

Abstract
Exposome factors that lead to stressed skin can be defined as any disturbance to homeostasis from environmental (meteorological factors, solar radiation, pollution or tobacco smoke) and/or internal exposure (unhealthy diet, hormonal variations, lack of sleep, psychosocial stress). The clinical and biological impact of chronic exposome effects on skin functions has been extensively reviewed, whereas there is a paucity of information on the impact of short‐term acute exposure. Acute stress, which would typically last minutes to hours (and generally no more than a week), provokes a transient but robust neuroendocrine‐immune and tissue remodelling response in the skin and can alter the skin barrier. Firstly, we provide an overview of the biological effects of various acute stressors on six key skin functions, namely the skin physical barrier, pigmentation, defences (antioxidant, immune cell‐mediated, microbial and microbiome maintenance), structure (extracellular matrix and appendages), neuroendocrine and thermoregulation functions. Secondly, we describe the biological and clinical effects on adult skin from individual exposome factors that elicit an acute stress response and their consequences in skin health maintenance. Clinical manifestations of acutely stressed skin may include dry skin that might accentuate fine lines, oily skin, sensitive skin, pruritus, erythema, pale skin, sweating, oedema and flares of inflammatory skin conditions such as acne, rosacea, atopic dermatitis, pigmentation disorders and skin superinfection such as viral reactivation. Acute stresses can also induce scalp sensitivity, telogen effluvium and worsen alopecia.
Introduction
The skin is one of the largest and most diverse barrier organs of the human body with the epidermis constituting the first line of defence against environmental stressors, e.g. meteorological factors (extreme heat, cold, humidity), solar radiation including ultraviolet radiation (UVR), pollution or tobacco smoke. At the same time, the skin is also affected by internal stressors, e.g. an unhealthy diet, hormonal variations, lack of sleep and psychosocial stress. Together, these challenges to the skin homeostasis constitute the skin exposome, a term which refers to the totality of exposures to such non‐genetic factors encountered by an individual over their lifetime. 1 , 2 The clinical and biological impact of chronic exposome aggressions on skin functions has been extensively reviewed, 1 , 2 , 3 , 4 , 5 , 6 whereas there is a paucity of information on the immediate effects of short‐term acute exposure, which is the aim of this review.
Exposome factors that lead to acutely stressed skin can be defined as any acute disturbance to homeostasis after environmental and/or internal exposure. Acute stress, which would typically last minutes to hours (and generally no more than a week in humans), thereby provokes a transient but robust response. This response from the key skin functions, including the skin barrier, pigmentation, defences (biochemical and immune/cellular), structure (extracellular matrix and skin appendages), neuroendocrine and thermoregulation functions, is aimed at protection or rapid elimination of the disturbance and return to homeostasis. In the present review, focussing on adult skin, we describe individual exposome factors that elicit an acute stress response and their corresponding impact on the key skin functions.
Skin functions affected by acute stress responses
Acute environmental and internal stressors can affect several skin functions. An overview of six key skin functions and their role in acute stress responses is shown in Fig. 1. Upon exposure to acute stress, these skin functions are coordinated by a transient activation of multiple biological mechanisms, some of which will be specific to different stressors, whereas others will be common to all of the stressors we review, as discussed below.
The epidermal physical barrier is the main mechanical defence against extrinsic factors including toxic damage, allergens and microbes, while it is also responsible for maintaining stratum corneum hydration by preventing unregulated trans‐epidermal water loss (TEWL). 7 , 8 , 9 The initial step in the repair response to an acute stressor is rapid secretion (within minutes) of the contents of the lamellar bodies from the outer stratum granulosum cells. 10 Rapidly acting skin barrier disruption recovery mechanisms include greater epidermal cell proliferation and lipogenesis, and increased adhesion molecule expression. 11
Skin pigmentation protects the basal keratinocytes from UV‐induced DNA damage. 12 The UVB‐induced DNA damage in keratinocytes promotes the activation of the p53 protein that binds the pro‐opiomelanocortin (POMC) promoter and ultimately induces the secretion of alpha‐melanocyte‐stimulating hormone (α‐MSH) that stimulates the MC1R melanocortin receptor on the melanocytes and activates the melanogenesis. 13 High energy visible light directly triggers pigmentation by stimulating a specific receptor called Opsin 3 at the melanocyte membrane. 14
The skin´s defence mechanisms are aimed at damage control. The biochemical defences to acute stress include antioxidant response, DNA repair and cellular osmolyte strategies. 15 An increase in reactive oxygen species (ROS) can induce the expression of matrix metalloproteinases (MMP) and promote the degradation of collagen, which can be attenuated by antioxidants.
Figure 1.
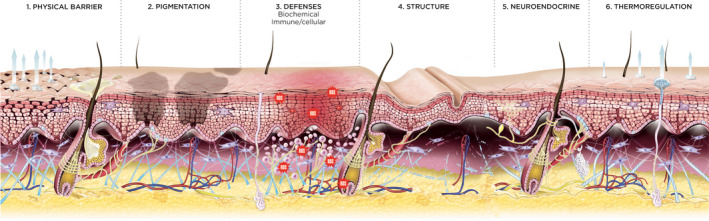
Skin functions affected by acute stressors. 1. The skin physical barrier or stratum corneum, consisting of differentiated keratinocytes (corneocytes) encased in lipid lamellae and tight junctions, is important for lipid synthesis, skin hydration and desquamation that play a role in skin dryness, tightness and skin sensitivity. 2. Melanocytes and interacting keratinocytes have a key role in skin pigmentation. 3. Skin biochemical defences have a role in antioxidant response, osmolyte strategy, DNA repair and pain, while immune/cellular defences have a role in skin immunity, including inflammation (e.g. in exacerbation of psoriasis, atopic dermatitis, seborrhoeic dermatitis, acne), Langerhans cells, decreased immunity with risk of superinfection and alteration of the skin microbiome (e.g. reactivation of viral infections under psychological stress and UV radiation). 4. The skin structure including the extracellular matrix and adnexa (hair follicles, sebaceous glands, sweat glands) plays a role in hypersudation and development of telogen effluvium. 5. Skin neuroendocrine delivery by vasculature and innervation involves local production of neurotransmitters, neurotrophins, neuropeptides, hormones with a role in the neurogenic inflammation (e.g. pain and pruritus), triggering the HPA axis, increased endocrine, vegetative and neuropeptidergic excitability levels. 6. The thermoregulation function involving blood vessels and fat has a role in vasoconstriction and flushes (e.g. rosacea)
Immune cell‐mediated defences are coordinated with the neuroendocrine response. 16 , 17 Acute stress primarily includes the innate immune system of the skin, involving antimicrobial peptides, Langerhans cells (LC), mast cells, monocytes and granulocytes as well as the epidermal keratinocytes and structural cells present deeper in the dermis, such as endothelial cells and fibroblasts. 18 Among them, the LC in the epidermis coordinate the role of stressor recognition. Acute stress additionally triggers the recruitment of natural killer (NK) cells, phagocytic cells, basophils and neutrophils into the dermis by the release of pro‐inflammatory cytokines. Immune activating damage‐associated molecular patterns (DAMPs), e.g. alarmins, activate Toll‐like receptor (TLR) signalling, 19 and increased production of pro‐inflammatory cytokines such as interleukin‐1 (IL‐1) β and tumour necrosis factor (TNF)‐α. 7 Mediators of subsequent acquired, adaptive innate immunity are pro‐inflammatory cytokines of T helper cell type 1 (Th1).
Skin microbiota play an integral role in the maturation and homeostatic regulation of keratinocytes and host immune networks. The skin microbiome and skin‐resident memory T cells may abrogate the immunosuppressive response following acute stress. 20 , 21
Defensive structures of the skin, especially the extracellular matrix, densely innervated and vascularized dermis and subcutis (nerve fibres), and skin appendages (hair follicles, sebaceous glands, sweat glands), provide the second line of defence from foreign intruders as well as internal damage. Sebaceous and eccrine glands secrete increased amounts of sebum and sweat in response to acute stress leading to moist, cool and slippery skin.
A heightened neuroendocrine‐immune response to acute stressors (e.g. heat, trauma, infestations) clinically manifests as erythema, oedema and hypersensitive responses such as pruritus or pain. The skin is one of the most densely innervated organs of the body. 22 , 23 , 24 Acute stress activates the sympathetic axis of the autonomous nervous system (SA), which reacts very quickly and leads to a transient release of adrenaline from the adrenals and a local release of noradrenaline from peripheral adrenergic nerve fibres. The SA triggers activation of the endocrine hypothalamus–pituitary–adrenal axis (HPA), resulting in a transient release of cortisol from the adrenals into the blood stream, while the release of corticotrophin‐releasing hormone (CRH), adrenocorticotropic hormone (ACTH) and cortisol from skin cells form a local equivalent of the central HPA. 25 Skin homeostasis is maintained by the additional activation of the cholinergic axis of the autonomic nervous system (CA). In addition, neuropeptides are either released from sensory nerve fibres or produced locally. Substance P (SP) and calcitonin gene related peptide (CGRP) are the main neuropeptides that modulate immediate‐type skin hypersensitivity reactions. This neuronal neuropeptidergic axis (NNA) response to acute stress regulates multiple tissue remodelling and inflammatory processes, some of which are acutely inflammatory such as mast cell activation and subsequent neurogenic inflammation, 18 , 26 , 27 while others down‐regulate inflammatory processes. Acute stressors can induce the neurotrophic factors such as artemin, nerve growth factor (NGF) or brain derived neurotrophic factor (BDNF) which contribute to hyper‐innervation, allokinesis and inflammation via, e.g. the aryl hydrocarbon receptor (AhR) and neurotrophin receptors. Acute stress triggers peripheral vasoconstriction by activating transient receptor potential (TRP) channels associated with neurogenic inflammation causing pale skin as well as rapid mobilization of immune cell trafficking into the skin. 28 , 29 It also leads to the release of melatonin, which contributes to immunomodulation, thermoregulation and tumour control.
