaMYB20 (C), LaMYB84 (D) and LaRAV1 (E)] in the active (9 July 2016) and dormant stages (21 December 2016) of Larix kaempferi seedlings (n ≥ 5) assayed by qRT-PCR with LaSYS as the internal control.
In the early spring of 2017, with the increase of temperature (Figure 7A), the levels of LaCDKB1;3, LaCYCB1;1, LaMYB20, LaMYB84 and LaRAV1 transcripts changed, but showed different patterns: LaCDKB1;3, LaCYCB1;1 and LaRAV1 increased from late February to March (P ≤ 0.05; Figure 7). Almost the same expression patterns occurred in the early spring of 2019 (see Figure S2 available as Supplementary data at Tree Physiology Online).
Figure 7.
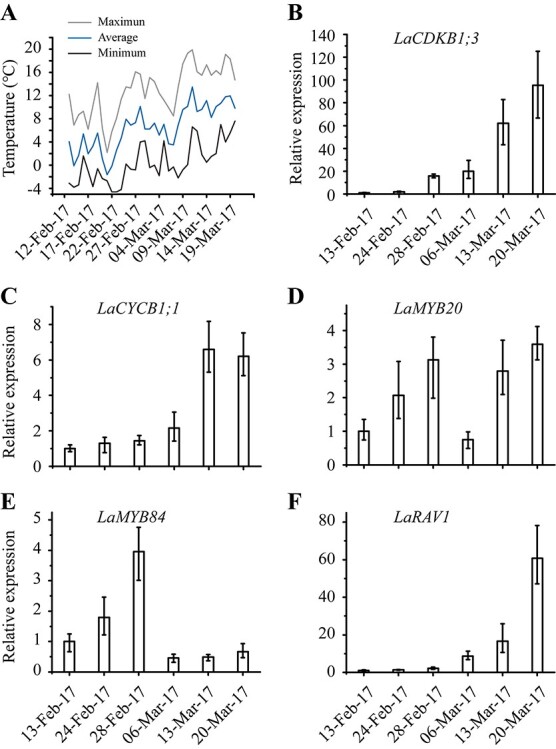
Expression patterns of two cell-cycle genes (LaCDKB1;3 and LaCYCB1;1) and three transcription factors (LaMYB20, LaMYB84 and LaRAV1) during reactivation naturally induced by temperature. (A) Climatic daily temperatures from 13 February to 20 March 2017. Variation of LaCDKB1;3 (B), LaCYCB1;1 (C), LaMYB20 (D), LaMYB84 (E) and LaRAV1 (F) expression from 13 February to 20 March 2017 during the reactivation of Larix kaempferi seedlings (n ≥ 5) assayed by qRT-PCR with LaSYS as the internal control. The P-value for LaCDKB1;3 was generated between samples harvested on 24 and 28 February 2017, and 6 and 13 March 2017; for LaCYCB1;1 on 6 and 13 March 2017; and for LaRAV1 on 28 February and 6 March 2017.
To assess the relationships between temperature and gene expression, we used Pearson correlation analysis. The results showed positive correlations between temperature and LaCDKB1;3, LaCYCB1;1, LaMYB20 and LaRAV1 transcript levels, but the correlation was different for each of them (Figure 8). The expression of LaMYB20 correlated with the daily average temperature (P ≤ 0.01; Figure 8C). The expression of LaRAV1 correlated with the mean minimum temperature at least for three sampling days (P ≤ 0.05; Figure 8B). The expression of LaCDKB1;3 correlated with the daily minimum temperature (P ≤ 0.05; Figure 8B), but it was also correlated with the mean minimum temperature for at least three sampling days (P ≤ 0.01; Figure 8B). The expression of LaCYCB1;1 correlated with the mean minimum temperature for at least two sampling days (P ≤ 0.05; Figure 8B).
Figure 8.

Correlations between gene expression and temperature. Pearson correlation coefficients between the expression levels of two cell-cycle genes (LaCDKB1;3 and LaCYCB1;1) and three transcription factors (LaMYB20, LaMYB84 and LaRAV1) and maximum temperature (A), minimum temperature (B) and average temperature (C) from 30 January to 20 March 2017. Pearson correlation coefficient values greater than those indicated by the green and red lines means that the correlations between gene expression and temperature were significant at P ≤ 0.05 and P ≤ 0.01, respectively.
The effects of temperature on LaCDKB1;3, LaCYCB1;1, LaMYB20, LaMYB84 and LaRAV1 expression were further determined in the temperature-treatment experiment. After the transfer of dormant seedlings from outdoor into a dark room at 23 °C, their transcript levels changed, but showed different patterns (Figure 9). LaCDKB1;3 and LaRAV1 expression increased after 4 days (P ≤ 0.05); in contrast, almost no regular increases occurred in the seedlings kept outdoors for 10 days (≤6 °C; P ≥ 0.05; Figure 9B and F). LaCYCB1;1 expression increased after 6 days (P ≤ 0.05; Figure 9C).
