

Abstract
Humankind has always been fascinated by venomous animals, as their toxic substances have transformed them into symbols of power and mystery. Over the centuries, researchers have been trying to understand animal venoms, unveiling intricate mixtures of molecules and their biological effects. Among venomous animals, Latrodectus Walckenaer, 1805 (widow spiders) have become feared in many cultures worldwide due to their extremely neurotoxic venom. The Latrodectus genus encompasses 32 species broadly spread around the globe, 14 of which occur in the Americas. Despite the high number of species found in the New World, the knowledge on these spiders is still scarce. This review covers the general knowledge on Latrodectus spp. from the Americas. We address widow spiders’ taxonomy; geographical distribution and epidemiology; symptoms and treatments of envenomation (latrodectism); venom collection, experimental studies, proteome and transcriptome; and biotechnological studies on these Latrodectus spp. Moreover, we discuss the main challenges and limitations faced by researchers when trying to comprehend this neglected group of medically important spiders. We expect this review to help overcome the lack of information regarding widow spiders in the New World.
Keywords: Latrodectus, Latrodectism, Widow spiders, Spider venom, Americas
Background
Among the members of the Araneae order, Latrodectus spp. (widow spiders) are well known for constituting a hazard to humans. The envenomation by these spiders is marked by a massive neurotransmitter release that leads to neurotoxic manifestations of high morbidity. Although Latrodectus spp. represent only 1.3% of the biodiversity of the Theridiidae family, their extremely neurotoxic venom has transformed this small group of spiders into a major symbol of araneism in different cultures worldwide. The negative emotions evoked by these animals are not limited to folk beliefs and superstitions, but also permeate the scientific field, as shown by the terms that identify the genus itself (Latrodectus = thief, bandit, aggressor, hidden, sneaky) and one of its most studied species (mactans = killer, deadly).
Widow spiders have been the subject of several studies since the 18th century, although there is still a lot to understand about these intricate animals due to the limitations found by researchers (e.g., little amounts of venom extracted, difficulty to keep the spiders in captivity, taxonomic instability, etc.). Around 44% of the validly recognized Latrodectus species can be found in the Americas, where they have been accountable for several cases of human envenomation [1, 2]. Nevertheless, little is known about the widow spiders from the American continent, as most of the early studies focused on the spiders from the Old World and there is currently no compilation of the knowledge about the American species. Therefore, this review covers the general knowledge on widow spiders, focusing on the species found in the Americas. Herein, we address widow spiders’ taxonomy; geographical distribution and epidemiology; symptoms and treatments of envenomation; venom collection, experimental studies, proteome and transcriptome; and biotechnological studies on Latrodectus spp.
Taxonomy
Theridiidae (Sundevall, 1833) is one of the ten most diverse and widely distributed spider families on the planet, comprising 124 genera and 2510 species. Members of this family, known as the cobweb spiders, are greatly diverse regarding their morphology, ecology, and behavior. These spiders are distributed in seven subfamilies: Hadrotarsinae, Spintharinae, Pholcommatinae, Argyrodinae, Anelosiminae, Theridiinae, and Latrodectinae. The latter group is composed of four genera: Asagena Sundevall, 1833; Crustulina Menge, 1868; Steatoda Sundevall, 1833; and Latrodectus Walckenaer, 1805, which is considered by many authors as some of the most dangerous spiders for humans [3-6]. The first species of this genus was described in Italy by Rossi in 1790, as Aranea 13-guttata. It was later included in the Latrodectus clade by Walckenaer, who made the first correlation between the bites of these spiders and the neurotoxic syndrome known as latrodectism [3].
Despite the small number of Latrodectus species when compared to other spider groups, the taxonomy of widow spiders is considered complex and difficult [7]. This difficulty has been attributed to a wide range of factors: the low interest of experienced arachnologists to study this group deeply; the wide geographical distribution of many species and the great variations observed among their populations; and the traditional use of morphological characteristics in the description and identification of species. This scenario leads to instability in the group’s taxonomy, a critical issue for the correct identification of widow spiders. Consequently, it is harder to conduct studies on bio-ecology and toxinology, as well as to propose strategies of environmental surveillance for public health.
Numerous initiatives have been carried out for a taxonomic organization of this important group of spiders, some with regional coverage and others almost global in scope. It is possible to observe cycles of expansion and reduction in the number of valid Latrodectus species over time; each cycle is directly influenced by the methods available at the periods when the studies were conducted [8-11].
Initially, Bettini and Marolli [3] identified three different moments of the Latrodectus taxonomic trajectory. In the first phase, the number of species was increased as a result of adopting body color as the main systematic character [8]. The adoption of genitalia morphology as a central taxonomic parameter started a second stage, characterized by the strong decrease in valid species [10]. In the third phase, the number of species increased once again, based on the association of morphological knowledge and other biological and biochemical aspects of these spiders [12, 13]. The application of molecular biology tools for phylogenetic analysis of this genus in the mid-2000s [11] is a landmark that started the fourth phase in the knowledge of the biodiversity of Latrodectus spiders, consolidating the use of multidisciplinary bio-ecological approaches in the study of widow spiders.
The tree of phylogenetic relationships proposed by Garb et al. [11] distributes the species of Latrodectus into two broad monophyletic clades: (1) the geometricus clade, comprising L. geometricus and L. rhodesiensis, both species originated in the African continent; and (2) the mactans clade, comprising L. mactans and all other species from Africa, Israel, Spain, Australia, New Zealand, and the Americas. Surprisingly, these findings revive and corroborate the hypothesis of Levi [10], who had classified most species as synonyms of L. mactans, thereby attributing a semi-cosmopolitan character to this species. Also, the little genetic variation observed among L. geometricus populations from very distant locations indicates that the dispersion and introduction of this species in new territories may have occurred very recently, suggesting the influence of human activity.
The taxonomy of American Latrodectus spp. was studied by distinct authors at different times, including widow spiders in North [14-19] and South America [9, 12, 20-26]. In March 2021, the World Spider Catalog [27] listed 32 valid species for the genus Latrodectus. Whereas L. geometricus CL Koch, 1841 is considered semi cosmopolitan, another 18 species occur throughout different regions of the world: L. cinctus Blackwall, 1865; L. dahli Levi, 1959; L. elegans Thorell, 1898; L. erythromelas Schmidt & Klaas, 1991; L. hasselti Thorell, 1870; L. hystrix Simon, 1890; L. indistinctus O. Pickard-Cambridge, 1904; L. karrooensis Smithers, 1944; L. katipo Powell, 1871; L. lilianae Melic, 2000; L. menavodi Vinson, 1863; L. obscurior Dahl, 1902; L. pallidus O. Pickard-Cambridge, 1872; L. revivensis Shulov, 1948; L. rhodesiensis Mackay, 1972; L. tredecimguttatus (Rossi, 1790); L. renivulvatus Dahl, 1902; and L. umbukwane BMOG Wright, CD Wright, Lyle & Engelbrecht, 2019.
In the Americas, 14 of these species can be found: L. geometricus CL Koch, 1841; L. antheratus (Badcock, 1932); L. apicalis Butler, 1877; L. bishopi Kaston, 1938; L. corallinus Abalos, 1980; L. curacaviensis (Müller, 1776); L. diaguita Carcavallo, 1960; L. hesperus Chamberlin & Ivie, 1935; L. mactans (Fabricius, 1775); L. mirabilis (Holmberg, 1876), L. quartus Abalos, 1980; L. thoracicus Nicolet, 1849; L. variegatus Nicolet, 1849; and L. variolus Walckenaer, 1837. Due to frequent taxonomic changes of Latrodectus spp. over the years, in this review, we respected the specific nomenclature adopted in the original sources.
Geographical distribution and epidemiology
The geographical and spatial distribution of Latrodectus spiders present great amplitude and variation [28-30]. Some species of this group have an almost exclusively endemic profile, such as L. katipo [31] and L. variolus [32]; other species have been introduced in two or more continents, like L. hesperus and L. hasselti [33-35]; there are also those with a highly invasive capacity, with semi-cosmopolitan distribution, such as L. geometricus [36-38]. This distribution is the result of the interaction of complex biological characteristics, including their reproductive efficacy associated with the invasion of new environments [39-43]; the development of semi-social behavior among members of certain populations [44-47]; dispersion models that allow a quick and widespread of spiderlings [48, 49]; tolerance and survivability within a wide thermal range [50, 51]; great variability in feeding habits [46, 52-54]; and the efficient plasticity of some species to environmental changes [15, 55-57]. This set of features results in the ability to infest and proliferate with great success even in environments strongly disturbed by humans [1, 37]. Therefore, the main barriers preventing the spread of terrestrial invertebrates [58] have little influence on the distribution limits of widow spiders, which find favorable conditions for survival and proliferation everywhere on the globe, except for Antarctica [31]. This phenomenon enhances contact with human populations and plays a great influence on the epidemiological profile of latrodectism [59-62].
Latrodectus spiders have been reported in almost all countries in the Americas, as have the accidents caused by them. Their occurrence has been confirmed all over North America, including Canada [63], the United States [64, 65], Mexico [66], the Caribbean Islands [3, 67], and the Lesser Antilles [17]. Although data on latrodectism in Central America are scarce, the presence of widow spiders in this region has been reported by some authors; Latrodectus spp. are present in Guatemala [68], Honduras [69, 70], El Salvador [71], Nicaragua [72], Costa Rica [73], and Panama [74]. In South America, spiders from this genus can be found in Ecuador [3], Galapagos Islands [75], Bolivia [76], Peru [26, 76], Venezuela [26, 77], Colombia [26], Chile [26, 76, 78], Argentina [26, 76, 79], Brazil [1, 26, 76], Paraguay [26, 76], Uruguay [26, 76], the Guianas [26, 80], and Suriname [80]. Figure 1 shows the geographical distribution of Latrodectus species in the Americas.
Figure 1. Geographical distribution of widow spiders in the Americas. The occurrence of fourteen Latrodectus species is currently recognized in the Americas. Symbols are placed on the countries where the presence of a given species has been reported. Brackets indicate an imprecise distribution across the continent. Latrodectus geometricus (white cross) has a wide distribution, occurring in many countries throughout the Americas. Latrodectus mactans (black square) is likely native to North America but has been introduced to South America. Latrodectus curacaviensis (white diamond) can be found in the Lesser Antilles and South America. Spiders from the Latrodectus genus have been reported in all the regions marked in green, although their identification at the species level is not always possible. No record of the presence of widow spiders is available for the regions marked in black (Belize and the state of Alaska, United States). This map is based on the list of Latrodectus species and their distribution available at The World Spider Catalog [27] as well as on the specialized literature on widow spiders, as mentioned before.
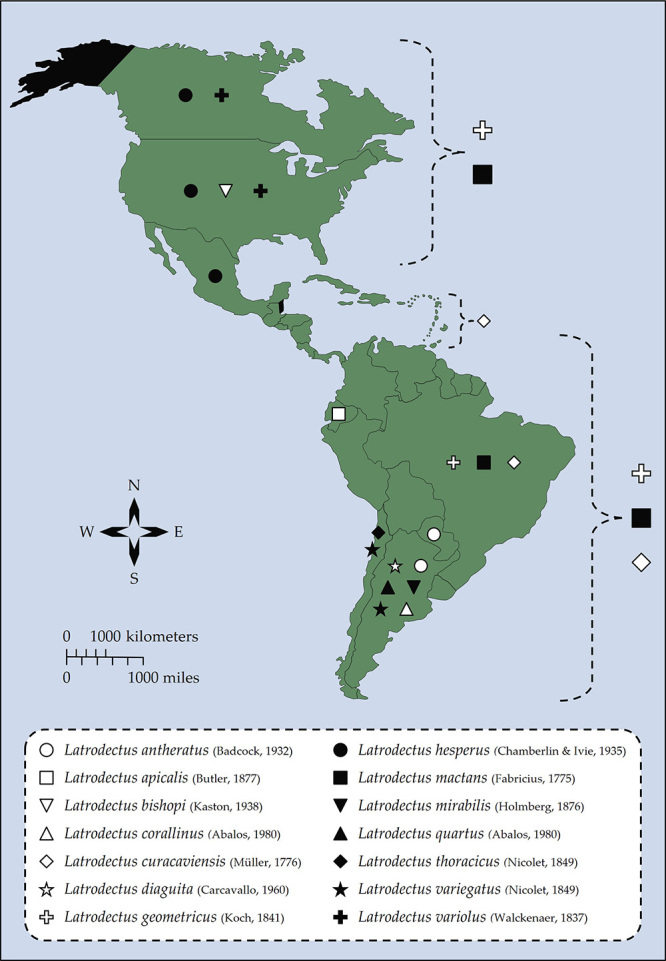
Due to the wide distribution of these spiders, latrodectism is considered a semi-cosmopolitan health grievance. Human envenomation by Latrodectus spiders can occur either by contact with isolated specimens [81] or with dense population aggregates that some species form under favorable environmental conditions [31, 59, 60, 82]. These differences influence the dynamics of accidents caused by both invasive and endemic species, which result in peculiar clinical characteristics [31, 59, 82].
The real profile and epidemiological importance of Latrodectus spider bites are still unknown for some reasons: (1) it is often difficult to accurately diagnose these accidents; (2) the taxonomic identification of the etiologic agent is infrequent [78, 81, 83, 84]; (3) these spiders exhibit non-aggressive behavior [57]; (4) the lethality rate is low and many accidents result in mild clinical manifestations [1]; (5) many accidents remain unreported due to the practice of unsupervised home treatments [64, 65, 82]; and (6) accidents are underestimated in several countries, some of which lack an official reporting system [59].
Even without precise coverage, latrodectism is usually considered one of the worst and most serious cases of araneism in the world due to its dramatic and prolonged morbidity [85]. This motivates frequent discussions on isolated cases or few reports [63, 86], epidemiological analysis of limited chronological or geographical scope [87], and the dilution of the latrodectism understanding by its inclusion in general analyses of animal envenomation [82, 88].
Bettini [59] published one of the first and broadest compilations of the epidemiological knowledge available on latrodectism. His work was based on the review of a previous study on the cases that occurred in Italy between 1946 and 1951 and on the analysis of the available literature on latrodectism epidemic outbreaks. In an attempt to understand the epidemiology of venomous bites, he discusses the biology of spiders along with environmental and human factors that correlate to envenomation. The concepts introduced by Bettini [59] were adopted and widely used in many later studies on latrodectism. His observations and inferences have been discussed and corroborated by subsequent epidemiological and biological studies [2, 37, 88].
This study also introduced the idea of possible time cycles of latrodectism, consisting of "outbreaks epidemic" and "silence" periods. During the outbreaks, more medical and biological papers were published due to the sense of emergency generated by the frequency of spider bites. The subsequent periods of silence were marked by the absence of accidents and a proportional lack of information on the subject. These cycles would often have heterogeneous behaviors, lasting for periods as short as a few weeks or longer than a decade. This pattern, which is influenced by biological, environmental, and human conditions, contributes to the difficulty of establishing a robust epidemiological picture of latrodectism [3, 64, 82, 89].
Due to these difficulties, the vast majority of the available analyses focus primarily on pathophysiological manifestations and the evolution of clinical cases, as seen in studies from Australia (L. hasselti, 68 cases) [90], South Africa (L. indistinctus and L. geometricus, 45 cases) [86], Brazil (Latrodectus spp., 77 cases) [1], Spain (L. tredecimguttatus, 12 cases) [91], and the United States (Latrodectus spp., 163 cases) [64].
Bettini [59] has drawn attention to the heterogeneous geographical distribution of epidemic outbreaks of latrodectism, which have a certain level of “endemism”. This phenomenon is the result of many factors, such as population variations and biological requirements like temperature ranges [92], the substrate for web construction [93], types and abundance of prey [53, 94], and factors of general impact on the spiders’ ecology (infectious agents, parasites, predators, etc.) [95]. Other determinants for latrodectism identified by Bettini [59] are the density of human populations and the activities that can favor their contact with spiders. Most studies indicate latrodectism as a grievance that affects mainly the rural area, due to the positive relationship found between certain types of culture and the increase in populations of these spiders, especially in Europe [88, 89], South America [76], and Oceania [31]. However, some publications indicate the urbanization of these spiders and accidents in different countries [37, 50].