Blood vessels and fat have a role in sympathetically mediated vasoconstriction and flushing under acute stress, causing a rapid drop in skin temperature. 30
Biological and clinical effects of distinct skin stressors
Acute solar radiation
Acute solar radiation not only stimulates pigmentation and induces DNA damage but also promotes oxidative stress, inflammation (e.g. sunburn, photosensitivity, photoallergy, flares of photodermatoses), decreased immunity against microbial challenges (e.g. herpes labialis photo‐immunosuppression with viral reactivation), Koebner phenomenon after acute sunburn, barrier function alteration, osmolyte strategy and microbiome alteration.
The damage response effect of UVR on the skin immune system has recently been reviewed. 19 The first‐line defence of UV‐induced oxidative stress is the acute activation of the oxidative pentose phosphate pathway to increase NADPH production, which is essential to prevent oxidative damage. 31 Exposure to UVB (290–315 nm) or UVA (340–400 nm) radiation significantly stimulates osmolyte uptake to protect cells against oxidative stress. 15 , 32 UVR can compromise epidermal barrier function causing skin dryness and enabling the penetration of bacteria and allergens. 33 Immediate short‐term response to UVR‐induced damage is mediated by the innate immune system of the skin and involves epidermal keratinocytes, melanocytes, LC, dermal endothelial cells, fibroblasts, mast cells, dendritic cells (DC), resident lymphocytes and neural elements, with subsequent recruitment of myeloid cell types, such as neutrophils, monocytes and macrophages. 19 In a mouse model, a single high dose of UV was shown to produce a deep inflammatory state characterized by the production of pro‐inflammatory cytokines and chemokines. 34 Sunburn from acute UVB exposure is characterized by epidermal cell necrosis, decreased antigen presentation and acute inflammation. UVR induces the release of DAMPs that activate TLR signalling. 19 Acute UVR rapidly activates skin‐resident T cells through mechanisms involving the release of ATP from keratinocytes to limit DNA damage in keratinocytes. UVR induces the epidermal recruitment of DC that compensate for the depletion of LC in human skin. 35 The effector functions of T cells depend on the activation state of LC by UVR. In response to UVB exposure, Treg cells are induced to maintain skin homeostasis and participate in epithelial stem cell differentiation of hair follicle cycles. 36 , 37 , 38 The skin microbiome may abrogate the immunosuppressive response following acute UV exposure. 21 Alternatively, direct UV‐induced DNA and membrane damage to the microbiome may result in pathogen associated molecular patterns that interfere with UV‐induced immune suppression. 20 In addition, acute UV exposure causing flushes and vasodilatation can decrease blood pressure. 39
Beneficial effects of acute UV exposure include synthesis of vitamin D, release of opioid factors and decrease in pain. 40 , 41 , 42
Acute solar radiation exposure leads to tanning and endothelial cells and fibroblasts activate melanocytes to induce pigmentation, which may cause postinflammatory hyperpigmentation (PIHP), especially for darker phototypes, or trigger photodermatoses, such as polymorphous light eruption. Melanocytes can protect from UV damage and contribute to the regulation of acute bursts of oxidative stress in the skin. 43
UVR, especially UVB, stimulates cutaneous neurogenic mediators that affect the central HPA and increase inflammation. 44 , 45 CRH is upregulated in human sebocytes and keratinocytes in vitro by UV. 46 In mice, cutaneous exposure to UVB rapidly stimulated systemic CRH, ACTH, β‐endorphin and corticosterone production accompanied by rapid immunosuppressive effects in splenocytes. 47 The rapid induction of immune suppression appears to be independent of the HPA axis (immunostimulation) and may be via direct neuronal activation. 47 In mice, pro‐inflammatory neuropeptides are released from skin nerve fibres in response to UV exposure. 48
UV radiation and heat (as well as cold, stress, spicy food and microbes) are trigger factors of rosacea that may modulate TLR signalling, induce ROS and enhance antimicrobial peptide and neuropeptide production. 49 , 50
The hypodermis contains a network of blood vessels that connects the local effects of UVR exposure to systemic immunosuppression effects and fever, if large body surfaces are exposed. 51 , 52 , 53 Acute UV exposure may inhibit the function of antigen‐presenting cells, induce T cells with suppressor activity and induce the release of immunosuppressive cytokines and the latter phenomenon is mainly responsible for systemic immunosuppression. 53 Acute UV total body exposure, psychosocial stress and hormonal variations can all cause a comparable mast cell activation and release of IL‐6 cytokine release with systemic effects. 54
Acute pollution
Airborne pollutants induce cutaneous oxidative stress and have been shown to damage skin barrier integrity by altering TEWL, inflammatory signalling, stratum corneum pH and the skin microbiome. Short‐term pollutant exposure has been linked to xerosis, pruritus and exacerbation of atopic dermatitis symptoms. 55
After 4 h exposure to volatile organic compounds (VOC), epidermal barrier damage was observed within 48 h in sensitized subjects with atopic eczema. 56
In an ex vivo skin model, diesel exhaust particles increased skin pigmentation, expression of pigmentation related genes and induced expression of MMP and pro‐inflammatory cytokines and these hyperpigmentation and inflammaging effects were reduced by application of an antioxidant mixture. 57
Among air pollutants, ozone is one of the most toxic due to its unstable structure and is able to initiate oxidative reactions and activate inflammatory response, leading to the onset of several skin conditions. 58 , 59 In vivo and in vitro studies have shown that short‐term acute exposure to ozone impacts skin defences by production of ROS, biomolecule oxidation (lipid peroxidation and protein carbonylation), depletion of cellular antioxidant defences, cell stress and cytotoxicity. 1 , 60 , 61 , 62 Ozone can induce inflammasome activation in a redox dependent manner in a mouse model, which may play a role in pollution‐induced inflammatory skin conditions. 63 , 64 When human forearm skin was exposed to ozone for 2 h, vitamin E decreased 70% with a concomitant increase in lipid hydroperoxides and a 50% decrease in the residual skin microflora in the superficial stratum corneum without producing a visible clinical response. 65 Ozone reacts with skin lipids and squalene peroxidation by‐products cause cytotoxic, pro‐inflammatory, immunological events and may lead to irritation, comedones and inflammatory acne. 66 Toxic effects of ozone are mediated through free radical reactions, leading to lipid peroxidation. In a clinical study in which skin was exposed ozone (0.8 ppm three times daily for 5 days), skin biopsies showed significant increases in α‐β unsaturated aldehyde 4‐hydroxynonenal and 8‐iso‐prostaglandin‐F(2a) protein adducts, while topical application of vitamin C appeared to prevent this oxidative modification of proteins. 60
Short‐term exposure to NO2 or VOC caused significantly increased TEWL in both healthy individuals and those with atopic dermatitis. 56 , 67 A time‐series study showed increased outpatient visits for acne vulgaris in Beijing when there was high air pollution (particulate matter [PM]10, PM2. 5, SO2, NO2). 68 Various organic components of pollutants interact with the AhR in keratinocytes to elicit an epidermal hyper‐innervation via induction of the neurotrophic factor artemin that causes nerve growth hypersensitivity, pruritus and an atopic dermatitis pathology. 69 Furthermore, retrospective time‐series studies on large populations of patients showed a relationship between a rising incidence of emergency department visits for urticaria and atopic dermatitis with an increased ambient level of ozone, 70 PM and SO2. 71 An effect of air pollution and meteorological factors (temperature and humidity) on the number of hospital outpatient visits for atopic dermatitis was also observed. 72
Combined acute challenges of UV and pollution
UV may act synergistically with particulate matter, causing an acute skin response with increased tissue peroxidation and decreased cutaneous α‐tocopherol causing additive oxidative stress in the stratum corneum. 73 , 74 Ozone, PM and UV radiation synergistically increased oxidative stress and oxinflammation changes in human skin explants. 59 , 64
Abrupt meteorological changes (humidity and temperature)
Dry environmental conditions can markedly enhance epidermal structure and function. 75 In hairless mice exposed 1–2 weeks in a dry environment (<10% relative humidity [RH]), TEWL was significantly lower, while epidermal hyperplasia, lamellar body secretory system and lamellar membranes were all increased, and barrier recovery was accelerated when compared to a humid environment (>80% RH). 75 A clinical study on dry facial skin found a higher dryness score with low temperatures, high wind speed and low humidity, and 15 min of exposure to cold and dry air led to a reduction in skin hydration. 76 These data suggest that lower temperatures lead to a decrease in skin hydration and TEWL and that this effect is stronger at low RH. Furthermore, a study to evaluate the effect of RH on the facial skin of Japanese volunteers observed lower skin conductance, lower elasticity and increased mean area of fine wrinkles after 30 min at low humidity (40% RH) compared to higher humidity (70% RH, all P < 0.05). 77
In an ex vivo study, stratum corneum water content increased 50% in vivo and pliability of skin when the temperature was raised from 20 to 35°C at RH below 60%. 78
Low humidity and low temperatures decrease skin barrier function and increase susceptibility towards mechanical stress, while the skin also becomes more reactive towards skin irritants and allergens as pro‐inflammatory cytokines and cortisol are released by keratinocytes, and the number of dermal mast cells increases. 79 Cold and dry weather appear to increase the prevalence and risk of flares in patients with atopic dermatitis. 67 , 79 However, cold alone for short periods of time (six cycles of 4°C for 90 s) did not affect TEWL or skin irritation. 80
Acute exposure to heat can cause erythema ab igne, a reticulated, hyperpigmentation of the skin. 81
Acute psychosocial stress
While chronic stress generally leads to pathogenic immune responses, 82 , 83 acute stress may induce a defensive response coordinated by a momentary and transient activation of the multiple stress response systems including activation of pro‐inflammatory mediators with immune‐enhancing effects. Acute psychosocial stress induces activation of the SA, HPA and NNA, triggering vasoconstriction, neurogenic inflammation and pro‐inflammatory mediator release and subsequently the anti‐inflammatory CA in an attempt to maintain homeostasis. 16 , 30 , 82 Inflammasome activation, upregulation of NK cell activity and upregulated release of Th1 cytokines via peripheral SA activation and via sensory nerves can protect against acute infectious agents as well as skin cancers; 83 this may contribute to better control of viral infections. 84 In mice, short‐term restrain stress before UV exposure also led to greater cutaneous T‐cell attracting chemokine, IFN‐γ gene expression and higher infiltrating T cell numbers. 85 Of note, physical pain (3‐min cold pressor pain stimulus) can cause acute psychological stress. Subjects who reported higher pain showed faster skin barrier recovery, and greater increase in norepinephrine (but not cortisol for HPA activation) was also associated with faster recovery and mediated the impact of pain on skin barrier repair. 86 , 87 The immune‐enhancing effect of acute stress is hence homeostatic. However, if the stress is intense or buffering resources low, the heightened neuroendocrine‐immune response becomes toxic and clinically manifests as erythematous rashes, oedema, pruritus or intense pain. Low neuropeptide oxytocin levels and high pro‐inflammatory cytokines are associated with both stress and pain, which may explain how psychological distress affects pain at skin level in patients with traumatic stress symptoms from burn wounds. 88