The effects of chilling on LaCDKB1;3, LaCYCB1;1, LaMYB20, LaMYB84 and LaRAV1 expression during tree reactivation were checked in another temperature-treatment experiment. After 6 weeks of natural chilling (≤10.2 °C) and the following 2 weeks of artificial warming in the greenhouse (≥13 °C), only LaCDKB1;3 and LaRAV1 showed increases in expression (P ≤ 0.05; Figure 10B and F). On 25 January 2018 when the treatment was ended, bud-break and new needles were visible in the seedlings with chilling treatment, and invisible in the seedlings without it (see Figure S3 available as Supplementary data at Tree Physiology Online). Notably, at the end of the treatment, higher levels of LaCDKB1;3 and LaRAV1 transcripts were measured in the seedlings with chilling treatment than in those without it (P ≤ 0.05; Figure 10B and F).
Discussion
Switch from the dormant to the active stage requires extensive and complex transcriptome reprogramming
When genes in at least four of the age categories and with an average normalized expression value >0.5 were considered to be expressed, 45,839 of 85,446 unigenes (53.65%) were expressed in the active stage and 28,603 (33.47%) in the dormant stage. Based on the comparison and GO enrichment analysis, a total of 2337 DEUs and 67 biological processes were identified (Figure 2C; see Table S4 available as Supplementary data at Tree Physiology Online). The number of unigenes up-regulated in the active stage was higher than that in the dormant stage, and more GO terms were enriched in the active stage (Figure 2B and C). These data indicate that more genes function in the active stage (Jokipii-Lukkari et al. 2018), and the complexity of the stem transcriptome increases from the dormant stage to the active stage (Schrader et al. 2004).
Wood formation involves cambium cell division, cell expansion, cell wall formation and cell death (Plomion et al. 2001). During the active stage, wood formation occurs actively (Figure 1B), and this was reflected and supported by our enrichment analysis, in which different processes of wood formation were identified. For example, the GO terms ‘mitotic cell cycle process’, ‘lignin metabolic process’, ‘lignin catabolic process’, ‘plant-type secondary cell wall biogenesis’, ‘cell wall organization’ and ‘sucrose metabolic process’ were enriched by the unigenes up-regulated in the active stage (see Table S5 available as Supplementary data at Tree Physiology Online). Entry into the dormant stage means cessation of wood formation and acquisition of other biological process, such as resistance to drought, and these were also found from our GO enrichment analysis. For example, the GO terms ‘response to water’ and ‘response to water deprivation’ were enriched for the unigenes up-regulated in the dormant stage (see Table S5 available as Supplementary data at Tree Physiology Online).
All RNA-Seq data in this study have been deposited in the NCBI SRA database (see Table S2 available as Supplementary data at Tree Physiology Online). Together with other transcriptome and genomic resources for Larix (Zhang et al. 2012, Men et al. 2013, Han et al. 2014, Li et al. 2014, He et al. 2015, 2020, Liu et al. 2016, Kuzmin et al. 2019), our data will serve as a useful tool to support investigation of the molecular basis of many aspects of larch tree growth and development, such as somatic embryogenesis, rooting, wood formation, aging, phase change, adaption and response to the environment. With these sequences, many molecular markers can also be developed to assist breeding and genetic diversity studies of Larix.
Control of cell-cycle genes is involved in the seasonal activity of cambium
In poplar and spruce, some cell-cycle genes are strongly expressed in cambium regions during the active stage (Schrader et al. 2004, Jokipii-Lukkari et al. 2017, Sundell et al. 2017), indicating their regulatory roles in cambium cell division; in addition, some of them have seasonal expression patterns (Jokipii-Lukkari et al. 2018, Li et al. 2009b), suggesting that their expression and function are synchronous with the season or environment, like cambium cell division and wood formation.
The seasonal synchronization of cell-cycle gene expression in temperate regions means that their expression is controlled by the seasonal environmental signals, such as temperature and day length, and this has been examined. For example, during the reactivation of poplar trees in spring, the increasing temperature induces the transcription of PtoCDKB and PtoCYCB (Li et al. 2009b); during the establishment of dormancy, the short-day signal decreases PttCDKA and PttCDKB translation (Espinosa-Ruiz et al. 2004). In addition, the short-day signal and low temperature decrease PcyclAt-gus and Pcdc2a-gus expression, respectively (Rohde et al. 1997). Here, 49 unigenes enriched in ‘mitotic cell cycle process’ might also be the targets of environmental signals and involved in the seasonal regulation of cambium activity in larch (see Table S5 available as Supplementary data at Tree Physiology Online). These data provide not only more information about the molecular basis of cambium activity and wood formation in gymnosperms, but also more candidate genes to study the seasonal control of tree growth and development via investigating the regulation of their expression.