Some authors discuss the importance of phenology of Latrodectus spp. for the occurrence of accidents [46, 96]. In general, accidents tend to increase in the hottest months of the year in both tropical and temperate countries [1, 62]. However, species that colonize environments modified by humans find appropriate conditions during the whole year, so the frequency of accidents remains stable throughout the months. On the other hand, for the species found in crops, the planting and harvesting cycles play an important role in the seasonality of widow spider bites [65].
Epidemiological studies in the Americas are still scarce. Grisolia et al. [62] published an analysis about the accidents that occurred in the province of Buenos Aires (Argentina) between 1979 and 1988, with a total of 281 reported cases (an average of 28 accidents per year). Because the accidents were related to the rural environment and men were more likely to work on crops than women, males (80%) were more affected by the spider bites than females (20%). The number of cases was higher in the hottest months of the year (November to March), which differed from what was observed in the province of Santiago del Estero, in the north of Argentina, where the cases concentrated from March to May [97].
Schenone and Correa [51] performed an analysis of 150 cases of latrodectism that occurred during the summer between 1983 and 1984 in various regions of Chile. In this study, men aged 10 to 39 years were more affected by these accidents, which occurred mostly between 10 a.m. and 7 p.m.
In Brazil, the retrospective study by Lira-da-Silva et al. [1] points out the relevance of latrodectism in the city of Salvador (state of Bahia). The analysis of the accident reports from 1980 to 1990 revealed a total of 77 cases of latrodectism. The main species involved in the accidents was L. curacaviensis, which proliferates in ravines, coconut shells, crevices of stones, gardens, and interiors of homes. The majority (70%) of the victims were men between 10 and 29 years old. Importantly, this study shows that envenomed people who received specific antivenom had a shorter hospital stay than those who had not. Moreover, the accidents occurred predominantly in the urban area (57%), similarly to isolated Latrodectus sp. spider bites reported in other Brazilian states [98, 99].
Clinical symptoms
Maretić [4] reported similarities in the clinical symptoms of the envenomation caused by distinct species of widow spiders, such as L. mactans, L. hesperus, L. variolus, and L. bishop (the Americas); L. tredecimguttatus (Eurasia); L. cinctus, L. indistinctus, and L. menavodi (Africa); L. hasselti (Oceania); and the semi-cosmopolitan L. geometricus. For this reason, in this review, we described the general clinical symptoms of latrodectism using reports from the American continent and comparing them, when necessary, with reports from elsewhere.
The venom of Latrodectus spp. is a complex mixture of components, composed mostly of proteins and peptides. These components play several biological roles, such as paralyzing, immobilizing, killing, liquefying prey, and restricting competitors. Additionally, Latrodectus spp. hold toxins not only in their venom glands but also in other body parts (legs and abdomen), as well as in their eggs and spiderlings [100].
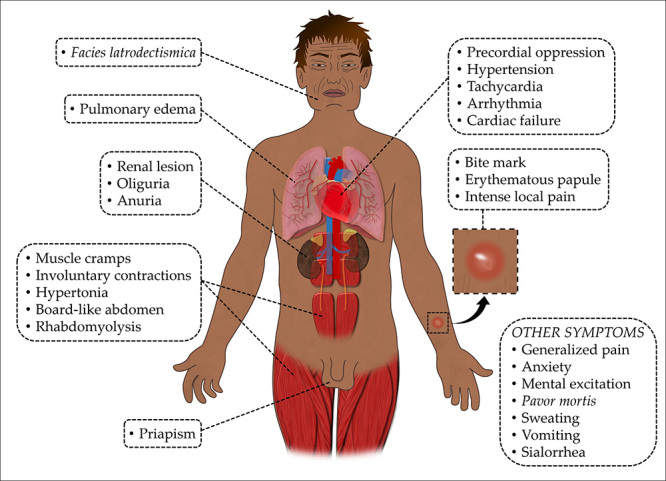
The most frequent symptom of Latrodectus bites is severe burning pain radiating from the inoculation site, and generalized muscle pain, due primarily to muscular spasm, which intensifies over time [1, 4, 64]. The neuromuscular manifestations, present in both experimental and human envenomation, are characterized by muscle cramps, involuntary contractions, and hypertonia, leading to stiffness in the abdomen and lower limbs. The facial manifestations, collectively called “facies latrodectismica”, include twisted facial muscles, blepharitis, rhinitis, cheilitis, and masseter trismus. Other symptoms include salivation, sweating, precordial oppression, anxiety, and mental excitation, which are responsible for the “pavor mortis” (fear of death) reported by the patients. Hypertension and renal alteration (oliguria) are also reported. Death is not common even without the use of serotherapy but may still occur, largely due to pulmonary edema and cardiac failure [101].
Latrodectus envenomation in humans can also result in hypertension and tachycardia. Although ECG changes may be present, damage to the myocardium is uncommon and cardiac manifestations are considered rare. In these uncommon cases, the concentration of troponin is increased, and echocardiographic changes are reported in all studies, but vary according to the report [61, 102-105]. Cardiogenic pulmonary edema that required mechanical ventilation has also been described [103]. Additionally, the venom of L. dahli from Iran causes myocytolysis, myocarditis, and coagulation necrosis of the heart under experimental conditions; the mechanisms of myocardial damage are nonetheless unclear [106]. The massive release of catecholamines is a direct toxic effect of the venom’s major component, α-latrotoxin (α-LTX), and hypersensitivity reactions have been suggested by some authors [5, 107, 108].
Rhabdomyolysis is another uncommon condition that can be observed from Latrodectus bites [102, 107]. It may be a consequence of generalized hypercontraction and intense muscle cramps that are typical of this envenomation; as a result, renal damage (oliguria and anuria) can occur. This lesion may also be caused by fluid loss (sweating, vomiting, or sialorrhea), or even by the direct action of the venom that causes paresis upon sphincter or hypercontraction at the abdominal musculature [4].
A few reports have described priapism caused by Latrodectus spp. bites [109-112]. Some authors have suggested that the mechanism involved in venom-induced priapism is probably of the high flow type. In this case, there is no ischemia since the arterial blood flow to the penis is not compromised; therefore, the priapism is not very painful and can even be painless [109]. They suggest that the exhaustion of the noradrenaline after its intense release compromises penile detumescence. Additionally, the increase in acetylcholine from cholinergic nerves causes smooth muscle relaxation and the synthesis of nitric oxide from non-adrenergic and non-cholinergic nerves that promote vasodilation [102, 111]. Figure 2 summarizes the symptoms of Latrodectus envenomation.
Figure 2. Local and systemic clinical manifestations of latrodectism.
Differential diagnosis
Understandably, for a correct management of the envenomed patient, some conditions that can be mistaken with Latrodectus bites must be recognized for a differential diagnosis. Considering their neuromuscular manifestations, the differential diagnoses include alcohol or opiate abuse, organophosphate or strychnine poisoning, tetanus, rabies, renal colic, acute abdomen conditions, appendicitis, and peritonitis. Considering the cardiovascular manifestations, latrodectism can closely mimic acute myocardial ischemia [61, 113-115]. Some authors occasionally suggest other conditions: acute intermittent porphyria, inorganic lead intoxication (Saturnine colic) [61], and food poisoning [115]. Envenomation by other animals should also be considered, especially those that cause severe pain as a major symptom. Lastly, it is also important to consider skin and soft tissue infections, allergic reactions, dermatoses, and other skin-related issues that can be misdiagnosed as widow spider bites [116].
Treatment
Several methods have been proposed for treating widow spider bites in the past. Most of these treatments tried to promote pain relief. Ancient therapies include the use of alcoholic beverages, hot baths, dancing, and even cocaine [117, 118]. However, even modern treatments are uncertain and variable [65], consisting mostly of muscle relaxants, opioids, benzodiazepines, calcium gluconate, and nonsteroidal anti-inflammatory drugs [64, 65, 119]. The efficacy of some of these treatments is nonetheless questionable, as shown by retrospective studies. In the case series by Clark et al. [64], 96% of the patients who received calcium gluconate, which has been extensively used since the mid-20th century, experienced no pain relief nor resolution of other symptoms; these patients required a subsequent administration of opioids. Moreover, the use of opioids seemed to be effective in some cases, whereas other patients were unresponsive to this treatment. Only 55% of the bitten patients had satisfying levels of analgesia with the use of opioids; this rate increased to 70% when opioids were associated with benzodiazepines [64]. On the other hand, Monte et al. [65] have found a correlation between the use of benzodiazepines and symptoms enduring longer than 24 hours.
Unlike other non-specific therapies, Latrodectus antivenom promotes a quick resolution of the envenomation symptoms with a 100% success rate [64, 65, 119]. The serotherapy is associated with effective pain relief [65, 119, 120] and a reduction in hospitalization time [64]. Additionally, antivenom is a cheap treatment, with the cost of one vial estimated to be U$ 33.25 in 2012 [119]. The standard dose for treating widow spider bites is one vial of antivenom, although some patients might require a second vial for a full resolution of symptoms [64, 120]. Despite the effectiveness of antivenom, its use has been discouraged by many health practitioners due to the fear of anaphylactic reactions [65, 118, 121, 122].
In retrospective studies performed in the United States, the rate of antivenom use in patients bitten by widow spiders is around 2.2 to 3.8% [65, 122]. The concern regarding antivenom’s safety increased after the first report of death following the use of Latrodectus antivenom in the United States by Clark et al. [64]. A second fatality was reported almost two decades later by Murphy et al. [122]. Importantly, both patients had asthma, which has been pointed out as a contraindication for antivenom [122]. The circumstances of these fatalities should be considered before discrediting the safety of Latrodectus antivenom. Moreover, most of the data used to determine the risk of Latrodectus antivenom is extrapolated from the experience with snake antivenom, which overestimates the chances of anaphylactic reaction because the anti-Latrodectus serum is given in a much smaller volume [122].
Many authors agree that antivenom is a safe and effective therapy that should be used in severe Latrodectus envenomations, although some still advise that antivenom must be used with caution and only when every other therapeutic approach has failed [64, 119, 120]. If an anaphylactic reaction does occur, the antivenom infusion must be stopped as quickly as possible and the patient must be given support therapy with antihistamines, steroids, and epinephrine [122]. When patients are not eligible for serotherapy, opioids associated with muscle relaxants are recommended [118].
Some methods have been developed aiming to have a better safety profile of antivenom. The precipitation of inactive proteins and the enzymatic cleavage of antibodies into F(ab) or F(ab)2 fragments are some strategies used to reduce the immunogenicity of horse serum-based products [120]. As new technologies arise, horse serum may eventually not be needed whatsoever. For instance, the IgY-technology is a promising new method based on the production of specific antibodies from egg yolks. One of its advantages is the low immunogenicity to mammals, making it a potential substitute for the current method [123].
Cross-reactivity studies have shown that Latrodectus antivenom is not species-specific, suggesting that any anti-Latrodectus serum could potentially be used to treat envenomations caused by any Latrodectus sp. [118, 121, 124] and other Theridiidae spiders such as Steatoda grossa [125], which also has Latrodectus-like toxins, as discussed in further detail in the section “venom components”. Consequently, the identification of the species is not clinically relevant, as all patients will respond well to antivenom. Thus, it would be interesting from the perspective of public health to discuss the production of antivenom that could be distributed and used anywhere in the Americas regardless of the species used for production.
The idea of an anti-Latrodectus serum dates to the early 1900s, when the blood of a patient who had recovered from a widow spider bite was given intramuscularly to another bitten patient, who then showed a significant improvement [117]. In 1936, Mulford Biological Laboratories of Sharp and Dohme, in the United States, produced the first commercially available Latrodectus horse serum-based antivenom [118]. Eventually, other companies from different countries made Latrodectus antivenom available, as shown in Table 1. It is worth mentioning that the Instituto Vital Brazil (Niterói, Brazil) has already registered an anti-Latrodectus serum produced against L. curacaviensis venom [126, 127], which could potentially be used in the near future to treat the accidents caused by widow spiders in the Brazilian territory.
Table 1. List of the anti-Latrodectus antivenoms produced in the world.
| Continent | Country | Producer | Species* |
|---|---|---|---|
| Oceania | Australia | Commonwealth Serum Laboratories (CSL) | L. hasselti |
| Africa | South Africa | National Health Laboratory Services (NHLS) | L. indistinctus |
| America | Argentina | Instituto Nacional de Producción de Biológicos (INPB) | L. variegatus, |
| L. antheratus, | |||
| L. diaguita, | |||
| L. corallinus, | |||
| L. mirabilis, | |||
| L. quartus | |||
| Brazil | Instituto Vital Brazil (IVB) | L. curacaviensis | |
| Mexico | Instituto Bioclon Laboratorio Silanes (BIOCLON) | L. mactans | |
| United States | Merck Sharp and Dohme International | L. mactans |
*This table considers only the licensed antivenoms and the species listed herein are indicated in the package insert or on the website of the producers.
Methods for venom collection
The small size of Latrodectus spiders resulted in the development of different techniques to obtain the small volume of venom they produce. The amount, composition, and properties of the venom obtained vary according to the extraction method. The most used techniques are crushing and washing the spider's cephalothorax followed by recovering the diluted venom; collecting venom in capillary tubes inserted in the glands’ lumen; surgically extracting the glands and homogenizing them in different buffers [128, 129]; electrically stimulating live specimens forcing venom ejection [130]; inducing spider bites on cotton or absorbent paper [131]; and stressing the specimens until they eject venom drops in capillary tubes [132]. Each of these techniques has its limitations such as the wide variation in individual responses of the extracted specimens; contamination by tissues or other body secretions; contamination by external agents; very low venom yield; and losses between processing steps [133].
Experimental studies on Latrodectus venom
The difficulties regarding Latrodectus genus taxonomy, obtaining a reasonable amount of venom, and the limited number of research groups that have systematically studied these spiders are limitations for understanding the characteristics of their venom and the effects of envenomation. There have been only a few studies regarding the effects of crude venom from widow spiders of the American continent, in isolated systems. Some of these studies are reported herein.
Toxicity
Latrodectus venom is widely known for its extreme toxicity. However, the LD50 data available in the literature are variable, considering the method of venom extraction, the test animals used, and the route of administration. Also, Latrodectus species have taxonomic inaccuracies that can compromise information.
McCrone [17] has found the following LD50 values after the administration of venom from Latrodectus spp. from the United States in mice by intraperitoneal route - L. mactans mactans: 1.3 (1.2 - 2.7) mg/kg; L. variolus: 1.8 (1.2 - 2.7) mg/kg; and L. bishop: 2.2 (1.29 - 3.74) mg/kg. De Roodt et al. [101], also using the intraperitoneal route in mice, has discovered a different range of LD50 values for spiders from different regions of Argentina - L. mirabilis: 0.155 to 0.6 mg/kg; L. diaguita: 0.31 to 1.08 mg/kg; and L. corallinus: 0.5 mg/kg 1 .
Different populations of L. geometricus, which generally causes only mild envenomation in humans, have been reported to have different LD50 values in experiments performed on mice: 0.43 mg/kg for spiders from Florida (United States) [17] and 0.225 mg/kg for spiders from Miranda (Venezuela) [134]. It is known that geographical differences can contribute to the intraspecific variability of venoms and their toxicity [135]; although there are no studies on the subject for Latrodectus spp., it is possible that variances in venom composition driven by different environmental pressures may have contributed to the discrepancy between the studies.
Cell culture experiments
Using neuronal hippocampal culture and human embryonic kidney cells (HEK 293 cells), Parodi and Romero [136] proposed that the Latrodectus venom from Chile closes potassium channels, extending the action potential similarly to tetraethylammonium (a non-selective potassium channel blocker). These changes induce a spontaneous synaptic activity in a concentration- and time-dependent manner. The effect is reversible after removing the venom; moreover, it is abolished by heating the venom at 96°C for 45 min, indicating its thermolability.