Acute psychosocial stress may negatively affect skin structure by inhibiting hair growth under the influence of the HPA via cortisol release in addition to a SP‐mast cell pathway. 18 , 89 Increased oxidative stress and redox impairment due to psychosocial stress could affect levels of pro‐inflammatory cytokines, as reported after short‐term (minutes) stress when students were preparing for an examination. 90
Autoimmune diseases, such as alopecia areata and vitiligo, may be triggered by acute stress with altered innate and adaptive responses and increased oxidative stress. 91 Acute emotional stress may precipitate alopecia areata by activation of overexpressed type 2β CRH receptors around the hair follicles leading to intense local inflammation. 92
Acute psychosocial stress may reactivate skin infectious diseases (herpes zoster, herpes labialis, herpes genitalis) via SA activation leading to increased vulnerability to infectious diseases due to skin barrier impairment and immune defence, as well as modified microbiota. 93
Acute emotional distress could lead to increased levels of glucocorticoids (GC) and androgens inducing increased sebum production in acne as well as increasing production and release of CRH from dermal nerves and sebocytes. 94 , 95 , 96 Increases in pro‐inflammatory cytokines, SP and lipids due to stress may also contribute to aggravation of acne. 97 , 98 , 99 Accordingly, in 22 subjects with acne vulgaris, severity was aggravated by emotional stress (evaluated 3 days before and 7 days after an examination). 100
Acute psychosocial stress, sleep deprivation and nutritional factors all have an effect on epidermal barrier integrity, host immune response and neurogenic factors, which can result in worsening of seborrhoeic dermatitis. 101 In psoriasis, stress promotes acute inflammation, driven by TNF‐α and epithelial hyperplasia through the SA and NNA. 102 Adults with atopic dermatitis show blunted HPA responsiveness to acute stress but hyperreactivity of the SA, 103 and an association between onset or flare of atopic dermatitis lesions and psychosocial stress has been observed. 104
If acute psychosocial stress persists and becomes chronic, it can cause severe and long‐lasting health problems in the case of chronification, as reviewed elsewhere. 16 , 105 Briefly, this stimulates a persistent increase in endogenous GCs that compromises permeability barrier homeostasis, stratum corneum cohesion, wound healing and epidermal innate immunity in normal skin. Stress and skin barrier injury then lead to better penetration of pathogenic microbes and increase vulnerability to cutaneous infectious diseases, such as superficial viral infections, mycosis and impetigo. This maladaptive state of the brain–skin connection may underlie inflammatory skin diseases caused or aggravated by stress, e.g. acne, rosacea, atopic dermatitis and psoriasis. 49 , 100 , 102
Acute sleep deprivation
Sleep loss results in an elevation of cortisol levels the next evening. 106 Acute total sleep deprivation significantly increases stress‐related hormones (with dysregulation of the HPA and activation of the NNA), making it difficult to differentiate between effects of stress and acute sleep deprivation. 107
Acute stress and one night of sleep deprivation may cause skin barrier impairment 7 that could aggravate skin dryness, intensify itch and worsen atopic dermatitis. 108 , 109 , 110
Sleep deprivation may increase oxidative stress and release of ROS. 111 Circadian imbalance could elevate levels of several potential somnogenic cytokines, including TNF‐α, interleukin (IL)‐10 and C‐reactive protein, that could be related to cortisol dysregulation due to poor sleep. 107 An impact on the immune system from lack of sleep could manifest as autoimmune diseases. 112 In Caucasian women (56 women aged 25–35 and 55 women aged 55–65 years old), fatigue from a working day induced mild changes in facial signs (infraorbicular dark circles) and slightly accentuated wrinkles. 113 In another study, in Chinese women (aged 20–40 years old), fatigue induced dull and tired‐looking skin and these signs were more pronounced in the older women aged 31–40 years old. 114
Acute sleep deprivation induced changes in thermoregulation in rats resulting in a decreased peripherical surface temperature due to SA activation during acute stress. 115 However, two nights of sleep deprivation with or without energy restriction did not impair the thermal response to cold in human subjects. 116 , 117
Acute nutrition/alcohol intake
Certain foods and dietary patterns can trigger acute changes that lead to visible skin effects. For example, consumption of alcohol, hot beverages, spicy food, capsaicin and cinnamaldehyde activate TRP channels, contributing to facial erythema and rosacea. 118 , 119 , 120 Changes in sebaceous gland composition have been documented after 5 to 7 days of fasting. In one trial, human subjects showed a marked change in forehead skin lipids, with suppression of sebaceous gland synthesis of all lipids (apart from squalene). 121
Other acute effects include biochemical and cellular changes, hormonal changes, changes in the gut microbiome and inflammatory cytokine effects. These acute changes may all impact skin disease, either directly or indirectly, even if clinical lesions will not necessarily be acutely visible. For example, large shifts in the gut microbiome have been documented to occur within 24 h, with potential implications on skin innate immunity and inflammation. 122
In acne, three major dietary components have been studied for their clinical impacts. These include hyperglycaemic carbohydrates, dairy products and certain patterns of fat consumption, including increases in saturated and trans fats and fewer ω‐3 polyunsaturated fatty acids (PUFAs). 123 In the cascade of events, cytokine production is acutely triggered by diet and the cellular changes occur acutely (inflammation, keratinocyte proliferation, hyperseborrhoea), as do hormonal effects, although clinically apparent acne lesions may not be acutely visible. Diet‐mediated changes include an increase in sebum production as well as a change in sebum composition. 124 This promotes the overgrowth of Cutibacterium acnes and increases levels of free palmitate. Free palmitate stimulates an inflammatory cascade, with resulting increases in IL‐1β, Th17 differentiation and IL‐17‐mediated keratinocyte proliferation. 123 , 125 This cascade contributes to the visible sebofollicular inflammation in acne vulgaris. 123
Hormonal changes have also been documented. In human subjects, a 7‐day controlled feeding trial reported short‐term effects of a low glycaemic load diet, suggesting that increases in dietary glycaemic load may increase the biological activity of sex hormones and IGF‐1. 126 Another randomized controlled crossover trial documented a significant increase in acne lesions (14.8 lesions) in a group consuming chocolate versus a jelly bean group (−0.7 lesions, P < 0.0001), when evaluated 48 h later. 127 The authors noted that dairy was a confounding factor, but hypothesized that chocolate components modulating cytokine production led to inflammation. 127
Food allergies are another example of acute effects and may worsen atopic dermatitis 128 , 129 , 130 due to immediate‐type IgE‐mediated hypersensitivity (rarely in adults; reactions occur within hours), systemic contact dermatitis (reactions typically occur 24–48 h later) 131 or delayed eczematous reactions (24–48 h later). 132
Acute hormonal variations
The skin is itself an endocrine organ, and all its components are constantly regulated by hormones. 133 The main hormones that affect the skin include sex hormones (oestrogens, progesterone, androgens), neuroendocrine hormones (GC, CRH, melatonin) and others (thyroid, growth hormones). 133 , 134 , 135 , 136 , 137 , 138
Acute postpartum hormonal variations may cause telogen effluvium. 139 Skin and hair follicles express melatonin that is a powerful antioxidant to combat ROS from acute stress responses. Oxytocin is released during labour and is a neuroendocrine mediator in human skin homoeostasis and modulates key processes which are dysregulated in atopic dermatitis such as proliferation, inflammation and oxidative stress responses. 140
Progesterone dermatitis hypersensitivity symptoms are associated with the progesterone surge during the luteal phase of the menstrual cycle or after exposure to exogenous progestins. 141 Hormonal variations can induce stimulation of the HPA, 142 , 143 GC 144 and modulation of skin neuropeptides. 145 Premenstrual variations in hormones have been reported to cause acne flares, 94 aphthous ulcers 146 and exacerbation of atopic dermatitis symptoms. 147
Medications and procedures
Procedures such as peels, botulinum neurotoxin, soft tissue fillers, lasers and microdermabrasion may cause skin barrier disruption, inflammation, PIHP and skin superinfection. Topical retinoids can cause retinoid irritation dermatitis, skin irritation, dryness and erythema due to skin barrier alteration, especially in the first days/weeks. 148 Topical and systemic retinoids have been reported to alter sebum quantity and quality and affect the facial skin microbiome, e.g. by reducing or eradicating the anaerobe Cutibacterium acnes which is involved in the complex pathogenesis of acne vulgaris. 149 , 150 , 151 , 152 , 153 Antibiotic use for acne and acute bacterial skin infections may alter the skin microbiome. 154 , 155
Acute stress challenges to skin during the COVID‐19 pandemic
The use of face masks, gloves and repeated hand sanitization has been associated with high rates of adverse skin reactions among healthcare professionals with reports of acute and chronic dermatitis, secondary infection and aggravation of underlying skin diseases such as acne, seasonal facial dermatitis, seborrhoeic dermatitis and rosacea. 156 , 157 In a cross‐sectional study in 34 healthcare workers, TEWL, temperature and erythema were all significantly increased after 2 h of glove and mask use, indicating impaired epidermal barrier function. 158 Possible mechanisms of aggravation of acne by mask wearing include rupture of comedones induced by pressure and friction, occlusion of pilosebaceous units, microcirculation dysfunction due to long‐term pressure, and bacterial proliferation due to higher temperature and humid environment caused by expired air and perspiration. 159
How to prevent and improve stressed skin
Understanding the pathogenesis of a maladaptive stress response is essential for the development of therapeutic strategies to improve skin health during acute exposome stress exposure.