There are two checkpoints in cell-cycle progression, and passing them requires special interaction between CDK and cyclin. For example, the CDKA and CYCD complex plays central roles at the G1−S checkpoint, and binding of CDKB to CYCB is required at the G2−M checkpoint (Scofield et al. 2014). Here, we found that the transcript level of LaCDKA was stable (Table 1), but those of genes putatively encoding its binding protein were not stable and strictly controlled; for example, LaCYCA, LaCYCD and LaCYCU were strongly expressed in the active stage, and weakly expressed in the dormant stage (Table 1). Notably, in poplar the transcript level of CDKA is also stable (Espinosa-Ruiz et al. 2004, Li et al. 2009a), while its protein level is controlled by the short-day signal and is not detectable when the tree enters into the endodormant stage (Espinosa-Ruiz et al. 2004); the regulatory mechanism is still unknown. For the cell-cycle genes controlling the G2-to-M transition, LaCDKB and LaCYCB had the same expression patterns, strongly expressed in the active stage and weakly in the dormant stage (Table 1), and these patterns have also been found in poplar (Li et al. 2009a, 2009b). These are different from those of the cell-cycle genes controlling the G1-to-S transition and might be orchestrated by the mechanism previously described (Kato et al. 2009, Haga et al. 2011). Notably, interaction indeed occurred between LaCDKB1;3 and LaCYCB1;1 (Figure 5), further showing that cell-cycle genes controlling the G2-to-M transition and functioning together are controlled by the same mechanisms downstream of environmental signals. However, further experiments are required to check the relationships between other CDKs and cyclins and their regulation.
Trees can modulate their growth and development with changes in ambient environmental conditions and forest management, because meristem forms one link in this response mechanism, in which we suppose that cell-cycle genes and their regulation play roles. In the context of climate change, the identification of 13 core cell-cycle genes provides a chance to study the environmental control of meristem activity during forest tree adaption and response to the environment, and the molecular basis of the effect of management on forest tree growth and development through analyzing the regulation of their expression, which is of great theoretical and practical importance.
The transcriptional regulatory network of cell-cycle genes during cambium cell division
In addition to cell-cycle genes, multiple transcription factors are also expressed synchronously with the changing seasons (Schrader et al. 2004, Ko et al. 2006, Druart et al. 2007, Galindo González et al. 2012, Qiu et al. 2013, Mishima et al. 2014, Yordanov et al. 2014, Jokipii-Lukkari et al. 2018, Azeez et al. 2021), such as members of the AP2/ERF, HD-ZIP and MYB families. Information is still limited as to whether these transcription factors control cell-cycle gene expression during cambium cell division. Here, we found that 31 transcription factors from 17 families were strongly expressed in the active stage and had the same expression patterns as 12 cell-cycle genes (Tables 1 and 2), indicating that regulation might occur between them and be involved in cambium cell division.
After analysis of the promoter sequences of the 12 cell-cycle genes we found that transcription factors from 10 families might control their expression (Figure 3B). Notably, the B3 family was predicted from all seven species (Figure 3B), suggesting that the control of cell-cycle gene expression by the B3 family is an evolutionarily conserved program for the regulation of cell-cycle progression between angiosperms and gymnosperms, and between annuals and perennials independent of the number of cotyledons. Altogether, 16 transcription factors from seven families might control the expression of 10 cell-cycle genes and 73 cases of regulation were predicted, while only three were confirmed by Y1H assays (Figure 3D): between LaMYB20 and LaCYCB1;1, LaMYB84 and LaCDKB1;3, and LaRAV1 and LaCDKB1;3, and two were further confirmed by dual-luciferase assays: between LaMYB20 and LaCYCB1;1, and LaRAV1 and LaCDKB1;3. Among these, one transcription factor was from the MYB family and one from the B3 family, and two cell-cycle genes include one CDKB and one CYCB and an interaction exists between them (Figure 5).
Notably, the homolog of LaMYB20 from A. thaliana, MYB20, is a member of the VIII-A-2 R2R3-MYB protein family (Jiang and Rao 2020), and regulates phenylalanine and lignin biosynthesis during secondary cell wall formation (Geng et al. 2020), suggesting that these homologs integrate cell-cycle progression and terminal differentiation to orchestrate the processes of wood formation. As to LaRAV1, although the functions of its homologs from other plants have received much attention (Moreno-Cortés et al. 2012, 2017, Matías-Hernández et al. 2014, Triozzi et al. 2018), to our knowledge, its regulation of cell-cycle genes is first reported here.
Together, these results not only add more information to construct the transcriptional regulatory network of cell-cycle genes during cambium cell division, but also offer a means of studying other processes of wood formation, such as cell expansion and secondary cell wall biogenesis, and adaption to the winter environment.
LaRAV1 regulation of LaCDKB1;3 expression is involved in the temperature-modulated dormancy release and reactivation of larch
To determine if the identified three forms of regulation mediate the control of tree reactivation by environment temperature, we detected the expression patterns of LaCDKB1;3, LaCYCB1;1, LaRAV1, LaMYB20 and LaMYB84 in early spring. Based on their expression patterns and relationships with temperature (Figure 8), we found that their responses to temperature and its effects were different. LaRAV1 and LaCDKB1;3 had almost the same response patterns and might belong to the same signaling pathway downstream of temperature.
To confirm these results, we measured their responses to temperature in another experiment, and found that only LaRAV1 and LaCDKB1;3 had almost the same response patterns in response to 23 °C, further suggesting that the LaRAV1-LaCDKB1;3 module functions in the signaling pathway downstream of temperature. These data not only add new functional information about RAV genes (Moreno-Cortés et al. 2012, 2017, Matías-Hernández et al. 2014, Triozzi et al. 2018), but also provide novel insights into the temperature control of cell-cycle gene expression during cambium reactivation.