Cardiovascular system and muscle tissue
Romero et al. [137, 138] showed positive inotropic and chronotropic effects of the extract of L. mactans venom glands from Chile on the cardiac papillary muscle of rats, similarly to sympathomimetics molecules. The authors suggested that the venom induces sympathetic tonus, which could explain the vascular and cardiac symptoms of the envenomation.
In some models of isolated smooth muscle, the venom induces a sustained tonic effect related to the permeability to Na+ and Ca2+ ions that modulate the contractile response. The response is characterized by a fast-phasic component followed by a slower and more sustained tonic component probably due to the release of adrenergic and cholinergic neurotransmitters [139]. However, other studies using the smooth muscle of the urethra and esophagus have shown that α-LTX can also release non-adrenergic and non-cholinergic mediators, particularly nitric oxide and intestinal vasoactive peptide [140, 141], thereby causing relaxation.
The ultrastructural analysis of skeletal muscle injuries caused by L. geometricus venom demonstrated the presence of eosinophil infiltration, edema of the sarcotubular systems, and rupture of cell membranes. The mitochondria and nuclei were degenerate and surrounded by necrotic areas and myofibrillar disorganization [134]. The mechanisms involved in this injury have not been investigated.
Liver and kidney
The experimental injection of L. dahli venom in rabbits (0.5 mg/kg by subcutaneous route) induced an increase in the levels of alanine aminotransferase, aspartate aminotransferase, alkaline phosphatase, and bilirubin, suggesting liver damage [142]. Hepatic damage had already been reported with the presence of liver edema, massive hyperemia, and lobular necrosis [143, 144]. Moreover, the increased levels of creatinine, albumin, and urea suggest the impairment of kidney function [142]. Maretić and Stani [145] have suggested that the kidney damage caused by Latrodectus envenomation is secondary to intense dehydration due to sweating and sialorrhea.
Pain and inflammation
Although the main symptom of latrodectism is pain, only recently Lauria et al. [146] described the mechanisms involved in the nociception induced by L. curacaviensis venom from Rio de Janeiro (Brazil). This venom causes intense and heat-sensitive spontaneous nociception, mediated by serotonin and bradykinin receptors, and TRPV1 channels. It has been suggested that the presence of serotonin in spider venoms can be associated with pain production, self-protection, and the facilitation of venom uptake by increasing local blood flow and cell permeability [147]. L. curacaviensis venom also induces mechanical allodynia, which is reduced by the pharmacological inhibition of H1 histamine receptors and TRPV1 channels. Additionally, the nociceptive effect of the venom is abolished when it is heated to 90°C for 10 min, indicating the thermolability of the nociceptive toxins [146].
The inflammatory profile of Latrodectus venoms has also been poorly investigated. L. curacaviensis venom induces paw edema of low intensity and short duration in mice, mast cell degranulation, and the increase in local IL-1β levels [146]. Liver and myocardium edema have also been observed after the inoculation of L. dahli venom in rabbits [106].
Venom components
Despite their vast geographical distribution and variety of species, all spiders from the Latrodectus genus express proteins of high molecular weight that are neurotoxic to vertebrates (α-LTX), insects (latroinsectotoxin; LIT), and crustaceans (latrocrustatoxin; LCT). These proteins have acid isoelectric points, ranging from pH 5.2 to 6.38. Since these molecules share similar features and mechanisms of action, suggesting that they are related, scientists decided to group them in a family named latrotoxins [148]. It is noteworthy that most of the information about the genus was acquired using the venoms from L. m. tredecimguttatus and L. tredecimguttatus.
Longenecker et al. [149] observed that the aqueous components of venom glands macerated (ACVGM) from L. m. tredecimguttatus caused the depletion of synaptic vesicles at the neuromuscular junction (NMJ) of frogs. Frontali et al. [150] were the first to isolate and characterize the B5 fraction, which contained a 130 kDa protein that was responsible for the neurotoxicity of the venom upon vertebrates. Tzeng and Siekevitz [151] improved the purification steps and named this protein α-LTX since it was the first characterized toxin of this genus. The toxin is a homodimer with three specific regions (wing, body, and head) that assumes a tetrameric form in the presence of divalent cations, becoming active. The oligomer resembles a four-bladed propeller with a channel in the center [152, 153].
Some of the characteristics of α-LTX include the ability to fuse into lipid bilayers [154]; the evocation of a substantial and fast influx of Ca2+ ions into different cells, promoting the release of neurotransmitters such as noradrenaline, dopamine, acetylcholine (Ach), and γ-aminobutyric acid (GABA) [155]; the depletion of synaptic vesicles at the NMJ [156], on slices of the cerebral cortex of mice [151, 157], and from rat synaptosomes preparations [156]. The toxin also induces the exocytosis of different hormones: catecholamines by chromaffin cells [158]; insulin by pancreatic ß-cells [159]; and antidiuretic hormone and oxytocin by the neurohypophysis [160], but this process depends on Ca2+ [161-163]. Hence, it is possible to assume that all secretory cells are sensitive to the action of α-LTX since the molecule was able to induce the secretion of hormones and several neurotransmitters.
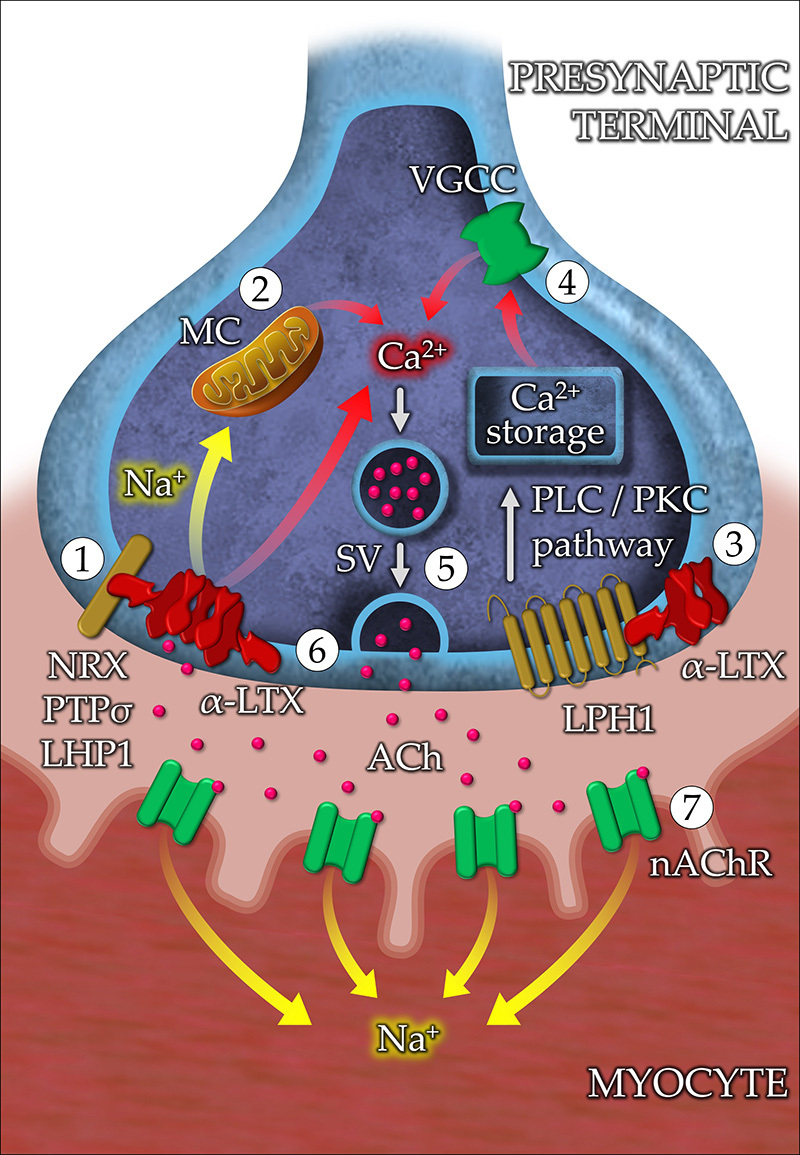
The mechanism of action of α-LTX is composed of complementary pathways: (1) the formation of a pore in the presynaptic membrane after the toxin insertion allowing cationic influx, and (2) the activation of intracellular signaling cascades that lead to a global increase in the amount of calcium (Figure 3). The interaction between the membrane receptors and α-LTX occurs through a Ca2+-dependent mechanism by neurexin-Iα (NXR-Iα) or Ca2+-independent mechanisms, by protein tyrosine phosphatase σ (PTPσ) and latrophilin 1 (LPH1), also named calcium-independent receptor of α-LTX (CIRL). The receptors interact with the toxin's upper portion, in its tetrameric form, while the base inserts itself in the membrane, permeating the lipid bilayer. These interactions facilitate the insertion of the toxin into the lipid bilayer and therefore pore formation. Additionally, the interaction between the toxin and LPH1, a G protein-coupled receptor, activates the protein kinase C, followed by the release of calcium from the endoplasmic reticulum [152, 153, 162, 164-168]. In summary, α-LTX interacts with specific receptors from neuronal and neuroendocrine terminals, resulting in a massive release of neurotransmitters based on pore formation and intracellular signaling cascades.
Figure 3. Proposed mechanisms of neurotoxicity of α-LTX in the neuromuscular junction. (1) In the presynaptic membrane, the wing region (N-terminal domain) of α-latrotoxin (α-LTX) interacts with neurexin-Iα (NXR-Iα), protein tyrosine phosphatase σ (PTPσ), and latrophilin 1 (LPH1). This interaction facilitates the insertion of the tetramer into the lipid bilayer, allowing the influx of monovalent (Li+, Cs+, Na+, K+) and divalent (Ca2+, Ba2+, Mg2+) alkali cations. (2) The excess of Na+ induces the release of more Ca2+ from the mitochondria into the cytoplasm. (3) α-LTX triggers LPH1 signaling, resulting in the activation of the phospholipase C (PLC)/protein kinase C (PKC) pathway. PLC increases the levels of inositol trisphosphate, promoting the release of intracellular Ca2+ stored in the endoplasmic reticulum. (4) The depolarization of the presynaptic terminal activates voltage-gated calcium channels (VGCC), allowing the influx of more Ca2+. (5) The massive increase in intracellular Ca2+ promotes the mobilization of synaptic vessels (SV) containing the neurotransmitter acetylcholine (Ach). (6) The release of Ach into the synaptic cleft can be either due to the fusion of SVs with the plasma membrane or due to the leakage of the neurotransmitter through the α-LTX pore. (7) The interaction of Ach with the nicotinic acetylcholine receptor (nAChR) present in the membrane of the myocyte causes depolarization and muscle contraction.
Frontali et al. [150] also demonstrated the toxicity of fraction C from ACVGM to insects, since it killed flies and blocked the heartbeat of cockroaches. Following on, Kovalevskaia et al. [169] isolated and identified a 120 kDa protein responsible for this effect and labeled it as α-latroinsectotoxin (α-LIT). This toxin displays specific characteristics: (1) the binding and fusion to membrane preparations, in a similar manner to α-LTX, but specific to insects [170]; (2) the increase of the frequency of glutamatergic miniature end-plate potentials at NMJs from insects, followed by a blockade of the synaptic transmission; (3) an absence of potentiation or antagonist action if associate to α-LTX; and (4) it is not recognized by polyclonal antibodies against α-LTX [169, 171, 172].
In addition to α-LIT, four other LIT subtypes were isolated: β-LIT (140 kDa), γ-LIT (120 kDa), ε-LIT (110 kDa), and δ-LIT (110 kDa). All subtypes were toxic to wax moths, with α-LIT being the most lethal molecule [148, 173]. Among the four subtypes, only δ-LIT had its primary sequence reported, which was shorter than those of α-LTX and α-LIT. This molecule also acts similarly to α-LIT [174].
According to Frontali et al. [150], fraction E from ACVGM is responsible for the depolarization of stretch receptors of crayfish. Fritz et al. [175] observed the specificity of this fraction against crustaceans by reporting the release of neurotransmitters in NMJs from lobsters, among other models. Grishin [148] isolated a 120 kDa protein responsible for these effects and proposed the name of α-latrocrustatoxin (α-LCT).
The paralogs α-LTX, α-LIT, δ-LIT, and α-LCT from L. m. tredecimguttatus venom are long polypeptides (1,200 to 1,400 amino acid residues) that share 30-60% of identity [148, 174, 176, 177]. These molecules have common characteristics such as moderate levels of amino acid homology (17%); proteolytic processing of both N- and C-terminal regions, by furin-like proteases, as steps of the toxin’s maturation [170, 178, 179]; and comparable domain organization. The mature toxins are composed of an N-terminal domain (with similar features to all latrotoxins, indicating an ancestral gene) and a C-terminal part (formed by two-thirds of ankyrin-like sequences that range from 13-22 repeats) [148, 171, 172, 174, 180]. Researchers believe that the ankyrin repeats, known for coordinating the interaction of several proteins, could act in the high-affinity binding between these molecules and their receptors, even though the receptors for LITs and LCT are unknown. The compilation of data about these toxins reveals a similar mechanism across phyla [172].
Other molecules described in the venoms of Latrodectus spp. are the Low Molecular Weight Proteins, also called latrodectins [181]. Latrodectin-1 (approximately 8,0 kDa) and latrodectin-2 (9,5 kDa) are acid proteins of unknown function that share structural homology and co-purify with α-LTX and LITs, respectively [181-183]. Studies demonstrate that latrodectins were derived molecules from the Crustacean Hyperglycaemic Hormone / Ion Transport Peptides superfamily that suffered positive and negative selections [184, 185]. Latrodectin-1 is present in the venom in equal amounts to α-LTX; it can be found free or forming a complex with α-LTX [183]. Studies revealed that a specific antibody against this molecule inhibited the effects triggered by α-LTX [186]. McCowan and Garb [185] proposed that latrodectin-1 could modulate the transport of calcium ions around or near nerve cells, thereby enhancing the toxicity of α-LTX. Other possible mechanisms of latrodectin-1 are stabilizing α-LTX and/or promoting its connection to the plasma membrane, facilitating the pore formation. Both hypotheses, however, have not yet been validated [162].
Some components from Latrodectus spp. venoms have been isolated or biochemically tested for their activity. In addition to latrotoxins and latrodectins, previous studies reported the presence of a variety of molecules, such as protease inhibitors and different types of enzymes (proteases and hyaluronidase) [187], and other components as part of the venom such as adenosine, guanosine, inosine, and serotonin [113, 147, 187], shown in Table 2.
Table 2. List of isolated or biochemically tested components (enzymes or inhibitors) from the venoms of Latrodectus spp.
| Molecules | Species |
|---|---|
| α-LTXs | L. m. tredecimguttatus/L. tredecimguttatus: α-LTX-Lt1a [176] |
| L. mactans: α-LTX-Lm1a [188] | |
| L. hesperus: α-LTX-Lhe1a [188] | |
| L. hasselti: α-LTX-Lh1a [188] | |
| LITs | L. m. tredecimguttatus/L. tredecimguttatus: α-LIT-Lt1a [177], |
| δ-LIT-Lt1a [174], ß-LIT, γ-LIT, and ε-LIT [148] | |
| L. hasselti: α-LIT-Lh1a and δ-LIT-Lh1a [188] | |
| L. hesperus: δ-LIT-Lhe1a [188] | |
| α-LCT | L. m. tredecimguttatus/L. tredecimguttatus: α-LCT-Lt1a [148] |
| Latrodectins | L. m. tredecimguttatus/L. tredecimguttatus [182, 183] |
| L. geometricus [185] | |
| L. hesperus [185] | |
| Peptide inhibitor of angiotensin-converting enzyme | L. tredecimguttatus [147] |
| Hyaluronidase | L. tredecimguttatus [113] |
| L. mactans [113] | |
| Collagenase | L. geometricus [134] |
| Phosphodiesterase | L. mactans (from North America*) [189] |
| Bradykinin-inactivating enzyme (kininase)-thiol endopeptidase | L. tredecimguttatus [190]** |
| Serotonin | L. tredecimguttatus [191] |
| Free amino acids (glutamic acid, aspartic acid, GABA, and taurine) | L. tredecimguttatus [3, 147] |
| Purine derivatives (adenosine, guanosine, inosine, and 2,4,6-trihydroxypurine) | L. menavodi [192] |
*Considering the great distribution and the taxonomic uncertainty of the species, the authors prefer to identify the origin of the venom when present in the original work.