The main biological effects of these acute stressors on skin include skin barrier alteration, subclinical microinflammation, inflammation, immunosuppression, DNA damage, melanogenesis, alteration of sebum and sweat production. The main clinical consequences include skin dryness or oiliness, dullness, redness, skin sensitivity, pruritus, sweating, flares of inflammatory skin conditions, skin superinfections such as viral reactivations, skin hyperpigmentation, as well as scalp sensitivity and hair loss. Repeated acute stressors may lead to enduring clinical effects contributing to skin ageing or skin cancers.
Global prevention measures include:
Avoidance of sunburn by adequate sun protection, e.g. sunscreen use, sun avoidance, protective clothing.
Adopting a healthy lifestyle, e.g. sleeping well, eating a well‐balanced diet, acquiring stress management skills, the use of psychosocial interventions.
Reinforcing the skin physical barrier and defences against exposome factors with, e.g. antioxidants, antipollution products, probiotics, moisturizers, unsaturated fatty acids.
Conclusions
Cutaneous perturbations created by acute exposures induce responses to protect the organism and re‐establish homeostasis. The main biological effects and resulting clinical manifestations of these acute exposures to individual external and internal exposome factors are summarized in Table 1. In adult skin, any acute stress effects should be contextualized against the background of chronic exposome exposures, especially in the case of chronic diseases, as well as the genome of the individual. Further research is required to elucidate individual effects from multiple stressors and may lead to a greater understanding of clinical presentations of skin disease at different times.
Table 1.
A summary table of the main biological mechanisms and clinical effects of acute exposure to exposome factors
| Acute stress | Skin function affected | Main biological mechanisms | Clinical manifestations |
|---|---|---|---|
| Solar radiation | Barrier | TEWL | Dryness |
| Pigmentation | Melanogenesis | PIHP, tanning, melasma exacerbation, dark spots | |
| Defences | Oxidative stress, 31 DNA damage, inflammation, 19 decrease in epidermal Langerhans cells, 35 Treg expansion, 36 , 37 microbiome alteration, 21 photoimmunosuppression 19 | Sunburn, photosensitivity, photoallergy, actinic keratoses, viral reactivation, herpes labialis. Improvement of some dermatoses (psoriasis, atopic dermatitis), Koebner phenomenon, photodermatoses (e.g. polymorphous light eruption) | |
| Neuroendocrinology |
Neurogenic inflammation, 44 , 45 upregulation of CRH, 46 , 47 vitamin D synthesis, opioid release, 42 decreased blood pressure 39 |
Pruritus, hypersensitivity, atopic dermatitis, rosacea | |
| Thermoregulation function and systemic effect | Vasodilatation 49 , 50 | Fever, erythema, rosacea flushes | |
| Skin structure | Hyaluronic acid degradation from epidermis and dermis extracellular matrix degradation via oxidative stress | Dryness, wrinkles, skin laxity | |
| Pollution | Barrier |
Change in sebum, squalene peroxidation 58 |
Dryness, skin sensitivity Flares of acne, atopic dermatitis |
| Pigmentation | Pigmentation 57 | Dark spots | |
| Defences | Oxidative stress, 60 , 61 microbiome alteration, 65 pro‐inflammatory immune response | Flares of acne, atopic dermatitis | |
| Neuroendocrinology | Neurotrophic factor artemin 69 | Pruritus, flares of atopic dermatitis | |
| Pollution and ultraviolet radiation | Defences | Oxidative stress 74 , pigmentation, oxinflammation 64 |
Photoaging, dark spots and wrinkles |
| Meteorological changes | Barrier | TEWL, 75 , 78 sebum production | Dryness, oily skin and scalp, pruritus, flares/improvement of atopic dermatitis, psoriasis |
| Structure | Hypersudation | Sweat | |
| Defences | Inflammation 67 | Skin sensitivity, flares of atopic dermatitis, rosacea | |
| Psychosocial stress | Barrier | TEWL, tight junction dysfunction | Dryness, transgression of microbes, toxins, allergens |
| Structure | Piloerection, sweating, hair loss by anagen termination and telogen effluvium, alopecia | ||
| Defences | Oxidative stress, inflammation 102 , immune suppression | Flares of acne, rosacea, psoriasis, alopecia areata, vitiligo, seborrhoeic dermatitis, atopic dermatitis, skin superinfection, viral reactivation | |
| Neuroendocrinology | Neurogenic inflammation, hyper‐innervation, 87 , 88 , 90 upregulation of CRH 94 , 95 , 96 | Erythema, oedema, pruritus, pain | |
| Thermoregulation | Vasodilation, vasoconstriction | Pale skin, hypothermia, redness | |
| Sleep deprivation | Barrier | TEWL 7 | Dryness, dullness |
| Defences | Oxidative stress, 111 inflammation 107 | Pruritus, flares of psoriasis, atopic dermatitis, seborrhoeic dermatitis, acne, skin superinfection, viral reactivation | |
| Nutrition | Barrier | Lipid/ sebum production 121 , 124 | Dry skin, oily skin |
| Defences | Antioxidant, inflammation, 123 allergic reactions, 130 microbiome 122 | Acne, atopic dermatitis, systemic contact dermatitis | |
| Neuroendocrinology | Flushing, rosacea exacerbations | ||
| Hormonal variations | Skin barrier | Oily skin, dry skin | |
| Structure | Hypersudation | Telogen effluvium, androgenic alopecia | |
| Defences | Oxidative stress, melatonin (antioxidant), oxytocin 140 , inflammation 136 , 138 | Acne, atopic dermatitis, aphtous ulcers | |
| Neuroendocrinology | Stimulation of the HPA, 142 , 143 GC, 144 modulation of skin neuropeptides 145 | Progesterone dermatitis | |
| Medications and procedures | Skin barrier | Irritation, dryness and erythema | |
| Defences | Inflammation, antimicrobial response, 152 , 153 changes in microbiome | Acne, superinfection, viral reactivation | |
| Mask use, disinfectants, frequent washing | Barrier and defence | Skin temperature, sebum, TEWL 158 | Dryness, pruritus, skin sensitivity, erythema, acne and rosacea flares |
CA, cholinergic axis; CRH, corticotrophin‐releasing hormone; HPA, hypothalamus–pituitary–adrenal axis; PIHP, postinflammatory hyperpigmentation; TEWL, trans‐epidermal water loss;
Acknowledgements
Writing and editorial assistance for the preparation of this manuscript was provided by Helen Simpson, PhD, ISMPP CMPP™, of My Word Medical Writing and funded by Vichy Laboratoires.
Funding source
An advisory board and editorial assistance were supported by Vichy Laboratoires.
Conflict of interest
DK is an employee of Vichy Laboratoires (L’Oréal), and LA is an employee of L’Oréal. All other authors have received honoraria as advisory board members for Vichy Laboratoires.