Notably, the poplar homolog of LaRAV1 increases FT2 expression in a transactivation assay in poplar leaf tissues as an activator, suggesting a new and divergent function for RAV1 with respect to its Arabidopsis homolog, TEMPRANILLO (TEM), which is a strong repressor of FT and flowering time (Triozzi et al. 2018); poplar RAV1 and Arabidopsis TEM show different diurnal expression patterns downstream of photoperiod signaling (Triozzi et al. 2018). Together with our results, these data indicate that RAV1/TEM functions as an activator or a repressor, and its expression is controlled by photoperiod and temperature in addition to pathogen infections and abiotic stresses (Matías-Hernández et al. 2014, Triozzi et al. 2018).
Chilling is required for dormancy release and is assumed to be the prerequisite for the induction of cell-cycle gene transcription by warmer temperatures (Li et al. 2009a). To test this speculation, we detected the expression patterns of LaCDKB1;3, LaCYCB1;1, LaRAV1, LaMYB20 and LaMYB84 in larch seedlings exposed to temperature treatments. After 6 weeks of chilling and 2 weeks of warming, dormant buds broke, new needles appeared (see Figure S3 available as Supplementary data at Tree Physiology Online), and the transcript levels of LaCDKB1;3 and LaRAV1 increased (Figure 10), while without chilling, dormant buds did not break and no new needles appeared (see Figure S3 available as Supplementary data at Tree Physiology Online). That is to say, bud dormancy can only be released after the chilling requirement is fulfilled, the expression of some key cell-cycle regulators can be induced by the following warm temperature, and finally bud-break occurs. These results support our speculation, not only providing new insights into the mechanism of the chilling induction of dormancy release (Brunner et al. 2014), but also offering evidence that tree growth can be affected by the climatic conditions of both the previous and current seasons. However, further experiments are required to establish links between the LaRAV1-LaCDKB1;3 module and the SVL-TCP18/BRC1 and EBB1-SVL-EBB3-CYCD3.1 modules (Singh et al. 2018, Azeez et al. 2021), which also mediate the control of tree reactivation.
Conclusions
Altogether, in this work we measured the expression of cell-cycle genes and transcription factors with up-regulation in the active stage and assessed their relationships and responses to temperatures. Based on the results, we proposed a model (Figure 11) in which increasing temperatures induce LaCDKB1;3, LaCYCB1;1, LaRAV1 and LaMYB20 expression after fulfillment of the chilling requirement. Meanwhile, LaRAV1 and LaMYB20 promote LaCDKB1;3 and LaCYCB1;1 expression, respectively, and LaCDKB1;3 interacts with LaCYCB1;1, together promoting the resumption of cell-cycle progression and cambium cell division. This model helps to reveal the mechanisms of genetic and temperature control of forest tree reactivation.
Supplementary Material
{kind=link}
Acknowledgments
The authors thank Dr I.C. Bruce (Peking University) for critical reading of the manuscript, and Dr Dan-Yang Wang (Northwest University) for help in yeast assays.
Contributor Information
Wan-Feng Li, State Key Laboratory of Tree Genetics and Breeding, Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Research Institute of Forestry, Chinese Academy of Forestry, No. 1, Dongxiaofu, Xiangshan Road, Haidian District, Beijing 100091, People's Republic of China.
Yanhui Kang, State Key Laboratory of Tree Genetics and Breeding, Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Research Institute of Forestry, Chinese Academy of Forestry, No. 1, Dongxiaofu, Xiangshan Road, Haidian District, Beijing 100091, People's Republic of China.
Yao Zhang, State Key Laboratory of Tree Genetics and Breeding, Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, No. 1, Dongxiaofu, Xiangshan Road, Haidian District, Beijing 100091, People's Republic of China.
Qiao-Lu Zang, State Key Laboratory of Tree Genetics and Breeding, Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Research Institute of Forestry, Chinese Academy of Forestry, No. 1, Dongxiaofu, Xiangshan Road, Haidian District, Beijing 100091, People's Republic of China.
Li-Wang Qi, State Key Laboratory of Tree Genetics and Breeding, Key Laboratory of Tree Breeding and Cultivation, National Forestry and Grassland Administration, Research Institute of Forestry, Chinese Academy of Forestry, No. 1, Dongxiaofu, Xiangshan Road, Haidian District, Beijing 100091, People's Republic of China.
Data availability statement
The data that support the findings of this study are openly available in NCBI at https://www.ncbi.nlm.nih.gov/, and reference numbers are listed in Tables S1 and S2 available as Supplementary data at Tree Physiology Online.
Figure 3.