**The authors reported that L. tredecimguttatus has no proteolytic activity against casein and hemoglobin. Moreover, Frontali et al. [150] did not detect lipolytic or proteolytic action of a toxic fraction of Latrodectus venom against Azocoll.
The studies of venom components from the American widow spiders are scarce and limited to some latrotoxin family members and latrodectins from L. mactans, L. hesperus, and L. geometricus. Graudins et al. [188] reported α-LTX, α-LIT, and δ-LIT molecules in the venom of the phylogenetically related spider Steatoda grossa (CL Coch, 1838) (Theridiidae). Additionally, Haney et al. [193] have described orthologous proteins from this family and other components in the venom glands from the genus Latrodectus and Steatoda. In the light of adaptive evolution, the authors reported that switches in the expression of these genes from other tissues to the venom gland associated with positive selection and gene duplication contribute to the diversity of molecules. Such evolutionary process could lead to variations in venoms’ toxicity. More recently, Dunbar et al. [194] have shown that Lactrodectus-like toxins are also present in the venom of Steatoda nobilis (Thorel, 1875), suggesting that the presence of these molecules among Theridiidae spiders could be more common than initially thought. Graudins et al. [188] have confirmed that α-LTX from L. mactans, L. hesperus, and L. hasselti elicited neurotransmitter release in the same manner as α-LTX from L. tredecimguttatus. However, the monoclonal antibody (4C4.1) against α-LTX from L. tredecimguttatus was unable to neutralize their toxicities. They deduced the sequence of α-LTX from L. hasselti by tracking the similarities regarding the number of amino acid residues, domain organizations, and proteolytic processing with α-LTX from L. tredecimguttatus. The sequences presented high identity (93%) with the presence of non-conservative substitutions at specific residues that precluded the neutralization by 4C4.1.
In another study, Parodi and Romero [136] reported the absence of α-LTX in L. mactans from Chile. The authors suggested that the latrodectins are responsible for the toxic effects observed in envenomation by this species. However, in a broader analysis with sequences of α-LTX from members of Theridiidae (Parasteatoda Archer, 1946; Steatoda; and Latrodectus), Garb and Hayashi [195] demonstrated the presence of α-LTX in venom samples from Chile and an identity of 94% among several Latrodectus species. They also declared that the occurrence of positive and negative selections in the gene α-LTX could explain the higher toxicity of this molecule in the Latrodectus genus. These findings confirm the existence of α-LTX homologs among species from the Theridiidae family. It also clarifies the cross-reactivity observed in antivenom studies [196]. McCowan and Garb [185] described latrodectins’ paralogs in the venoms of L. hesperus and L. geometricus, discussing the higher similarity of these toxins within the mactans clade and therefore the closer phylogenetic relationship between L. hesperus and L. tredecimguttatus when compared to L. geometricus (geometricus clade).
Proteome and transcriptome analyses of Latrodectus venom
Even though widow spiders are medically relevant, the number of studies that investigate and characterize the molecules of their venoms is scarce, compromising the understanding of all mechanisms involved in the envenomation. The implementation of high-throughput methods such as transcriptomic and proteomic was a breakthrough step in the development of studies related to venom. There are currently six studies that characterize the venom and/or venom glands using proteomic and/or transcriptomic approaches. Two of them describe the venoms from two American species (L. hesperus and L. mactans), whereas one reported the venom components of the semi-cosmopolitan species L. geometricus. These data reinforce the impressive lack of information about the venoms from American widow spiders.
In recent years, high-throughput methods have gained notoriety in the study of widow spiders due to the limitations regarding the amount and purity of the venom that can be obtained from widow spiders, as discussed in the section “methods for venom collection”. Researchers have employed this approach to different biological materials, amounting to ten studies to gather more information about this genus. These studies encompass eggs [197, 198], silk glands [199-204], newly hatched spiderlings [205], and different tissues from adult spiders [206]. Notably, the use of genome and/or transcriptome techniques make it easier to define the protein content of these biological tissues. Here we discuss the results of the analyses of the venom or venom glands. Table 3 presents a compilation of the different studies conducted to date using transcriptomics and proteomics for the analysis of Latrodectus spp.
Table 3. Compilation of the studies on Latrodectus spiders using proteomics and transcriptomics approaches.
| Species | Origin | Analyzed material | P/T |
|---|---|---|---|
| L. tredecimguttatus | Not informed | Eggs [198] | P |
| Not informed | Eggs [197] | T | |
| China | Newly-hatched spiderlings and adult spiders [205] | P | |
| Not informed | Venom glands (F) [207] | T | |
| China | Venom extracted from dissected venom glands (F) [129] | P | |
| China | Venom extracted by ES (F) [208] | P | |
| L. hesperus | Not informed | Silk glands (M and F) [199] | T |
| United States | Silk glands (F) [200] | P | |
| Database | Different tissue types [206] | T | |
| Database | Different tissue types [201] | T | |
| Database | Different tissue types [202] | T | |
| United States | Different tissue types (F) [203] | T | |
| United States | Venom glands, silk glands, and cephalothorax (F) [204] | T | |
| Not informed | Silk glands, cephalothorax minus venom glands, venom glands, and venom extracted by ES (F) [209] | P/T | |
| L. geometricus | United States | Different tissue types (F) [203] | T |
| Not informed | Silk glands (M and F) [199] | T | |
| Thailand | Venom extracted by the cold shock method (F) [210] | P | |
| L. mactans | Alphabiotoxine Laboratory | Venom glands and venom extracted by ES [211] | P/T |
P: proteome analysis; T: transcriptome analysis; M: male; F: female; ES: electrical stimulation.
Duan and coworkers [129, 208] conducted the initial proteomics studies using the venom from L. tredecimguttatus. In the first work, the authors obtained the sample from dissected venom glands punctured with a needle [129], whereas in the second work, the authors used electrostimulation [208]. These strategies allowed the identification of 86 and 122 molecules in each study, respectively. The authors classified these molecules according to their biological functions. The main classes were latrotoxins (α-LTX, α- and δ-LIT, and α-LCT precursors), hydrolases, other enzymes, and proteins, some of which have an unknown function. Each study, however, presented exclusive categories. In the first work, these molecules were in the cytoskeletal and related proteins class; the researchers reported the presence of contaminants, such as histiocytic proteins [129]. In the second study, the authors have described binding proteins and other specific molecules (leech-derived tryptase inhibitor trypsin complex, venom allergen antigen 5-like protein, and fucolectin) [208]. Recently, Khamtorn et al. [210] reported a partial proteome profile of L. geometricus venom using a cold shock method during the collection step. The authors described, similarly to Duan et al. [129, 208], the presence of members from the latrotoxin family, hemocyanins, proteases, and other enzymes among the venom components.
Data generated from transcriptome analyses partially overcome the issue of obtaining venom in high amounts, for it allows the large-scale production of specific toxins in gene expression systems. There are currently only three transcriptome studies using venom glands from the Latrodectus genus, and all applied next-generation sequencing, followed by de novo assembly. The pioneers were He et al. [207], who searched for novel toxins from L. tredecimguttatus, enabling the identification of 146 toxin-like proteins sequences grouped in the following categories: (1) neurotoxins (members of α-LTX-Lt1a family 1, α-LTX-Lt1a family 2, α-LIT-Lt1a, δ-LIT-Lt1a, ankyrin family, sperm-coating glycoprotein, and lycotoxin families); (2) assistant toxins (theriditoxin family); (3) proteases (trypsin family); (4) protease inhibitors (ctenitoxin family); and (5) toxins with unknown functions (scorpion toxin-like and orphan families). In a multi-tissue analysis of L. hesperus, Haney et al. [209] compared selected venom gland-specific transcripts with the venom. This strategy allowed the discovery of 62 transcripts sequences with homology to known proteins and the identification of 61 proteins; the molecules identified were classified as toxins or enzymes. In the first group of molecules, there were latrotoxins, latrodectins, inhibitor cystine knots (ICK) toxins, cysteine-rich secretory proteins (CRISPs), and leucine-rich repeats (LRR) proteins. In the second group of molecules, there were metalloproteases, serine proteases, chitinases, and hyaluronidases. Oldrati et al. [211] compared the transcripts from venom glands and the venoms of phylogenetically distant species: L. mactans and another three species belonging to different genera. The authors focused the analysis on a subgroup of molecules, the cysteine-rich peptide toxins, differing from the previous studies.
Despite their distinct origins, there is evident conservation of the molecules present in the venom glands of L. tredecimguttatus (Asia) and L. hesperus (America). The presence of these components is related to their similar envenomation symptoms and could be partially explained by the high identity among the α-LTXs sequences. In general, proteins and peptides with homology to known toxins were present in the venoms. Latrotoxins (ankyrin superfamily), latrodectins (theriditoxin members), ctenitoxins, and CRISPs were the toxins or toxin families found. Additionally, metalloproteinases, hyaluronidases, and a serine protease were also present in both venoms. However, some differences were also observed between them, for instance, the venom glands of L. tredecimguttatus contain molecules with a characteristic pattern of six highly conserved cysteines, classified as members of the scorpion toxin-like family [207]. Moreover, LRR proteins and chitinases were exclusively found in the venom of L. hesperus by Haney et al. [209]. Besides, although both species present latrotoxins, these molecules, and their motifs diverge phylogenetically [209, 212]. Haney et al. [209] reported more than 20 divergent latrotoxins paralogs, expressed in the venom glands of L. hesperus (the Americas), one of which had already been described by Graudins et al. [188]. They also identified orthologs of α-LTX, but the biological action of these molecules still needs to be tested. Whether these differences are relevant in the distinction among them (Old World vs. New World) is yet to be confirmed.
Haney et al. [209] analyzed the whole venom from L. hesperus, whereas Oldrati et al. [211] focused on the cysteine-rich peptide toxins from L. mactans. Both researchers used species found in the Americas. The authors compared the venom components (obtained by electrostimulation) with the sequences found in the transcriptome of the venom glands in an attempt to pinpoint the secreted molecules. Among these molecules, Haney et al. [209] identified latrotoxins, latrodectins, LRR proteins, ICK toxins, putative ICKs, CRISPs, and several enzymes (metalloproteases, serine proteases, hyaluronidases, and chitinase). Oldrati et al. [211] identified peptides from three distinct families as Cupiennius salei (Keyserlingi, 1877) toxins (CSTX), peptidase S1, and arthropod CHH/MIH/GIH/VIH hormone. In both studies, the researchers noticed the CSTX molecules, classified as ICK toxins, in the analysis of L. hesperus venom.
Curiously, the only similarity among the proteomic studies on the venoms of L. tredecimguttatus [129, 208], L. hesperus [209], and L. geometricus [210] is the presence of latrotoxins, latrodectins, and some enzymes. We believe that some of those differences are related to the partial proteomic profile obtained by Khamtorn et al. [210] and the different databases used in the L. tredecimguttatus and L. hesperus studies. Another difference lies in the parameters used to classify the molecules as toxins. Duan et al. [129, 208] loosely classified their molecules using generic biological functions. Haney et al. [209] used a set of defined parameters such as the presence of signal peptide and conserved domains in the sequences and the prediction of potential toxicity for the molecules; the use of these parameters was only possible due to the gain of information provided by the transcriptome analysis. No transcriptome nor genome data were available at the time Duan et al. [129, 208] performed their work.
The reasons why the envenomation varies remain unexplained. The interspecific differences of the Latrodectus genus could explain some of these variations. Even if α-LTX sequences present a high identity among Latrodectus spp., the variability (5.8%) within the mactans clade cannot be ignored [188, 195]. Also, the actions of the remaining components of the venom should not be disregarded since they lack characterization. Therefore, combining analyses of transcriptomic and proteomic data and comparing them among species is essential. Ultimately, the association of these techniques will allow a better understanding of the complexity of the venom and the mechanisms involved in the envenomation.
Beyond biological and biochemical characterization: biotechnology opportunities
The molecules from the venom and body parts of Latrodectus spp. have also been studied for their biotechnological potential and some patents have been applied/granted. For instance, a fragment of a peptide isolated from L. mirabilis from Chile has been proposed as a spermicide contraceptive drug [213]. Another example is the use of polypeptides derived from α-LTX in the treatment of erectile dysfunction [214]. However, the biotechnological uses are not restricted to these products and the possibilities are many, as shown by the studies from different fields described in the subsections below.
Insecticidal activity
Because of their feeding habits, the venom of spiders contains toxins that directly affect insects; therefore, some of these molecules can potentially be used as insecticides. A clear example is the LITs from widow spiders, as proposed by Rohou et al. [172]. Toxins from this family act specifically toward insects, either by a pore-forming mechanism or by the interaction with membrane receptors leading to neurotransmitter release. Moreover, latrodectins have been shown to increase α-LTXs activity toward insects. Nevertheless, there are limitations to the biotechnological use of LITs. Firstly, their receptors are still unknown; and secondly, LITs have high molecular weight, so they are quickly cleaved and deactivated by insects’ proteases [215]. The insecticide molecules are not restricted to the venom; a component purified from L. tredecimguttatus eggs (latroeggtoxin-III, a 36 kDa protein) showed a potent toxicity on cockroaches [216].
Antimicrobial activity
Widow spider-derived molecules have also been shown to display antimicrobial activity. Khamtorn et al. [210] have described the antibiotic action of L. geometricus venom against gram-positive bacteria. Makover et al. [217], also working with L. geometricus, have shown that components from both extract and surface of eggs play an important role in the protection of eggs against infection. Moreover, latroeggtoxin-IV, an egg toxin from L. tredecimguttatus, exerts a potent antibiotic activity against gram-positive and gram-negative bacteria; this effect is comparable to that from ampicillin, a broad-spectrum antibiotic [216]. These results altogether indicate the presence of molecules in Latrodectus spp. venom and eggs with potential application as antibiotics.
Antitumor activity
A few articles analyzed the antitumor action of components from widow spiders’ venom and eggs. The recombinant latroeggtoxin-V inhibits the proliferation and migration of breast cancer cells (MDA-MB-231) and also causes their apoptosis; the toxin inhibits the activity of the Na+/K+-ATPase in a concentration-dependent manner, suggesting that its action affects ion transport [218]. In a short communication, Mousavi et al. [219] reported that L. dahli crude venom induces cell death in a breast cancer cell line (MCF-7), possibly by apoptosis. Rivera-de-Torre et al. [220] have proposed that conjugating spider toxins with antibodies that target tumor cells could enhance the selectivity of the toxins toward the tumor. Pore-forming toxins such as α-LTX are ideal candidates for this strategy, as they deal damage to the cell without the need for internalization. Although a deeper understanding of the mechanisms of toxins and putative immunotoxin complexes is still required, this strategy could represent an advance in cancer treatment.
Novel ICK toxins from widow spider venom
As discussed in the section “proteome and transcriptome analyses of Latrodectus venom”, all of the studies performed have reported the presence of ICK-motif toxins, which in spider venoms are cystine-stabilized peptides with a specific pattern of four sequential disulfide bridges (C1-C4, C2-C5, C3-C8, C6-C7). The presence of these toxins was unprecedented in the Theridiidae family until He et al. [207] reported that L. tredecimguttatus venom contains ICK peptides with sequence homology to members of the CSTX superfamily; similar results were later found for L. hesperus [209] and L. mactans [211]. CSTX-1 has been reported to selectively block L-type Ca2+ channels in neurons [221] and to exert cytolytic activity on both prokaryotic and eukaryotic cells [222]. Molecules that are structurally related to CSTX-1, such as the novel ICK peptides from Latrodectus spp., are likely to promote similar biological activities. ICK toxins can act as hemolytic, antiviral, and antibacterial agents as well as ion channel blockers [223, 224]. Moreover, the ICK motif gives stability to the molecule, a very important characteristic for molecular engineering applications [224]. Hence, ICK toxins from widow spiders can potentially be used as pharmacological tools for multiple studies.