References
- 1. Krutmann J, Bouloc A, Sore G, Bernard BA, Passeron T. The skin aging exposome. J Dermatol Sci 2017; 85(3): 152–161. [DOI] [PubMed] [Google Scholar]
- 2. Passeron T, Krutmann J, Andersen ML, Katta R, Zouboulis CC. Clinical and biological impact of the exposome on the skin. J Eur Acad Dermatol Venereol 2020; 34(Suppl 4): 4–25. [DOI] [PubMed] [Google Scholar]
- 3. Dreno B, Bettoli V, Araviiskaia E, Sanchez Viera M, Bouloc A. The influence of exposome on acne. J Eur Acad Dermatol Venereol 2018; 32(5): 812–819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Cecchi L, D'Amato G, Annesi‐Maesano I. External exposome and allergic respiratory and skin diseases. J Allergy Clin Immunol 2018; 141(3): 846–857. [DOI] [PubMed] [Google Scholar]
- 5. Stefanovic N, Flohr C, Irvine AD. The exposome in atopic dermatitis. Allergy 2020; 75(1): 63–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Gracia‐Cazaña T, González S, Parrado C, Juarranz Á, Gilaberte Y. Influence of the Exposome on Skin Cancer. Actas Dermosifiliogr 2020; 111(6): 460–470. [DOI] [PubMed] [Google Scholar]
- 7. Altemus M, Rao B, Dhabhar FS, Ding W, Granstein RD. Stress‐induced changes in skin barrier function in healthy women. J Invest Dermatol 2001; 117(2): 309–317. [DOI] [PubMed] [Google Scholar]
- 8. Schmuth M, Feingold KR, Elias PM. Stress test of the skin: the cutaneous permeability barrier treadmill. Exp Dermatol 2020; 29(1): 112–113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Maarouf M, Maarouf CL, Yosipovitch G, Shi VY. The impact of stress on epidermal barrier function: an evidence‐based review. Br J Dermatol 2019; 181(6): 1129–1137. [DOI] [PubMed] [Google Scholar]
- 10. Feingold KR. Thematic review series: skin lipids. The role of epidermal lipids in cutaneous permeability barrier homeostasis. J Lipid Res 2007; 48(12): 2531–2546. [DOI] [PubMed] [Google Scholar]
- 11. Bäsler K, Brandner JM. Tight junctions in skin inflammation. Pflugers Arch 2017; 469(1): 3–14. [DOI] [PubMed] [Google Scholar]
- 12. Plonka PM, Passeron T, Brenner M et al. What are melanocytes really doing all day long..? Exp Dermatol 2009; 18(9): 799–819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Cui R, Widlund HR, Feige E et al. Central role of p53 in the suntan response and pathologic hyperpigmentation. Cell 2007; 128(5): 853–864. [DOI] [PubMed] [Google Scholar]
- 14. Regazzetti C, Sormani L, Debayle D et al. Melanocytes sense blue light and regulate pigmentation through opsin‐3. J Invest Dermatol 2018; 138(1): 171–178. [DOI] [PubMed] [Google Scholar]
- 15. Warskulat U, Reinen A, Grether‐Beck S, Krutmann J, Häussinger D. The osmolyte strategy of normal human keratinocytes in maintaining cell homeostasis. J Invest Dermatol 2004; 123(3): 516–521. [DOI] [PubMed] [Google Scholar]
- 16. Peters EM. Stressed skin?–a molecular psychosomatic update on stress‐causes and effects in dermatologic diseases. J Dtsch Dermatol Ges 2016; 14(3): 233–252. quiz 53. [DOI] [PubMed] [Google Scholar]
- 17. Shepherd AJ, Downing JE, Miyan JA. Without nerves, immunology remains incomplete ‐in vivo veritas. Immunology 2005; 116(2): 145–163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Shin H, Choi SJ, Cho AR, Kim DY, Kim KH, Kwon O. Acute stress‐induced changes in follicular dermal papilla cells and mobilization of mast cells: implications for hair growth. Ann Dermatol 2016; 28(5): 600–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bernard JJ, Gallo RL, Krutmann J. Photoimmunology: how ultraviolet radiation affects the immune system. Nat Rev Immunol 2019; 19(11): 688–701. [DOI] [PubMed] [Google Scholar]
- 20. Patra V, Byrne SN, Wolf P. The skin microbiome: is it affected by UV‐induced immune suppression? Front Microbiol 2016; 7: 1235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Patra V, Wagner K, Arulampalam V, Wolf P. Skin microbiome modulates the effect of ultraviolet radiation on cellular response and immune function. iScience 2019; 15: 211–222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Pereira MR, Leite PE. The involvement of parasympathetic and sympathetic nerve in the inflammatory reflex. J Cell Physiol 2016; 231(9): 1862–1869. [DOI] [PubMed] [Google Scholar]
- 23. Buffoli B, Rinaldi F, Labanca M et al. The human hair: from anatomy to physiology. Int J Dermatol 2014; 53(3): 331–341. [DOI] [PubMed] [Google Scholar]
- 24. Roosterman D, Goerge T, Schneider SW, Bunnett NW, Steinhoff M. Neuronal control of skin function: the skin as a neuroimmunoendocrine organ. Physiol Rev 2006; 86(4): 1309–1379. [DOI] [PubMed] [Google Scholar]
- 25. Slominski AT, Zmijewski MA, Skobowiat C, Zbytek B, Slominski RM, Steketee JD. Sensing the environment: regulation of local and global homeostasis by the skin's neuroendocrine system. Adv Anat Embryol Cell Biol 2012;212:v, vii: 1–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Arck PC, Slominski A, Theoharides TC, Peters EM, Paus R. Neuroimmunology of stress: skin takes center stage. J Invest Dermatol 2006; 126(8): 1697–1704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Singh LK, Pang X, Alexacos N, Letourneau R, Theoharides TC. Acute immobilization stress triggers skin mast cell degranulation via corticotropin releasing hormone, neurotensin, and substance P: a link to neurogenic skin disorders. Brain Behav Immun 1999; 13(3): 225–239. [DOI] [PubMed] [Google Scholar]
- 28. Ince LM, Weber J, Scheiermann C. Control of leukocyte trafficking by stress‐associated hormones. Front Immunol 2018; 9: 3143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Dhabhar FS. The short‐term stress response ‐ Mother nature's mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Front Neuroendocrinol 2018; 49: 175–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Herborn KA, Graves JL, Jerem P et al. Skin temperature reveals the intensity of acute stress. Physiol Behav 2015; 152(Pt A): 225–230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Kuehne A, Emmert H, Soehle J et al. Acute activation of oxidative pentose phosphate pathway as first‐line response to oxidative stress in human skin cells. Mol Cell 2015; 59(3): 359–371. [DOI] [PubMed] [Google Scholar]
- 32. Rockel N, Esser C, Grether‐Beck S et al. The osmolyte taurine protects against ultraviolet B radiation‐induced immunosuppression. J Immunol 2007; 179(6): 3604–3612. [DOI] [PubMed] [Google Scholar]
- 33. Biniek K, Levi K, Dauskardt RH. Solar UV radiation reduces the barrier function of human skin. Proc Natl Acad Sci USA 2012; 109(42): 17111–17116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Cela EM, Friedrich A, Paz ML, Vanzulli SI, Leoni J, González Maglio DH. Time‐course study of different innate immune mediators produced by UV‐irradiated skin: comparative effects of short and daily versus a single harmful UV exposure. Immunology 2015; 145(1): 82–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Achachi A, Vocanson M, Bastien P et al. UV radiation induces the epidermal recruitment of dendritic cells that compensate for the depletion of langerhans cells in human skin. J Invest Dermatol 2015; 135(8): 2058–2067. [DOI] [PubMed] [Google Scholar]
- 36. Yamazaki S, Odanaka M, Nishioka A et al. Ultraviolet B‐induced maturation of CD11b‐type langerin(‐) dendritic cells controls the expansion of Foxp3(+) regulatory T cells in the skin. J Immunol 2018; 200(1): 119–129. [DOI] [PubMed] [Google Scholar]
- 37. Shime H, Odanaka M, Tsuiji M et al. Proenkephalin(+) regulatory T cells expanded by ultraviolet B exposure maintain skin homeostasis with a healing function. Proc Natl Acad Sci USA 2020; 117(34): 20696–20705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Ali N, Zirak B, Rodriguez RS et al. Regulatory T cells in skin facilitate epithelial stem cell differentiation. Cell 2017; 169(6): 1119–29.e11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Weller RB. Sunlight has cardiovascular benefits independently of vitamin D. Blood Purif 2016; 41(1–3): 130–134. [DOI] [PubMed] [Google Scholar]
- 40. Garmyn M, Young AR, Miller SA. Mechanisms of and variables affecting UVR photoadaptation in human skin. Photochem Photobiol Sci 2018; 17(12): 1932–1940. [DOI] [PubMed] [Google Scholar]