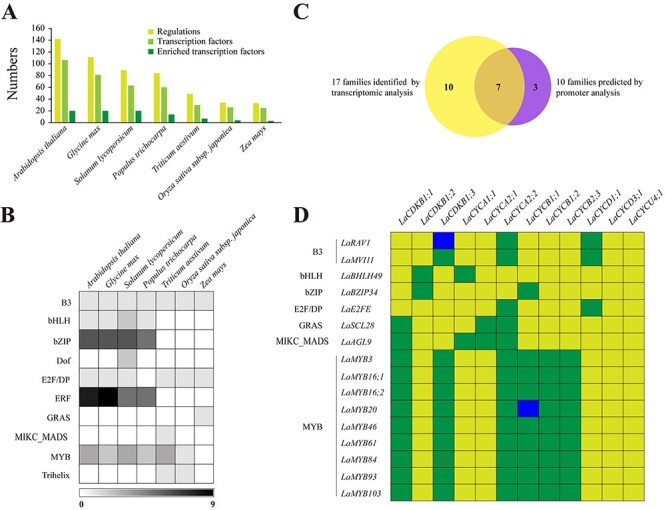
Prediction and verification of transcriptional regulators of 12 differentially expressed cell-cycle genes. (A) Cases of regulation, transcription factors and enriched transcription factors with over-represented targets in the input gene set under the cutoff P-value ≤0.05 predicted using the data from seven plant species: Arabidopsis thaliana, Glycine max, Solanum lycopersicum, Populus trichocarpa, Triticum aestivum, Oryza sativa subsp. japonica and Zea mays, with the PlantRegMap server (http://plantregmap.cbi.pku.edu.cn/regulation_prediction.php; Jin et al. 2017). (B) Summary of the enriched transcription factors from 10 families. (C) Venn analysis of transcription factor families identified by prediction and transcriptomic analysis. (D) Seventy-three cases of regulation (highlighted in green and blue) between 16 transcription factors and 12 expressed cell-cycle genes, both of which were strongly expressed in the active stage of Larix kaempferi, two of which (highlighted in blue) were confirmed by yeast one-hybrid and dual-luciferase assays.
Funding
The National Natural Science Foundation of China (Grant/Award Number: 31770714); the Basic Research Fund of Research Institute of Forestry (Grant/Award Number: RIF2014-07); and the National Transgenic Major Program (Grant/Award Number: 2018ZX08020-003).
Authors’ contributions
W.-F.L. conceived, designed, and carried out the study, analyzed the data, and wrote the manuscript. Y.K. conducted the quantitative reverse transcription polymerase chain reaction assay and statistical analysis. Y.Z. cloned the full-length cDNA sequences of transcription factors. Q.-L.Z. conducted the bimolecular fluorescent complementation and dual-luciferase assays. L.-W.Q. provided suggestions on the experimental design and analysis.
Conflict of interest
The authors declare no conflicts of interest.
References
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410. [DOI] [PubMed] [Google Scholar]
- Anh Tuan P, Bai S, Saito T, Imai T, Ito A, Moriguchi T (2016) Involvement of EARLY BUD-BREAK, an AP2/ERF transcription factor gene, in bud break in Japanese pear (Pyrus pyrifolia Nakai) lateral flower buds: expression, histone modifications and possible target genes. Plant Cell Physiol 57:1038–1047. [DOI] [PubMed] [Google Scholar]
- Azeez A, Zhao YC, Singh RK et al. (2021) EARLY BUD-BREAK 1 and EARLY BUD-BREAK 3 control resumption of poplar growth after winter dormancy. Nat Commun 12:1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Begum S, Kudo K, Rahman MH et al. (2018) Climate change and the regulation of wood formation in trees by temperature. Trees 32:3–15. [Google Scholar]
- Berckmans B, De Veylder L (2009) Transcriptional control of the cell cycle. Curr Opin Plant Biol 12:599–605. [DOI] [PubMed] [Google Scholar]
- Brunner AM, Evans LM, Hsu CY, Sheng X (2014) Vernalization and the chilling requirement to exit bud dormancy: shared or separate regulation? Front Plant Sci 5:732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busov V, Carneros E, Yakovlev I (2016) EARLY BUD-BREAK1 (EBB1) defines a conserved mechanism for control of bud-break in woody perennials. Plant Signal Behav 11:e1073873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Druart N, Johansson A, Baba K et al. (2007) Environmental and hormonal regulation of the activity-dormancy cycle in the cambial meristem involves stage-specific modulation of transcriptional and metabolic networks. Plant J 50:557–573. [DOI] [PubMed] [Google Scholar]
- Espinosa-Ruiz A, Saxena S, Schmidt J, Mellerowicz E, Miskolczi P, Bakó L, Bhalerao RP (2004) Differential stage-specific regulation of cyclin-dependent kinases during cambial dormancy in hybrid aspen. Plant J 38:603–615. [DOI] [PubMed] [Google Scholar]
- Fischer M, Müller GA (2017) Cell cycle transcription control: DREAM/MuvB and RB-E2F complexes. Crit Rev Biochem Mol Biol 52:638–662. [DOI] [PubMed] [Google Scholar]
- Forzani C, Aichinger E, Sornay E, Willemsen V, Laux T, Dewitte W, Murray JAH (2014) WOX5 suppresses CYCLIN D activity to establish quiescence at the center of the root stem cell niche. Curr Biol 24:1939–1944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galindo González LM, el Kayal WALID, Ju CJT, Allen CCG, King-Jones S, Cooke JEK (2012) Integrated transcriptomic and proteomic profiling of white spruce stems during the transition from active growth to dormancy. Plant Cell Environ 35:682–701. [DOI] [PubMed] [Google Scholar]
- Geng P, Zhang S, Liu J, Zhao C, Wu J, Cao Y, Fu C, Han X, He H, Zhao Q (2020) MYB20, MYB42, MYB43, and MYB85 regulate phenylalanine and lignin biosynthesis during secondary cell wall formation. Plant Physiol 182:1272–1283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabherr MG, Haas BJ, Yassour M et al. (2011) Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat Biotechnol 29:644–652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hachez C, Ohashi-Ito K, Dong J, Bergmann DC (2011) Differentiation of Arabidopsis guard cells: analysis of the networks incorporating the basic helix-loop-helix transcription factor, FAMA. Plant Physiol 155:1458–1472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haga N, Kato K, Murase M et al. (2007) R1R2R3-Myb proteins positively regulate cytokinesis through activation of KNOLLE transcription in Arabidopsis thaliana. Development 134:1101–1110. [DOI] [PubMed] [Google Scholar]