Conclusion
Although the real epidemiological profile of latrodectism in the Americas is still unknown, the reports of envenomation by Latrodectus spp. date back to the 18th century. Widow spiders from the Americas encompass 44% of the valid Latrodectus species, yet the venom from most of these species remains uncharacterized. Most of the data obtained from venom analyses focus on latrotoxins and latrodectins from a few species. For example, α-LTX is the most studied toxin so far. It is known to play an important role in envenomation by inducing massive non-specific neurotransmitter release. Recently, high-throughput methods have been used in the study of widow spider venoms, allowing a deeper comprehension of the molecules present in Latrodectus spp. Nevertheless, the current knowledge on the subject is still limited, so the exploration and comparison among different venoms are necessary.
Future studies should focus not only on the venom, but also on the silk fibers, eggs, and body parts. This way, not only a better understanding of the envenomation picture would be possible, but also new molecules of biotechnological interest could be uncovered. Moreover, these analyses could clarify the phylogenetic relationships within the Latrodectus genus and among widow spiders and other Araneae. Lastly, even though the use of antivenom in the treatment of Latrodectus bites is still controversial, we believe that the production of a Pan American anti-Latrodectus serum would be beneficial from a public health perspective, given the good results of serotherapy reported in the literature. However, more studies are necessary to reduce the immunogenicity of antivenom and therefore the chances of unwanted side effects.
Abbreviations
Ach: acetylcholine; ACVGM: aqueous components of venom glands macerated; CHH: crustacean hyperglycaemic hormone; CIRL: calcium-independent receptor of α-LTX; CRISPs: cysteine-rich secretory proteins; CSTX: Cupiennius salei toxin; GABA: γ-aminobutyric acid; GIH: gonad-inhibiting hormone; ICK: inhibitor cystine knot; LCT: latrocrustatoxin; LIT: latroinsectotoxin; LPH1: latrophilin 1; LRR: leucine-rich repeats; LTXs: latrotoxins; MIH: molt-inhibiting hormone; nAChR: nicotinic acetylcholine receptor; NMJ: neuromuscular junction; NXR-Iα: neurexin-Iα; PKC: protein kinase C; PLC: phospholipase C; PTPσ: protein tyrosine phosphatase σ; SV: synaptic vessels; VGCC: voltage-gated calcium channels; VIH: vitellogenesis-inhibiting hormone; α-LIT: α-latroinsectotoxin; α-LTX: α-latrotoxin.
Acknowledgments
The authors thank Sidney Prytherch for the English review of the original manuscript.
To allow the comparison between works, the LD50 values reported by De Roodt et al. [101] were recalculated and expressed as mg/kg, considering 20 g as the mean weight of mice.
Availability of data and materials: All data generated or analyzed during this study are included in this article.
Funding: This work was supported by FAPERJ (grant no. E-26/ 202.829/2017 and -E26/010.003016/2014), CNPq (grant no. 308179/2015-3), and IVB. The PNPD-CAPES 2017 scholarship was granted to MBC.
Ethics approval : Not applicable.
Consent for publication: Not applicable.
References
- Lira q1-da-Silva RM, Matos GB, Sampaio RO, Nunes TB. Estudo retrospectivo de latrodectismo na Bahia, Brasil. Rev Soc Bras Med Trop. 1995;28:205–210. doi: 10.1590/s0037-86821995000300007. [DOI] [PubMed] [Google Scholar]
- Sutherland SK, Trinca JC. Survey of 2144 cases of red‐back spider bites: Australia and New Zealand, 1963‐1976. Med J Australia. 1978;2(14):620–623. doi: 10.5694/j.1326-5377.1978.tb131783.x.. [DOI] [PubMed] [Google Scholar]
- Bettini S, Maroli M. In: Handbook of experimental pharmacology. Bettini S, editor. New York: Springer-Verlag; 1978. Venoms of Theridiidae, genus Latrodectus; pp. 149–184. [Google Scholar]
- Maretić Z. Latrodectism: variations in clinical manifestations provoked by Latrodectus species of spiders. Toxicon. 1983;21(4):457–466. doi: 10.1016/0041-0101(83)90123-x.. [DOI] [PubMed] [Google Scholar]
- Golcuk Y, Velibey Y, Gonullu H, Sahin M E. K. Acute toxic fulminant myocarditis after a black widow spider envenomation: case report and literature review. Clin Toxicol. 2013;51(3):191–192. doi: 10.3109/15563650.2013.774010. [DOI] [PubMed] [Google Scholar]
- Agnarsson I. Morphological phylogeny of cobweb spiders and their relatives (Araneae, Araneoidea, Theridiidae) Zool J Linn Soc. 2004;141(4):447–626. doi: 10.1111/j.1096-3642.2004.00120.x.. [DOI] [Google Scholar]
- Aguilera-Ardiles MA. Delimitación de especies del género Latrodectus (Araneae : Theridiidae) a partir de evidencia molecular y morfológica. Concepción: Universidad de Concepción; 2016. dissertation. [Google Scholar]
- Pickard-Cambridge FO. On the spiders of the genus Latrodectus, Walckenaer. P Zool Soc Lond. 1902;1:247–261. [Google Scholar]
- Gerschman BS, Schiapelli RD. In: Latrodectus mactans y latrodectismo. Sampayo R, editor. Buenos Aires: Museo Argentino de Ciencias Naturales; 1943. Revisión del género Latrodectus Walckenaer 1805; pp. 1–23. [Google Scholar]
- Levi HW. The spider genus Latrodectus (Araneae, Theridiidae) T Am Micros Soc. 1959;78(1):7–43. [Google Scholar]
- Garb JE, Gonzalez A, Gillespie RG. The black widow spider genus Latrodectus (Araneae: Theridiidae): phylogeny, biogeography, and invasion history. Mol Phylogenet Evol. 2004;31(3):1127–1142. doi: 10.1016/j.ympev.2003.10.012.. [DOI] [PubMed] [Google Scholar]
- Abalos JW, Báez EC. Las arañas del genero Latrodectus en Santiago del Estero. Rev Fac Cienc Exact Fis Nat. 1967;27:6–30. [Google Scholar]
- McCrone JD. Biochemical differentiation of the sibling black widow spiders, Latrodectus mactans and L. variolus. Psyche. 1967;74(3):212–217. doi: 10.1155/1967/878547.. [DOI] [Google Scholar]
- Kaston BJ. Notes on a new variety of black widow spider from Southern Florida. Fla Entomol. 1938;21(4):60–62. [Google Scholar]
- Kaston BJ. Comparative biology of American black widow spiders. T San Diego Soc Nat Hist. 1970;16:33–82. [Google Scholar]
- Gonzalez TL. Latrodectus mactans mexicanus subsp. nov. An Inst Biol Univ Nac México. 1954;24:455–457. [Google Scholar]
- McCrone JD, Levi HW. North American widow spiders of the Latrodectus curacaviensis group (Araneae: Theridiidae) Psyche. 1964;71(1):12–27. doi: 10.1155/1964/86469.. [DOI] [Google Scholar]
- Levi HW. Cosmopolitan and pantropical species of theridiid spiders (Araneae: Theridiidae) Pac Insects. 1967;9:175–186. [Google Scholar]
- Levi HW, Randolph DE. A key and checklist of American spiders of the family Theridiidae north of Mexico (Araneae) J Arachnol. 1975;3:31–51. [Google Scholar]
- Gerschman BS, Schiapelli RD. El género Latrodectus Walckenaer, 1805 (Araneae: Theridiidae) en la Argentina. Rev Soc Entomol Arg. 1965;27:51–59. [Google Scholar]
- Mello-Leitão CF. Arañas de la provincia de Buenos Aires y de las gobernaciones de La Pampa, Neuquén, Río Negro y Chubut. Rev Mus La Plata. 1940;2:3–62. [Google Scholar]
- Mello-Leitão CF. Arañas del Paraguay. Notas Mus La Plata. 1946;11(91):17–50. [Google Scholar]
- Carcavallo RU. Una nueva Latrodectus y consideraciones sobre las especies del género en la Republica Argentina (Arach. Theridiidae) Neotropica. 1960;5:85–94. [Google Scholar]
- Roth VD, Craig PR. In: Mission zoologique belge aux iles Galapagos et en Ecuador Résultats scientifiques Premiere partie. Leleup N, Leleup J, editors. 1970. Arachnida of the Galapagos Islands; pp. 107–124. [Google Scholar]
- Aguilera MA, D'Elía G, Casanueva ME. Revalidation of Latrodectus thoracicus Nicolet, 1849 (Araneae:Theridiidae): biological and phylogenetic antecedents. Gayana. 2009;73:161–171. [Google Scholar]
- Simó M, Dias MFR, Jorge C, Castro M, Dias MA, Laborda A. Habitat, redescription and distribution of Latrodectus geometricus in Uruguay (Araneae: Theridiidae) Biota Neotrop. 2013;13(1):371–375. doi: 10.1590/s1676-06032013000100040. [DOI] [Google Scholar]
- World Spider Catalog. Version 22.0. Natural History Museum Bern. 2021. [DOI] [Google Scholar]
- De Biasi P. Variações em aranhas do complexo Latrodectus mactans - Latrodectus curacaviensis (Araneae; Theridiidae) Rev Bras Biol. 1970;30:233–244. [Google Scholar]
- Burt CE. A review of the biology and distribution of the hourglass spider. J Kansas Entomol Soc. 1935;8(4):117–130. [Google Scholar]
- Forster RR, Platnick NI. A review of the archaeid spiders and their relatives, with notes on the limits of the superfamily Palpimanoidea (Arachnida, Araneae) Bull Am Mus Nat Hist. 1984;178 [Google Scholar]
- Mowery M. Invasion in the light of evolution: an integrative approach using two widespread widow spider species. Toronto, ON: Toronto, ON: University of Toronto; 2020. dissertation. [Google Scholar]
- Wang Y, Casajus N, Buddle C, Berteaux D, Larrivee M. Predicting the distribution of poorly-documented species, Northern black widow (Latrodectus variolus) and black purse-web spider (Sphodros niger), using museum specimens and citizen science data. PLoS One. 2018;13(8):e0201094. doi: 10.1371/journal.pone.0201094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi M, Lee S, Yoo JS, Jun J, Kwon O. First record of the Western black widow spider Latrodectus hesperus Chamberlin & Ivie, 1935 (Araneae: Theridiidae) in South Korea: Western black widow spider in Korea. Entomol Res. 2019;49(3):141–146. doi: 10.1111/1748-5967.12350.. [DOI] [Google Scholar]
- Goyal V, Malik V. The first record of the redback spider Latrodectus hasselti Thorell (Araneae: Theridiidae) from Haryana, North India. JEBS. 2017;31:441–442. [Google Scholar]
- Vink CJ, Derraik JGB, Phillips CB, Sirvid PJ. The invasive Australian redback spider, Latrodectus hasseltii Thorell 1870 (Araneae: Theridiidae): current and potential distributions, and likely impacts. Biol Invasions. 2011;13(4):1003–1019. doi: 10.1007/s10530-010-9885-6.. [DOI] [Google Scholar]
- Bayram A, Danisman T, Yigit N, Kunt KB, Sankac A. A brown widow spider new for the Turkish araneo-fauna: Latrodectus geometricus C. L. Koch, 1841 (Araneae; Theridiidae) Turk J Arachnol. 2008;1:98–103. [Google Scholar]
- Taucare-Rios A, Bizama G, Bustamante RO. Using global and regional species distribution models (SDM) to infer the invasive stage of Latrodectus geometricus (Araneae: Theridiidae) in the Americas. Environ Entomol. 2016;45(6):1379–85. doi: 10.1093/ee/nvw118. [DOI] [PubMed] [Google Scholar]
- Muslimin M, Wilson JJ, Ghazali AR, Braima KA, Jeffery J, Wan-Nor F, et al. First report of brown widow spider sightings in Peninsular Malaysia and notes on its global distribution. J Venom Anim Toxins Incl Trop Dis. 2015;21:11. doi: 10.1186/s40409-015-0010-2.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biaggio MD, Sandomirsky I, Lubin Y, Harari AR, Andrade MC. Copulation with immature females increases male fitness in cannibalistic widow spiders. Biol Lett. 2016;12(9) doi: 10.1098/rsbl.2016.0516.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guimarães IC, Cardoso CA, Lima SM, Andrade LH, Antonialli WF., Junior Chemical signals might mediate interactions between females and juveniles of Latrodectus geometricus (Araneae: Theridiidae) Behavioural processes. 2016;126:27–35. doi: 10.1016/j.beproc.2016.03.001. [DOI] [PubMed] [Google Scholar]
- Abalos JW. The egg-sac in the identification of species of Latrodectus (black-widow spiders) Psyche. 1962;69(4):268–270. doi: 10.1155/1962/36967.. [DOI] [Google Scholar]
- Andrade MCB. In: Advances in the study of behavior. Naguib M, Barrett L, Healy SD, Podos J, Simmons LW, Zuk M, editors. Academic Press; 2019. Sexual selection and social context: web-building spiders as emerging models for adaptive plasticity; pp. 177–250. [Google Scholar]
- Deevey GB. The developmental history of Latrodectus mactans (Fabr.) at different rates of feeding. Am Midl Nat. 1949;42(1) doi: 10.2307/2421795.. [DOI] [Google Scholar]
- Andrade M, MacLeod E. In: Cryptic female choice in arthropods. Peretti A, Aisenberg A, editors. Springer; 2015. Potential for CFC in black widows (genus Latrodectus): mechanisms and social context; pp. 27–53. [Google Scholar]
- Bertani R, Fukushima C, Martins R. Sociable widow spiders? Evidence of subsociality in Latrodectus Walckenaer, 1805 (Araneae, Theridiidae) J Ethol. 2008;26(2):299–302. doi: 10.1007/s10164-007-0082-8.. [DOI] [Google Scholar]
- Salomon M, Vibert S, Bennett RG. Habitat use by Western black widow spiders (Latrodectus hesperus) in coastal British Columbia: evidence of facultative group living. Can J Zool. 2010;88(3):334–346. doi: 10.1139/z10-004.. [DOI] [Google Scholar]
- Morley EL, Sivalinghem S, Mason AC. Developmental morphology of a lyriform organ in the Western black widow (Latrodectus hesperus) Zoomorphology. 2016;135(4):433–440. doi: 10.1007/s00435-016-0324-9.. [DOI] [Google Scholar]
- Antonialli-Junior W, Guimarães I. Aggregation behavior in spiderlings: a strategy for increasing life expectancy in Latrodectus geometricus (Araneae: Theridiidae) Sociobiology. 2014;59(2):463. doi: 10.13102/sociobiology.v59i2.611.. [DOI] [Google Scholar]
- Johnson J, Halpin R, Stevens D, Vannan A, Lam J, Bratsch K. Individual variation in ballooning dispersal by black widow spiderlings: the effects of family and social rearing. Curr Zool. 2015;61(3):520–528. doi: 10.1093/czoolo/61.3.520.. [DOI] [Google Scholar]