- 41. Juzeniene A, Moan J. Beneficial effects of UV radiation other than via vitamin D production. Dermatoendocrinol. 2012; 4(2): 109–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Fell GL, Robinson KC, Mao J, Woolf CJ, Fisher DE. Skin β‐endorphin mediates addiction to UV light. Cell 2014; 157(7): 1527–1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Panzella L, Leone L, Greco G et al. Red human hair pheomelanin is a potent pro‐oxidant mediating UV‐independent contributory mechanisms of melanomagenesis. Pigment Cell Melanoma Res 2014; 27(2): 244–252. [DOI] [PubMed] [Google Scholar]
- 44. Slominski AT, Zmijewski MA, Plonka PM, Szaflarski JP, Paus R. How UV light touches the brain and endocrine system through skin, and why. Endocrinology 2018; 159(5): 1992–2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Han M, Ban JJ, Bae JS, Shin CY, Lee DH, Chung JH. UV irradiation to mouse skin decreases hippocampal neurogenesis and synaptic protein expression via HPA axis activation. Sci Rep 2017; 7(1): 15574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Fimmel S, Glas E, Zouboulis C. Possible interaction between neuropeptides and UV light in the development of rosacea. In Zouboulis CC, Picardo M, eds. 2nd International Conference Sebaceous Gland, Acne, Rosacea and Related Disorders – Basic and Clinical Research, Clinical Entities and Treatment, 2009: 75–80. [Google Scholar]
- 47. Skobowiat C, Postlethwaite AE, Slominski AT. Skin exposure to ultraviolet B rapidly activates systemic neuroendocrine and immunosuppressive responses. Photochem Photobiol 2017; 93(4): 1008–1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Camponogara C, Brum ES, Pegoraro NS et al. Neuronal and non‐neuronal transient receptor potential ankyrin 1 mediates UVB radiation‐induced skin inflammation in mice. Life Sci. 2020; 262: 118557. [DOI] [PubMed] [Google Scholar]
- 49. Gerber PA, Buhren BA, Steinhoff M, Homey B. Rosacea: the cytokine and chemokine network. J Investig Dermatol Symp Proc 2011; 15(1): 40–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Zouboulis CC. Acne vulgaris and rosacea. In Granstein RD, Luger TA, eds. Neuroimmunology of the Skin: Basic Science to Clinical Practice, Berlin, Heidelberg, Springer, Berlin Heidelberg, 2009: 219–232. [Google Scholar]
- 51. Norval M, Halliday GM. The consequences of UV‐induced immunosuppression for human health. Photochem Photobiol 2011; 87(5): 965–977. [DOI] [PubMed] [Google Scholar]
- 52. Hanneman KK, Cooper KD, Baron ED. Ultraviolet immunosuppression: mechanisms and consequences. Dermatol Clin 2006; 24(1): 19–25. [DOI] [PubMed] [Google Scholar]
- 53. Schwarz A, Schwarz T. Molecular determinants of UV‐induced immunosuppression. Exp Dermatol 2002; 11(Suppl 1): 9–12. [DOI] [PubMed] [Google Scholar]
- 54. Kagiwada K, Chida D, Sakatani T et al. Interleukin (IL)‐6, but not IL‐1, induction in the brain downstream of cyclooxygenase‐2 is essential for the induction of febrile response against peripheral IL‐1alpha. Endocrinology 2004; 145(11): 5044–5048. [DOI] [PubMed] [Google Scholar]
- 55. Hendricks AJ, Eichenfield LF, Shi VY. The impact of airborne pollution on atopic dermatitis: a literature review. Br J Dermatol 2020; 183(1): 16–23. [DOI] [PubMed] [Google Scholar]
- 56. Huss‐Marp J, Eberlein‐Konig B, Breuer K et al. Influence of short‐term exposure to airborne Der p 1 and volatile organic compounds on skin barrier function and dermal blood flow in patients with atopic eczema and healthy individuals. Clin Exp Allergy 2006; 36(3): 338–345. [DOI] [PubMed] [Google Scholar]
- 57. Grether‐Beck S, Valois A, Brenden H et al. An antioxidant cocktail‐containing cosmetic product (Liftactive cureR) prevents air pollution‐induced skin hyperpigmentation: ex vivo Düsseldorf Pollution Skin Test. American Academy of Dermatology annual meeting; Virtual,2020, June 12‐14.
- 58. Lefebvre MA, Pham DM, Boussouira B, Bernard D, Camus C, Nguyen QL. Evaluation of the impact of urban pollution on the quality of skin: a multicentre study in Mexico. Int J Cosmet Sci 2015; 37(3): 329–338. [DOI] [PubMed] [Google Scholar]
- 59. Ferrara F, Pambianchi E, Woodby B et al. Evaluating the effect of ozone in UV induced skin damage. Toxicol Lett 2021; 338: 40–50. [DOI] [PubMed] [Google Scholar]
- 60. Valacchi G, Pecorelli A, Belmonte G et al. Protective effects of topical vitamin c compound mixtures against ozone‐induced damage in human skin. J Invest Dermatol 2017; 137(6): 1373–1375. [DOI] [PubMed] [Google Scholar]
- 61. Valacchi G, van der Vliet A, Schock BC et al. Ozone exposure activates oxidative stress responses in murine skin. Toxicology 2002; 179(1–2): 163–170. [DOI] [PubMed] [Google Scholar]
- 62. Fussell JC, Kelly FJ. Oxidative contribution of air pollution to extrinsic skin ageing. Free Radic Biol Med 2020; 151: 111–122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Ferrara F, Pambianchi E, Pecorelli A et al. Redox regulation of cutaneous inflammasome by ozone exposure. Free Radic Biol Med 2020; 152: 561–570. [DOI] [PubMed] [Google Scholar]
- 64. Ferrara F, Woodby B, Pecorelli A et al. Additive effect of combined pollutants to UV induced skin OxInflammation damage. Evaluating the protective topical application of a cosmeceutical mixture formulation. Redox Biol 2020; 34: 101481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. He QC, Tavakkol A, Wietecha K, Begum‐Gafur R, Ansari SA, Polefka T. Effects of environmentally realistic levels of ozone on stratum corneum function. Int J Cosmet Sci 2006; 28(5): 349–357. [DOI] [PubMed] [Google Scholar]
- 66. Pham DM, Boussouira B, Moyal D, Nguyen QL. Oxidization of squalene, a human skin lipid: a new and reliable marker of environmental pollution studies. Int J Cosmet Sci 2015; 37(4): 357–365. [DOI] [PubMed] [Google Scholar]
- 67. Kantor R, Silverberg JI. Environmental risk factors and their role in the management of atopic dermatitis. Expert Rev Clin Immunol 2017; 13(1): 15–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Liu W, Pan X, Vierkötter A et al. A time‐series study of the effect of air pollution on outpatient visits for acne vulgaris in Beijing. Skin Pharmacol Physiol 2018; 31(2): 107–113. [DOI] [PubMed] [Google Scholar]
- 69. Hidaka T, Ogawa E, Kobayashi EH et al. The aryl hydrocarbon receptor AhR links atopic dermatitis and air pollution via induction of the neurotrophic factor artemin. Nat Immunol 2017; 18(1): 64–73. [DOI] [PubMed] [Google Scholar]
- 70. Xu F, Yan S, Wu M et al. Ambient ozone pollution as a risk factor for skin disorders. Br J Dermatol 2011; 165(1): 224–225. [DOI] [PubMed] [Google Scholar]
- 71. Baek JO, Cho J, Roh JY. Associations between ambient air pollution and medical care visits for atopic dermatitis. Environ Res 2021; 195: 110153. [DOI] [PubMed] [Google Scholar]
- 72. Guo Q, Xiong X, Liang F et al. The interactive effects between air pollution and meteorological factors on the hospital outpatient visits for atopic dermatitis in Beijing, China: a time‐series analysis. J Eur Acad Dermatol Venereol 2019; 33(12): 2362–2370. [DOI] [PubMed] [Google Scholar]
- 73. Marrot L. Pollution and sun exposure: a deleterious synergy. mechanisms and opportunities for skin protection. Curr Med Chem 2018; 25(40): 5469–5486. [DOI] [PubMed] [Google Scholar]
- 74. Soeur J, Belaïdi J‐P, Chollet C et al. Photo‐pollution stress in skin: Traces of pollutants (PAH and particulate matter) impair redox homeostasis in keratinocytes exposed to UVA1. J Dermatol Sci 2017; 86(2): 162–169. [DOI] [PubMed] [Google Scholar]
- 75. Denda M, Sato J, Masuda Y et al. Exposure to a dry environment enhances epidermal permeability barrier function. J Invest Dermatol 1998; 111(5): 858–863. [DOI] [PubMed] [Google Scholar]
- 76. Roure R, Lanctin M, Nollent V, Bertin C. Methods to assess the protective efficacy of emollients against climatic and chemical aggressors. Dermatol Res Pract 2012; 2012: 864734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Tsukahara K, Hotta M, Fujimura T, Haketa K, Kitahara T. Effect of room humidity on the formation of fine wrinkles in the facial skin of Japanese. Skin Res Technol 2007; 13(2): 184–188. [DOI] [PubMed] [Google Scholar]
- 78. Spencer TS, Linamen CE, Akers WA, Jones HE. Temperature dependence of water content of stratum corneum. Br J Dermatol 1975; 93(2): 159–164. [DOI] [PubMed] [Google Scholar]
- 79. Engebretsen KA, Johansen JD, Kezic S, Linneberg A, Thyssen JP. The effect of environmental humidity and temperature on skin barrier function and dermatitis. J Eur Acad Dermatol Venereol 2016; 30(2): 223–249. [DOI] [PubMed] [Google Scholar]