- Haga N, Kobayashi K, Suzuki T et al. (2011) Mutations in MYB3R1 and MYB3R4 cause pleiotropic developmental defects and preferential down-regulation of multiple G2/M-specific genes in Arabidopsis. Plant Physiol 157:706–717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han H, Sun X, Xie Y, Feng J, Zhang S (2014) Transcriptome and proteome profiling of adventitious root development in hybrid larch (Larix kaempferi × Larix olgensis). BMC Plant Biol 14:305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han S, Qi X, Sugihara K, Dang JH, Endo TA, Miller KL, Kim E, Miura T, Torii KU (2018) MUTE directly orchestrates cell-state switch and the single symmetric division to create stomata. Dev Cell 45:303–315. [DOI] [PubMed] [Google Scholar]
- He S, Xie Y, Sun X, Zhang S (2020) Comparative transcriptome analyses reveal candidate genes regulating wood quality in Japanese larch (Larix kaempferi). J Forestry Res 31:65–73. [Google Scholar]
- Hochberg Y (1988) A sharper Bonferroni procedure for multiple tests of significance. Biometrika 75:800–802. [Google Scholar]
- Hu X, Yang J, Li C (2015) Transcriptomic response to nitric oxide treatment in Larix olgensis Henry. Int J Mol Sci 16:28582–28597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito M, Araki S, Matsunaga S, Itoh T, Nishihama R, Machida Y, Doonan JH, Watanabe A (2001) G2/M-phase-specific transcription during the plant cell cycle is mediated by c-Myb-like transcription factors. Plant Cell 13:1891–1905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang C, Rao G (2020) Insights into the diversification and evolution of R2R3-MYB transcription factors in plants. Plant Physiol 183:637–655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin J, Tian F, Yang D, Meng Y, Kong L, Luo J, Gao G (2017) PlantTFDB 4.0: toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res 45:D1040–D1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jokipii-Lukkari S, Sundell D, Nilsson O, Hvidsten TR, Street NR, Tuominen H (2017) NorWood: a gene expression resource for evo-devo studies of conifer wood development. New Phytol 216:482–494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jokipii-Lukkari S, Delhomme N, Schiffthaler B, Mannapperuma C, Prestele J, Nilsson O, Street NR, Tuominen H (2018) Transcriptional roadmap to seasonal variation in wood formation of Norway spruce. Plant Physiol 176:2851–2870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlberg A, Bako L, Bhalerao RP (2011) Short day–mediated cessation of growth requires the downregulation of AINTEGUMENTALIKE1 transcription factor in hybrid aspen. PLoS Genet 7:e1002361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato K, Gális I, Suzuki S et al. (2009) Preferential up-regulation of G2/M phase-specific genes by overexpression of the hyperactive form of NtmybA2 lacking its negative regulation domain in tobacco BY-2 cells. Plant Physiol 149:1945–1957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko J, Prassinos C, Han K (2006) Developmental and seasonal expression of PtaHB1, a Populus gene encoding a class III HD-Zip protein, is closely associated with secondary growth and inversely correlated with the level of microRNA (miR166). New Phytol 169:469–478. [DOI] [PubMed] [Google Scholar]
- Kobayashi K, Suzuki T, Iwata E et al. (2015) Transcriptional repression by MYB3R proteins regulates plant organ growth. EMBO J 34:1992–2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Komaki S, Sugimoto K (2012) Control of the plant cell cycle by developmental and environmental cues. Plant Cell Physiol 53:953–964. [DOI] [PubMed] [Google Scholar]
- Kuzmin DA, Feranchuk SI, Sharov VV, Cybin AN, Makolov SV, Putintseva YA, Oreshkova NV, Krutovsky KV (2019) Stepwise large genome assembly approach: a case of Siberian larch (Larix sibirica Ledeb). BMC Bioinformatics 20:37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langmead B, Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2. Nat Methods 9:357–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li B, Dewey CN (2011) RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 12:323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li W, Han S, Qi L, Zhang S (2014) Transcriptome resources and genome-wide marker development for Japanese larch (Larix kaempferi). Front Agric Sci Eng 1:77. [Google Scholar]
- Li WF, Cui KM, He XQ (2009a) Regulation of cell cycle regulators by environmental signals during growth-dormancy cycle of trees. Plant Signal Behav 4:959–961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li WF, Ding Q, Chen JJ, Cui KM, He XQ (2009b) Induction of PtoCDKB and PtoCYCB transcription by temperature during cambium reactivation in Populus tomentosa Carr. J Exp Bot 60:2621–2630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li WF, Yang WH, Zhang SG, Han SY, Qi LW (2017) Transcriptome analysis provides insights into wood formation during larch tree aging. Tree Genet Genomes 13:19. [Google Scholar]