- Johnson JC, Urcuyo J, Moen C, Stevens DR. Urban heat island conditions experienced by the Western black widow spider (Latrodectus hesperus): extreme heat slows development but results in behavioral accommodations. PLoS One. 2019;14(9):e0220153. doi: 10.1371/journal.pone.0220153.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schenone H, Correa LE. Algunos conocimientos prácticos sobre la biología de la araña Latrodectus mactans y el síndrome del latrodectismo en Chile. Bol Chil Parasitol. 1985;40(1-2):18–23. [PubMed] [Google Scholar]
- Blair AW. Life history of Latrodectus mactans. Arch Intern Med. 1934;54(6) doi: 10.1001/archinte.1934.00160180016002.. [DOI] [Google Scholar]
- Hódar JA, Sánchez-Piñero F. Feeding habits of the blackwidow spider Latrodectus lilianae (Araneae: Theridiidae) in an arid zone of south-east Spain. J Zool. 2002;257(1):101–109. doi: 10.1017/s0952836902000699.. [DOI] [Google Scholar]
- Noriega J. First report of Latrodectus geometricus Koch (Araneae: Theridiidae) as a predator of Digitonthophagus gazella (Fabricius) (Coleoptera: Scarabaeidae) Coleopts Bull. 2016;70(2):407–408. doi: 10.1649/0010-065X-70.2.407.. [DOI] [Google Scholar]
- Brandley N, Johnson M, Johnsen S. Aposematic signals in North American black widows are more conspicuous to predators than to prey. Behav Ecol. 2016;27(4):1104–1112. doi: 10.1093/beheco/arw014.. [DOI] [Google Scholar]
- Cariaso BL. A biology of the black widow spider, Latrodectus hasselti Thorell (Araneida: Theridiidae) Philippine Agric. 1967;51:171–180. [Google Scholar]
- DiRienzo N, Aonuma H. Plasticity in extended phenotype increases offspring defence despite individual variation in web structure and behaviour. Anim Behav. 2018;138:9–17. doi: 10.1016/j.anbehav.2018.01.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anthony SE. Thermal biology of temperate and high-latitude arachnids. London, ON: London, ON: The University of Western Ontario; 2019. dissertation. [Google Scholar]
- Bettini S. Epidemiology of Latrodectus. Toxicon. 1964;2:93–102. doi: 10.1016/0041-0101(64)90009-1. [DOI] [PubMed] [Google Scholar]
- Maretić Z. In: Handbook of experimental pharmacology. Bettini S, editor. New York: Springer-Verlag; 1978. Venoms of Theridiidae, genus Latrodectus: epidemiology of envenomation, symptomatology, pathology and treatment; pp. 185–212. [Google Scholar]
- Lazarte PEO, Samur NPO, Martinez RAO. Latrodectismo. Rev Cientif Cienc Med. 2009;12(1):25–28. [Google Scholar]
- Grisolia CS, Peluso FO, Stanchi NO, Francini F. Epidemiologia del latrodectismo en la Provincia de Buenos Aires, Argentina. Rev Saúde Pública. 1992;26(1):1–5. doi: 10.1590/s0034-89101992000100001.. [DOI] [PubMed] [Google Scholar]
- Curran-Sills G, Wilson SM, Bennett R. A review of black widow (Araneae: Theridiidae) envenomation, epidemiology, and antivenom utilization in Canada. J Med Entomol. 2021;58(1):99–103. doi: 10.1093/jme/tjaa148. [DOI] [PubMed] [Google Scholar]
- Clark RF, Wethern-Kestner S, Vance MV, Gerkin R. Clinical presentation and treatment of black widow spider envenomation: a review of 163 cases. Ann Emerg Med. 1992;21(7):782–787. doi: 10.1016/s0196-0644(05)81021-2.. [DOI] [PubMed] [Google Scholar]
- Monte AA, Bucher-Bartelson B, Heard KJ. A US perspective of symptomatic Latrodectus spp. envenomation and treatment: a National Poison Data System review. Ann Pharmacother. 2011;45(12):1491–1498. doi: 10.1345/aph.1Q424. [DOI] [PubMed] [Google Scholar]
- Castañeda-Gómez J, Pinkus-Rendón M, Arisqueta-Chablé C, Barrera-Pérez M, Ortiz-Martínez D, Manrique-Saide P. Nuevos registros del género Latrodectus en Yucatán, México. Rev Bioméd. 2012;23:105–111. [Google Scholar]
- Baerg WJ. The brown widow and the black widow spiders in Jamaica (Araneae, Theridiidae) Ann Entomol Soc Am. 1954;47(1):52–60. doi: 10.1093/aesa/47.1.52.. [DOI] [Google Scholar]
- Castellanos E, Rodríguez L. Loxoscelismo cutáneo y cutáneo visceral. Rev Méd Col Méd Cir Guatem. 2017;156(1):47–50. [Google Scholar]
- Oviedo AH, Marín M, Martínez O, Martínez H. Una especie de viuda negra (Latrodectus hesperus Chamberlain & Ivie) encontrada en Olancho, Honduras. Rev Cienc Tecnol. 2009;5:37–46. [Google Scholar]
- Cruz OAC, Barrera RES. Síndrome neurotóxico por mordedura de Latrodectus mactans: reporte de un caso pediátrico y revisión de literatura. CIMEL. 2014;19(2):87–90. [Google Scholar]
- Ramos JJL. Distribucion y frecuencia de Latrodectus mactans Fabricio y sus enemigos naturales en El Salvador. San Salvador: Universidad de El Salvador; 1977. dissertation. [Google Scholar]
- Vado VV. Envenenamiento por araña viuda negra en un niño: reporte de un caso. Nicaragua Pediátrica. 2013;1(3):33–35. [Google Scholar]
- Barrantes-Montero G, Valerio-Pizarro R. Lactrodectismo en Costa Rica. Acta Méd Costarric. 2017;59(2):73–75. [Google Scholar]
- Petrunkevitch A. The spider fauna of Panama and its Central American affiliation. Am Nat. 1929;63(688):455–469. [Google Scholar]
- Jäger H, E Causton C, Rodríguez J, Baert L, Buchholz S. Spiders in Galapagos: diversity, biogeography and origin. Biol J Linn Soc. 2020;130(1):41–48. doi: 10.1093/biolinnean/blaa019. [DOI] [Google Scholar]
- Bucherl W. Biology and venoms of the most important South American spiders of the genera Phoneutria, Loxosceles, Lycosa, and Latrodectus. Am Zool. 1969;9(1):157–159. doi: 10.1093/icb/9.1.157. [DOI] [PubMed] [Google Scholar]
- Kiriakos D, Nunez P, Parababire Y, Garcia M, Medina J, Sousa LD. First case of human latrodectism in Venezuela. Rev Soc Bras Med Trop. 2008;41(2):202–204. doi: 10.1590/s0037-86822008000200015. [DOI] [PubMed] [Google Scholar]
- Canals M, Casanueva ME, Aguilera M. ¿Cuáles son las especies de arañas peligrosas en Chile? Rev Med Chil. 2004;132(6):773–776. doi: 10.4067/s0034-98872004000600016. [DOI] [PubMed] [Google Scholar]
- Abalos JW. Las arañas del género Latrodectus en la Argentina. Obra Cent Mus La Plata. 1980;6:29–52. [Google Scholar]
- Oostburg BFJ, Hiwat H. Araneism in Suriname. Surinaams Med Bull. 1998;13:37–47. [Google Scholar]
- Corsi-Sotelo OF, Del-Río-Otto E, Peña-Riveros A, Acuña-Ramírez D. Latrodectismo: caso clínico y revisión de la literatura. ARS Med Rev Cienc Méd. 2017;42(3):26–30. doi: 10.11565/arsmed.v42i3.1003. [DOI] [Google Scholar]
- Diaz JH. The global epidemiology, syndromic classification, management, and prevention of spider bites. Am J Trop Med Hyg. 2004;71(2):239–250. doi: 10.4269/ajtmh.2004.71.2.0700239. [DOI] [PubMed] [Google Scholar]
- Ali H, Baker I, Fadhil H. Taxonomic and molecular study of the widow spider genus Latrodectus Walckenaer, 1805 (Araneae: Theridiidae) in Iraq. Pak Entomol. 2018;40(1):25–29. [Google Scholar]
- Alizadeh A, Vahabzadeh M, Azarfar A, Ravanshad Y. Misdiagnosis of spider bite in a 3-year-old child. J Res Med Sci. 2018;23:28. doi: 10.4103/1735-1995.228585.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanaei-Zadeh H. Spider bite in Iran. Electron Physician. 2017;9(7):4703. doi: 10.19082/4703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Müller GJ. Black and brown widow spider bites in South Africa: a series of 45 cases. S Afr Med J. 1993;83:399–405. [PubMed] [Google Scholar]
- Afshari R, Khadem-Rezaiyan M, Balali-Mood M. Spider bite (latrodectism) in Mashhad, Iran. Human & experimental toxicology. 2009;28(11):697–702. doi: 10.1177/0960327109350668.. [DOI] [PubMed] [Google Scholar]
- Cesaretli Y, Ozkan O. A clinical and epidemiological study on spider bites in Turkey. Asian Pac J Trop Med. 2011;4(2):159–162. doi: 10.1016/s1995-7645(11)60060-6. [DOI] [PubMed] [Google Scholar]
- Antoniou G, Iliopoulos D, Kalkouni R, Iliopoulou S, Rigakos G, Baka A. Latrodectus envenomation in Greece. Perm J. 2014;18(4):e155–e158. doi: 10.7812/TPP/14-028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isbister GK, Gray MR. Latrodectism: a prospective cohort study of bites by formally identified redback spiders. Med J Australia. 2003;179(2):88–91. doi: 10.5694/j.1326-5377.2003.tb05442.x. [DOI] [PubMed] [Google Scholar]
- Díez-García F, Laynez-Bretones F, Gálvez-Contreras MC, Mohd H, Collado-Romacho A, Yélamos-Rodríguez F. Mordedura por araña viuda negra (Latrodectus tredecimguttatus). Presentación de 12 casos. Medicina clinica. 1996;106(9):344–346. [PubMed] [Google Scholar]
- Barnes CL, Blay NW, Wilder SM. Upper thermal tolerances of different life stages, sexes, and species of widow spiders (Araneae, Theridiidae) J Insect Physiol. 2019;114:10–14. doi: 10.1016/j.jinsphys.2019.02.004.. [DOI] [PubMed] [Google Scholar]
- Benjamin S, Zschokke S. Webs of theridiid spiders: construction, structure and evolution. Biol J Linn Soc. 2003;78(3):293–305. doi: 10.1046/j.1095-8312.2003.00110.x.. [DOI] [Google Scholar]
- Forster LM, Kavale J. Effects of food deprivation on Latrodectus hasselti Thorell (Araneae: Theridiidae), the Australian redback spider. New Zeal J Zool. 1989;16(3):401–408. doi: 10.1080/03014223.1989.10422906.. [DOI] [Google Scholar]
- Eason RR, Peck WB, Whitcomb WH. Notes on spider parasites, including a reference list. J Kansas Entomol Soc. 1967;40(3):422–434. [Google Scholar]
- Matsuse IT, Kamimura K, Yoshida M. The tolerance of Latrodectus hasseltii (Araneae: Theridiidae) to low temperatures. Med Entomol Zool. 1999;50(1):71–73. doi: 10.7601/mez.50.71_1.. [DOI] [Google Scholar]
- Urtubey N, Baez E, Magarinos E. Epidemiologia del Latrodectismo en Santiago del Estero, Argentina. Rev Inst Animales Venenosos. 1984;2:1–28. [Google Scholar]
- Souza AR, Bührnheim PF, Lima CS. Relato de um caso de latrodectismo ocorrido em Manaus, Amazonas, Brasil. Rev Soc Bras Med Trop. 1998;31(1):95–98. [PubMed] [Google Scholar]
- Almeida R, Ferreira-Junior RS, Chaves CR, Barraviera B. Envenomation caused by Latrodectus geometricus in São Paulo state, Brazil: a case report. J Venom Anim Toxins Incl Trop Dis. 2009;15:562–571. [Google Scholar]
- Yan S, Wang X. Recent advances in research on widow spider venoms and toxins. Toxins. 2015;7(12):5055–5067. doi: 10.3390/toxins7124862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Roodt AR, Lanari LC, Laskowicz RD, Costa de Oliveira V, Irazu LE, González A, et al. Toxicity of the venom of Latrodectus (Araneae: Theridiidae) spiders from different regions of Argentina and neutralization by therapeutic antivenoms. Toxicon. 2017;130:63–72. doi: 10.1016/j.toxicon.2017.02.029. [DOI] [PubMed] [Google Scholar]
- Bush SP, Davy JV. Troponin elevation after black widow spider envenomation. CJEM. 2015;17(5):571–575. doi: 10.1017/cem.2015.82. [DOI] [PubMed] [Google Scholar]
- Dendane T, Abidi K, Madani N, Benthami A, Gueddari FZ, Abouqal R, et al. Reversible myocarditis after black widow spider envenomation. Case Rep Med. 2012;2012:794540. doi: 10.1155/2012/794540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pneumatikos IA, Galiatsou E, Goe D, Kitsakos A, Nakos G, Vougiouklakis TG. Acute fatal toxic myocarditis after black widow spider envenomation. Ann Emerg Med. 2003;41(1):158. doi: 10.1067/mem.2003.32. [DOI] [PubMed] [Google Scholar]
- Pulignano G, Del Sindaco D, Giovannini M, Zeisa P, Faia M, Soccorsi M, et al. Myocardial damage after spider bite (Latrodectus tredecimguttatus) in a 16-year-old patient. G Ital Cardiol. 1998;28(10):1149–1153. [PubMed] [Google Scholar]
- Valikhanfard-Zanjani E, Zare-Mirakabadi A, Zayerzadeh E. Antivenom efficacy in neutralizing histopathological complications following Latrodectus dahli envenomation. J Arthropod-Borne Di. 2017;11(1):42–49. [PMC free article] [PubMed] [Google Scholar]
- Erdur B, Turkcuer I, Bukiran A, Kuru O, Varol I. Uncommon cardiovascular manifestations after a Latrodectus bite. Am J Emerg Med. 2007;25(2):232–235. doi: 10.1016/j.ajem.2006.11.005.. [DOI] [PubMed] [Google Scholar]
- Sari I, Zengin S, Davutoglu V, Yildirim C, Gunay N. Myocarditis after black widow spider envenomation. Am J Emerg Med. 2008;26(5):630 e1–630 e3. doi: 10.1016/j.ajem.2007.09.012. [DOI] [PubMed] [Google Scholar]
- Bush S, McCune R, Phan T. Priapism after Western black widow spider (Latrodectus hesperus) envenomation. Wild Environ Med. 2014;25(1):80–81. doi: 10.1016/j.wem.2013.10.008.. [DOI] [PubMed] [Google Scholar]
- Hoover NG, Fortenberry JD. Use of antivenin to treat priapism after a black widow spider bite. Pediatrics. 2004;114(1):e128–e129. doi: 10.1542/peds.114.1.e128. [DOI] [PubMed] [Google Scholar]
- Quan D, Ruha AM. Priapism associated with Latrodectus mactans envenomation. Am J Emerg Med. 2009;27(6):759 e1–759 e2. doi: 10.1016/j.ajem.2008.10.033. [DOI] [PubMed] [Google Scholar]
- Stiles AD. Priapism following a black widow spider bite. Clin Pediatr. 1982;21(3):174–175. doi: 10.1177/000992288202100308.. [DOI] [PubMed] [Google Scholar]
- Bonnet MS. The toxicology of Latrodectus tredecimguttatus: the mediterranean black widow spider. Homeopathy. 2004;93(1):27–33. doi: 10.1016/j.homp.2003.10.001. [DOI] [PubMed] [Google Scholar]
- Saracco S, De Roodt A. Latrodectismo. Rev Med Universitaria. 2010;6(3):1–13. [Google Scholar]
- Shackleford R, Veillon D, Maxwell N, LaChance L, Jusino T, Cotelingam J, et al. The black widow spider bite: differential diagnosis, clinical manifestations, and treatment options. J Louisiana St Med Soc. 2015;167(2):74–78. [PubMed] [Google Scholar]
- Fusto G, Bennardo L, Duca ED, Mazzuca D, Tamburi F, Patruno C, et al. Spider bites of medical significance in the Mediterranean area: misdiagnosis, clinical features and management. J Venom Anim Toxins Incl Trop Dis. 2020;26:e20190100. doi: 10.1590/1678-9199-JVATITD-2019-0100.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogen E. Arachnidism: a study in spider poisoning. J Am Med Assoc. 1926;86(25):1894–1896. [Google Scholar]
- Monte AA. Black widow spider (Latrodectus mactans) antivenom in clinical practice. Curr Pharm Biotechno. 2012;13(10):1935–1939. doi: 10.2174/138920112802273335.. [DOI] [PubMed] [Google Scholar]