- 80. Fluhr JW, Bornkessel A, Akengin A et al. Sequential application of cold and sodium lauryl sulphate decreases irritation and barrier disruption in vivo in humans. Br J Dermatol 2005; 152(4): 702–708. [DOI] [PubMed] [Google Scholar]
- 81. Baltazar D, Brockman R, Simpson E. Kotatsu‐induced erythema ab igne. An Bras Dermatol 2019; 94(2): 253–254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Pondeljak N, Lugović‐Mihić L. Stress‐induced interaction of skin immune cells, hormones, and neurotransmitters. Clin Ther 2020; 42(5): 757–770. [DOI] [PubMed] [Google Scholar]
- 83. Dhabhar FS. Acute stress enhances while chronic stress suppresses skin immunity. The role of stress hormones and leukocyte trafficking. Ann N Y Acad Sci 2000; 917: 876–893. [DOI] [PubMed] [Google Scholar]
- 84. Peters EMJ, Schedlowski M, Watzl C, Gimsa U. To stress or not to stress: Brain‐behavior‐immune interaction may weaken or promote the immune response to SARS‐CoV‐2. Neurobiol Stress 2021; 14: 100296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Dhabhar FS, Saul AN, Daugherty C, Holmes TH, Bouley DM, Oberyszyn TM. Short‐term stress enhances cellular immunity and increases early resistance to squamous cell carcinoma. Brain Behav Immun 2010; 24(1): 127–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Graham JE, Song S, Engeland CG. Acute pain speeds skin barrier recovery in healthy men and women. J Psychosom Res 2012; 73(6): 452–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Foley P, Kirschbaum C. Human hypothalamus‐pituitary‐adrenal axis responses to acute psychosocial stress in laboratory settings. Neurosci Biobehav Rev 2010; 35(1): 91–96. [DOI] [PubMed] [Google Scholar]
- 88. Van Loey NE, Hofland H, Vlig M et al. Associations between traumatic stress symptoms, pain and bio‐active components in burn wounds. Psychoneuroendocrinol 2018; 96: 1–5. [DOI] [PubMed] [Google Scholar]
- 89. Chebotaev DV, Yemelyanov AY, Lavker RM, Budunova IV. Epithelial cells in the hair follicle bulge do not contribute to epidermal regeneration after glucocorticoid‐induced cutaneous atrophy. J Invest Dermatol 2007; 127(12): 2749–2758. [DOI] [PubMed] [Google Scholar]
- 90. Matalka KZ. Neuroendocrine and cytokines‐induced responses to minutes, hours, and days of mental stress. Neuro Endocrinol Lett 2003; 24(5): 283–292. [PubMed] [Google Scholar]
- 91. Villasante Fricke AC, Miteva M. Epidemiology and burden of alopecia areata: a systematic review. Clin Cosmet Investig Dermatol 2015; 8: 397–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Katsarou‐Katsari A, Singh LK, Theoharides TC. Alopecia areata and affected skin CRH receptor upregulation induced by acute emotional stress. Dermatology 2001; 203(2): 157–161. [DOI] [PubMed] [Google Scholar]
- 93. Ives AM, Bertke AS. Stress hormones epinephrine and corticosterone selectively modulate herpes simplex virus 1 (HSV‐1) and HSV‐2 productive infections in adult sympathetic, but not sensory, neurons. J Virol 2017; 91(13): e00582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Zouboulis CC, Böhm M. Neuroendocrine regulation of sebocytes – a pathogenetic link between stress and acne. Exp Dermatol 2004; 13(Suppl 4): 31–35. [DOI] [PubMed] [Google Scholar]
- 95. Ganceviciene R, Böhm M, Fimmel S, Zouboulis CC. The role of neuropeptides in the multifactorial pathogenesis of acne vulgaris. Dermatoendocrinol 2009; 1(3): 170–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Zouboulis CC, Seltmann H, Hiroi N et al. Corticotropin‐releasing hormone: an autocrine hormone that promotes lipogenesis in human sebocytes. Proc Natl Acad Sci USA 2002; 99(10): 7148–7153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Albuquerque RG, Rocha MA, Bagatin E, Tufik S, Andersen ML. Could adult female acne be associated with modern life? Arch Dermatol Res 2014; 306(8): 683–688. [DOI] [PubMed] [Google Scholar]
- 98. McEwen BS, Karatsoreos IN. Sleep deprivation and circadian disruption: stress, allostasis, and allostatic load. Sleep Med Clin 2015; 10(1): 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Toyoda M, Nakamura M, Makino T, Kagoura M, Morohashi M. Sebaceous glands in acne patients express high levels of neutral endopeptidase. Exp Dermatol 2002; 11(3): 241–247. [DOI] [PubMed] [Google Scholar]
- 100. Chiu A, Chon SY, Kimball AB. The response of skin disease to stress: changes in the severity of acne vulgaris as affected by examination stress. Arch Dermatol 2003; 139(7): 897–900. [DOI] [PubMed] [Google Scholar]
- 101. Borda LJ, Wikramanayake TC. Seborrheic dermatitis and dandruff: a comprehensive review. J Clin Investig Dermatol 2015; 3(2). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Peters EM, Liezmann C, Klapp BF, Kruse J. The neuroimmune connection interferes with tissue regeneration and chronic inflammatory disease in the skin. Ann N Y Acad Sci 2012; 1262: 118–126. [DOI] [PubMed] [Google Scholar]
- 103. Buske‐Kirschbaum A, Ebrecht M, Hellhammer DH. Blunted HPA axis responsiveness to stress in atopic patients is associated with the acuity and severeness of allergic inflammation. Brain Behav Immun 2010; 24(8): 1347–1353. [DOI] [PubMed] [Google Scholar]
- 104. Seiffert K, Hilbert E, Schaechinger H, Zouboulis CC, Deter HC. Psychophysiological reactivity under mental stress in atopic dermatitis. Dermatology 2005; 210(4): 286–293. [DOI] [PubMed] [Google Scholar]
- 105. Martin SF, Esser PR, Weber FC et al. Mechanisms of chemical‐induced innate immunity in allergic contact dermatitis. Allergy 2011; 66(9): 1152–1163. [DOI] [PubMed] [Google Scholar]
- 106. Leproult R, Copinschi G, Buxton O, Van Cauter E. Sleep loss results in an elevation of cortisol levels the next evening. Sleep 1997; 20(10): 865–870. [PubMed] [Google Scholar]
- 107. Wright KP, Drake AL, Frey DJ et al. Influence of sleep deprivation and circadian misalignment on cortisol, inflammatory markers, and cytokine balance. Brain Behav Immun 2015; 47: 24–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Xerfan EMS, Tomimori J, Andersen ML, Tufik S, Facina AS. Sleep disturbance and atopic dermatitis: a bidirectional relationship? Med Hypotheses 2020; 140: 109637. [DOI] [PubMed] [Google Scholar]
- 109. Fenner J, Silverberg NB. Skin diseases associated with atopic dermatitis. Clin Dermatol 2018; 36(5): 631–640. [DOI] [PubMed] [Google Scholar]
- 110. Korkina L, Pastore S. The role of redox regulation in the normal physiology and inflammatory diseases of skin. Front Biosci (Elite Ed) 2009; 1: 123–141. [DOI] [PubMed] [Google Scholar]
- 111. Kahan V, Andersen ML, Tomimori J, Tufik S. Can poor sleep affect skin integrity? Med Hypotheses 2010; 75(6): 535–537. [DOI] [PubMed] [Google Scholar]
- 112. Gómez‐González B, Domínguez‐Salazar E, Hurtado‐Alvarado G et al. Role of sleep in the regulation of the immune system and the pituitary hormones. Ann N Y Acad Sci 2012; 1261: 97–106. [DOI] [PubMed] [Google Scholar]
- 113. Flament F, Pierre J, Delhommeau K, Adam AS. How a working day‐induced‐tiredness may alter some facial signs in differently‐aged Caucasian women. Int J Cosmet Sci 2017; 39(5): 467–475. [DOI] [PubMed] [Google Scholar]
- 114. Flament F, Qiu H, Abric A, Charbonneau A. Assessing changes in some facial signs of fatigue in Chinese women, induced by a single working day. Int J Cosmet Sci 2019; 41(1): 21–27. [DOI] [PubMed] [Google Scholar]
- 115. Landis CA, Bergmann BM, Ismail MM, Rechtschaffen A. Sleep deprivation in the rat: XV. Ambient temperature choice in paradoxical sleep‐deprived rats. Sleep 1992; 15(1): 13–20. [DOI] [PubMed] [Google Scholar]
- 116. Oliver SJ, Harper Smith AD, Costa RJ, Maassen N, Bilzon JL, Walsh NP. Two nights of sleep deprivation with or without energy restriction does not impair the thermal response to cold. Eur J Appl Physiol 2015; 115(10): 2059–2068. [DOI] [PubMed] [Google Scholar]
- 117. Esmat TA, Clark KE, Muller MD, Juvancic‐Heltzel JA, Glickman EL. Fifty‐three hours of total sleep deprivation has no effect on rewarming from cold air exposure. Wilderness Environ Med 2012; 23(4): 349–355. [DOI] [PubMed] [Google Scholar]
- 118. National Rosacea Society . Hot Sauce, Wine and Tomatoes Cause Flare‐ups, Survey Finds 2005. URL https://www.rosacea.org/rosacea‐review/2005/fall/hot‐sauce‐wine‐and‐tomatoes‐cause‐flare‐ups‐survey‐finds (last accessed 01 April 2021).