- Liu Y, Han S, Ding X, Li X, Zhang L, Li W, Xu H, Li Z, Qi L (2016) Transcriptome analysis of mRNA and miRNA in somatic embryos of Larix leptolepis subjected to hydrogen treatment. Int J Mol Sci 17:1951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lloyd AD, Little CHA, Mellerowicz EJ, Riding RT (1996) Changes in nuclear genome size and relative ribosomal RNA gene content in cambial region cells of Abies balsamea shoots during the development of dormancy. Can J Bot 74:290–298. [Google Scholar]
- Locosselli GM (2018) The cambium activity in a changing world. Trees 32:1–2. [Google Scholar]
- Ma Q, Dai X, Xu Y et al. (2009) Enhanced tolerance to chilling stress in OsMYB3R-2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol 150:244–256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matías-Hernández L, Aguilar-Jaramillo AE, Marín-González E, Suárez-López P, Pelaz S (2014) RAV genes: regulation of floral induction and beyond. Ann Bot 114:1459–1470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mellerowicz EJ, Riding RT, Little CHA (1989) Genomic variability in the vascular cambium of Abies balsamea. Can J Bot 67:990–996. [Google Scholar]
- Mellerowicz EJ, Riding RT, Little C (1992) Periodicity of cambial activity in Abies balsamea. II. Effects of temperature and photoperiod on the size of the nuclear genome in fusiform cambial cells. Physiol Plant 85:526–530. [Google Scholar]
- Mellerowicz EJ, Riding RT, Greenwood MS (1995) Nuclear and cytoplasmic changes associated with maturation in the vascular cambium of Larix laricina. Tree Physiol 15:443–449. [DOI] [PubMed] [Google Scholar]
- Men L, Yan S, Liu G (2013) De novo characterization of Larix gmelinii (Rupr.) Rupr. transcriptome and analysis of its gene expression induced by jasmonates. BMC Genomics 14:548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menges M, De Jager SM, Gruissem W, Murray JAH (2005) Global analysis of the core cell cycle regulators of Arabidopsis identifies novel genes, reveals multiple and highly specific profiles of expression and provides a coherent model for plant cell cycle control. Plant J 41:546–566. [DOI] [PubMed] [Google Scholar]
- Mishima K, Fujiwara T, Iki T, Kuroda K, Yamashita K, Tamura M, Fujisawa Y, Watanabe A (2014) Transcriptome sequencing and profiling of expressed genes in cambial zone and differentiating xylem of Japanese cedar (Cryptomeria japonica). BMC Genomics 15:219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno-Cortés A, Hernández-Verdeja T, Sánchez-Jiménez P, González-Melendi P, Aragoncillo C, Allona I (2012) CsRAV1 induces sylleptic branching in hybrid poplar. New Phytol 194:83–90. [DOI] [PubMed] [Google Scholar]
- Moreno-Cortés A, Ramos-Sánchez JM, Hernández-Verdeja T et al. (2017) Impact of RAV1-engineering on poplar biomass production: a short-rotation coppice field trial. Biotechnol Biofuels 10:110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikolayeva O, Robinson MD (2014) edgeR for differential RNA-seq and ChIP-seq analysis: an application to stem cell biology. Methods Mol Biol 1150:45–79. [DOI] [PubMed] [Google Scholar]
- Peng L, Skylar A, Chang PL, Bisova K, Wu X (2014) CYCP2;1 integrates genetic and nutritional information to promote meristem cell division in Arabidopsis. Dev Biol 393:160–170. [DOI] [PubMed] [Google Scholar]
- Plomion C, Leprovost G, Stokes A (2001) Wood formation in trees. Plant Physiol 127:1513–1523. [PMC free article] [PubMed] [Google Scholar]
- Qiu Z, Wan L, Chen T, Wan Y, He X, Lu S, Wang Y, Lin J (2013) The regulation of cambial activity in Chinese fir (Cunninghamia lanceolata) involves extensive transcriptome remodeling. New Phytol 199:708–719. [DOI] [PubMed] [Google Scholar]
- Rohde A, van Montagu M, Inzé D, Boerjan W (1997) Factors regulating the expression of cell cycle genes in individual buds of Populus. Planta 201:43–52. [Google Scholar]
- Schrader J, Moyle R, Bhalerao R, Hertzberg M, Lundeberg J, Nilsson P, Bhalerao RP (2004) Cambial meristem dormancy in trees involves extensive remodelling of the transcriptome. Plant J 40:173–187. [DOI] [PubMed] [Google Scholar]
- Scofield S, Jones A, Murray JAH (2014) The plant cell cycle in context. J Exp Bot 65:2557–2562. [DOI] [PubMed] [Google Scholar]
- Singh RK, Maurya JP, Azeez A, Miskolczi P, Tylewicz S, Stojkovic K, Delhomme N, Busov V, Bhalerao RP (2018) A genetic network mediating the control of bud break in hybrid aspen. Nat Commun 9:4173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skirycz A, Radziejwoski A, Busch W et al. (2008) The DOF transcription factor OBP1 is involved in cell cycle regulation in Arabidopsis thaliana. Plant J 56:779–792. [DOI] [PubMed] [Google Scholar]