- Nordt SP, Clark RF, Lee A, Berk K, Lee-Cantrell F. Examination of adverse events following black widow antivenom use in California. Clin Toxicol. 2012;50(1):70–3. doi: 10.3109/15563650.2011.639714. [DOI] [PubMed] [Google Scholar]
- Heard K, O'Malley GF, Dart RC. Antivenom therapy in the Americas. Drugs. 1999;58(1):5–15. doi: 10.2165/00003495-199958010-00002. [DOI] [PubMed] [Google Scholar]
- Isbister GK, Graudins A, White J, Warrell D. Antivenom treatment in arachnidism. J Toxicol Clin Toxicol. 2003 doi: 10.1081/clt-120021114. [DOI] [PubMed] [Google Scholar]
- Murphy CM, Hong JJ, Beuhler MC. Anaphylaxis with Latrodectus antivenin resulting in cardiac arrest. J Med Toxicol. 2011;7(4):317–321. doi: 10.1007/s13181-011-0183-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leiva CL, Gallardo MJ, Casanova N, Terzolo H, Chacana P. IgY-technology (egg yolk antibodies) in human medicine: a review of patents and clinical trials. Int Immunopharmacol. 2020;81:106269. doi: 10.1016/j.intimp.2020.106269. [DOI] [PubMed] [Google Scholar]
- Daly FFS, Hill RE, Bogdan GM, Dart RC. Neutralization of Latrodectus mactans and L. hesperus venom by redback spider (L. hasseltii) antivenom. J Toxicol Clin Toxicol. 2001;39(2):119–123. doi: 10.1081/clt-100103826.. [DOI] [PubMed] [Google Scholar]
- Graudins A, Gunja N, Broady KW, Nicholson GM. Clinical and in vitro evidence for the efficacy of Australian red-back spider (Latrodectus hasselti) antivenom in the treatment of envenomation by a Cupboard spider (Steatoda grossa) Toxicon. 2002;40(6):767–775. doi: 10.1016/s0041-0101(01)00280-x.. [DOI] [PubMed] [Google Scholar]
- Souza CM. Notas sobre o Latrodectismo no Brasil. Gaz Méd Bahia. 2012;82:90–96. [Google Scholar]
- Lucas SM. The history of venomous spider identification, venom extraction methods and antivenom production: a long journey at the Butantan Institute, São Paulo, Brazil. J Venom Anim Toxins Incl Trop Dis. 2015;21:21. doi: 10.1186/s40409-015-0020-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frontali N, Grasso A. Separation of three toxicologically different protein components from the venom of the spider Latrodectus tredecimguttatus. Arch Biochem Biophys. 1964;106:213–218. doi: 10.1016/0003-9861(64)90178-x.. [DOI] [PubMed] [Google Scholar]
- Duan ZG, Yan XJ, He XZ, Zhou H, Chen P, Cao R, et al. Extraction and protein component analysis of venom from the dissected venom glands of Latrodectus tredecimguttatus. Comp Biochem Physiol B Biochem Mol Biol. 2006;145(3-4):350–357. doi: 10.1016/j.cbpb.2006.08.006. [DOI] [PubMed] [Google Scholar]
- Garb JE. Extraction of venom and venom gland microdissections from spiders for proteomic and transcriptomic analyses. J Vis Exp. 2014;(93):e51618. doi: 10.3791/51618.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lebez D. Beiträge zum studium des giftes von Latrodectus tredecimguttatus Rossi. Biol Chem. 1954;298:73–76. [PubMed] [Google Scholar]
- Hayes WK, Fox GA, Nelsen DR. Venom collection from spiders and snakes: voluntary and involuntary extractions ("milking") and venom gland extractions. Meth Mol Biol. 2020;2068:53–71. doi: 10.1007/978-1-4939-9845-6_3. [DOI] [PubMed] [Google Scholar]
- Ichtchenko K, Khvotchev M, Kiyatkin N, Simpson L, Sugita S, Sudhof TC. alpha-latrotoxin action probed with recombinant toxin: receptors recruit alpha-latrotoxin but do not transduce an exocytotic signal. EMBO J. 1998;17(21):6188–6199. doi: 10.1093/emboj/17.21.6188.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reyes-Lugo M, Sanchez T, Finol HJ, Sanchez EE, Suarez JA, Guerreiro B, et al. Neurotoxic activity and ultrastructural changes in muscles caused by the brown widow spider Latrodectus geometricus venom. Rev Inst Med Trop São Paulo. 2009;51(2):95–101. doi: 10.1590/s0036-46652009000200007. [DOI] [PubMed] [Google Scholar]
- Chippaux JP, Williams V, White J. Snake venom variability: methods of study, results and interpretation. Toxicon. 1991;29(11):1279–1303. doi: 10.1016/0041-0101(91)90116-9.. [DOI] [PubMed] [Google Scholar]
- Parodi J, Romero F. Synaptic effects of low molecular weight components from Chilean black widow spider venom. Neurotoxicology. 2008;29(6):1121–1126. doi: 10.1016/j.neuro.2008.08.006. [DOI] [PubMed] [Google Scholar]
- Romero F, Salvatici R, Schulz C, Cabezas M, Standen D, Altieri E. Efecto del veneno de Latrodectus mactans sobre la respuesta contráctil del músculo papilar de rata. Rev Chil Cardiol. 1996;15(3):112 [Google Scholar]
- Romero F, Altieri E, Quiñehual C, Cayuqueo A. Actividad contráctil de músculo papilar cardíaco y conducto deferente de rata inducido por veneno de la araña Lactrodectus mactans de Chile. Gayana. 2000;64:161–170. [Google Scholar]
- Romero F, Altieri E, Urrutia M, Jara J. Veneno de Latrodectus mactans de Chile (Araneae, Theridiidae): su efecto sobre músculo liso. Rev Biol Trop. 2003;51(2):305–312. [PubMed] [Google Scholar]
- Ny L, Waldeck K, Carlemalm E, Andersson KE. Alpha-latrotoxin-induced transmitter release in feline oesophageal smooth muscle: focus on nitric oxide and vasoactive intestinal peptide. Brit J Pharmacol. 1997;120(1):31–38. doi: 10.1038/sj.bjp.0700882.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werkstrom V, Ny L, Persson K, Andersson KE. Neurotransmitter release evoked by alpha-latrotoxin in the smooth muscle of the female pig urethra. Naunyn-Schmiedeberg's Arch Pharmacol. 1997;356(2):151–158. doi: 10.1007/pl00005035. [DOI] [PubMed] [Google Scholar]
- Valikhanfard-Zanjani E, Zare-Mirakabadi A, Oryan S, Goodarzi HR, Rajabi M. Specific antivenom ability in neutralizing hepatic and renal changes 24 hours after Latrodectus dahli envenomation. J Arthropod-Borne Di. 2016;10(2):237–244. [PMC free article] [PubMed] [Google Scholar]
- Maretić Z. Arachnidism treated with cortisone: report of a case. J Am Med Assoc. 1953;152 doi: 10.1001/jama.1953.63690070005010d. [DOI] [PubMed] [Google Scholar]
- Marzan B. Pathologic reactions associated with bite of Latrodectus tredecimguttatus: observations in experimental animals. Arch Pathol. 1955;59(6):727–728. [PubMed] [Google Scholar]
- Maretić Z, Stanic M. The health problem of arachnidism. Bull World Health Org. 1954;11(6):1007–1022. [PMC free article] [PubMed] [Google Scholar]
- Lauria PSS, Casais-e-Silva LL, Espirito-Santo RF, Souza CMV, Zingali RB, Caruso MB, et al. Pain-like behaviors and local mechanisms involved in the nociception experimentally induced by Latrodectus curacaviensis spider venom. Toxicol Lett. 2018;299:67–75. doi: 10.1016/j.toxlet.2018.09.008.. [DOI] [PubMed] [Google Scholar]
- Rash LD, Hodgson WC. Pharmacology and biochemistry of spider venoms. Toxicon. 2002;40(3):225–254. doi: 10.1016/s0041-0101(01)00199-4. [DOI] [PubMed] [Google Scholar]
- Grishin EV. Black widow spider toxins: the present and the future. Toxicon. 1998;36(11):1693–1701. doi: 10.1016/s0041-0101(98)00162-7. [DOI] [PubMed] [Google Scholar]
- Longenecker HEJ, Hurlbut WP, Mauro A, Clark AW. Effects of black widow spider venom on the frog neuromuscular junction: effects on end-plate potential, miniature end-plate potential and nerve terminal spike. Nature. 1970;225(5234):701–703. doi: 10.1038/225701a0. [DOI] [PubMed] [Google Scholar]
- Frontali N, Ceccarelli B, Gorio A, Mauro A, Siekevitz P, Tzeng MC, et al. Purification from black widow spider venom of a protein factor causing the depletion of synaptic vesicles at neuromuscular junctions. J Cell Biol. 1976;68(3):462–479. doi: 10.1083/jcb.68.3.462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tzeng MC, Siekevitz P. The effect of the purified major protein factor (α-latrotoxin) of black widow spider venom on the release of acetylcholine and norepinephrine from mouse cerebral cortex slices. Brain Research. 1978;139(1):190–196. doi: 10.1016/0006-8993(78)90073-2.. [DOI] [PubMed] [Google Scholar]
- Orlova EV, Rahman MA, Gowen B, Volynski KE, Ashton AC, Manser C, et al. Structure of alpha-latrotoxin oligomers reveals that divalent cation-dependent tetramers form membrane pores. Nat Struct Mol Biol. 2000;7(1):48–53. doi: 10.1038/71247. [DOI] [PubMed] [Google Scholar]
- Ashton AC, Rahman MA, Volynski KE, Manser C, Orlova EV, Matsushita H, et al. Tetramerisation of α-latrotoxin by divalent cations is responsible for toxin-induced non-vesicular release and contributes to the Ca2+-dependent vesicular exocytosis from synaptosomes. Biochimie. 2000;82(5):453–468. doi: 10.1016/s0300-9084(00)00199-1.. [DOI] [PubMed] [Google Scholar]
- Sokolov YV, Chanturia AN, Lishko VK. Latrotoxin-induced fusion of liposomes with bilayer phospholipid membranes. BBA Biomembranes. 1987;900(2):295–299. doi: 10.1016/0005-2736(87)90345-2.. [DOI] [PubMed] [Google Scholar]
- Grasso A, Alema S, Rufini S, Senni MI. Black widow spider toxin-induced calcium fluxes and transmitter release in a neurosecretory cell line. Nature. 1980;283(5749):774–776. doi: 10.1038/283774a0.. [DOI] [PubMed] [Google Scholar]
- Grasso A. Preparation and properties of a neurotoxin purified from the venom of black widow spider (Latrodectus mactans tredecimguttatus) BBA Protein Structure. 1976;439(2):406–412. doi: 10.1016/0005-2795(76)90077-5. [DOI] [PubMed] [Google Scholar]
- Tzeng MC, Cohen RS, Siekevitz P. Release of neurotransmitters and depletion of synaptic vesicles in cerebral cortex slices by alpha-latrotoxin from black widow spider venom. P Natl Acad Sci USA. 1978;75(8):4016–4020. doi: 10.1073/pnas.75.8.4016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picotti GB, Bondiolotti GP, Meldolesi J. Peripheral catecholamine release by alpha-latrotoxin in the rat. Naunyn-Schmiedeberg's Arch Pharmacol. 1982;320(3):224–229. doi: 10.1007/BF00510132.. [DOI] [PubMed] [Google Scholar]
- Lang J, Ushkaryov Y, Grasso A, Wollheim CB. Ca2+-independent insulin exocytosis induced by alpha-latrotoxin requires latrophilin, a G protein-coupled receptor. EMBO J. 1998;17(3):648–657. doi: 10.1093/emboj/17.3.648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hlubek M, Tian D, Stuenkel EL. Mechanism of alpha-latrotoxin action at nerve endings of neurohypophysis. Brain Research. 2003;992(1):30–42. doi: 10.1016/j.brainres.2003.08.025.. [DOI] [PubMed] [Google Scholar]
- Henkel AW, Sankaranarayanan S. Mechanisms of alpha-latrotoxin action. Cell Tissue Res. 1999;296(2):229–233. doi: 10.1007/s004410051284. [DOI] [PubMed] [Google Scholar]
- Ushkaryov YA, Rohou A, Sugita S. Handbook of experimental pharmacology. New York: Springer-Verlag; 2008. alpha-Latrotoxin and its receptors; pp. 171–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenthal L, Meldolesi J. α-latrotoxin and related toxins. Pharmacol Therap. 1989;42(1):115–134. doi: 10.1016/0163-7258(89)90024-7. [DOI] [PubMed] [Google Scholar]
- Ushkaryov YA, Petrenko AG, Geppert M, Sudhof TC. Neurexins: synaptic cell surface proteins related to the alpha-latrotoxin receptor and laminin. Science. 1992;257(5066):50–56. doi: 10.1126/science.1621094. [DOI] [PubMed] [Google Scholar]
- Davletov BA, Shamotienko OG, Lelianova VG, Grishin EV, Ushkaryov YA. Isolation and biochemical characterization of a Ca2+-independent alpha-latrotoxin-binding protein. J Biol Chem. 1996;271(38):23239–23245. doi: 10.1074/jbc.271.38.23239.. [DOI] [PubMed] [Google Scholar]
- Lelianova VG, Davletov BA, Sterling A, Rahman MA, Grishin EV, Totty NF, et al. Alpha-latrotoxin receptor, latrophilin, is a novel member of the secretin family of G protein-coupled receptors. J Biol Chem. 1997;272(34):21504–21508. doi: 10.1074/jbc.272.34.21504. [DOI] [PubMed] [Google Scholar]
- Krasnoperov VG, Bittner MA, Beavis R, Kuang Y, Salnikow KV, Chepurny OG, et al. α-latrotoxin stimulates exocytosis by the interaction with a neuronal G-protein-coupled receptor. Neuron. 1997;18(6):925–937. doi: 10.1016/s0896-6273(00)80332-3. [DOI] [PubMed] [Google Scholar]
- Krasnoperov V, Bittner MA, Mo W, Buryanovsky L, Neubert TA, Holz RW, et al. Protein-tyrosine phosphatase-sigma is a novel member of the functional family of alpha-latrotoxin receptors. J Biol Chem. 2002;277(39):35887–35895. doi: 10.1074/jbc.M205478200. [DOI] [PubMed] [Google Scholar]
- Kovalevskaia GI, Pashkov VN, Bulgakov OV, Fedorova IM, Magazanik LG, Grishin EV. Identifikatsiia i vydelenie belkovogo insektotoksina (alpha-latroinsektotoksina) iz iada pauka Latrodectus mactans tredecimguttatus [Identification and isolation of the protein insect toxin (alpha-latroinsectotoxin from venom of the spider Latrodectus mactans tredecimguttatus] Bioorganicheskaya Khimiya. 1990;16(8):1013–1018. [PubMed] [Google Scholar]
- Kiyatkin NI, Kulikovskaya IM, Grishin EV, Beadle DJ, King LA. Functional characterization of black widow spider neurotoxins synthesised in insect cells. Eur J Biochem. 1995;230(3):854–859. doi: 10.1111/j.1432-1033.1995.tb20628.x.. [DOI] [PubMed] [Google Scholar]
- Magazanik LG, Fedorova IM, Kovalevskaya GI, Pashkov VN, Bulgakov OV, Grishin EV. Selective presynaptic insectotoxin (α-latroinsectotoxin) isolated from black widow spider venom. Neuroscience. 1992;46(1):181–188. doi: 10.1016/0306-4522(92)90017-v. [DOI] [PubMed] [Google Scholar]
- Rohou A, Nield J, Ushkaryov YA. Insecticidal toxins from black widow spider venom. Toxicon. 2007;49(4):531–549. doi: 10.1016/j.toxicon.2006.11.021.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krasnoperov VG, Shamotienko OG, Grishin EV. Vydelenie i svoĭstva insektospetsificheskikh neĭrotoksinov iz iada pauka karakurta Latrodectus mactans tredecimguttatus [Isolation and properties of insect-specific neurotoxins from venoms of the spider Lactodectus mactans tredecimguttatus] Bioorganicheskaya Khimiya. 1990;16(8):1138–1140. [PubMed] [Google Scholar]