- 119. Weiss E, Katta R. Diet and rosacea: the role of dietary change in the management of rosacea. Dermatol Pract Concept 2017; 7(4): 31–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Buddenkotte J, Steinhoff M. Recent advances in understanding and managing rosacea. F1000Res 2018; 7: 1885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Downing DT, Strauss JS, Pochi PE. Changes in skin surface lipid composition induced by severe caloric restriction in man. Am J Clin Nutr 1972; 25(4): 365–367. [DOI] [PubMed] [Google Scholar]
- 122. Singh RK, Chang H‐W, Yan DI et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med 2017; 15(1): 73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Melnik BC. Linking diet to acne metabolomics, inflammation, and comedogenesis: an update. Clin Cosmet Investig Dermatol 2015; 8: 371–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Zouboulis CC, Jourdan E, Picardo M. Acne is an inflammatory disease and alterations of sebum composition initiate acne lesions. J Eur Acad Dermatol Venereol 2014; 28(5): 527–532. [DOI] [PubMed] [Google Scholar]
- 125. Choi CW, Kim Y, Kim JE et al. Enhancement of lipid content and inflammatory cytokine secretion in SZ95 sebocytes by palmitic acid suggests a potential link between free fatty acids and acne aggravation. Exp Dermatol 2019; 28(2): 207–210. [DOI] [PubMed] [Google Scholar]
- 126. Smith R, Mann N, Mäkeläinen H, Roper J, Braue A, Varigos G. A pilot study to determine the short‐term effects of a low glycemic load diet on hormonal markers of acne: a nonrandomized, parallel, controlled feeding trial. Mol Nutr Food Res 2008; 52(6): 718–726. [DOI] [PubMed] [Google Scholar]
- 127. Delost GR, Delost ME, Lloyd J. The impact of chocolate consumption on acne vulgaris in college students: a randomized crossover study. J Am Acad Dermatol 2016; 75(1): 220–222. [DOI] [PubMed] [Google Scholar]
- 128. Katta R, Schlichte M. Diet and dermatitis: food triggers. J Clin Aesthet Dermatol 2014; 7(3): 30–36. [PMC free article] [PubMed] [Google Scholar]
- 129. Boyce JA, Assa'ad A, Burks AW et al. Guidelines for the diagnosis and management of food allergy in the United States: summary of the NIAID‐Sponsored Expert Panel report. J Am Acad Dermatol 2011; 64(1): 175–192. [DOI] [PubMed] [Google Scholar]
- 130. Worm M, Forschner K, Lee HH et al. Frequency of atopic dermatitis and relevance of food allergy in adults in Germany. Acta Derm Venereol 2006; 86(2): 119–122. [DOI] [PubMed] [Google Scholar]
- 131. Baruffi FY, Venkatesh KP, Nelson KN, Powell A, Santos DM, Ehrlich A. Systemic contact dermatitis: a review. Dermatol Clin 2020; 38(3): 379–388. [DOI] [PubMed] [Google Scholar]
- 132. Breuer K, Heratizadeh A, Wulf A et al. Late eczematous reactions to food in children with atopic dermatitis. Clin Exp Allergy 2004; 34(5): 817–824. [DOI] [PubMed] [Google Scholar]
- 133. Zouboulis CC. The skin as an endocrine organ. Dermatoendocrinol 2009; 1(5): 250–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Theoharides TC, Stewart JM, Taracanova A, Conti P, Zouboulis CC. Neuroendocrinology of the skin. Rev Endocr Metab Disord 2016; 17(3): 287–294. [DOI] [PubMed] [Google Scholar]
- 135. Ganceviciene R, Graziene V, Böhm M, Zouboulis CC. Increased in situ expression of melanocortin‐1 receptor in sebaceous glands of lesional skin of patients with acne vulgaris. Exp Dermatol 2007; 16(7): 547–552. [DOI] [PubMed] [Google Scholar]
- 136. Oeff MK, Seltmann H, Hiroi N et al. Differential regulation of Toll‐like receptor and CD14 pathways by retinoids and corticosteroids in human sebocytes. Dermatology 2006; 213(3): 266. [DOI] [PubMed] [Google Scholar]
- 137. Krause K, Schnitger A, Fimmel S, Glass E, Zouboulis CC. Corticotropin‐releasing hormone skin signaling is receptor‐mediated and is predominant in the sebaceous glands. Horm Metab Res 2007; 39(2): 166–170. [DOI] [PubMed] [Google Scholar]
- 138. Schagen SK, Ganceviciene R, Krause K et al. Update on cutaneous stress: The sebocyte own corticotropin‐releasing hormone system is an amplifier of inflammation. In Wollina U, ed. COSMODERM XVI ‐ European Society for Cosmetic and Aesthetic Dermatology (ESCAD) – International Proceedings, 2011: 63–66. [Google Scholar]
- 139. Rebora A. Telogen effluvium: a comprehensive review. Clin Cosmet Investig Dermatol 2019; 12: 583–590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Deing V, Roggenkamp D, Kühnl J et al. Oxytocin modulates proliferation and stress responses of human skin cells: implications for atopic dermatitis. Exp Dermatol 2013; 22: 399–405. [DOI] [PubMed] [Google Scholar]
- 141. Buchheit KM, Bernstein JA. Progestogen hypersensitivity: heterogeneous manifestations with a common trigger. J Allergy Clin Immunol Pract 2017; 5(3): 566–574. [DOI] [PubMed] [Google Scholar]
- 142. Ziegler CG, Krug AW, Zouboulis CC, Bornstein SR. Corticotropin releasing hormone and its function in the skin. Horm Metab Res 2007; 39(2): 106–109. [DOI] [PubMed] [Google Scholar]
- 143. Zouboulis CC, Angres S. Macrophage‐activating lipopeptide‐2 and corticotropin‐releasing hormone stimulate the inflammatory signalling in human sebocytes through activation of stearoyl‐CoA desaturase and fatty acid desaturase 2. J Eur Acad Dermatol Venereol 2021; 35(2): 493–501. [DOI] [PubMed] [Google Scholar]
- 144. Nikolakis G, Zouboulis CC. Skin and glucocorticoids: effects of local skin glucocorticoid impairment on skin homeostasis. Exp Dermatol 2014; 23(11): 807–808. [DOI] [PubMed] [Google Scholar]
- 145. Jockers‐Scherübl MC, Zouboulis CC, Boegner F, Hellweg R. Is nerve growth factor a serum marker for neurological and psychiatric complications in Behcet's disease? Lancet 1996; 347(9006): 982. [DOI] [PubMed] [Google Scholar]
- 146. Zouboulis CC, Katsantonis J, Ketteler R et al. Adamantiades‐Behçet's disease: interleukin‐8 is increased in serum of patients with active oral and neurological manifestations and is secreted by small vessel endothelial cells. Arch Dermatol Res 2000; 292(6): 279–284. [DOI] [PubMed] [Google Scholar]
- 147. Kiriyama K, Sugiura H, Uehara M. Premenstrual deterioration of skin symptoms in female patients with atopic dermatitis. Dermatology 2003; 206(2): 110–112. [DOI] [PubMed] [Google Scholar]
- 148. Sator PG. Skin treatments and dermatological procedures to promote youthful skin. Clin Interv Aging 2006; 1(1): 51–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. McCoy WH, Otchere E, Rosa BA, Martin J, Mann CM, Mitreva M. Skin Ecology during Sebaceous Drought‐How Skin Microbes Respond to Isotretinoin. J Invest Dermatol. 2019; 139(3): 732–735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Ryan‐Kewley AE, Williams DR, Hepburn N, Dixon RA. Non‐antibiotic isotretinoin treatment differentially controls propionibacterium acnes on skin of acne patients. Front Microbiol 2017; 8: 1381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Kelhälä H‐L, Aho VTE, Fyhrquist N et al. Isotretinoin and lymecycline treatments modify the skin microbiota in acne. Exp Dermatol 2018; 27(1): 30–36. [DOI] [PubMed] [Google Scholar]
- 152. Lee D‐Y, Yamasaki K, Rudsil J et al. Sebocytes express functional cathelicidin antimicrobial peptides and can act to kill propionibacterium acnes. J Invest Dermatol 2008; 128(7): 1863–1866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Borrel V, Thomas P, Catovic C et al. Acne and stress: impact of catecholamines on cutibacterium acnes. Front Med (Lausanne). 2019; 6: 155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Bassetti M, Almirante B, Giamarellos‐Bourboulis EJ et al. The interplay between acute bacterial skin and skin structure infections and depression: a vicious circle of major clinical importance. Curr Opin Infect Dis 2020; 33(2): 155–165. [DOI] [PubMed] [Google Scholar]
- 155. Xu H, Li H. Acne, the skin microbiome, and antibiotic treatment. Am J Clin Dermatol 2019; 20(3): 335–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Yan Y, Chen H, Chen L et al. Consensus of Chinese experts on protection of skin and mucous membrane barrier for health‐care workers fighting against coronavirus disease 2019. Dermatol Ther 2020; 33(4): e13310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157. Lin P, Zhu S, Huang Y et al. Adverse skin reactions among healthcare workers during the coronavirus disease 2019 outbreak: a survey in Wuhan and its surrounding regions. Br J Dermatol 2020; 183(1): 190–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Montero‐Vilchez T, Martinez‐Lopez A, Cuenca‐Barrales C, Rodriguez‐Tejero A, Molina‐Leyva A, Arias‐Santiago S. Impact of gloves and mask use on epidermal barrier function in health care workers. Dermatitis 2021; 32(1): 57–62. [DOI] [PubMed] [Google Scholar]
- 159. Han C, Shi J, Chen Y, Zhang Z. Increased flare of acne caused by long‐time mask wearing during COVID‐19 pandemic among general population. Dermatol Ther 2020; 33(4): e13704. [DOI] [PMC free article] [PubMed] [Google Scholar]