- Sozzani R, Cui H, Moreno-Risueno MA et al. (2010) Spatiotemporal regulation of cell-cycle genes by SHORTROOT links patterning and growth. Nature 466:128–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sundell D, Street NR, Kumar M et al. (2017) AspWood: high-spatial-resolution transcriptome profiles reveal uncharacterized modularity of wood formation in Populus tremula. Plant Cell 29:1585–1604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Triozzi PM, Ramos-Sánchez JM, Hernández-Verdeja T, Moreno-Cortés A, Allona I, Perales M (2018) Photoperiodic regulation of shoot apical growth in poplar. Front Plant Sci 9:1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umeda M, Aki SS, Takahashi N (2019) Gap 2 phase: making the fundamental decision to divide or not. Curr Opin Plant Biol 51:1–6. [DOI] [PubMed] [Google Scholar]
- Vanneste S, Coppens F, Lee E et al. (2011) Developmental regulation of CYCA2s contributes to tissue-specific proliferation in Arabidopsis. EMBO J 30:3430–3441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weimer AK, Matos JL, Sharma N, Patell F, Murray JAH, Dewitte W, Bergmann DC (2018) Lineage- and stage-specific expressed CYCD7;1 coordinates the single symmetric division that creates stomatal guard cells. Development 145:v160671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wisniewski M, Norelli J, Artlip T (2015) Overexpression of a peach CBF gene in apple: a model for understanding the integration of growth, dormancy, and cold hardiness in woody plants. Front Plant Sci 6:85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie Z, Lee E, Lucas JR, Morohashi K, Li D, Murray JAH, Sack FD, Grotewold E (2010) Regulation of cell proliferation in the stomatal lineage by the Arabidopsis MYB FOUR LIPS via direct targeting of core cell cycle genes. Plant Cell 22:2306–2321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang K, Wang H, Xue S, Qu X, Zou J, Le J (2014) Requirement for A-type cyclin-dependent kinase and cyclins for the terminal division in the stomatal lineage of Arabidopsis. J Exp Bot 65:2449–2461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Cao Y, Xu HQ, Gao L, Guo X, Liu XQ, Zhang L, Zhang XX, Wang DY (2018) TADEA-PCR is a highly efficient method of amplifying unknown flanking fragments of T-DNA transformants. Physiol Plant 164:242–250. [DOI] [PubMed] [Google Scholar]
- Yordanov YS, Ma C, Strauss SH, Busov VB (2014) EARLY BUD-BREAK 1 (EBB1) is a regulator of release from seasonal dormancy in poplar trees. Proc Natl Acad Sci USA 111:10001–10006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, Zhou W, Chen Q, Fang M, Zheng S, Scheres B, Li C (2018) Mediator subunit MED31 is required for radial patterning of Arabidopsis roots. Proc Natl Acad Sci USA 115:E5624–E5633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Zhang S, Han S, Li X, Qi L (2012) Transcriptome profiling and in silico analysis of somatic embryos in Japanese larch (Larix leptolepis). Plant Cell Rep 31:1637–1657. [DOI] [PubMed] [Google Scholar]
- Zhang Y, Zang QL, Qi LW, Han SY, Li WF (2020) Effects of cutting, pruning, and grafting on the expression of age-related genes in Larix kaempferi. Forests 11:218. [Google Scholar]
- Zhao X, Han X, Wang Q, Wang X, Chen X, Li L, Fu X, Gao D (2020) EARLY BUD BREAK 1 triggers bud break in peach trees by regulating hormone metabolism, the cell cycle, and cell wall modifications. J Exp Bot 71:3512–3523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong Y, Mellerowicz EJ, Lloyd AD, Leinhos V, Riding RT, Little CHA (1995) Seasonal variation in the nuclear genome size of ray cells in the vascular cambium of Fraxinus americana. Physiol Plant 93:305–311. [Google Scholar]
- Zhou W, Lozano-Torres JL, Blilou I, Zhang X, Zhai Q, Smant G, Li C, Scheres B (2019) A jasmonate signaling network activates root stem cells and promotes regeneration. Cell 177:942–956.e14. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data that support the findings of this study are openly available in NCBI at https://www.ncbi.nlm.nih.gov/, and reference numbers are listed in Tables S1 and S2 available as Supplementary data at Tree Physiology Online.
Figure 3.
Prediction and verification of transcriptional regulators of 12 differentially expressed cell-cycle genes. (A) Cases of regulation, transcription factors and enriched transcription factors with over-represented targets in the input gene set under the cutoff P-value ≤0.05 predicted using the data from seven plant species: Arabidopsis thaliana, Glycine max, Solanum lycopersicum, Populus trichocarpa, Triticum aestivum, Oryza sativa subsp. japonica and Zea mays, with the PlantRegMap server (http://plantregmap.cbi.pku.edu.cn/regulation_prediction.php; Jin et al. 2017). (B) Summary of the enriched transcription factors from 10 families. (C) Venn analysis of transcription factor families identified by prediction and transcriptomic analysis. (D) Seventy-three cases of regulation (highlighted in green and blue) between 16 transcription factors and 12 expressed cell-cycle genes, both of which were strongly expressed in the active stage of Larix kaempferi, two of which (highlighted in blue) were confirmed by yeast one-hybrid and dual-luciferase assays.