- Dulubova IE, Krasnoperov VG, Khvotchev MV, Pluzhnikov KA, Volkova TM, Grishin EV, et al. Cloning and structure of delta-latroinsectotoxin, a novel insect-specific member of the latrotoxin family: functional expression requires C-terminal truncation. J Biol Chem. 1996;271(13):7535–7543. doi: 10.1074/jbc.271.13.7535. [DOI] [PubMed] [Google Scholar]
- Fritz LC, Tzen MC, Mauro A. Different components of black widow spider venom mediate transmitter release at vertebrate and lobster neuromuscular junctions. Nature. 1980;283(5746):486–487. doi: 10.1038/283486a0.. [DOI] [PubMed] [Google Scholar]
- Kiyatkin NI, Dulubova IE, Chekhovskaya IA, Grishin EV. Cloning and structure of cDNA encoding α-latrotoxin from black widow spider venom. FEBS Lett. 1990;270(1-2):127–131. doi: 10.1016/0014-5793(90)81250-r. [DOI] [PubMed] [Google Scholar]
- Kiyatkin N, Dulubova I, Grishin E. Cloning and structural analysis of alpha-latroinsectotoxin cDNA: abundance of ankyrin-like repeats. Eur J Biochem. 1993;213(1):121–127. doi: 10.1111/j.1432-1033.1993.tb17741.x. [DOI] [PubMed] [Google Scholar]
- Volkova TM, Galkina TG, Kudelin AB, Grishin EV. Structure of tryptic fragments of a neurotoxin from black widow spider venom. Bioorganicheskaya Khimiya. 1991;17(4):437–441. [PubMed] [Google Scholar]
- Volynski KE, Nosyreva ED, Ushkaryov YA, Grishin EV. Functional expression of α-latrotoxin in baculovirus system. FEBS Lett. 1999;442(1):25–28. doi: 10.1016/s0014-5793(98)01624-x. [DOI] [PubMed] [Google Scholar]
- Ushkaryov YA, Volynski KE, Ashton AC. The multiple actions of black widow spider toxins and their selective use in neurosecretion studies. Toxicon. 2004;43(5):527–542. doi: 10.1016/j.toxicon.2004.02.008. [DOI] [PubMed] [Google Scholar]
- Pescatori M, Bradbury A, Bouet F, Gargano N, Mastrogiacomo A, Grasso A. The cloning of a cDNA encoding a protein (latrodectin) which co-purifies with the alpha-latrotoxin from the black widow spider Latrodectus tredecimguttatus (Theridiidae) Eur J Biochem. 1995;230(1):322–328. doi: 10.1111/j.1432-1033.1995.tb20566.x.. [DOI] [PubMed] [Google Scholar]
- Kiyatkin N, Dulubova I, Chekhovskaya I, Lipkin A, Grishin E. Structure of the low molecular weight protein copurified with α-latrotoxin. Toxicon. 1992;30(7):771–774. doi: 10.1016/0041-0101(92)90012-t. [DOI] [PubMed] [Google Scholar]
- Volkova TM, Pluzhnikov KA, Woll PG, Grishin EV. Low molecular weight components from black widow spider venom. Toxicon. 1995;33(4):483–489. doi: 10.1016/0041-0101(94)00166-6.. [DOI] [PubMed] [Google Scholar]
- Gasparini S, Kiyatkin N, Drevet P, Boulain JC, Tacnet F, Ripoche P, et al. The low molecular weight protein which co-purifies with alpha-latrotoxin is structurally related to crustacean hyperglycemic hormones. J Biol Chem. 1994;269(31):19803–19809. [PubMed] [Google Scholar]
- McCowan C, Garb JE. Recruitment and diversification of an ecdysozoan family of neuropeptide hormones for black widow spider venom expression. Gene. 2014;536(2):366–375. doi: 10.1016/j.gene.2013.11.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grishin EV, Himmelreich NH, Pluzhnikov KA, Pozdnyakova NG, Storchak LG, Volkova TM, et al. Modulation of functional activities of the neurotoxin from black widow spider venom. FEBS Lett. 1993;336(2):205–207. doi: 10.1016/0014-5793(93)80803-3.. [DOI] [PubMed] [Google Scholar]
- Rueda A, Realpe E, Uribe A. Toxicity evaluation and initial characterization of the venom of a Colombian Latrodectus sp. Toxicon. 2017;125:53–58. doi: 10.1016/j.toxicon.2016.11.255. [DOI] [PubMed] [Google Scholar]
- Graudins A, Little MJ, Pineda SS, Hains PG, King GF, Broady KW, et al. Cloning and activity of a novel alpha-latrotoxin from red-back spider venom. Biochem Pharmac. 2012;83(1):170–183. doi: 10.1016/j.bcp.2011.09.024.. [DOI] [PubMed] [Google Scholar]
- Russell FE. Phosphodiesterase of some snake and arthropod venoms. Toxicon. 1966;4(2):153–154. doi: 10.1016/0041-0101(66)90009-2.. [DOI] [PubMed] [Google Scholar]
- Akhunov A, Golubenko Z. Isolation and study of the properties of kininase from the venom of spider Latrodectus tredecimguttatus. Adv Exp Med Biol. 1989;247A:313–318. doi: 10.1007/978-1-4615-9543-4_46. [DOI] [PubMed] [Google Scholar]
- Pansa MC, Natalizi GM, Bettini S. 5-hydroxytryptamine content of Latrodectus mactans tredecimguttatus venom from gland extracts. Toxicon. 1972;10(1):85–86. doi: 10.1016/0041-0101(72)90097-9.. [DOI] [PubMed] [Google Scholar]
- Horni A, Weickmann D, Hesse M. The main products of the low molecular mass fraction in the venom of the spider Latrodectus menavodi. Toxicon. 2001;39(2-3):425–428. doi: 10.1016/s0041-0101(00)00147-1. [DOI] [PubMed] [Google Scholar]
- Haney RA, Clarke TH, Gadgil R, Fitzpatrick R, Hayashi CY, Ayoub NA, et al. Effects of gene duplication, positive selection, and shifts in gene expression on the evolution of the venom gland transcriptome in widow spiders. Genome Biol Evol. 2016;8(1):228–242. doi: 10.1093/gbe/evv253.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunbar JP, Fort A, Redureau D, Sulpice R, Dugon MM, Quinton L. Venomics approach reveals a high proportion of Lactrodectus-like toxins in the venom of the noble false widow spider Steatoda nobilis. Toxins. 2020;12(6):402. doi: 10.3390/toxins12060402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garb JE, Hayashi CY. Molecular evolution of alpha-latrotoxin, the exceptionally potent vertebrate neurotoxin in black widow spider venom. Mol Biol Evol. 2013;30(5):999–1014. doi: 10.1093/molbev/mst011.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graudins A, Padula M, Broady K, Nicholson GM. Red-back spider (Latrodectus hasselti) antivenom prevents the toxicity of widow spider venoms. Ann Emerg Med. 2001;37(2):154–160. doi: 10.1067/mem.2001.113033. [DOI] [PubMed] [Google Scholar]
- Xu D, Wang X. Transcriptome analysis to understand the toxicity of Latrodectus tredecimguttatus eggs. Toxins. 2016;8(12) doi: 10.3390/toxins8120378.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Liu H, Duan Z, Cao R, Wang X, Liang S. Protein compositional analysis of the eggs of black widow spider (Latrodectus tredecimguttatus): implications for the understanding of egg toxicity. Journal of biochemical and molecular toxicology. 2012;26(12):510–515. doi: 10.1002/jbt.21460. [DOI] [PubMed] [Google Scholar]
- Correa-Garhwal SM, Chaw RC, Clarke TH, Ayoub NA, Hayashi CY. Silk gene expression of theridiid spiders: implications for male-specific silk use. Zoology. 2017;122:107–114. doi: 10.1016/j.zool.2017.04.003.. [DOI] [PubMed] [Google Scholar]
- Chaw RC, Correa-Garhwal SM, Clarke TH, Ayoub NA, Hayashi CY. Proteomic evidence for components of spider silk synthesis from black widow silk glands and fibers. J Proteome Res. 2015;14(10):4223–4231. doi: 10.1021/acs.jproteome.5b00353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christie AE. In silico characterization of the neuropeptidome of the Western black widow spider Latrodectus hesperus. Gen Comp Endocr. 2015;210:63–80. doi: 10.1016/j.ygcen.2014.10.005.. [DOI] [PubMed] [Google Scholar]
- Christie AE, Chi M. Neuropeptide discovery in the Araneae (Arthropoda, Chelicerata, Arachnida): elucidation of true spider peptidomes using that of the Western black widow as a reference. Gen Comp Endocr. 2015;213:90–109. doi: 10.1016/j.ygcen.2015.02.003. [DOI] [PubMed] [Google Scholar]
- Clarke TH, Garb JE, Hayashi CY, Arensburger P, Ayoub NA. Spider transcriptomes identify ancient large-scale gene duplication event potentially important in silk gland evolution. Genome Biol Evol. 2015;7(7):1856–1870. doi: 10.1093/gbe/evv110.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke TH, Garb JE, Hayashi CY, Haney RA, Lancaster AK, Corbett S, et al. Multi-tissue transcriptomics of the black widow spider reveals expansions, co-options, and functional processes of the silk gland gene toolkit. BMC Genomics. 2014;15:365. doi: 10.1186/1471-2164-15-365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng X, Dai Z, Wang X. Comparative proteomic analysis to probe into the differences in protein expression profiles and toxicity bases of Latrodectus tredecimguttatus spiderlings and adult spiders. Comp Biochem Physiol C Toxicol Pharmacol. 2020;232:108762. doi: 10.1016/j.cbpc.2020.108762.. [DOI] [PubMed] [Google Scholar]
- Veenstra JA. Neuropeptide evolution: chelicerate neurohormone and neuropeptide genes may reflect one or more whole genome duplications. Gen Comp Endocr. 2016;229:41–55. doi: 10.1016/j.ygcen.2015.11.019. [DOI] [PubMed] [Google Scholar]
- He Q, Duan Z, Yu Y, Liu Z, Liu Z, Liang S. The venom gland transcriptome of Latrodectus tredecimguttatus revealed by deep sequencing and cDNA library analysis. PLoS One. 2013;8(11):e81357. doi: 10.1371/journal.pone.0081357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duan Z, Yan X, Cao R, Liu Z, Wang X, Liang S. Proteomic analysis of Latrodectus tredecimguttatus venom for uncovering potential latrodectism-related proteins. Journal of biochemical and molecular toxicology. 2008;22(5):328–336. doi: 10.1002/jbt.20244. [DOI] [PubMed] [Google Scholar]
- Haney RA, Ayoub NA, Clarke TH, Hayashi CY, Garb JE. Dramatic expansion of the black widow toxin arsenal uncovered by multi-tissue transcriptomics and venom proteomics. BMC Genomics. 2014;15:366. doi: 10.1186/1471-2164-15-366.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khamtorn P, Rungsa P, Jangpromma N, Klaynongsruang S, Daduang J, Tessiri T, et al. Partial proteomic analysis of brown widow spider (Latrodectus geometricus) venom to determine the biological activities. Toxicon X. 2020;8:100062. doi: 10.1016/j.toxcx.2020.100062.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oldrati V, Koua D, Allard PM, Hulo N, Arrell M, Nentwig W, et al. Peptidomic and transcriptomic profiling of four distinct spider venoms. PLoS One. 2017;12(3):e0172966. doi: 10.1371/journal.pone.0172966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhere KV, Haney RA, Ayoub NA, Garb JE. Gene structure, regulatory control, and evolution of black widow venom latrotoxins. FEBS Lett. 2014;588(21):3891–3897. doi: 10.1016/j.febslet.2014.08.034.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mejia FR, Gutierrez RS, Obregón EB, De Miranda A, Fontaine AR. Contraceptive peptides derived from the venom of the spider Latrodectus mirabilis, nucleotide sequences to transform a microorganism to produce said peptides: methods to obtain the peptides: pharmaceutical compositions containing thereof and use thereof. United States patent. 2015 [Google Scholar]
- Mejia FGR, Patricio R, Salazar S, Miranda A. Compositions and methods for treating erectile dysfunction. United States patent. 2011 [Google Scholar]
- Wang X, Tang X, Xu D, Yu D. Molecular basis and mechanism underlying the insecticidal activity of venoms and toxins from Latrodectus spiders. Pest Manag Sci. 2019;75(2):318–323. doi: 10.1002/ps.5206. [DOI] [PubMed] [Google Scholar]
- Lei Q, Yu H, Peng X, Yan S, Wang J, Yan Y, et al. Isolation and preliminary characterization of proteinaceous toxins with insecticidal and antibacterial activities from black widow spider (L. tredecimguttatus) eggs. Toxins. 2015;7(3):886–899. doi: 10.3390/toxins7030886.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makover V, Ronen Z, Lubin Y, Khalaila I. Eggshell spheres protect brown widow spider (Latrodectus geometricus) eggs from bacterial infection. J Roy Soc Interface. 2019;16(150):20180581. doi: 10.1098/rsif.2018.0581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu D, Tang X, Wu X, Yu D, Tang P, Wang X. Anti-breast cancer activity of latroeggtoxin-V mined from the transcriptome of spider Latrodectus tredecimguttatus eggs. Toxins. 2018;10(11) doi: 10.3390/toxins10110451.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mousavi M, Zargan J, Haji-Noor M. A, Goudarzi HR, Dezianian S, Keshavarz-Alikhani H, et al. Anticancer effects of the Latrodectus dahli crude venom on MCF-7 breast cancer cell line. Breast J. 2019;25(4):781–782. doi: 10.1111/tbj.13332. [DOI] [PubMed] [Google Scholar]
- Rivera-de-Torre E, Palacios-Ortega J, Gavilanes JG, Martinez-Del-Pozo A, Garcia-Linares S. Pore-forming proteins from cnidarians and arachnids as potential biotechnological tools. Toxins. 2019;11(6) doi: 10.3390/toxins11060370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubista H, Mafra RA, Chong Y, Nicholson GM, Beirão PSL, Cruz JS, et al. CSTX-1, a toxin from the venom of the hunting spider Cupiennius salei, is a selective blocker of L-type calcium channels in mammalian neurons. Neuropharmacology. 2007;52(8):1650–1662. doi: 10.1016/j.neuropharm.2007.03.012. [DOI] [PubMed] [Google Scholar]
- Kuhn-Nentwig L, Fedorova IM, Lüscher BP, Kopp LS, Trachsel C, Schaller J, et al. A venom-derived neurotoxin, CsTx-1, from the spider Cupiennius salei exhibits cytolytic activities. J Biol Chem. 2012;287(30):25640–25649. doi: 10.1074/jbc.M112.339051.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siemens J, Zhou S, Piskorowski R, Nikai T, Lumpkin EA, Basbaum AI, et al. Spider toxins activate the capsaicin receptor to produce inflammatory pain. Nature. 2006;444(7116):208–212. doi: 10.1038/nature05285. [DOI] [PubMed] [Google Scholar]
- Craik DJ, Daly NL, Waine C. The cystine knot motif in toxins and implications for drug design. Toxicon. 2001;39(1):43–60. doi: 10.1016/s0041-0101(00)00160-4.. [DOI] [PubMed] [Google Scholar]